 Tian Li
Tian Li Zehao Zhang
Zehao Zhang Jingkuan Sun
Jingkuan Sun Zhanyong Fu
Zhanyong Fu Yinghan Zhao
Yinghan Zhao Wenjing Xu
Wenjing Xu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 24 June 2022
Sec. Conservation and Restoration Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.948682
This article is part of the Research Topic Disturbance, Resilience and Restoration of Wetlands View all 20 articles
The seasonal dynamic characteristics of C, N, and P stoichiometry and water use efficiency (WUE) of Messerschmidia sibirica and the soil in the Yellow River Delta (YRD) were studied. The correlations of stoichiometric characteristics and WUE between organs of M. sibirica and soil were analyzed. The results showed that: (1) the contents of C, N, and P and their stoichiometric ratios in various organs of M. sibirica varied with seasons. The seasonal dynamics of leaf N:P showed that the degree of nutrient restriction by N decreased in July and increased in September. (2) The contents of C, N, and P, as well as their stoichiometric ratio, showed a high correlation throughout the growing season, with N:P showing a significantly positive correlation among organs and the lowest stoichiometric correlation between leaf and root. (3) C13 stable isotope analysis showed that the WUE of M. sibirica in May was significantly higher than that of other months (July and September). The WUE had a significantly positive correlation with leaf C and N content and a significantly negative correlation with leaf C:N, indicating that M. sibirica can compensate for the decline in N use efficiency through the improvement of WUE. The structural equation model (SEM) showed that the leaf N and P contents were affected by the joint effect of season and WUE, and the leaf C content was mainly directly affected by WUE. (4) Redundancy analysis (RDA) analysis showed that soil P content and soil N:P were the main factors affecting the variation of stoichiometry and WUE in various organs of M. sibirica. This study is helpful to deeply understand the adaptive mechanism of plant nutrient and water use, which provides a theoretical basis for vegetation protection and restoration in the study area.
Change in plant stoichiometry not only reveal the nutrient uptake and utilization status of different plants but also indicate the constraint relationship between different nutrients, which is an important indicator to determine whether plants can renew themselves and recycle nutrients (Crous et al., 2019). Changes in plant carbon, nitrogen, and phosphorus stoichiometry are influenced not only by environmental factors and plant physiological processes but also by differences in structure and function in different organs of the same plant (Luo et al., 2021; Yang and Xu, 2021). Observing changes in environmental and plant stoichiometry can help identify nutrient-limiting factors for plant growth and is an important complement to investigating the relationships between ecosystem stoichiometry and plant function and environmental adaptation mechanisms (Cao and Chen, 2017; Tang et al., 2018).
Various factors influence plant stoichiometry, and the soil C: N: P ratio directly reflects the soil nutrient status and can be used indirectly as an indicator of plant nutrient status (Elser et al., 2010). Studying the stoichiometric characteristics of plants and soils and their interrelationships provides insight into their feedback processes in terms of nutrients and improves our understanding of the adaptation mechanisms of plants. However, due to the study scale, scope, plant species, and other reasons, there is no unified research conclusion on whether plant stoichiometry is influenced by soil stoichiometry (Wang et al., 2015). Therefore, the relationship between plant and soil stoichiometry needs to be further explored.
There is a close relationship between water and nutrient use by plants, and plants need to constantly coordinate the balance between nutrient and water use during growth to adapt to environmental influences (Li et al., 2021). In recent years, stable isotope 13C has been widely used to characterize plant water use efficiency (WUE), which provides important technical support for the study of the plant WUE. Researching the relationship between plant water use and plant stoichiometry can enrich the theory of plant water-fertilizer relationship (Yan et al., 2016). However, studies on the relationship between plant nutrient use characteristics, WUE, and soil nutrients are still very limited.
Messerschmidia sibirica is a perennial herb with salt secretion characteristics and strong adaptability. It is a widely grown constructive species in the chenier of the Yellow River Delta (YRD) and plays an important role in species diversity and ecological function. Previous research on M. sibirica mainly focused on the distribution survey of plant communities, the growth and physiological characteristics under salt and sand burial stresses, and diversity of endophytic bacteria (Lee et al., 2007; Suzuki et al., 2011; Xie et al., 2015; Zhang et al., 2019a). The ecological stoichiometry and its dynamic change characteristics of M. sibirica are not clear, especially from the perspective of water and nutrient use. It is necessary to further strengthen the research on the adaptation strategies of M. sibirica to chenier habitats. Therefore, the seasonal changes of C, N, and P stoichiometry of M. sibirica and soils in the chenier of YRD and WUE of M. sibirica were analyzed to (1) clarify the nutrient characteristics and seasonal dynamics of nutrient use in different organs of M. sibirica; (2) reveal the seasonal characteristics of WUE and its relationship with leaf stoichiometry; and (3) explain the correlation between different organ stoichiometric characteristics and WUE of M. sibirica and soil nutrients. The results contribute to a better understanding of the adaptation of M. sibirica to the chenier coastal environment and provide a theoretical basis for the protection and restoration of vegetation in the coastal zone of YRD.
The study area is located in the chenier of the YRD in Wudi County, Shandong Province, China (37°35′∼38°12′ N, 118°33′∼119°20′ E), with a semi humid continental monsoon climate. The average annual temperature ranges from approximately 11.7 °C to 12.6 °C, and the average annual precipitation is 550 mm.
Sample lines were set along the vertical coastline through grid distribution, that based on soil texture, geomorphology, and land use type of chenier of YRD and the distance from the coast. Three sample points were set on the sample ground wire, the spacing of each sample point was not less than 1 km, and three quadrats (each with a size of 5 m × 5 m) were set at each sampling point.
The samples were collected in May, July, and September on the seaward side, representing the seasons of spring, summer, and autumn, respectively. Representative M. sibirica communities were selected and a random sample method was used to collect three to five complete plant samples, including the root system, from each sample point, which were placed inside ventilated bags. A total of 36 plant samples were collected during the sampling period. The soil samples were collected by using a 4.5 cm diameter special earth drill in four soil depths: 0–10 cm, 10–20 cm, 20–40 cm, and 40–60 cm. Three parallel soil samples were taken at each sampling point adjacent to the plant. The soil was placed inside sealed bags and brought back to the lab. The soil samples were naturally air-dried, sieved, and stored for extraction and determination. The plant samples were separated into root, stem, and leaf, then dried at 105°C for 20 min, and then transferred to 70°C for 48 h. The dried samples were grinded, and then selected by 80 mesh sieving.
The leaf δ13C determination was carried out by the Finnigan DELTAPlus XP stable isotope mass spectrometer (Thermo Electron Corp., Waltham, MA, United States). The value of plant leaf δ13C was calculated by the following formula:
in which 13C/12Csample is the 13C/12C ratio of plant leaf samples, and 13C/12Cstandard is the 13C/12C ratio of glycine in the determination process.
The total carbon (TC) and total nitrogen (TN) of soil and plant were determined by element analyzer (Vario EL III, Elementar, Germany), while total phosphorus (TP) was determined by molybdenum-antimony colorimetric method (Lu, 1999).
Excel 2010 (Microsoft Corp., Redmond, WA, United States) and SPSS19.0 (SPSS Inc., Chicago, IL, United States) were used to analyze the variance of the data. One-way ANOVAs were conducted on the C, N, and P contents and their stoichiometric ratios of soil and roots, stems and leaves of M. sibirica, respectively. Duncan’s test, Pearson method and maximum likelihood method was used for multiple comparison, correlation analysis, and structural equation modeling, respectively.
The C, N, and P contents of each organ of M. sibirica showed different variation patterns with seasonal changes (Figure 1). The C content of root was significantly higher than that of the stem and leaf (P < 0.05), and there was no significant difference between the stem and leaf (P > 0.05). The C content of the root did not change significantly with seasonal changes, while the C content of both stem and leaf showed a significant decrease with the increase of months (P < 0.05). The N content of all organs showed a similar change pattern in each month, that was leaf > stem > root, and the differences were significant (P < 0.05). The dynamic changes of N showed that N of both root and stem increased and then decreased with the increase of the month, and both had the highest value in July, which was significantly higher than that in May and September. The maximum leaf N content was in May, and decreased with the increase of the months, which was different from the change of root and stem N content. There were differences in the P changes of each organ of M. sibirica, in which the P showed leaf > stem > root in May and July (P < 0.05). In September, it showed stem > leaf > root (P < 0.05). In the whole growing season, P content showed stem = leaf > root. According to the dynamic change characteristics, the P content of the root and stem was highest in September, while the P content of the leaf decreased and then increased with the month, with the lowest in July and no significant difference between May and September.
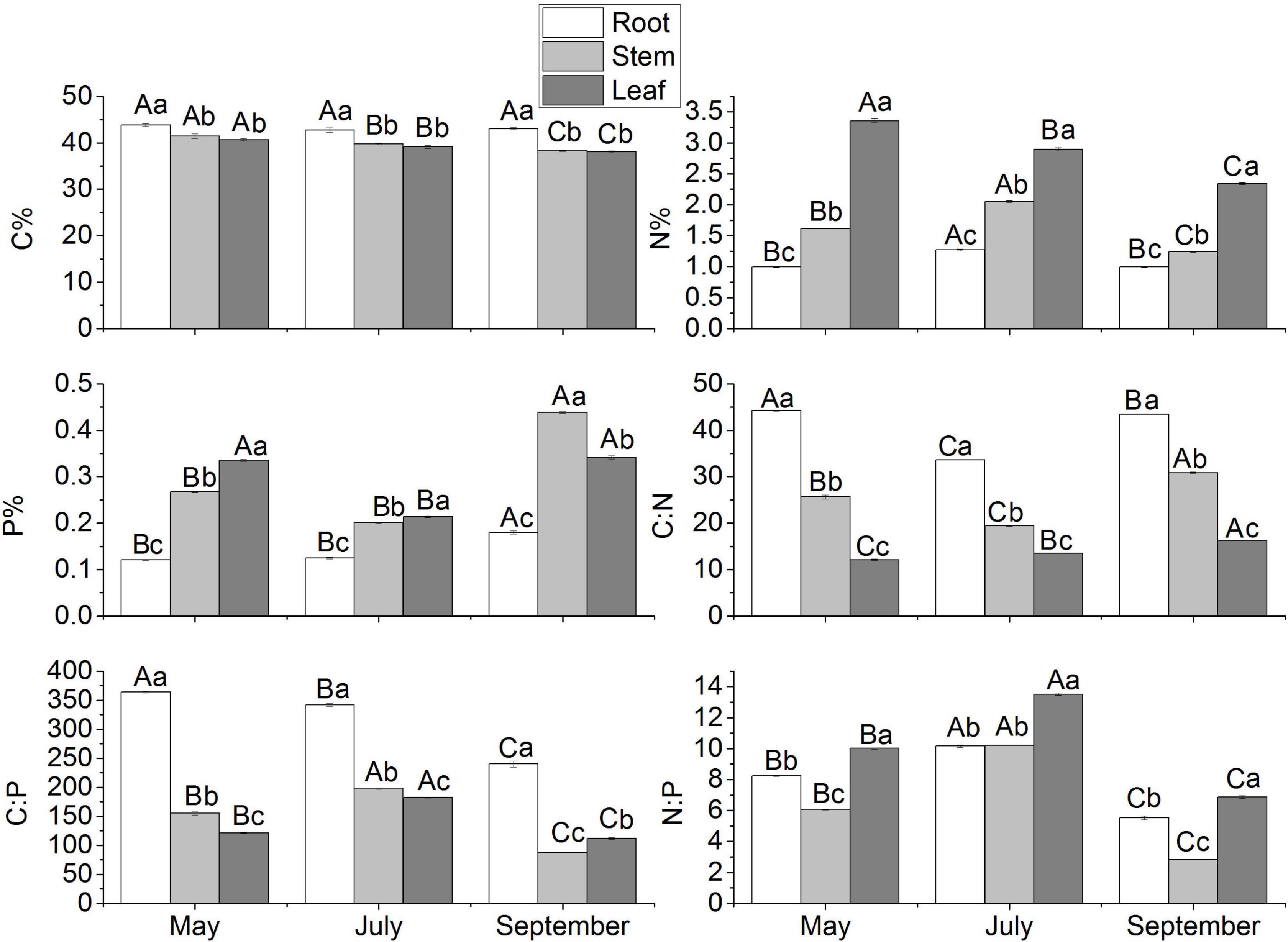
Figure 1. Seasonal dynamics of C, N, and P stoichiometry in different organs of M. sibirica. Different capital letters indicate the significant difference in the same organ in different months, and different small letters indicate the significant difference between different organs in the same month.
The seasonal pattern of C:N:P stoichiometric ratio was different for each organ (Figure 1). The C: N ratio showed root > stem > leaf, and all the differences were significant (P < 0.05). The C: N of both root and stem tended to decrease and then increase with increasing months. The maximum of root C:N was in May, the maximum of stem C:N was in September, and the lowest of both root and stem occurred in July with 33.64 and 19.38, respectively. Leaf C:N showed an increasing trend with increasing months. Root C:P was significantly higher than that of both stem and leaf throughout the growing season (P < 0.05), and its C:P tended to decrease. Stem and leaf C:P showed similar trends, both increasing first and then decreasing. C:P of root, stem, and leaf was lowest in September and highest in May, July, and July, respectively. N:P in different organs showed that leaf > root > stem (P < 0.05), and the dynamic change pattern of N:P in different organs was similar. The N:P of root, stem, and leaf increased and then decreased with increasing months, with the highest in July and the lowest in September, respectively.
There were high correlations in C, N, P content and C:N:P ratios of M. sibirica (Table 1). The correlations were different between different seasons. In May, the correlations among all indicators were significant except for N:P. The correlations between July and May were similar, and N: P was significantly positively and negatively correlated with N content and C:N, respectively. In September, significant correlations were reduced. There was no correlation between C and N, N and P, N and C:P, P and C:N. In all months, there were no significant correlations among C, N, and P of M. sibirica, but there were significant correlations between C, N, and P contents and C:N, C:P, and N:P.
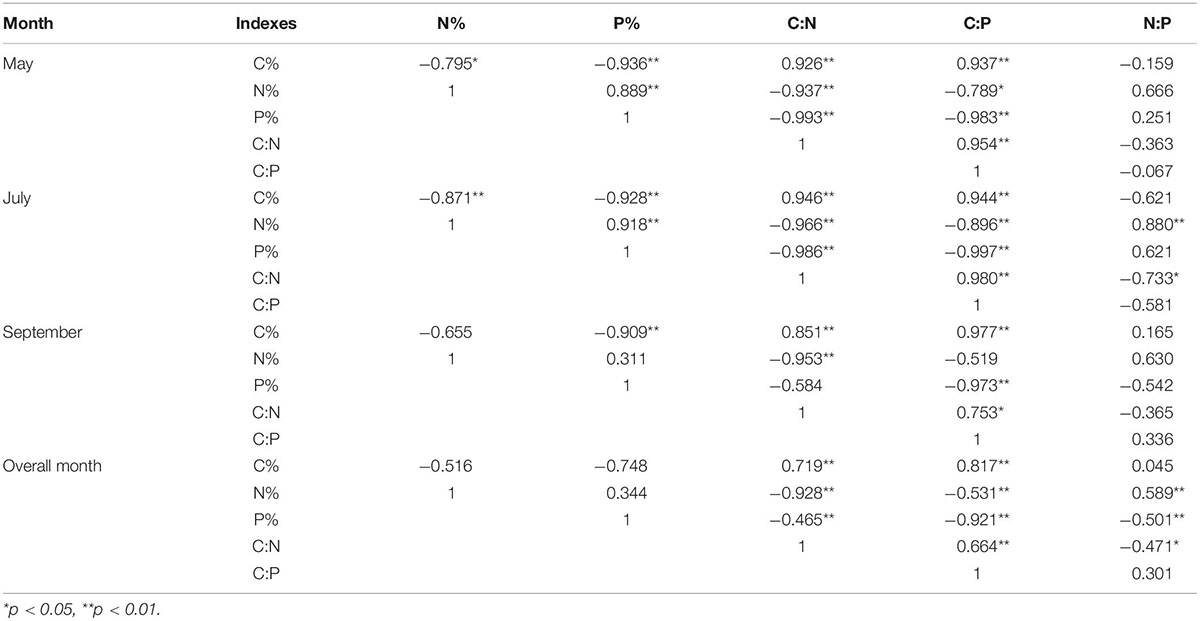
Table 1. Correlation of C, N, and P Stoichiometry in different months of M. sibirica.
The C, N, and P stoichiometric correlations among the organs of M. sibirica are shown in Table 2. Except for the C which was not significant between the stem and the root, the other indexes were significantly positively correlated, and the correlation coefficient was high, ranging from 0.843 to 0.983. C, P, C:P, and N:P were significantly and positively correlated between stem and leaf, and the correlation coefficient was 0.940, 0.744, 0.863, and 0.998, respectively. However, only N:P was significantly and positively correlated between leaf and root with a correlation of 0.989. N:P had the highest correlation in all organs and was most closely related to each organ.
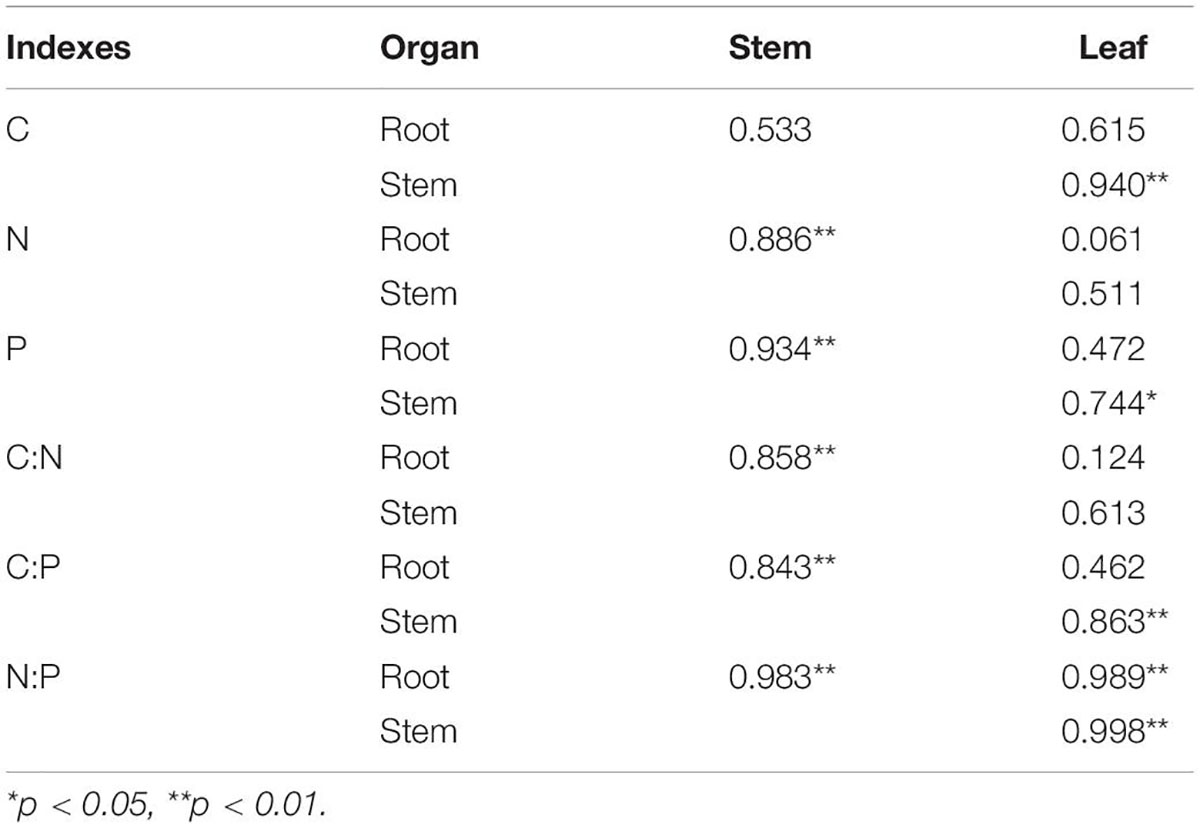
Table 2. Correlation of C, N, and P stoichiometry among roots, stems and leaves of M. sibirica.
The seasonal variation of WUE of M. sibirica leaf is shown in Figure 2. WUE in May was significantly higher than in July and September (P < 0.05). There was no significant difference between July and September. The average δ13C during the growing season was −29.98‰, which was significantly lower than the global plant δ13C average of −28.74‰.
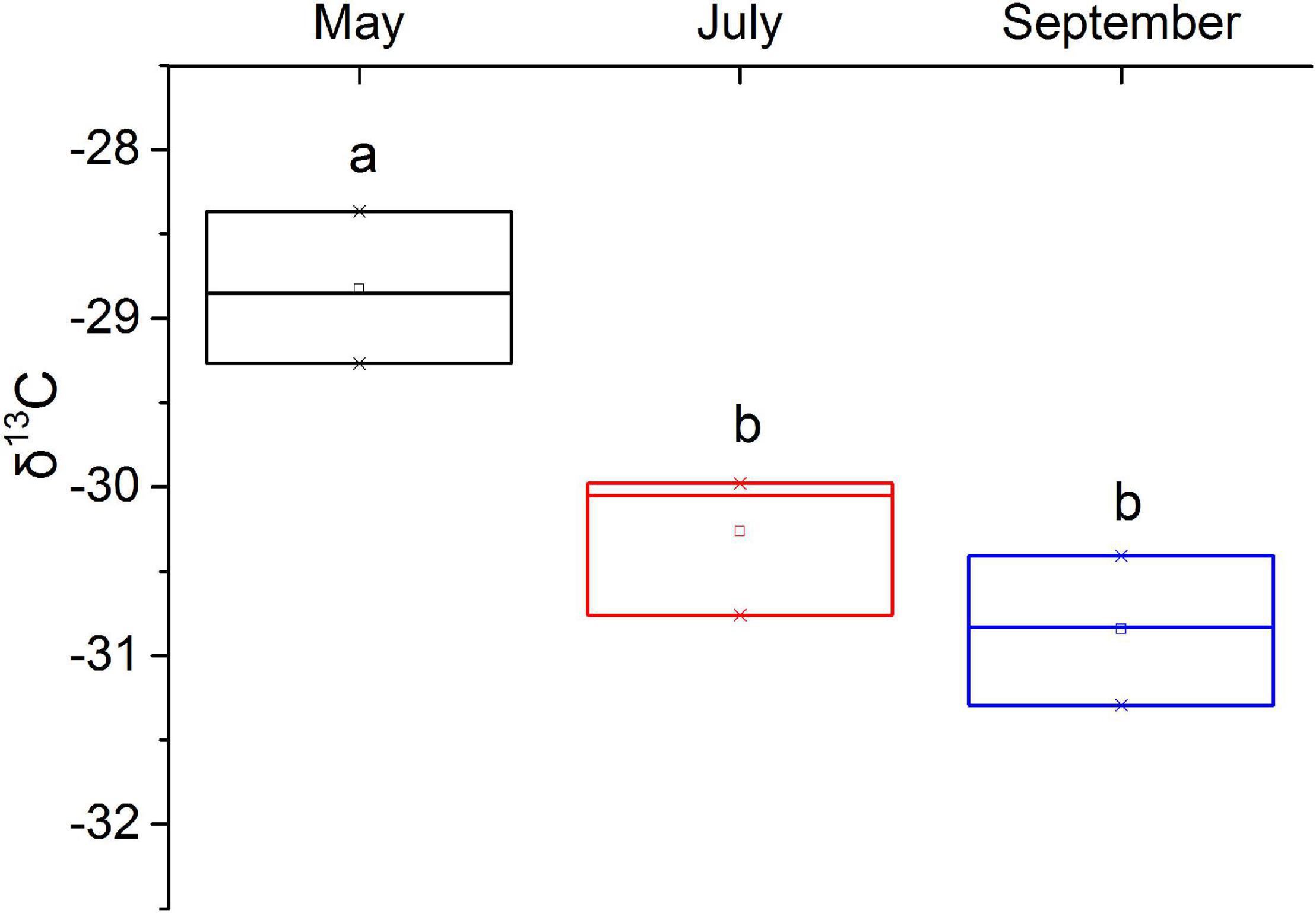
Figure 2. Seasonal variation characteristics of δ13C in leaves of M. sibirica.
Correlation analysis (Table 3) showed that the WUE of M. sibirica was significantly positively correlated with leaf C and leaf N (P < 0.01) and significantly negatively correlated with leaf C:N. In addition, WUE was positively correlated with leaf P content and N:P but did not reach a significant level.
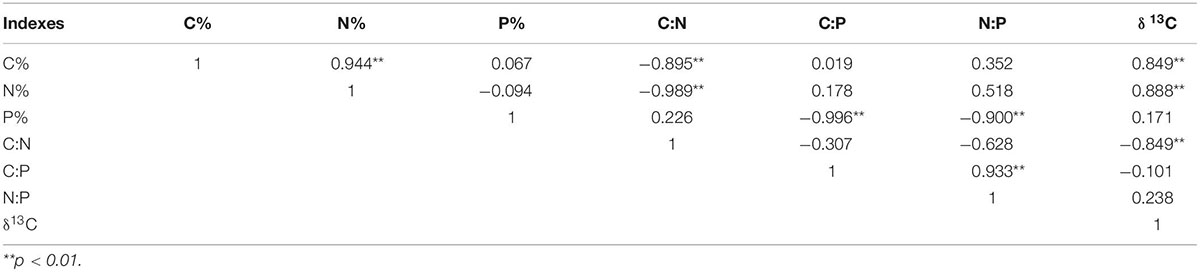
Table 3. Correlation between water use efficiency (WUE) and leaf stoichiometry.
Structural equation modeling was used to analyze the effect of water utilization and seasonal variation on C, N, and P nutrient characteristics (Figure 3). Seasonal variation had a significant direct negative effect on leaf P (P < 0.001) and a significant positive effect on leaf N (P < 0.05), while the effect on C content and δ13C was not significant (P > 0.05). The direct effects of δ13C on the C, N, and P content of leaves were positive. The results of structural equation modeling were consistent with the results of correlation analysis, and all the above effects reached a highly significant level. The effect coefficients showed that the negative effect of seasonal variation on the leaf stoichiometric P was greater than the positive effect of WUE on it, and the positive effect of WUE on the leaf N content was greater than the positive effect of seasonal variation on it. In addition, leaf C content was more significantly influenced by the direct effect of WUE. Leaf P content was influenced by the opposite effects of seasonal variation and WUE, while leaf N content was influenced by the same positive effect of both.
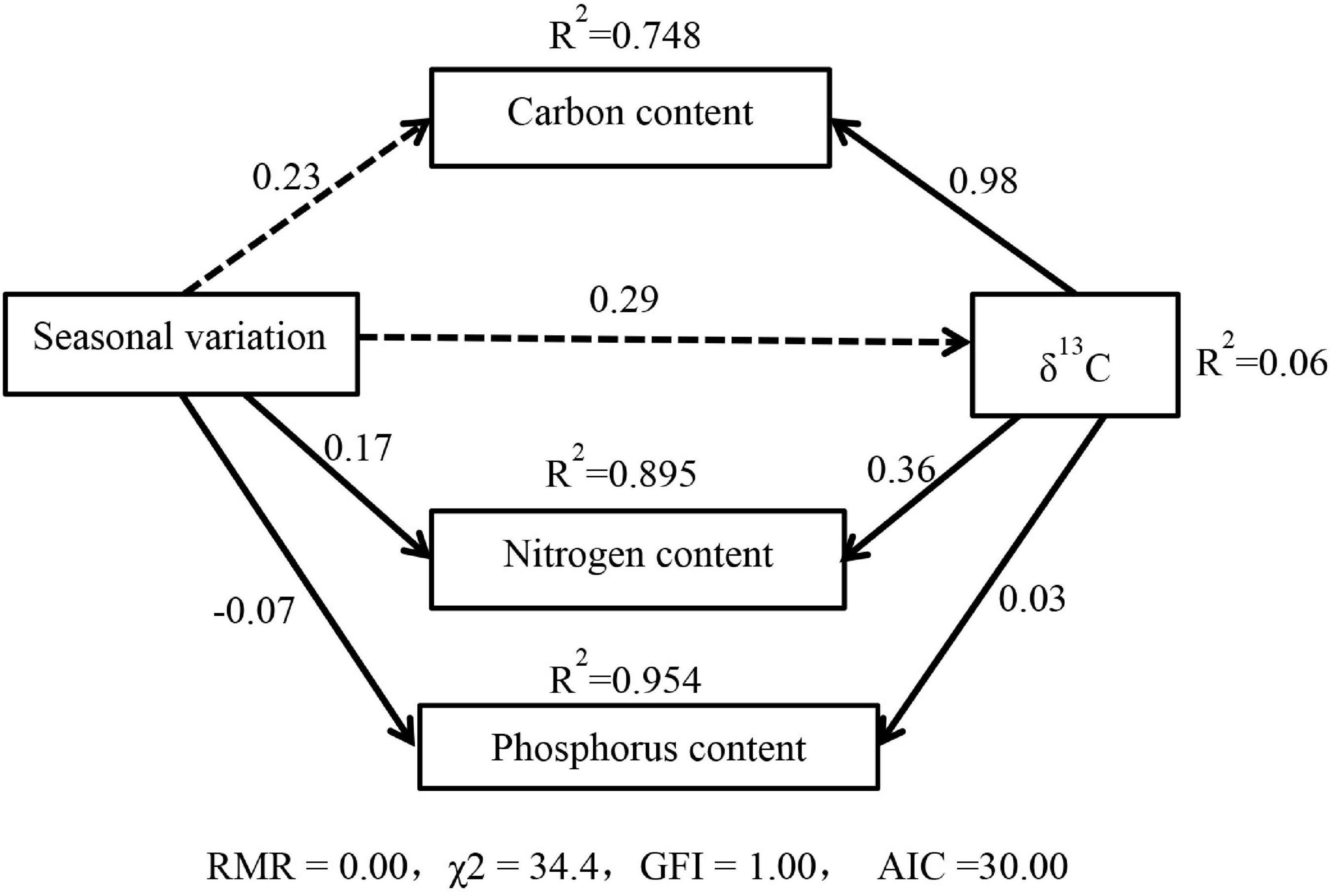
Figure 3. Relationships between season, water use efficiency, and C, N, P content in the leaves of M. sibirica. Solid lines are statically different at P < 0.001, while dotted lines are not significant at P > 0.05.
The C, N, and P stoichiometric characteristics of each soil layer at different seasons are shown in Figure 4. Overall, there was no significant change in C content with seasonal change in the same soil layer. However, the C of different soil layers showed a trend of increasing and then decreasing with increasing depth. The highest C content was in the 20–40 cm soil layer, which was significantly higher than that in the 0–10 cm soil layer. In the same soil layer, the N of the 40–60 cm soil layer showed a trend of decreasing and then increasing with seasonal changes, while the differences of N content of other soil layers with seasonal changes were not significant. Vertically, the pattern of variation in N content between soil layers in each month was consistent, showing a decrease with increasing soil depth. P content tended to increase in the 10–20 cm soil layer with increasing months, and was significantly higher in September than in July, while there was no significant difference in P content between May and July. The overall P content variation in each soil layer was similar to the N variation characteristics.
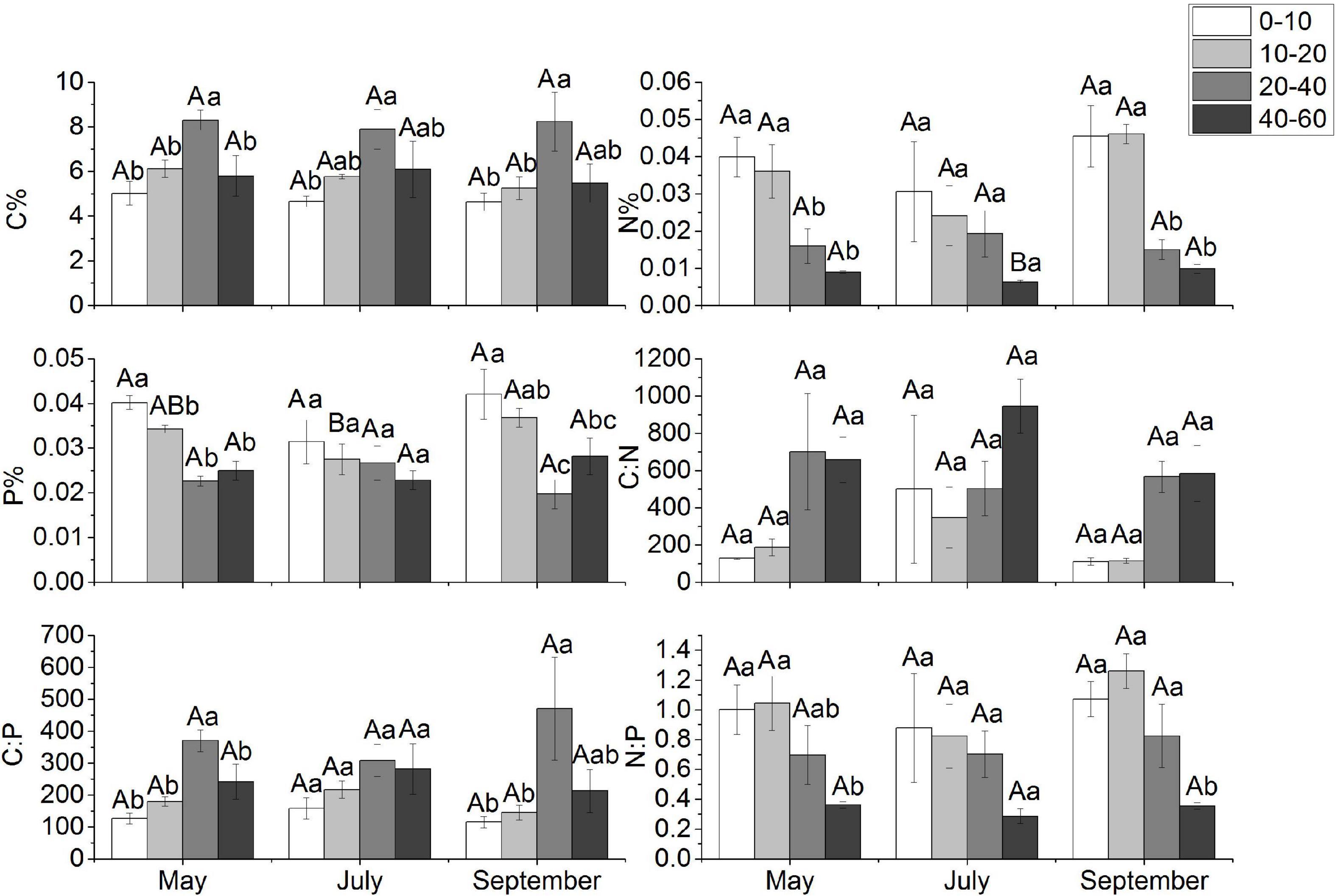
Figure 4. Seasonal dynamics of C, N, and P stoichiometry in different soil layers. Different capital letters indicate the significant difference in the same soil layer in different months, and different small letters indicate the significant difference of different soil layers in the same month.
In soil stoichiometric ratios, the seasonal variation of C:N in the same soil layer showed a general trend of increasing and then decreasing, but the seasonal variation in the same soil layer did not reach the level of significant difference. In the same month, the differences between soil layers were not significant. Similarly, both C:P and N:P did not reach significant levels of difference among months, but there were some differences among soil layers under the same month. In May, the C:P of 20–40 cm soil layer was significantly higher than that of the other three soil layers, and the N:P of 0–20 cm soil layer was significantly higher than that of 40–60 cm soil layer. In July, C:P and N:P were similar between different soil layers and were not significantly different. In September, C:P in 20–40 cm soil layer was significantly higher than that in 0–20 cm soil layer, and N:P in 0–40 cm soil layer was significantly higher than that in 40–60 cm soil layer.
Redundancy analysis (RDA) of stoichiometry, WUE in M. sibirica and soils are shown in Figure 5. The longer rays of soil N:P and soil P content suggested that both were important factors affecting the ecological stoichiometric characteristics and WUE of each organ of M. sibirica. In contrast, the ray of soil C, soil C:P, and soil C:N were shorter, indicated that the influences of these three factors were less. Soil P content was positively correlated with root C:N, root C content, stem C:N and leaf P content, and negatively correlated with root N content, stem N, C:P, N:P and leaf δ13C. Soil N:P was positively correlated with root P content, stem P content, and leaf C:N, and was negatively correlated with root C:P, stem C content, leaf C content, leaf N content, and leaf δ13C. The stoichiometric characteristics of root, stem, and leaf were greatly affected by soil N:P in September.
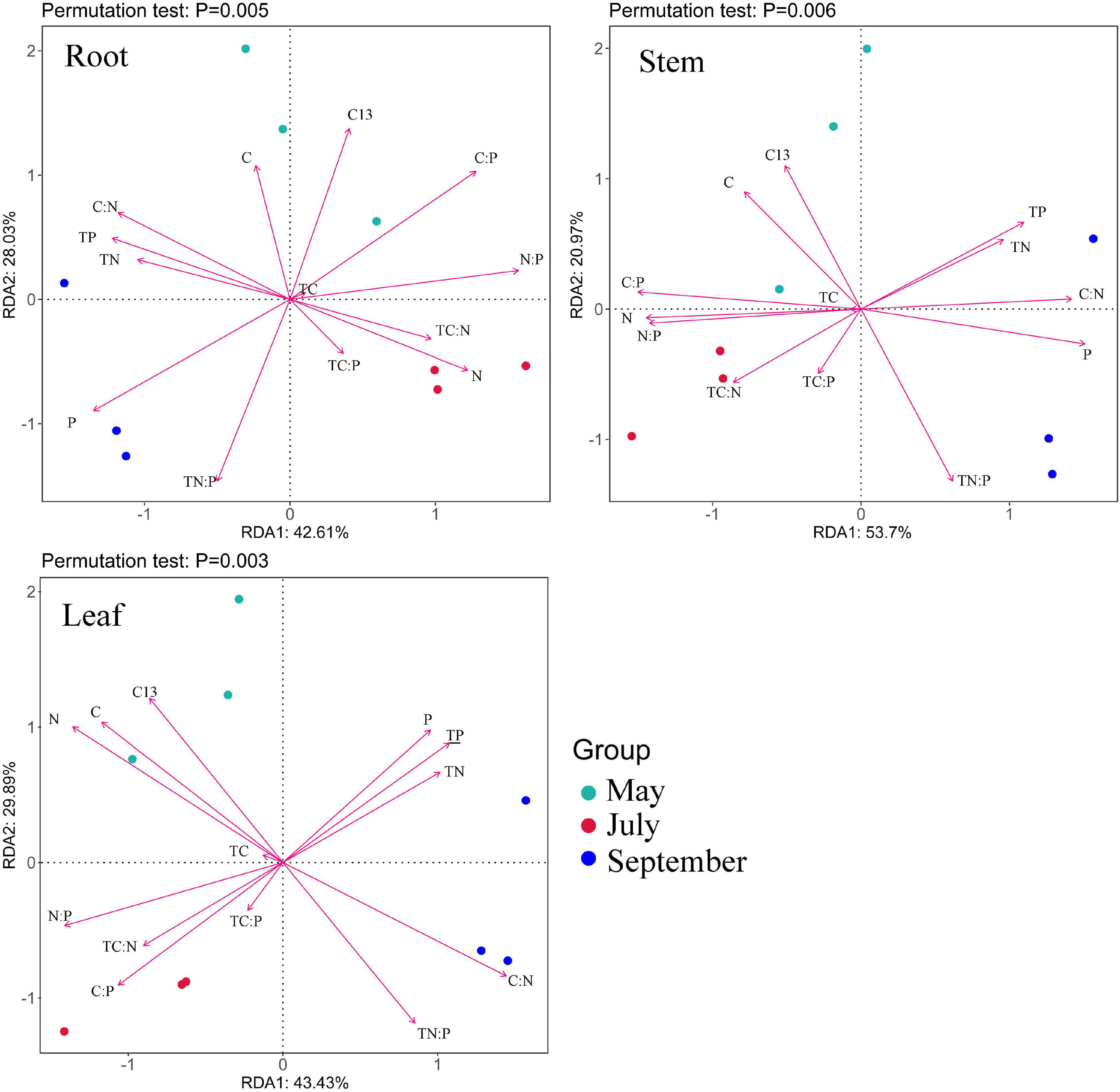
Figure 5. Redundancy analysis of stoichiometry, WUE in M. sibirica and soil nutrients. C, Plant carbon content; N, Plant nitrogen content; P, Plant phosphorus content; C: N, Plant C: N; C:P, Plant C:P; N:P, Plant N:P; C13, Plant WUE; TC, Soil carbon content; TN, Soil nitrogen content; TP, Soil phosphorus content; TC:N, Soil C:N; TC:P, Soil C:P; TN:P, Soil N:P.
The ecological stoichiometric characteristics of plants under seasonal changes reflect the inherent response and adaptation of plants to environmental changes. For example, a study on the dominant shrubs in a dry-hot valley showed that the contents of C, N, and P in leaves varied with seasons, and the seasonal variation of C content was less than that of N and P content (Liu et al., 2020). Xiong et al. (2020) found that the leaf N and P contents of typical dicotyledons in the Mongolian steppe showed a decreasing trend throughout the growing season.
In this study, C content of root, stem and leaf of M. sibirica did not vary significantly with the seasons, but N and P contents of each organ showed significant variability with the seasons, with leaf N gradually decreasing and the P content first decreasing and then increasing with the seasons. This was consistent with some previous studies (Niu et al., 2013; Jing et al., 2020). N and P were regarded as the most important nutrient factors in limiting plant growth (Drinkwater and Snapp, 2007). With the change of seasons, plants optimized their growth as much as possible through the rational distribution and adjustment of nutrients in various tissues (Weih et al., 2016). In this study, the N content of M. sibirica was more distributed in the leaf during the early growth period, the N content of root and stem was more dominant during the peak growth period, and N contents in stem and leaf were affected during the late period, with little effect on the root. The P content in each organ was the most in September, indicating that the accumulation of P in each organ had an advantage in the later growth stage. The response patterns of N and P content of each organ of M. sibirica with the seasons reflected the distribution and transfer characteristics of nutrients at different growth stages. In addition, the path analysis showed that seasonal changes had different effects on N and P contents in leaves, which was related to the element concentration pattern in plant was affected by plant developmental stage and weather (Weih et al., 2016; Zhang et al., 2016).
Plant C:N:P was an important physiological index, that reflected the growth rate of plants. Plant C:N and C:P were used to reflect plant utilization of nutrients (Wang et al., 2014). There was a significant correlation between the C, N, and P content and the stoichiometric ratios of C:N, C:P, and N:P in different seasons, which reflected one of the general laws of nutrient stoichiometry in higher terrestrial plants, that was, the dominant role of plant leaf N and P contents on leaf C:N and C:P (Sardans et al., 2012). It reflected the strong coupling mechanism between plant nutrients (Zhou et al., 2019). Previous studies have found that the distribution of N and P content in leaf followed a certain stoichiometric law. Plants maintained a dynamic balance of C:N:P through stoichiometric homeostasis mechanism. Leaf N:P increased with increasing plant age, reflecting the transition from N-limited to P-limited, a dynamic mechanism usually observed in both short- and long-term time series (Fan et al., 2015). Dynamic nutrient stoichiometry and limitation implied plant nutrition status at different growth stages (Bratt et al., 2020). In this research, the seasonal dynamics of leaf N:P reflected the variation of the N nutrient limitation degree of M. sibirica. In July, leaf N:P increased significantly compared with May, and N:P was close to 14, indicating that the nutrient limitation degree of N content was reduced in this period, and the growth of M. sibirica was in a relatively good nutritional state. In September, N:P was significantly lower than in May and July, indicating an increase in N limitation at the end of growth. This was consistent with the trend of increasing and then decreasing N:P in aboveground organs of Imperata Cylindrica with seasons (Niu et al., 2013).
Physiological differences in the structure and function of different organs may lead to differences in nutrient uptake and accumulation in different organs of plants (Feng et al., 2019; Li et al., 2019). In the present study, there was no significant correlation in C among the root, stem, and leave of M. sibirica, which was consistent with the results of Liu et al. (2020). And there was a correlation between the utilization of N and P nutrients by various plant organs (Han et al., 2005; Ping et al., 2014). In our study, the N:P between root and stem, stem and leaf, and leaf and root of M. sibirica showed highly significant positive correlations, which were consistent with the previous studies (Yuan et al., 2011), and reflected the uniformity of growth among plant organs. More correlations were seen between stem and leaf, root and stem than those between leaf and root, and this result was similar to that of Platycladus Orientalis (Feng et al., 2019).
Plant nutrients regulated the plant’s photosynthetic capacity by influencing leaf stomatal opening and photosynthetic rate, which in turn influenced plant δ13C (Zhang et al., 2019b). In this study, the significant correlations of δ13C with C, N contents, and C:N of leaf indicated that there were important effects of leaf WUE on stoichiometric characteristics. The WUE of M. sibirica was positively correlated with the leaf C content, which was consistent with the physiological characteristics that carbon fixation and WUE were closely related in the plant photosynthesis process, and reflected the relationship between them. The higher the plant WUE, the higher the amount of leaf carbon assimilated into organic matter (Basso and Ritchie, 2018). Within a certain range, the increase in plant N content promoted photosynthesis. The significant positive correlation between WUE and leaf N content in this study was due to a positive correlation of leaf N content with carboxylation efficiency and photosynthetic capacity, and therefore there was a positive correlation between δ13C and leaf N content (Hamerlynck et al., 2004). The structural equation model (SEM) analysis showed that seasonal variation did not contribute to the leaf stoichiometry by directly affecting its WUE. This reflects the effect of plant growth stage on WUE, which mainly depends on the change of external environmental conditions, such as precipitation, temperature, and light.
Plant leaf C:N ratios were found to be an important indicator of N distribution and utilization, as well as C acquisition and assimilation (Hedges et al., 1986). Plants with higher WUE had lower nitrogen use efficiency in natural ecosystems (Sheng et al., 2011). Xia et al. (2020) studied two types of halophytes and found that δ13C and C:N in the leaf of both salt-dilute halophyte and salt-repellent halophyte were significantly negatively correlated. This study showed that M. sibirica was similar to the other two types of halophyte in terms of growth adaptation strategies. Salt-secreting plants could also compensate for the reduced N use efficiency by increasing WUE, especially in barren environments, plant water and nutrient use efficiencies might compensate for each other (Salazar et al., 2018).
Soil was the main source of plant nutrients, and the C, N, and P content and stoichiometry of plants were influenced by soil fertility (Lambers et al., 2008). Plant leaf P content was dominated by soil P content, and Chinese plant leaf P content was lower than global plant leaf P content, which was probably due to the lower soil P content of China (He et al., 2008). Our study found that the leaf P content of M. sibirica was significantly and positively correlated with soil P content, suggesting that soil P content was one of the important factors affecting M. sibirica nutrient, which was consistent with the theory that low soil P content caused low leaf P content in plants (He et al., 2008). However, Geng et al. (2011) found that there was no significant correlation between leaf P content and soil P content in Inner Mongolia grasslands. The same phenomenon was found between Robinia pseudoacacia L. and soil in the Loess Plateau (Li et al., 2013). This suggests that the plant–soil nutrient relationship varies depending on the region or species.
The C:N:P ratio in soil directly reflected soil fertility and nutrient status, and influenced plant growth to a certain extent. Studies on the leaf and soil nutrients of nearly two thousand plant species in China showed a positive correlation between the stoichiometry of soil and plant leaf (Han et al., 2011). Previous studies showed that the N:P of subtropical eucalyptus leaves was significantly correlated with soil N:P (Fan et al., 2015). Both C:P and N:P were positively correlated between soil and Robinia pseudoacacia L. in the Loess Plateau (Cao and Chen, 2017). In addition, in the YRD, C:P of Phragmites communis leaf and soil N:P were positively correlated, C:P of Limonium bicolor leaf and both C:P and N:P of soil were positively correlated, and C:P of Suaeda salsa leaf and soil N were positively correlated (Li et al., 2021). In this study, we found that the soil N:P in the chenier of YRD was significantly lower than the soil N:P of forest ecosystems in the country, indicating a soil N deficiency in this region. The leaf C:N of M. sibirica was strongly and positively correlated with soil N:P, indicating that N:P was more closely related to leaf building efficiency than N and P content in soil. Chen et al. (2016) found that the nutritional variation of different organs of Cunninghamia lanceolata had a relative consistency with that of soil, which ensured the stable growth and development of Cunninghamia lanceolata. In our work, the correlations between root N content and soil N: P, between root and stem C: N and soil N content, and between leaf P content and soil P content were significant and positive, indicating a good correlation between each organ of M. sibirica and soil stoichiometry, and embodying a close relationship between M. sibirica and soil nutrient environment with mutual constraints and interactions, which enhances the adaptability of M. sibirica to grow in this region.
The accumulation of N and P by above-ground leaves and stem was higher than that by below-ground roots in various growing seasons of M. sibirica. The plant N was mainly distributed to the leaves in the early growth stage and to the roots and stem in the peak growth stage of M. sibirica. The accumulation of P in each organ was dominant in the later stages of growth. The ratio of C:N, C:P, N:P in different growing seasons showed the highest utilization efficiency of N and P in the root and the lowest limitation of N in the leaf. The degree of N restriction of M. sibirica decreased in July, and increased in September. The overall C, N, and P content and stoichiometric ratio of M. sibirica were strongly correlated, reflecting the coupling between plant nutrient elements. The N:P of root and stem, stem and leaf, and leaf and root were strongly significantly and positively correlated, indicating the same nutrient-limited growth of each organ. The stoichiometric correlations between stem and leaf and between root and stem were higher than those between leaf and root. The δ13C of M. sibirica was significantly higher in May than in July and September, and it was strongly and positively correlated with leaf C and N content, indicating that WUE played an important role in nutrient accumulation. The highly significant negative correlation between WUE and leaf C:N indicated that M. sibirica could not optimize both water and nutrient use in the relatively poor soil habitat, and there was a compensatory effect between water and nutrient use. RDA showed that there was mutual influence and constraint between the root, stem, and leaf of M. sibirica and soil in terms of stoichiometry. Soil P content and soil N:P were the main factors affecting the ecological stoichiometric characteristics and WUE of M. sibirica. This study provides valuable information for the protection and restoration of M. sibirica.
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
TL completed data analysis and wrote the manuscript. ZZ was in charge of the modification of the manuscript. JS conceived and designed the study, and fund support. ZF conducted experiments and data sorting. YZ and WX participated in data sorting and revision of the manuscript. All authors contributed to the article and approved the submitted version.
This funding was supported by the National Natural Science Foundation of China (41871089, 41971119, and 42171059), the “Collection, Conservation, and Accurate Identification of Forest Tree Germplasm Resources” of Shandong Provincial Agricultural Elite Varieties Project (2019LZGC01805), the Natural Science Foundation of Shandong Province (ZR2019MD024 and ZR2020QD004), and the Science and Technology Support Plan for Youth Innovation of Colleges and Universities (2019KJD010).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Basso, B., and Ritchie, J. T. (2018). Evapotranspiration in high-yielding maize and under increased vapor pressure deficit in the US Midwest. Agricultural Environ. Lett. 3:170039.
Bratt, A. R., Finlay, J. C., Welter, J. R., Vculek, B. A., and Van Allen, R. E. (2020). Co-limitation by N and P characterizes phytoplankton communities across nutrient availability and land Use. Ecosystems 23, 1121–1137. doi: 10.1007/s10021-019-00459-6
Cao, Y., and Chen, Y. M. (2017). Coupling of plant and soil C: N: P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau. China. Trees 31, 1559–1570. doi: 10.1007/s00468-017-1569-8
Chen, C., Wang, G. J., Zhao, Y., Zhou, G. X., Li, L., and Gao, J. Q. (2016). Seasonal dynamics and allometric growth relationships of C, N, and P stoichiometry in the organs of Cunninghamia lanceolata from Huitong. Acta Ecol. Sinica 36, 7614–7623.
Crous, K. Y., Wujeska, A., Jiang, M., Medlyn, B. E., and Ellsworth, D. (2019). Nitrogen and phosphorus retranslocation of leaves and stemwood in a mature eucalyptus forest exposed to 5 years of elevated CO2. Front. Plant Sci. 10:664. doi: 10.3389/fpls.2019.00664
Drinkwater, L. E., and Snapp, S. S. (2007). Nutrients in agroecosystems: rethinking the management paradigm. Adv. Agronomy 92, 163–186. doi: 10.1016/s0065-2113(04)92003-2
Elser, J. J., Fagan, W. F., Kerkhoff, A. J., Swenson, N. G., and Enquist, B. J. (2010). Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol. 186, 593–608. doi: 10.1111/j.1469-8137.2010.03214.x
Fan, H. B., Wu, J. P., Liu, W. F., Yuan, Y. H., Hu, L., and Cai, Q. K. (2015). Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil. 392, 127–138.
Feng, H. Y., Du, M. Y., Xin, X. B., Gao, X., Zhang, L. J., Kong, Q. Y., et al. (2019). Seasonal variation in C, N, and P stoichiometry of Platycladus orientalis plantation in the rocky mountainous areas of North China. Acta Ecol. Sinica 39, 1572–1582. doi: 10.5846/stxb201803120483
Geng, Y., Wu, Y., and He, J. S. (2011). Relationship between leaf phosphorus concentration and soil phosphorus availability across Inner Mongolia grassland. Chinese J. Plant Ecol. 35, 1–8.
Hamerlynck, E. P., Huxman, T. E., Mcauliffe, J. R., and Smith, S. D. (2004). Carbon isotope discrimination and foliar nutrient status of Larrea tridentata (creosote bush) in contrasting Mojave Desert soils. Oecologia 138, 210–215. doi: 10.1007/s00442-003-1437-7
Han, W. X., Fang, J. Y., Reich, P. B., Woodward, F. I., and Wang, Z. H. (2011). Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 14, 788–796. doi: 10.1111/j.1461-0248.2011.01641.x
Han, W., Fang, J., Guo, D., and Zhang, Y. (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 168, 377–385. doi: 10.1111/j.1469-8137.2005.01530.x
He, J. S., Wang, L., Flynn, D. F. B., Wang, X. P., Ma, W. H., and Fang, J. Y. (2008). Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes. Oecologia 155, 301–310. doi: 10.1007/s00442-007-0912-y
Hedges, J. I., Clark, W. A., Quay, P. D., Richey, J. E., Devol, A. H., and Santos, M. (1986). Compositions and fluxes of particulate organic material in the Amazon River. Limnol. Oceanogr. 31, 717–738.
Jing, H. X., Sun, N., Umair, M., Liu, C., and Du, H. (2020). Stoichiometric characteristics of soils and dominant shrub leaves and their responses to water addition in different seasons in degraded karst areas in southern yunnan of China. Chinese J. Plant Ecol. 44, 56–69.
Lambers, H., Raven, J. A., Shaver, G. R., and Smith, S. E. (2008). Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 23, 95–103. doi: 10.1016/j.tree.2007.10.008
Lee, J. S., Ihm, B. S., Du, S. C., Son, D. Y., and Kim, J. W. (2007). Soil particle sizes and plant communities on coastal dunes. J. Plant Biol. 50, 475–479. doi: 10.1007/BF03030685
Li, H., Li, J., He, Y. L., Li, S. J., Liang, Z. S., Peng, C. H., et al. (2013). Changes in carbon, nutrients and stoichiometric relations under different soil depths, plant tissues and ages in black locust plantations. Acta Physiol. Plan. 35, 2951–2964. doi: 10.1007/s11738-013-1326-6
Li, M., Huang, C., Yang, T., Drosos, M., Wang, J. Z., Kang, X. M., et al. (2019). Role of plant species and soil phosphorus concentrations in determining phosphorus: nutrient stoichiometry in leaves and fine roots. Plant Soil. 445, 231–242.
Li, T., Sun, J. K., and Fu, Z. Y. (2021). Halophytes differ in their adaptation to soil environment in the Yellow River Delta: effects of water source, soil depth, and nutrient stoichiometry. Front. Plant Sci. 12:675921. doi: 10.3389/fpls.2021.675921
Liu, Y., He, J. W., Yu, H., Lin, Y. M., and Wang, D. J. (2020). Nutrients (C, N, P) contents and stoichiometric ratios of fine root, coarse root and leaf in dominant shrubs in dry-hot valley. Mountain Res. 38, 668–678.
Lu, R. K. (1999). Analytical Methods for Soil and Agro-Chemistry. Beijing: China Agricultural Science and Technology Press.
Luo, Y., Peng, Q., Li, K., Gong, Y., and Han, W. (2021). Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in xinjiang, China. Catena 199:105100.
Niu, D. C., Li, Q., Jiang, S. G., Chang, P. J., and Fu, H. (2013). Seasonal variations of leaf C:N:P stoichiometry of six shrubs in desert of China’s Alxa Plateau. Chinese J. Plant Ecol. 37, 317–325.
Ping, C., Wang, C. K., and Quan, X. K. (2014). Influence of environmental changes on stoichiometric traits of nitrogen and phosphorus for Larix gmelinii trees. Acta Ecol Sinica 34, 1965–1974. doi: 10.5846/stxb201306301805
Salazar, T. D., Castro, J., Villar, S. P., Vinegla, B., Matías, L., Michelsen, A., et al. (2018). The “isohydric trap”: a proposed feedback between water shortage, stomatal regulation, and nutrient acquisition drives differential growth and survival of European pines under climatic dryness. Global Change Biol. 24, 4069–4083. doi: 10.1111/gcb.14311
Sardans, J., Rivas, A., and Penelas, J. (2012). The C: N: P stoichiometry of organisms and ecosystems in a changing world: a review and perspectives. Perspect. Plant Ecol. 14, 33–47.
Sheng, W. P., Ren, S. J., Yu, G. R., Fang, H. J., Jiang, C. M., and Zhang, M. (2011). Patterns and driving factors of WUE and NUE in natural forest ecosystems along the North-South Transect of Eastern China. J. Geograph. Sci. 21, 651–665. doi: 10.1007/s11442-011-0870-5
Suzuki, S., Abe, M., and Motokawa, M. (2011). Allometric comparison of skulls from two closely related weasels, Mustela itatsi and M. sibirica. Zoology 28, 676–688. doi: 10.2108/zsj.28.676
Tang, Z., Xu, W., Zhou, G., Bai, Y., Li, J., Tang, X., et al. (2018). Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. U S A. 115, 4033–4038.
Wang, M., Murphy, M. T., and Moore, T. R. (2014). Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog. Oecologia 174, 365–377. doi: 10.1007/s00442-013-2784-7
Wang, Z. N., Lu, J. Y., Yang, H. M., and Zhang, Q. P. (2015). Stoichiometric characteristics of carbon, nitrogen, and phosphorus in leaves of differently aged lucerne (Medicago sativa) stands. Front. Plant Sci. 6:1062. doi: 10.3389/fpls.2015.01062
Weih, M., Pourazari, F., and Vico, G. (2016). Nutrient stoichiometry in winter wheat: element concentration pattern reflects developmental stage and weather. Sci. Rep. 6:35958. doi: 10.1038/srep35958
Xia, D. J., Liu, Q. R., Zou, L., Ge, Z. W., Xue, J. H., and Peng, S. L. (2020). Foliarδ13C correlates with elemental stoichiometry in halophytes of coastal wetlands. Acta Ecol. Sinica 40, 2215–2224. doi: 10.5846/stxb201901170140
Xie, W. H., Zhou, R. L., Liang, H. M., Qu, H., and Qiang, S. B. (2015). Physiological difference in Messerschmidia sibirica grown in inland and coastal sand land in nature environment and under sand burial. J. Desert Res. 35, 1538–1548.
Xiong, X. S., Cai, H. Y., Li, Y. Q., Ma, W. H., Niu, K. C., Chen, D. M., et al. (2020). Seasonal dynamics of leaf C, N and P stoichiometry in plants of typical steppe in Nei Mongol, China. Chinese J. Plant Ecol. 44, 1138–1153.
Yan, W. M., Zhong, Y. Q., Zheng, S. X., and Shangguan, Z. P. (2016). Linking plant leaf nutrients/stoichiometry to water use efficiency on the Loess Plateau in China. Ecol. Eng. 87, 124–131. doi: 10.1016/j.ecoleng.2015.11.034
Yang, C., and Xu, W. (2021). Seasonal variations in carbon, nitrogen, and phosphorus stoichiometry of a Robinia pseudoacacia plantation on the Loess Hilly Region. China. Forests 12:214. doi: 10.3390/f12020214
Yuan, Z. Y., Chen, H. Y. H., and Reich, P. B. (2011). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat. Commun. 2:344. doi: 10.1038/ncomms1346
Zhang, P., Bakker, E. S., Zhang, M., and Xu, J. (2016). Effects of warming on Potamogeton crispus growth and tissue stoichiometry in the growing season. Aquatic Botany 128, 13–17.
Zhang, C. W., Tian, X. Y., and Zhang, C. S. (2019a). Diversity and probiotic activities of endophytic bacteria associated with the coastal halophyte Messerschmidia sibirica. Appl. Soil Ecol. 143, 35–44.
Zhang, X. S., Joachimski, M. M., Over, D. J., Ma, K. Y., Huang, C., and Gong, Y. M. (2019b). Late Devonian carbon isotope chemostratigraphy: a new record from the offshore facies of South China. Global Planet. Change 182:103024.
Keywords: stoichiometry, plant organ, stable carbon isotope, growing season, Yellow River Delta
Citation: Li T, Zhang Z, Sun J, Fu Z, Zhao Y and Xu W (2022) Seasonal Variation Characteristics of C, N, and P Stoichiometry and Water Use Efficiency of Messerschmidia sibirica and Its Relationship With Soil Nutrients. Front. Ecol. Evol. 10:948682. doi: 10.3389/fevo.2022.948682
Received: 20 May 2022; Accepted: 01 June 2022;
Published: 24 June 2022.
Edited by:
Chuanyu Gao, Northeast Institute of Geography and Agroecology (CAS), ChinaReviewed by:
Zhongsheng Zhang, Key Laboratory of Mollisols Agroecology, Northeast Institute of Geography and Agroecology (CAS), ChinaCopyright © 2022 Li, Zhang, Sun, Fu, Zhao and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jingkuan Sun, c3VuamluZ2t1YW5AMTI2LmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.