 Yucheol Shin
Yucheol Shin Kyungmin Kim3
Kyungmin Kim3 Jordy Groffen
Jordy Groffen Amaël Borzée
Amaël Borzée
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 12 August 2022
Sec. Conservation and Restoration Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.944318
This article is part of the Research TopicAmphibian and Reptile Road EcologyView all 12 articles
Roadkills are a major threat to the wildlife in human-modified landscapes. Due to their ecology, relatively small size, and slow movement, amphibians and reptiles are particularly prone to roadkill. While standardized roadkill surveys provide valuable data for regional roadkill trends, such surveys are often resource-intensive and have limited geographic coverage. Herein, we analyzed a roadkill dataset of the Korean herpetofauna derived from the citizen science database iNaturalist and compared the overall roadkill trends detected in the iNaturalist data with standardized survey-based literature data. Our results show that the overall citizen science data provide a good picture of roadkill trends for the Korean herpetofauna in terms of recorded species. We detected both similarities and notable differences between the iNaturalist and literature data. The most notable differences between the two datasets were found in the number of recorded species, distribution across habitat types, and distribution across elevational ranges. Even with spatially biased sampling, the iNaturalist data had a considerably broader geographic coverage compared to standardized surveys. In addition, we related the presence of roadkills of amphibians and reptiles to the presence of agricultural lands, forests, and grassland. While the unstandardized nature of the citizen science data can be criticized, we argue that this feature also acts as an advantage for this type of data, as citizen science can better detect roadkills of rare species or seasonal events, such as mass migration of amphibians, and inform population trends and threats. Thus, our results highlight the importance of spatially biased and unstandardized citizen science data for roadkill detection. This study builds on previous studies demonstrating citizen science as a viable method of roadkill surveys.
Anthropogenic alterations of the landscapes result in numerous negative pressures on wildlife (Swaisgood, 2020). Among these stresses are urbanization and the associated increase in road density and traffic volume (Vercayie and Herremans, 2015). Roads tend to promote human activity (Yue et al., 2019) and they have a negative impact on populations from their construction to their daily use (Trombulak and Frissell, 2000). After the harvesting of animals, roads are the largest source of anthropogenic mortality worldwide for vertebrates (Hill et al., 2019). In addition to mortality due to wildlife-vehicle collisions (hereafter roadkill), roads also result in reduced animal movement and dispersal (Forman, 1998; Forman and Alexander, 1998; Holderegger and Di Giulio, 2010), reduced population connectivity altering ecosystem dynamics (Benítez-López et al., 2010), habitat loss (Forman, 2005), and habitat fragmentation (Delgado et al., 2007).
While roadkills present challenges for conservation (Swaisgood, 2020), they can be used as an effective tool to monitor population trends, behavioral ecology, range delineation of native and invasive species, or contaminants and pathogens (reviewed by Schwartz et al., 2020). However, roadkills do not affect all wildlife equally, and some taxa are more prone to being killed (Fahrig and Rytwinski, 2009). Reptiles and amphibians are among the species most threatened by roadkills because they are generally small-bodied and relatively slow-moving (Glista et al., 2007; Andrews et al., 2015). Furthermore, their ecology increases the susceptibility to roadkills, as amphibians migrate long distances to and from breeding sites and take advantage of linear unobstructed structures, while snakes and other reptiles utilize warm road surfaces for thermoregulation and foraging (e.g., Glista et al., 2007; Colino-Rabanal and Lizana, 2012; Mccardle and Fontenot, 2016).
In this study, we focused on the overall roadkill trends for the terrestrial herpetofauna of the Republic of Korea (R Korea hereafter). The R Korea is heavily populated and anthropogenic modifications of the landscapes are widespread (Lee and Miller-Rushing, 2014; Borzée et al., 2019b), including road construction (Andersen and Jang, 2021). Currently, there is a total of 112,977 km of roads in R Korea (KOSIS, 2020), posing a significant conservation concern for the Korean wildlife (Byun et al., 2016; Choi, 2016; Kim et al., 2019, 2021a,b; Andersen and Jang, 2021). While the Korean Expressway Corporation records mammal roadkills on expressways in R Korea (36.863 between 2004 and 2019; Kim et al., 2021b), little attention has been given to the roadkills affecting the herpetofauna. The terrestrial herpetofauna of R Korea is composed of 23 amphibian (14 frog species and 10 salamander species) and 27 reptile species (12 species of snakes, six species of lizards, and 10 species of freshwater turtles). Among the 23 amphibian species, 12 are endemic to the Korean Peninsula and one is invasive (the American Bullfrog, Lithobates catesbeianus). In addition, 11 species are either currently listed as nationally threatened or have been identified as threatened by previous studies (Borzée et al., 2020, 2022; Borzée and Min, 2021), and two are listed as globally threatened (NIBR, 2019; https://www.iucnredlist.org/; accessed October 4, 2021). Among reptiles, none is endemic to the Korean Peninsula, eight are invasive (freshwater turtles; Lee et al., 2016), five are listed as nationally threatened, and one is listed as globally threatened (NIBR, 2019; https://www.iucnredlist.org/; accessed October 4, 2021). On the whole, the terrestrial herpetofauna of R Korea represents a relatively high endemicity (22% of the total species diversity) with many threatened species under intense anthropogenic pressure. Therefore, identifying the various threats to the Korean herpetofauna, including roadkills, is the first important step for an effective conservation planning.
Previous studies on the herpetofauna roadkills in R Korea have used standardized roadkill surveys within relatively small geographic areas (Won et al., 2005; Yang et al., 2006; Song et al., 2009; Min and Han, 2010; Kim et al., 2018b). While such studies provide valuable insights into the regional roadkill trends, they are time-consuming and resource-intensive, consequently restricted to focal taxonomic groups and geographic areas, while the threats to the herpetofauna continue to crop up (Lee and Miller-Rushing, 2014; Macias et al., 2021). However, roadkill surveys do not need to be conducted solely by professional herpetologists (Englefield et al., 2020), and the role of citizen scientists in helping herpetological research is increasingly being recognized (Borzée et al., 2019a; Gardner et al., 2019; Rowley et al., 2019). In this regard, incorporating data from citizen science programs has significant advantages when taxonomically and geographically sampling the herpetofauna roadkills. Furthermore, citizen science initiatives are continuous, not restricted by the resources constraining research, and not geographically bound. The data is therefore generated simultaneously across multiple habitat types by multiple contributors and generally this approach results in much larger datasets (Lintott et al., 2010). Detecting herpetofauna roadkills is strongly affected by spatiotemporal factors (Vercayie and Herremans, 2015), even when surveys are specifically focused on herpetofauna roadkills. Therefore, the ability of citizen science programs to record roadkills without the constraints of resources and geographic extent provides significant advantages over conventional and standardized roadkill surveys. Indeed, previous studies have shown promising benefits of utilizing citizen science programs to detect herpetofauna roadkills (Heigl et al., 2017; Chyn et al., 2019). The utility and characteristics of citizens science-based roadkill data however have not been investigated for the Korean terrestrial herpetofauna.
Therefore, our study has multiple objectives. First, we wanted to determine and highlight the threats caused by roadkills to the Korean terrestrial herpetofauna, in terms of species threatened, geographic extent, landscape characteristics, and elevation. We then aimed to determine the importance of roadkill data points from citizens science platforms, published literature, and national databases to assess the overall herpetofauna roadkill trends in R Korea and the possible use for other studies. We also compared the monthly roadkill trend for each species to their ecology, thus highlighting the ability of citizen science data to detect key ecological characteristics of the Korean herpetofauna. Therefore, our study aims to highlight the utility and benefits of citizen science in detecting the broad trends of roadkills threatening the Korean terrestrial herpetofauna.
In order to rapidly collect previously uploaded roadkill observations, we created a project in iNaturalist titled “Herpetofauna Roadkill in the Republic of Korea” (https://www.inaturalist.org/projects/herpetofauna-roadkill-in-the-republic-of-korea). By generating a project with specific criteria for inclusion, the observations of dead reptiles and amphibians were automatically flagged and included in the project, if fitting the criteria. The observations were included in the project only if they had photographic records, were annotated as dead animals, and were recorded from within R Korea. From the dataset thus generated, we manually removed the observations of dead reptiles and amphibians that were not roadkills, and also removed duplicate observations of the same animal made by two or more observers. For these processes, we visually inspected each photograph uploaded with the observation. We also conducted species-by-species searches for unflagged roadkill observations and we manually included them to the project database by annotating them as dead animals. We used the datapoints that were observed between 2 October 2005 to 13 August 2021. We organized the dataset by species name, year, month of observation, observer, GPS coordinates, data quality grade (either Research Grade or Needs ID, a data validation process embedded in the platform, https://www.inaturalist.org/pages/help), and Class- and Order-level classifications (Supplementary Data 1). The project we created does not have any standardized survey protocols, and therefore the data generated from the project can be regarded as opportunistically collected.
In this study, we used the list of Korean herpetofauna provided by the Korean Society of Herpetologists (http://www.krsh.co.kr/html/sub0201.html) as a reference taxonomic framework. To this list we added the three recently described Hynobius species (Borzée and Min, 2021) and Bufo sachalinensis (reassigned from B. gargarizans for population on the Korean Peninsula; Othman et al., 2022), but we did not include the newly described Onychodactylus species (Borzée et al., 2022). However, the taxonomy used herein differs slightly from that used by iNaturalist. Most notably, iNaturalist recognizes a population of Gloydius ussuriensis on Jeju Island as a different species “Gloydius tsushimaensis”. Although G. tsushimaensis is a valid taxon originally described from the Tsushima Island in Japan (Isogawa et al., 1994), there is currently no published study to support the presence of G. tsushimaensis on Jeju Island. Thus, we consider occurrences of “G. tsushimaensis” from Jeju as G. ussuriensis in this study, following previous studies (Song, 2007; Lee et al., 2012; Jang et al., 2016; Shin and Borzée, 2020). Similarly, while iNaturalist recognizes the presence of both Elaphe schrenckii and Elaphe anomala in the Korean Peninsula, we only recognize the former name based on previous studies using genetic and morphological data (An et al., 2010; Lee, 2011). Thus, we included the occurrences of E. anomala from R Korea to that of E. schrenckii.
To generate an independent dataset to compare against the iNaturalist data, we utilized the roadkill records of Korean herpetofauna reported in the literature. We searched for the relevant literature by using the Environment Digital Library Search service provided by the Korean Ministry of Environment (https://library.me.go.kr/#/), Google Scholar (https://scholar.google.com/), Research Information Sharing Service (http://www.riss.kr/index.do), and the library search engine of the Kangwon National University (https://library.kangwon.ac.kr/#/). We used the following combinations of keywords (both in Korean and English): roadkill Korea, reptile roadkill Korea, amphibian roadkill Korea, herpetofauna roadkill, and wildlife roadkill. We accessed the search engines between 24 August 2021 and 5 October 2021. The search resulted in five peer-reviewed research articles, four governmental research reports, one conference proceeding, and one dissertation containing information on herpetofauna roadkills. From each publication, we extracted the frequency of roadkill detection per reported species. We then pooled the number of records for each species to generate a literature-based dataset. We compared the species-level diversity of terrestrial herpetofauna roadkills in this dataset with that of the iNaturalist dataset. The list of literature used to generate the literature dataset is shown in Supplementary Table 1. We did not include a study describing mass movement events and associated roadkills in Onychodactylus koreanus (Shin et al., 2020), as the observations described in that study were uploaded to iNaturalist prior to its publication. Thus, the observations described therein were included as a part of the iNaturalist data.
As the iNaturalist observations were not normally distributed, we primarily used chi-square tests to investigate monthly trends in the number of recorded roadkills. We focused on monthly roadkill trends as many reptile and amphibian species show highly seasonal movement patterns (Shepard et al., 2008; Garriga et al., 2017), and since previous studies suggest strong monthly patterns associated with the recorded number of herpetofauna roadkills in R Korea (Song et al., 2009; Park et al., 2017). We analyzed the monthly data at different levels. First, we conducted chi-square tests for each species of amphibians and reptiles and plotted monthly roadkill trends for each species. We then superimposed the known key life-history aspects (e.g., hibernation and breeding seasons) over the monthly roadkill trends to investigate the ability of citizen science-based roadkill trends in capturing ecological signals associated with species life histories. For information on life histories, we primarily consulted the works of Lee et al. (2012) for reptiles, and Lee and Park (2016) for amphibians. For two species of snakes (Sibynophis chinensis and Gloydius intermedius) with limited life histories documented in Lee et al. (2012), we supplemented relevant information derived from recently published studies (Do et al., 2017; Koo et al., 2018; Banjade et al., 2020).
Next, as some species had small sample sizes (n ≤ 5), we used ecological groupings to further analyze monthly roadkill patterns. For amphibians, this grouping was based on breeding ecology (spring and summer breeders), as seasonal migration to and from breeding sites is a major aspect of amphibian ecology, and it is also associated with high road mortality (Hels and Buchwald, 2001). Using breeding ecology to group species is further justified because the hibernation period (another major factor of seasonal migration) is generally similar among Korean amphibians, spanning from October to March (Lee and Park, 2016). We excluded species with highly specialized (e.g., Bufo stejnegeri; Lee and Park, 2016) or poorly known (e.g., O. koreanus; Lee and Park, 2016; Shin et al., 2020) breeding ecologies that could not be assigned to either of the two groups. Thus, we conducted separate chi-square tests on spring and summer breeders to investigate the seasonal patterns of roadkill occurrences. The group assignment based on the breeding ecology for each amphibian species is based on the study by Lee and Park (2016) and is provided in Supplementary Table 2. Regarding reptiles, we further analyzed the monthly trends at the Order level (Squamata and Testudines) and by taxonomic groups (snake, lizards, and turtles), using these groupings as a general ecological proxy. We then analyzed the monthly trends at the Class level for both amphibians and reptiles (Amphibia and Reptilia).
In addition, we plotted the monthly roadkill observations against the total monthly observations of reptiles and amphibians. We conducted this procedure separately for amphibians and reptiles to visualize whether the temporal bias of the total herpetofauna observations was also reflected in the roadkill observations. We matched the time period of total observations to that of the roadkill observations to ensure comparability of data. Finally, we assessed the overall yearly sampling efforts of the iNaturalist dataset through years 2005 to 2021, expressed as the number of observations divided by the number of participants (sensu Chyn et al., 2019). We performed these biostatistical analyses in R version 4.1.0 (R Core Team, 2021), implemented in the RStudio environment.
To visualize the spatial sampling intensity of iNaturalist roadkill observations, we generated kernel density maps separately for reptiles and amphibians, using the MASS package (Venables and Ripley, 2002) in R version 4.1.0 (R Core Team, 2021). We also visualized the spatial sampling intensity of roadkill records based on the literature data. However, we were unable to employ kernel density analysis on the literature dataset as the exact GPS coordinates of roadkill occurrence points were not provided. Thus, we extracted information on the number of provinces surveyed in each publication, thereby generating provincial “counts”. We then summed up these counts for each province to obtain the “total provincial count,” which we used as a literature-based representation of spatial sampling bias broadly comparable to that of the iNaturalist data (Supplementary Data 2).
Next, we investigated the elevational distribution of iNaturalist roadkill records by using the 30-arc-s resolution (~1 km) elevation raster downloaded from WorldClim 2.1 (https://www.worldclim.org/data/worldclim21.html; Fick and Hijmans, 2017) as the template. Using the raster package (Hijmans, 2019) in R, we extracted elevation values from the elevation raster for each occurrence point of both amphibian and reptile roadkill records. We then categorized the obtained elevation values into range classes divided by 100 m interval, resulting in 10 classes for amphibians (spanning 0–1,241 m asl) and nine classes for reptiles (spanning 0–810 m asl). We conducted chi-square tests on the obtained elevation ranges, separately for the amphibian and reptile datasets, to test for differences in the elevational distribution of roadkill records.
We also investigated the distribution of roadkill records across different habitat types. To do so, we first obtained the 30-m resolution broad-scale land cover raster of R Korea provided by the Ministry of Environment (https://egis.me.go.kr/main.do). This land cover map is classified into seven discrete habitat categories such as, residential, agriculture, forest, grassland, wetland, barren ground, and water bodies (http://egis.me.go.kr/map/map.do?type=land). We extracted habitat types from the land cover map for each occurrence point of amphibian and reptile roadkills using the raster package (Hijmans and Etten, 2012) in R, and we categorized the habitat types according to the classification code of the land cover map. We then conducted separate chi-square tests for amphibian and reptile datasets to test for differences in the distribution of roadkill records across different habitat types.
For comparative purposes, we extracted information on elevational and habitat distribution of roadkill records reported in the literature (see section Literature data for the methods of literature search). To generate literature-based elevation and habitat type datasets, we first excluded four studies from the initial set of 11 studies because they either did not provide elevational and/or habitat information or the provided information was too coarse to make comparisons with the iNaturalist data (Table 1). Thus, the literature-based elevation dataset was generated from two studies for amphibians, and from three studies for reptiles (Table 1 and Supplementary Data 3), and the literature-based habitat dataset for amphibians was generated from four studies, and that of reptiles was generated from five studies (Table 1 and Supplementary Data 4). We categorized elevation data extracted from the literature in the same way as the iNaturalist data, with different upper elevation limits (due to differences in data). However, we could not apply the same habitat type categories to the literature-based data, mainly because the method of categorization used in the literature was different from our GIS-based approach. Our approach can only identify one habitat type falling in the same 30-m raster grid as our roadkill occurrence data, describing the general habitat settings of each roadkill record, whereas two habitat types immediately adjacent to the roadkill observation (with road expressed as an intersection of two habitat types) are usually recorded in the literature (Song et al., 2009; Park et al., 2017). Therefore, we categorized the habitat type of the literature-based data into the following nine classes, utilizing the classification scheme most frequently used in the literature: agriculture-agriculture, agriculture-residential, agriculture-water, forest-agriculture, forest-forest, forest-water, grassland-forest, residential-residential, and water-residential. Next, we conducted chi-square tests separately on literature-based amphibian and reptile datasets to test for differences in the distribution of roadkill records across elevational ranges and habitat types.
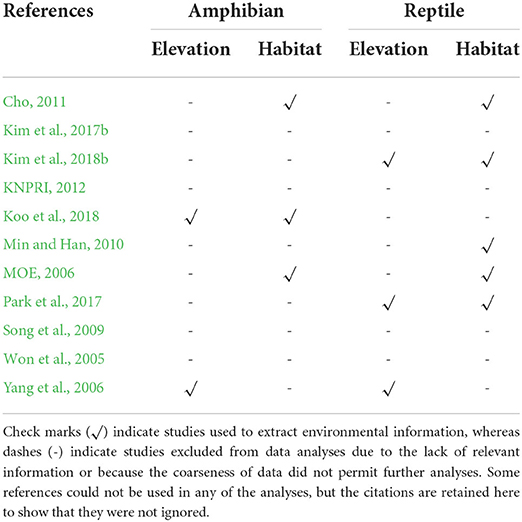
Table 1. The list of published literature used to generate literature-based datasets.
For further comparisons on the spatial aspects of the roadkill occurrences, we utilized another independent dataset generated by the National Institute of Ecology (NIE) in R Korea. The NIE dataset is based on the Korea Roadkill Observation System (KROS) launched in 2018 by the Ministry of Environment and the Ministry of Land, Infrastructure, and Transport (Kim et al., 2019). The roadkill data were recorded between March 2019 and September 2021. The resulting dataset is comprised of GPS coordinates and species-level identification for each roadkill occurrence point. After removing the entries with missing species-level identification, we used the same methods applied to the iNaturalist dataset to categorize elevational ranges and habitat types of the NIE data, separately for amphibians and reptiles. Next, we conducted chi-square tests separately for amphibian and reptile datasets to test for differences in the distribution of NIE roadkill records across habitat types and elevational ranges. As the NIE dataset had a considerably shorter time span compared to both the iNaturalist and literature datasets, we restricted the use of this dataset to the broad-scale comparisons of landscape and elevational aspects only.
In addition, we measured the Euclidean distance between each roadkill point and seven types of land cover: residential area, agricultural land, forest, grassland, wetland, barren ground, and water bodies. This method highlights spatial patterns of roadkill and makes comparisons between land cover types more convenient (Kim et al., 2019). This analysis was conducted in QGIS 3.4.5 with the land cover map provided by the Ministry of Environment (http://egis.me.go.kr/). Prior to the statistical analyses, we excluded datapoints that were more than 5,000 m away from the focal feature. As a result, we analyzed 361 data points for amphibians and 273 points for reptiles. Then, we analyzed the data between and within classes to determine the difference in roadkill occurrence in relation to the landscape type. As the data were not normally distributed (Kolmogorov–Smirnov test, K = 0.21, df = 3,490, p < 0.001), we used a Bayesian inference for a linear regression to test for the difference between class, with distance as dependent variables and landscape class and species as independent variables. This analysis was conducted in SPSS v.28.0 (SPSS Inc., Chicago, USA). The steps of data analyses are briefly summarized in Figure 1.
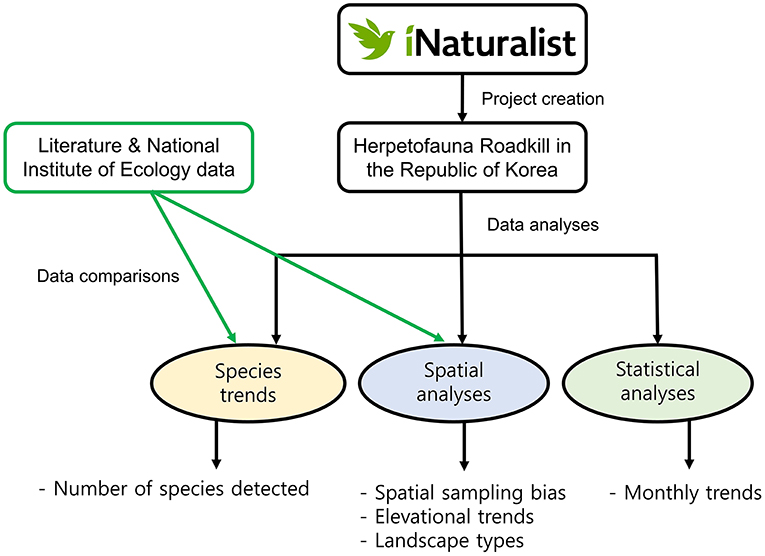
Figure 1. Flow chart of data analyses on the roadkill dataset collected through iNaturalist. The project can be retrieved at: https://www.inaturalist.org/projects/herpetofauna-roadkill-in-the-republic-of-korea.
A total of 638 observations were compiled into the project, spanning the years from 2005 to 2021 (Figure 2 and Table 2). Of the 638 recorded observations, 588 records were of research grade quality and 50 records needed further identification to be listed as Research Grade. Records in the latter category generally consisted of highly damaged Rana and snake roadkills, making confident identifications at the species level difficult. A total of 634 observations were retained after removing redundant observations and non-roadkill observations of deceased reptiles and amphibians (reptile n = 273; amphibian n = 361). Regarding reptiles, the iNaturalist project detected roadkills in all 11 species of terrestrial snakes native to R Korea, in 3 out of 6 lizard species (Takydromus amurensis, Takydromus wolteri, and Scincella vandenburghi), and in all native freshwater turtle species (Mauremys reevesii and Pelodiscus maackii). For amphibians, the project detected roadkills in all anuran species (including the invasive L. catesbeianus) with the exception of a recently described hylid Dryophytes flaviventris (Borzée et al., 2020; but refer to Section Discussion for a personal observation of roadkills in the species), and detected roadkills in two out of 10 salamander species (O. koreanus and Hynobius leechii). In total, the iNaturalist project observations detected roadkills in 76.2% of the terrestrial herpetofaunal diversity of R Korea (32 out of 42 described species). The observations also detected roadkills in several nationally endangered and near threatened species. These species include the nationally endangered Chinese Many-toothed Snake (Sibynophis chinensis; EN; n = 3), the Suweon Treefrog (Dryophytes suweonensis; EN; n = 2), the Russian Ratsnake (Elaphe schrenckii; VU; n = 10), the Gold-spotted Pond Frog (Pelophylax chosenicus; VU; n = 2), the Boreal Digging Frog (Kaloula borealis; VU; n = 6), the Reeves' Turtle (M. reevesii; VU; n = 1), and the Amur Softshell Turtle (P. maackii; VU; n = 1) (NIBR, 2019). Moreover, the project detected a roadkill of the Slender Racer (Orientocoluber spinalis; NT; n = 1), a species that is rarely and infrequently observed in R Korea (NIBR, 2019; Macias et al., 2021).
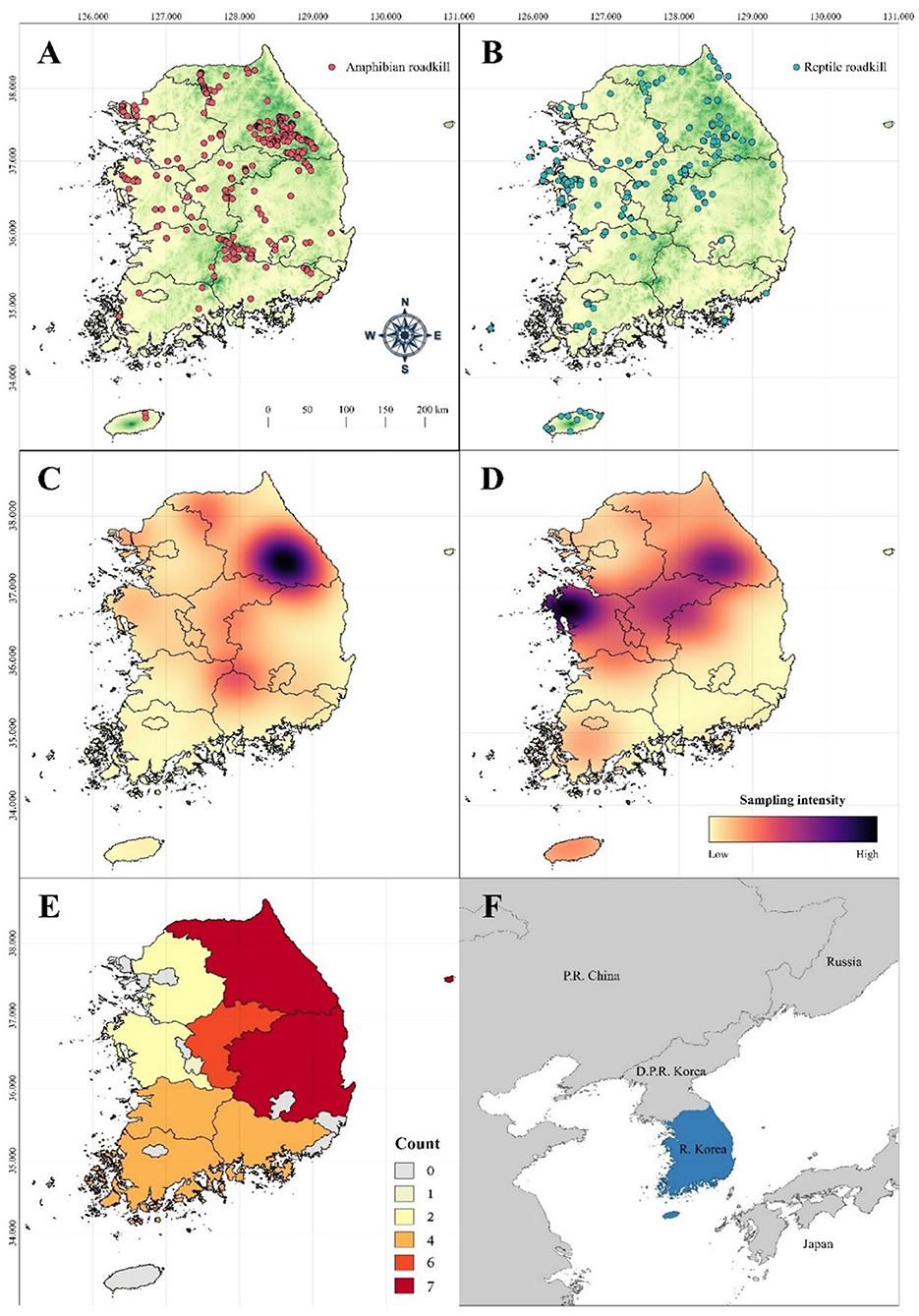
Figure 2. Spatial distributions of herpetofauna roadkill observations across the Republic of Korea (R Korea). (A) GPS coordinates of the iNaturalist amphibian roadkill observations. (B) GPS coordinates of the iNaturalist reptile roadkill observations. (C) A kernel density map of the iNaturalist amphibian roadkill observations showing the spatial sampling intensity across R Korea. (D) A kernel density map of the iNaturalist reptile roadkill observations showing the spatial sampling intensity across R Korea. The spatial bias of roadkill observations is apparent from the cluster of records in the northern and western portions and the relative paucity of observations in the southeastern region of R Korea. (E) Spatial bias of the regional representation of herpetofauna roadkills based on the available literature. The unit of spatial bias (count) is a simple count of the general region surveyed (refer to Section Materials and methods for details). (F) The location of the study area (R Korea) in northeast Asia highlighted in blue. The map was generated in QGIS 3 with the elevation raster downloaded from the WorldClim database (Fick and Hijmans, 2017) as a base map for (A,B). Darker coloration in the base map (green shade) indicates higher elevation, whereas lighter coloration (yellow shade) indicates lower elevation.
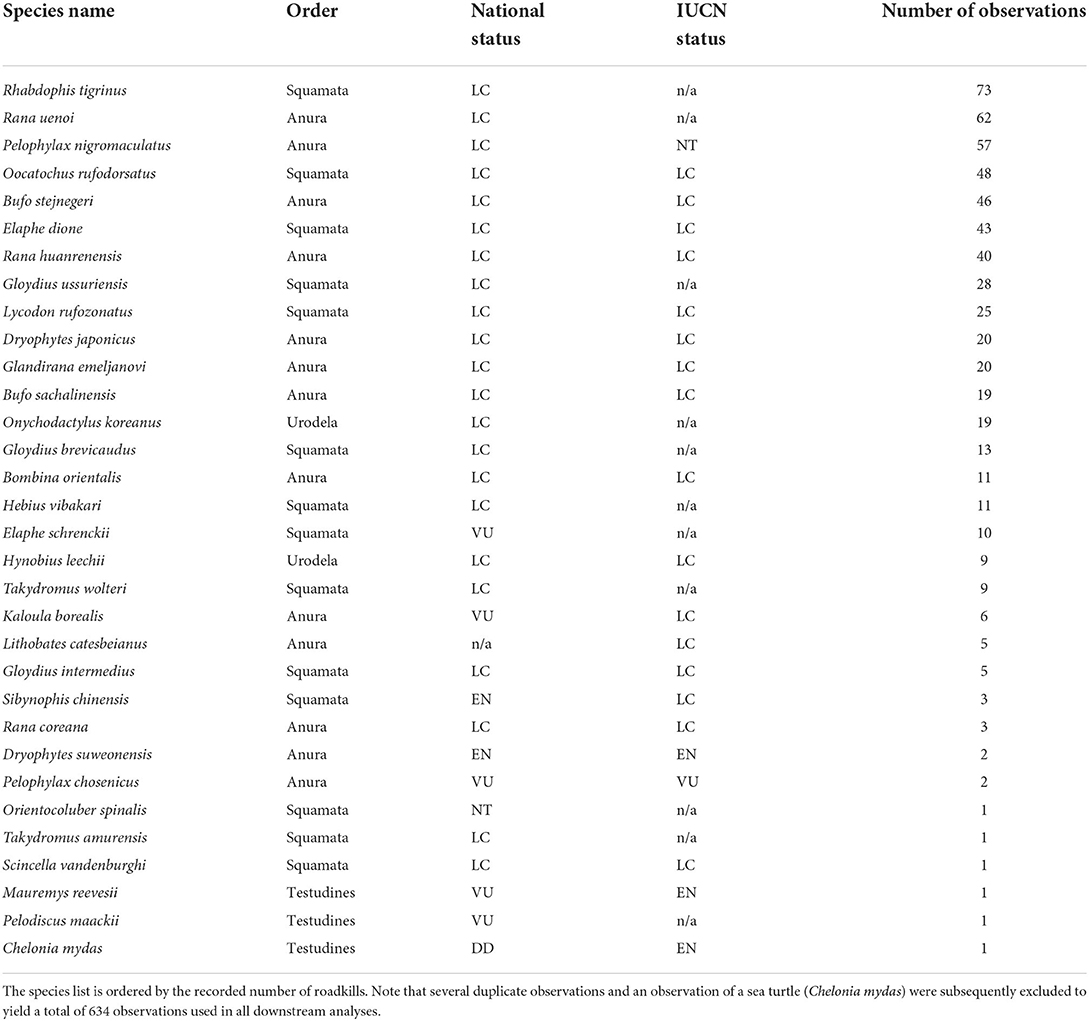
Table 2. The list of Korean herpetofauna detected in the raw iNaturalist roadkill dataset (n = 638) at the point of data analyses, shown alongside Order-level classification, national threat status based on NIBR (2019), and the global threat status based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed 4 October 2021), if applicable.
The literature data encompassed 10,618 observations spanning years from 2003 to 2017 (Supplementary Table 1). Regarding reptiles, the literature had recorded roadkills in 10 out of 11 species of snakes (all species except S. chinensis), one lizard species (T. amurensis), one native freshwater turtle species (P. maackii), and one invasive freshwater turtle species (Trachemys scripta elegans). For amphibians, the literature data had recorded roadkills in 10 out of 14 species of anurans (all species except D. suweonensis, D. flaviventris, K. borealis, and P. chosenicus), and two out of 10 species of salamanders (O. koreanus and H. leechii). In total, the literature data detected 59.5% of the terrestrial herpetofaunal diversity of R Korea (25 out of 42 species). The only two nationally threatened species detected in the literature data were the Russian Ratsnake (E. schrenckii; VU; n = 5) and the Amur Softshell Turtle (P. maackii; VU; n = 1). The literature data also detected roadkills of the Slender Racer (O. spinalis; NT; NIBR, 2019), with higher frequency (n = 16) than the iNaturalist data (n = 1). There were no roadkill records of globally threatened species reported in the literature.
Considering the taxonomic representation in both citizen science and literature data, anuran amphibians were detected at the highest frequency (iNaturalist: n = 333; literature: n = 7,828), followed by snakes (iNaturalist: n = 260; literature: n = 2,750), salamanders (iNaturalist: n = 28; literature: n = 33), lizards (iNaturalist: n = 11; literature: n = 5), and turtles (iNaturalist: n = 2; literature: n = 2; Figure 3). We converted the raw frequency to percentage to apply the same unit of comparison between the two datasets (iNaturalist and literature). Overall, the two datasets were comparable in terms of the percentage of species detected (Figure 4).
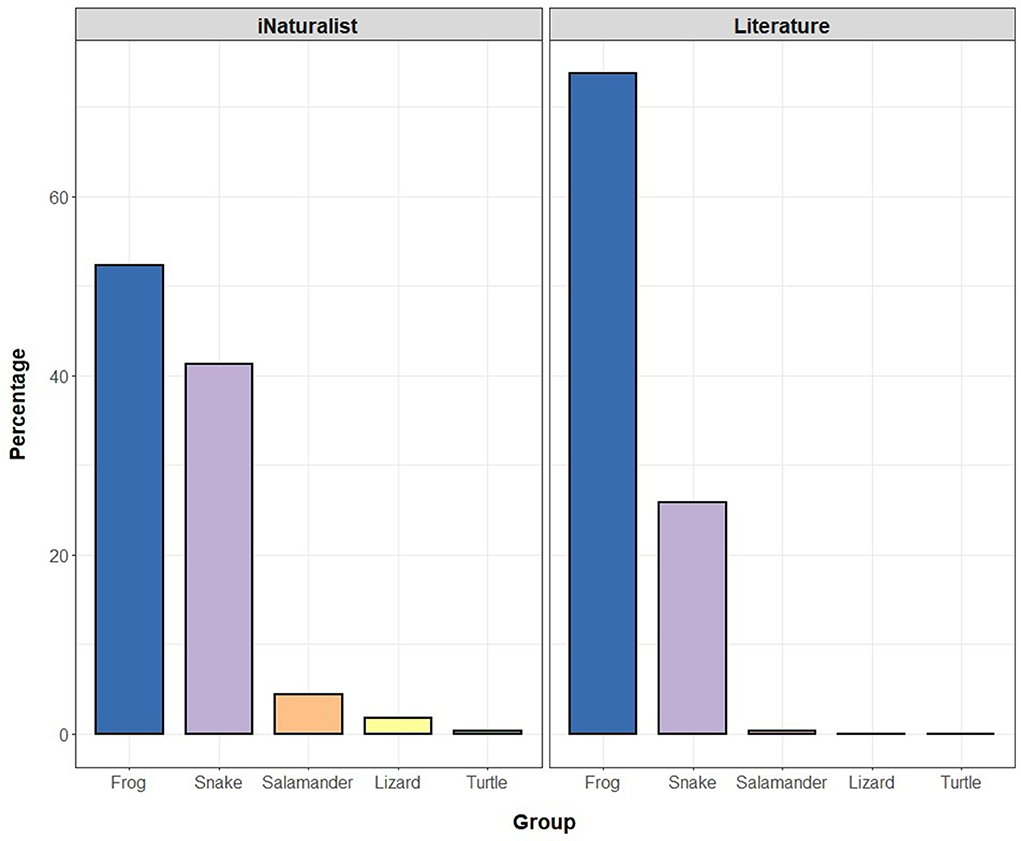
Figure 3. Taxonomic sampling of roadkill records according to the citizen science (iNaturalist) and literature data. The overall taxonomic representation, expressed as percentages was similar between the datasets, despite the much larger total sample size of the literature data. The anuran amphibians showed the highest roadkill frequency (iNaturalist: n = 333; literature: n = 7,828), followed by snakes (iNaturalist: n = 260; literature: n = 2,750), salamanders (iNaturalist: n = 28; literature: n = 33), lizards (iNaturalist: n = 11; literature: n = 5), and turtles (iNaturalist: n = 2; literature: n = 2).
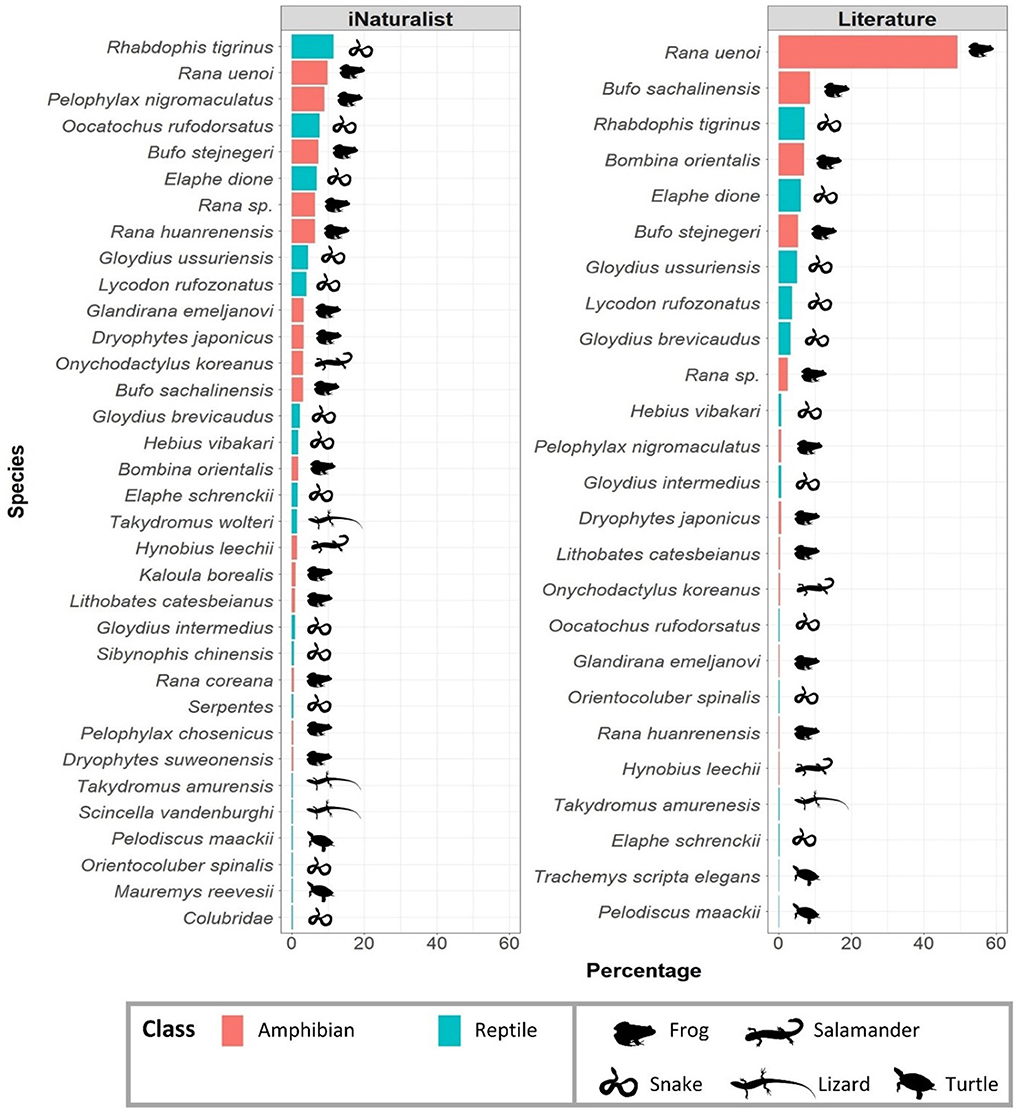
Figure 4. Comparison of the taxonomic composition of roadkill observations between the iNaturalist and literature-based data. Due to the different sample sizes between data types, we converted the raw count of frequency to percentages to apply the same scale of comparison. The most heavily affected species were generally congruent between datasets, with several differences associated with taxonomic diversity detected in each dataset as well as the sampled percentage of each species. Note that the iNaturalist data detected roadkills in more species (n = 32; 76.2% of total herpetofaunal diversity of R Korea) compared to the literature-based data (n = 25; 59.5% of total herpetofaunal diversity of R Korea). The silhouette figures were obtained from PhyloPic (http://phylopic.org/) under the Public Domain Dedication 1.0 license or the Public Domain Mark 1.0 license (for the turtle silhouette). Frog silhouette (Leptodactylus melanonotus) by Jose Carlos Arenas-Monroy; Salamander silhouette (Onychodactylus fischeri) by zoosnow; Snake silhouette (Vipera berus) by Beth Reinke; Lizard silhouette (Eumeces fasciatus) by David Orr; Turtle silhouette (Chrysemys picta) by an uncredited author.
In general, the expected levels of activity associated with key ecological aspects were reflected in the monthly roadkill trends of each reptile and amphibian species (Figure 5, 6). Focusing on the analyses based on the ecological groupings of amphibians, the roadkill trends were significantly different between months for both spring breeding (n = 173; χ2 = 231.08; df = 10; p < 0.001) and summer breeding (n = 123; χ2 = 116.94; df = 10; p < 0.001) amphibians. For both groups, the highest peak of roadkill records was in September and October. Regarding the Order-level groupings for reptiles, only the squamate grouping had the sufficient sample size at the Order level for statistical analysis, and the number of monthly roadkill observations differed significantly between seasons for this grouping (n = 271; χ2 = 266.3; df = 10; p < 0.001), with the peak of roadkill observations in October. The data analyzed at the Class level (Amphibia and Reptilia) also suggested a significant difference in the number of monthly roadkill observations for amphibians (n = 361; χ2 = 446.53; df = 10; p < 0.001) and reptiles (n = 273; χ2 = 265.27; df = 10; p < 0.001), with peaks of roadkill records in October and September, respectively.

Figure 5. The monthly roadkill trends for Korean amphibians (n = 361), based on iNaturalist observations. The roadkill records were collected from 1 January 2005 to 13 August 2021 through the iNaturalist project “Herpetofauna Roadkill in the Republic of Korea”. Ambiguous observations with identifications above the genus level were excluded. The information on breeding season and hibernation period for each species is based on Lee and Park (2016), and summarized in Supplementary Table 2. Photographs of amphibians by AB.

Figure 6. The monthly roadkill trends for Korean reptiles based on iNaturalist observations (n = 273). The roadkill records were collected from 1 January 2005 to 13 August 2021 through the iNaturalist project “Herpetofauna Roadkill in the Republic of Korea”. We excluded observations with ambiguous identifications above the species level. The information on breeding and hibernation periods are primarily derived from Lee et al. (2012). The breeding season of Sibynophis chinensis was inferred from Koo et al. (2018) and Banjade et al. (2020), whereas information on the breeding season of Gloydius intermedius was further supplemented from Do et al. (2017). Photographs of reptiles courtesy of Hyun-Tae Kim, modified and used under permission.
The two peaks of roadkill frequency generally matched with the two peaks of total observations for reptiles. However, for amphibians, the two peaks of total observations were decoupled with a peak of roadkill occurrences in October (Figure 7). The yearly sampling effort of the iNaturalist data, as expressed by the yearly number of observations divided by the yearly number of participants, fluctuated through the years 2005 to 2021. Nevertheless, the overall sampling intensity showed a steadily increasing trend (Figure 8).
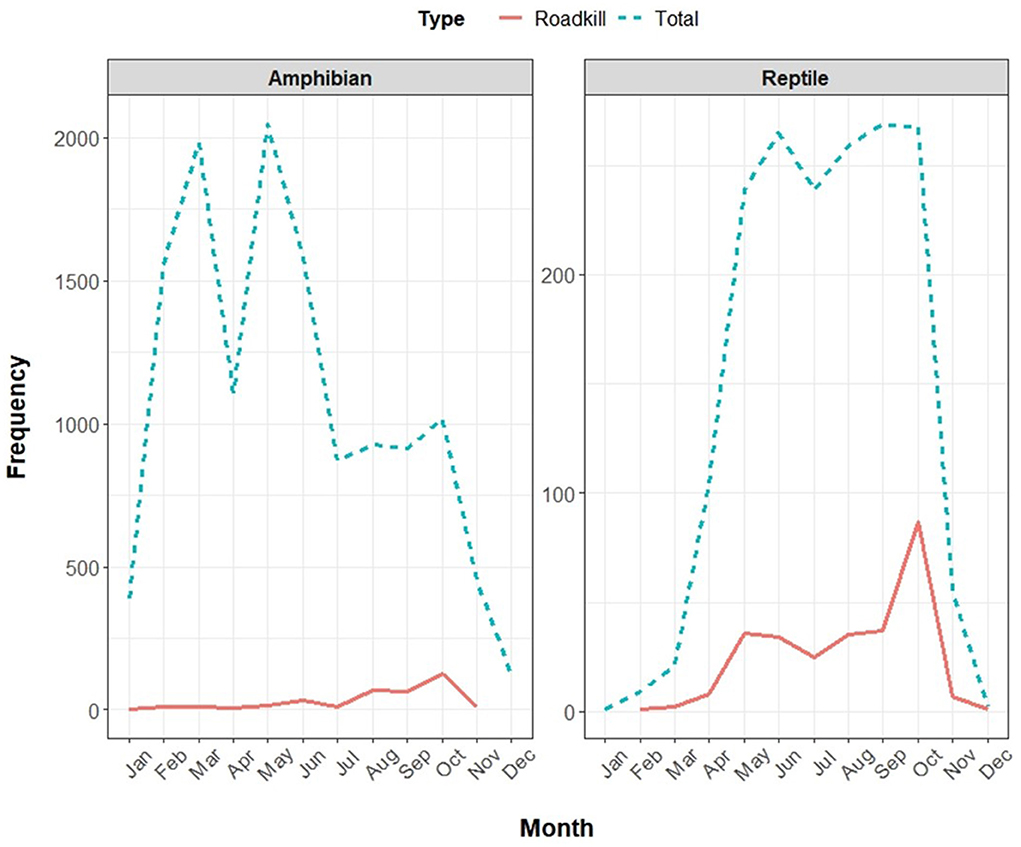
Figure 7. The total monthly roadkill records for the Korean herpetofauna (total n = 634; amphibian n = 361; reptile n = 273) plotted against the total monthly observations (total n = 14,699; amphibian n = 12,969; reptile n = 1,730), based on the pooled iNaturalist dataset collected from 1 January 2005 to 13 August 2020. Distinctly different monthly trends are notable between the total observations and roadkill observations for both amphibian and reptile datasets, highlighting that the peaks in the number of roadkills do not match the peaks in the number of total observations.
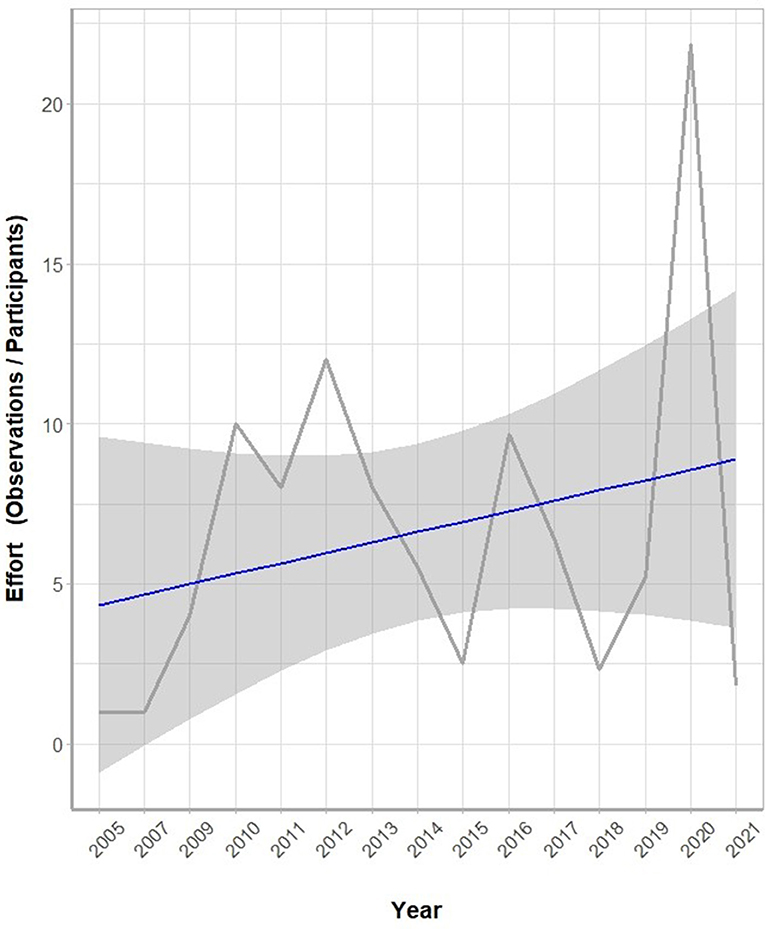
Figure 8. The yearly sampling efforts of the iNaturalist roadkill observations through the years 2005 to 2021 (from 1 January 2005 to 13 August 2020). Here, the yearly sampling effort is expressed as the number of observations per year divided by the number of participants per year. Overall, the sampling efforts show an increasing trend through the years. The trend line (blue) is shown with 95% confidence interval.
The kernel density maps generated from amphibian and reptile roadkill observation points revealed considerable spatial bias of the iNaturalist dataset (Figures 2C,D). Regarding the amphibian roadkills from the iNaturalist database, most of R Korea had an intermediate sampling intensity. The regions with low sampling intensity were generally located in the southwestern provinces and offshore islands. Meanwhile, the area of the highest sampling intensity was located in the east-central region of R Korea. Regarding the reptile roadkills, the regions with low sampling intensity based on the iNaturalist data were in the southeastern provinces. The regions with high sampling intensity were located along the central R Korea, with the west-central region showing the highest sampling intensity. Other regions fell within the range of intermediate sampling intensity. The spatial sampling bias of herpetofauna roadkills based on the literature dataset was distinctly different from that based on the iNaturalist dataset. Based on the literature data, the survey intensity (measured by provincial “counts”) were the highest in the eastern and central provinces (6 ~ 7 counts), followed by the southern provinces (~ 4 counts), and the western provinces (~ 2 counts). Some provinces were either not surveyed, or there were no published literature that surveyed those provinces.
In the iNaturalist data, the frequency of roadkill records was significantly different across elevational ranges for both amphibians (n = 292; χ2 = 91.105; df = 9; p < 0.001) and reptiles (n = 272; χ2 = 550.11; df = 8; p < 0.001). The highest number of amphibian roadkills was recorded between 300 and 399 m asl. (n = 69), followed by 0–99 m asl (n = 54), 200–299 m asl (n = 45), 400–499 m asl (n = 44), 500–599 m asl (n = 43), 100–199 m asl (n = 39), 700–799 m asl (n = 24), 600–699 m asl (n = 23), 900–1,241 m asl (n = 12), and 800–899 m asl (n = 8). Meanwhile, the highest number of reptile roadkills was recorded between 0 and 99 m asl (n = 147), followed by 100–199 m asl (n = 39), 300–399 m asl (n = 26), 200–299 m asl (n = 25), 400–499 m asl (n = 18), 600–699 m asl (n = 7), 500–599 m asl (n = 5), 700–799 m asl (n = 4), and 800–810 m asl (n = 1). Therefore, according to the iNaturalist data, the number of herpetofauna roadkill observations generally showed a decreasing trend with increasing elevation (Figure 9).
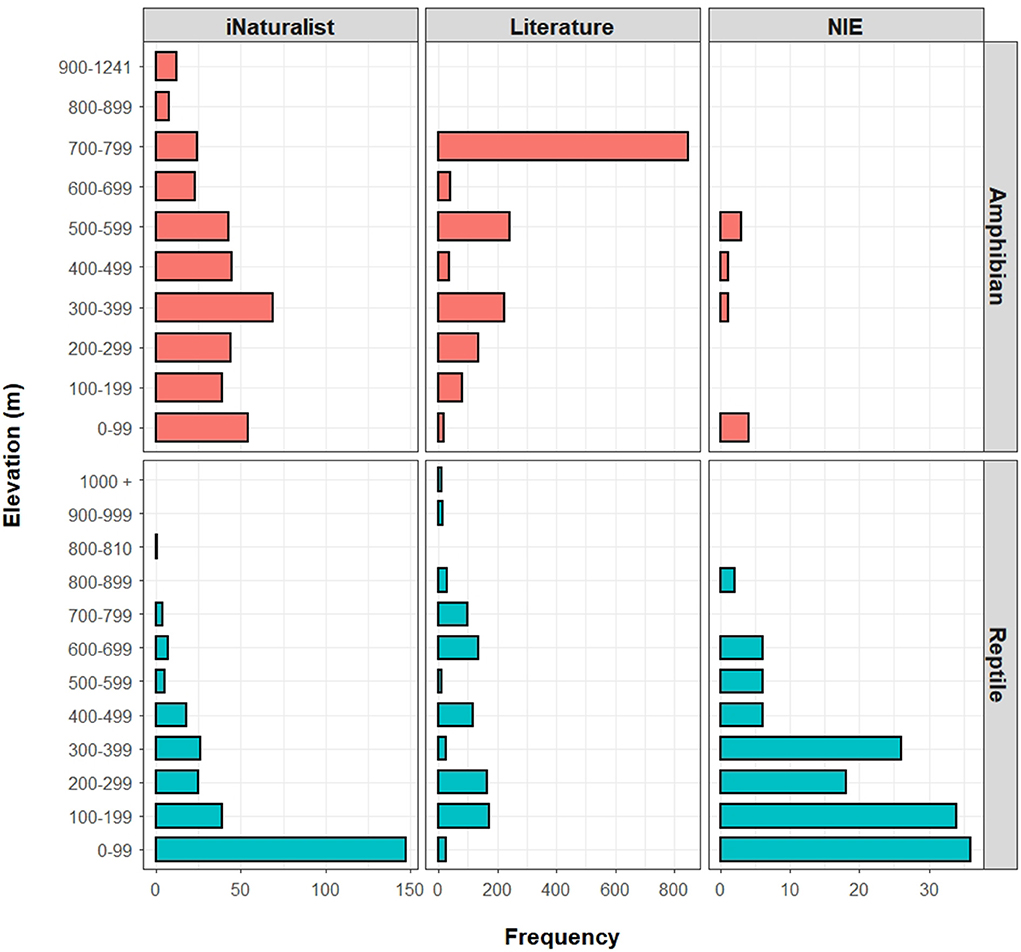
Figure 9. Elevational distributions of roadkill records compared among the iNaturalist, literature, and governmental (National Institute of Ecology; NIE) data. Regarding the iNaturalist data, the amphibian roadkill frequency was the highest around low to mid elevations, whereas the reptile roadkill frequency was the highest in low elevations, with clear decrease toward higher elevations. The literature-based data shows a different trend, with amphibian roadkill frequency roughly increasing toward higher elevations, and with reptile roadkill frequency showing fluctuating patterns. The sample size of NIE data is insufficient to detect elevational patterns for amphibians, but there is an overall decreasing trend with increasing elevation for reptiles.
Regarding the literature data, the distribution of roadkill records was also significantly different across elevational ranges for both amphibians (n = 1,617; χ2 = 2,592; df = 7; p < 0.001) and reptiles (n = 802; χ2 = 586.85; df = 10; p < 0.001; Figure 9). The highest number of amphibian roadkills was recorded from 700 to 799 m asl (n = 846), followed by 500–599 m asl (n = 241), 300–399 m asl (n = 222), 200–299 m asl (n = 134), 100–199 m asl (n = 81), 600–699 m asl (n = 40), 400–499 m asl (n = 35), and 0–99 m asl (n = 18). For reptiles, the highest number of roadkills was recorded from 100 to 199 m asl (n = 173), followed by 200 −299 m asl (n = 166), 600–699 m asl (n = 134), 400–499 m asl (n = 117), 700–799 m asl (n = 98), 800–899 m asl (n = 30), 0–99 m asl (n = 25), 300–399 m asl (n = 24), 900 −999 m asl (n = 15), 500–599 m asl (n = 10), and 1,000 m + asl (n = 10). Therefore, according to the literature data, the amphibian roadkill frequency generally showed an increasing trend with increasing elevation, while the reptile roadkill frequency showed a decreasing trend with increasing elevation (Figure 9).
The sample size of NIE data was insufficient to test for differences in the elevational distribution of amphibian roadkills (n = 10), although most roadkills were recorded in elevational ranges of 0–99 m asl (n = 4) and 500–599 m asl (n = 3). On the other hand, the distribution of NIE reptile roadkill records was significantly different across elevational ranges (n = 134; χ2 = 78.776; df = 7; p < 0.001; Figure 9). The highest number of reptile roadkills was recorded in 0–99 m asl (n = 36), followed by 100–199 m asl (n = 34), 300–399 m asl (n = 26), 200–299 m asl (n = 18), 400–499 m asl (n = 6), 500–599 m asl (n = 6), 600–699 m asl (n = 6), and 800–899 m asl (n = 2).
The distribution of iNaturalist roadkill records was significantly different across habitat types, for both amphibians (n = 325; χ2 = 540.79; df = 5; p < 0.001; Figure 10) and reptiles (n = 260; χ2 = 287.29; df = 6; p < 0.001; Figure 10). The highest number of amphibian roadkills was recorded in forest environments (n = 198), followed by agriculture (n = 84), grassland (n = 20), residential area (n = 14), barren ground (n = 5), and water bodies (n = 4). Meanwhile, the highest number of reptile roadkills was recorded in agricultural landscapes (n = 106), followed by forest (n = 90), wetland (n = 21), residential area (n = 15), barren ground (n = 14), grassland (n = 11), and water bodies (n = 3).
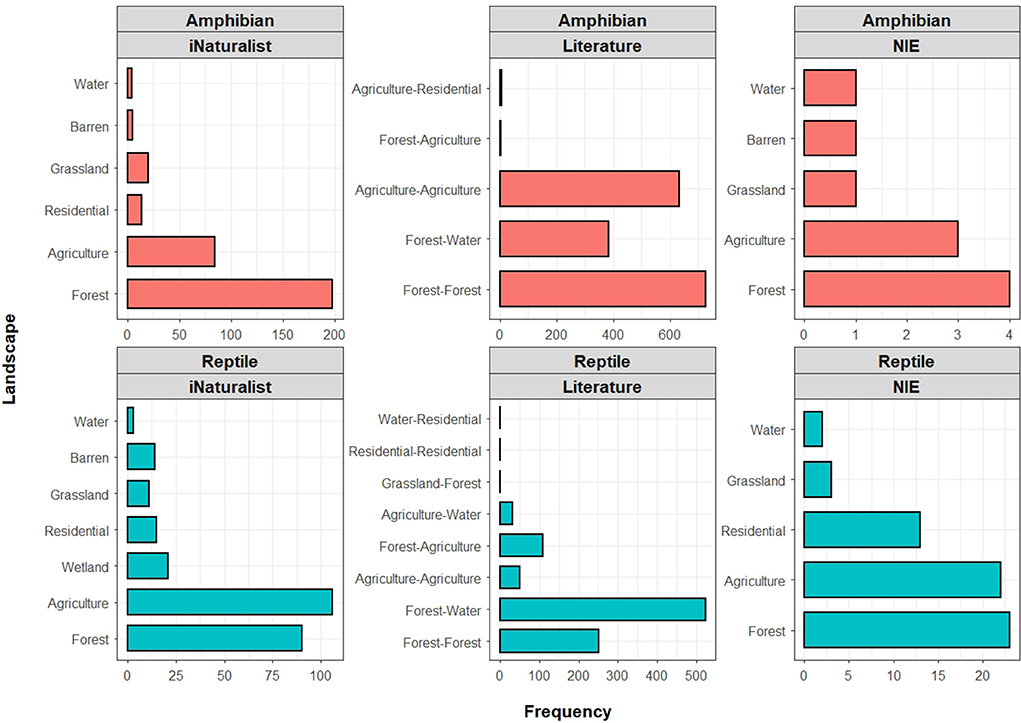
Figure 10. Distribution of roadkill records across different habitat types compared among the iNaturalist, literature, and governmental (National Institute of Ecology; NIE) data. While the specific classification of habitat types differ between datasets, all three datasets generally converge on a congruent pattern by detecting the highest frequency of herpetofauna roadkills in either forest or agricultural landscapes.
The distribution of literature-based roadkill records was also significantly different across habitat types, for both amphibians (n = 1,750; χ2 = 1311.7; df = 4; p < 0.001) and reptiles (n = 966; χ2 = 1941.8; df = 7; p < 0.001; Figure 10). For amphibians, the highest number of roadkills was recorded from the intersection of forest environments (forest-forest; n = 724), followed by the intersection of agricultural landscapes (agriculture-agriculture; n = 633), intersection of forests and water bodies (forest-water; n = 383), intersection of agricultural landscapes and residential areas (agriculture-residential; n = 6), and the intersection of forests and agricultural landscapes (forest-agriculture; n = 4). Regarding reptiles, the highest number of roadkills was recorded from the intersection of forests and water bodies (forest-water; n = 522), followed by the intersection of forest environments (forest-forest; n = 252), the intersection of forests and agricultural landscapes (forest-agriculture; n = 108), the intersection of agricultural landscapes (agriculture-agriculture; n = 50), the intersection of agricultural landscapes and water bodies (agriculture-water; n = 31), the intersection of grasslands and forests (grassland-forest; n = 1), the intersection of residential areas (residential-residential; n = 1), and the intersection of water bodies and residential areas (water-residential; n = 1; Figure 10).
The sample size of NIE data was insufficient to test for the differences in habitat distribution of amphibian roadkills (n = 10), although most roadkills were recorded in forests (n = 4) and agricultural landscapes (n = 3). Meanwhile, the distribution of NIE reptile roadkill records was significantly different in habitat types (n = 61; χ2 = 31.841; df = 4; p < 0.001). The highest number of reptile roadkills was recorded in forests (n = 23), followed by agricultural landscapes (n = 22), residential areas (n = 13), grasslands (n = 3), and near water bodies (n = 2; Figure 10).
We measured and compared the distance between all roadkill points and the seven land cover types. The Bayesian inference for the Linear Regression Model showed that the distances between roadkill observations and the seven land cover types were different between reptiles and amphibians (Bayes Factor = 0.00, R = 0.17; R2 = 0.03, SE = 237.27). The average distance between roadkill observations and wetlands was nearly twice higher in amphibians than reptiles (2,023 m for amphibians and 1,021 m for reptiles). In addition, the three habitat types the closest to the occurrences of roadkills were agricultural lands (48.82 ± 104.06 m; mean ± SD), wetlands (86.02 ± 142.47 m), and forests (216.86 ± 210.41 m) for amphibians, and agricultural lands (29.49 ± 94.78 m), forests (91.23 ± 148.04 m), and grasslands (200.51 ± 231.70 m) for reptiles.
Our results demonstrate that the citizen science data can detect roadkills in the majority of the Korean terrestrial herpetofauna with broad geographic coverage and can capture patterns related to the ecology of individual species given sufficient sample size. Therefore, citizen science-based roadkill surveys have significant benefits that can complement traditional survey methods. Here we show that the iNaturalist roadkill dataset is capturing the key points of the behavioral ecology of the Korean herpetofauna in terms of monthly observation trends, habitat use, and especially for species with large sample sizes (Figure 5, 6 and Supplementary Figures 1, 2). In general, the peak(s) of roadkill for each species of reptiles and amphibians coincided with the periods immediately before the breeding season, during the breeding seasons, and periods immediately before the hibernation period. Increased movements can be expected in many reptile and amphibian species during these periods, as they often migrate long distances in search of breeding habitats or suitable hibernacula. For example, the frequency of total monthly observations captured two life history aspects for the three Rana species (plus a non-identifiable group of “Rana sp.” Supplementary Figure 1). The peaks in monthly roadkill observations between February and March also correspond to the breeding season (Figure 5), as this is the period when adults emerge from hibernation and migrate over the landscape to reach their breeding habitats (Song et al., 2009; Park et al., 2017). In addition, the lack of roadkill observations after the breeding seasons followed by a second peak of roadkill observations in October (mostly composed of R. uenoi and R. huanrenensis), correspond to the adults migrating to their hibernacula across roads. The iNaturalist data also captured species-specific roadkill trends in amphibian species with highly specialized ecologies. For instance, the frequency of roadkills in B. stejnegeri, as well as total monthly observations, was the highest immediately before hibernation, rather than during the breeding season as in most other amphibian species (Figure 5 and Supplementary Figure 1). This is likely because B. stejnegeri hibernates underwater as amplectant pairs and breed as they emerge from hibernation without long-distance migration to breeding habitats. Therefore, the roadkill patterns of B. stejnegeri are generally consistent with the patterns for other amphibian species while also reflecting aspects of the ecology specific to the species. However, outside the breeding period, B. stejnegeri migrates back to summer habitats in the mountains. Therefore, the lack of a second peak of recorded roadkills in the summer (as expected from the ecology of species) likely reflects the low sampling efforts in this period (refer to the paragraph below).
Both species-specific behavioral ecology and the sampling mode can explain why some species were not detected in the iNaturalist dataset. For some species, there are clear biases stemming from the lack of sampling. For instance, H. leechii was the only Hynobius species detected by both the iNaturalist and literature datasets, even though there are seven species of Hynobius described in R Korea (Borzée and Min, 2021). H. leechii is the most common and widely distributed species of the genus in R Korea and it migrates across roads in large numbers during the breeding season (A Borzée, pers. obs.), which is partly reflected in the peak of roadkill records in the iNaturalist data. However, as only a few roadkills of this species were detected, the true magnitude of roadkill affecting this species is likely to be greatly underrepresented in the iNaturalist data. On the other hand, all seven species of Hynobius in R Korea form a clade of lentic breeders with similar ecologies (Borzée and Min, 2021). However, the other species of the genus less common than H. leechii have a relatively small distribution range and are found in more rural areas, which in combination with their secretive lifestyles and habitat characteristics might reduce the potential risk of roadkill and explain the lack of records. This low representation of the species is further supported by the spatial distribution of sampling intensity across R Korea (southern R Korea; Figure 2C). The same pattern is also found in other rare species with restricted ranges, such as the Critically Endangered treefrog D. flaviventris (Pers. Obs.; Borzée et al., 2020).
Habitat characteristics of roadkills were different for amphibians and reptiles, although agricultural lands and forests were among the top three habitat types where roadkills were recorded for both classes. This similarity at the macro scale despite the difference is likely related to the broadly similar habitat use of many reptile and amphibian species. Habitat characteristics could also explain the lack of roadkill detection for the three lizard species (Eremias argus, Gekko japonicus, and Scincella huanrenensis). For instance, G. japonicus has a restricted distribution in the southern coastal regions (Kim et al., 2017a) and is usually observed on walls, fences, and roofs in city parks and residential areas (Lee et al., 2012; Kim et al., 2018a). Similarly, E. argus have small home ranges (Kim et al., 2012) and they are restricted to coastal sand dunes, inland grasslands along lakes, and mountain slopes with low vegetation cover (Lee et al., 2012). Finally, the distribution of the third species, S. huanrenensis, is poorly known although potentially distributed across a broader range than currently known (Shin et al., 2021), and the species is usually found in the forest interiors (Lee et al., 2012). The behavioral characteristics and habitat use of the three lizard species is likely the cause for the reduced risk of roadkill.
Some of the discrepancies in the spatial distributions of herpetofauna roadkills between the iNaturalist and literature datasets originate from apparent sampling biases. For example, the elevational distributions of amphibian roadkills based on the iNaturalist and literature data show a roughly inversed pattern (Figure 9), noting that the elevational distribution from the iNaturalist dataset better represents the actual elevational distribution of the species (Andersen et al., 2022). Although fully disentangling the conflicting patterns between datasets can be difficult due to the characteristics of different data types, differences in sample sizes, and associated sampling artifacts, the reason for the conflicting elevational distribution of amphibian roadkills can be partly attributed to an apparent sampling bias in the literature dataset, but also partly explained by the general scarcity of literature on the herpetofauna roadkill in R Korea. Therein, the high frequency of amphibian roadkills at high elevations mostly originated from a single study (Yang et al., 2006), which recorded an exceptionally large number of roadkills (n = 568) for a single species (B. sachalinensis) at 700 m + asl, an elevation at which they are not common (Andersen et al., 2022). In addition, due to the large sample size collected through the citizen science project, and the demonstrated match between large citizen science datasets collected on roads and real species distribution for toads in the United Kingdom (Petrovan et al., 2020), we expect the iNaturalist project to be representative of the distribution of the species when the sample size is large enough, and we expect the literature data to be skewed toward the high-value sites selected for the studies.
Importantly, there are also similarities between the iNaturalist and literature datasets. For example, the five snake species with the highest roadkill frequency are generally congruent between the datasets, with Rhabdophis tigrinus scoring the highest in both datasets. The same general pattern can be found among anuran species, with Rana uenoi having the highest frequency in both data types. However, there were also some notable differences. First, the five snake and anuran species with the highest roadkill frequencies, although generally similar, were not identical between the datasets (Figure 4). For example, Pelophylax nigromaculatus has the second highest roadkill frequency recorded for anurans after R. uenoi and Oocatochus rufodorsatus has the second highest roadkill frequency recorded for snakes after R. tigrinus in the iNaturalist dataset. However, both P. nigromaculatus and O. rufodorsatus have low recorded roadkill frequencies in the literature. This discrepancy can be traced to an apparent under-sampling of agricultural landscapes in standardized surveys (Figure 10). It is also notable that the iNaturalist data detected roadkills in seven more species than the literature data. Moreover, the iNaturalist dataset recorded more roadkills of threatened species than the literature data. For example, S. chinensis, K. borealis, and P. chosenicus were only detected by the iNaturalist data, and E. schrenckii was detected more frequently by the iNaturalist data (n = 10) than the literature data (n = 5).
It is important to note that the frequency of roadkills is linked with detection rate, which is highly dependent on weather conditions (Kosmala et al., 2016), as bad weather impacts the activity of both citizen science and standardized surveys, and good weather impacts the preservation of roadkill before decomposition or scavenging, resulting in variations in body size, color, state of preservation, and potential for identification (Chyn et al., 2019). In addition, while roadkills of juveniles are less likely to be preserved long enough to be detected compared to adults, the roadkills of the E. schrenckii were composed mostly of juveniles hatched that same year. The high ratio of juveniles is likely linked to the seasonal dispersal by the species, highlighting that a large enough sample size can compensate the weaknesses of citizen science data. Here, we analyzed terrestrial roadkills, including that of species with aquatic life-stages. However, it is more difficult for citizen science to detect deaths related to human activities in the aquatic habitat, and thus creating a bias in our data. Nevertheless, citizen science can detect deaths of aquatic herpetofauna caused by human activities as the iNaturalist dataset also captured an observation of a dead Green sea turtle (Chelonia mydas), likely resulting from a collision with a boat (Denkinger et al., 2013).
Even with its unstandardized nature, spatial bias, and observer bias inherent to the iNaturalist data, or any citizen science dataset, this type of data has considerable advantages and values when investigating the broad-scale trends of roadkill. For example, one distinct advantage of using public platforms such as iNaturalist in roadkill surveys is the ability to make multiple validations on species-level identifications, and the ability to re-evaluate identifications based on photographic data accompanying each uploaded observation. This can increase the confidence of species-level identification, and thereby improve the overall quality of data. On the other hand, such validations are rarely possible for standardized surveys, as only the results of the surveys are reported without reference to photographic data. For instance, any reader is able to confirm or reject the identification of the observations included in this research (https://www.inaturalist.org/projects/herpetofauna-roadkill-in-the-republic-of-korea), which is not possible with other published research. In addition, another significant advantage of citizen science-based roadkill surveys is the ability to rapidly accumulate roadkill data from across broad geographic regions and habitats with minimal resource inputs.
We also highlight the unique and different advantages of citizen science-based roadkill surveys and typical standardized roadkill surveys that can be mutually beneficial. For example, some detailed aspects of roadkill (e.g., the age demography of roadkill) might be difficult to study with citizen science data if these data are not standardized to some degree. This issue of data collection can be overcome by citizen science initiatives by adopting the standardized or semi-standardized survey protocols from typical, standardized roadkill surveys (similar to Gardner et al., 2019). The standardized surveys conducted by professional researchers can also benefit from including the unstandardized citizen science data. The standardized roadkill surveys previously conducted and reported from R Korea were usually conducted at fixed temporal intervals in set routes (Yang et al., 2006; Song et al., 2009; Min and Han, 2010). While these surveys and related studies have provided important insights into the broad patterns of herpetofauna roadkills in R Korea, they were generally limited to certain stretches of roads penetrating key habitats (e.g., National Parks; Song et al., 2009; Cho, 2011; Koo et al., 2018). Therefore, citizen science data can help researchers to make informed decisions on survey locations and timing. For example, the spatial density, habitat, and elevational representations of roadkill sampling in citizen science platforms can be used by researchers to target specific habitat types and/or under-sampled regions to broaden the survey extent. Furthermore, researchers can utilize the citizen science data to specifically target otherwise opportunistic elements of roadkill detections, such as mass migration of amphibians associated with movements to and from breeding sites and hibernacula which are often affected by short-term or even hourly environmental conditions (Shin et al., 2020). Therefore, the advantages of citizen science and standardized survey data can complement each other to better characterize the true magnitude of roadkill affecting the herpetofauna of R Korea.
In conclusion, our study demonstrated citizen science as a viable method to survey roadkill trends of herpetofauna, adding to a growing number of studies highlighting the benefits of citizen science in biodiversity and conservation research. However, the estimates of roadkill trends derived from the citizen science, as well as targeted road surveys, still underestimate the true magnitude, and subsequent impacts, of roadkill to the Korean herpetofauna. Therefore, we encourage the users of citizen science platforms to upload more roadkill observations and encourage researchers to utilize such data to get a fuller picture of the impacts of roadkill affecting the Korean herpetofauna.
The original contributions presented in the study are included in the article/Supplementary material. Further inquiries can be directed to the corresponding author/s.
The animal study was reviewed and approved by Ethic Committee of Nanjing Forestry University.
YS, KK, JG, and AB: conceptualization and visualization. YS and KK: formal analysis. AB: funding acquisition. YS, KK, and JG: writing the original draft. YS, KK, JG, DW, ES, and AB: writing—review and editing. All authors contributed to the article and approved the submitted version.
This work was supported by the Foreign Youth Talent Program (QN2021014013L) from the Ministry of Science and Technology of the People's Republic of China to AB.
We are grateful for the efforts of the citizen scientists who entered herpetofauna roadkill observations to iNaturalist, and especially @pintail for their numerous observations. We also thank the contributors of the PhyloPic silhouettes database. Finally, we thank the handling editor Dr. Silviu O. Petrovan and two reviewers for providing constructive comments that significantly improved this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.944318/full#supplementary-material
An, J., Park, D., Lee, J., Kim, K.-S., Lee, H., and Min, M.-S. (2010). No genetic differentiation of Elaphe schrenckii subspecies in Korea based on 9 microsatellite loci. Korean J. Syst. Zool. 26, 15–19. doi: 10.5635/KJSZ.2010.26.1.015
Andersen, D., Chuang, M.-F., Choi, M., Kim, A., Kwon, S., Jang, Y., et al. (2022). Altitudinal distribution of Korean amphibians. Zool. Stud. 61:25.
Andersen, D., and Jang, Y. (2021). Biodiversity and transportation infrastructure in the Republic of Korea: a review on impacts and mitigation in developing the country. Diversity 13, 519. doi: 10.3390/d13110519
Andrews, K. M., Langen, T. A., and Struijk, R. (2015). “Reptiles: overlooked but often at risk from roads,” in Handbook of Road Ecology, eds R. V. D. Ree, D. J. Smith, and C. Grilo (Chichester: Wiley Blackwell), 271–280. doi: 10.1002/9781118568170.ch32
Banjade, M., Jeong, Y.-H., Han, S.-H., Kim, Y.-K., Kim, B. S., and Oh, H.-S. (2020). First report on the reproduction of captive Chinese many-toothed snake (Sibynophis chienensis) in Jeju Island, South Korea. J. Ecol. Environ. 44, 6. doi: 10.1186/s41610-020-0150-3
Benítez-López, A., Alkemade, R., and Verweij, P. A. (2010). The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol. Conserv. 143, 1307–1316. doi: 10.1016/j.biocon.2010.02.009
Borzée, A., Baek, H. J., Lee, C. H., Kim, D. Y., Song, J.-Y., Suh, J.-H., et al. (2019a). Scientific publication of georeferenced molecular data as an adequate guide to delimit the range of Korean Hynobius salamanders through citizen science. Acta Herpetol. 14, 27–33. 10.13128/Acta_Herpetol-24102
Borzée, A., Messenger, K. R., Chae, S., Andersen, D., Groffen, J., Kim, Y. I., et al. (2020). Yellow sea mediated segregation between North East Asian Dryophytes species. PLoS ONE 15, e0234299. doi: 10.1371/journal.pone.0234299
Borzée, A., and Min, M.-S. (2021). Disentangling the impact of speciation, sympatry and island effect on the morphology of seven Hynobius sp. salamanders. Animals 11, 187. doi: 10.3390/ani11010187
Borzée, A., Shin, Y., Poyarkov, N. A., Jeon, J. Y., Baek, H. J., Lee, C. H., et al. (2022). Dwindling in the mountains: description of a critically endangered and microendemic Onychodactylus species (Amphibia, Hynobiidae) from the Korean Peninsula. Zool. Res. 43, 750–755. doi: 10.24272/j.issn.2095-8137.2022.048shu
Borzée, A., Struecker, M.-Y., Yi, Y., Kim, D., and Kim, H. (2019b). Time for Korean wildlife conservation. Science 363, 1161–1162. doi: 10.1126/science.aaw9023
Byun, Y. S., Kwon, J. N., Kim, J. H., Shin, M. H., and Lee, S. D. (2016). How do landscape and road barriers affect road crossing of multihabitat mammals. J. Korean Assoc. Geograph. Inform. Stud. 19, 89–101. doi: 10.11108/kagis.2016.19.3.089
Cho, D. (2011). The Study on the Road-Kill Occurred on the Old National Highway, Route 1 within Gyeryongsan National Park. Woosuk University.
Chyn, K., Lin, T. E., Chen, Y. K., Chen, C. Y., and Fitzgerald, L. A. (2019). The magnitude of roadkill in Taiwan: patterns and consequences revealed by citizen science. Biol. Conserv. 237, 317–326. doi: 10.1016/j.biocon.2019.07.014
Colino-Rabanal, V., and Lizana, M. (2012). Herpetofauna and roads: a review. Basic Appl. Herpetol. 26, 5–31. doi: 10.11160/bah.12008
Delgado, J. D., Arroyo, N. L., Arevalo, J. R., and Fernandez-Palacios, J. M. (2007). Edge effects of roads on temperature, light, canopy cover, and canopy height in laurel and pine forests (Tenerife, Canary Islands). Landscape Urban Plan. 81, 328–340. doi: 10.1016/j.landurbplan.2007.01.005
Denkinger, J., Parra, M., Muñoz, J., Carrasco, C., Murillo, J., Espinosa, E., et al. (2013). Are boat strikes a threat to sea turtles in the Galapagos Marine Reserve? Ocean Coast. Manage. 80, 29–35. doi: 10.1016/j.ocecoaman.2013.03.005
Do, M. S., Nam, K. B., and Yoo, J. C. (2017). First observation on courtship behavior of short-tailed viper snake, Gloydius saxatilis (Squamata: Viperidae) in Korea. J. Asia Pac. Biodivers. 10, 583–586. doi: 10.1016/j.japb.2017.08.003
Englefield, B., Starling, M., Wilson, B., Roder, C., and Mcgreevy, P. (2020). The Australian roadkill reporting project-applying integrated professional research and citizen science to monitor and mitigate roadkill in Australia. Animals 10, 1112. doi: 10.3390/ani10071112
Fahrig, L., and Rytwinski, T. (2009). Effects of roads on animal abundance: an empirical review and synthesis. Ecol. Soc. 14, 21. doi: 10.5751/ES-02815-140121
Fick, S. E., and Hijmans, R. J. (2017). WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Forman, R. (1998). Road ecology: a solution for the giant embracing us. Landscape Ecol. 13, 3–5. doi: 10.1023/A:1008036602639
Forman, R.T. (2005). “Good and bad places for roads: effects of varying road and natural pattern on habitat loss, degradation, and fragmentation,” in ICOET 2005 Proceedings (Davis, CA: Road Ecology Center).
Forman, R. T., and Alexander, L. E. (1998). Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 29, 207–231. doi: 10.1146/annurev.ecolsys.29.1.207
Gardner, E., Julian, A., Monk, C., and Baker, J. (2019). Make the adder count: population trends from a citizen science survey of UK adders. Herpetol. J. 29, 57–70. doi: 10.33256/hj29.1.5770
Garriga, N., Franch, M., Santos, X., Montori, A., and Llorente, G. (2017). Seasonal variation in vertebrate traffic casualties and its implications for mitigation measures. Landscape Urban Plan. 157, 36–44. doi: 10.1016/j.landurbplan.2016.05.029
Glista, D. J., Devault, T. L., and Dewoody, J. A. (2007). Vertebrate road mortality predominantly impacts amphibians. Herpetol. Conserv. Biol. 3, 77–87.
Heigl, F., Horvath, K., Laaha, G., and Zaller, J. G. (2017). Amphibian and reptile road-kills on tertiary roads in relation to landscape structure: using a citizen science approach with open-access land cover data. BMC Ecol. 17, 24. doi: 10.1186/s12898-017-0134-z
Hels, T., and Buchwald, E. (2001). The effect of road kills on amphibian populations. Biol. Conserv. 99, 331–340. doi: 10.1016/S0006-3207(00)00215-9
Hijmans, R. J. (2019). Raster: Geographic Data Analysis and Modeling. R package version 3.0-7. Available online at: https://CRAN.R-project.org/package=raster
Hijmans, R. J., and Etten, J. V. (2012). Raster: Geographic Analysis and Modeling with Raster Data. R package version 2.0-12. Available online at: http://CRAN.R-project.org/package=raster
Hill, J., Devault, T., and Belant, J. (2019). Cause-specific mortality of the world's terrestrial vertebrates. Global Ecol. Biogeogr. 28, 680–689. doi: 10.1111/geb.12881
Holderegger, R., and Di Giulio, M. (2010). The genetic effects of roads: a review of empirical evidence. Basic Appl. Ecol. 11, 522–531. doi: 10.1016/j.baae.2010.06.006
Isogawa, K., Moriya, A., and Mitsui, S. (1994). A new snake of the genus Agkistrodon (Serpentes: Viperidae) from Tsushima Island, Nagasaki Prefecture, Japan. Jpn. J. Herpetol. 15, 101–111. doi: 10.5358/hsj1972.15.3_101
Jang, H.-J., Kim, D.-I., and Chang, M.-H. (2016). Distribution of reptiles in South Korea - based on the 3rd National Ecosystem Survey. Korean J. Herpetol. 7, 30–35.
Kim, D.-I., Choi, W.-J., Park, I.-K., Kim, J.-S., Kim, I.-H., and Park, D. (2018a). Comparisons of microhabitat use of Schlegel's Japanese gecko (Gekko japonicus) among three populations and four land cover types. J. Ecol. Environ. 42, 24. doi: 10.1186/s41610-018-0088-x
Kim, H., Kim, J., Lee, J., Song, J.-Y., Hwang, T., and Kim, E. (2017b). “Wildlife roadkill occurrences in Korean National Parks,” in Proceedings of the Korea Environmental Sciences Society Conference (Gyeongju).
Kim, H.-T., Bae, Y. H., Kim, H., Kim, D., and Borzée, A. (2017a). Gekko japonicus (Schlegel's Japanese Gecko). Herpetol. Rev. 48, 82.
Kim, I.-H., Ra, N.-Y., and Park, D. S. (2012). Habitat use, home range, and hibernaculum of the Mongolian racerunner, Eremias argus (Lacertidae, Reptilia) in a coastal sand dune in South Korea. Asian Herpetol. Res. 3, 133–140. doi: 10.3724/SP.J.1245.2012.00133
Kim, K., Serret, H., Clauzel, C., Andersen, D., and Jang, Y. (2019). Spatio-temporal characteristics and predictions of the endangered leopard cat Prionailirus bengalensis euptilura road-kills in the Republic of Korea. Global Ecol. Conserv. 19, e00673. doi: 10.1016/j.gecco.2019.e00673
Kim, K., Yi, Y., Woo, D., Park, T., and Song, E. (2021a). Factors influencing roadkill hotspot in the Republic of Korea. Proc. Natl. Inst. Ecol. 2, 274–278. doi: 10.22920/PNIE.2021.2.4.274
Kim, M., Park, H., and Lee, S. (2021b). Analysis of roadkill on the Korean Expressways from 2004 to 2019. Int. J. Environ. Res. Public Health 18, 10252. doi: 10.3390/ijerph181910252
Kim, S.-B., Lee, J.-H., and Park, D. (2018b). Patterns of snake roadkills on the roads in the national parks of South Korea. Korean J. Ecol. Environ. 51, 234–244. doi: 10.11614/KSL.2018.51.3.234
KNPRI (2012). Wildlife Roadkill Occurrences and the Usage of Wildlife Corridors on the Roads Penetrating National Parks: Comprehensive Results of 2006-2011 Roadkill Monitoring. Wonju: Korea National Park Research Institute.
Koo, K., Kim, T., Yang, K., and Oh, H. (2018). First observation on the breeding behavior of endangered species, Chinese many-tooth snake, Sibynophis chinensis. J. Asia Pac. Biodivers. 11, 305–307. doi: 10.1016/j.japb.2018.02.003
KOSIS (2020). Yearly Road Statistics. Available online at: https://kosis.kr/eng/statisticsList/statisticsListIndex.do?menuId=M_01_01andvwcd=MT_ETITLEandparmTabId=M_01_01andstatId=1976027andthemaId=#SelectStatsBoxDiv: Korean statistical information service
Kosmala, M., Wiggins, A., Swanson, A., and Simmons, B. (2016). Assessing data quality in citizen science. Front. Ecol. Environ. 14, 551–560. doi: 10.1002/fee.1436
Lee, D.-H., Kim, Y.-C., Chang, M.-H., Kim, S., Kim, D., and Kil, J. (2016). Current status and management of alien turtles in Korea. J. Environ. Impact Assess. 25, 319–332. doi: 10.14249/eia.2016.25.5.319
Lee, J.-H. (2011). Taxonomic status, habitat use, and suitability modeling of the Amur ratsnake (Elaphe schrenckii) (Ph.D. dissertation). Kangwon National University, Chuncheon, South Korea.
Lee, J.-H., Jang, H.-J., and Suh, J.-H. (2012). Ecological Guide Book of Herpetofauna in Korea. Incheon: National Institute of Environmental Research.
Lee, J.-H., and Park, D. (2016). The Encyclopedia of Korean Amphibians. Seoul: Checklist of Organisms in Korea.
Lee, S.-D., and Miller-Rushing, A. J. (2014). Degradation, urbanization, and restoration: a review of the challenges and future of conservation on the Korean Peninsula. Biol. Conserv. 176, 262–276. doi: 10.1016/j.biocon.2014.05.010
Lintott, C., Schawinski, K., Bamford, S., Slosar, A., Land, K., Thomas, D., et al. (2010). Galaxy Zoo 1: data release of morphological classifications for nearly 900 000 galaxies. Monthly Notices R. Astron. Soc. 410, 166–178. doi: 10.1111/j.1365-2966.2010.17432.x
Macias, D., Shin, Y., and Borzée, A. (2021). An update on the conservation status and ecology of Korean terrestrial squamates. J. Nat. Conserv. 60, 125971. doi: 10.1016/j.jnc.2021.125971
Mccardle, L. D., and Fontenot, C. L. J. (2016). The influence of thermal biology on road mortality risk in snakes. J. Thermal Biol. 56, 39–49. doi: 10.1016/j.jtherbio.2015.12.004
Min, J.-H., and Han, G.-S. (2010). A study on the characteristics of road-kills in the Odaesan National Park. Korean J. Environ. Ecol. 24, 46–53.
MOE (2006). Research Report on the Wildlife Roadkill Occurrences on Major Roads in North Gyeongsang Province. Daegu: Ministry of Environment Daegu Regional Office.
NIBR (2019). Red Data Book of Republic of Korea. Volume 2: Amphibians and Reptiles. Incheon: National Institute of Biological Resources, Ministry of Environment.
Othman, S. N., Litvinchuk, S. N., Maslova, I., Dahn, H., Messenger, K. R., Andersen, D., et al. (2022). From Gondwana to the Yellow Sea, evolutionary diversifications of true toads Bufo sp. in the Eastern Palearctic and a revisit of species boundaries for Asian lineages. eLife 11, e70494. doi: 10.7554/eLife.70494
Park, D., Jeong, S.-M., Kim, S.-K., Ra, N.-Y., Lee, J.-H., Kim, J.-K., et al. (2017). Patterns of snake roadkills on the roads in the northeast region of South Korea. Korean J. Environ. Ecol. 31, 42–53. doi: 10.13047/KJEE.2017.31.1.042
Petrovan, S. O., Vale, C. G., and Sillero, N. (2020). Using citizen science in road surveys for large-scale amphibian monitoring: are biased data representative for species distribution? Biodivers. Conserv. 29, 1767–1781. 10.1007/s10531-020-01956-0 doi: 10.1007/s,10531-020-01956-0
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rowley, J., Callaghan, C., Cutajar, T., Portway, C., Potter, K., Mahoney, S., et al. (2019). FrogID: citizen scientists provide validated biodiversity data on frogs of Australia. Herpetol. Conserv. Biol. 14, 155–170.
Schwartz, A. L., Shilling, F. M., and Perkins, S. E. (2020). The value of monitoring wildlife roadkill. Eur. J. Wildlife Res. 66, 1–12. doi: 10.1007/s10344-019-1357-4
Shepard, D., Dreslik, M., Jellen, B., and Phillips, C. (2008). Reptile road mortality around an oasis in the Illinois corn desert with emphasis on the endangered Eastern Massasauga. Copeia 2, 350–359. doi: 10.1643/CE-06-276
Shin, Y., and Borzée, A. (2020). Melanism in the Ussuri pitviper (Gloydius ussuriensis) from the Republic of Korea, with remarks on color variations in the species. Jordan J. Nat. History 7, 60–63.
Shin, Y., Jeong, D., and Borzée, A. (2020). Mass displacement of the Korean clawed salamander (Onychodactylus koreanus) and the threat of road-kill. Herpetol. Bull. 151, 28–31. doi: 10.33256/hb151.2831
Shin, Y., Messenger, K. R., Koo, K. S., Lee, S. C., Hou, M., and Borzée, A. (2021). How threatened is Scincella huanrenensis? An update on threats and trends. Conservation 1, 58–72.
Song, J.-Y., Kim, M.-S., Kim, I.-S., Kim, T.-H., Roh, I., Seo, S.-W., et al. (2009). Roadkill of amphibians in the Korean National Parks. Korean J. Environ. Ecol. 23, 187–193.
Song, J. Y. (2007). Current status and distribution of reptiles in the Republic of Korea. Korean J. Environ. Biol. 25, 124–138.
Swaisgood, R. (2020). Grand challenges in animal conservation. Front. Conserv. Sci. 1, 602856. doi: 10.3389/fcosc.2020.602856
Trombulak, S. C., and Frissell, C. A. (2000). Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol. 14, 18–30. doi: 10.1046/j.1523-1739.2000.99084.x
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics with S. New York, NY: Springer. doi: 10.1007/978-0-387-21706-2
Vercayie, D., and Herremans, M. (2015). “Citizen science and smartphones take roadkill monitoring to the next level,” in Proceedings of IENE 2014 International Conference on Ecology and Transportation, eds A. Seiler and J.-O. Helldin (Malmö). doi: 10.3897/natureconservation.11.4439
Won, C., Yoo, B., Yang, B., Kim, W., Moon, J., Oh, K., et al. (2005). Wildlife Survey. Incheon: National Institute of Environmental Research.
Yang, B., Oh, H., Ku, Y., Hur, W., and Cho, Y. (2006). Improvement of Road's Facilities for Wildlife Road-Kill Reduction - in the Case of Baekdudaegan Region. Incheon: National Institute of Environmental Research.
Keywords: citizen science, conservation, herpetofauna, Republic of Korea, wildlife vehicle collisions
Citation: Shin Y, Kim K, Groffen J, Woo D, Song E and Borzée A (2022) Citizen science and roadkill trends in the Korean herpetofauna: The importance of spatially biased and unstandardized data. Front. Ecol. Evol. 10:944318. doi: 10.3389/fevo.2022.944318
Received: 15 May 2022; Accepted: 11 July 2022;
Published: 12 August 2022.
Edited by:
Silviu O. Petrovan, University of Cambridge, United KingdomReviewed by:
Laurentiu Rozylowicz, University of Bucharest, RomaniaCopyright © 2022 Shin, Kim, Groffen, Woo, Song and Borzée. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amaël Borzée, YW1hZWxib3J6ZWVAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.