 Raimundo Real
Raimundo Real José C. Báez
José C. Báez Julia E. Fa
Julia E. Fa Jesús Olivero
Jesús Olivero Pelayo Acevedo
Pelayo Acevedo- 1Biogeography, Diversity, and Conservation Research Team, Department of Animal Biology, Universidad de Málaga, Málaga, Spain
- 2Instituto Español de Oceanografía (CSIF), Centro Oceanográfico de Málaga, Fuengirola, Spain
- 3Instituto Iberoamericano de Desarrollo Sostenible, Universidad Autónoma de Chile, Temuco, Chile
- 4Department of Natural Sciences, Manchester Metropolitan University, Manchester, United Kingdom
- 5Instituto de Investigación en Recursos Cinegéticos (IREC), UCLM-CSIC-JCCM, Ciudad Real, Spain
In biogeography the competitive exclusion principle (CEP) has been confirmed in some cases but not in others. This has fueled an unresolved debate between those advocating niche theory or the neutral theory in biodiversity and biogeography. We suggest that this situation mainly arises from the use of crisp logic, where the CEP is defined as either completely true or false. We propose the application of the fuzzy concepts of favorability (the degree to which environmental conditions are propitious for the occurrence of individual species) and favorableness (the degree to which environmental conditions are simultaneously favorable for competing species) to operationalize a fuzzy version of the CEP. Favorability was obtained by performing species distribution models applying favorability functions, while favorableness was derived from the application of the fuzzy intersection between the favorability for competing species. Then we plotted individual favorability values along the gradient of favorableness. Two potentially competing species would coexist in high-favorableness locations, as the demands of both species would be well fulfilled. In locations of low favorableness, the result would be either autecological exclusion of both species or autecological segregation, as abiotic conditions are unfavorable for at least one of the species. Competitive exclusion would occur at the intermediate stretch of the favorableness gradient, as the conditions would be good enough for persistence of each species separately but not enough for permanent coexistence. According to this theoretical framework, the observed probability that a location belongs to the intermediate favorableness area given that the two species co-occur in this location should be lower than expected according to the environmental probability models for the two species. We tested this prediction on published data about the distribution of pairs of native and introduced deer species in Great Britain, using a Bayesian approach. In two thirds of comparisons between a native and an introduced deer species the predictions of the fuzzy CEP were corroborated, which suggests that these are the pairs of species and the specific geographical areas affected by competitive exclusion. This is important both theoretically and for biodiversity conservation planning.
Introduction
Currently, the conciliation of the well-known competitive exclusion principle (CEP) and the well-known hypothesis of full functional redundancy (HFR), is an unresolved crucial question in ecology and evolutionary theory (Rodriguez et al., 2015; Kalmykov and Kalmykov, 2016; Cui et al., 2020; Hening and Nguyen, 2020; Wang and Liu, 2020; Wang and Wu, 2020). According to Rodriguez et al. (2015) the CEP and the HFR are simultaneously true and false in the same measure, because the only feasible option to keep the functional stability of ecosystems is a wave-like combination of both options.
The CEP suggests that two species with similar ecological requirements or functions cannot coexist. Gause (1934) proposed the concept in a way that could be experimentally tested. However, attempts to generalize the results of these experiments at a biogeographical scale have not been successful (Brown and Maurer, 1989).
Hardin (1960) argued that competitive exclusion occurs in nature under certain conditions as follows: “i) if two non-inter breeding populations “do the same thing”- that is, occupy precisely the same ecological niche in Elton’s sense - and ii) if they are “sympatric” - that is, if they occupy the same geographic territory - and iii) if population A multiplies even the least bit faster than population B, then ultimately A will completely displace B, which will become extinct. This is the “weak form” of the principle. Always in practice a stronger form is used based on the removal of the hypothetical character of condition (iii)”. Hardin (1960) further advocated that the stronger form of the principle is based on “what may be called the axiom of inequality, which states that no two things or processes in a real world, are precisely equal”. In this situation, as Hardin (1960) recognized, the CEP is a tautology that cannot be disproved by facts, although it could be valuable as a null model against which to contrast real data.
However, if the axiom of inequality is used to remove the hypothetical character of condition (iii), it may be used as well to remove conditions (i) and (ii) altogether, making the CEP unrealistic and, thus, of little use. This definition of the CEP is consequently insufficient, as reflected in mathematical modeling studies that had to deal with the difficulty of defining “similar ecology” (Munkemuller et al., 2009; Montes de Oca and Perez, 2012; Sari, 2013; Zhao et al., 2013; Kalmykov and Kalmykov, 2016; Xue and Goldenfeld, 2017; Cui et al., 2020; Hening and Nguyen, 2020; Wang and Wu, 2020).
Empirical evidence abounds both in favor (Metzger et al., 2009; Khelifa et al., 2013; Koelsch and Kubiak, 2013; Navarro et al., 2013; Rodriguez et al., 2015; Rapaport and Veruete, 2019) and against (Andersen et al., 2013; Smetacek and Zingone, 2013; Rodriguez et al., 2015; Lee et al., 2019; Antell et al., 2020; Wang and Liu, 2020) the CEP. These contradictory results have generated a debate between advocates of the CEP (Darlington, 1972; Gordon, 2000; Wang et al., 2005), and those supporting the full functional redundancy hypothesis (Walker, 1992; Lawton and Brown, 1993; Wohl et al., 2004; Petchey and Gaston, 2006; Mayfield and Levine, 2010; Rodriguez et al., 2015) and the neutral theory of biodiversity and biogeography (Hubbell, 2001; Rosindell et al., 2011).
The so-called ‘plankton paradox’, proposed by Hutchinson (1961), is a prime example of unexpected species coexistence according to the CEP. Hutchinson (1961) argued that the large number of different plankton species (mainly phytoplankton) that persist in a uniform environment, does not agree with the CEP, because they live in homogeneous environments, compete for the same resources, but still coexist. Several possible explanations of the plankton paradox have been proposed (Huisman and Weissing, 1999; Bracco et al., 2000; Scheffer et al., 2003; Shoresh et al., 2008; Fox et al., 2010; Segura et al., 2013; Li and Chesson, 2016), which basically agree with Hutchinson’s (1961) idea that plankton communities are not in equilibrium due to weather-driven variations (Hutchinson, 1961; Scheffer et al., 2003). Nonetheless, equilibrium may be as difficult to obtain in the real world as the conditions in Hardin’s (1960) CEP. The phenomenon was later generalized as the ‘biodiversity paradox’, which refers to the frequent contradiction between CEP predictions and the species richness found in nature at a biogeographical scale (Mayfield and Levine, 2010). The recent concern for biodiversity conservation has made this contradiction more central to ecological and biogeographical debates, because this situation does not clarify, for example, if, or where, invasive species may competitively exclude native species (Acevedo et al., 2010). As a result, resolving the biodiversity paradox has been considered one of the central issues in theoretical ecology (Tilman and Kareiva, 1998; Sommer, 1999).
Classical ad hoc accounts of the biodiversity paradox suggest that assumptions of the CEP have been violated (Schoener, 1976; Tilman, 1987; Palmer, 1994; Chunco et al., 2012; Xue and Goldenfeld, 2017; Roeleke et al., 2018) which, combined with the fact that these assumptions are extremely difficult to fulfill, renders the crisp definition of the CEP impractical. In an almost ironic conclusion to a simulation-based study, Kalmykov and Kalmykov (2013) stated that a species would be excluded from the habitat if each and every one of its individuals always had a direct conflict of interest with an individual of the fittest species and always loses, all other things being equal for all individuals of the competing species. This is tantamount to denying the applicability of the CEP, which should practically never be observed to occur (Kalmykov and Kalmykov, 2016). However, the fact that competitive exclusion has been also observed in nature seems to concur with Hardin’s (1960) assertion that the CEP has merit even if we still do not comprehend the exact limits of the principle.
Other observations advocate partial agreement and disagreement with the CEP, with the outcome depending on environmental conditions. The experimental introduction of the lizards Lacerta melisellensis and L. sicula in three Adriatic islands (Radovanovic, 1965), for instance, resulted in displacement of L. sicula by L. melisellensis in one of the islands, displacement of L. melisellensis by L. sicula in another island, and coexistence of the two species in the third island (Nevo et al., 1972). This suggests that competitive exclusion was acting only in two islands, favoring a different species in each. Results of experiments with the beetles Tribolium confusum and T. castaneum also showed that the dominance of one species over the other varied according to abiotic conditions (Park, 1954), with some instances of coexistence of both species that researchers considered difficult to interpret (Leslie et al., 1968). In a completely different scenario, patients infected with Hepatitis C were cleared of this virus when super-infected with Hepatitis A (Amaku et al., 2013), which was attributed to the CEP. However, when the action of the immune system was considered, both the elimination of one virus as well as the coexistence of both infections was possible within a range of parameters (Amaku et al., 2013).
Despite this empirical evidence, there is no theoretical model to predict (1) the simultaneous occurrence of coexistence and exclusion in different environmental conditions in a consistent way for any set of species, or (2) that a different competing species is expected to dominate when competitive exclusion is acting in differing environmental contexts. Here we provide a redefinition of the CEP based on fuzzy logic that can be better used to this end.
Fuzzy definition of the competitive exclusion principle
The classic Gause’s definition of the CEP is based on crisp (binary) logic, and therefore can only be applied with difficulty to living beings that are distributed in time and space essentially in a gradual and non-discrete manner. Evolving from the need to model the type of vague, ill-defined systems difficult to handle using the conventional binary valued logic, Zadeh (1965) proposed the fuzzy set theory, which is the basis of the multi-valued fuzzy logic. Fuzzy logic avoids the discrete true-or-false syllogisms, thus conferring a conceptual malleability that is not possible under a rigid definition of, for example, equality in the traditional meaning of the CEP. A fuzzy logic perspective has been shown to be particularly useful for processing environmental data, which tend to be uncertain or imprecise (Salski, 2006). Fuzzy logic uses concepts that are inherently gradual and imply a degree of membership in two antithetic states such as, for example, similar-dissimilar or favorable-unfavorable. The fuzzy notions of favorability for a species presence and favorableness for several species are particularly useful for redefining the CEP in a more realistic and operational way.
Favorableness was defined as “the effect of the mean of the environmental variables on diversity”, or as the degree to which the physical conditions needed to maintain the basic biochemical machinery shared by living systems range from the optimum (highest favorableness) to the lethal (lowest favorableness) limit (Richerson and Lum, 1980). This implicitly fuzzy term was applied to multiple species under the hypothesis that when environmental conditions are very unfavorable for the species, organisms must increase the variety of adaptations, energy and material resources devoted to coping with these conditions, at the expense of microhabitat-specific specializations and co-evolutionary accommodation with other species. Conversely, when environmental conditions are very favorable, organisms can dedicate a higher amount of energy, matter and genome to co-adaptive adjustments to other species and, therefore, will coexist more easily. Favorableness is then the degree to which the environment is simultaneously favorable for the set of competing species. This concept was used as an abstract notion with no operational method to measure it, or any explicit range of favorableness values (Richerson and Lum, 1980).
Favorability was defined as the degree (between 0 and 1) to which local environmental conditions are propitious for the presence of a species (Real et al., 2006), and the term was applied to the occurrence of individual species at different locations. Favorability is a concept related to, but different from, probability of occurrence (Supplementary material 1), being the membership function in the fuzzy set of locations favorable for the occurrence of a species. Thus, the fuzzy intersection between several such fuzzy sets could be used to identify areas simultaneously favorable to several species (Real et al., 2006), thus able to obtain favorableness values (i.e., minimum of the favorability values) ranging between 0 and 1 for any set of species. Both concepts can then be assigned values directly drawn from observations of the species distributions in different environmental conditions. This allows favorability and favorableness to be combined in a redefinition of the CEP in the framework of fuzzy set theory.
The degree to which autecological conditions (i.e., the living and non-living factors of the environment directly influencing a single species) are favorable for each species, i.e., favorability (Real et al., 2006), can be plotted alongside the degree to which conditions are simultaneously favorable for overlapping species, i.e., favorableness (the function SharedFav in the R package FuzzySim can be used to this end, Barbosa, 2015). This can help to understand a range of likely biogeographical outcomes, such as autecological exclusion or segregation, competitive exclusion, or species co-existence, in different parts of the ranges of competing species (Figure 1). In this theoretical framework, two potentially competing species would coexist in high-favorableness locations (Figure 1). These high-favorableness areas could also be seen as places where the demands of both species are well fulfilled. If competition is defined as a process that happens only when several species make the same or similar demands in the absence of an adequate supply (Clements and Shelford, 1939), then this process would be less pronounced in high favorableness areas where, consequently, competitive exclusion is unlikely to occur. In locations of low favorableness, the result would be either autecological exclusion of both species or autecological segregation, as abiotic conditions are unfavorable for at least one of the species (Figure 1). Competitive exclusion would occur at the intermediate stretch of the favorableness gradient, as the conditions are good enough for persistence of each species separately but not enough for permanent coexistence (Figure 1). The species with the highest favorability in this intermediate-favorableness stretch is expected to dominate over the other. This is a more nuanced and operational theory against which to contrast empirical data.
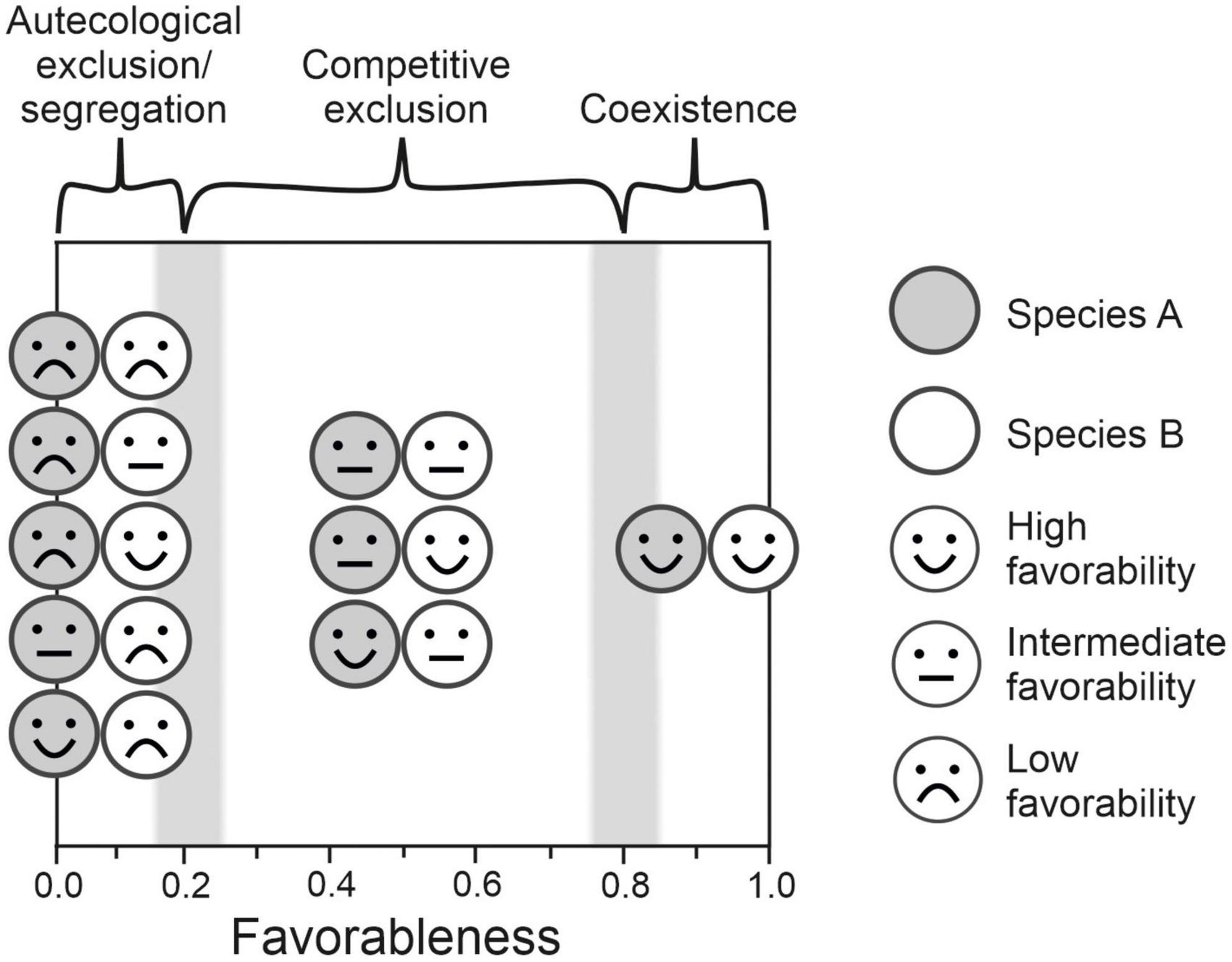
Figure 1. Expected biogegraphical outcomes (autecological exclusion/segregation, competitive exclusion and coexistence) of species interaction according to the proposed theoretical framework combining favorability and favorableness. Favorability is defined as the degree (between 0 and 1) to which autecological conditions that are favorable for the occurrence of individual species. Favorableness is defined as the fuzzy intersection of favorability values (i.e., minimum) for several species, i.e., the degree to which conditions are auspicious for all species together. If the favorableness for two species A and B together is low, at least one of them is exposed to low environmental favorability, and they would experience exclusion or segregation due to autecological causes; if the favorableness for both species together is high, both enjoy high environmental favorability, and the two species would be expected to coexist as the demands of both species would be well fulfilled; competitive exclusion would occur if the favorableness for both species together is intermediate, as environmental conditions are favorable enough for persistence of each species separately but the presence of a competitor become detrimental. Fuzzy columns represent the expected fuzzy thresholds between autecological exclusion/segregation, competitive exclusion and coexistence.
Theoretical connection of the original and fuzzy-logic definitions of the competitive exclusion principle
To better understand the theoretical setting of this proposal, we may connect it with the formulae used by Gause (1932) in his experimental studies about competitive exclusion. These formulae rest on the assumption that the population growth of any species on a given surface has an intrinsic stationary limit K, and that the population growth proceeds at a slower pace as the number of the individuals already produced (y) approaches the stationary number (Gause, 1934 in the pp. 8-9). In other words, the population growth is proportional to the unutilized potential of growth, which is given by (K-y)/K (Gause, 1932). In mixed populations with a second competing species this potential is affected by a new quantity m that represents the places (sic) of the first species that are taken up by the competing species, and the potential for growth is given by [K-(y + m)]/K = (K- m - y)/K (Gause, 1932). In this situation, the effective stationary limit is reduced in the numerator to K-m, which represents the degree of the competitive exclusion.
K is intrinsic for any species, genetically determined by its metabolism and by density-dependent factors such as territoriality or other spatio-temporal configurations of the population. Density-dependent limiting factors tend to be biotic rather than physical features of the environment. Among them are predation (higher-density populations may particularly attract predators), infectious diseases and parasites (infectious diseases are more likely to break out and result in deaths and parasites are more likely to spread in denser populations), or waste accumulation (high population densities can lead to the accumulation of harmful waste products). Santini et al. (2022), for example, found that the variation of population density across mammal species is largely explained, apart from resource availability and accessibility, by their body mass and by genetically determined metabolism and energy demand. Other examples of genetically determined limits to the use of resources may be seen in the fact that the feeding of animals in captivity under ad libitum conditions often results in a shorter lifespan and a decreased health, due to an increase in metabolic and degenerative diseases (Moraal et al., 2012). Thus, the local environmental favorability F may allow for a theoretical limit K’ that can be higher or lower than the K value, constituting an autecological limit for growth in the latter case. Then, the possibilities taken up by the competing species (m) may be considered to detract from K’ (the potential allowed by F). When the local favorableness allows for a higher K’ than the actual intrinsic K for two competing species, the effect of each competitor affects the surplus in possibilities of the other but does not detract from K, and the competitive exclusion does not happen. At the point where the local F allows exactly for the intrinsic K or lower (K’ ≤ K), the value of m detracts from K, exactly as Gause (1932, 1934) proposed, and the competitive exclusion does occur. When the local F, and thus K’, is extremely low, the autecological limitation may be enough to explain the exclusion of the species from the territory, making the additional effect of competition negligible.
The transition from autecological exclusion to competitive exclusion would be gradual, but around an F value of 0.2, as the response of the favorability function to environmental conditions is logistic, and F values approach 0 asymptotically from 0.2 as environmental conditions worsen. The response of the favorability function to environmental conditions is quasi linear for F values ranging between 0.2 and 0.8, and then this response is again asymptotically approaching 1 as environmental conditions improve further. This is why the fuzzy transition between competitive exclusion and coexistence is expected to be around an F value of 0.8 (Figure 1). Note that these values 0.2 and 0.8 represent the fuzzy equivalent of the odds ratios 1:4 and 4:1, respectively. As the complement of favorability (F) is unfavorability (1-F), at F = 0.2 unfavorability is equal to 0.8 and the ratio of favorability to unfavorability is 0.2:0.8 = 1:4, i.e., the environmental conditions are four times more unfavorable than favorable. Similarly, at F = 0.8 unfavorability is equal to 0.2 and the ratio of favorability to unfavorability is 0.8:0.2 = 4:1 and environmental conditions are four times more favorable than unfavorable. This is like this irrespective of the parametrization of the logistic model.
This unified theory of favorability and favorableness may be helpful for distinguishing competitive exclusion from the mere identity of species’ niches (Warren et al., 2008). As inferred by Hardin (1960), two species (or populations) can compete if they occupy the same ecological niche. To understand our view, it is necessary to apply fuzzy logic also to the niche concept. If the niche, either Eltonian, Ginnellian or Hutchinsonian, is conceived crisply, defined by its position, boundaries, size, shape and overlap, then there is no practical difference for the species in any location within this crisp theoretical niche. Interestingly, Hutchinson (1957, p. 416-418) applied the classical set theory to define the niche and admitted there were limitations in this mode of expression. Such limitations included the supposition that all points in each niche implied equal probability of species persistence, and all points outside the niche implied zero probability of survival. Here, the application of the fuzzy set theory overcame these limitations and allowed to identify an internal structure of the niche. This is in line with the ideas of Maguire (1973), who defined the niche as the evolutionarily-determined gradual responses of species in function of the habitat conditions. Favorability models were in fact functions and treated the niche as an interaction between the species and the environment, as Maguire (1973) did (Gouveia et al., 2020). Using fuzzy logic, the sameness of the niches occupied by two species is replaced by the similarity in their interaction with the environmental conditions, in terms of their respective functions of favorability for occurrence (Real et al., 2006).
Biogeographical implications
This theory may be also helpful for defining explicit competitors differentially in diverse parts of their geographical ranges. By visualizing species interactions in a biogeographic framework using fuzzy logic, we are better able to consider the CEP as deriving from the geographic distribution of different degrees of favorability enjoyed by competing species throughout their geographical ranges. Hence, the set of environmental conditions favorable for each species can be geographically present in a gradient from very favorable to very unfavorable, and from simultaneously favorable to differentially favorable to competing species. This means that ecological interactions may act differently in contrasting situations (and in different locations). The local intensity of the competition between two species will then depend on the similarity of their respective functions of favorability for occurrence and on the actual availability of shared (more competition) or exclusive (less competition) resources in the local sites.
Empirical examples of this kind of geographically differentiated biotic interaction have been hypothesized at a continental scale for parapatric hares (Lepus species) in Europe (Figure 2A; Acevedo et al., 2012), at a regional scale for native and introduced deer species in Great Britain (Figure 2B; Acevedo et al., 2010), and at a local scale for the native Mediterranean pond turtle (Mauremys leprosa) and the sympatric invasive red-eared slider (Trachemys scripta) in Spain (Romero et al., 2014). In Supplementary material 2 we show with a detailed example that the predictions of the fuzzy logic definition of the CEP about the distribution of pairs of native and introduced deer species in Great Britain can be operationally tested using a Bayesian approach. In two thirds of pairs of a native and an introduced deer species the predictions of the fuzzy CEP were corroborated, which suggests that these pairs of species are affected by competitive exclusion, being also able to locate the areas where competitive exclusion is acting.
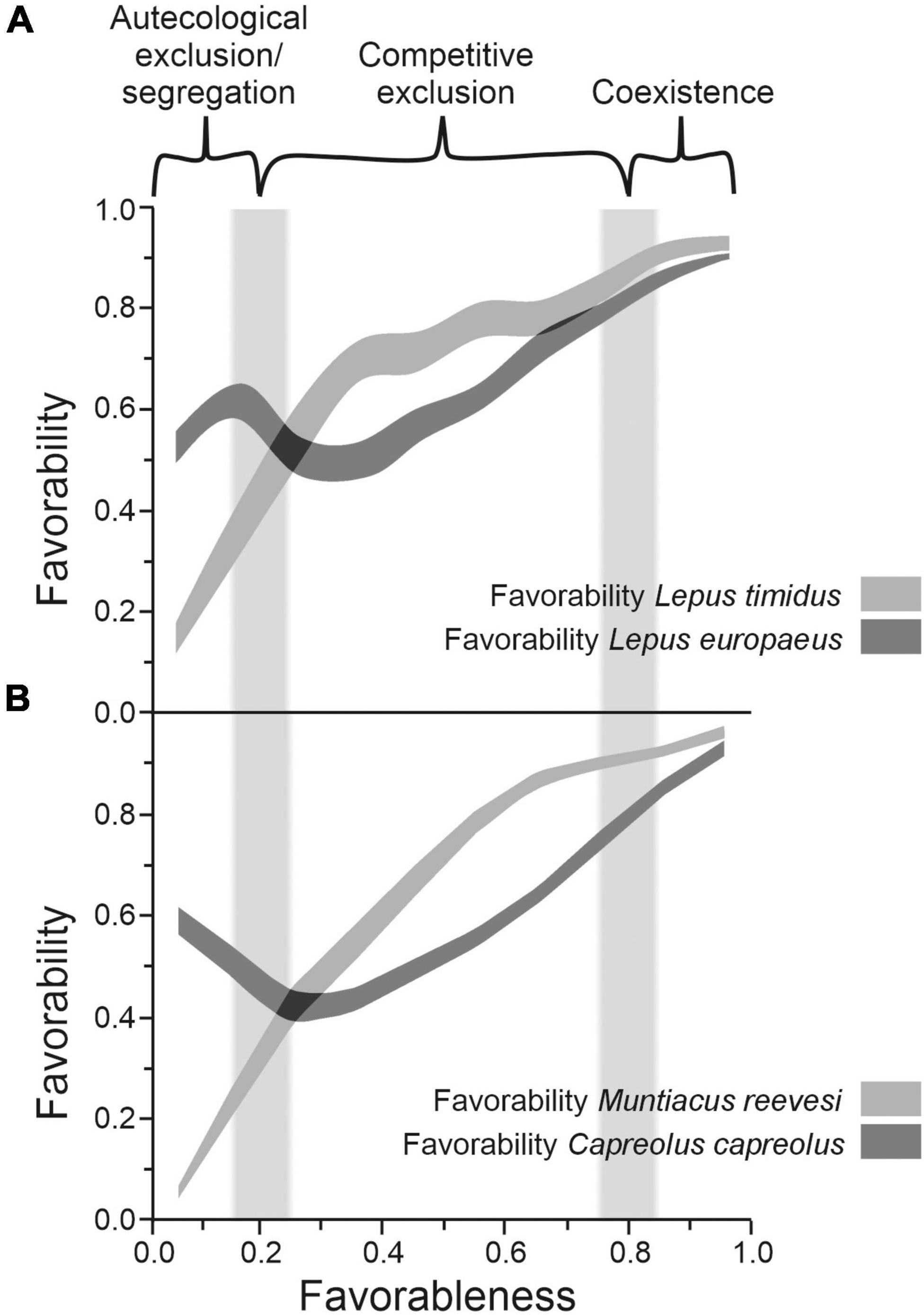
Figure 2. Examples of biogeographical outcomes of species interaction after representing the mean favorability of individual competing species along their favorableness grandient. Favorableness and favorability are defined as in Figure 1. (A) Lepus timidus and L. europaeus in Europe (Acevedo et al., 2012). (B) Muntiacus reevesi and Capreolus capreolus in Great Britain (Acevedo et al., 2010). Plots result of joining the 95% confidence intervals of mean favorability values of the locations grouped in 10 bins of equal range along the favorableness gradient. Fuzzy columns as in Figure 1.
Discussion
The above examples corroborate that a model of differing biogeographical outcomes of ecological interactions, generalizable to any two or more competing species, can be generated. Favorability is a function of environmental factors, and the identification of these factors (absence of disturbance, nutrient supply rate, energy or water availability, climatic conditions, habitat connectivity…) is the realm of biogeography and species distribution modeling. At smaller ecological scales it is still necessary to investigate how individual fitness and/or demography depend on population densities, or how the presence of more individuals of one species affects the favorability for conspecifics and heterospecifics. However, favorability may be operationally obtained from probability of occurrence and species prevalence at biogeographic scales, and favorableness for a set of species may be operationally derived from individual favorability values using fuzzy intersection.
Within this framework, competitive exclusion between two species is not expected to occur always or not occur at all, but the outcome will depend on the environmental conditions. The CEP is valid but restricted to areas of intermediate favorableness. Given this, the biodiversity paradox can be resolved; several competing species may coexist if environmental conditions are favorable for all of them. Our approach responds to Hubbell’s (2001) call for better theories for species presence-absence, relative abundance and persistence times in communities that can be confronted with real data. Our proposal produces differential observable expectations that may be corroborated or refuted with observations. It provides a more realistic and operational framework that yields more detailed predictions of coexistence and competitive exclusion at geographical scale and, thus, is more falsifiable than the classical CEP. We also argue that rendering the CEP operational with fuzzy logic is crucial for the conservation of biodiversity. More generally, the use of fuzzy approaches should be advocated to better understand the ecological interaction between overlapping species. If the ecological universe seems fuzzy and unpredictable (Turnbull, 2014), then the use of fuzzy logic may be appropriate to forecast the different outcomes of the complex ecological interactions between species in a heterogeneous geographical environment.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author/s.
Author contributions
RR and JB conceived the project. PA provided the data. RR conducted the analyses. All authors contributed to writing and discussions.
Acknowledgments
We are grateful to Benjamin Bolker for his insights and comments to a previous version of the manuscript. This work has been supported by the Project P20_01063 of the Junta de Andalucía (Spain). PA was currently supported by the Spanish Ministry of Economy, Industry and Competitiveness and Universidad de Castilla-La Mancha (UCLM) through a ‘Ramón y Cajal’ contract (RYC- 2012-11970).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.991344/full#supplementary-material
References
Acevedo, P., Jiménez-Valverde, A., Melo-Ferreira, J., Real, R., and Alves, P. C. (2012). Parapatric species and the implications for climate change studies: a case study on hares in Europe. Glob. Change Biol. 18, 1509–1519. doi: 10.1111/j.1365-2486.2012.02655.x
Acevedo, P., Ward, A. I., Real, R., and Smith, G. C. (2010). Assessing biogeographical relationships of ecologically related species using favourability functions: a case study on British deer. Divers. Distrib. 16, 515–528. doi: 10.1111/j.1472-4642.2010.00662.x
Amaku, M., Coutinho, F. A. B., Chaib, E., and Massad, E. (2013). The impact of hepatitis A virus infection on hepatitis C virus infection, A competitive exclusion hypothesis. Bull. Mathe. Biol. 75, 82–93. doi: 10.1007/s11538-012-9795-0
Andersen, N. A., Arnan, X., and Sparks, K. (2013). Limited niche differentiation within remarkable co-occurrences of congeneric species: monomorium ants in the Australian seasonal tropics. Austral Ecol. 38, 557–567. doi: 10.1111/aec.12000
Antell, G. S., Kiessling, W., Aberhan, M., and Saupe, E. E. (2020). Marine biodiversity and geographic distributions are independent on large scales. Curr. Biol. 30, 115–121.e5. doi: 10.1016/j.cub.2019.10.065
Barbosa, A. M. (2015). FuzzySim: applying fuzzy logic to binary similarity indices in ecology. Methods Ecol. Evol. 6, 853–858. doi: 10.1111/2041-210X.12372
Bracco, A., Provenzale, A., and Scheuring, I. (2000). Mesoscale vortices and the paradox of the plankton. Proc. R. Soc. Biol. Sci. 267, 1795–1800. doi: 10.1098/rspb.2000.1212
Brown, J. H., and Maurer, B. A. (1989). Macroecology: the division of food and space among species on continents. Science 243, 1145–1150. doi: 10.1126/science.243.4895.1145
Chunco, A. J., Jobe, T., and Pfennig, K. S. (2012). Why do species co-occur? A test of alternative hypotheses describing abiotic differences in sympatry versus allopatry using spadefoot toads. PLoS One 7:e32748. doi: 10.1371/journal.pone.0032748
Cui, W. P., Marsland, R., and Mehta, P. (2020). Effect of resource dynamics on species packing in diverse ecosystems. Phys. Rev. Lett. 125:48101. doi: 10.1103/PhysRevLett.125.048101
Darlington, P. J. Jr. (1972). Competition, competitive repulsion, and coexistence. Proc. Natl. Acad. Sci. U.S.A. 69, 3151–3315. doi: 10.1073/pnas.69.11.3151
Fox, J. W., Nelson, W. A., and McCauley, E. (2010). Coexistence mechanisms and the paradox of the plankton, quantifying selection from noisy data. Ecology 91, 1774–1786. doi: 10.1890/09-0951.1
Gause, G. F. (1932). Experimental studies on the struggle for existence. I. mixed population of two species of yeast. J. Exp. Biol. 9, 389–402. doi: 10.1242/jeb.9.4.389
Gause, G. F. (1934). The Struggle for Existence. Baltimore: Williams & Wilkins. doi: 10.5962/bhl.title.4489
Gordon, C. E. (2000). The coexistence of species. Rev. Chil. Hist. Nat. 73, 175–198. doi: 10.4067/S0716-078X2000000100016
Gouveia, S. F., Rubalcaba, J. G., Soukhovolsky, V., Tarasova, O., Barbosa, A. M., and Real, R. (2020). Ecophysics reload — exploring applications of theoretical physics in macroecology. Ecol. Modell. 424:109032. doi: 10.1016/j.ecolmodel.2020.109032
Hardin, G. (1960). The competitive exclusion principle. Science 13, 1292–1297. doi: 10.1126/science.131.3409.1292
Hening, A., and Nguyen, D. H. (2020). The competitive exclusion principle in stochastic environments. J. Math. Biol. 80, 1323–1351. doi: 10.1007/s00285-019-01464-y
Hubbell, S. P. (2001). The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press.
Huisman, J., and Weissing, F. J. (1999). Biodiversity of plankton by species oscillations and chaos. Nature 402, 407–410. doi: 10.1038/46540
Hutchinson, G. E. (1957). Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 22, 415–427. doi: 10.1101/SQB.1957.022.01.039
Kalmykov, L. V., and Kalmykov, V. L. (2013). Verification and reformulation of the competitive exclusion principle. Chaos Solit. Fract. 56, 124–131. doi: 10.1016/j.chaos.2013.07.006
Kalmykov, V. L., and Kalmykov, L. V. (2016). On ecological modelling problems in the context of resolving the biodiversity paradox. Ecol. Modell. 329, 1–4. doi: 10.1016/j.ecolmodel.2016.03.005
Khelifa, R., Zebsa, R., Moussaoui, A., Kahalerras, A., Ben-Souilah, S., and Mahdjoub, H. (2013). Niche partitioning in three sympatric congeneric species of dragonfly, Orthetrum chrysostigma, O. coerulescens anceps, and O. nitidinerve: the importance of microhabitat. J. Insect Sci. 13:71. doi: 10.1673/031.013.7101
Koelsch, G., and Kubiak, M. (2013). The aquatic leaf beetle species Macroplea mutica and M. appendiculata (Coleoptera, Chrysomelidae, Donaciinae) differ in their use of Myriophyllum spicatum as a host plant. Aquat. Insects 33, 13–26. doi: 10.1080/01650424.2011.572558
Lawton, J. H., and Brown, V. K. (1993). “Redundancy in ecosystems,” in Biodiversity and Ecosystem Function, eds E. D. Schulze and H. A. Mooney (Berlin: Springer), 255–270. doi: 10.1007/978-3-642-58001-7_12
Lee, M. R., Powell, J. R., Oberle, B., Cornwell, W. K., Lyons, M., Rigg, J. L., et al. (2019). Good neighbors aplenty: fungal endophytes rarely exhibit competitive exclusion patterns across a span of woody habitats. Ecology 100:e02790. doi: 10.1002/ecy.2790
Leslie, P. H., Park, T., and Mertz, D. B. (1968). The effect of varying the initial numbers on the outcome of competition between two Tribolium species. J. Anim. Ecol. 37, 9–23. doi: 10.2307/2708
Li, L., and Chesson, P. (2016). The effects of dynamical rates on species coexistence in a variable environment: the paradox of the plankton revisited. Am. Nat. 188, 46–58. doi: 10.1086/687111
Maguire, B. Jr. (1973). Niche response structure and the analytical potentials of its relationship to the habitat. Am. Nat. 107, 213–246. doi: 10.1086/282827
Mayfield, M. M., and Levine, J. M. (2010). Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecol. Lett. 13, 1085–1093. doi: 10.1111/j.1461-0248.2010.01509.x
Metzger, C., Ursenbacher, S., and Christe, P. (2009). Testing the competitive exclusion principle using various niche parameters in a native (Natrix maura) and an introduced (N. tessellata) colubrid. Amphib. Reptil. 30, 523–531. doi: 10.1163/156853809789647031
Montes de Oca, F., and Perez, L. (2012). Extinction in non autonomous competitive Lotka-Volterra systems with infinite delay. Nonlinear Anal. 75, 758–768. doi: 10.1016/j.na.2011.09.009
Moraal, M., Leenaars, P. P. A. M., Arnts, H., Smeets, K., Savenije, B. S., Curfs, J. H. A. J., et al. (2012). The influence of food restriction versus ad libitum feeding of chow and purified diets on variation in body weight, growth and physiology of female Wistar rats. Lab. Anim. 46, 101–107. doi: 10.1258/la.2011.011011
Munkemuller, T., Bugmann, H., and Johst, K. (2009). Hutchinson revisited: patterns of density regulation and the coexistence of strong competitors. J. Theor. Biol. 259, 109–117. doi: 10.1016/j.jtbi.2009.03.010
Navarro, J., Votier, S. C., Aguzzi, J., Chiesa, J. J., Forero, M. G., and Phillips, R. A. (2013). Ecological segregation in space, time and trophic niche of sympatric planktivorous Petrels. PLoS One 8:e0062897. doi: 10.1371/journal.pone.0062897
Nevo, E., Gorman, G., Soulé, M., Yang, S., Clover, R., and Jovanoviæ, V. (1972). Competitive exclusion between insular Lacerta species (Sauria, Lacertidae). Oecologia 10, 183–190. doi: 10.1007/BF00347990
Palmer, M. W. (1994). Variation in species richness: towards a unification of hypotheses. Folia Geobot. Phytotaxon. 29, 511–530. doi: 10.1007/BF02883148
Park, T. (1954). Experimental studies of interspecies competition II. Temperature, humidity, and competition in two species of Tribolium. Physiol. Zool. 27, 177–238. doi: 10.1086/physzool.27.3.30152164
Petchey, O. L., and Gaston, K. J. (2006). Functional diversity: back to basics and looking forward. Ecol. Lett. 9, 741–758. doi: 10.1111/j.1461-0248.2006.00924.x
Radovanovic, M. (1965). Experimentelle beiträge zum problem der kompetition. Zool. Anzeiger 29, 534–539.
Rapaport, A., and Veruete, M. (2019). A new proof of the competitive exclusion principle in the chemostat. Discr. Cont. Dyn. Syst. Series B 24, 3755–3764. doi: 10.3934/dcdsb.2018314
Real, R., Barbosa, A. M., and Vargas, J. M. (2006). Obtaining environmental favourability functions from logistic regression. Environ. Ecol. Stat. 13, 237–245. doi: 10.1007/s10651-005-0003-3
Richerson, P. J., and Lum, K. (1980). Patterns of plant species diversity in California: relation to weather and topography. Am. Nat. 116, 504–536. doi: 10.1086/283645
Rodriguez, R. A., Herrera, A. M., Santander, J., Miranda, J. V., Fernández-Rodríguez, M. J., Quirós, A., et al. (2015). Uncertainty principle in niche assessment: a solution to the dilemma redundancy vs. competitive exclusion, and some analytical consequences. Ecol. Modell. 316, 87–110. doi: 10.1016/j.ecolmodel.2015.07.032
Roeleke, M., Johannsen, L., and Voigt, C. C. (2018). How bats escape the competitive exclusion principle—seasonal shift from intraspecific to interspecific competition drives space use in a bat ensemble. Front. Ecol. Evol. 6:101. doi: 10.3389/fevo.2018.00101
Romero, D., Báez, J. C., Ferri-Yáñez, F., Bellido, J. J., and Real, R. (2014). Modelling favourability for invasive species encroachment to identify areas of native species vulnerability. Sci. World J. 2014:519710. doi: 10.1155/2014/519710
Rosindell, J., Hubbell, S. P., and Etienne, R. S. (2011). The unified neutral theory of biodiversity and biogeography at age ten. Trends Ecol. Evol. 26, 340–348. doi: 10.1016/j.tree.2011.03.024
Salski, A. (2006). “Scope, techniques and applications,” in Ecological Informatics, ed. F. Recknagel (Berlin: Springer-Verlag), 3–14.
Santini, L., Benítez-López, A., Dormann, C. F., and Huijbregts, M. A. J. (2022). Population density estimates for terrestrial mammal species. Glob. Ecol. Biogeogr. 31, 978–994. doi: 10.1111/geb.13476
Sari, T. (2013). Competitive exclusion for chemostat equations with variable yields. Acta Appl. Math. 123, 201–219. doi: 10.1007/s10440-012-9761-8
Scheffer, M., Rinaldi, S., Huisman, J., and Weissing, F. J. (2003). Why plankton communities have no equilibrium: solutions to the paradox. Hydrobiologia 491, 9–18. doi: 10.1023/A:1024404804748
Schoener, T. W. (1976). Alternatives to lotka-volterra competition, models of intermediate complexity. Theor. Popul. Biol. 10, 309–333. doi: 10.1016/0040-5809(76)90022-8
Segura, A. M., Kruk, C., Calliari, D., García-Rodriguez, F., Conde, D., Widdicombe, C. E., et al. (2013). Competition drives clumpy species coexistence in estuarine phytoplankton. Sci. Rep. 3:1037. doi: 10.1038/srep01037
Shoresh, N., Hegreness, M., and Kishony, R. (2008). Evolution exacerbates the paradox of the plankton. Proc. Natl. Acad. Sci. U.S.A. 105, 12365–12369. doi: 10.1073/pnas.0803032105
Smetacek, V., and Zingone, A. (2013). Green and golden seaweed tides on the rise. Nature 504, 84–88. doi: 10.1038/nature12860
Tilman, D. (1987). The importance of the mechanisms of interspecific competition. Am. Nat. 129, 769–774. doi: 10.1086/284672
Tilman, D., and Kareiva, P. M. (1998). The Role of Space in Population Dynamics and Interspecific Interactions. Princeton, NJ: Princeton University Press.
Turnbull, L. A. (2014). Ecology’s dark matter: the elusive & enigmatic niche. Basic Appl. Ecol. 15, 93–100. doi: 10.1016/j.baae.2013.10.007
Walker, B. H. (1992). Biodiversity and ecological redundancy. Conserv. Biol. 6, 18–22. doi: 10.1046/j.1523-1739.1992.610018.x
Wang, X., and Liu, Y. Y. (2020). Overcome competitive exclusion in ecosystems. iScience 23:101009. doi: 10.1016/j.isci.2020.101009
Wang, Y. S., and Wu, H. (2020). Global dynamics of Lotka-Volterra equations characterizing multiple predators competing for one prey. J. Math. Anal. Appl. 491:124293. doi: 10.1016/j.jmaa.2020.124293
Wang, Z. L., Zhang, D. Y., and Wang, G. (2005). Does spatial structure facilitate coexistence of identical competitors? Ecol. Modell. 181, 17–23. doi: 10.1016/j.ecolmodel.2004.06.020
Warren, D. L., Glor, R. E., and Turelli, M. (2008). Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution 62, 2868–2883. doi: 10.1111/j.1558-5646.2008.00482.x
Wohl, D. B., Arora, S., and Gladstone, J. R. (2004). Functional redundancy supports biodiversity and ecosystem function in a closed and constant environment. Ecology 85, 1534–1540. doi: 10.1890/03-3050
Xue, C., and Goldenfeld, N. (2017). Coevolution maintains diversity in the stochastic “Kill the Winner” model. Phys. Rev. Lett. 119, 268101. doi: 10.1103/PhysRevLett.119.268101
Keywords: coexistence, ecological interactions, favorability, favorableness, macroecology, niche, species distribution
Citation: Real R, Báez JC, Fa JE, Olivero J and Acevedo P (2022) Making the competitive exclusion principle operational at the biogeographical scale using fuzzy logic. Front. Ecol. Evol. 10:991344. doi: 10.3389/fevo.2022.991344
Received: 11 July 2022; Accepted: 22 August 2022;
Published: 23 September 2022.
Edited by:
Sidney F. Gouveia, Federal University of Sergipe, BrazilReviewed by:
Paulo Campos, Federal University of Pernambuco, BrazilNiels Hellwig, Thünen Institute of Biodiversity, Germany
Copyright © 2022 Real, Báez, Fa, Olivero and Acevedo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raimundo Real, cnJnaW1lbmV6QHVtYS5lcw==