
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 20 October 2022
Sec. Phylogenetics, Phylogenomics, and Systematics
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.969263
 Arthur D. Abegg1,2*
Arthur D. Abegg1,2* Alfredo P. Santos Jr.3
Alfredo P. Santos Jr.3 Henrique C. Costa4
Henrique C. Costa4 Jaqueline Battilana5
Jaqueline Battilana5 Roberta Graboski6
Roberta Graboski6 Fernanda S. L. Vianna7
Fernanda S. L. Vianna7 Weverton S. Azevedo1,2
Weverton S. Azevedo1,2 Nelson J. R. Fagundes7,8
Nelson J. R. Fagundes7,8 Clément M. Castille8
Clément M. Castille8 Pedro C. Prado9
Pedro C. Prado9 Sandro L. Bonatto10
Sandro L. Bonatto10 Hussam Zaher5
Hussam Zaher5 Felipe G. Grazziotin2
Felipe G. Grazziotin2Although the recent advances on the relationship of its major groups, the systematics of the rich fauna of Neotropical snakes is far from being a consensus. In this sense, derived groups presenting continental distributions have represented a main challenge. The taxonomy of the snake tribe Echinantherini is one of the most contentious among the diverse family known as Dipsadidae. The tribe is poorly sampled in phylogenetic studies, resulting in conflicting hypotheses of relationships among its taxa. Moreover, several rare and micro endemic species of Echinantherini have never been evaluated within a comprehensive phylogenetic framework. Here, we assess for the first time the phylogenetic position of the rare Echinanthera amoena within Echinantherini. We based our analyses on a comprehensive multilocus dataset including 14 of the 16 species described for the tribe. Our results support the monophyly of Echinantherini and strongly indicate E. amoena as a unique lineage, phylogenetically positioned apart from all other congeners. From the three current genera (Echinanthera, Taeniophallus, and Sordellina) our results indicate that Echinanthera and Taeniophallus are paraphyletic, since the T. affinis species group is positioned as sister to Echinanthera (except E. amoena) clustering apart from the clade formed by the T. brevirostris and T. occipitalis groups. We describe new genera for the T. affinis and T. occipitalis species groups and an additional monospecific genus for E. amoena. Although we did not evaluate the phylogenetic position of T. nebularis, we described a new genus and removed it from Echinantherini since its morphology strikingly departs from all species now included in the tribe. Finally, we redefine the genera Echinanthera and Taeniophallus and we provide comments about further directions to study the biogeography and the evolution of morphological traits in Echinantherini.
Recent advances in the systematic of the South American herpetofauna have allowed the exploration of a variety of biological questions, with a direct impact on our understanding of the biogeographical patterns, phenotypic evolution, and natural history (Cadle and Greene, 1993; Werneck, 2011; Simon et al., 2016; Correa et al., 2017). Despite these advances, available hypotheses of phylogenetic relationships are still affected by a range of limitations that includes, among others, incomplete lineage sorting, introgression and hybridization, methodological discordance, convergences and parallelisms, and poor taxon and gene sampling (Joly et al., 2009; Nixon and Carpenter, 2012; Mindell, 2013).
In particular, poor taxon sampling can produce pervasive effects that are usually difficult to identify beforehand in empirical studies (Zwickl and Hillis, 2002; Heath et al., 2008). The lack of a comprehensive sampling can generate long branches that may bias the resulting tree topology (Felsenstein, 1978; Siddall and Whiting, 1999; Pol and Siddall, 2001; Bergsten, 2005). Usually, the missing lineages in phylogenetic studies represent rare, divergent, and morphologically distinct taxa, which are difficult to obtain or find in scientific collections (Boakes et al., 2010). These features can make even more difficult to assess the phylogenetic position of a missing taxon or the effect it can produce on the general tree topology.
Recently, while conducting fieldwork in southeastern Brazil, we (ADA and WSA) obtained additional material for Echinanthera amoena (Jan, 1863), one of the most enigmatic snake from South America. With few specimens deposited in scientific collections and no molecular data available, E. amoena represents a missing piece in the systematic puzzle of Dipsadidae, the largest Neotropical snake family (Zaher et al., 2019). This species is allocated in Echinantherini Zaher et al., 2009 due to its general hemipenial similarity, although it does not share some of the synapomorphies of the genus Echinanthera (Schargel et al., 2005). Besides its morphology, E. amoena also presents a unique biology, being intimately associated to microhabitats restricted to the margins of streams in highlands in the Brazilian Atlantic Forest. This habitat is not shared by any other Echinantherini and makes E. amoena particularly difficult to obtain in the field (Cassimiro et al., 2013; Azevedo et al., 2018).
Echinantherini was recently described to accommodate one of the groups of dipsadids with an enduring unstable generic taxonomic history. The tribe was proposed to allocate 15 strictly South American species belonging to the genera Echinanthera Cope, 1894 and Taeniophallus Cope, 1895. Zaher et al. (2009) included two species of Taeniophallus in their molecular analysis, but recognized the monophyly of the tribe including both Echinanthera and Taeniophallus based on the putative hemipenial synapomorphies shared by these genera (Di-Bernardo, 1992; Myers and Cadle, 1994; Zaher, 1999; Schargel et al., 2005). In the next year, Vidal et al. (2010) corroborated the close affinities of Echinanthera and Taeniophallus by adding a species of Echinanthera to their molecular analysis. Grazziotin et al. (2012) significantly expanded the taxonomic sampling of South American dipsadids, and recovered the enigmatic monotypic genus Sordellina Proctor, 1923, nested within Echinantherini as the sister group of Taeniophallus (Figure 1). Such unexpected result was acknowledged by the authors who included Sordellina in the tribe Echinantherini. Subsequent studies have included sequences of Echinantherini without expanding the taxon sampling for the tribe (Figure 1; Pyron et al., 2013; Figueroa et al., 2016; Tonini et al., 2016; Zaher et al., 2018). The only exception was Zaher et al. (2019) who added Taeniophallus occipitalis to the previously sequenced species (Figure 1). Although all recent studies recovered Echinantherini as monophyletic, the relationships between its constituent parts are still unsolved (Figure 1). Until now, even the largest phylogeny of Dipsadidae (Zaher et al., 2018) has included less than half of the known diversity of Echinantherini.
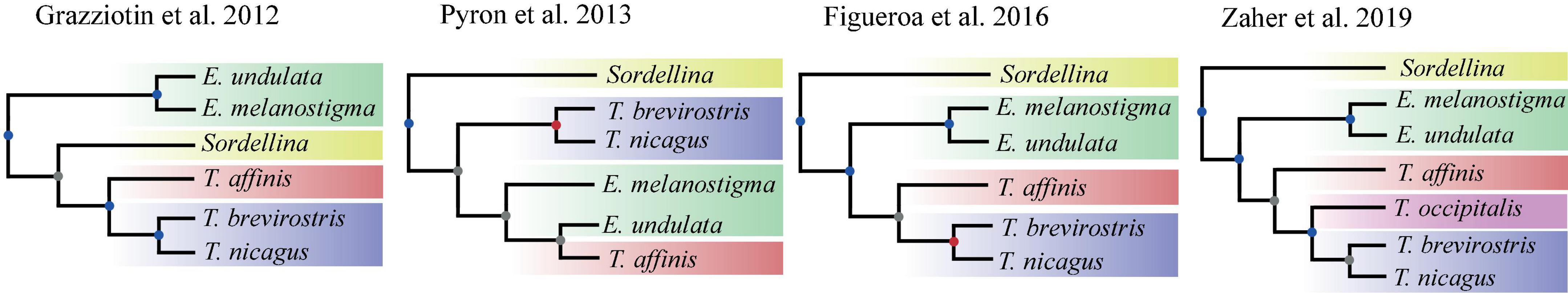
Figure 1. Comparison between previous topologies on Echinantherini systematics, showing the state of flux on genus-level relationships. Colored circles on each node correspond to bootstrap support values: red (100%), blue (70–99%), and gray (<70%).
Nonetheless, difficulties in classifying most species currently allocated to Echinantherini precede the era of molecular phylogenies. The taxonomic status of Echinanthera and Taeniophallus has been contentious since the mid-1970s (Myers, 1974; Di-Bernardo, 1992, 1994, 1996; Myers and Cadle, 1994). In the last few decades, molecular techniques, mainly based on DNA sequences, started to solve some of these longstanding morphological-based problems of relationships within Dipsadidae. As a consequence, several taxonomic changes at generic and suprageneric levels were advanced as a way to adequately reflect the recovered evolutionary history (e.g., Zaher et al., 2009, 2018; Grazziotin et al., 2012; Pyron et al., 2015; Arredondo et al., 2020; Trevine et al., 2022). However, several tribes that are well-supported molecularly and morphologically are still poorly sampled in molecular phylogenies, hampering any attempt to provide reliable hypotheses of phylogenetic relationships within these groups. The expansion of the taxon sampling for Echinantherini, including particularly the rare E. amoena, represents a key factor to elucidate the phylogenetic affinities within the group and helps clarify the contradictory molecular signals reached so far.
Here we evaluate the phylogenetic position of E. amoena within Echinantherini, through the analysis of a multilocus dataset. Besides E. amoena, for the first time, we sequenced and analyzed six other unsampled species of Echinantherini, representing 87% of the total number of species described for the tribe. Based on our results, we redefine the taxonomy of Echinantherini to accommodate the tribe diversity, and we additionally discuss and redefine a number of morphological characters for all valid genera within the tribe.
We generated DNA sequences for 35 individuals representing all three genera (Echinanthera, Taeniophallus, and the monospecific Sordellina) and 14 out of the 16 known species of Echinantherini, missing only Taeniophallus nebularis Schargel, Rivas and Myers, 2005 and Echinanthera cephalomaculata Di-Bernardo, 1994. We sampled five out of six species of Echinanthera, including E. amoena, E. cephalostriata Di-Bernardo, 1996, E. cyanopleura (Cope, 1885), E. melanostigma (Wagler, 1824), and E. undulata (Wied, 1824). We also covered all species of Taeniophallus within the three known species groups, as follow: (1) affinis group—T. affinis (Günther, 1858), T. bilineatus (Fischer, 1885), and T. persimilis (Cope, 1869), T. poecilopogon (Cope, 1863); (2) brevirostris group—T. brevirostris (Peters, 1863) and T. nicagus (Cope, 1868); (3) occipitalis group—T. occipitalis (Jan, 1863), and T. quadriocellatus Santos, Di-Bernardo and Lema, 2008. Taeniophallus quadriocellatus, T. bilineatus, T. persimilis, T. poecilopogon, E. amoena, E. cephalostriata, and E. cyanopleura were sequenced here for the first time. All new sequences were deposited in GenBank (Supplementary Appendix 1).
We extracted DNA from scales, muscles, and liver tissues using the PureLinkVR Genomic DNA kit (ThermoFisher, Waltham, MA, USA). We amplified by PCR partial sequences for six genes, including three mitochondrial (12S, small subunit ribosomal RNA; 16S, large subunit ribosomal RNA; cytb, cytochrome b) and three nuclear (bdnf, brain derived neurotrophic factor; c-mos, oocyte maturation factor Mos; nt3, neurotrophin-3). We performed the PCRs using the protocols described in Grazziotin et al. (2012). The primer sequences are given in Supplementary Appendix 2: Table S1. Both DNA strands were sequenced using the sequencing service of Macrogen (Seoul, South Korea). Chromatograms were evaluated and assembled using Geneious v.8.1.31 (Kearse et al., 2012). Possible contaminations were accessed by using the BLAST tool (Altschul et al., 1997) at the NCBI website. We aligned the sequences using MAFFT (Katoh and Standley, 2013) as implemented in Geneious. We used the MAFFT default settings to align all loci, except for the loop regions of the RNA genes, for which we used the G-INS-i algorithm. The concatenated alignment can be found at doi:10.6084/m9.figshare.15116877.
The phylogenetic position of Echinanthera amoena and monophyly of Echinantherini were tested using the expanded data set from Zaher et al. (2018), which comprises a concatenated alignment of 3,938 base pairs from the same six genes used in the present study. The dataset was composed by 338 outgroups from the following caenophidian families (number of terminals in parentheses): Acrochordidae (2), Atractaspididae (4), Boidae (1), Calamariidae (2), Elapidae (6), Erycidae (1), Colubridae (5), Dipsadidae (281), Grayidae (1), Homalopsidae (3), Natricidae (5), Lamprophiidae (2), Pareidae (3), Psammophiidae (2), Pseudoxenodontidae (3), Pseudoxyrhophiidae (2), Sibynophiidae (2), Viperidae (9), and Xenodermidae (3). The dataset includes representatives from most currently recognized dipsadid tribes, except Amnesteophiini and Incaspidini.
We used Geneious to generate our concatenated matrix, setting 14 initial partitions, as follows: each protein-coding gene was partitioned by codon position; and each rRNA was analyzed as a separate partition. We defined the best model of nucleotide substitutions and the best partition scheme using PartitionFinder2 v.2.1.1 (Lanfear et al., 2012). We only allowed PartitionFinder2 to select the GTR model as implemented in RAxML v.8 (Stamatakis, 2014). The Akaike Information Criteria with correction (AICc) was used as the selection criteria and the “greedy” algorithm was used as the searching option.
We performed maximum likelihood (ML) analyses using RAxML v.8 through the CIPRES portal (Miller et al., 2010) using the concatenated dataset and the best partition schemes selected by PartitionFinder2. We conducted a rapid bootstrap analysis and search for the best scoring ML tree in the same run (option -f a). Branch support was assessed by using 1,000 rapid bootstrap iterations, whose robustness was classified as follows: unambiguously supported (100%), strongly supported (80–99%), moderately supported (70–79%), and weakly supported (<70%).
We revisited the limited literature providing relevant morphological information on the genera Echinanthera, Sordellina, and Taeniophallus (e.g., Myers, 1974; Hoge and Romano, 1978; Di-Bernardo and Lema, 1986, 1987, 1988; Di-Bernardo, 1991, 1992, 1994, 1996; Myers and Cadle, 1994; Schargel et al., 2005; Santos-Jr et al., 2008; Miranda and Fernandes, 2012). Specimens examined are listed in the Supplementary Appendix 3. Scale counts followed Myers (1974) and Dowling (1951). Hemipenial preparation followed the method of Pesantes (1994) with the modifications proposed by Zaher and Prudente (2003). The terminology for hemipenial morphology followed Zaher (1999) and Schargel et al. (2005).
To compare the cranial osteology, we analyzed prepared skulls deposited in scientific collections and generated 3D images using high-resolution micro computed tomography (μ-CT). Specimens were scanned in a 300-kV μ-focus X-ray source micro computed tomography GE Phoenix v| tome| x M 300 (General Electric Measurement and Control Solutions, Wunstorf, Germany) at the Laboratório de Microtomografia of the Museu de Zoologia da Universidade de São Paulo (MZUSP). The acquired scan data was processed on a high-end computer HP Z820 workstation with eight core Intel Xeon E5-2660, 2.20 GHz and 128 GB of memory. Reconstruction of raw data was performed using the system supplied software phoenix datos| x reconstruction v. 2.3.0 (General Electric Measurement and Control Solutions, Wunstorf, Germany). Three-dimensional visualization, segmentation, and analysis of the reconstructed data was performed using myVGL for Windows 64 bits, v3.0. We followed Cundall and Irish (2008) for skull terminology. For external morphology we evaluated all the 16 species currently attributed to Echinantherini; for hemipenial morphology we analyzed 15 species (E. cephalomaculata was not sampled); and for cranial osteology we analyzed 14 species (E. cephalomaculata and T. nebularis were not sampled).
Since detailed morphological descriptions are already available in the literature for most species of Echinantherini (see references above), we provided for Echinanthera amoena the following additional descriptions: external morphology, color pattern variation in preservative and in life, hemipenial morphology, cranial osteology, and meristic variation. We measured specimens with a digital caliper to the nearest 0.1 mm, except for snout–vent length (SVL) and tail length (TL), which were measured to the nearest 1.0 mm with a flexible ruler. Sex was verified by an incision at the base of the tail to check for the presence/absence of a hemipenis. Color descriptions followed Köhler (2012). Sexual dimorphism in number of ventral and subcaudal scales were tested using Student’s t-test, significance was defined using alpha < 0.05. The statistical tests were implemented in the R environment (R Core Team, 2015). Descriptions and results of these analyses are provided in Supplementary Appendix 4.
We mapped the trait evolution of some morphological characters frequently used to define species groups within Echinantherini. We built a morphological matrix with 13 characters for 14 species of Echinantherini, and we also included four outgroups, which represent the four successive sister tribes of Echinantherini in the molecular tree. We optimized the morphological matrix on the tree topology derived from the analysis of the combined molecular dataset. We used the software Mesquite (Maddison and Maddison, 2009) to build the morphological matrix and TNT v.1.5 (Goloboff and Catalano, 2016) to map the trait evolution on the molecular tree. Only the unambiguous synapomorphies (shared or private) are described and commented in the Results and Discussion section. The result of this analysis is provided in Supplementary Appendix 5.
The concatenated matrix encompassed 4,002 base pairs of aligned sequences. The best fit partition scheme grouped our initial partitions into 12 subsets. All the partitions and the selected models are shown in Supplementary Appendix 2: Table S2. The complete phylogenetic tree based on all concatenated genes is available in Supplementary Appendix 6.
The overall topology of our ML tree is similar to recent studies (e.g., Zaher et al., 2009, 2018; Di-Bernardo, 1996; Pyron et al., 2011, 2013; Grazziotin et al., 2012; Figueroa et al., 2016). Our results unambiguously recovered the superfamilies Elapoidea and Colubroidea (sensu Zaher et al., 2009), as well as the families Acrochordidae, Xenodermidae, Pareidae, Viperidae, Homalopsidae, Lamprophiidae, Pseudoxyrhophiidae, Elapidae, Natricidae, Calamariidae, and Pseudoxenodontidae. The following families were strongly supported: Atractaspididae (97%), Psammophiidae (81%), Colubridae (84%), and Sibynophiidae (90%). Only Dipsadidae (< 60%) was weakly recovered.
Within Dipsadidae, we recovered the genera Thermophis Malnate, 1953 and Stichophanes Wang et al., 2014 as successive sister groups of all other New-World dipsadids. Both subfamilies Dipsadinae and Xenodontinae were recovered as monophyletic but with weak support, while Carphophiinae was paraphyletic, with Farancia Gray, 1842 nested within its diversity (<70%).
We recovered most of the tribes of Dipsadidae previously recognized as monophyletic: Diaphorolepidini (99%), Imantodini (97%), Dipsadini (sensu Grazziotin et al., 2012; 81%), Conophiini (sensu Zaher et al., 2018, <60%), Echinantherini (90%), Elapomorphini (96%), Tropidodryadini (100%), Philodryadini (94%), Hydropsini (98%), Tachymenini (99%), Hydrodynastini (100%), Pseudoboini (99%), Psomophiini (100%), Alsophiini (sensu Grazziotin et al., 2012, 71%), and Xenodontini (100%).
The tribe Caaeteboini grouped with the genus Xenopholis Peters, 1869 (<70%); and Nothopsini was positioned as the sister group of Dipsadini (<70%). The tribe Saphenophiini was not retrieved as monophyletic, since the genus Pseudalsophis Zaher et al., 2009 was positioned as the sister group of Xenodontini (<70%), whereas Saphenophis Myers 1973 was recovered as the sister group of a clade including Tropidodryadini and Philodryadini (<70%) (Supplementary Figure 1). The tribe Echinantherini was positioned as the sister group of Elapomorphini (<70%) (Figure 2 and Supplementary Figure 1).
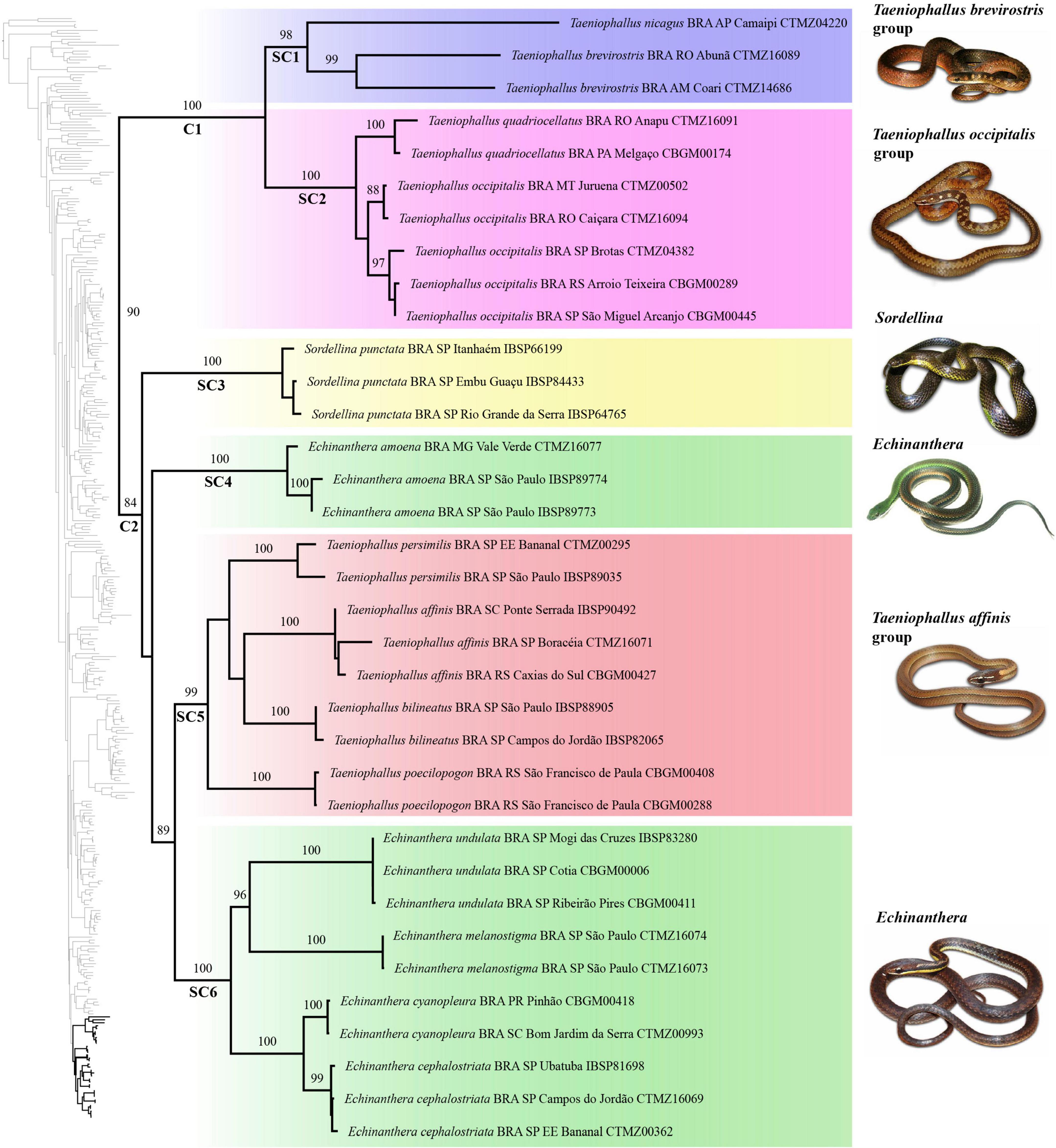
Figure 2. Maximum likelihood tree based on the concatenated matrix (RAxML) of the tribe Echinantherini. Only bootstrap support values > 70 are shown for clarity. The topology of the complete ML tree is provided on the left; the clade highlighted in black corresponds to the tribe Echinantherini (Supplementary Appendix 6).
Although we did not evaluate the phylogenetic position of T. nebularis based on molecular evidence, our preliminary analysis of the morphological diversity within the tribe strongly suggests that this species does not belong to Echinantherini. Taeniophallus nebularis has an external general morphology similar to Echinantherini (Figures 3, 4), but departs strikingly from all species included in this tribe by the symmetrically ornamented hemipenis and the presence of 19 dorsal scales instead of 17 or 15. Moreover, the deeply bilobed hemipenis, with lobes bearing one or more enlarged apical calyces, and the presence of a proximal naked pocket are conditions present only in T. nebularis, not shared with any other species of Echinantherini. Finally, T. nebularis does not share the pair of small light dots on the parietal scales, which is the main putative synapomorphy for the tribe. Therefore, we describe below a new genus for T. nebularis and remove it from Echinantherini. We provisionally allocate the new genus as Dipsadidae incertae sedis to indicate the uncertain affinity within the family.

Figure 3. General morphological aspects of some species representing the diversity of the genera Myersinia, Adelphostigma, Amnisiophis and Dibernardia. (A) Myersinia nebularis; (B) Adelphostigma cf. occipitalis; (C) Adelphostigma quadriocellata; (D) Amnisiophis amoenus; (E) Dibernardia affinis; (F) Dibernardia bilineata; (G) Dibernardia persimilis; and (H) Dibernardia poecilopogon.

Figure 4. General morphological aspects of some species representing the diversity of the genera Echinanthera, Sordellina and Taeniophallus. (A) Echinanthera cephalomaculata; (B) Echinanthera cephalostriata; (C) Echinanthera cyanopleura; (D) Echinanthera melanostigma; (E) Echinanthera undulata; (F) Sordellina punctata; (G) Taeniophallus brevirostris; and (H) Taeniophallus nicagus.
Myersinia Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus
Zoobank registration. urn:lsid:zoobank.org:act:9F960D61-AB0D-4C88-AA33-B9205E1CF3A8
Type species: Taeniophallus nebularis Schargel et al., 2005.
Etymology: Named after the North American herpetologist Charles W. Myers (1936–2018). Charles Myers was one of the most notable herpetologists of his generation, and his research on taxonomy and systematics of amphibians and reptiles has expressively increased our knowledge on the herpetofauna of the Americas. Together with Marcos Di-Bernardo, he was one of the main experts on the systematics of Echinantherini. The genus name is feminine.
Distribution and Habitat: Myersinia is endemic to Península de Paria, in northern Venezuela (see Figure 1 in Schargel et al., 2005). The vegetation in this region is represented by Coastal Cloud Forests (Schargel et al., 2005).
Diagnosis: (1) eight supralabials, with 2nd–3rd touching the loreal and 3rd–5th touching the orbit; (2) 19/19/17 dorsal scale rows, usually with one or two apical pits; (3) lateral ends of each ventral bearing triangle-shaped markings (Figure 3A); (4) midventral portion immaculate; (6) hemipenis noncapitate, deeply bilobed and with a prolonged asulcate interspinal gap extending distally from a transverse pair of large spines onto medial sides of the lobes, which bears enlarged apical calyces (Schargel et al., 2005) (Figure 5A).

Figure 5. Comparative hemipenial morphology among the evaluated genera. (A) Myersinia nebularis (MHNLS 15743) redrawn from Schargel et al. (2005); (B) Taeniophallus nicagus (IBSP 91656); (C) Taeniophallus brevirostris (MPEG 8639); (D) Adelphostigma quadriocellata (MPEG 8272); (E) Adelphostigma occipitalis (DZUFRGS 6466); (F) Sordellina punctata (IBSP 84433); (G) Amnisiophis amoenus (UFMG 3272); (H) Dibernardia poecilopogon (DZUFRGS 5973); (I) Dibernardia persimilis (IBSP 55279); (J) Dibernardia affinis (IBSP 51561); (K) Dibernardia bilineata (IBSP 82065); (L) Echinanthera undulata (IBSP 92283); (M) Echinanthera melanostigma (UFMG 3273); (N) Echinanthera cyanopleura (IBSP 81751); and (O) Echinanthera cephalostriata (URCA 4103). Scale bars: 2 mm.
Putative Synapomorphies: Hemipenis deeply bilobed with a prolonged asulcate interspinal gap extending distally from a transverse pair of large spines onto medial sides of the lobes, which bears one or more enlarged apical calyces (Figure 5A).
Maximum likelihood analysis recovered Echinantherini as comprising the following two main clades (Figure 2): clade C1 (100%), composed by the T. brevirostris and T. occipitalis species group; clade C2 (84%), formed by Sordellina and the species of Echinanthera and T. affinis species group.
The clade C1 was structured in two subclades, as follows: SC1 (98%), comprising only the generic type species T. nicagus and T. brevirostris; and SC2 (100%), including T. occipitalis and T. quadriocellatus. Clade C2 was formed by the following four subclades: SC3 (100%), comprising S. punctata (Peters, 1880); SC4 (100%), formed by E. amoena; SC5 (99%), including T. affinis, T. bilineatus, T. persimilis, and T. poecilopogon; and SC6 (100%), comprising E. cephalostriata, E. cyanopleura, E. melanostigma, and E. undulata. Subclades SC3 and SC4 are the successive sister groups of the clade (89%) formed by subclades SC5 and SC6. Within SC5, T. poecilopogon and T. persimilis are positioned as successive sister taxa of a clade formed by T. bilineatus and T. affinis, although all of these relationships are weakly supported (<70%). Among the species from SC6, the generic type E. cyanopleura and E. cephalostriata grouped in an unambiguously supported clade that is sister of a clade including E. melanostigma and E. undulata, retrieved with strong support (96%).
Our phylogenetic results indicate that both genera, Echinanthera and Taeniophallus, are paraphyletic. Additionally, the morphological comparison among taxa also indicates that both genera are composed by different evolutionary entities. To retrieve a monophyletic classification for Echinantherini we are proposing below a new taxonomic scheme for the tribe. We are erecting a new genus to accommodate E. amoena and redefining Echinanthera to include only E. cephalostriata, E. cyanopleura, E. melanostigma, E. undulata (subclade SC6; Figure 2), and tentatively the unsampled E. cephalomaculata. We are also proposing the recognition of the three species groups of Taeniophallus (subclades SC1, SC2, and SC5; Figure 2) as different genera. Therefore, we are redefining the genus Taeniophallus to include only T. nebularis and the brevirostris species group (SC1); and we are describing two new genera to accommodate the occipitalis (SC2) and affinis (SC5) species groups.
Here we provide a new taxonomic scheme for Echinantherini classifying the species in the following six genera: Adelphostigma Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus, including A. occipitalis new combination and A. quadriocellata new combination; Amnisiophis Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus, including A. amoenus new combination; Dibernardia Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus, including D. affinis new combination, D. bilineata new combination, D. persimilis new combination, and D. poecilopogon new combination; Echinanthera, including E. cephalomaculata, E. cephalostriata, E. cyanopleura, E. melanostigma, and E. undulata; Taeniophallus, including T. brevirostris, and T. nicagus; and the monospecific Sordellina that only includes S. punctata.
Below we expanded the description and comparison among the genera of Echinantherini based on our new classification.
Adelphostigma Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus
Zoobank registration. urn:lsid:zoobank.org:act:425AB6EE-D9A5-4801-80AB-3541DE66EC7C
Type species. Enicognathus occipitalis Jan, 1863.
Contents. Adelphostigma occipitalis (Jan, 1863) new comb.; A. quadriocellata (Santos-Jr et al., 2008) new comb.
Etymology: From the composition of the Ancient Greek words ′αδελφóς (adelphos [twin, brother]) and στíγμα (stigma [mark, brand, spot]) (Liddell and Scott, 1883; Brown, 1954), in reference to the paired dorsal spots typical of the species here included in this new genus. The genus name is feminine.
Distribution and Habitat: Adelphostigma occurs from the state of Pará, northern Brazil, southwestward to Peru, Bolivia and Paraguay, southeastward to Brazilian coast and southward to Argentina and Uruguay (Nogueira et al., 2019) (Figure 6C). It is distributed in many ecoregions, ranging from the Amazon and Atlantic forests to the Pantanal floodplains, Cerrado, Pampa grasslands and coastal dunes (Myers, 1974; Santos-Jr et al., 2008; Nogueira et al., 2019).
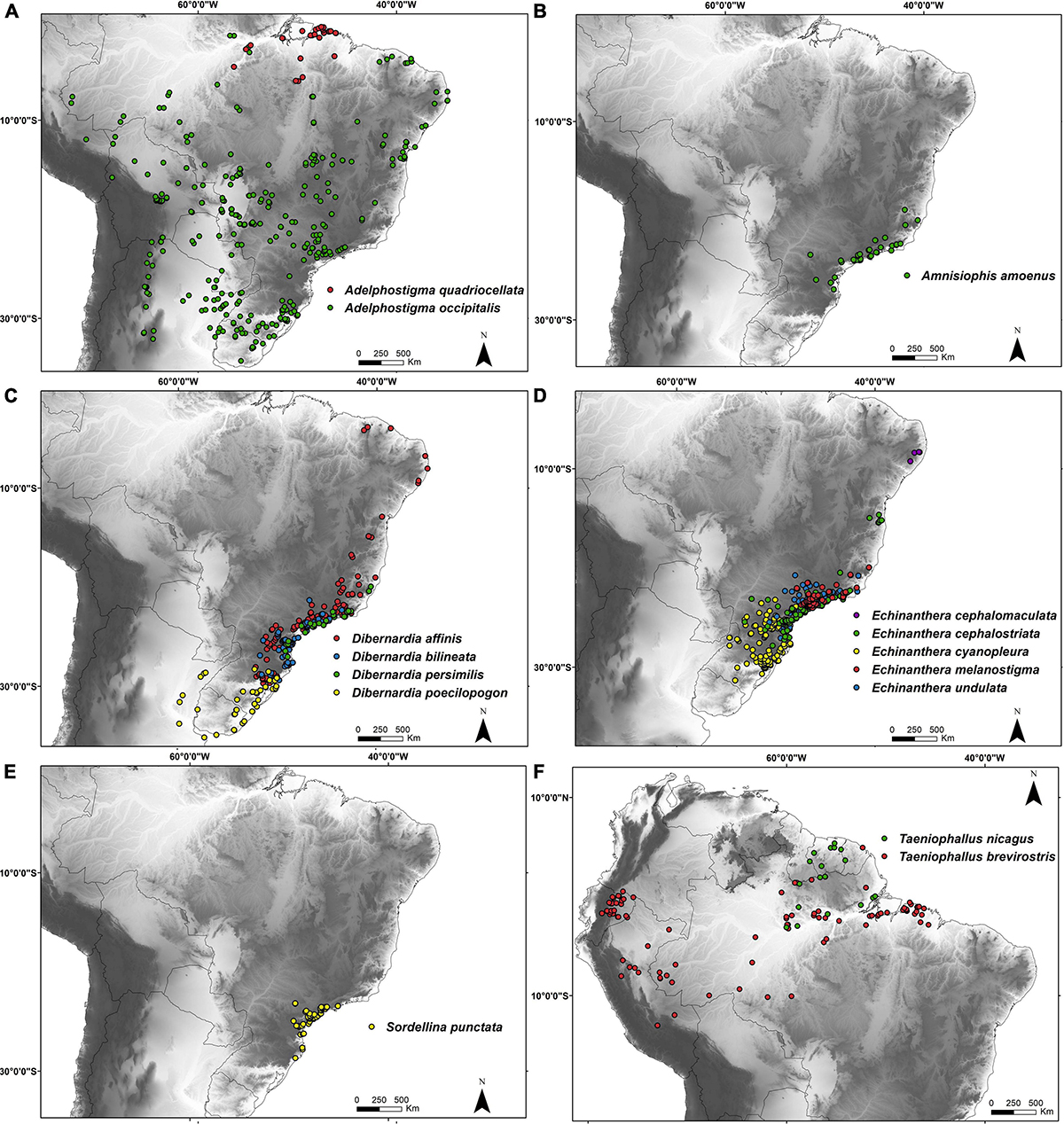
Figure 6. Geographical distribution of Echinantherini according to the taxonomic rearrangement proposed in this study. Locality data are based on Nogueira et al. (2019). (A) Adelphostigma; (B) Amnisiophis; (C) Dibernardia; (D) Echinanthera; (E) Sordellina; and (F) Taeniophallus.
Diagnosis: Adelphostigma can be recognized by presenting (1) usually eight supralabials, with 2nd touching the loreal and 3rd–5th touching the orbit; (2) 15/15/15 or 15/15/13 dorsal scale rows without apical pits; (3) a conspicuous pale canthal line and a middorsal pattern composed of dark-brown blotches in the anterior region that are gradually replaced by paired spots toward the tail; (4) immaculate midventral portion of the body; (5) hemipenis unilobed or slightly bilobed, unycaliculate and unicapitate, with an asulcate interspinal gap interrupted by an enlarged central spine approximately at the midorgan; (6) a nude, usually deep and proximally open pocket immediately below the capitulum (Figures 4B,C, 5D, E,7D and Table 1).

Figure 7. Comparative midventral pattern in Echinantherini. (A) Adelphostigma cf. occipitalis; (B) Amnisiophis amoenus; (C) Dibernardia persimilis; (D) Echinanthera cephalostriata; (E) Sordellina punctata; and (F) Taeniophallus nicagus.
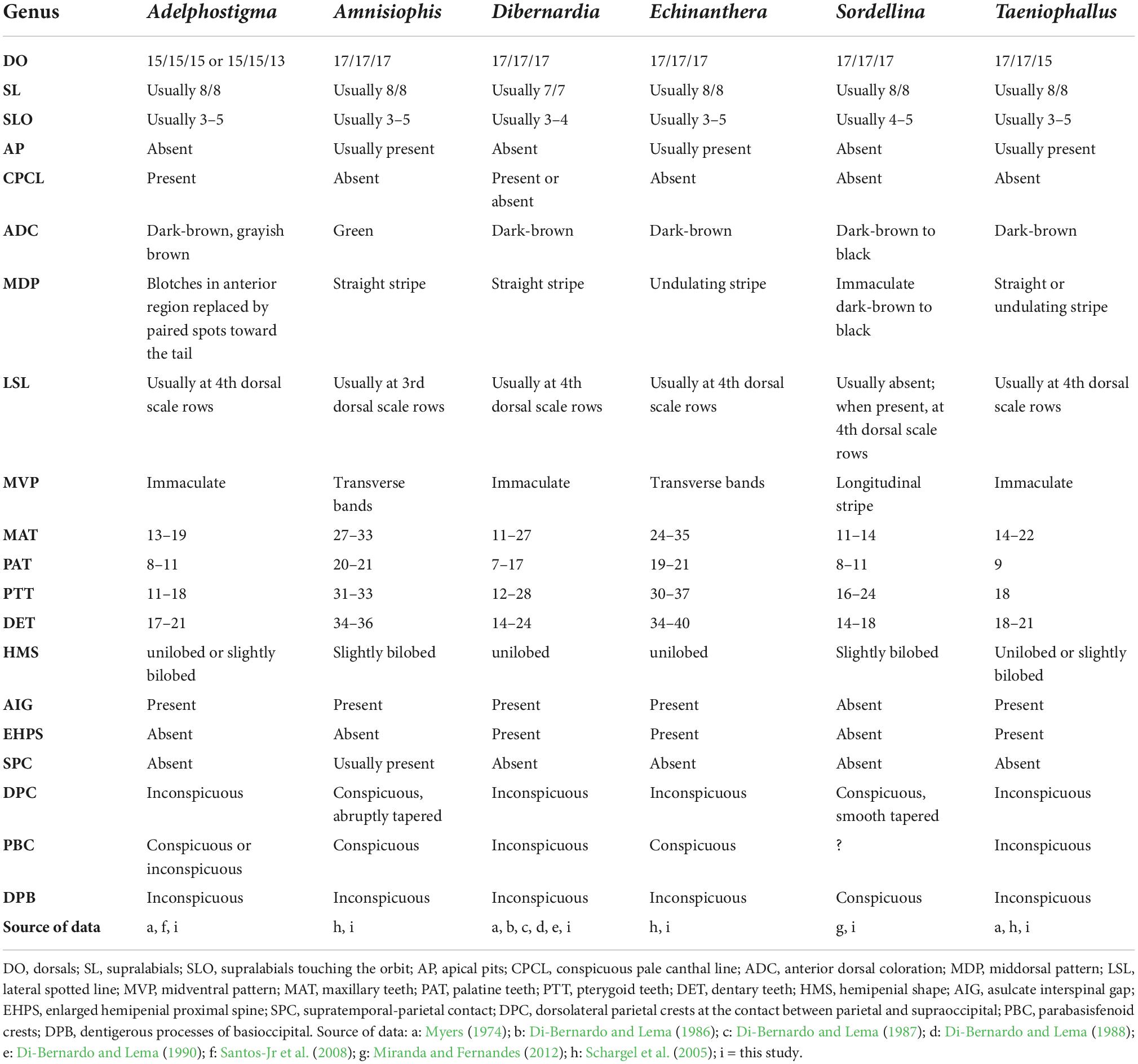
Table 1. Comparative morphology within the new taxonomic arrangement for Echinantherini genera.
Synapomorphies: (1) middorsal pattern composed of dark-brown blotches in the anterior region that are gradually replaced by paired spots toward the tail.
Remarks: This genus corresponds to the occipitalis group of Taeniophallus (sensu Schargel et al., 2005).
Amnisiophis Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus
Zoobank registration. urn:lsid:zoobank.org:act:30A32D0F-061B-4CFC-9895-A96E32371F69
Type species: Enicognathus amoenus Jan, 1863.
Etymology: From the combination of the Latin word amnis (stream of water, river) and the Greek word oφις (ophis [snake]) (Liddell and Scott, 1883; Brown, 1954). This name was chosen in reference to the habit of the monotypic species A. amoenus, whose individuals inhabit the margins of forest streams. The genus name is masculine.
Distribution and Habitat: Amnisiophis amoenus occurs in southeastern and southern Brazil, in regions of ombrophylous forests throughout the Brazilian Atlantic Forest between 500 and 1440 m (Azevedo et al., 2018; Nogueira et al., 2019; Figure 6B).
Diagnosis: Amnisiophis amoenus can be distinguished by (1) usually eight supralabials, with 2nd–3rd touching the loreal and 3rd–5th touching the orbit; (2) 17/17/17 dorsal scale rows, usually with one or two apical pits; (3) a dorsal color that shifts from green on the back of the head and first third of the body to brown on the midbody and dark brown on the posterior part; (4) midventral portion with transverse bands at the base of the ventral scales, especially observable in adults; (5) dorsal scales of the 3rd row with a light spot forming a continuous line from the anterior part of the body toward the tail; (6) hemipenis slightly bilobed, with an interspinal asulcate gap and absence of an enlarged proximal spine; (7) supratemporals usually contact parietal; (8) dorsolateral crests of the parietal conspicuous and abruptly tapered in the posterior region, close to the contact with supraoccipital; (9) parabasisfenoid lateral crests conspicuous; (10) large number of maxillary (27–33), palatine (20–21), pterygoid (31–33) and dentary teeth (34–36) (Figures 3D, 5G, 7B, 8A and Table 1).
Synapomorphies: (1) dorsal color that shifts from green on the back of the head and first third of the body to brown on the midbody and dark brown on the posterior part; (2) dorsolateral crests of the parietal conspicuous and abruptly tapered in the posterior region, close to the contact with supraoccipital.
Remarks: The natural history of A. amoenus is largely unknown, although based on field observations the species is particularly associated to streams, where it putatively preys on torrent frogs, genus Hylodes (Azevedo et al., 2018; Muscat and Moroti, 2020).
Dibernardia Abegg, Santos-Jr, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin, new genus.
Zoobank registration. urn:lsid:zoobank.org:act:DCE66544-7BEB-46EB-95C3-EB8BB879B482
Type species: Dromicus affinis Günther, 1858.
Contents. Dibernardia affinis (Günther, 1858) new comb., D. bilineata (Fischer, 1885) new comb., D. persimilis (Cope, 1869) new comb., and D. poecilopogon (Cope, 1863) new comb.
Etymology: Named after the Brazilian herpetologist Marcos Di Bernardo (1963–2006). During his short and prolific career, he published several studies on the taxonomy of Echinantherini, including the revalidation of Echinanthera (Di-Bernardo, 1992) and the description of two species (Di-Bernardo, 1994, 1996). We honor Marcos Di Bernardo by giving his name to this genus, in recognition to his contributions to South American herpetology. The genus name is feminine.
Distribution and Habitat: Species of Dibernardia occurs from province of Buenos Aires, in northeastern Argentina, throughout Uruguay northward to northeastern Brazil in the state of Ceará (Nogueira et al., 2019; Figure 6C). Dibernardia also inhabits the Pampa grasslands (Uruguayan savanna) and the dense and mixed ombrophilous forests of Atlantic Forest (Bérnils, 2009; Nogueira et al., 2019).
Diagnosis: Dibernardia can be recognized by (1) usually seven supralabials, with 2nd touching the loreal and 3rd–4th touching the orbit; (2) 17/17/17 dorsal scale rows without apical pits; (3) immaculate midventral portion of the body; (4) hemipenis unilobed, unicalyculate, and semicapitate, with an asulcate interspinal gap extending from the distal to the proximal region of the organ; in its most distal portion, it terminates as an abruptly cluster of several flaplike enlarged papillae (Schargel et al., 2005); (5) dorsolateral crests of the parietal bone inconspicuous in the region near the contact with the supraoccipital; (6) parabasisphenoid lateral crests inconspicuous (Figures 5H–K, 7B and Table 1).
Synapomorphy: asulcate interspinal gap bearing an abruptly cluster of several flaplike enlarged papillae in its most distal portion.
Remarks: This genus corresponds to the affinis group of Taeniophallus (sensu Schargel et al., 2005).
Echinanthera Cope, 1894.
Type species: Aporophis cyanopleurus Cope, 1885, by original designation.
Contents. Echinanthera cephalomaculata (Di-Bernardo, 1994), E. cephalostriata (Di-Bernardo, 1996), E. cyanopleura (Cope, 1885), E. melanostigma (Wagler, 1824), and E. undulata (Wied, 1824).
Etymology: Not informed by Cope (1894), but maybe a combination of the Greek words εχις (echis [adder, viper]) and ανθηρóς (antheros [flowery, blooming]) (Liddell and Scott, 1883; Carreira et al., 2005), probably in reference to the color pattern. The genus name is feminine.
Distribution and Habitat: The genus Echinanthera occurs from the province of Misiones, in northeastern Argentina, to northeastern Brazil, in the state of Pernambuco (Nogueira et al., 2019; Figure 6D). They inhabit the Atlantic Forest from sea level to 1730 meters elevation (Di-Bernardo, 1994, 1996; Menezes et al., 2018).
Diagnosis: Echinanthera can be recognized by (1) usually eight supralabials, with 2nd–3rd or 2nd only touching the loreal and 3rd–5th touching the orbit; (2) 17/17/17 dorsal scale rows usually with one or two apical pits; (3) middorsal pattern composed by an undulating dark-brown stripe (vestigial in E. cephalostriata, E. cyanopleura and E. melanostigma); (4) midventral portion with transverse bands at the base of the ventral scales, especially observable in adults; (5) hemipenis unilobed, unicalyculate and semicapitate, with an asulcate interspinal gap extending from the distal to the proximal region of the organ; in its distal end, it terminates in a homogeneous group of small to medium-sized papillae; (6) dorsolateral crests of the parietal bone inconspicuous in the region close to the contact with supraoccipital; (7) parabasisphenoid lateral crests conspicuous; (8) large number of maxillary (24–35), palatine (19–21), pterygoid (30–37) and dentary teeth (34–40) (Figures 4A–E, 5L–O, 7D, 8B–E and Table 1).
Synapomorphy: middorsal pattern composed of a dark-brown undulating stripe.
Remarks: With the new taxonomic arrangement, the morphological characters generally used to define Echinanthera are also shared by Amnisiophis and Taeniophallus. The recognition of Echinanthera as an independent taxon incorporates in the taxonomy our knowledge about the morphology, phylogenetic relationships, and biogeographic patterns of Echinantherini (see discussion below for an extended justification).
Sordellina Procter, 1923.
Type species: Sordellina brandon-jonesii Procter, 1923, by monotypy.
Etymology: Named after Ferdinando Sordelli (1837–1916), Italian artist and naturalist, “for whose beautiful work in the ‘Iconographie Générale des Ophidiens’ all workers at snakes will ever be grateful” (Procter, 1923). The genus name is feminine.
Distribution and Habitat: Sordellina punctata is endemic to southeastern and southern Brazil, inhabiting open areas and riparian vegetation in the Atlantic Forest (Nogueira et al., 2019) (Figure 6E).
Diagnosis: Sordellina punctata can be recognized by having (for additional characters, see Hoge and Romano, 1978; Miranda and Fernandes, 2012): (1) usually eight supralabials; (2) 17/17/17 dorsal scale rows, with no apical pits; (3) head dark brown in dorsal view with supralabials mottled with white or yellow; (4) dorsum uniformly dark brown to black; (5) hemipenis slightly bilobed, the lobular region covered by papillae with little or no tissue interconnection; (6) 11–14 prediastemal maxillary teeth and two postdiastemal teeth; (7) dorsolateral crests of the parietal conspicuous and smoothly tapered in the posterior region, close to the contact with supraoccipital (Figures 4F, 5F).
Synapomorphies: (1) a uniform dark brown to black dorsum, without a vertebral straight/undulating stripe, or blotches; (2) dorsolateral crests of the parietal conspicuous and smoothly tapered in the posterior region, close to the contact with supraoccipital.
Remarks: Although Sordellina departs clearly from other Echinantherini in terms of hemipenial morphology, it shares some external morphology features with its tribal counterparts (see section Discussion). Taeniophallus Cope, 1895
Type species: Lygophis nicagus Cope, 1868, by original designation.
Contents. Taeniophallus brevirostris (Peters, 1863) and T. nicagus (Cope, 1868).
Etymology: From the Greek words ταινíα (taenia [band, fillet]) and ϕαλλóς (phallus [penis]) (Liddell and Scott, 1883; Brown, 1954). The genus name is masculine.
Distribution and Habitat: Species of Taeniophallus occur in Guyana, Suriname, French Guiana and northern Brazil, and toward west in Peru, Ecuador and Colombia (Nogueira et al., 2019; Figure 6F). They inhabit the Guianan Savanna and Amazon Forest, from the sea level to 1500 meters elevation (Myers, 1974; Martins and Oliveira, 1998; Starace, 2015).
Diagnosis: Taeniophallus can be recognize by (1) usually eight supralabials, with the 2nd–3rd or 2nd only touching the loreal and 3rd–5th touching the orbit; (2) 17/17/15 dorsal scale rows usually with one or two apical pits; (3) midventral portion immaculate; (4) hemipenis unilobed or slightly bilobed and unicapitate, with an asulcate interspinal gap interrupted by an enlarged central spine approximately at the midorgan; (5) sulcus spermaticus asymmetrically divided, with shortening of one of the branches (T. brevirostris) or undivided (T. nicagus); (6) a pair of nude pockets delimited by small spines immediately below the capitulum on the asulcate face (7) dorsolateral crests of the parietal bone inconspicuous close the contact with supraoccipital (Figures 5B,C, 7F and Table 1).
Synapomorphies: (1) 17/17/15 dorsal scale rows, (2) dorsal scales usually with one or two apical pits; (3) sulcus spermaticus asymmetrically divided, with shortening of one of the branches (T. brevirostris) or undivided (T. nicagus).
Remarks: This genus corresponds to the brevirostris group of Taeniophallus (sensu Schargel et al., 2005). Although the type species is T. nicagus, the description of the genus was based on the morphology of the hemipenis of T. brevirostris (Myers, 1974).
Although our taxonomic proposal is based on compelling morphological and phylogenetic evidence, one might argue that would be more desirable just erecting a new name for Echinanthera amoena and transferring the Taeniophalus affinis group to the genus Echinanthera to retrieve a monophyletic classification for the tribe. Or, in an even more parsimonious way, just transferring the T. affinis group to Echinanthera and keeping E. amoena in this genus as well. However, we claim that taxonomic ranks (in our case, genera) must contain meaningful taxonomic information in order to facilitate communication, serving as proxies for our understanding about the diversity of forms and evolutionary histories included in the tree of life. By splitting the traditional taxonomic concepts of Echinanthera and Taeniophallus (see next item below) into four different genera, we are approaching this goal by reflecting the morphological, phylogenetic, and biogeographic patterns through the taxonomic classification of the tribe. These four genera represent: (1) four subclades/lineages highly supported by our molecular phylogenetic analysis (Figure 2); (2) four cohesive morphological groups that share several phenotypic traits (e.g., color, scutellation, hemipenial and cranial morphology; see section “Morphological comparison”); and (3) four evolutionary lineages that indicate specific biogeographic events related to their diversification in the South America (see section “Patterns of geographical distribution”).
Although several molecular evaluations have been published including species of Echinantherini (Figure 1), the taxonomic scheme for the tribe follows classical morphological studies. Myers (1974) reviewed the large and heterogeneous Neotropical genus Rhadinaea, dividing it in eight species groups, including the strictly South American “brevirostris group”. Di-Bernardo (1992) suggested the revalidation of the genus Echinanthera Cope, 1894 for Enicognathus amoenus Jan 1863, Natrix melanostigma Wagler 1824, and Coluber undulatus Wied 1824, and for the Rhadinaea brevirostris group (sensu Myers, 1974). By doing so, Di-Bernardo (1992) demonstrated that these species were more closely related to each other than to any other dipsadid, a concept that prevails until today. Myers and Cadle (1994) failed to recognize the innovative aspects of Di-Bernardo’s (1992) contribution, criticizing the latter for not providing strong evidence for the monophyly of his expanded Echinanthera. Myers and Cadle (1994) then further divided Echinanthera by resurrecting Taeniophallus Cope, 1895 for the species originally allocated in the brevirostris group. The apparent paraphyly of Taeniophallus (sensu Myers and Cadle, 1994) with respect to Echinanthera was commented by Zaher (1999), who agreed with Di-Bernardo’s (1992) broader conclusions and suggested a more thorough review of the arguments.
Schargel et al. (2005) acknowledged the arguments of Myers and Cadle (1994) and decided to recognize three species groups within Taeniophallus: the brevirostris group (including T. brevirostris and T. nicagus), the occipitalis group (with only T. occipitalis), and the affinis group (including T. affinis, T. bilineatus, T. persimilis, and T. poecilopogon). Their hemipenial review and arrangement of Taeniophallus served as the basis for the description of Taeniophallus nebularis, a rare species known only from the holotype and allocated tentatively in this genus by the authors (Schargel et al., 2005, p. 1).
Schargel et al. (2005) considered Echinanthera as comprising the six remaining species of Echinantherini (i.e., E. amoena, E. cephalomaculata, E. cephalostriata, E. cyanopleura, E. melanostigma, and E. undulata). According to these authors, monophyly of Echinanthera was supposedly supported by the putative morphological synapomorphy of a large number of prediastemal maxillary teeth (Schargel et al., 2005, p. 18). An undulating middorsal stripe pattern and ventral crossbanding were also mentioned as possible additional synapomorphies for the group.
By including almost all species of Echinantherini in our phylogenetic analysis, it was finally possible to shed light on the longstanding problems about the relationship among the tribe’s diversity. The tree topology, exclusively derived from the molecular evidence, is different from all phylogenetic hypotheses published so far (Figures 1, 2).
The monotypic genus Sordellina has been found as the sister group of Taeniophallus (Grazziotin et al., 2012), or as the sister group of all other Echinantherini (Pyron et al., 2013, 2015; Figueroa et al., 2016; Zaher et al., 2019). Our results, on the other hand, strongly support Sordellina as a member of a clade composed by Amnisiophis, Echinanthera, and Dibernardia. Although the position of Sordellina in clade C2 is strongly supported, our tree indicates that the clade containing Amnisiophis, Dibernardia, and Echinanthera received low bootstrap values, suggesting that the relationship between Sordellina, Amnisiophis and the clade composed by Dibernardia and Echinanthera may be unstable, pending future analysis with more data.
Echinanthera and Taeniophallus have been found mostly as monophyletic groups (Zaher et al., 2009, 2019; Vidal et al., 2010; Grazziotin et al., 2012; Figueroa et al., 2016), except for Pyron et al. (2013) who recovered both genera as paraphyletic with respect to each other. Our analyzes also provide strong evidence regarding the paraphyletic condition of both, Echinanthera and Taeniophallus, and furnishing new grounds for the interpretations of the morphological diversity of these genera. This result partially corroborates the hypotheses raised by Schargel et al. (2005), regarding the paraphyly of Taeniophallus (sensu lato) and the closest affinity between the occipitalis and brevirostris species groups.
Although our phylogenetic framework was the most comprehensive within Echinantherini, we were unable to test the phylogenetic position of E. cephalomaculata. This species was described in 1994 based on two specimens from Reserva Biológica de Pedra Talhada, northeastern Brazil (Di-Bernardo, 1994). A third specimen was collected just some years later in the same place (Roberto et al., 2015). Recently, Freitas et al. (2019) expanded the geographic range of this species in about 150 km northeast from its type locality, recording three specimens that were not collected. The general pattern of E. cephalomaculata fits the morphological diagnosis of Echinanthera described by Schargel et al. (2005) and by us. Although Schargel et al. (2005) have raised some concerns about the position of E. cephalomaculata within Echinanthera, based on the apparent absence of bands at the base of the ventral scales in E. cephalomaculata, the existence of individuals who clearly show this characteristic (A. Abegg, personal observation) gives support to our taxonomic scheme. Nevertheless, this provisory arrangement for E. cephalomaculata is pending future phylogenetic tests.
Although we did not implement a fully integrative approach (e.g., total evidence phylogenetic analysis, sensu Kluge, 1989), the morphological comparison based on our phylogenetic framework provides grounds for inference and discussion about the evolution of some major traits within Echinantherini. Below, we comment on some of these traits, and we provide a complete description of morphological differences among genera in the Supplementary Appendix 7.
Considering the similarities formerly used to include Adelphostigma, Dibernardia, and Taeniophallus in the same genus (Taeniophallus sensu lato), Schargel et al. (2005) have already suggested that despite the conspicuous pale canthal line shared by A. occipitalis and D. poecilopogon, the species of Adelphostigma (the occiptalis group) are more closely related to Taeniophallus (the brevirostris group) than to Dibernardia (the affinis group). More specifically, Myers (1974) previously observed that A. occipitalis and T. brevirostris share occasional hemipenis bilobation, spinulated calyces, asulcate wall of the capitulum forming single or double folds, and thick and knoblike spinules on the asulcate side of the capitulum in retracted hemipenis, and that the pocket observed in the lower edge of the capitulum of T. brevirostris could be homologous to the deep capitulate overhang of A. occipitalis (Schargel et al., 2005). Our analysis gives support to this hypothesis, since Adelphostigma and Taeniophallus are sister groups.
In addition to the characteristics commented by Schargel et al. (2005), we observed that T. brevirostris and T. nicagus share with the species of Adelphostigma the presence of an enlarged hemipenial spine approximately at the middle of the organ, and a reduction in the number of dorsal scales near the cloaca. These traits support the relationship and uniqueness of both genera within Echinantherini.
Myers and Cadle (1994) indicated the absence of an asulcate interspinal gap in the hemipenis as a unique trait common only to T. brevirostris and T. nicagus (Schargel et al., 2005; Myers, 2011). Regarding the topology of our tree and the analysis of the morphological variation, we consider the distribution of this trait particularly puzzling, because all Echinantherini but Sordellina present a clear asulcate interspinal gap in the hemipenis. We disagree with Myers and Cadle (1994) and we provide a reinterpretation of this hemipenial trait.
Myers and Cadle (1994) defined the asulcate interspinal gap as “… the arrangement of spines plus differential tissue expansion that permits a characteristic interspinal asulcal gap to appear upon eversion” and they emphasize that this trait is particularly pronounced in Echinanthera and absent in T. brevirostris and T. nicagus. We acknowledge that the asulcate face of the hemipenis of both T. brevirostris and T. nicagus bears relatively small spines, although there is a clear row of enlarged, vertically oriented spines arranged on each side of the organ of these two species delimiting an asulcate gap. We consider such gap homologous to the interspinal asulcal gap as defined by Myers and Cadle (1994). Moreover, the small, sparsely distributed spines visible in T. nicagus (Figure 5B), are also present in Dibernardia and Echinanthera, although especially distributed in the proximal region of the hemipenis (Figures 5I,K–M).
In spite of the similarities shared between Adelphostigma and Taeniophallus, they also depart remarkably from each other in several traits. Among them, the most noteworthy certainly refers to the asymmetry of the spermatic branches in Taeniophallus (presumably derived from the single condition in T. nicagus), which is, as far as we known, an unique condition among dipsadids (Supplementary Appendix 5; Zaher, 1999; Schargel et al., 2005). Still in terms of hemipenial morphology, while Adelphostigma presents a deep and proximally open nude pocket immediately below the capitulum on the asulcate side, the putative geographical homologous structure in Taeniophallus corresponds to a pair of nude pockets proximally surrounded by rows of spinulate papillae. Other general distinguishing features between them include the unique dorsal scale pattern of each genus within Echinantherini (15/15/15 or 15/15/13 in Adelphostigma and 17/17/15 in Taeniophallus), the absence of apical pits in Adelphostigma (present in Taeniophallus), the conspicuous pale canthal line in Adelphostigma (absent in Taeniophallus) and the unique middorsal pattern composed of dark-brown blotches in the anterior region that are gradually replaced by paired spots toward the tail in Adelphostigma, versus a straight or undulating dark-brown stripes ornamenting the middorsal region in Taeniophallus.
The allocation of Sordellina in Echinantherini has been a matter of debate (Grazziotin et al., 2012) and the interpretation of its relationship based only on morphological evidence has been problematic (Miranda and Fernandes, 2012). The hemipenial similarities shared between Sordellina and other species of the tribe are not informative because they are also present in different lineages within Dipsadidae (Zaher, 1999). On the other hand, the differences are easily recognizable. The asulcate side of the hemipenis lacks any structure comparable to the typical asulcate interspinal gap, since the hemipenial body is covered by spines of similar sizes from the proximal region to the base of the capitulum. Additionally, the capitulum possesses several rows of papillae with little or no tissue interconnection (vestigial calyces; Zaher, 1999).
Although the hemipenial traits observed in Sordellina are unique within Echinantherini, our examination of external characters revealed unexpected similarities between Sordellina and its tribal counterparts. Some specimens of Sordellina, specifically the juveniles, present a pair of light spots on the parietal scales (Figure 4F, Miranda and Fernandes, 2012), a characteristic frequently present in all other species of the tribe. Additionally, some individuals have light yellow spots or dashes in the 4th dorsal scale rows (Figure 4F), a generalized pattern for Echinantherini (presumably changed to spots in the 3rd row in Amnisiophis). Finally, Sordellina has no reduction in the number of dorsal scales, arranged in 17/17/17 rows. Although this feature is present in other lineages of Dipsadidae, within Echinantherini it is only shared with Amnisiophis, Dibernardia and Echinanthera, which forms with Sordellina the clade C2 in our phylogenetic tree (Figure 2 and Supplementary Appendix 5). The reevaluation of this morphological traits represents a significant advance in the understanding of the evolution of Sordellina within Echinantherini.
The phylogenetic relationship between Amnisiophis, Dibernardia, and Echinanthera is supported by at least one morphological trait, the asulcate interspinal gap extending from the distal to the proximal region of the organ, bearing small papillae on its midline. Moreover, Dibernardia and Echinanthera are similar considering the general pattern of hemipenial morphology. These two genera share a unilobed and semicapitate organ, with an interspinal asulcate gap that usually ends proximally in an enlarged and laterally positioned spine (Figure 8 and Supplementary Appendix 5; Di-Bernardo, 1992; Zaher, 1999). Although similar, these genera present several unique characteristics that were used here to define them within clade C2.

Figure 8. Comparative cranial osteology among Echinantherini. (A) Amnisiophis amoenus (IBSP 88359); (B) Echinanthera cephalostriata, (IBSP 84971); (C) Echinanthera cyanopleura (IBSP 81751); (D) Echinanthera melanostigma (IBSP 56933); and (E) Echinanthera undulata (IBSP 22904). Colored bones: blue = parietal; green = supratemporal. Scale bars: 2 mm.
Dibernardia presents the unique derived condition of an asulcate interspinal gap that ends distally at an abruptly cluster of flaplike enlarged papillae, whereas in Echinanthera and Amnisiophis this gap ends distally at a homogeneous group of small to medium-sized papillae (Figure 5 and Supplementary Appendix 5; Schargel et al., 2005). Another well-known diagnostic character of Dibernardia is the number and scutellation of the supralabials, which are consistently present as seven scales, with the third and fourth touching the orbit and only the second touching the loreal (Myers, 1974; Di-Bernardo and Lema, 1986, 1987, 1988, 1990; Schargel et al., 2005). Other general differences of Dibernardia with respect to Echinanthera and Amnisiophis include the reduced number of maxillary, palatine, pterygoid and dentary teeth, the absence of dorsal apical pits, the immaculate medioventral portion of the body, and the inconspicuousness of parabasisfenoid crests (Figures 3, 4, 7, 8; Schargel et al., 2005).
Similarly, Echinanthera (sensu lato) has been considered a well-defined genus, supported by the unique derived condition of a large number of maxillary teeth (24–35) (Schargel et al., 2005). The undulating middorsal stripe and ventral transverse bands (specially in adults) have also been speculated as potential synapomorphies for this genus (Schargel et al., 2005). However, with the description of Amnisiophis, both genera share the unusual high number of teeth (not just maxillaries, but also palatines, pterygoids and dentaries, see Table 1), as well as the presence of ventral transverse bands (Figure 4). On the other hand, although sometimes also observable in Taeniophallus, the undulating middorsal stripe was recovered as a synapomorphy for Echinanthera (Supplementary Appendix 5). Therefore, as redefined in this study, the genus Echinanthera is unambiguously supported by molecular evidence, easily diagnosed by the combination of morphological traits (Table 1) and supported by at least one unambiguous, non-private morphological synapomorphy.
On the other hand, the newly erected Amnisiophis presents several distinctiveness considering both morphology and natural history aspects. The dorsal coloration of Amnisiophis is unique in presenting a green pattern on the head and the anterior region of the body, with a line of light spots on the 3rd dorsal scale row along the body (Figures 2, 3 and Supplementary Appendix 5). Additionally, it differs from all other Echinantherini in the use of the microhabitat, since Amnisiophis lives strictly associated with lotic water bodies in the interior of forests (Azevedo et al., 2018), while its close relatives are mostly leaf-litter inhabitants or open area dwellers. Sordellina punctata represents another species historically associated with aquatic microhabitats, but differently from Amnisiophis it lives in wetlands and other soak soils surrounding lakes or rivers (Pereira et al., 2007). Another autapomorphy for Amnisiophis is the conspicuousness and abruptly tapering of the parietal dorsolateral crests in the region close to the contact with the supraoccipital bone (Supplementary Appendices 4, 5). Finally, the enlarged supratemporals, which usually contacts the parietal-prootic suture are a unique condition within Echinantherini (Figure 8 and see also Appendices 4, 5).
Our morphological comparison based on a phylogenetic framework provides a substantial advance in the understanding of the trait evolution of Echinantherini. However, to obtain the complete scenario of morphological changes within the tribe, a proper integrative approach through an analysis of total evidence is necessary. By implementing such approach, it will be also possible to test the phylogenetic positioning of E. cephalomaculata, for which we only obtained morphological data.
Our results strongly indicate that the phylogenetic diversity within Echinantherini is geographically structured (Figures 2, 6). Although presenting species widely distributed throughout South America, from the Amazon rainforest in northern Suriname to the province of Córdoba in central Argentina (Cacivio et al., 1999; Schargel et al., 2005), our phylogenetic tree suggest two main geographical patterns of distribution that are related to clades C1 and C2.
Clade C1— Adelphostigma and Taeniophallus—is composed by species occurring mainly in the Amazon rainforest. Except for A. occipitalis, the four remaining species within this clade are endemic to the Amazonian region (Figures 6A,F). This pattern may suggest that the evolution of the clade is linked to the biogeographical processes that shape the Amazon rainforest. On the other hand, the wide geographic range of A. occipitalis is particularly puzzling and may reflect the evolutionary history of a more generalist species. Adelphostigma occipitalis is currently distributed in very distinct habitats from the Amazonia to the Patagonian grasslands (Figure 6A). The broad morphological diversity of A. occipitalis also reinforces this idea, by linking a generalist species with a very variable phenotype. Moreover, such morphological variation may indicate that A. occipitalis represents a species complex (Santos-Jr, 2009).
Clade C2 encompasses four genera mostly distributed throughout the Brazilian Atlantic Forest (BAF; Figures 2, 6). The monotypic genera Amnisiophis and Sordellina present similar geographic ranges and are endemic to the southern and southeastern portion of BAF. While Amnisiophis occurs exclusively in elevated areas (>500 asl) and also in relatively less forested areas (e.g., Araucaria moist forests), Sordellina inhabits both highland and lowland (at sea level) and seems to be restricted to the coastal forests of Serra do Mar.
Dibernardia and Echinanthera present a relatively wider range in the BAF (Figure 6). Both genera have a similar geographic pattern, ranging from the subtropical Uruguayan savannah throughout the entire tropical BAF. These genera also occur in highland relictual forests known as “brejos de altitude” in the semi-arid Caatinga of northeastern Brazil. Although all species in clade C2 can be considered forest dwellers, D. poecilopogon is an exception, inhabiting only the open areas of southern Brazil, Uruguay and Argentina. Such range also indicates a basal dichotomy within Dibernardia, since D. poecilopogon is the sister group of all other species of the genus restricted to the BAF.
The strong signal of allopatric distribution within the different clades of Echinantherini suggests a clear connection between the cladogenic events that shaped the phylogenetic diversity of the tribe and the formation of ecosystems and ecoregions in cis-Andean South America. These links between geographical and phylogenetic patterns suggest that Echinantherini is an ideal model for studies on the biogeography of South American snakes.
Here we evaluate for the first time the phylogenetic position of the rare Echinanthera amoena within the most comprehensive phylogenetic framework of the tribe Echinantherini. We based our analysis on six gene fragments, including 14 of the 16 species described for the tribe, half of them sequenced for the first time. By increasing taxon sampling, it was finally possible to test more accurately the relationship between the constituents of Echinantherini, a subject that has been contradictory in recent studies that have addressed the systematics of Neotropical snakes.
Our maximum likelihood analysis retrieved E. amoena as a unique lineage, phylogenetically positioned apart from all other congeners. Besides Echinanthera our results also retrieved Taeniophallus as paraphyletic, since the T. affinis species group was positioned as sister to Echinanthera (except E. amoena) clustering apart from the clade formed by the T. brevirostris and T. occipitalis groups. In brief the retrieved relationship can be summarized as ((T. brevirostris group, T. occipitalis group), (Sordellina, (E. amoena, (Echinanthera spp., T. affinis group)))). To better reflect the phylogenetic relationship and the amount of morphological variation within Echinantherini, we provide a new taxonomic rearrangement composed of six genera. We described the new genera Dibernardia and Adelphostigma for the former T. affinis and T. occipitalis species groups, respectively, and the monospecific genus Amnisiophis for E. amoena, in addition to redefining Echinanthera and Taeniophallus. Although we did not evaluate the phylogenetic position of T. nebularis, we described the new genus Myersinia to accommodate it and we removed it from Echinantherini since its morphology remarkably departs from all species included in the tribe.
Even though we could not include all the species in our molecular phylogenetic analysis, the classification provided herein encompassed the major lineages, and was complemented by the discussion of distinct aspects regarding the morphological diversity of Echinantherini. Furthermore, the phylogenetic arrangement found for the tribe presents a high level of geographical concordance, which suggests a common scenario shaping the ecoregions of cis-Andean South America and the evolutionary history of Echinantherini.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: doi: 10.6084/m9.figshare.15116877 and the sequences are deposited at GenBank under the accessions: MZ746963–MZ746995, MZ746747–MZ746777, MZ768815–MZ768849, MZ733987–MZ734020, MZ734021–MZ734049, and MZ734050–MZ734078.
The animal study was reviewed and approved by CEUAIB 4479020217.
AA, SB, HZ, and FG conceived the research project. AA, WA, PP, JB, RG, and FV gathered the molecular data. AA, WA, PP, AS, and HC gathered the morphological data. AA and FG performed the analyses. AA, HC, HZ, and FG wrote the manuscript. NF, CC, and SB contributed intellectually and discussed the text. SB, HZ, and FG funded the research project. All authors revised the text.
Funding for this study was provided to HZ by Fundação de Amparo à Pesquisa do Estado de São Paulo (BIOTA/FAPESP grant numbers 2002/13602-4 and 2011/50206-9); FG was supported by scholarships from FAPESP (numbers 2007/52781-5 and 2016/50127-5); and AA received a Master’s fellowship from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (130115/2019–3).
We thank all curators and technicians for allowing access to the herpetological collections under their care. We are grateful to Cladinara Sarturi for her laboratory assistance during acquisition of molecular data. We thank Adnan Farhat, Valquiria O. Pereira, and Paulo R. Machado-Filho for their help with figures edition. We are grateful to Glauco Oliveira, Daniel Loebmann, Omar M. Entiauspe-Neto, Renato Gaiga, Wolfgang Wüster, Mayke de Freitas, Gilson Rivas, Ivo Ghizoni-Jr, Rafael de Fraga, Rodrigo Tavares-Pinheiro, Carlos E. Costa-Campos, and the Projeto Dacnis research team for providing photographs of live specimens of Echinantherini species. We are indebted to Alberto B. Carvalho for scanning specimens at the Laboratório de Microtomografia and providing 3-D reconstructions of the raw data. We also thank Pedro M. S. Nunes for providing images of the hemipenis of S. punctata; Jairo Mendoza-Roldan for reviewing the Italian text.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.969263/full#supplementary-material
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Arredondo, J. C., Grazziotin, F. G., Scrocchi, G. J., Rodrigues, M. T., Bonatto, S. L., and Zaher, H. (2020). Molecular phylogeny of the tribe Philodryadini Cope, 1886 (Dipsadidae: Xenodontinae): rediscovering the diversity of the South American Racers. Papéis Avulsos Zool. 60:e20206053. doi: 10.11606/1807-0205/2020.60.53
Azevedo, W. S., Amorim, L. G. S., Menezes, F. A., and Abegg, A. D. (2018). Natural history notes and geographic distribution of the poorly known Echinanthera amoena (Serpentes: Dipsadidae). Herpetol. Notes 11, 925–928.
Bergsten, J. (2005). A review of long-branch attraction. Cladistics 21, 163–193. doi: 10.1111/j.1096-0031.2005.00059.x
Bérnils, R. S. (2009). Composição e Padrões de Distribuição de Caenophidia (Squamata, Serpentes) das Serras Atlânticas e Planaltos do Sudeste da América do Sul. doctorate’s thesis. Rio de Janeiro: Universidade Federal do Rio de Janeiro.
Boakes, E. H., McGowan, P. J., Fuller, R. A., Chang-Qing, D., Clark, N. E., O’Connor, K., et al. (2010). Distorted views of biodiversity: spatial and temporal bias in species occurrence data. PLoS Biol. 8:e1000385. doi: 10.1371/journal.pbio.1000385
Cacivio, P., Scrocchi, G., and Giraudo, A. R. (1999). Echinantera occipitalis (Serpentes, Colubridae) nuevos datos de escamación, hemipenes y distribuición. Cuad. Herpetol. 13, 81–87.
Cadle, J. E., and Greene, H. W. (1993). “Phylogenetic patterns, biogeography, and the ecological structure of Neotropical snake assemblages,” in Species Diversity in Ecological Communities: Historical and Geographical Perspectives, eds R. E. Ricklefs and R. Schluter (Chicago, IL: University of Chicago Press).
Carreira, S., Meneghel, M., and Achaval, F. (2005). Reptiles de Uruguay. Montevideo: Universidad de la República.
Cassimiro, J., Teixeira, M. Jr., Recoder, R. S., and Rodrigues, M. T. U. (2013). Echinanthera amoena (Corredeira-do-mato). Herpetol. Rev. 44:625.
Correa, C., Vásquez, D., Castro-Carrasco, C., Zúñiga-Reinoso, Á, Ortiz, J. C., and Palma, R. E. (2017). Species delimitation in frogs from South American temperate forests: the case of Eupsophus, a taxonomically complex genus with high phenotypic variation. PLoS One 12:e0181026. doi: 10.1371/journal.pone.0181026
Cundall, D., and Irish, F. (2008). “The snake skull,” in Biology of Reptilia vol. 20: The Skull of Lepidosauria, eds C. Gans, A. S. Gaunt, and K. Adler (Washington: Society for the Study of Amphibians and Reptiles Press), 349–692.
Di-Bernardo, M. (1991). Estudo Revisivo de Natrix melanostigma Wagler, 1824, com a revalidação de Echinanthera Cope, 1894 e análise cladística dos táxons afins (Serpentes, Colubridae, Xenodontinae). [dissertation/master’s thesis]. Porto Alegre: Pontifícia Universidade Católica do Rio Grande do Sul.
Di-Bernardo, M. (1992). Revalidation of the genus Echinanthera Cope, 1894, and its conceptual amplification (Serpentes, Colubridae). Comunicações do Museu de Ciências e Tecnologia da PUCRS 5, 225–257.
Di-Bernardo, M. (1994). Uma nova espécie de Echinanthera Cope, 1894 (Serpentes, Colubridae) do nordeste do Brasil. Biociências 2, 75–81.
Di-Bernardo, M. (1996). A new species of the Neotropical snake genus Echinanthera Cope, 1894 from southeastern Brazil (Serpentes, Colubridae). Snake 27, 120–126.
Di-Bernardo, M., and Lema, T. (1986). O gênero Rhadinaea Cope, 1863, no Brasil meridional. II [sic]. Rhadinaea persimilis (Cope, 1869) (Serpentes, Colubridae). Acta Biol. Leopoldensia 8, 101–122.
Di-Bernardo, M., and Lema, T. (1987). O gênero Rhadinaea Cope, 1863, no Brasil meridional. I. Rhadinaea poecilopogon Cope, 1863 (Serpentes, Colubridae). Acta Biol. Leopoldensia 9, 203–224.
Di-Bernardo, M., and Lema, T. (1988). O gênero Rhadinaea Cope, 1863, no Brasil meridional. III. Rhadinaea affinis (Günther, 1858) (Serpentes, Colubridae). Acta Biol. Leopoldensia 10, 223–252.
Di-Bernardo, M., and Lema, T. (1990). O gênero Rhadinaea Cope, 1863, no Brasil meridional. IV. Rhadinaea bilineata (Fischer, 1885) (Serpentes, Colubridae). Acta Biol. Leopoldensia 12, 359–392.
Dowling, H. G. (1951). A proposed standard system of counting ventrals in snakes. J. Herpetol. 1, 97–99.
Felsenstein, J. (1978). Cases in which parsimony or compatibility methods will be positively misleading. Syst. Zool. 27, 401–410. doi: 10.1093/SYSBIO/27.4.401
Figueroa, A., McKelvy, A. D., Grismer, L. L., Bell, C. D., and Lailvaux, S. P. (2016). A species-level phylogeny of extant snakes with description of a new colubrid subfamily and genus. PLoS One 11:e0161070. doi: 10.1371/journal.pone.0161070
Freitas, M. A., Barbosa, G. G., Bernardino, K. P., Pinheiro Filho, J. D., and Abegg, A. D. (2019). First records of the rare snake Echinanthera cephalomaculata Di-Bernardo, 1994 in the state of Pernambuco, Brazil (Serpentes: Dipsadidae). Herpetol. Notes 12, 1005–1009.
Goloboff, P., and Catalano, S. (2016). TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238. doi: 10.1111/cla.12160
Grazziotin, F. G., Zaher, H., Murphy, R. W., Scrocchi, G., Benavides, M. A., Zhang, Y. P., et al. (2012). Molecular phylogeny of the New World Dipsadidae (Serpentes: Colubroidea): a reappraisal. Cladistics 28, 437–459. doi: 10.1111/j.1096-0031.2012.00393.x
Heath, T. A., Hedtke, S. M., and Hillis, D. M. (2008). Taxon sampling and the accuracy of phylogenetic analyses. J. Syst. Evol. 3, 239–257. doi: 10.3724/SP.J.1002.2008.08016
Hoge, A. R., and Romano, S. (1978). Redescription and range of Sordellina punctata (Peters) (Serpentes: Colubridae). Mem. Instit. Butantan 40, 63–70.
Joly, S., McLenachan, P. A., and Lockhart, P. J. (2009). A statistical approach for distinguishing hybridization and incomplete lineage sorting. Am. Nat. 174, 54–70. doi: 10.1086/600082
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., and Sturrock, S. (2012). Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kluge, A. G. (1989). A concern for evidence and a phylogenetic hypothesis of relationships among Epicrates (Boidae, Serpentes). Syst. Biol. 38, 7–25. doi: 10.1093/sysbio/38.1.7
Lanfear, R., Calcott, B., Ho, S. Y. W., and Guindon, S. (2012). PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 29, 1695–1701. doi: 10.1093/molbev/mss020
Liddell, H. G., and Scott, R. (1883). Greek-English Lexicon, Seventh Edn. New York, NY: Harper and Brothers.
Maddison, W. P., and Maddison, D. R. (2009). Mesquite: A Modular System for Evolutionary Analysis, Version 2.7.1. Available online at: https://mesquiteproject.org/ (accessed August 26, 2022).
Martins, M., and Oliveira, M. E. (1998). Natural history of snakes in forests of the Manaus region, central Amazonia, Brazil. Herpetol. Nat. Hist. 6, 78–150.
Menezes, F. A., Abegg, A. D., Silva, B. R., Franco, F. L., and Feio, R. N. (2018). Composition and natural history of the snakes from the Parque Estadual da serra do papagaio, southern minas gerais, serra da mantiqueira, Brazil. ZooKeys 797, 117–160. doi: 10.3897/zookeys.797.24549
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceedings of the Gateway Computing Environments Workshop, New Orleans, 1–8.
Mindell, D. P. (2013). The tree of life: metaphor, model, and heuristic device. Syst. Biol. 62, 479–489. doi: 10.1093/sysbio/sys115
Miranda, B. H. V., and Fernandes, D. S. (2012). Morphological variation of Sordellina punctata (Peters, 1880) with notes on the phylogenetic position of the genus within the family Dipsadidae (Serpentes). Int. J. Zool. 2012, 1–10. doi: 10.1155/2012/827820
Muscat, E., and Moroti, M. (2020). Filling gaps: new record of Echinanthera amoena (Jan, 1863) for the Serra da Mantiqueira, state of São Paulo, Brazil. Herpetol. Notes 13, 429–431.
Myers, C. W. (1974). The systematics of Rhadinaea (Colubridae), a genus of new world snakes. Bull. Am. Museum Nat. Hist. 153, 1–272.
Myers, C. W. (2011). A new genus and new tribe for Enicognathus melanauchen Jan, 1863, a neglected South American snake (Colubridae: Xenodontinae), with taxonomic notes on some Dipsadinae. Am. Museum Novitates 3715, 1–34.
Myers, C. W., and Cadle, J. E. (1994). A new genus for South American snakes related to Rhadinaea obtusa Cope (Colubridae) and resurrection of Taeniophallus Cope for the “Rhadinaea” brevirostris group. Am. Museum Novitates 3102, 1–33.
Nixon, K. C., and Carpenter, J. M. (2012). On homology. Cladistics 28, 160–169. doi: 10.1111/j.1096-0031.2011.00371.x
Nogueira, C. C., Argôlo, A. J., Arzamendia, V., Azevedo, J. A., Barbo, F. E., Bérnils, R. S., et al. (2019). Atlas of Brazilian snakes: verified point-locality maps to mitigate the Wallacean shortfall in a megadiverse snake fauna. South Am. J. Herpetol. 14, 1–274. doi: 10.2994/SAJH-D-19-00120
Pereira, D. N., Stender-Oliveira, F., Rodrigues, M. G., and Bérnils, R. S. (2007). Distribution and habitat use of Sordellina punctata (Serpentes, Colubridae), with a new record from State of São Paulo, Brazil. Herpetol. Bull. 100, 18–22.
Pesantes, O. S. (1994). A method for preparing the hemipenis of preserved snakes. J. Herpetol. 28, 93–95. doi: 10.2307/1564686
Pol, D., and Siddall, M. E. (2001). Biases in maximum likelihood and parsimony: a simulation approach to a 10-taxon case. Cladistics 17, 266–281. doi: 10.1006/clad.2001.0172
Procter, J. B. (1923). On a new genus and species of colubrine snake from SE Brazil. J. Nat. Hist. 11, 227–230.
Pyron, R. A., Burbrink, F. T., Colli, G. R., Montes de Oca, A. N., Vitt, L. J., Kuczynski, C. A., et al. (2011). The phylogeny of advanced snakes (Colubroidea), with discovery of a new subfamily and comparison of support methods for likelihood trees. Mol. Phylogenet. Evol. 58, 329–342. doi: 10.1016/j.ympev.2010.11.006
Pyron, R. A., Burbrink, F. T., and Wiens, J. J. (2013). A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 3:93. doi: 10.1186/1471-2148-13-93
Pyron, R. A., Guayasamin, J. M., Peñafiel, N., Bustamante, L., and Arteaga, A. (2015). Systematics of Nothopsini (Serpentes, Dipsadidae), with a new species of Synophis from the Pacific Andean slopes of southwestern Ecuador. ZooKeys 541, 109–147.
R Core Team (2015). R: A Language and Environment for Statistical Computing. Version 3.0.1. Available from: http://www.r-project.org/ (accessed October 07, 2016).
Roberto, I. J., Melgarejo, A., and Ávila, R. W. (2015). Répteis (Testudines, Squamata, Crocodylia) da Reserva Biológica de Pedra Talhada. Biodiversidade da Reserva Biológica de Pedra Talhada (Alagoas-Pernambuco, Brasil). Boissiera 68, 357–375.
Santos-Jr, A. F., Di-Bernardo, M., and Lema, T. (2008). New species of the Taeniophallus occipitalis group (Serpentes, Colubridae) from eastern Amazonia, Brazil. J. Herpetol. 42, 419–426.
Santos-Jr, A. P. (2009). Revisão Taxonômica Do Grupo de Taeniophallus occipitalis e o Relacionamento Filogenético da Tribo Echinantherini (Serpentes, Dipsadidae, Xenodontinae). doctorate’s thesis. Porto Alegre: Pontifícia Universidade Católica do Rio Grande do Sul.
Schargel, W. E., Rivas, G. F., and Myers, C. W. (2005). An enigmatic new snake from cloud forest of the Península de Paria, Venezuela (Colubridae: Genus Taeniophallus?). Am. Museum Novitates 3484, 1–24.
Siddall, M. E., and Whiting, M. F. (1999). Long-branch abstractions. Cladistics 15, 9–24. doi: 10.1111/j.1096-0031.1999.tb00391.x
Simon, M. N., Machado, F. A., and Marroig, G. (2016). High evolutionary constraints limited adaptive responses to past climate changes in toad skulls. Proc. R. Soc. B 283:20161783. doi: 10.1098/rspb.2016.1783
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Tonini, J. F. R., Beard, K. H., Ferreira, R. B., Jetz, W., and Pyron, R. A. (2016). Fully-sampled phylogenies of squamates reveal evolutionary patterns in threat status. Biol. Conserv. 204, 23–31. doi: 10.1016/j.biocon.2016.03.039
Trevine, V. C., Grazziotin, F. G., Giraudo, A., Sallesbery–Pinchera, N., Vianna, J. A., and Zaher, H. (2022). The systematics of Tachymenini (Serpentes, Dipsadidae): An updated classification based on molecular and morphological evidence. Zoologica Scripta. doi: 10.1111/zsc.12565
Vidal, N., Dewynter, M., and Gower, D. J. (2010). Dissecting the major American snake radiation: a molecular phylogeny of the Dipsadidae Bonaparte (Serpentes, Caenophidia). Comptes Rendus Biol. 333, 48–55. doi: 10.1016/j.crvi.2009.11.003
Werneck, F. P. (2011). The diversification of eastern South American open vegetation biomes: historical biogeography and perspectives. Quatern. Sci. Rev. 30, 1630–1648. doi: 10.1016/j.quascirev.2011.03.009
Zaher, H. (1999). Hemipenial morphology of the South American xenodontine snakes, with a proposal for a monophyletic Xenodontinae and a reappraisal of colubroid hemipenes. Bull. Am. Museum Nat. Hist. 240, 1–170.
Zaher, H., Grazziotin, F. G., Cadle, J. E., Murphy, R. W., Moura-Leite, J. C., and Bonatto, S. L. (2009). Molecular phylogeny of advanced snakes (Serpentes, Caenophidia) with an emphasis on South American xenodontines: a revised classification and descriptions of new taxa. Papéis Avulsos Zool. 49, 115–153. doi: 10.1590/S0031-10492009001100001
Zaher, H., Murphy, R. W., Arredondo, J. C., Graboski, R., Machado-Filho, P. R., Mahlow, K., et al. (2019). Large-scale molecular phylogeny, morphology, divergence-time estimation, and the fossil record of advanced caenophidian snakes (Squamata: Serpentes). PLoS One 14:e0216148. doi: 10.1371/journal.pone.0216148
Zaher, H., and Prudente, A. L. C. (2003). Hemipenes of Siphlophis (Serpentes, Xenodontinae) and techniques of hemipenial preparation in snakes: a response to Dowling. Herpetol. Rev. 34:302.
Zaher, H., Yánez-Muñoz, M. H., Rodrigues, M. T., Graboski, R., Machado, F. A., Altamirano-Benavides, M., et al. (2018). Origin and hidden diversity within the poorly known Galápagos snake radiation (Serpentes: Dipsadidae). Syst. Biodivers. 16, 614–642. doi: 10.1080/14772000.2018.1478910
Keywords: Echinanthera, comparative morphology, molecular systematics, snake taxonomy, Sordellina, Taeniophallus, Xenodontinae
Citation: Abegg AD, Santos AP Jr, Costa HC, Battilana J, Graboski R, Vianna FSL, Azevedo WS, Fagundes NJR, Castille CM, Prado PC, Bonatto SL, Zaher H and Grazziotin FG (2022) Increasing taxon sampling suggests a complete taxonomic rearrangement in Echinantherini (Serpentes: Dipsadidae). Front. Ecol. Evol. 10:969263. doi: 10.3389/fevo.2022.969263
Received: 14 June 2022; Accepted: 20 September 2022;
Published: 20 October 2022.
Edited by:
Diego Baldo, CONICET Institute of Subtropical Biology (IBS), ArgentinaReviewed by:
Santiago Andres Catalano, CCT CONICET Tucuman, ArgentinaCopyright © 2022 Abegg, Santos, Costa, Battilana, Graboski, Vianna, Azevedo, Fagundes, Castille, Prado, Bonatto, Zaher and Grazziotin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Arthur D. Abegg, YXJ0aHVyX2FiZWdnQGhvdG1haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.