
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 08 September 2022
Sec. Ecophysiology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.971429
This article is part of the Research TopicBehavioral and Physiological Adaptations of Mammals and Birds to Anthropogenic DisturbancesView all 11 articles
 Caijun Zhang1,2
Caijun Zhang1,2 Yukun Kang1,2Baohui Yao1,2Kang An1,2Qiangsheng Pu1,2Zhicheng Wang1,2Xiaomei Sun2,3
Yukun Kang1,2Baohui Yao1,2Kang An1,2Qiangsheng Pu1,2Zhicheng Wang1,2Xiaomei Sun2,3 Junhu Su1,2*
Junhu Su1,2*The increased population density of rodent species during ongoing grassland degradation further deteriorates its conditions. Understanding the effects of grassland degradation on rodent feeding habits is of great value for optimizing grassland management strategies. In this study, lightly degraded (LD), moderately degraded (MD), severely degraded (SD), and reseeded grassland (RG) were selected and their plant resources and soil physical properties were investigated. In addition, the study used ITS2 barcode combined with the Illumina MiSeq sequencing method to analyze the food composition and proportion of plateau zokors in different grassland conditions. The results showed that, with grassland degradation, plant biomass decreased, but the relative proportion of forbs increased (LD: 32.05 ± 3.89%; MD: 28.97 ± 2.78%; SD: 49.16 ± 4.67% and RG: 10.93 ± 1.53%). Forbs were the main food of the plateau zokor, accounting for more than 90% of their diet, and the animal had a clear preference for Potentilla species; the soil compaction of feeding habits showed a decreasing trend in the 10–25 cm soil layer, suggesting a decreased foraging cost. Nutritional analysis showed that the stomach content of crude protein in zokors feeding on MD grassland was significantly higher than that of animals feeding on the other grassland types. Structural equation modeling showed that soil physical properties and the relative biomass of forbs had significant (P < 0.05) and extremely significant (P < 0.001) impacts on the population density of plateau zokors, with direct impact contribution rates of 0.20 and 0.63. As the severity of grassland degradation increased, although the aboveground and underground biomass of the plants decreased, the proportion of food preferred by the plateau zokor increased, and the corresponding changes in the feeding environment resulted in decreased foraging energy expenditure, thereby increasing the suitability of the degraded grassland for the plateau zokor. Compared with degraded grassland, the food diversity and evenness of zokors increased, the food niche width enlarged, and the proportion of weeds decreased in RG, which increased the difficulty of obtaining food. Reseeding in grassland management is therefore an effective way to control plateau zokors.
Grassland degradation from climate change and human activity is a global environmental problem (Wang and Wesche, 2016). Grassland degradation is a process of retrograde succession of grassland ecosystems, during which the structures, processes and functions within the system have undergone significant changes (Dong et al., 2020; Yu et al., 2022). Owing to the high altitude and unique alpine climate, the alpine ecosystem of the Qinghai-Tibet Plateau (QTP) is very sensitive and fragile, and the impact of grassland degradation is more prominent (Klein et al., 2004; Schleuss et al., 2015). This not only hinders the development of animal husbandry in the country, but also seriously affects the stability of the regional ecosystem and the functioning of the plateau ecological barrier (Harris, 2010; Liu et al., 2021). Recently, the Chinese government has implemented numerous ecological protection projects (Xu et al., 2006; Quan et al., 2011; Zhao et al., 2022). Dealing with the environmental problems of grassland degradation, restoring and improving functions, and connecting ecological protection projects, such as the construction of ecological security barriers, is of great significance to the sustainable development of the region and the construction of ecological security barriers in the future.
Grassland degradation is often accompanied by an increase in the number of rodents, and a large number of rodents aggravate the process of grassland degradation (Miehe et al., 2008; Li et al., 2014). Studying the mechanisms of rodent populations during grassland degradation is of great significance for rodent control and the optimal management of grasslands (Wei et al., 2020). Changes in environmental conditions play an important role in population fluctuations and can have important effects on population numbers either directly or through their interactions (Harris et al., 2015). After grassland degradation, the original environment underwent significant change. Grassland height and coverage have decreased, vegetation community structure and dominant species have changed, aboveground and belowground biomass has decreased, soil bulk density and compactness have increased, and soil water content has decreased (Harris, 2010; Wang et al., 2014). Relevant studies have shown that degraded grassland provides a good open habitat for rodents who prefer to live in these conditions, which is conducive to observing natural enemies from a distance and reducing the risk of predation (Liu et al., 2013). As the main body of rodents builds burrows, soil properties are important factors affecting rodent habitat selection. Soil compaction determines the firmness of the rodent tunnel system and affects the energy consumption in the process of building the tunnel system (Zhang, 2007). Changes in vegetation community structure and dominant species directly affect animal food resources (Tang et al., 2015).
The abundance and quality of food directly affect the population density and distribution areas of animals in the environment (Chua et al., 2021). Optimal foraging theory indicates that animals usually tend to choose food resources of the highest nutritional quality and concurrently modify their foraging strategies in terms of energy and time. They tend to minimize food handling time, reduce exposure to predation threats while decreasing their energy expenditure, and maximize their own nutritional and energy gain (Forbey et al., 2018; Abrahms et al., 2021; Demi et al., 2021). Therefore, when food resources are abundant, animals only select the best dietary items. Under food shortages, generalist herbivores can respond by expanding their dietary selection to low-ranked food types, regardless of whether forage is preferred (Li et al., 2019; Goldberg et al., 2020). However, food types of different qualities are spatially distributed in a mosaic pattern. High-quality food resources often exist on a small scale and are often surrounded by large-scale, low-quality food resources (Hope et al., 2021). The primary vegetation of alpine meadows on the QTP is dominated by grasses and sedges; however, as grassland degradation advances, the proportion of forbs significantly increases (Zhang et al., 2018). In the case of severely degraded (SD) grasslands, a plant community with forbs as the dominant species is often formed, and the soil surface becomes bare and loosely compacted (Wang et al., 2020). Optimal foraging theory also assumes that animals need to spend energy in the search for food, and the difference between the energy gained from food consumption and the energy expended during foraging is the net income of energy gain. The greater the income, the more beneficial the food will be to foragers; therefore, animals prefer to forage in patches of high-quality food (Schai-Braun et al., 2020). The underground niche may impose some constraints on the food selection process of subterranean species, and their foraging mainly relies on digging tunnels, which necessitates extra energy expenditure compared with aboveground animals (Su et al., 2018). For example, the amount of energy an animal uses to move a unit distance underground by digging is 360–3400 times greater than that of moving aboveground (Xie et al., 2013).
The plateau zokor (Eospalax baileyi), a small mammal in the alpine meadow, is a typical subterranean rodent endemic to the QTP (Su et al., 2015). As a key species in the alpine meadow ecosystem of the QTP, it performs several vital ecological functions. In natural habitats, digging activities can promote soil renewal, which is helpful for the circulation of soil nutrients and germination of seeds and underground buds. In addition, the underground activities of plateau zokors may be beneficial for the colonization of exotic plants in alpine meadow ecosystems. Therefore, it deserves the reputation of “Ecosystem engineer” (Zhang et al., 2010; Eldridge et al., 2016; Miranda and Rothen, 2019). However, when the population density of zokors is high, a series of activities, such as digging and eating plant roots, seriously damage plants and thus affect grassland ecosystems. Burrowing activity causes grassland vegetation to be covered by mounds of soil for a long time, changes the structure of the plant community, reduces the productivity of the grassland, and further aggravates its degradation (Li et al., 2021). In practice, it has been found that the population density of the plateau zokor increases with the aggravation of grassland degradation (Wang et al., 2000a). However, the factors that lead to the increase in the plateau zokor population during grassland degradation remain unclear. The change in the food resources of the plateau zokor under grassland degradation might be a key factor. The adequacy of food resources and environmental safety are priorities for animal habitat selection (Ṧumbera et al., 2012). For subterranean rodents, security in underground tunnels is guaranteed because of the absence of predation risks. Therefore, food resources may be regarded as crucial factors for subterranean dwellers. High-quality food and loose soil structures are important for the reproduction and development of subterranean rodent populations. Thus, we speculate that degraded grassland provides an optimum foraging environment for plateau zokors (the cost of foraging is reduced after grassland degradation) and a reliable food source.
Our working hypothesis was that the total biomass of plants decreases after grassland degradation, but the proportion of food that the plateau zokor prefers increases, and the cost of obtaining food decreases, making it more suitable for foraging. This might be the main reason for the increase in the population density of plateau zokors in degraded grasslands. In this study, soil bulk density, compactness, and water content were used to indicate the difficulty of food acquisition. Food composition and proportion were used as indicators to analyze whether food changes and how diversity changes. Finally, the stomach nutrients contents were analyzed. The aim of this study was to determine how grassland degradation affects the dietary selection of plateau zokors and to provide a new perspective on the mechanism of rodent damage to the grassland ecosystem.
The experimental area was located in the Luqu County, Gansu Province, QTP (34°14′07″–34°48′48″N, 102°10′80″–102°58′15″E), with an average altitude of 3550 m. The annual average temperature was 2.6 °C, and the annual frost-free period was 56 days. The annual precipitation ranges from 632 to 781 mm, mainly from May to September. The soil was mainly subalpine and alpine meadow soils. Alpine shrub meadows and alpine meadows accounted for 88.68% and 11.32% of the county’s grassland area, respectively. The alpine meadow is one of the main grazing grasslands in the region, and is also the area with the most severe prairie rat infestation. The area of rodent infestation in the county is 14.18 × 104 hm2, accounting for 33.84% of the county’s grasslands. The plateau zokor is one of the dominant groups of rodents in the Luqu County, accounting for ∼13.58% of rodent species (Wu et al., 2020).
According to the Grading Standard of Natural Grassland Degradation, Desertification and Salinization (GB19377-2003) (Su et al., 2003), vegetation coverage was selected as a classification index of degraded grassland, and the vegetation coverage of each type of degraded grassland was as follows: lightly degraded (LD), 80–100%; moderately degraded (MD), 60–80%; severely degraded (SD), < 60%. The degree of grassland degradation in the study area, LD, MD, and SD grasslands were scored and selected. Another reseeded grassland (RG) was selected (Figure 1 and Table 1). The reseeding was Elymus nutans + Poa pratensis, and the reseeded period was 4 years.
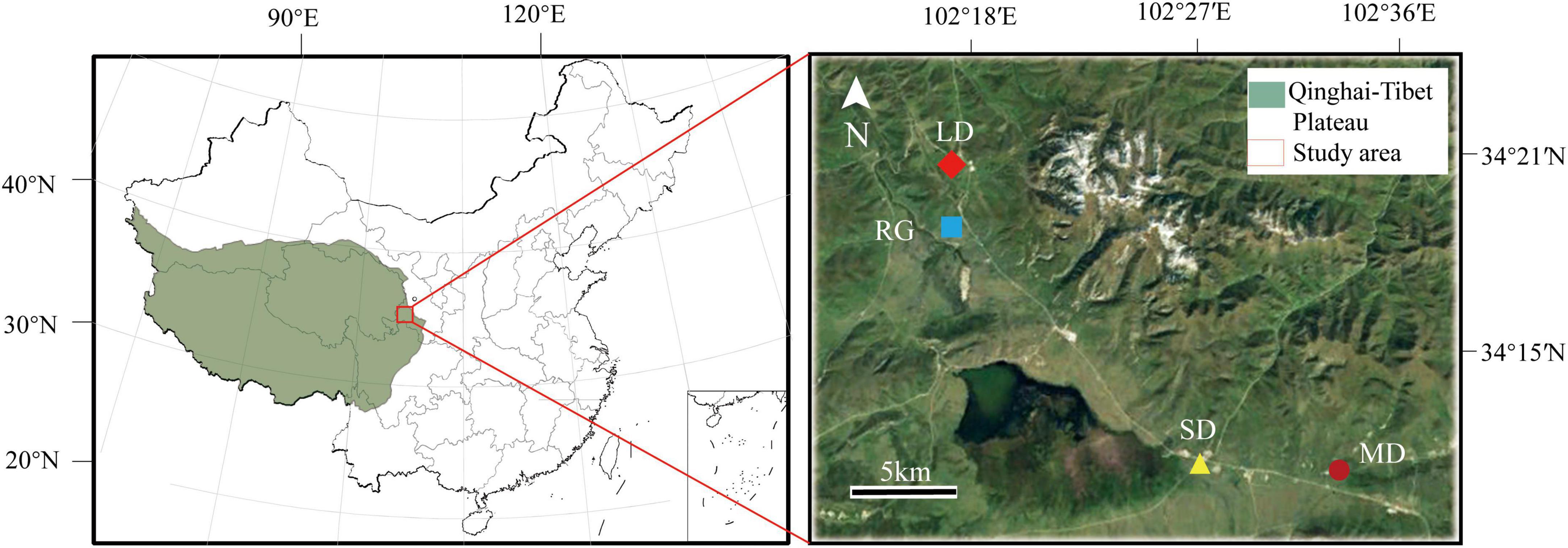
Figure 1. Location of the sampling site. LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland.

Table 1. Study area.
Five (0.5 m × 0.5 m) quadrats were randomly selected from different plots to investigate the density, coverage, and height of each plant. All plants in the square were collected and classified, brought back to the laboratory for drying at 95°C, and weighed separately for the aboveground and underground parts to obtain the corresponding biomasses of each species. The coverage, density, height, and biomass of each species in the plant community were standardized using the maximum-standardization method.
Species importance value = (relative coverage + relative density + relative height of species + relative biomass)/4. The importance value was used to calculate the diversity (Shannon-Wiener index) of the plant community in the quadrat (Niu et al., 2019).
The moisture content and bulk density were determined by the aluminum box drying method, which was repeated three times. Soil compaction at a depth of 0–30 cm was measured using a soil compaction meter (SC-900), recorded every 2.5 cm, and repeated 15 times.
In the study area, 122 plateau zokors were captured using bow traps, including 25 individuals in RG, 24 individuals in LD, 38 individuals in MD, and 35 individuals in SD. The population density of the plateau zokor per unit area was calculated based on the specific plot area of the captured plateau zokor (Table 1). Ten adult individuals were selected from each grassland type (LD: 4 males and 6 females; MD: 5 males and 5 females; SD: 5 males and 5 females; RG: 3 males and 7 females), euthanized by decapitation, and their gastric contents were collected. Forty samples were used for food habit analysis. Institutional guidelines for the use of animals were followed, the Animal Ethics Committee of Gansu Agricultural University approved the experimental procedure (approval no. GAU-LC-2020-014), and local authorities’ approval was also obtained.
Referring to GB/T14924.9-2001 (Zhou et al., 2001), the semi-micro Kjeldahl method was used to determine the crude protein (CP) content, the crude fat was determined by the Soxhlet extraction method, and Van’s fiber method was used to determine the acid detergent fiber (ADF) and neutral detergent fiber (NDF) contents.
Genetic analysis of 40 samples was performed using Genepioneer Biotechnologies (Nanjing, China), including DNA extraction, ITS2 gene amplification, and sequencing of the amplified products on an Illumina MiSeq platform.
The QIIME2 software was used for species annotation with default parameters, and the RDP3 Classifier v. 2.2 algorithm were used to identify the taxonomic composition (Edgar, 2004). The diversity analyses of the diet were conducted according to output-normalized sequence data. The significance analysis in this study was carried out using one-way ANOVA in SPSS 23.0 (IBM®, New York, United States). The R version 2.6-2 (The R Foundation for Statistical Computing, Vienna, Austria) “vegan” packages was used for non-metric multidimensional scaling (NMDS) analysis (Oksanen et al., 2018), and the Amos Graphics 24.0 (IBM®, New York, United States) software was used to draw structural equations (Grace, 2006). The level of significance at which the null hypothesis was rejected was set at α = 0.05.
The survey found that there were 18 families, 37 genera, and 41 plant species in the RG; 16 families, 33 genera, and 38 plant species in LD; 15 families, 30 genera, and 31 species in MD; and 11 families, 16 genera, and 16 species in SD. The aboveground and underground biomasses of RG were the highest, and the aboveground and underground biomasses decreased with increasing degree of grassland degradation (Figure 2A). The aboveground biomass of different functional groups and underground biomass of grasses and sedges decreased with an increase in the degree of degradation. The relative biomass of forbs under RG was significantly lower than that of other grassland varieties (P < 0.05), the relative biomass of forbs under SD was the highest, and the relative biomass of grasses under SD was significantly lower than that of other grassland types (Figure 2B). The biomass proportions of the top 15 genera were 96.46% in RG, 86.59% in LD, 88.35% in MD, and 93.77% in SD. The top four genera of relative biomass in RG were Elymus (29.63%), Poa (17.82%), Scirpus (16.15%), and Kobresia (10.62%); Elymus (27.86%), Kobresia (23.08%), Poa (10.92%), and Potentilla (7.51%) in LD; Elymus (24.07%), Kobresia (20.20%), Scirpus (12.79%), and Potentilla (9.13%) in MD; and Potentilla (36.52%), Elymus (30.54%), Poa (12.12%), and Astragalus (6.32%) in SD (Figure 3).
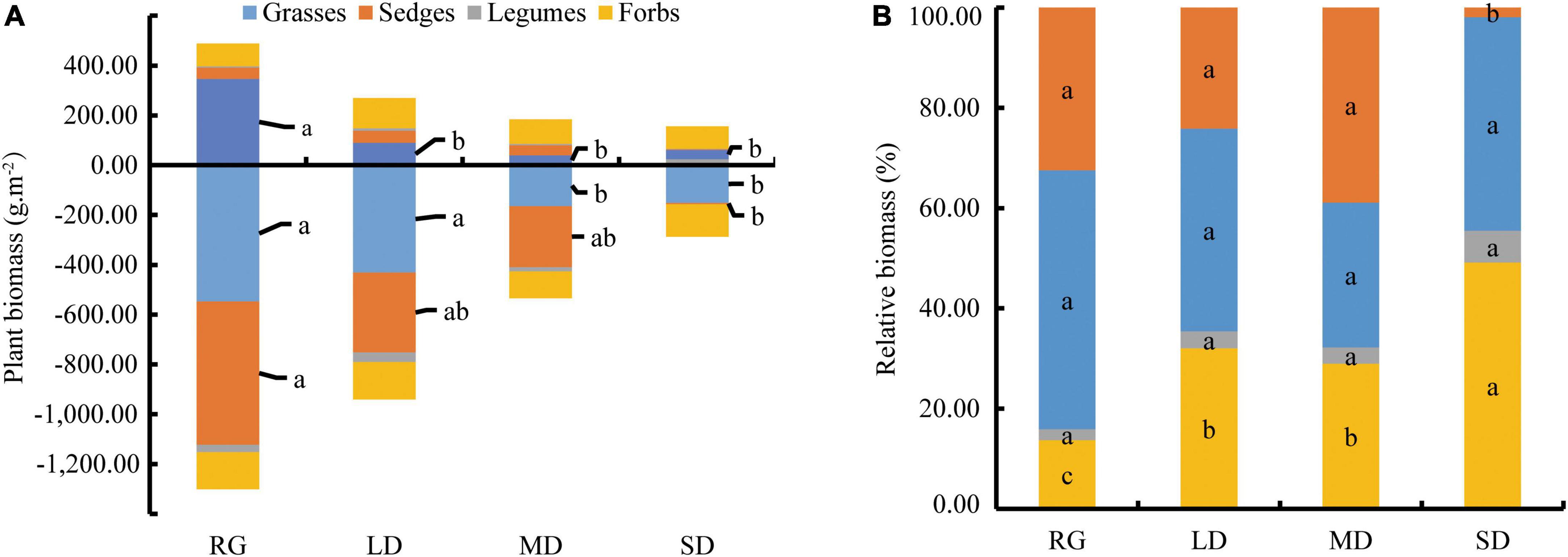
Figure 2. Biomass (A) and relative biomass (B) of different functional groups. LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland. Different letters indicate significant differences between groups (P < 0.05).
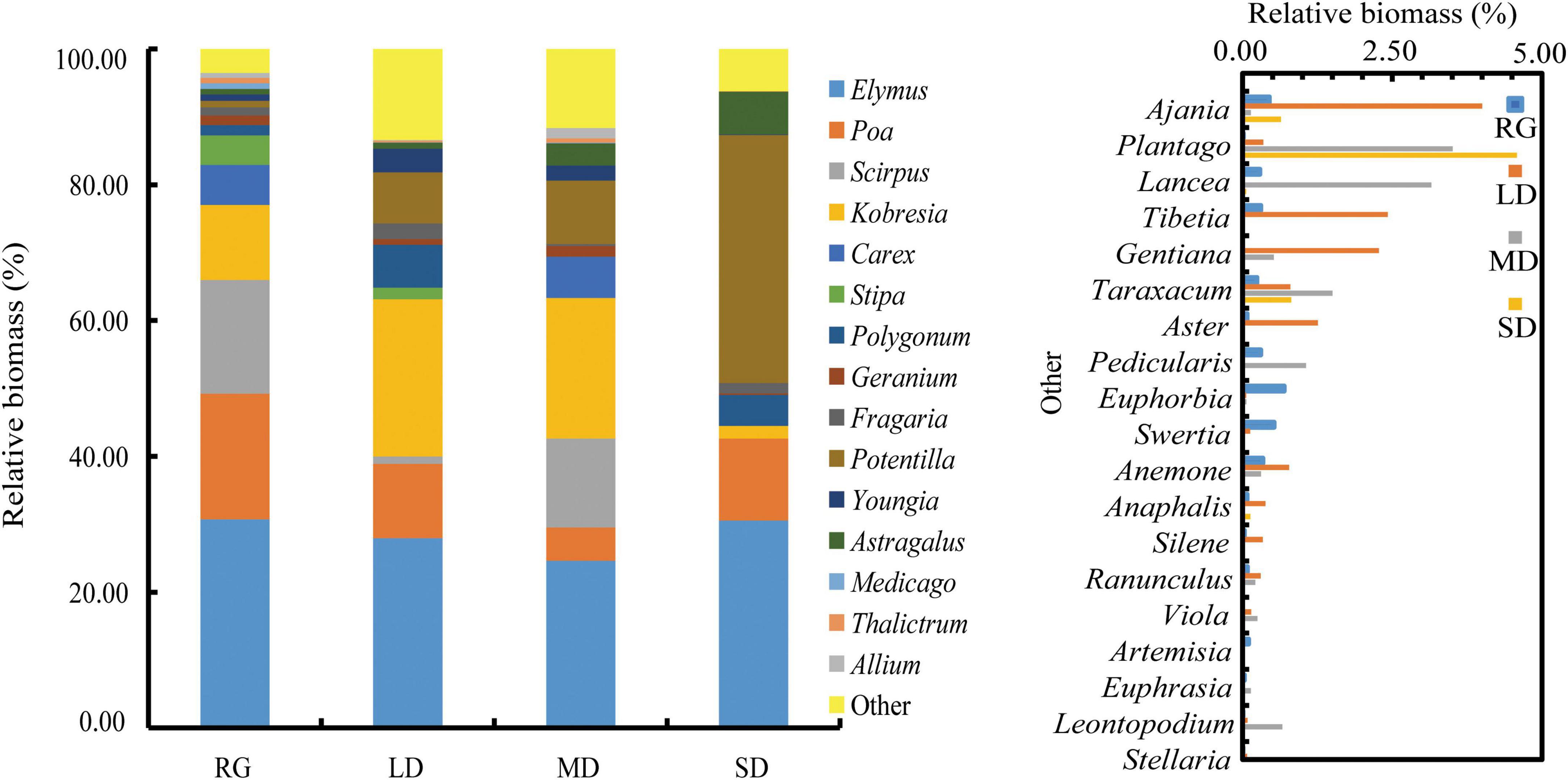
Figure 3. Relative biomass of plants at the genus level. LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland.
The soil water content at LD within 0–10 cm depth was significantly higher than that of other grasslands (P < 0.05), whereas the soil moisture at SD at 10–20 cm and 20–30 cm depths was the lowest (Figure 4A). There were no significant differences in the soil bulk density at different depths among the different grassland conditions. Under SD, soil bulk densities at depths of 0–10 cm and 20–30 cm were the lowest, and soil bulk density at depths of 10–20 cm increased with the deterioration of grassland conditions (Figure 4B). The soil compaction degree of the grassland under different conditions was significantly different at 0–10 cm depth (P < 0.05), and the soil compaction degree at 0–10 cm of RG was significantly smaller than those of MD and SD. Soil compaction was the smallest in RG at 10–20 cm depth and the largest in MD at 20–30 cm depth, LD had the smallest soil compaction, and the largest was recorded in SD (Figure 4C). The soil compaction degree under different grassland conditions increased gradually at 2.5–7.5 cm, decreased at 7.5–12.5 cm, and showed an increased gradual trend at 12.5–25 cm (Figure 5).
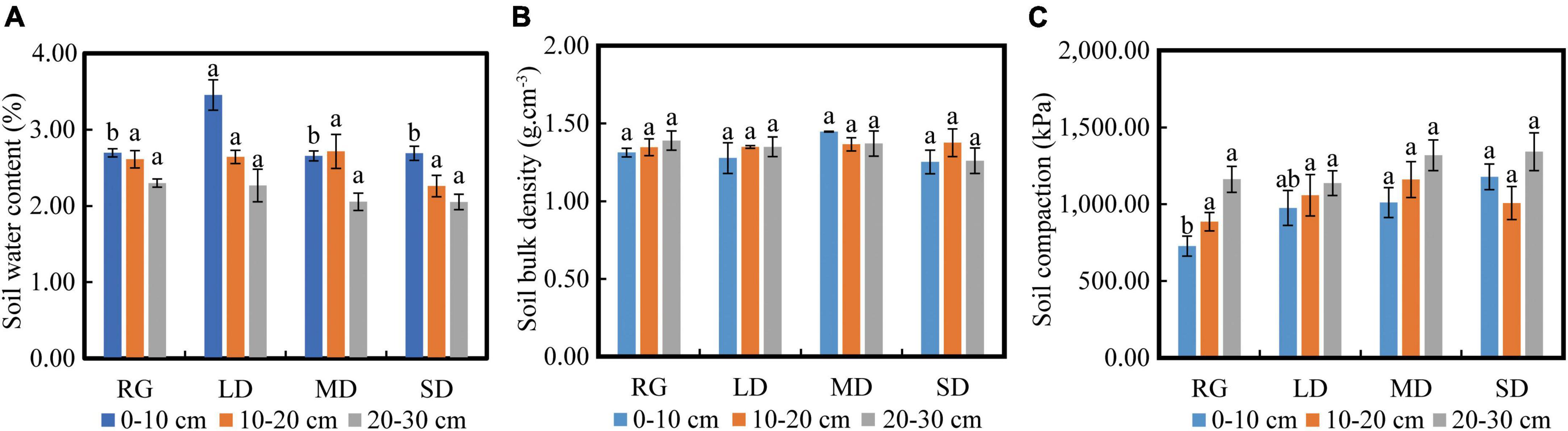
Figure 4. Changes in soil water content (A), soil bulk density (B), and soil compaction (C) of different grasslands. LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland. Different letters indicate significant differences between groups (P < 0.05).
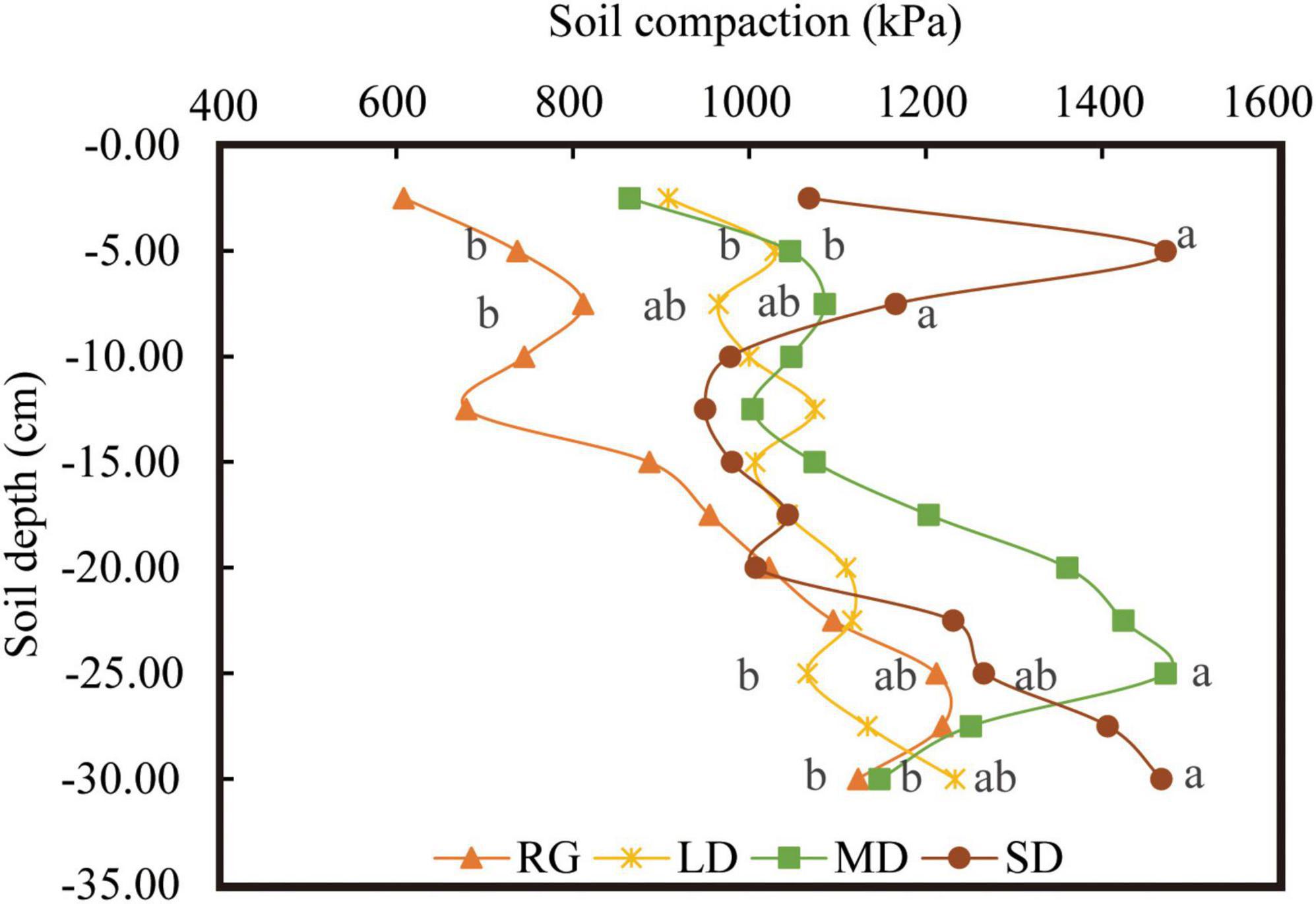
Figure 5. Change in soil compactness between different soil layers. LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland. Different letters indicate significant differences between groups (P < 0.05).
The plateau zokor fed on 21 families and 30 genera of plants in RG, 18 families and 25 genera in LD, 22 families and 35 genera in MD, and 19 families and 30 genera in SD. The plant species (genus level) eaten by the plateau zokor in various grasslands were roughly the same. The proportions of the top 15 genera in their diets were 99.53% in RG, 99.61% in LD, 97.92% in MD, and 97.51% in SD. Potentilla had a large proportion, accounting for 40.57 % in RG, 72.96 in LD, 77.17% in MD, and 86.13% in SD. In addition, other genera with a higher proportion in the diet of plateau zokors in RG were Geranium (22.07%), Taraxacum (8.68%), and Tibetia (7.12%); in LD were Geranium (7.56%), Lancea (4.94%), and Taraxacum (4.49%); in MD were Youngia (10.29%), Tibetia (4.36%), and Taraxacum (3.27%); and in SD were Stellaria (6.85%), Youngia (1.65%), and Lepidium (1.53%) (Figure 6B). According to the statistics of plant functional groups, forbs under the four different grassland types were the main diet of plateau zokors, and their proportions were more than 90% (Figure 6A). The food diversity of the plateau zokor in different types of grasslands was significantly different (P < 0.05). The richness and Shannon-Wiener index of RG were significantly greater than those of LD and SD, and the Shannon-Wiener index of SD was significantly lower than that of LD (P < 0.05) (Figures 7B,C). The evenness index of RG was significantly larger than those of MD and SD (P < 0.05) (Figure 7D). In addition, the food niche breadth of RG was significantly larger than that of other grasslands (Figure 7E). The NMDS results show that the food composition of plateau zokors under different grassland conditions was roughly divided into two groups: the food compositions of RG and LD were similar, and those for MD and SD grasslands were similar (Figure 7A).
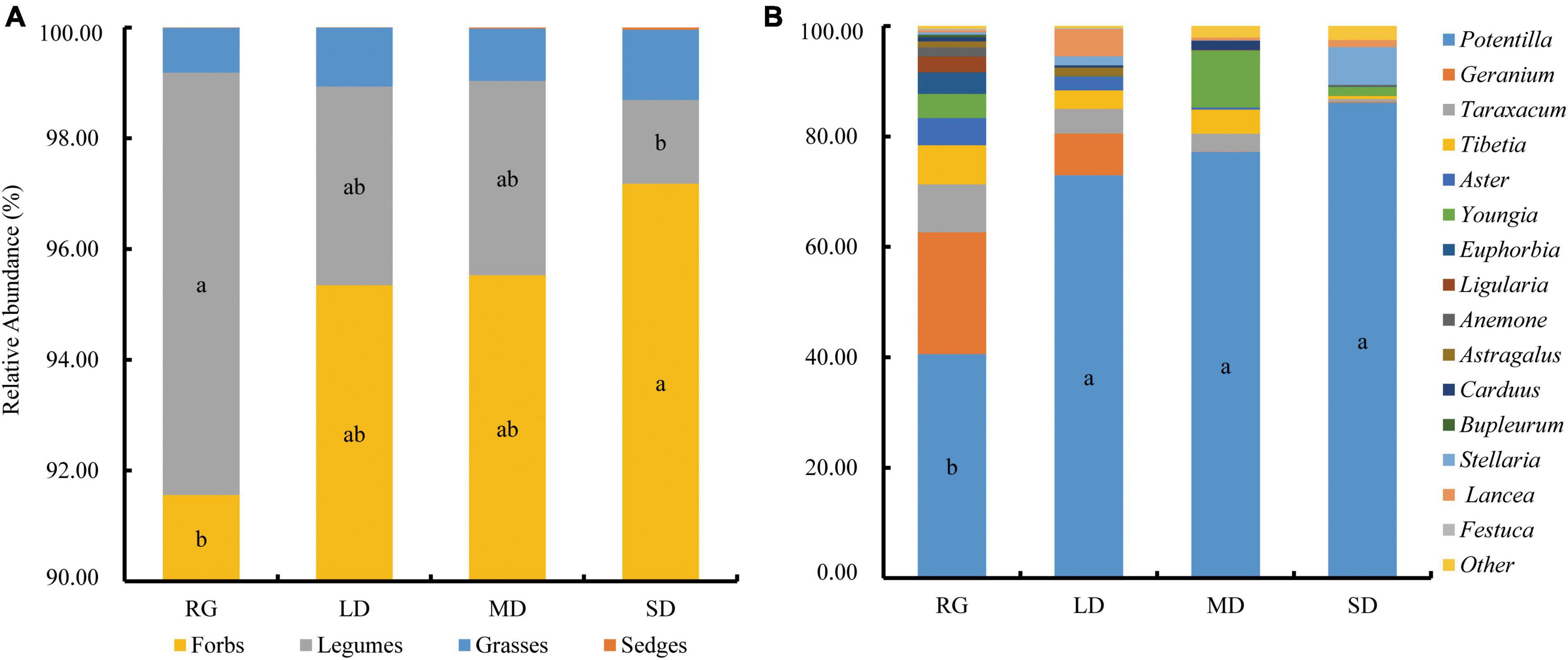
Figure 6. Food composition and proportion of plateau zokors with different functional groups (A) and plants at the genus level (B). LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland. Different letters indicate significant differences between groups (P < 0.05).
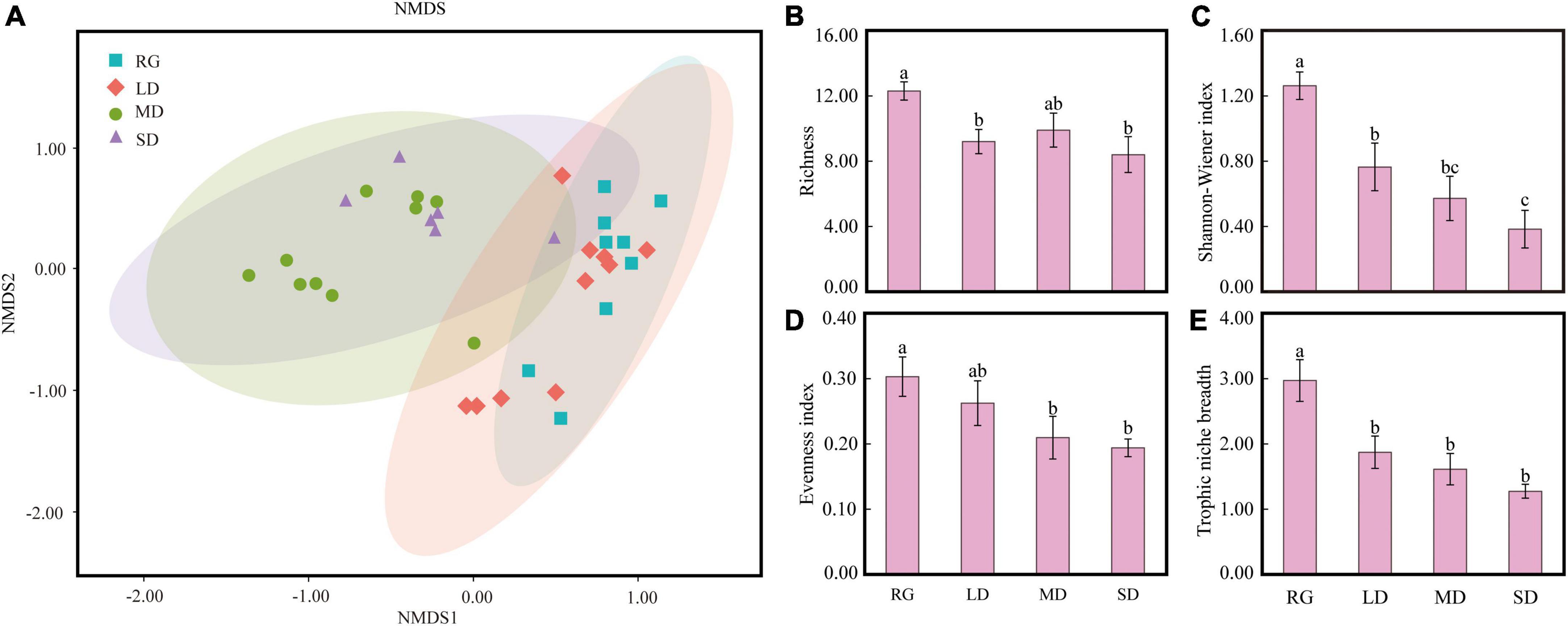
Figure 7. NMDS analysis of food structure of plateau zokors (A), food richness (B), Shannon-Wiener index (C), evenness index (D), and food niche width (E). LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland. Different letters indicate significant differences between groups (P < 0.05).
The ADF content was the highest in RG and decreased with an increase in grassland degradation degree, which was significantly the lowest in SD (P < 0.05). NDF did not show any significant differences among the plots. Crude fat was the highest in RG (P < 0.05) and the lowest in MD (P < 0.05). The CP contents of MD and LD were significantly higher than those of RG and SD (P < 0.05) (Figure 8).
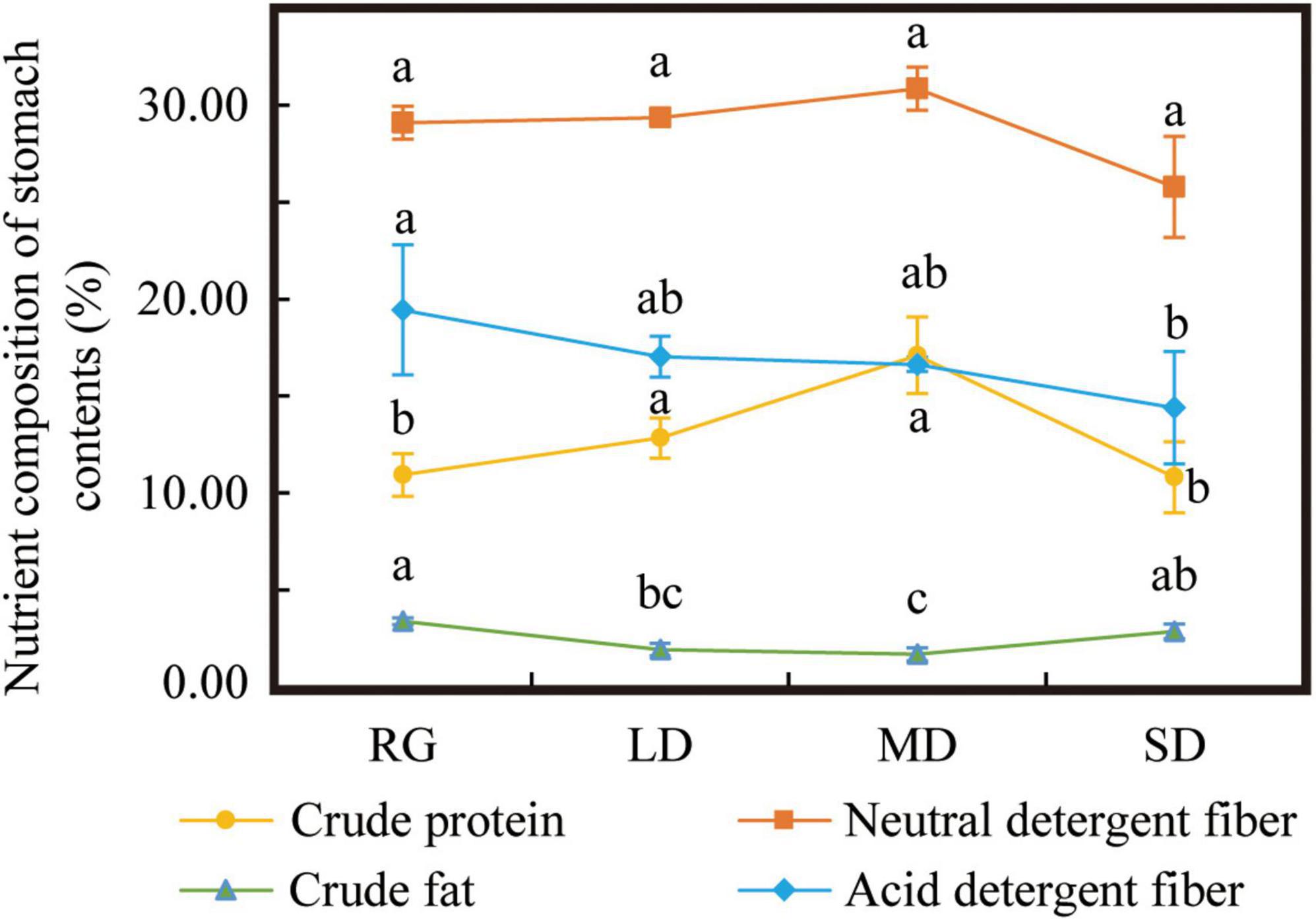
Figure 8. Nutrient composition of stomach contents. LD, lightly degraded grassland; MD, moderately degraded grassland; SD, severely degraded grassland; RG, reseeded grassland. Different letters indicate significant differences between groups (P < 0.05).
During grassland degradation, soil physical properties had a significant impact on the population density of the plateau zokor and the relative biomass of forbs (P < 0.05), and the direct impact contribution rates were 0.20 and 0.30, respectively. Plant diversity had an extremely significant negative impact on the nutrient composition of gastric contents (P < 0.001), and the contribution rate of the direct impact was 0.48. The relative biomass of forbs had an extremely significant negative correlation with feeding habits (P < 0.001) with a direct impact contribution rate of 0.56, and an extremely significant impact on the population density of plateau zokors (P < 0.001) with a direct impact contribution rate of 0.63 (Figure 9).
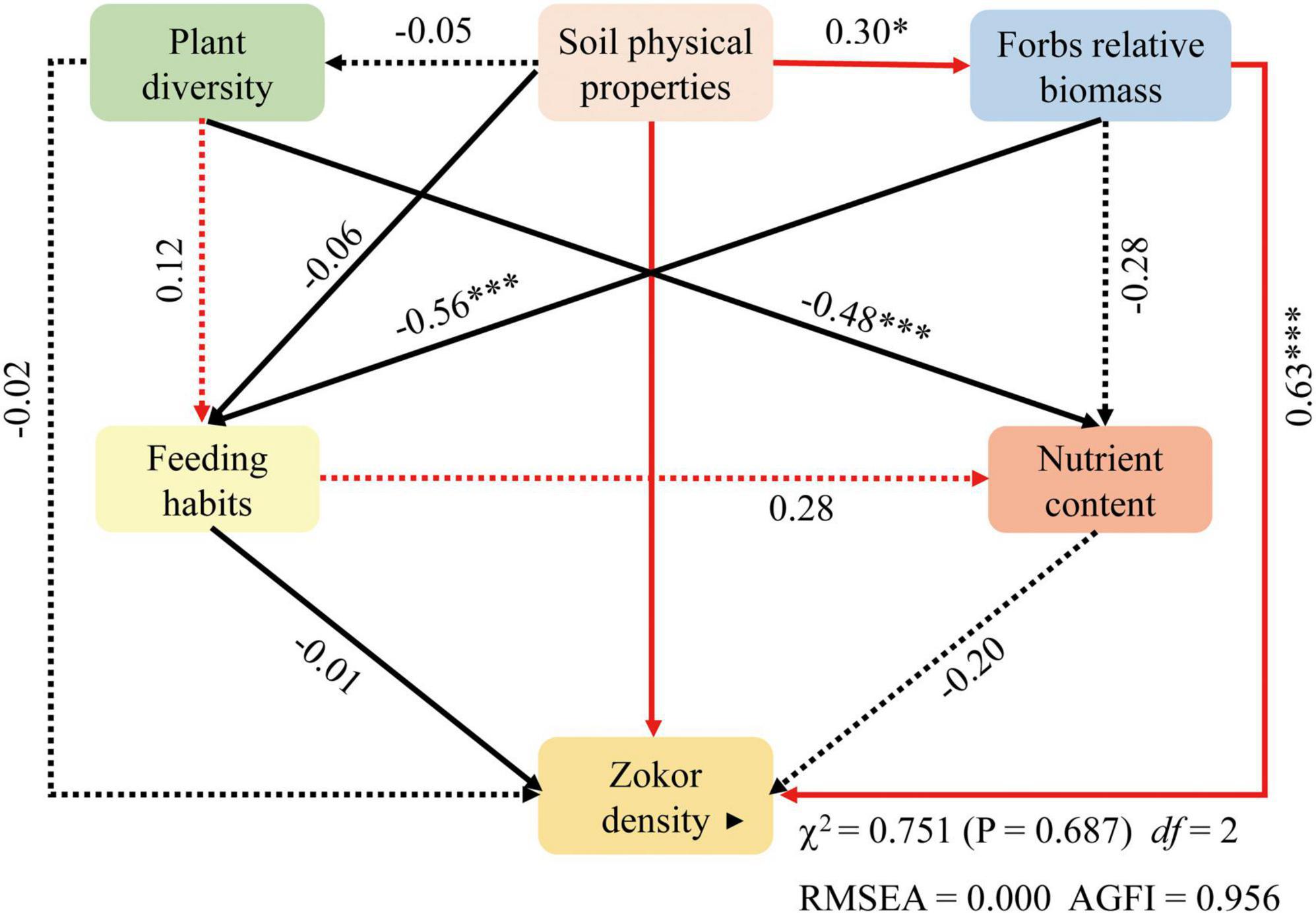
Figure 9. Structural equation model between plateau zokor density and soil physical properties, plant resources, and feeding habits. Dotted line indicates that the effect is insignificant and the black line indicates a negative correlation. *P < 0.05, ***P < 0.001.
In this study, 24 families and 48 genera were identified in all stomachs, whereas only 21 families and 45 genera were found in the research area. Our results show that the ITS2 barcode combined with Illumina MiSeq sequencing can effectively identify animal feeding habits. Among the four types of grasslands, forbs were the main food source for plateau zokors. Plateau zokors have a clear preference for Potentilla plants, which occupy a considerable proportion of their diet. This result is in agreement with that of Wang et al. (2000a). Plateau zokors dwell in special underground circumstances that require higher energy expenditures for excavation and mound building during foraging (Hu et al., 2017; Lopes et al., 2020). Therefore, forbs with a relatively large biomass were selected as food. Generally, the lower parts of the forbs are enlarged and their nutrient content is more abundant. The distribution of the biomass of forbs determines the population density of plateau zokors in different regions to a certain extent (Zhang et al., 2010, 2022). Plateau zokors at different degradation levels and RG feed on the same species of plants but with different proportions. Animals can adjust their food intake ratios according to changes in food resources and nutritional needs (Egert-Berg et al., 2021; Guo et al., 2021). As the degree of grassland degradation increased, the feed intake of the plateau zokors of forbs increased. The proportion of forbs of plateau zokors under RG was the smallest, which increased the amount of leguminous plant intake to meet their dietary needs.
In the present study, plateau zokors preferred to inhabit degraded grasslands, and the population density of plateau zokors increased with the severity of degradation. Habitat selection is the most important step for animals to survive and reproduce successfully, and the adequacy of food resources is a crucial factor in habitat selection (Ṧumbera et al., 2012). The foraging activities of the plateau zokor were not random but highly selective. Although the diet of plateau zokors contains numerous plant species, Potentilla accounted for a large proportion of their dietary choices. Owing to the high energetic cost of burrowing, non-directional underground search patterns, and low food availability, plateau zokors need to consider the benefits and costs of foraging (Xie et al., 2014), but diets of different qualities are distributed in a mosaic model (Hope et al., 2021). The dominant species of RG and LD grassland were mainly grasses and sedges, whereas the forbs favored by plateau zokors were relatively small and embedded within. Among them, forb rhizomes are surrounded by tightly connected rhizomes of grasses and sedges. With the aggravation of grassland degradation, the proportion of grasses and sedges decreased significantly, whereas the proportion of forbs increased. Especially in the SD grassland, the vegetation cover was mainly forbs, and the forbs are the primary dietary selection of plateau zokors, which qualifies the degraded grassland as a suitable habitat for these animals. The nutrient composition of the stomach contents of plateau zokors in different grasslands was analyzed, and it was found that the contents of acidic and neutral detergent fibers in the degraded grassland were lower, and the content of CP was higher, in light and moderately degraded grasslands. Proteins play a key role in growth and metabolism. Too low a protein content in the diet seriously deteriorates the fitness of an individual, leading to weight loss and immune impairment (Kohl et al., 2016). Nutrients in the food of the plateau zokor under the degraded grassland can better ensure and maintain the normal physiological processes of the animal. Plateau zokors live underground and rely on digging tunnels for daily foraging. Soil hardness not only affects the firmness of rodent tunnels but is also directly linked to the level of energy expenditure during digging, both for foraging and tunnel building. In this study, we found that the compactness of 10–25 cm in SD grassland was less than that in moderately degraded grassland, and the tunnels of the plateau zokor were concentrated at 10–20 cm underground. Therefore, zokors may consume less energy during digging in SD grasslands than in moderately degraded ones. Although the compactness of the RG and LD grassland is low, which is favorable for excavation, the population density of plateau zokor is low. Therefore, we may assume that the relative proportion of high-quality food is the main factor that adjusts the population density of plateau zokors, and that low soil compactness may be an auxiliary factor for the regulation of the plateau zokor population when food is plentiful. After grassland degradation, the relative proportion of forbs preferred by the plateau zokor increased with the low digging costs required for foraging. Therefore, foraging efficiency is improved, and more protein is consumed as the protein content in the stomach is higher. This might help explain, from a dietary point of view, why the population density of the zokors increases after grassland degradation.
In this study, compared with degraded grasslands, the population density of plateau zokors in reseeding grasslands was the lowest. Habitat selection in rodents is imperative for their optimal survival and fitness, and changes in vegetation communities often have an impact on rodent populations in grasslands (Fox and Monamy, 2007).
Additional explanations and mechanisms, such as food resource theory (Ostoja and Schupp, 2009) and movement obstruction theory (Gitzen et al., 2001; Rieder et al., 2010), might help unravel increasing zokor population density with advanced grassland degradation. Plateau zokors live underground, and soil hardness determines the energy expenditure of subterranean rodents when they forage and build burrows. Wang et al. (2000b) found that there is a significant negative correlation between the population density of plateau zokors and soil hardness, and the zokor prefers to live in areas with less soil hardness. In the present study, soil compaction under the RG tended to develop in the direction of the habitat preferred by the plateau zokor. A reasonable explanation is that when the soil compactness is too low, the tunnels constructed by the plateau zokor may collapse readily, and it is difficult to maintain the original state. Therefore, plateau zokors need to re-pair or reconstruct the tunnel, which requires extra time and energy (Zhang, 2007).
Changes in the plant community structure also alter the original food resources (Ostoja and Schupp, 2009). In this study, the vegetation composition of the grassland in the original habitat of the plateau zone changed significantly after the grassland was reseeded. Vegetation coverage and height increased, and the reseeded grass species, Elymus nutans and Poa pratensis, became the dominant species. In general, the proportion of grasses and sedges increased, whereas the proportion of forbs favored by plateau zokors decreased, resulting in food availability reduction and food shortage for the animals. Therefore, the plateau zokor increased the food intake of legume plants in the RG, its food diversity and evenness increased, and the vegetative niche breadth was significantly widened. Owing to the characteristics of subterranean life, foraging underground adopts different mechanisms compared to aboveground species, and its normal foraging activities need to rely on tunnels. However, the dominant groups of grasses and sedges in the RG have developed underground rhizomes and are closely connected, and the favorite forbs are often embedded in the sedges and grasses, which greatly increases the foraging difficulty and time of plateau zokors. Colloquially, the decrease in population density of plateau zokors in RG is closely linked to the reduction of preferred food availability and increasing foraging difficulty, which qualifies the grassland replantation strategy as an optimum policy to, on the one hand, decrease the population density, and, on the other hand, to combat grassland degradation.
Briefly, grassland degradation aggravates the occurrence of rodent pests, the aggravation of rodent pests in degraded grasslands should be regarded as the result of the comprehensive effect of a series of factors such as food resources, soil characteristics, intraspecies relationship, and aboveground environment. The present study focused on two factors, food resources and soil characteristics. Subterranean mammals live mainly in open habitats, and their evolution is tightly connected with the emergence of seasonal habitats and prolonged droughts (Nevo, 1999). In these habitats, plants storing energetic reserves in bulbs, roots, and rhizomes have evolved, and these parts of plants are staple foods for subterranean mammals (Nevo, 1999; Busch et al., 2000); in addition, plants in arid regions allocate more underground biomass, providing them with more food resources. Nevertheless, soil water content needs to be kept within a reasonable threshold, because the extremely dry or humid habitat cannot form the tunnel for subterranean mammals to inhabit, and the underground biomass will also be greatly reduced, resulting in a shortage of food resources. Therefore, soil properties have always been an essential factor affecting the distribution of subterranean mammals, and the plateau zokor is no exception.
This study showed that plateau zokors prefer to inhabit degraded grasslands, and the population of plateau zokors increased with grassland deterioration. The mechanisms underlying this phenomenon were explored from the perspective of feeding habits. ITS2 barcoding combined with Illumina MiSeq sequencing was used to analyze the feeding habits of different degrees of degraded and reseeded grasslands. Forbs were found to be the main food source of plateau zokors, and Potentilla occupies a considerable proportion of its diet. After grassland degradation, the relative proportion of forbs that plateau zokors preferred increased, the feeding cost reduced, the feeding efficiency improved, and the content of CP in the stomach contents increased. The foraging difficulties for plateau zokors to obtain the plants they preferred significantly increased after the grassland was reseeded. Reduced food availability has led to a decline in the population density of plateau zokors. Affecting rodent eating habits through reseeding is an effective way to prevent and control rodent ecology. In the future, it is necessary to perform monitoring trial to observe the long-term effects of grassland degradation and reseeding on plateau zokor population.
The data presented in this study are deposited in the NCBI repository, bioProject ID: PRJNA867532, accession numbers: SAMN30207155–SAMN30207194.
The animal study was reviewed and approved by Institutional guidelines for the use of animals were followed, the Animal Ethics Committee of Gansu Agricultural University approved the experimental procedure (approval no. GAU-LC-2020-014), and local authorities’ approval was also obtained.
KA, QP, and ZW were responsible for the majority of data collection. JS guided the primary writing. YK, XS, and BY assisted with revisions of the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (31760706), the Program for Longyuan Youth Innovation Talents of Gansu Province of China (LYRC2019-5), Science and Technology Support Project of Forestry and Grassland Bureau of Gansu Province (LCJ2021020), and “Fuxi Talent” Plan of Gansu Agricultural University (Gaufx-02J03).
We thank Tan Y and Hou Q for assistance in conducting experiments.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abrahms, B., Aikens, E. O., Armstrong, J. B., Deacy, W. W., Kauffman, M. J., and Merkle, J. A. (2021). Emerging perspectives on resource tracking and animal movement ecology. Trends. Ecol. Evo. 36, 308–320. doi: 10.1016/j.tree.2020.10.018
Busch, C., Antinuchi, C. D., del Valle, J. C., Kittlein, M. J., Malizia, A. I., Vassallo, A. I., et al. (2000). “Population ecology of subterranean rodents,” in Life Underground: The Biology of Subterranean Rodents, eds E. A. Lacey, J. L. Patton, and G. N. Cameron (Chicago: University Chicago Press), 183–226.
Chua, P., Lammers, Y., Menoni, E., Ekrem, T., Bohmann, K., Boessenkool, S., et al. (2021). Molecular dietary analyses of western capercaillies (Tetrao urogallus) reveal a diverse diet. Environ. DNA 3, 1156–1171. doi: 10.1002/edn3.237
Demi, L. M., Taylor, B. W., Reading, B. J., Tordoff, M. G., and Dunn, R. R. (2021). Understanding the evolution of nutritive taste in animals: Insights from biological stoichiometry and nutritional geometry. Ecol. Evol. 11, 8441–8455. doi: 10.1002/ece3.7745
Dong, S., Zhang, J., Li, Y., Liu, S., Dong, Q., and Zhou, H. (2020). Effect of grassland degradation on aggregate-associated soil organic carbon of alpine grassland ecosystems in Qinghai-Tibetan Plateau. Eur. J. Soil Sci. 71, 69–79. doi: 10.1111/ejss.12835
Edgar, R. C. (2004). MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Egert-Berg, K., Handel, M., Goldshtein, A., Eitan, O., Borissov, I., and Yovel, Y. (2021). Fruit bats adjust their foraging strategies to urban environments to diversify their diet. BMC Biol. 19:123. doi: 10.1016/j.cub.2018.09.064
Eldridge, D. J., Delgado-Baquerizo, M., Woodhouse, J. N., and Neilan, B. A. (2016). Mammalian engineers drive soil microbial communities and ecosystem functions across a disturbance gradient. J. Anim. Ecol. 85, 1636–1646. doi: 10.1111/1365-2656.12574
Forbey, J. S., Liu, R., Caughlin, T. T., Matocq, M. D., Vucetich, J. A., Kohl, K. D., et al. (2018). Review: Using physiologically based models to predict population responses to phytochemicals by wild vertebrate herbivores. Animal 12:s383–s398. doi: 10.1017/S1751731118002264
Fox, B. J., and Monamy, V. (2007). A review of habitat selection by the swamp rat, Rattus lutreolus (Rodentia: Muridae). Austral Ecol. 32, 837–849. doi: 10.1111/j.1442-9993.2007.01849.x
Gitzen, R. A., West, S. D., and Trim, B. E. (2001). Additional information on the distributions of small mammals at the Hanford Site, Washington. Northwest Sci. 75, 350–362. doi: 10.1046/j.1471-8278.2001.00062.x
Goldberg, A. R., Conway, C. J., Tank, D. C., Andrews, K. R., Gour, D. S., and Waits, L. P. (2020). Diet of a rare herbivore based on DNA metabarcoding of feces: Selection, seasonality, and survival. Ecol. Evol. 10, 7627–7643. doi: 10.1002/ece3.6488
Grace, J. (2006). Structural Equation Modeling And Natural Systems. Cambridge: Cambridge University Press.
Guo, N., Wu, Q., Shi, F., Niu, J., Zhang, T., Degen, A., et al. (2021). Seasonal dynamics of diet-gut microbiota interaction in adaptation of yaks to life at high altitude. NPJ Biofilms Microbiomes 7:38. doi: 10.1038/s41522-021-00207-6
Harris, R. B. (2010). Rangeland degradation on the Qinghai-Tibetan Plateau: A review of the evidence of its magnitude and causes. J. Arid Environ. 74, 1–12. doi: 10.1016/j.jaridenv.2009.06.014
Harris, R. B., Wenying, W., Smith, A. T., and Bedunah, D. J. (2015). Herbivory and competition of Tibetan steppe vegetation in winter pasture: Effects of livestock exclosure and plateau pika reduction. PLoS One 10:e0132897. doi: 10.1371/journal.pone.0132897
Hope, A. G., Gragg, S. F., Nippert, J. B., and Combe, F. (2021). Consumer roles of small mammals within fragmented native tallgrass prairie. Ecosphere 12:e03441. doi: 10.1002/ecs2.3441
Hu, L., Zi, H. B., Ade, L. J., Lerdau, M., and Wang, C. (2017). Effects of zokors (Myospalax baileyi) on plant, on abiotic and biotic soil characteristic of an alpine meadow. Ecol. Eng. 103, s95–s105. doi: 10.1016/j.ecoleng.2017.03.010
Klein, J. A., Harte, J., and Zhao, X. Q. (2004). Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecol. Lett. 7, 1170–1179. doi: 10.1111/j.1461-0248.2004.00677.x
Kohl, K. D., Stengel, A., and Dearing, M. D. (2016). Inoculation of tannin-degrading bacteria into novel hosts increases performance on tannin-rich diets. Environ. Microbiol. 18, 1720–1729. doi: 10.1111/1462-2920.12841
Li, G., Li, X., Li, J., Chen, W., Zhu, H., Zhao, J., et al. (2019). Influences of plateau zokor burrowing on soil erosion and nutrient loss in alpine meadows in the Yellow river source zone of west China. Water 11:2258. doi: 10.3390/w11112258
Li, G., Shi, C., Song, Y., Chu, H., and Zhang, Z. (2021). The role transition of dietary species richness in modulating the gut microbial assembly and postweaning performance of a generalist herbivore. mSystems 6:e0097921. doi: 10.1128/mSystems.00979-21
Li, X. L., Perry, G., Brierley, G., Sun, H. Q., Li, C. H., and Lu, G. X. (2014). Quantitative assessment of degradation classifications for degraded alpine meadows (Heitutan), Sanjiangyuan, Western China. Land Degrad. Dev. 25, 417–427. doi: 10.1002/ldr.2154
Liu, J., Xin, Z., Huang, Y., and Yu, J. (2021). Climate suitability assessment on the Qinghai-Tibet Plateau. Sci. Total Environ. 816:151653. doi: 10.1016/j.scitotenv.2021.151653
Liu, Y., Fan, J., Harris, W., Shao, Q., Zhou, Y. C., Wang, N., et al. (2013). Effects of plateau pika (Ochotona curzoniae) on net ecosystem carbon exchange of grassland in the Three Rivers Headwaters region, Qinghai-Tibet, China. Plant Soil. 366, 491–504. doi: 10.1007/s11104-012-1442-x
Lopes, C. M., De Barba, M., Boyer, F., Mercier, C., Galiano, D., Kubiak, B. B., et al. (2020). Ecological specialization and niche overlap of subterranean rodents inferred from DNA metabarcoding diet analysis. Mol. Ecol. 29, 3143–3153. doi: 10.1111/mec.15549
Miehe, G., Mlehe, S., Kaiser, K., Liu, J., and Zhao, X. (2008). Status and dynamics of the Kobresiapygmaea ecosystem on the Tibetan plateau. Ambio 37, 272–279. doi: 10.1579/0044-7447200837
Miranda, V., and Rothen, C. (2019). Subterranean desert rodents (Genus Ctenomys) create soil patches enriched in root endophytic fungal propagules. Microb. Ecol. 77, 451–459. doi: 10.1007/s00248-018-1227-8
Nevo, E. (1999). Mosaic Evolution Of Subterranean Mammals: Regression, Progression And Global Convergence. New York, NY: Oxford University Press.
Niu, Y., Yang, S., Zhou, J., Chu, B., Ma, S., Zhu, H., et al. (2019). Vegetation distribution along mountain environmental gradient predicts shifts in plant community response to climate change in alpine meadow on the Tibetan Plateau. Sci. Total. Environ. 650, 505–514. doi: 10.1016/j.scitotenv.2018.08.390
Oksanen, J., Simpson, G. L., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., et al. (2018). Vegan: Community ecology package. R package version 2.6-2. Available online at: https://CRAN.R-project.org/package=vegan (accessed May 1, 2022).
Ostoja, S. M., and Schupp, E. W. (2009). Conversion of sagebrush shrublands to exotic annual grasslands negatively impacts small mammal communities. Divers. Distrib. 15, 863–870. doi: 10.1111/j.1472-4642.2009.00593.x
Quan, J., Ouyang, Z., Xu, W., and Miao, H. (2011). Assessment of the effectiveness of nature reserve management in China. Biodivers. Conserv. 20, 779–792. doi: 10.1007/s10531-010-9978-7
Rieder, J. P., Newbold, T., and Ostoja, S. M. (2010). Structural changes in vegetation coincident with annual grass invasion negatively impacts sprint velocity of small vertebrates. Biol. Invasions. 12, 2429–2439. doi: 10.1007/s10530-009-9653-7
Schai-Braun, S. C., Lapin, K., Bernhardt, K. G., Alves, P., and Hackländer, K. (2020). Effect of landscape type, elevation, vegetation period, and taxonomic plant identification level on diet preferences of Alpine mountain hares (Lepus timidusvarronis). Eur. J. Wildl. Res. 66:57. doi: 10.1007/s10344-020-01398-7
Schleuss, P. M., Heitkamp, F., Sun, Y., Miehe, G., Xu, X., Kuzyakov, Y., et al. (2015). Nitrogen uptake in an alpine kobresia pasture on the Tibetan Plateau: Localization by 15N labeling and implications for a vulnerable ecosystem. Ecosyst 25, 946–957. doi: 10.1007/s10021-015-9874-9
Su, D. X., Zhang, Z. H., Chen, Z. Z., and Hu, X. Z. (2003). Parameters for degradation, sandification and salification of rangelands. China. GB19377-2003. Beijing: China Standard Press.
Su, J., Achyut, A., Nan, Z., and Ji, W. (2015). Climate change-induced range expansion of a subterranean rodent: Implications for rangeland management in Qinghai-Tibetan Plateau. PLoS One. 10:e0138969. doi: 10.1371/journal.pone.0138969
Su, J., Hegab, I. M., Ji, W., and Nan, Z. (2018). Function-related drivers of skull morphometric variation and sexual size dimorphism in a subterranean rodent, Plateau zokor (Eospalax baileyi). Ecol. Evol. 8, 4631–4643. doi: 10.1002/ece3.3986
Ṧumbera, R., Mazoch, V., Patzenhauerová, H., Lövy, M., Skliba, J., Bryja, J., et al. (2012). Burrow architecture, family composition and habitat characteristics of the largest social African mole-rat: The giant mole-rat constructs really giant burrow systems. Acta. Theriol. 57, 121–130. doi: 10.1007/s13364-011-0059-4
Tang, L., Dong, S., Sherman, R., Liu, S., Liu, Q., Wang, X., et al. (2015). Changes in vegetation composition and plant diversity with rangeland degradation in the alpine region of Qinghai-Tibet Plateau. Rangeland J. 37, 107–115. doi: 10.1071/RJ14077
Wang, H., Qi, Y., Huang, C., Li, X., Deng, X., and Zhang, J. (2020). Analysis of vegetation changes and dominant factors on the Qinghai-Tibet Plateau, China. Sci. Cold. Arid. Reg. 11, 150–158. doi: 10.3724/SP.J.1226.2019.00150
Wang, Q. Y., Zhang, Y. M., Wei, W. H., and Bian, J. H. (2000a). Food habit of the plateau zokor. Acta Theriol. Sinica. 20, 193–199. doi: 10.16829/j.slxb.2000.03.005
Wang, Q. Y., Zhou, W. Y., Wei, W. H., Zhnag, Y. M., and Fan, N. C. (2000b). The digging behavior of plateau zokor and its relationship with soil hardness. Acta Theriol. Sinica. 20, 277–283. doi: 10.16829/j.slxb.2000.04.006
Wang, X., Dong, S., Yang, B., Li, Y., and Su, X. (2014). The effects of grassland degradation on plant diversity, primary productivity, and soil fertility in the alpine region of Asia’s headwaters. Environ. Monit. Assess. 186, 6903–6917. doi: 10.1007/s10661-014-3898-z
Wang, Y., and Wesche, K. (2016). Vegetation and soil responses to livestock grazing in Central Asian grasslands: A review of Chinese literature. Biodivers. Conserv. 25, 2401–2420. doi: 10.1007/s10531-015-1034-1
Wei, W. R., He, J. D., and Zheng, Q. Y. (2020). Plateau pikas (Ochotona curzoniae) at low densities have no destructive effect on winter pasture in alpine meadows. Rangeland J. 42, 55–61. doi: 10.1071/RJ19042
Wu, J., Ma, W., Li, G., Alhassan, A. R., Wang, H., and Chen, G. (2020). Vegetation degradation along water gradient leads to soil active organic carbon loss in Gahai wetland. Ecol. Eng. 145:105666. doi: 10.1016/j.ecoleng.2019.105666
Xie, J. X., Lin, G. H., Liu, C. X., Yang, C., Deng, X. G., Cui, X. F., et al. (2013). Diet selection in overwinter caches of plateau zokor (Eospalax baileyi). Acta. Theriol. 59, 337–345. doi: 10.1007/s13364-013-0168-3
Xie, J. X., Lin, G. H., Zhang, T. Z., and Su, J. (2014). Foraging strategy of plateau zokors (Eospalax baileyi) when collecting food for winter caches. Pol. J. Ecol. 62, 173–182. doi: 10.3161/104.062.0116
Xu, J., Yin, R., Li, Z., and Liu, C. (2006). China’s ecological rehabilitation: Unprecedented efforts, dramatic impacts, and requisite policies. Ecol. Econ. 57, 595–607. doi: 10.1016/j.ecolecon.2005.05.008
Yu, J., Wan, L., Liu, G., Ma, K., Cheng, H., and Shen, Y. (2022). A meta-analysis on degraded alpine grassland mediated by climate factors: Enlightenment for ecological restoration. Front. Plant Sci. 12:821954. doi: 10.3389/fpls.2021.821954
Zhang, X., Zou, Y., Zou, X., Xu, Z., Nan, X., and Han, C. (2022). DNA metabarcoding uncovers the diet of subterranean rodents in China. PLoS One. 17:e0258078. doi: 10.1371/journal.pone.0258078
Zhang, Y. (2007). “The biology and ecology of plateau zokors (Myospalax fontanierii),” in Subterranean Rodents: News from Underground, eds S. Begall, H. Burda, and C. Schleich (Verlag Berlin Heidelberg: Springer), 239–251.
Zhang, Y., Zhang, Z., and Liu, J. (2010). Burrowing rodents as ecosystem engineers: The ecology and management of plateau zokors (Myospalax fontanierii) in alpine meadow ecosystems on the Tibetan Plateau. Mammal Rev. 33, 284–294. doi: 10.1046/j.1365-2907.2003.00020.x
Zhang, Z., Zhou, H., Zhao, X., Yao, B. Q., Ma, Z., Dong, Q., et al. (2018). Relationship between biodiversity and ecosystem functioning in alpine meadows of the Qinghai-Tibet Plateau. Biodivers. Sci. 26, 111–129. doi: 10.17520/biods.2017021
Zhao, S., Zhao, X., Zhao, J., Liu, N., Sun, M., and Mu, B. (2022). Grassland conservation effectiveness of national nature reserves in northern China. Remote Sens. 14:1760. doi: 10.3390/rs14071760
Keywords: plateau zokor, feeding habits, degraded and reseeded grassland, ITS2 barcode, illumina MiSeq sequencing
Citation: Zhang C, Kang Y, Yao B, An K, Pu Q, Wang Z, Sun X and Su J (2022) Increased availability of preferred food and decreased foraging costs from degraded grasslands lead to rodent pests in the Qinghai-Tibet Plateau. Front. Ecol. Evol. 10:971429. doi: 10.3389/fevo.2022.971429
Received: 17 June 2022; Accepted: 11 August 2022;
Published: 08 September 2022.
Edited by:
Wei Wanrong, China West Normal University, ChinaCopyright © 2022 Zhang, Kang, Yao, An, Pu, Wang, Sun and Su. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Junhu Su, c3VqaEBnc2F1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.