 Jeffery A. Wilkinson*John M. Romansic
Jeffery A. Wilkinson*John M. Romansic- H. T. Harvey & Associates, Los Gatos, CA, United States
Numerous studies have documented high numbers of amphibians killed by vehicular strikes on roads. This direct mortality can cause population declines and extirpations, but in some cases the declines might be masked, initially, by large population sizes. Population viability analysis can help discern population trajectories and identify incipient declines. We applied this tool to a situation in Santa Clara County, California where a dead-on-the-road carcass survey in 2017 demonstrated that a large number of newts in the genus Taricha were being killed by vehicles using a small two-lane road (Alma Bridge Road) most likely during annual breeding migrations to a local reservoir (Lexington Reservoir). To help determine the effect of this road-based mortality on the California newt (T. torosa) population, we conducted a drift fence/pitfall trap array study on the road during the 2020–2021 breeding season. Drift fence/pitfall trap arrays were installed at six locations along a 6.6-km stretch of the road and daily surveys were conducted at these arrays over a 148-day period from mid-November to end of March. Daily traffic and precipitation data were also recorded to help inform timing of proposed mitigation efforts. Concurrently, a group of community scientists conducted a dead-on-the-road carcass survey over the entire 6.6-km study area. We calculated the number of adult newts attempting to cross Alma Bridge Road at the arrays and the associated road-based mortality rates. Then, we combined our array results with road mortality data provided by the community scientists to estimate the number of adult California newts attempting to cross the road and their mortality rates over the entire study area during the survey period. We then incorporated this data into a population viability model to determine whether this road-based mortality rate might, if left unabated, lead to a reduction in, and possibly the eventual extirpation of, the local population of T. torosa breeding in Lexington Reservoir. The model indicated that this population would be extirpated in approximately 92 years. Because the road has been in use for approximately 67 years, we discussed the possible reasons why this population is currently extant and experiencing this high level of road-based mortality.
Introduction
Roads and traffic have long been known to have major impacts on animal populations and communities (Stoner, 1925; Trombulak and Frissell, 2000; Forman et al., 2003). One study estimated that one million vertebrates are killed each day on roads in the United States (Lalo, 1987), and another emphasized that road-kill is a factor in the overall decline of amphibians (Glista et al., 2007). The negative effects of roads and traffic on amphibian populations are well known (Beebee, 2013), and although road mortality may not have a substantial effect on very large populations, it can negatively affect populations of threatened or endangered species over time (Glista et al., 2007). For this reason, three threatened and endangered amphibian species with ranges in or near the San Francisco Bay Area in California have received much attention with respect to the negative impacts of roads: the California tiger salamander (Ambystoma californiense), which is listed under both the Federal and California Endangered Species Acts as threatened, with the Sonoma and Santa Barbara County populations listed federally as endangered (Bain et al., 2017); the Santa Cruz long-toed salamander (A. macrodactytum croceum), which is federally and state listed as endangered (Hobbs, 2013); and the California red-legged frog (Rana draytonii), which is federally listed as threatened, and is state listed as a species of special concern. Concern about potential impacts of roads on their populations has had some influence on how roads are currently constructed and used within the ranges of these species, with some road designs incorporating measures such as barriers to over-the-road movement coupled with undercrossings to allow amphibians to safely cross the roads (Bain et al., 2017). In some areas, compensatory mitigation is required to reduce the impacts of new road construction on these species through land conservation (e.g., U.S. Fish and Wildlife Service, 2005).
A study by Brehme et al. (2018) ranked species of amphibians and reptiles in California based on the risk of roads to their populations. Each species was given a road risk score based on movement distance, frequency of movement, habitat preference, road use, movement speed, fecundity, proportion of population at risk, size of range or amount of isolation, and conservation status, and then grouped into five broad categories of very high risk, high risk, medium risk, low risk, and very low risk. Of the 33 species in the very high risk category, eight were amphibians and 25 were reptiles. Of the eight amphibian species in this category, the top four were salamanders: red-bellied newt (Taricha rivularis) with a score of 561, California newt (T. torosa) with a score of 532, then California tiger salamander, and Sierra newt (T. sierrae), which both received a score of 437.
The placement of the California tiger salamander in the very high risk category is understandable, given its threatened and endangered status as mentioned above. However, the placement of the red-bellied newt and the California newt above, and the Sierra newt at the same level as, the California tiger salamander is less intuitively obvious, as the red-bellied newt and the California newt are listed only as California species of special concern (only the southern California populations of the California newt are listed as such), and the Sierra newt has no special listing status. Everything being equal, the California tiger salamander should have a higher risk factor than these other species because of its threatened and endangered status. These rankings reflect higher individual-level and population-level risk scores in the California newt and the Sierra newt due to the other factors in the risk model, such as longer movement distances and higher frequency of movement than other species, which would increase their chances of crossing (and encountering risk) on a road, along with evidence of high mortality on roads and early disappearance following landscape fragmentation (Brehme et al., 2018). Another interesting finding of Brehme et al. (2018) is the placement of the rough-skinned newt (T. granulosa) within the high risk category along with the Santa Cruz long-toed salamander, with road risk scores of 304 and 308, respectively. The rough-skinned newt has no federal or state protective status, and has a relatively large range of four or more states and four geographic regions within California, whereas the Santa Cruz long-toed salamander is restricted to the southern Santa Cruz and northern Monterey Counties (U.S. Fish and Wildlife Service, 2004). Again, these rankings reflect high individual-level and population-level risk scores.
Both the rough-skinned newt and the California newt inhabit the mountainous parts of the San Francisco Bay Area, but have been mostly extirpated from the lower elevations because of habitat loss caused by development. The distribution of the rough-skinned newt extends south along the San Francisco Peninsula to southern Santa Cruz and southwestern Santa Clara counties. On the other hand, the California newt has several disjunct distributions from Mendocino County south to southern California, one of which is along the San Francisco Peninsula similar to the rough-skinned newt; the California newt also occurs east of the bay in the hills and mountains of western Contra Costa and Alameda counties, south to southern Santa Clara and southwestern San Joaquin counties. Both the rough-skinned newt and California newt are found in sympatry within San Mateo, western Santa Clara, and Santa Cruz counties (Stebbins and McGinnis, 2012).
Even though these two species do not have state or federal protective status in the Bay Area, there is some recognition that development (including roads) is having an impact on their survivorship. Accordingly, some measures have been introduced to protect these species from road mortality. For example, each year the East Bay Regional Parks District closes South Park Drive in Tilden Park from November to March to protect California newts crossing the road during the winter rainy season from vehicle strikes1, and volunteers in Marin assist newts in safely crossing Chilleno Valley Road.2
Other newt populations in the region may be in need of similar intervention to prevent or reduce vehicle strikes. Indeed, during surveys conducted over four breeding seasons spanning 2017 to 2021, a community scientist (Parsons, 2021) documented over 15,000 vehicle-killed newts, including both Bay Area Taricha species, along Alma Bridge Road, a small two-lane road in western Santa Clara County. These results have generated media interest and prompted investigation of potential effects of traffic mortality on the local newt populations.3 This case is particularly intriguing because it appears that large numbers of newts have been killed on the road for many years, yet neither species have become extirpated from the area, at least not yet. Thus, it is unclear whether the newt populations can sustain these levels of road mortality and persist long-term or whether they are on a downward trajectory towards extirpation. Indeed, considerable lag time might elapse between the onset of population decline and ultimate population extirpation.
Alma Bridge Road is a 7.4-kilometer (4.6 mile) long, two-lane road along the east side of Lexington Reservoir (Figure 1). The northern section of the road is used by trucks to transport sandstone from the Lexington Quarry (Vulcan Materials Company) to the east, recreationalists (hikers, boaters, and bicyclist) that use the road for travel and parking to access Lexington Reservoir County Park and surrounding open space preserves, and private residents that have property to the south and east. The road is also used by commuters that choose to bypass a section of Highway 17, especially during times of high congestion. Observations of high newt mortality on Alma Bridge Road suggest that, if left unchecked, such traffic impacts could have an impact on this population (Parsons, 2021).
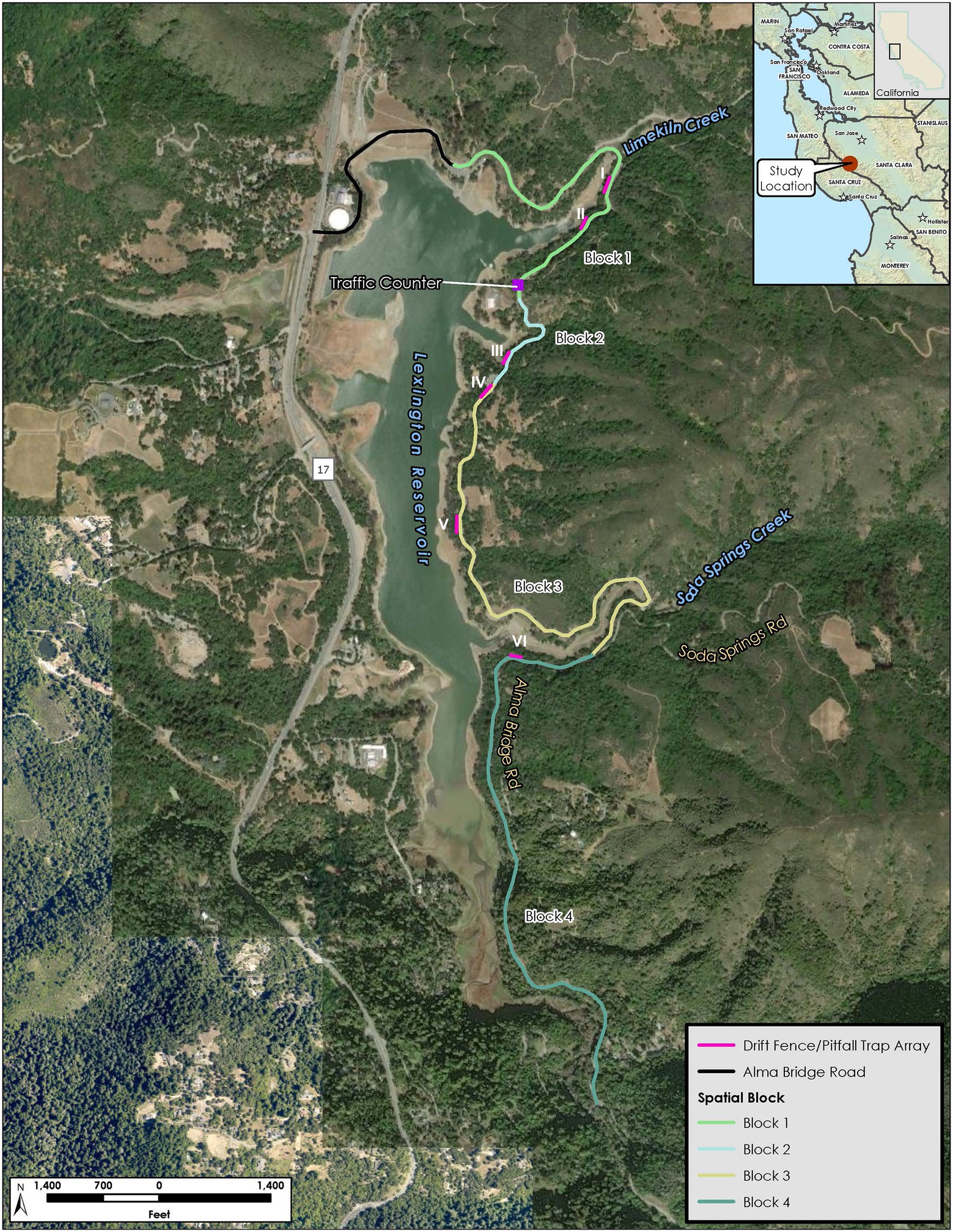
Figure 1. Alma Bridge Road study area, showing Lexington Reservoir, upland habitat, the road, and trapping arrays and spatial blocks along the road.
The purpose of our study is to estimate the effect of current levels of vehicular strikes on the local population of California newts, the newt species that predominates in the study area. Knowledge of these population-level effects will allow better understanding of the conservation value of any mitigation of this impact. Thus, we conducted a drift fence/pitfall trap array study to estimate the number of adult California newts attempting to cross Alma Bridge Road to breed in Lexington Reservoir and estimate the percentage that were killed by vehicular strikes during a single breeding season (2020/2021). We then incorporated these estimates into a population viability model to simulate the long-term impacts of road mortality on the local California newt population.
Materials and methods
Study area
The study area encompassed the approximately 6.6-kilometer (4.1-mile) section of Alma Bridge Road from the Saint Joseph Hill Open Space Preserve (OSP) trail (37.200364°, −121.987036°) to the junction of Aldercroft Heights Road (37.168124°, −121.980288°) (Figure 1). This section of road mostly separated the eastern shore of Lexington Reservoir from the adjacent expanse of upland habitat that is a mixture of public open space and private property. Based on the high dead-on-road (DOR) counts (Parsons, 2021), we suspected that the newt mortality on Alma Bridge Road was a result of adult newts crossing the road from upland habitat to aquatic breeding habitat during annual breeding migrations. California newts breed in ponds, streams, and reservoirs (Kuchta, 2005; Thomson et al., 2016), including Lexington Reservoir and at least one of its inlet streams, downstream from Alma Bridge Road (section 1 of Supplementary material). The area between the reservoir and the road contains little upland habitat and appears to lack other potential breeding sites. Therefore, for the purposes of our study, we defined the local population of California newts subjected to mortality on Alma Bridge Road to consist of those that inhabit upland habitat east of Lexington Reservoir, breed exclusively in Lexington Reservoir or in its inlet streams between Alma Bridge Road and the reservoir, and must cross Alma Bridge Road in order to breed. We acknowledge that some newts breeding in the reservoir may use upland habitat to the southwest (e.g., south of the reservoir or between the reservoir and Highway 17). It is also our understanding that the extension of Lexington Reservoir in Lyndon Canyon west of Highway 17 is not freely connected to the reservoir proper under Highway 17 but instead water between this extension and the reservoir must pass through a high-water Lexington Culvert under the highway north of Black Road that would restrict adult newts and preclude larval newts from freely crossing under Highway 17 between the extension and the reservoir. Therefore, due to residential development southwest of the reservoir and the presence of Highway 17 (a barrier to newt dispersal) to the west, it is our opinion that population dynamics in Lexington Reservoir as a whole are driven primarily by the newts that were explicitly included in our study. Therefore, our references to “population” in this paper refer interchangeably to the group of newts that were explicitly included in our study and the overall population breeding in the reservoir.
Drift fence/pit-fall trap survey
In late October and early November 2020, we installed six drift fence/pitfall trap arrays at the locations indicated in Figure 1. These six arrays were distributed across four spatial blocks (1–2 arrays per block). These blocks were delineated to distinguish potential differences in road use by the public and thus possible differences in traffic volume. Blocks are separated by three destinations along Alma Bridge Road. Block 1 extends from the Saint Joseph Hill OSP trail to the Los Gatos Rowing Club and includes the road segments at Arrays I and II; Block 2 extends from the Los Gatos Rowing Club to the entrance of the Miller Point Parking Lot and includes the road segment at Array III; Block 3 extends from the Miller Point Parking Lot to the Soda Springs Road junction and includes the road segments at Arrays IV and V; and Block 4 extends from the Soda Springs Road junction to the intersection of Alma Bridge Road and Aldercroft Heights Road and includes the road segment at Array VI (Figure 1). Mortality rates are expected to differ between the four blocks due to differences in various aspects of vehicular traffic including the number of vehicles and the timing of travel with respect to newt activity. For example, we expect that most of the traffic to and from the Los Gatos Rowing Club comes from Highway 17, which could cause differences in traffic patterns between Block 1 and the other blocks. Although we did not measure differences in traffic volume between blocks (because we utilized only one traffic counter, see below), it is likely that they are largely responsible for the between-block differences we detected in newt mortality rates (see section Results), although other factors might also contribute to the between-block differences.
We placed each array at the downslope (reservoir) side of Alma Bridge Road to intercept and capture newts that moved from the uplands, and that successfully crossed the road, towards the reservoir. The barrier consisted of silt fence buried at least 15.2 cm (6 inches) below ground and extending 61 cm (24 inches) above ground (Figure 2). All arrays were initially planned to be of equal length, but due to site-specific limitations (e.g., the lack of a sufficiently wide shoulder in which to construct the arrays), the arrays ranged in length from approximately 36.6 to 57.9 m (120 to 190 feet) as follows: Array I at 39.6 m (130 feet), Array II at 57.9 meters (190 feet), Array III at 57.9 meters (190 feet), Array IV at 51.8 m (170 feet), Array V at 36.6 m (120 feet), and Array VI at 36.6 m (120 feet). We installed a series of paired pitfall traps at each array; at each pair, one trap was on the upslope side (front trap) and the other trap was on the downslope side (back trap) of the array. The outermost pairs of pitfall traps were located 1.5 m from the ends of each array, with intervening pairs of traps spaced at 3-m intervals. There were a total of 24 traps (12 pairs) at Array I, 36 traps (18 pairs) at Arrays II and III, 32 traps (16 pairs) at Array IV, and 22 traps (11 pairs) at Arrays V and VI. This placement was designed to intercept both newts traveling to the reservoir to breed and newts traveling from the reservoir back to the uplands upslope of the road after breeding. The pitfall traps were flush with the array so that a newt would fall into the trap as it was walking along the array in an attempt to cross to the other side (Figure 2).

Figure 2. (A) Array V showing pairs of traps on the front (upslope) and back (downslope) side of the array. (B) Pitfall trap at Array V flush with ground and against the array.
We began the surveys on 4 November 2020, conducting daily inspections of the arrays and adjacent road segments at the arrays, and ending the surveys on 31 March 2021. California and rough-skinned newts breed during the wet season, moving from upland refugia to breeding waterbodies after the fall/winter rains begin and breeding before moving back into upland areas (Stebbins and McGinnis, 2012). Therefore, we expected that the period of early November through March would encompass the breeding season for the newts and would allow us to determine the peak(s) of newt movement across the road. During each daily inspection, we inspected each road segment adjacent to an array for newt carcasses, and the road segment 15.2 m (50 ft) to the north and 15.2 m to the south of the array (north and south “wings”). As a newt carcass was identified on the road, it was photographed, georeferenced, and then removed from the road so that it was not counted subsequently. We did not attempt to identify dead newts to species, sex, or life stage, because the poor condition of most DOR individuals made determination of these variables difficult or impossible. Thus, DOR counts at each array likely include some individuals of each of the two newt species present in the area. We conducted the inspections at the same time each morning to standardize results (with a starting time of 9 a.m. at Array I). This design followed the recommendations of Santos et al. (2011), who advise that road mortality surveys for salamanders be conducted daily to minimize the negative bias in counts that arises if carcass persistence times are short. At each array, after inspecting the adjacent road segment and its “wings,” we then inspected the pitfall traps for newts. We counted and categorized each newt (i.e., species, age-class, and sex) in each pitfall trap on the upslope (road) side of the array (front trap) and released the newt to the nearest suitable habitat on the downslope (reservoir) side of the array, under the assumption that newts captured in the front (road side) traps were moving toward the reservoir when they encountered the array. We then repeated this procedure for newts in the pitfall traps on the downslope (reservoir) side of the array (back trap), releasing the newts to the nearest suitable habitat on the upslope side (across the road) under the assumption that these newts were moving away from the reservoir.
Strategies to reduce wildlife road mortality have become a component of many conservation efforts (Glista et al., 2009; Beebee, 2013). However, their success depends on knowledge of the spatiotemporal patterns of mortality. Therefore, we also investigated hourly variation in traffic level and the relationship between newt road crossing attempts and weather to better understand where the ‘hotspots” (i.e., areas of high mortality) and when the “hot moments” (i.e., times of high newt movement) occur to help inform the timing and location of mitigation efforts. To provide information on vehicular activity within the study area, we installed a TRAFx vehicle counter on a guard rail post at 37.196278° -121.984222° between Arrays II and III (Figure 1). The counter was able to take hourly counts of vehicles in both directions over the survey period. We tabulated the hourly counts into daily totals corresponding to the periods in which newt observations were tallied (i.e., the 24 h period from 9:00 a.m. of the previous day to 9:00 a.m. of the day of each newt survey). We also took daily rainfall data from the National Ocean and Atmospheric Administration Lake Kittridge, CA KDGC1 station, approximately 2.6 km west of the study area. The period of recorded precipitation associated with each particular date of newt observations was from 9:15 a.m. of the previous day to 9:00 a.m. of the date of newt survey.
Data analysis
Determining mortality rates and numbers of adult California newts attempting to cross Alma bridge road at arrays
The main analytical objectives of this study were to measure the road-induced mortality rate of California newts attempting to cross Alma Bridge Road and determine whether this mortality is likely to adversely affect the Lexington Reservoir California newt population. Accordingly, we (1) measured the mortality rates of newts at multiple road segments (the arrays), (2) investigated a set of statistical models that related mortality rate to potential explanatory variables, (3) selected from this set a well-supported model that related mortality rate to road location (block), and (4) applied array results and the model to estimate the total number of adult California newts for the whole road (population size) and derive overall road mortality results for the population. We arrived at a single ratio of adult newts killed on the road to the total number of adult newts attempting to cross the road during the breeding migration. We also estimated the ratio of juvenile newts killed on the road to the total number of juveniles attempting to disperse across the road to access upland habitat following metamorphosis. We then incorporated these ratios and our estimate of population size into the population viability model of Gibbs and Shriver (2005) to test the influence of road mortality.
Fortunately for us, annual DOR carcass surveys, started in 2017, were continued by community scientists during the 2020/2021 breeding season concurrently with our pitfall trap surveys.4 Therefore, we were able to combine the 2020/2021 DOR data with our 2020/2021 pitfall trap results to estimate an adult newt breeding number and overall road-based mortality rate of the newts crossing Alma Bridge Road to breed in Lexington Reservoir. The two studies were closely coordinated to avoid duplication of efforts, to avoid double counting of carcasses, and to better inform both studies. Based on the pitfall trap captures (see section Results), which indicated that the vast majority of newts crossing Alma Bridge Road were California newts, we decided to restrict further analysis to this species.
To facilitate data analysis, we made a series of assumptions based on our understanding of California newt breeding biology, breeding migration, array position, survey methodology, and model requirements (Assumptions S1-S7, section 2 in Supplementary material). With these assumptions, we calculated the number of adult California newts attempting to cross the road (Aat,ur) at each array to breed in the reservoir with the following equation:
where Tat,ur is the number of adult California newts captured in the front traps (traps on the upland side of the array), Dur is the number of newts DOR at the array, and Cat,ur is the proportion all newts (including both species, both sexes, and all life stages) captured in the front traps that were adult California newts. This equation applies Assumption S4 (section 2, Supplementary material).
Cat,ur is calculated separately for each array using the following equation:
where Tjt,ur is the number of juvenile California newts, Tag,ur is the number of adult rough-skinned newts, and Tjg,ur is the number of juvenile rough-skinned newts captured in the front traps. We then calculated the survival rate of breeding adult California newts crossing the road at each array to breed in the reservoir (Sat,ur) with:
The corresponding mortality rate (Mat,ur) was calculated thus:
The number of adult California newts that died attempting to cross the road in the upland-to-reservoir direction (Dat,ur) was estimated as:
We were able to directly calculate the number of adult California newts attempting to cross the road in the reservoir-to-upland direction after breeding in the reservoir (Aat,ru) at each array by totaling the number of adult California newts captured in the back traps (traps on the reservoir side of the array) over the entire survey period. However, because the arrays prevented newts from crossing the road at the arrays, and surveyors transported newts captured in the back traps across the road, we used an indirect method to calculate the mortality rate of adult California newts crossing the road after breeding in the reservoir as follows.
We first divided the survey period into 144 one-day periods; as described above, this was done to control for potential differences in mortality rate between directions arising from daily variation in traffic levels and differential timing and location of newt movement between the two directions. Although the survey period lasted 148 days (21 weeks and 1 day), we excluded the first 4 days because traffic counts necessary for the regression modeling of newt mortality did not begin until the fifth day, and no newts were observed in the traps or on the road at the arrays during the initial four-day period. Each one-day period began after the morning check and lasted to the end of the morning check on the following day.
For each combination of day and array, we obtained a prediction of the mortality rate of all newts (all species, life stages, and sexes) attempting to cross the road in the upland-to-reservoir direction on that particular day at that particular array (Md,ur) using an array-based logistical regression model (array model, sections 3 and 4 of Supplementary material).
We then applied Assumption S6 (section 2 of Supplementary material), wherein we assumed that the mortality rate for adult California newts attempting to cross the road at an array in the reservoir-to-upland direction on a particular day (Md,ru) would be equal to Md,ur (the mortality rate predicted by the array model for that particular array on that particular day). Next, for each day/array combination, we multiplied the number of adult California newts captured in the back traps by the appropriate Md,ru value to estimate how many would have been killed on that part of the road that day if the array was not present (i.e., “rescued” adult California newts). For day/array combinations in which no adult California newts were captured in the back traps, the number of rescued adult California newts was zero.
For each array, we summed the number of “rescued” adult California newts across all 144 day periods to derive the total number of “rescued” adult California newts (Rat,ru). We then calculated the estimated mortality rate for the array over the study period for the reservoir-to-upland direction (Mat,ru) by dividing Rat,ru by the total number adult California newts attempting to cross the road in that direction (total number of adult California newts captured in the back traps) (Aat,ru).
Determining mortality rates and numbers of adult California newts attempting to cross Alma Bridge Road in the entire study area
We used the array results, combined with the community scientist data from the same season (2020/2021), to estimate the number of adult California newts attempting to cross Alma Bridge Road in the entire study area when migrating to Lexington Reservoir to breed and the number killed during this breeding migration to, and the reverse migration from, Lexington Reservoir due to vehicular strikes. Because the arrays represent only a portion of Alma Bridge Road, we used statistical models to (1) investigate factors potentially associated with mortality of California newts attempting to cross the road at the arrays, and (2) select the best approach for applying the array-based mortality rates to the other parts of the road.
Statistical models of road mortality and calculation of whole-road mortality rates
We estimated overall, whole-road mortality rates for adult California newts attempting to cross Alma Bridge Road in each direction and the number of adult California newts attempting to breed by combining array results with the results of community scientist observations of DOR newts in road areas outside of the arrays. Rather than assume that mortality rates were constant in time and space, we used statistical models to derive the relationship between mortality and environmental variables. These models, which indicated that mortality rates were not constant, helped us derive rigorous estimates of the whole-road mortality rates and the number of California newts attempting to breed.
In brief, we used regression to analyze the outcome of individual road-crossing attempts at arrays to investigate block and other environmental variables (day, daily number of vehicles, percent cover of uncanopied grassland, canopy cover, average slope, and average aspect) as potentially explanatory variables (sections 3 and 4 of Supplementary material). Based on Aikaike information criterion (AIC) scores, the “complex” model, which contained block, day, and daily number of vehicles as explanatory variables, was the best model for describing mortality of newts attempting to cross the road in the upland-to-reservoir direction (Table 1; section 4 of Supplementary material). This model was selected as the “array” model and was used to estimate mortality rates for adult newts traveling in the opposite (reservoir-to-upland) direction (see Assumption S6, section 2 of Supplementary material) and to build the binomial dataset for that direction (See section 3 of Supplementary material). The “non-block” model, which contained day, daily number of vehicles, and canopy cover as explanatory variables, was the best model for describing mortality of newts attempting to cross the road in the reservoir-to-upland direction (Supplementary Table S1).
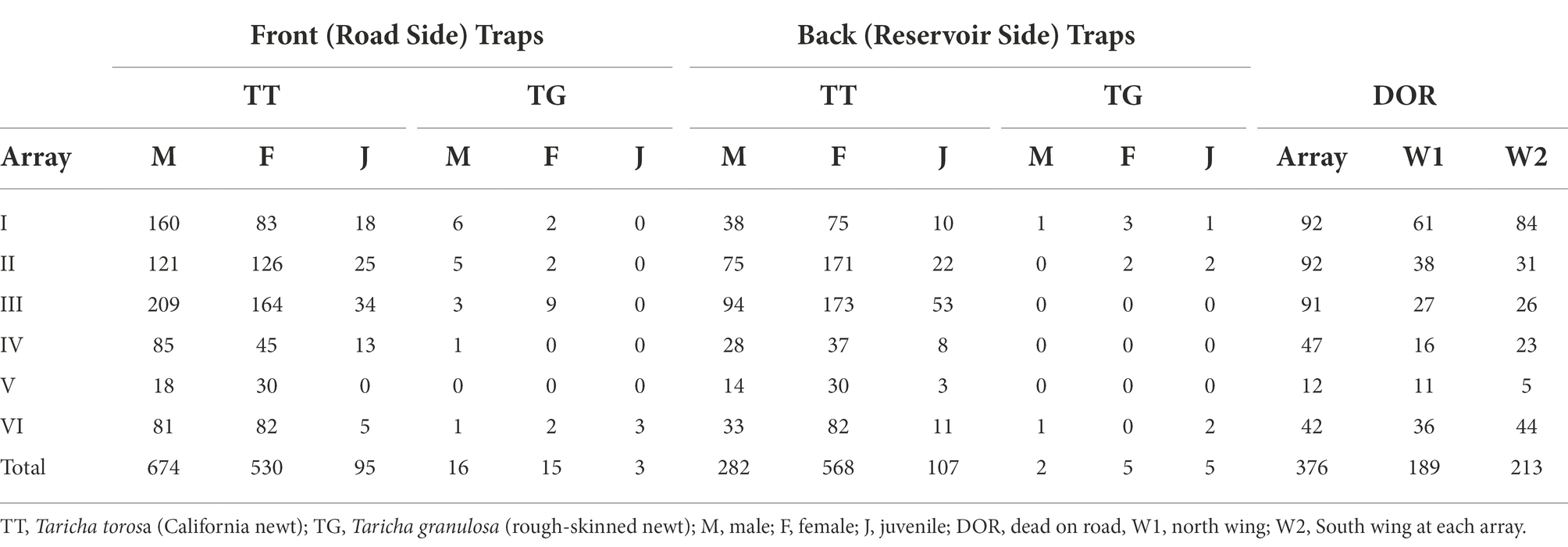
Table 1. Number of newts captured and observed DOR at arrays.
Ideally, we would apply the complex and non-block models to the community scientist observations to estimate mortality rates for the non-array road areas. However, this approach was not possible because both models included day and number of vehicles (a daily count) as explanatory variables. These variables are absent from the community scientist dataset and cannot be incorporated into it, because the community scientist observations were not made every day. Thus, we applied the “block-only” models (sections 3 and 4 of Supplementary material), which outperformed the non-block models and did not require day or daily number of vehicles as model inputs.
We applied the block-only models to obtain direction-specific estimates of the number of California newts attempting to cross the road and the number that died in the attempt for areas located outside of the arrays, using (1), the array-derived mortality rates that were specific to block as well as the newts’ direction of travel, and (2) community scientist counts of DOR newts outside of the arrays (section 5 of Supplementary material). Then, for each direction of newt travel, we totaled the array and non-array estimates of dead and “rescued” adult California newts across all four blocks to obtain an entire-road estimate of overall adult California newts that died on the road. Thus, “rescued” adult California newts were counted as dead, to best estimate what the overall mortality would have been without intervention. We also, for each direction of newt travel, totaled the array and non-array results for number of adult California newts attempting to cross the road across all four blocks. We then calculated an overall mortality rate and an overall survival rate (1 – overall mortality rate) for each direction of newt travel from the entire-road estimates of the number of adult California newts that died on the road and the number that attempted to cross the road. The overall mortality rate for adults attempting to cross the road to breed during the 2020/2021 breeding season was calculated by subtracting the product of the survival rates in each direction from one.
We then applied the estimated number of adult California newts attempting to cross Alma Bridge Road in the upland-to-reservoir direction in order to breed in Lexington Reservoir and the overall mortality rate as two parameters from this study to be used in the aforementioned population viability model under the following assumption:
1. The local population of California newts that reside in the upland habitat at Lexington Reservoir consists of individuals that exclusively breed in Lexington Reservoir and are required to cross Alma Bridge Road in order to breed.
Projecting the impact of road-based mortality on the Lexington reservoir California newt population using a population viability model
We used a model of population viability in order to investigate the impact of this road-related mortality on the local newt population (Gibbs and Shriver, 2005). We identified the parameters and input data that were required to construct this model of population viability to determine the threshold level at which such mortality would be great enough to risk long-term population viability specific for the population at Alma Bridge Road. Some of the data were available from the literature, but other data would need to be obtained through additional fieldwork or by making assumptions.
The model from Gibbs and Shriver (2005) is:
where Na,t is the number of adults in the current year, Ne,t is the number of eggs produced in a given year, σa is the adult annual survival rate, σm is the survival rate from egg to metamorphosis, σj is the survival rate of juveniles through their first winter, and Kl is the carrying capacity of larval habitat.
The number of eggs produced in a given year is calculated as:
where ϕm is the average eggs produced per individual, calculated by multiplying the number of eggs per mass by the number of masses laid per year, and multiplying this number by the probability of successful breeding by females after arrival at the breeding site divided by 2 (to account for the presence of both sexes).
The starting adult population (or current population during a given year) was provided in this study. From this starting adult population, a starting average number of eggs ϕm was calculated by multiplying the number of individuals with the average clutch size and the average number of clutches per year. For the California newt, this average number of eggs was not available in the literature; instead the literature provided a range of 7–47 eggs per egg mass and a range of 3–6 egg masses laid by a female during the breeding season (Brame Jr., 1968) in addition to an ovarian count range of 130–160 per female (Thomson et al., 2016). We therefore made the assumption that the average number of egg masses laid is 4.5, with an average number of eggs per egg mass of 32.2, to arrive at a midrange ovarian count of 145.
A study by Jones et al. (2017) used a much lower average of 60 eggs (24 eggs in 2.5 egg masses) laid per female during a breeding season. They base this average 60 eggs laid per female on Kats et al. (2013) who state in the introduction that “after breeding, female newts remain in the streams to deposit small (2–3 cm diameter, 15–30 embryos each) egg masses…”; and on Brame Jr (1968) who referenced Ritter (1897) stating that the average number of eggs produced at any one laying… “seems to be about 60 for each female, these being distributed in three or four of the masses.” Yet, Brame in the same study observed one female laying six egg masses, with an average of 22.1 eggs per mass for a total of 133 eggs, and suggested that his higher counts may reflect differences in geographic regions between his observations and those of Ritter. We however, consider our average of 145 eggs (32.2 eggs per 4.5 egg masses) to be a better indicator of average number of eggs laid by females based on an egg mass range of 3–6 egg masses and an ovarian count range of 130–160 per female as provided in Thomson et al. (2016). Therefore, we kept our average number of 145 eggs per female instead of the much lower 60 eggs per female.
Jones et al. (2017) suggested that females migrate to breeding sites annually and breed through most of their adult lives but that the probability of successful breeding depends on the availability of appropriate breeding habitat. However, Thomson et al. (2016) state that it is unknown whether females breed every year or skip years, and Stebbins and McGinnis (2012) state that “individual newts probably do not breed every year.” Similarly, they state that for red-bellied newts “males breed every year, but females rarely breed more frequently than at two- or three-year intervals or longer.” This statement is based on mark-recapture research on homing ability in red-bellied newts, with an interpretation of results speculating on the frequency of breeding by males and females (Twitty, 1961, 1964). Therefore, to be conservative, we decided to set the probability of successful breeding by females, given that they survive the breeding migration and arrive at the breeding site, to 0.5. However, we assumed that females and males in our study population migrate to breeding sites annually because the sex ratio we observed in migrating adults was approximately equal (see section Results).
Also, because we were unable to find a previously reported survival rate from egg to metamorphosis in the literature, we followed Jones et al. (2017) in using a proxy of 0.025 for tiger salamanders (Ambystoma tigrinum). This rate may correspond to the potential effects of predation by fish in the reservoir. Crayfish and mosquitofish (Gambusia affinis) are known to prey on newt larvae (Gamradt and Kats, 1996), which do not retain the toxin present in the egg mass after yolk absorption (Twitty and Johnson, 1934; Twitty, 1937). Other predatory fishes, such as native rainbow trout (Oncorhynchus mykiss) and nonnative green sunfish (Lepomis cyanellus), are commonly abundant in streams devoid of newts, indicating that these fish may exclude newts in some situations (Kuchta, 2005). These fishes are present in Lexington Reservoir5, suggesting that predation might be a significant source of larval mortality in our study population.
Carrying capacity of the reservoir for California newts is unknown. Due to the size of the reservoir, one could assume that the carrying capacity is very large (e.g., into the millions of larvae). However, carrying capacity depends on the abundance of aquatic invertebrate food source for the larvae in the reservoir, which may be quite small, as reservoirs are considered less productive than other natural waterbodies of the same size typically due to a limited littoral zone (Moss, 2008). We therefore, provided a carrying capacity of the reservoir as the larval habitat of 2,000,000 (an admittedly speculative number estimated as four times the starting number of eggs).
To model the population in the absence of road mortality, we used a baseline value of σj (survival rate of juveniles through their first winter) equal to 0.7917, which was taken from the results of Jones et al. (2017). Following Jones et al. (2017), baseline σa (adult annual survival) was set at 0.91, a value based on a study on red-bellied newts by Twitty (1966). To model the population in the presence of mortality on Alma Bridge Road, we recalculated σj and σa under the following assumptions:
2. A juvenile newt will cross the road exactly once when dispersing to the upland from the reservoir after metamorphosis.
3. Adult newts attempting to breed attempt to cross the road once in the upland to reservoir direction. The adults that attempt to cross the road in the upland to reservoir direction during a breeding migration subsequently attempt to cross the road in the other direction to return to the upland habitat.
4. Each attempted road crossing is a separate event unrelated to the baseline mortality rate.
Thus, for the road mortality scenario, σj was calculated as the baseline σj multiplied by the overall survival rate of newts crossing in the reservoir to upland direction, and σa was set equal to baseline σa multiplied by the overall survival rate of newts crossing in the reservoir to upland direction and the overall rate of survival rate of newts crossing in the other direction.
Results
We captured a total of 2,302 newts in the pit fall traps: 1,333 newts in the upslope or road side (front) traps, and 969 newts in the downslope or reservoir side (back) traps. We also counted 376 newts DOR at the arrays and an additional 402 newts DOR at the wings. Of the newts captured in the traps, 2,256 (98%) were California newts and 45 (2%) were rough-skinned newts. Of the California newts captured, 956 (42%) were adult males, 1,098 (49%) were adult females, and 202 (9%) were juveniles (individuals that were under 2.5 inches total length and lacked breeding adult characteristics such as smooth skin and flattened tail in both sexes, and enlarged tailfin, swollen cloaca, and nuptial pads on finger tips in males). Of the rough-skinned newts captured, 18 (40%) were adult males, 20 (44%) were adult females, and 8 (16%) were juveniles. A breakdown of the numbers of newts observed DOR and captured at each array is provided in Table 1. Here, because the arrays differed in length, we standardized results for each array by multiplying the number of individuals capture at that array and DOR at that array by 57.9 (the length of the longest array in meters) divided by the length of the target array. These standardized numbers are provided in Table 2. These standardized numbers indicate that Array I has the highest DOR counts, followed by Arrays II and III, then Array VI, Array IV, and finally Array V. At least 17 other vertebrate species were observed during array-based and community scientist surveys during the 2020/2021 newt breeding season, but the carcasses of each non-newt vertebrate species were at least two orders of magnitude less abundant than newt carcasses (section 9 of Supplementary material).
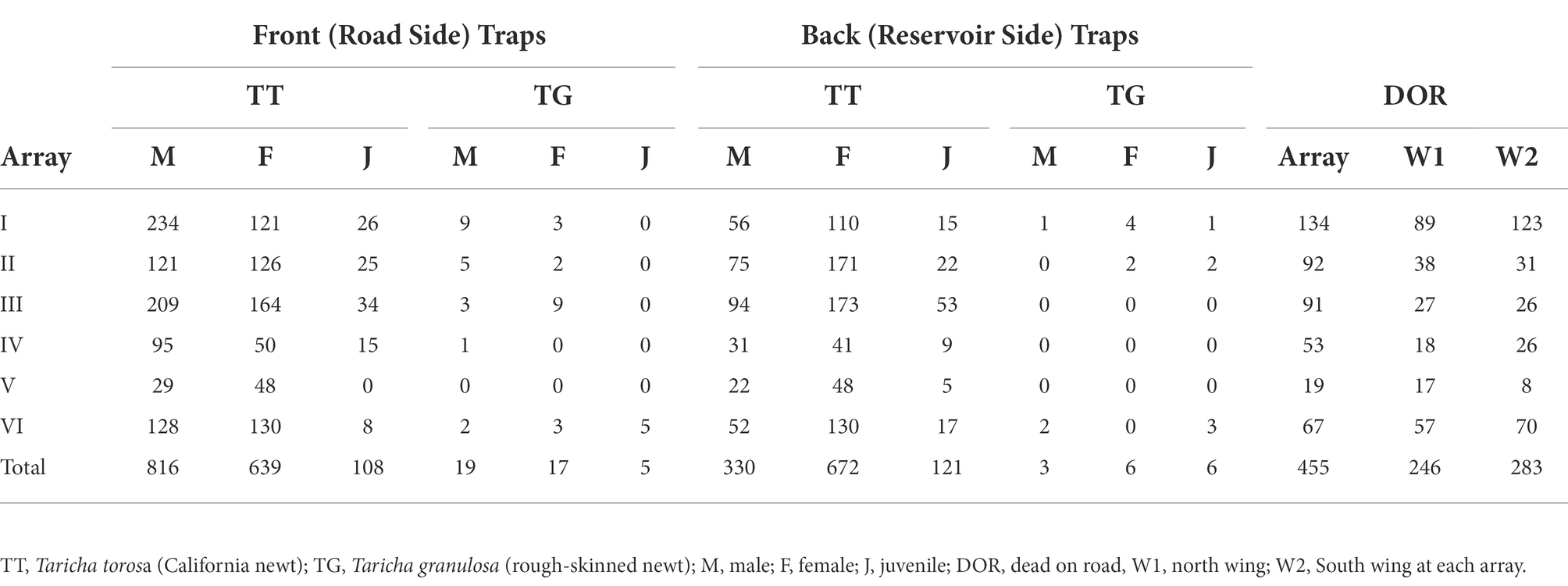
Table 2. Standardized numbers of newts captured and observed DOR at arrays.
Adult California newt crossing attempts and road crossing mortality rates at arrays
Table 3 provides the estimates of the number of adult California newts attempting to cross Alma Bridge Road at the arrays in the upland-to-reservoir direction and the mortality rates due to vehicular strikes. These adult California newt numbers and mortality rates are directly calculated by the number of adult California newts captured in the front traps, contribution of adult California newts to all newts captured in the front traps, and newts DOR at each array. The mortality rates are relatively similar between the six arrays (range of 18 to 25 percent) but are higher at Arrays I, II, and IV than at Arrays III, V, and VI. The higher mortality rates at Arrays I, II, and IV, even though the total number of newts crossing the road at Array III is the highest among all arrays, indicate that the road segments at Arrays I, II, and IV were possibly experiencing higher vehicular use due to their locations at or near recreation-associated parking areas (which are not present in the other segments).
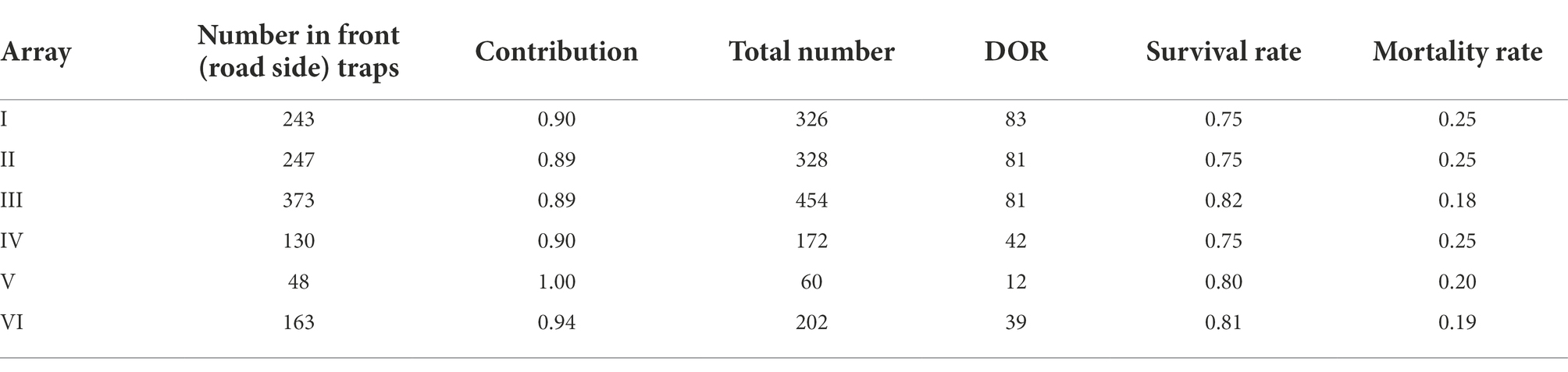
Table 3. Number of adult California newts caught in front (road side) traps, contribution to all newts caught in front (road side) traps, estimated number attempting to cross Alma Bridge Road in upland to reservoir direction at each array, and survival and mortality rates for that direction.
Table 4 provides the number of adult California newts attempting to cross Alma Bridge Road at the arrays in the reservoir-to-upland direction based on the number of adult California newts captured in the back (reservoir side) traps, and the estimated number of “rescued” adult California newts based on the day-specific rates of mortality from vehicular strikes for newts attempting to cross the road predicted by the array model. Estimated mortality rates for Arrays I, IV, V, and VI were very similar, falling within a range of 6 percentage points (19–25%). Estimated mortality rate was relatively high at Array II (27%) and relatively low at Array III (16%) because of two factors: (1) the directly observed mortality rates for the upland-to-reservoir direction were relatively high at Block 1 arrays (Arrays I and II) and relatively low at the single array in Block 2 (Array III), which heavily influenced the array model upon which the reservoir-to-upland estimates were based; and (2) newts attempting to cross the road in the reservoir-to-upland direction at Arrays II and III happened to time their crossings on days that also happened to be relatively high and relatively low, respectively, in risk of road mortality. Nevertheless, mortality rates for the reservoir-to-upland direction were fairly consistent; all rates for this direction fell within a range of 11 percentage points (16 to 27%). Thus, across our estimates of mortality for each direction at each of the six arrays, there was a difference of only 11 percentage points between the highest and lowest estimate. This consistency suggests that our sampling regime was sufficient in geographic scope to represent the range of variation in mortality rate along the entire road and further suggests that our estimates of whole-road mortality rates for each direction of travel, which are derived from the array-level estimates, are reasonably accurate.
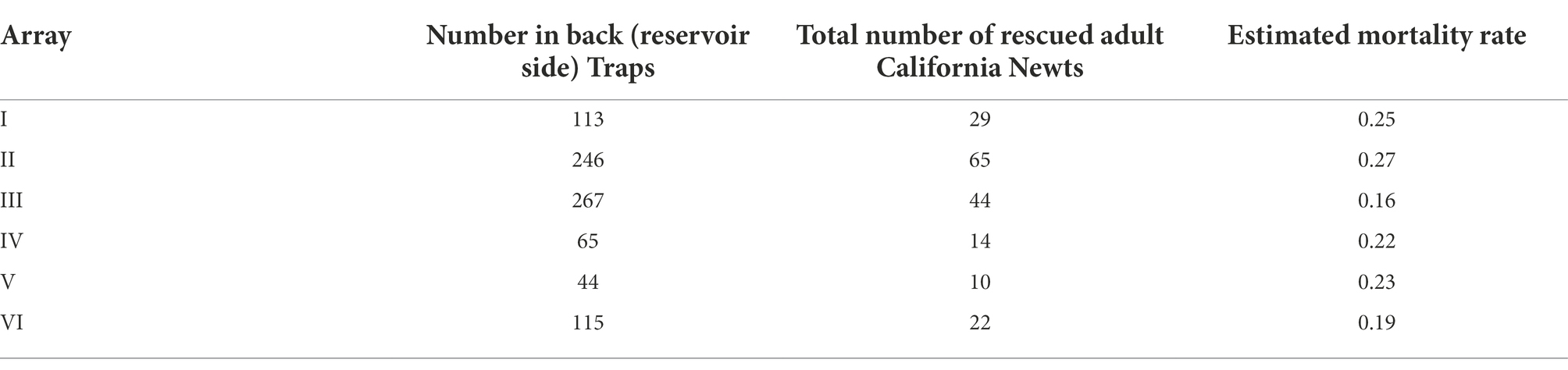
Table 4. Number of adult California newts caught in back (reservoir side) traps, estimated number of “rescued adult California newts”, and mortality rate at each array for adult California newts attempting to cross Alma Bridge Road in reservoir-to-upland direction.
Adult California newt crossing attempts and overall mortality rate
Based on our analyses, which use the newt capture results from the arrays in combination with observations of newts DOR at the arrays and segments of Alma Bridge Road located outside of the arrays, we estimate that at least 13,786 adult California newts attempted to cross the entire Alma Bridge Road in the study area in order to migrate to Lexington Reservoir to breed during the survey period. This number is the sum of our estimates of the number of adult California newts that attempted to cross the road at each array and the number that attempted to cross at the road segments of the four different blocks that are outside of the arrays. Applying the mortality rates estimated for each array and each non-array road segment, we also estimate that 3,066 adult California newts died on the road due to vehicular strikes during this migration, for a road-based mortality rate of 22.2%. Also, based on our captures of newts moving in the reservoir-to-upland direction, in combination with DOR observations from the non-array road segments, we estimate road-based mortality rate for this return migration to be 21.9%. These estimates indicate that of the 13,786 adult California newts migrating from the upland across Alma Bridge Road to the reservoir to breed, 10,720 adult California newts successfully crossed and potentially bred in the reservoir. We presume that a large proportion of these adult California newts then attempted to cross Alma Bridge Road on the return migration back to the uplands, though many were likely still on the reservoir side of the road at the end of the survey period. It is likely that these adult California newts will continue to migrate back to the uplands as conditions allow during the summer and fall, for example, during rain events or periods of high humidity at night (i.e., foggy nights).
The areas of high mortality, shown in Figure 3, were at the sections of Alma Bridge Road from Limekiln Creek south to half way between Arrays II and III, then from the curve and road section north of the Miller Point parking lot, including Array III but before Array IV, and from the junction with Soda Springs Road westward along the south side of Soda Springs Creek (including Array VI), with “hotspots” of very high mortality before and after Array I (Figure 3). Because an adult breeding California newt is required to cross the road twice to successfully survive a round trip migration, it is subject to an upland-to-reservoir mortality and a reservoir-to-upland mortality. We calculated the roundtrip survival rate of crossing the road to be 60.8% percent and therefore estimate the overall road mortality rate for migrations in both directions during the survey period to be 39.2%.
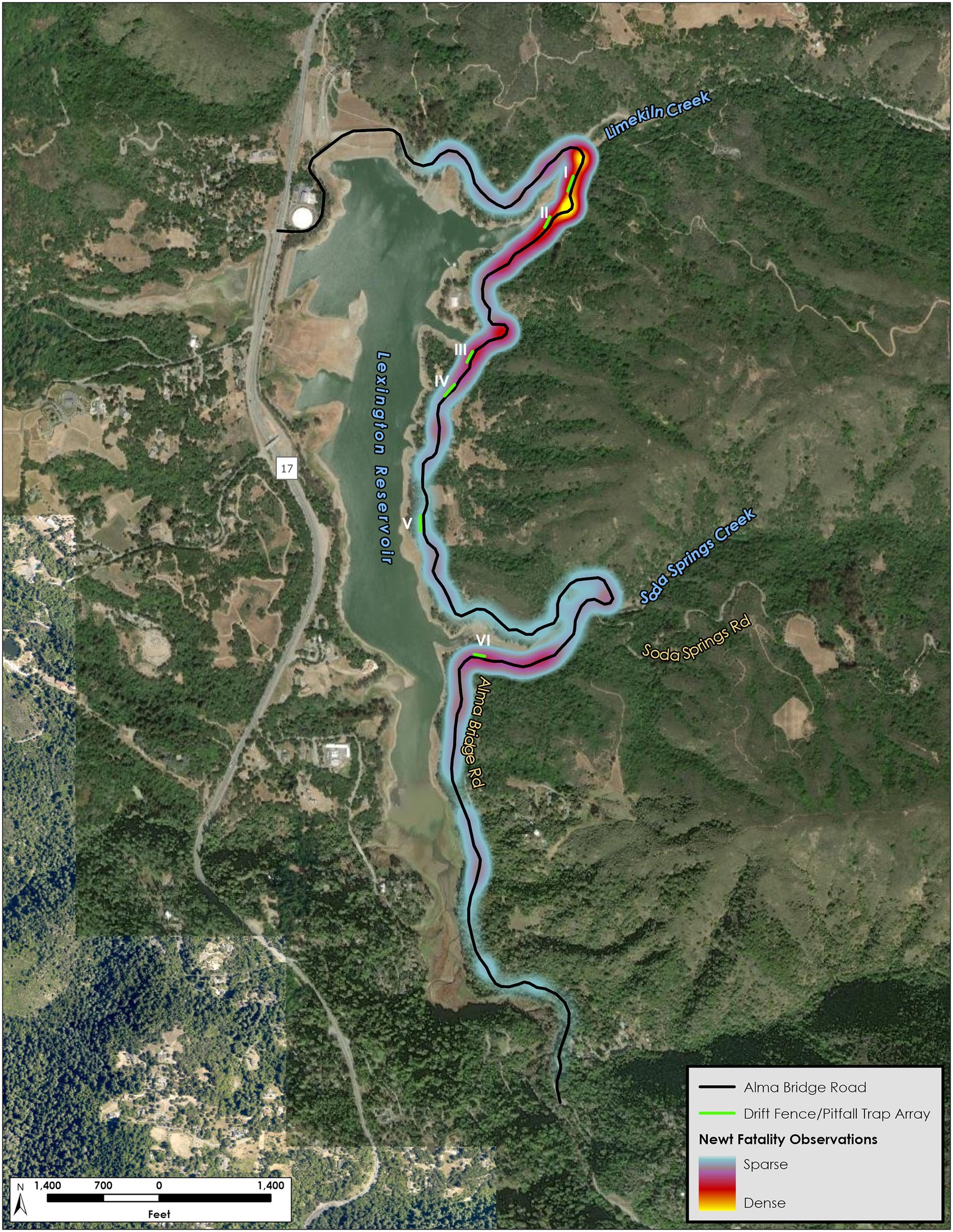
Figure 3. Newt mortality density map.
Population viability model and expected impact of road mortality on the Lexington reservoir California newt population
We incorporated these values of adult California newts attempting to cross the road in the upland-to-reservoir direction (13,786) and the estimated road mortality rates into the parameters of the population viability model of Gibbs and Shriver (2005), with all other values coming from the literature as provided in the section Materials and Methods, to determine how the Lexington Reservoir California newt population is expected to change given these new model inputs.
The graph in Figure 4 illustrates the results. As indicated in this graph, the population is predicted to quickly decline to under 1,000 adult California newts in 23 years and may be extirpated in 92 years (red trend line in Figure 4).
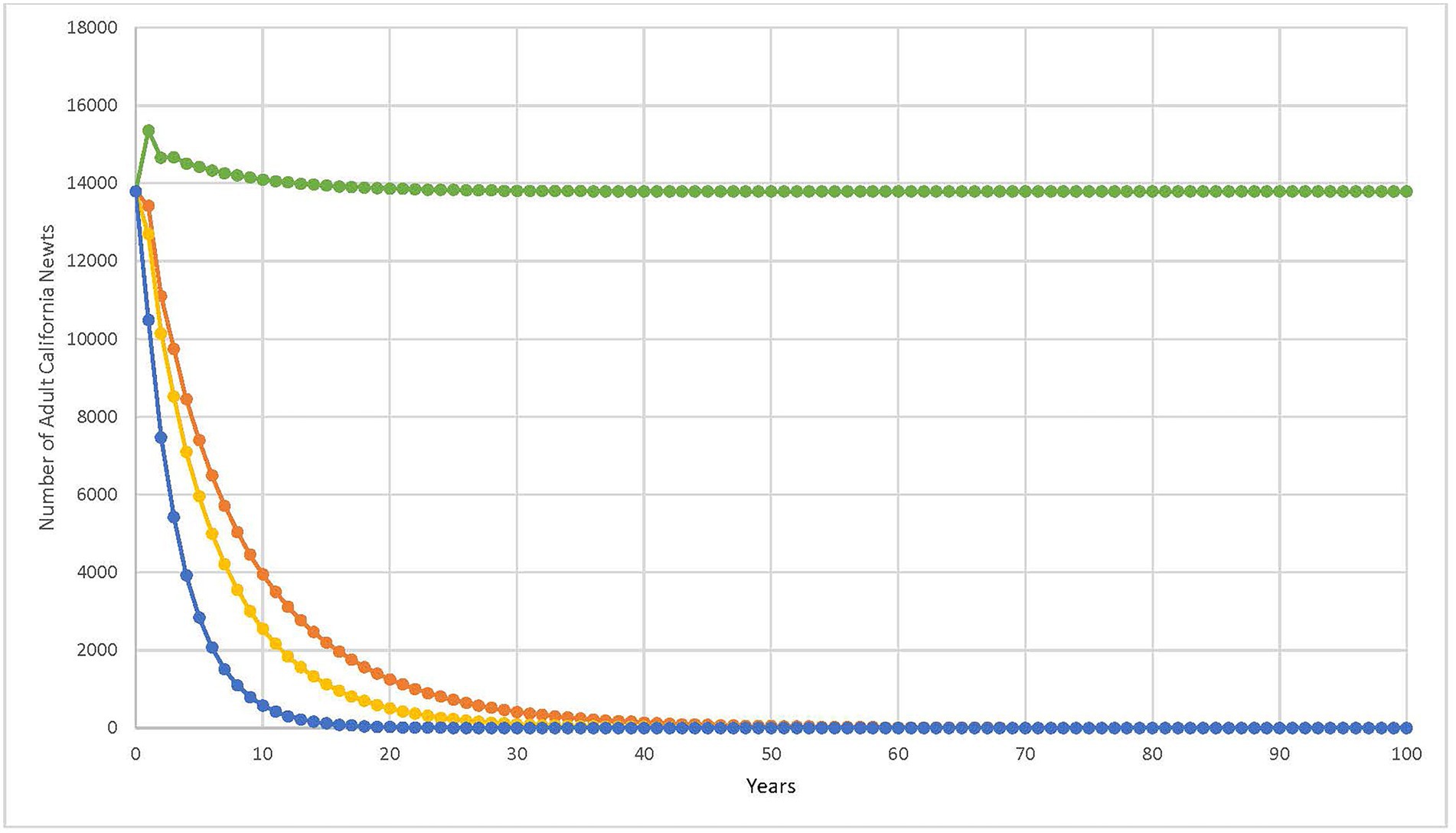
Figure 4. Population viability model with starting adult California newt number and mortality rate from current study. Red trend line using parameters in current study. Green trend line using parameters in current study, but changing to breeding every year. Yellow trend line using parameters in current study, but changing breeding every year and average of 60 eggs per female. Blue trend line using parameters in current study, but changing average of 60 eggs per female.
Correlation of newt movement with precipitation
Of the 148 days of the survey period, 45 days received precipitation. These “rain days” were distributed as 11 single days, four pairs of consecutive rain days, three groups of three, three groups of four, and a single group of five consecutive rain days, interspersed with days of no measurable precipitation “dry days.” The longest period of consecutive dry days was a 22-day period from 20 November 2020 to 11 December 2020, followed by two 13-day periods from 9 January 2021 to 21 January 21 2021 and from 21 February 2021 to 4 March 2021, and a 10-day period from 22 March 2021 to the end of the survey period of 31 March 2021.
As expected, the majority of newt capture and newt DOR observations at arrays occurred during and after rain days, and the counts of newt capture and/or DOR observations decreased (with a few exceptions) as the period of consecutive dry days lengthened. The longest string of consecutive days with counts of newt captures and DOR is 34 days from 22 January to 24 February 2021, with a total count of 1,287 newts. This period also contained 18 rain days, with two of four consecutive rain days (27 and 28 January) receiving 2.96 and 4.21 inches of rain, respectively, the highest single-day rain totals in the survey period. This period also contained the only five consecutive rain day period from 12 February to 16 February 2021.
The highest single-day count of newts captured and DOR was 6 March 2021 at 278. This date was near the beginning of a 26-day string of consecutive days with counts of newt captures and DOR, totaling 836. This period also contained nine rain days (three single rain days, one pair of consecutive rain days, and one group of three consecutive rain days) interspersed with 18 dry days. In fact, 59 newts were captured and observed DOR in the 10 consecutive dry day period that extended to the end of the survey period (Figure 5), indicating that newts continued to move until the end of the survey period even though major rain events were over for the season.
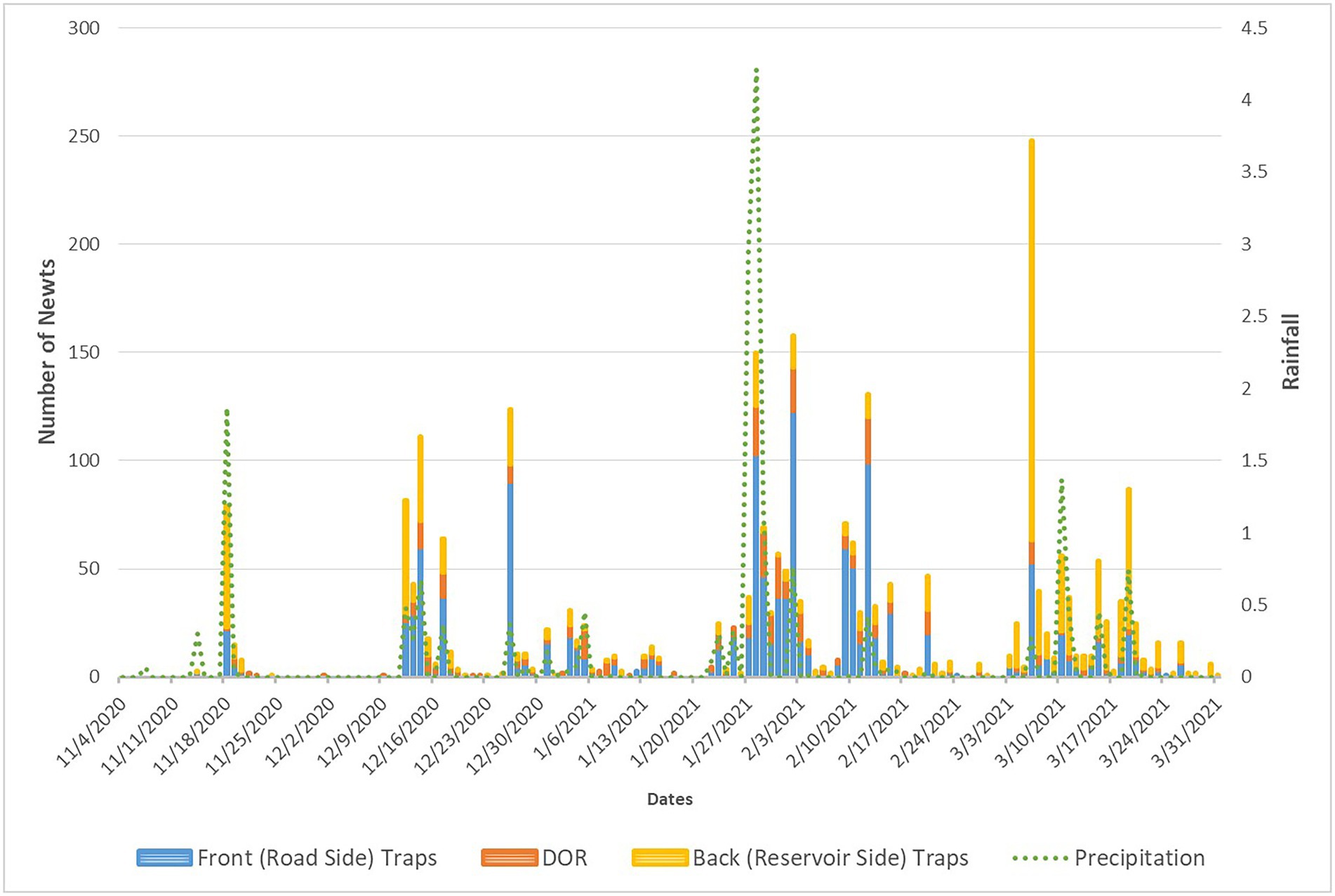
Figure 5. Graph of newt capture and DOR at arrays in relation with precipitation.
Approximately half of the newts were observed crossing the road at the arrays (i.e., observed captured in traps and/or DOR at arrays and wings) during just 12 of the 148 days (8%) of the survey period, all associated with rain events (only one of these 12 days was immediately before a rain event). If we were to include all rain events plus two dry days after the last rain day in order to include a lag time of newt movement after a rain event, this would encompass 81 days (or 55% of the survey period) and, 91% of newt movement across the road at the arrays. So, approximately 50% of newt movement occurred during 8% of the survey period, and 91% occurred during 55% of the survey period, associated with rain events.
There also appeared to be a switch in the relative abundance of newt movement from one direction to the other at the beginning of the survey period and near the end of the survey period during the last week in February. During the initial rain events from November 2020 to 12 December 2020 more newts were captured in the back (reservoir side) traps than DOR at the arrays and/or captured in the front (road side) traps (Figure 5). Then from 13 December 2020 to 20 February 2021, more newts were typically recorded DOR at the arrays and/or captured in the front (road side) traps than captured in the back (reservoir side) traps (Figure 5). Then, from 21 February 2021 to the end of the survey period on 31 March 2021 more newts were typically captured in the back (reservoir side) traps than DOR at the arrays and/or captured in the front (road side) traps (Figure 5).
We interpret these patterns as follows: (1) newts were moving in both directions and crossing the road throughout the rainy season; however, (2) from the initiation of rain events in the fall to 12 December 2020 (day 39 of the survey period), most newts crossing the road were adult newts that most likely bred in the previous year and may have been continuing to migrate away from the reservoir to the upland; then, (3) from 13 December 2020 (day 40) to 20 February 2021 (day 109) most newts crossing the road were adults migrating from the upland to the reservoir to breed during this breeding season, with the peak of this migration occurring around 2 February 2021 (day 91); finally (4) from 21 February 2021 (day 110) to the end of the survey period (day 148), most newts crossing the road were adults migrating from the reservoir to the uplands after having bred during this breeding season, with a peak of this reverse migration occurring on 3 March 2021 (day 121; Figure 6). We also conclude that the rain events and 2 day lag time during early February and early March would constitute “hot moments” of newt movement.
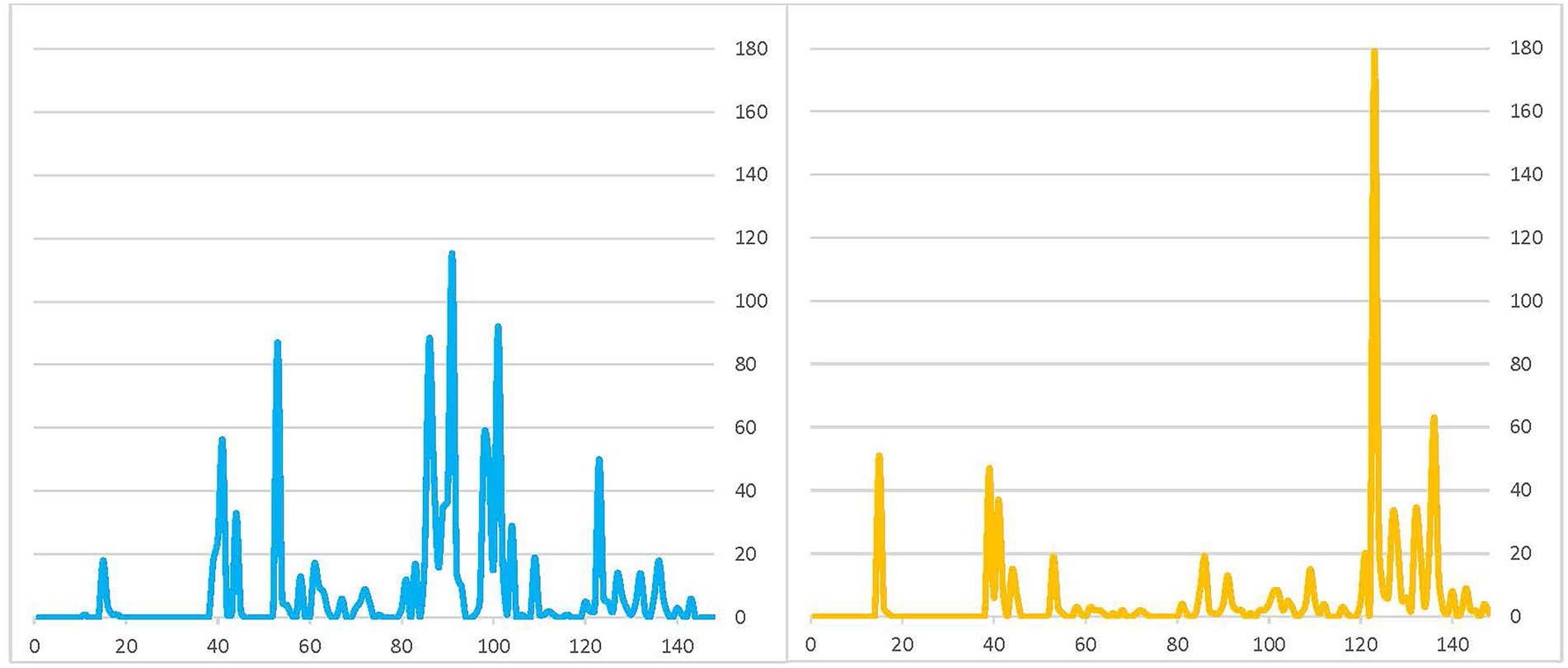
Figure 6. Graph of numbers of adult newts caught in front (road side) traps (blue) versus back (reservoir side) traps (orange) over the 148-day survey period.
Discussion
The results of the population viability model indicate that the local California newt population using Lexington Reservoir for breeding may be extirpated in 92 years. However, Alma Bridge Road has been in use for approximately 67 years. Therefore, it is important to discuss why this population is still extant and experiencing such a potentially high road-related mortality rate.
One reason may be that the road-based mortality rate was much lower in the past and has increased due to increased traffic volume on Alma Bridge Road. During our study there was an average of 577 vehicles per day (see section 7 of Supplementary material). This current traffic volume may have resulted from the start of and increased operations of the Lexington Quarry, increased recreational traffic with the opening of the Los Gatos Rowing Club and trails and park through the open space, and an expansion of private development in the south. The road may also have received increased overflow traffic from Highway 17 when it has become congested over the years. We have very little historical data on traffic using this road since its construction. For example, one set of data over a three-day period from 13 October through 15 October 2015 that we have examined (internal records from Santa Clara County) gives an average daily count of 194 vehicles. This count is below the lowest daily count in our study (233). Another more recent two-day count from 15 March through 16 March 2019 has an average daily count of 289 (internal records from Midpeninsula Regional Open Space District provided on 3 June 2021). If more data are available and show that traffic volume has steadily or dramatically increased over the decades then it is likely that the road-based mortality rate has also increased in concert with this increased traffic volume.
Another possibility of why the newt population is extant is a higher annual breeding potential of the females than we used in the model. If we were to re-run the population viability model, increasing the probability of success in a female’s breeding attempt, given that she reaches the breeding site, from 0.5 (as assumed in our original model) to 1.0 (all breeding attempts by females at the breeding site are successful) the adult population size would be sustained at approximately the same level in 60 years (green trend line in Figure 4). Jones et al. (2017), like our study, assumed that females attempt breeding annually. But Jones et al. (2017) also reasoned that the success of female breeding attempts is limited by the amount of habitat available for egg laying, particularly in the streams they were investigating; egg laying habitat decreases in these streams during drought conditions. We assume that in the case of Lexington Reservoir, there is not a limitation in egg-laying habitat, as the reservoir will maintain the littoral zone that newts would use regardless of annual rainfall, and therefore would be available every time a female attempts to breed and lay eggs. Therefore, it is possible that adult females achieve close to 100% success during breeding attempts after reaching the breeding site, given that Lexington Reservoir is available to them for breeding and egg laying each year, and that this breeding frequency is sufficient to sustain the population (though at a lower number) despite the high mortality rates we estimated.
However, if we were to decrease the average number of eggs per female to 60 as in the Jones et al. (2017) study, even with successful breeding occurring annually, the model results would be similar to our original results (yellow trend line in Figure 4), with the population becoming extirpated in 63 years; and if successful breeding occurred every other year, the population would be extirpated in 32 years (blue trend line in Figure 4). This exercise illustrates the importance of knowing these other parameters, particularly the average annual reproductive output of the female.
Our study assumes that all adults in the California newt population attempt to breed (i.e., undergo the breeding migration) every year. In some salamander populations, males may attempt to breed every year while females skip at least some years between attempts, foregoing the breeding migration in some years in order to avoid unfavorable conditions or to acquire energy for use in later breeding attempts. Such a scenario should result in a male-skewed sex ratio among adult salamanders undergoing the breeding migration. In contrast, we observed an approximately equal sex ratio (1.15 females, 1 male) among migrating adults, consistent with males and females alike undergoing the breeding migration every year.
The model is also conditioned on Lexington Reservoir and inlet streams on the reservoir side of Alma Bridge Road being the only breeding source for this population and the adult newts in this population crossing Alma Bridge Road to breed (see Assumption 1 in section Materials and Methods). However, it is possible that there are adult newts in other upland areas around Lexington Reservoir that are breeding in the reservoir without crossing Alma Bridge Road. We suspect though that the number of newts in these other areas (i.e., areas south and west of Lexington Reservoir) is much lower than those in the upland east of Lexington Reservoir, on the other side of Alma Bridge Road, due to the relatively limited amount of upland habitat available to these newts for dispersal and refugia as a result of more residential development and Highway 17 in these areas. Also, there may be other breeding sources for this population besides Lexington Reservoir. For example, newts may breed in the upper reaches of Limekiln Creek (approximately 16 km of creek distance upstream of Alma Bridge Road) and Soda Springs Creek (approximately 19 km of creek distance upstream of Alma Bridge Road). If newts are breeding in Lexington Reservoir without crossing Alma Bridge Road or are also breeding in these other locations, annual recruitments from Lexington Reservoir or these other sources might be sustaining or supplementing the population, even though the high mortality rate of crossing Alma Bridge Road to breed in Lexington Reservoir would represent a population sink for the overall metapopulation. However, if these other breeding sources do exist, they may become less reliable in the future due to increased drought conditions as a result of climate change (Jones et al., 2017).
Assuming that all parameters and assumptions for the population viability model are correct, resulting in a possible extirpation of the local California newt population at Lexington Reservoir in 92 years, then reducing the roundtrip road-based mortality rate from the current 39.2 to 24.95% would allow the population to be sustained at approximately its current size after 132 years. These road-based mortality rates should therefore be considered when examining potential corrective measures to reduce the negative impact of this road-based mortality on this population.
Our population projections use our best estimates of the true survival rates of juveniles and adults in the Lexington Reservoir population in an average year given the available data. However, our juvenile survival rate might be less accurate than our adult survival rate. Adult California newts travelling to breeding sites have been observed during both daytime and nighttime hours, primarily during wet months of the year (Stebbins and McGinnis, 2012). In contrast, larvae of this species metamorphose and leave the aquatic breeding site for the terrestrial environment during the dry season (summer and fall; Kuchta, 2005). They likely travel at night, when temperatures and risk of desiccation are lowest. Traffic levels should also be lower at night (see section 8 of Supplementary material), resulting in lower risk of vehicular strike for juveniles attempting to cross Alma Bridge Road during the dry season. Dry season movement of juveniles appears to explain why we captured few juvenile California newts attempting to cross the road during the 2020/2021 wet season; juvenile captures accounted for only 9% of all California newt captures at the trap arrays. This runs counter to expectation, based on observations of another drift fence/pit fall trap study of California tiger salamanders in Alameda County (H. T. Harvey & Associates, 2004), where juveniles outnumbered adults in the captures, suggesting that the juveniles we captured represent only a small fraction of the total number of juveniles in the Lexington Reservoir population. The juveniles we captured metamorphosed the previous year and likely emerged from underground retreats to continue migrating away from the reservoir or forage for prey at the surface. Most juveniles in this cohort probably crossed the road in the previous dry season. In essence, we applied the reservoir-to-upland road mortality rate measured during the wet season to these dry-season crossings in preference to ignoring juvenile mortality. To our knowledge, no studies have directly measured the success rate of juvenile salamanders that attempt to cross a road during the dry season, but incorporation of these direct measurements would increase confidence in the parameterization of our model.
Another source of uncertainty in our population viability model is the estimate of the number of adults attempting to cross the road to breed. The actual number was likely somewhat higher. Data from carcass persistence trials conducted by the community scientists in the study area during the same breeding season6 (accessed on February 8, 2022) suggest that some carcasses were removed by subsequent vehicle strikes, rain, or scavengers before they could be counted in the community scientist DOR surveys. Because these surveys were generally conducted twice per week (Parsons, 2021), carcass removal between surveys likely lead to underestimation of the true number of DOR in non-array road segments, underestimation of the total number of newts attempting to cross the road, and overestimation of time-to-extirpation. In contrast, our estimates of mortality rate for road-crossing attempts are largely robust to potential undercounting from carcass removal and imperfect searcher efficiency, because they were measured using array-based surveys, which were conducted every 24 h. Our examination of the carcass persistence results (M. Vonshak, unpublished data) suggests that the daily schedule allowed surveyors several chances to detect most carcasses.
Our study demonstrates how the population viability analysis can identify amphibian populations in danger of extirpation from direct mortality caused by vehicular strikes on roads. This technique may be particularly useful for elucidating population trajectory in non-intuitive situations where large populations undergo heavy losses of individuals on roads over long periods of time. We recommend that researchers undertaking these analyses account the roundtrip nature of (successful) adult migrations to and from breeding sites in situations where roads intervene between upland habitat and breeding sites. Furthermore, we recommend that researchers also incorporate potential vehicular strikes on juvenile life history stages (Petrovan and Schmidt, 2019), especially in light of evidence that mitigation efforts focused on juveniles can have large effects on population-level outcomes (Sterrett et al., 2018). Successful implementation of such relatively complex, including road mortality of multiple life history stages, models for diverse species in a variety of ecological situations will require demographic and natural history studies to obtain robust model parameters.
Summary and conclusion
In summary, based on the results of this study, over 13,700 adult California newts attempted to cross Alma Bridge Road during the 2020/2021 winter season in order to breed in Lexington Reservoir, experiencing a 39.2% roundtrip road-based mortality rate. Our population viability model indicates that this mortality of adults, combined with mortality of juveniles dispersing across the road following metamorphosis, is having a negative impact on the population using Lexington Reservoir to breed and if unmitigated may cause this population to become extirpated in 92 years. This model-derived estimate of time-to-extirpation might be somewhat too short or too long, but is based on the best available data to inform the model parameters. Further investigation of demographic parameters for the Lexington Reservoir California newt population could be used to refine the model in the future. These investigations could include: (1) a mark-recapture component to more accurately determine the frequency of breeding attempts by males and females; (2) a detection probability factor based on the results of a carcass persistence study, as such a factor may influence both the overall road-based mortality rate and the population size in the model, and may direct future survey efforts to include more frequent surveys (e.g., twice a day); (3) an extended drift fence/pitfall trap array survey period throughout the year to capture newts dispersing during the non-breeding season, for example juvenile newts and non-migrating adult newts, to arrive at a more accurate road-based mortality rate for these life stages and seasons; and (4) multiple years of similarly procured capture and mortality data to reflect possible differences in numbers of newts migrating during drought years as opposed to years of average or above average rainfall, which would better estimate the adult breeding population.
It is likely that adult rough-skinned newts experienced similar rates of mortality during their migrations to and from the Lexington Reservoir breeding site. However, only 38 adult rough-skinned newts were observed in the traps compared to 2,054 adult California newts, indicating that the rough-skinned newt population breeding at the reservoir is far smaller than the corresponding California newt population, perhaps because this locality is near the southernmost limit of the rough-skinned newt’s range. High levels of mortality on the road combined with a small population size might make the rough-skinned newt population more susceptible to potential extirpation in the near term than California newt. Although we did not estimate overall population size and mortality rates for rough-skinned newts due to the very low number of captures, such estimates based on a larger survey effort, combined with the same quantitative population modeling approach applied to the California newt population, may provide more information on the risk of extirpation to the rough-skinned newt population and how long it would take such extirpation to occur without intervention. Regardless, our study results indicate that temporary and permanent actions should be implemented to help preserve the local population of California newts, and these actions will also benefit rough-skinned newts.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical review and approval was not required for the animal study because this study involved capture and release of live individuals in the field under a California Department of Fish and Wildlife Scientific Collection Permit No. S-200350006-20041-001.
Author contributions
JW developed the study design, supervised the surveys, analyzed the data, and wrote portions of the manuscript. JR analyzed the data, performed the statistical analysis, and wrote portions of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was partially funded by Regional Peninsula Open Space Trust and Midpeninsula Regional Open Space District.
Acknowledgments
The basis for this study was identified through the considerable efforts of Anne Parsons, surveying for road-killed newts along Alma Bridge Road and entering the data into iNaturalist. We would like to thank Blair Pagano, Dana Page, Jackson Bramham, Jared Bond, Mason Hyland, Michael Rhoades, and Vanessa Clayton of the Santa Clara County Parks Department for assisting with the installation of the drift fence/pitfall trap arrays and coordinating with the volunteers. We would also like to thank Anderson Wang, Ariel Starr, Chuck Wilson, Freya Prissberg, Joan Pinder, Kevin Yokoo, Marika Powers, Marisa Dobkins, Mojgan Mahdizadeh, Pam Hansen, Sadie Hine, Matt Bozzo, Shawn Lockwood, Thomas Wang, K. Perry, and Vanessa Roy, who, as volunteers, partnered with H. T. Harvey & Associates biologists Steve Carpenter, Matt Louder, Christian Knowlton, Will Lawton, Emily Malkauskas, Jane Lien, Craig Fosdick, Zach Hampson, Jazmine Jensen, and Stephen Peterson to conduct the daily surveys during the survey period. We would also like to thank Merav Vonshak of the BioBlitz Club and San Jose State University and her team of community scientists, particularly Robin Agarwal, who performed the surveys for newt mortalities along Alma Bridge Road and coordinated closely with our efforts during the 2020/2021 breeding season to ensure that our data were complementary rather than overlapping, inconsistent, or confounding. We would also like to thank Abra Kaiser for producing the maps. Finally, we would like to thank Neal Sharma of the Peninsula Open Space Trust, Julie Andersen, Brian Malone, Jane Mark, Tina Hugg, Alex Casbara, and Kirk Lenington of the Midpeninsula Open Space District, Harry Freitas and Ananth Prasad of Santa Clara County Roads and Airports, Jared Bond, Jeremy Farr of Santa Clara County Parks, Jae Abel and Shawn Lockwood of Valley Water, Merav Vonshak, Shani Kleinhaus of the Santa Clara Valley Audubon Society, Cheryl Brehme of the U.S. Geological Survey, Tiffany Yap of the Center for Biological Diversity, Fraser Shilling of the University of California at Davis Road Ecology Center, Terris Kasteen of the California Department of Fish and Wildlife, Dashiell Leeds of the Sierra Club, and Steve Rottenborn of H. T. Harvey & Associates for participating in the multiple discussions on the study, analytical methods, and our results and conclusions, and for providing comments on earlier drafts of this paper.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.944848/full#supplementary-material
Footnotes
1. ^https://www.ebparks.org/about-us/whats-new/news/tildens-south-park-drive-closed-annual-newt-migration
2. ^https://www.pressdemocrat.com/article/news/newt-brigade-shuttles-salamanders-to-safety/
3. ^https://www.openspace.org/visit-a-preserve/plants-wildlife/newts
4. ^https://www.inaturalist.org/projects/pacific-newt-roadkill-main-project-lexington-reservoir
5. ^https://parks.sccgov.org/plan-your-visit/activities/fishing
6. ^https://www.inaturalist.org/projects/pacific-newt-roadkill-longevity-study-2020-2021-lexington-reservoir
References
Bain, T. K., Cook, D. G., and Girman, D. J. (2017). Evaluating the effects of abiotic and biotic factors on movement through wildlife crossing tunnels during migration of the California tiger salamander Ambystoma californiense. Herpetol. Conserv. Biol. 12, 192–201.
Beebee, T. C. (2013). Effects of road mortality and mitigation measures on amphibian populations. Conserv. Biol. 27, 657–668. doi: 10.1111/cobi.12063
Brame, A. H. Jr. (1968). The number of egg masses and eggs laid by the California newt, Taricha torosa. J. Herpetol. 2, 169–170. doi: 10.2307/1563126
Brehme, C. S., Hathaway, S. A., and Fisher, R. N. (2018). An objective road risk assessment method for multiple species: ranking 166 reptiles and amphibians in California. Landsc. Ecol. 33, 911–935. doi: 10.1007/s10980-018-0640-1
Forman, R. T. T., Sperling, D., Bissonette, J. A., Clevenger, A. P., Cutshall, C. D., Dale, V. H., et al. (2003). Road ecology: Science and solutions. Washington, D.C: Island Press.
Gamradt, S. C., and Kats, L. B. (1996). Effects of introduced crayfish and mosquitofish on California newts. Conserv. Biol. 10, 1155–1162. doi: 10.1046/j.1523-1739.1996.10041155.x
Gibbs, J. P., and Shriver, W. G. (2005). Can road mortality limit populations of pool-breeding amphibians? Wetl. Ecol. Manag. 13, 281–289. doi: 10.1007/s11273-004-7522-9
Glista, D. J., DeVault, T. L., and DeWoody, J. A. (2007). Vertebrate road mortality predominantly impacts amphibians. Herpetol. Conserv. Biol. 3, 77–87.
Glista, D. J., DeVault, T. L., and DeWoody, J. A. (2009). A review of mitigation measures for reducing wildlife mortality on roadways. Landsc. Urban Plan. 91, 1–7. doi: 10.1016/j.landurbplan.2008.11.001
H. T. Harvey & Associates. (2004). Dublin ranch West California Tiger salamander survey and salvage report, winter 2003/2004. Prepared for Ms. Jennifer Lin c/o Martin Inderbitzen [].
Hobbs, M. T. (2013). Amphibian mortality on roads: A case study in Santa Cruz long-toed salamander habitat. [master' thesis] San Jose, CA: San Jose State University.
Jones, M. T., Milligan, W. R., Kats, L. B., Vandergon, T. L., Honneycutt, R. L., Fisher, R. N., et al. (2017). A discrete stage-structured model of California newt population dynamics during a period of drought. J. Theor. Biol. 414, 245–253. doi: 10.1016/j.jtbi.2016.11.011
Kats, L. B., Bucciarelli, G., Vandergon, T. L., Honeycutt, R. L., Mattiasen, E., Sanders, A., et al. (2013). Effects of natural flooding and manual trapping on the facilitation of invasive crayfish-native amphibian coexistence in a semi-arid perennial stream. J. Arid Environ. 98, 109–112. doi: 10.1016/j.jaridenv.2013.08.003
Kuchta, S. R. (2005). “Taricha torosa,” in Amphibian declines: The conservation status of United States species. ed. M. J. Lannoo (Berkeley, CA: University of California Press).
Moss, B. (2008). The kingdom of the shore: achievement of good ecological potential in reservoirs. Fr. Rev. 1, 29–42. doi: 10.1608/FRJ-1.1.2
Parsons, A. (2021). Mass mortality of Pacific newts at Lexington reservoir bearing witness to the decimation of two populations: Summary of four migration seasons (November 2017 – May 2021). Available online at: https://4f81b87a-f0a5-4d8f-93b9-ca628f6c8f26.usrfiles.com/ugd/4f81b8_fb79203c08d14fa2802d76c42594b416.pdf
Petrovan, S. O., and Schmidt, B. R. (2019). Neglected juveniles; a call for integrating all amphibian life stages in assessments of mitigation success (and how to do it). Biol. Conserv. 236, 252–260. doi: 10.1016/j.biocon.2019.05.023
Ritter, W. E. (1897). Diemyctylus torosus Esch.: the life-history and habits of the Pacific coast newt. Proc. Calif. Acad. Sci. 1, 73–114.
Santos, S. M., Carvalho, F., and Mira, A. (2011). How long do the dead survive on the road? Carcass persistence probability and implications for road-kill monitoring surveys. PLoS One 6:e25383. doi: 10.1371/journal.pone.0025383
Stebbins, R. C., and McGinnis, S. M. (2012). Field guide to amphibians and reptiles of California. Berkeley, CA: University of California Press.
Sterrett, S. C., Katz, R. A., Fields, W. R., and Campbell Grant, E. H. (2018). The contribution of road-based citizen science to the conservation of pond-breeding amphibians. J. Appl. Ecol. 56, 988–995. doi: 10.1111/1365-2664.13330
Thomson, R. C., Wright, A. N., and Shaffer, H. B. (2016). California amphibian and reptile species of special concern. Berkeley, CA: University of California Press.
Trombulak, S. C., and Frissell, C. A. (2000). Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol. 14, 18–30. doi: 10.1046/j.1523-1739.2000.99084.x
Twitty, V. C. (1937). Experiments on the phenomenon of paralysis produced by a toxin occurring in Triturus embryos. J. Exp. Zool. 76, 67–104. doi: 10.1002/jez.1400760105
Twitty, V. C. (1961). “Experiments on homing behavior and speciation in Taricha,” in Vertebrate speciation. ed. W. F. Blair (Austin, TX: University of Texas Press)
Twitty, V. C. (1966). Of scientists and salamanders. W. H. Freeman and Company, Stanford University.
Twitty, V. C., and Johnson, H. H. (1934). Motor inhibition in Amblystoma produced by Triturus transplants. Science 80, 78–79. doi: 10.1126/science.80.2064.78
U.S. Fish and Wildlife Service (2004). Draft revised recovery plan for the Santa Cruz long-toed salamander (Ambystoma macrodactylum croceum). U.S. Fish and Wildlife Service, Portland, Oregon.
U.S. Fish and Wildlife Service (2005). Santa Rosa plain conservation strategy. Available online at: https://www.fws.gov/sites/default/files/documents/1-Santa-Rosa-Plain-Conservation-Strategy-Main%20Body-508.pdf
Keywords: California newt, rough-skinned newt, road-based mortality, drift fence/pitfall trap array, carcass survey, population viability model
Citation: Wilkinson JA and Romansic JM (2022) The effect of road-based mortality on a local population of newts along a narrow two-lane road in California. Front. Ecol. Evol. 10:944848. doi: 10.3389/fevo.2022.944848
Edited by:
Viorel Dan Popescu, Ohio University, United StatesReviewed by:
Rodney L. Honeycutt, Pepperdine University, United StatesGiulia Tessa, Museo civico di Storia Naturale di Morbegno, Italy
Copyright © 2022 Wilkinson and Romansic. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeffery A. Wilkinson, jwilkinson@harveyecology.com