
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 28 September 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.928277
This article is part of the Research Topic Fish as Sentinels of Urbanization Impacts in Aquatic Ecosystems View all 6 articles
 Piatã Marques1*†
Piatã Marques1*† Eugenia Zandonà1
Eugenia Zandonà1 Jeferson Amaral1Yasmin Selhorst1
Jeferson Amaral1Yasmin Selhorst1 Rana El-Sabaawi2
Rana El-Sabaawi2 Rosana Mazzoni1Letícia Castro1
Rosana Mazzoni1Letícia Castro1 Andrea Pilastro3
Andrea Pilastro3Urbanization transforms natural and agricultural areas into built landscapes. Such profound habitat alteration imposes strong pressure on phenotypic trait changes through processes related to natural and/or sexual selection. Evidence of how natural selection drives changes to traits in urban biota is increasing, but little is known about the role of sexual selection. In this study, we assessed the effect of urbanization on the expression and interaction of males' pre-mating traits (body size and color) and a post-mating trait (sperm load). We used a widespread invasive species, the guppy (Poecilia reticulata), which is a wellknown model for studying sexual selection, but have never been studied in urban systems for this purpose. We found that urbanization did not affect mean body size or condition, but it resulted in size-dependent reductions in the expression of orange and iridescent colors, as well as sperm load. The orange color was reduced in small urban guppies, while the iridescent colors were reduced in large urban guppies compared to non-urban guppies. The difference in sperm load was only found in large males, with lower sperm load in urban guppies. The relationship between orange color and sperm load was positive in urban guppies but negative in non-urban guppies, while the association between iridescent color and sperm load followed the opposite pattern. Our findings suggest that sexual selection on pre- and post-mating traits is weaker in urban than in non-urban systems and that interactions between such traits are context dependent. These responses can be related to the pollution and altered visual environment of urban systems and provide an opportunity to advance our understanding of the mechanisms determining adaptation in cities.
To satisfy the needs of an increasing urban population, cities are growing at an alarming rate. This urbanization process causes profound habitat modification through deforestation, construction, and pollution (Alberti, 2008). Such changes to the habitat lead to marked biodiversity loss, but some species such as the house sparrow (Passer domesticus) are able to flourish in cities (Shochat et al., 2010).
Understanding how species persist in cities has been a recent focus of urban ecology studies. The changes in characteristics (i.e., traits) that are either heritable (evolutionary) or non-heritable (plasticity) can help organisms adapt and persist in urban environments (Lambert et al., 2021). Urbanization can drive rapid, contemporary changes in traits ranging from behavior to morphology, through processes involving natural and/or sexual selection (Alberti et al., 2017; Tüzün et al., 2017). However, evidence on how natural selection operates in the urban systems and the resulting trait change is scarce, and even less is known about the role of sexual selection (Rivkin et al., 2019; Sepp et al., 2020).
Sexual selection tends to operate more strongly on males, either before or after mating (Janicke et al., 2016). Pre-mating sexual selection often involves traits related to mating success, such as body size and conspicuous ornaments (e.g., bright-colored feathers) that are used to attract females. Post-mating sexual selection involves traits that determine fertilization success following copulation, such as sperm motility and sperm number. Understanding the expression and interaction of pre- and post-mating traits is therefore fundamental for exposing mechanisms shaping sexual selection (Evans and Garcia-Gonzalez, 2016).
A well-known process in sexual selection is the interaction between body size and coloration (pre-mating traits) with fertility (post-mating trait). In many species, larger, colorful males tend to be more fertile (Rogers et al., 2008; Smith et al., 2014). This is because individuals with larger body size are often assumed to be in better condition (i.e., large nutrient storage, better health), which allows increased investment in mating-related traits, such as color and fertility, rather than survival traits (Cotton et al., 2004; Rahman et al., 2013).
Mating-related traits can be strongly affected by the environment. The availability of food and the presence of predators can determine the expression of body size, color, and fertility (Rahman et al., 2013; Schmitz, 2017). Similarly, ambient light conditions, background color, and transmission properties can affect mating color signaling (Hutton et al., 2015). Despite the existing evidence, how the environment affects the interaction between pre- and post-mating traits remains poorly known (Evans and Garcia-Gonzalez, 2016). Such information is central to understanding how human disturbances, such as urbanization, modulate the strength and direction of sexual selection. This is especially important in aquatic urban ecosystems, such as urban streams, which are much less studied than terrestrial urban systems (Marques et al., 2019; Langerhans and Kern, 2020).
The urbanization of streams causes profound habitat modifications, such as pollution, altered hydrology, and food availability, which have the potential to change traits (Marques et al., 2019). However, the extent to which urbanization affects the expression and interaction of pre- and post-mating traits is poorly known (Sepp et al., 2020; Cronin et al., 2022). This information is critical to expose the mechanisms through which aquatic organisms, such as fish, respond and adapt to urbanization (Evans and Garcia-Gonzalez, 2016; Sepp et al., 2020).
In this study, we assessed the effect of urbanization on the expression and interaction of pre-mating traits (body size and mating color) and post-mating traits (sperm load) to explore the strength and direction of sexual selection in urban systems. We tested this by using a widespread invasive species, the guppy (Poecilia reticulata). Guppies are a wellknown model for studying pre- and post-mating sexual selection in their non-urban, native range in Trinidad (Endler, 1984; Kemp et al., 2018). Sexual selection favors larger and more colorful male guppies with larger sperm loads (Pitcher and Evans, 2001). However, existing evidence comes exclusively from non-urban ecosystems, and the effect of urbanization on the expression and interaction between pre- and post-mating traits in guppies and other aquatic animals is unknown. We hypothesize the profound changes urbanization imposes on streams disrupting the expression and interaction between pre- and post-mating traits, as detailed below.
We initially compared pre-mating traits (body size and coloration) between urban and non-urban guppy populations to explore trait changes. Urbanization increases food availability for guppies in urban streams, which can allow greater investment in body size (Snell-Rood et al., 2015; Marques et al., 2020). However, this can be counterbalanced by investment toward survival in the polluted urban environment (Sepp et al., 2018). We expect male urban guppies to be similar in size to non-urban guppies. Also, studies on guppies from non-urban streams in Trinidad suggest pigment-based orange coloration, which can be seen at distance, is the main color used to attract mates (Kodric-Brown, 1989; Long and Houde, 1989). However, urbanization impairs the visual environment (high turbidity and low light incidence) (Walsh et al., 2005), with potential consequences for visual communication. We expect urban guppies to either express less orange coloration or to invest more in iridescent colors, which are more efficient for communicating under poor visual conditions, at proximity (Doucet and Meadows, 2009).
Then, we tested for differences in the post-mating trait (sperm load) between urban and non-urban guppies. Despite the increased food availability, the harsh conditions of the urban environment (e.g., pollution and altered hydrology) can force animals to invest more in self-maintenance traits than in reproduction (Sepp et al., 2018). We expected urban male guppies to show no change or to deliver smaller sperm loads than non-urban guppies.
Finally, we assessed the relationship between size, color, and sperm load to investigate the interaction between pre- and post-mating traits. Urbanization has the potential to affect the strength and direction of the interaction between pre- and post-mating traits, but the evidence is lacking (Evans and Garcia-Gonzalez, 2016; Cronin et al., 2022). In the future, we expect urbanization to alter the relationship between mating traits as seen in non-urban systems.
We used a study system located in Rio de Janeiro, Brazil, as detailed by Marques et al. (2020). In a previous study, we used this system to explore the effect of urbanization on female guppy life history traits (Marques et al., 2020). Now, we took advantage of the same system to focus on male guppy traits. We used a subset of the stream reaches previously considered in Marques et al. (2020). We selected three urban (CAR, CATO, and ELSU) and three non-urban stream reaches (ELLI, JOA, and WPL) invaded by guppies (Supplementary Figure 1). In all sampled reaches, guppies are known to be the only fish species. Each sampling reach was 30 m in length. Urban stream reaches differ from non-urban stream reaches mainly by having a high concentration of fecal coliforms and ammonium, indicating contamination with sewage (Marques et al., 2020).
We collected guppies using an electro fisher, as detailed in Marques et al. (2020). Only adult male guppies (sexually mature and having fully developed sexual organs) were analyzed in this study.
We collected a total of 256 male guppies (140 urban and 116 non-urban) in 2015, 2016, 2017, and 2021. Sampling was replicated at each reach every year, whenever possible. Guppies were transported live in aerated containers to the aquatic facility at the Universidade do Estado do Rio de Janeiro, where they were either anesthetized or euthanized in Tricaine (MS222). Individual guppies were then photographed using Nikon D80®, Nikon Z50®, and Canon G12® cameras. To remove any color variation introduced by the difference in camera models, we calibrated the color spectrum of the photographs to a common standard. For that, we used a color chart with graded colors (X-Rite ColorChecker Passport, X-Rite Inc.) in the background of each photograph. We used the color chart to generate a standard color profile for each camera using the Adobe® Photoshop Light room Classic software (v. 11.0.1). This profile was used to calibrate the colors and white balance in each photograph. No other changes were made to the photographs following color calibration. We photographed only the left side of each individual guppy.
Photographs were uploaded to the Fiji platform of the open source software ImageJ (v.2.3.0/1.53f) for color analysis (Schindelin et al., 2012). We set the scale (pixels/mm) of each image based on a ruler embedded in each photograph, using the command “Set scale” in ImageJ. We calculated body size as the area of the left side of each guppy (mm2) using the “Measure tool” in ImageJ. Guppies have both pigment-based (carotenoid colors: orange, reds, and yellows) and structurally based colors (iridescent colors: blues, greens, and purples) (Kemp et al., 2018). We performed automated color analyses for the two color categories (orange and iridescent) separately, by using the command “Color Threshold” in ImageJ. Thresholding was performed using the HSB color space. The orange category was given by Hue = 44, Saturation = 148–255, and Brightness = 92–255. The iridescent category was defined by Hue = 46–230, Saturation = 63–255, Brightness = 74–255. Thresholding allowed us to select pixels with the specified HSB values for each color category. Then, we defined the minimum size of the particle (i.e., group of pixels) considered to be > 0.2 mm2, using the “Particle size” command in ImageJ. Based on that, we estimated the area (mm2) of each color category per individual. Color analysis was automated by building a macro function, using the “Macro” command in ImageJ, which repeats color thresholding and particle analysis for each color category on each individual photograph.
We tested for differences in body size between urban and non-urban guppies with linear mixed effect models (LMM) using stream reach as a random factor. We tested for an effect of body size on the area of each color category (orange and iridescent) and if the effect of body size and the area of each color category differed between urban and non-urban guppies using the LMM models that included an interaction term between habitat (urban and non-urban) and body size. The color area was square-root transformed. We included data from all years in the models because color and body size have high heritability across generations (Reynolds and Gross, 1992; Brooks and Postma, 2011). All statistical analysis and plots were performed using the R software (R Core Team, 2020).
In addition to color analysis, guppies collected in 2021 (34 urban and 35 non-urban) were assessed for sperm load. At the aquatic facility, guppies were housed for 10 days in six male-only tanks (40 L each) separated by reach identity. The laboratory housing period is important to allow males to replenish sperm reserves depleted right before capture. Tanks were maintained at constant temperature (24°C) and aeration. Guppies were fed daily with brine shrimp.
Following the housing period, individual guppies were retrieved from the tanks, anesthetized in Tricaine (MS222), and photographed following the procedures described above. Then, guppies were weighted (g) and measured for body length (standard length, mm). Under a dissecting microscope, we stripped the sperm bundles (i.e., spermatozeugmata and packages of sperm) by gently massaging the abdomen, as described in Cattelan et al. (2016). Sperm bundles retrieved from each individual guppy were counted, and the total number was used to estimate sperm load. Because an individual's condition (i.e., health) can determine investment in sperm traits, we also estimated individual guppy condition using a weight-length regression (Rahman et al., 2013; Lloret et al., 2014).
We tested for differences in sperm load and condition between habitats (urban and non-urban) with LMMs using stream reach as a random factor. The LMM for sperm load also included body size as a covariate and the interaction between habitat and body size as a fixed factor. All statistical analyses and plots were performed using the R software (R Core Team, 2020).
We built predictive linear mixed models to assess the relationship between size, color, and sperm load using data collected in 2021. We built four separate models, using the area of either color (orange or iridescent) as the response variable, separated by habitat (urban or non-urban). We separated models by habitat because the mechanisms affecting the interaction between color and mating-related traits can widely differ between urban and non-urban streams (Marques et al., 2019). For each model, we used sperm load and body size as fixed factors and reach identity as random factors. Fixed variables were scaled to improve model fit. From each model, we generated a set of sub models having all possible combinations of predictors by removing or including different fixed factors (Grueber et al., 2011). The sub models were subjected to model selection based on AICc criteria (Grueber et al., 2011). Only the models with substantial empirical support (ΔAICc < 2) were retrieved and averaged to estimate the coefficient of each predictor (coefavg; Burnham and Anderson, 2002).
The models were fitted with the nlme package of the R software (Pinheiro et al., 2022). Model fit was evaluated visually using Q-Q plots. We tested the models for multicollinearity with the VIF function of the car package for R (Fox and Weisberg, 2011). The variance explained only by the fixed factors (marginal R square, ) and the variance explained by both fixed plus random factors (conditional R square, ) were estimated using the “r.squaredGLMM” function of the R package MuMIn (Barton, 2008). Model selection and averaging were performed with the “dredge”, “get.models”, and “model.avg” functions of the MuMIn package.
We found that the pigment-based orange coloration was determined by the interaction between habitat (urban and non-urban) and body size (LMM t = −4.1, p < 0.001). This indicated that a reduction in orange color occurred for urban guppies with small body sizes (< 85 mm2) but not for large ones (Figure 1A; Table 1). Similarly, the structurally based iridescent colorations were determined by the interaction between habitat and body size (LMM t = 2.6, p = 0.01), where large urban guppies tended to be less iridescent than non-urban guppies of similar size (Figure 1B; Table 1).
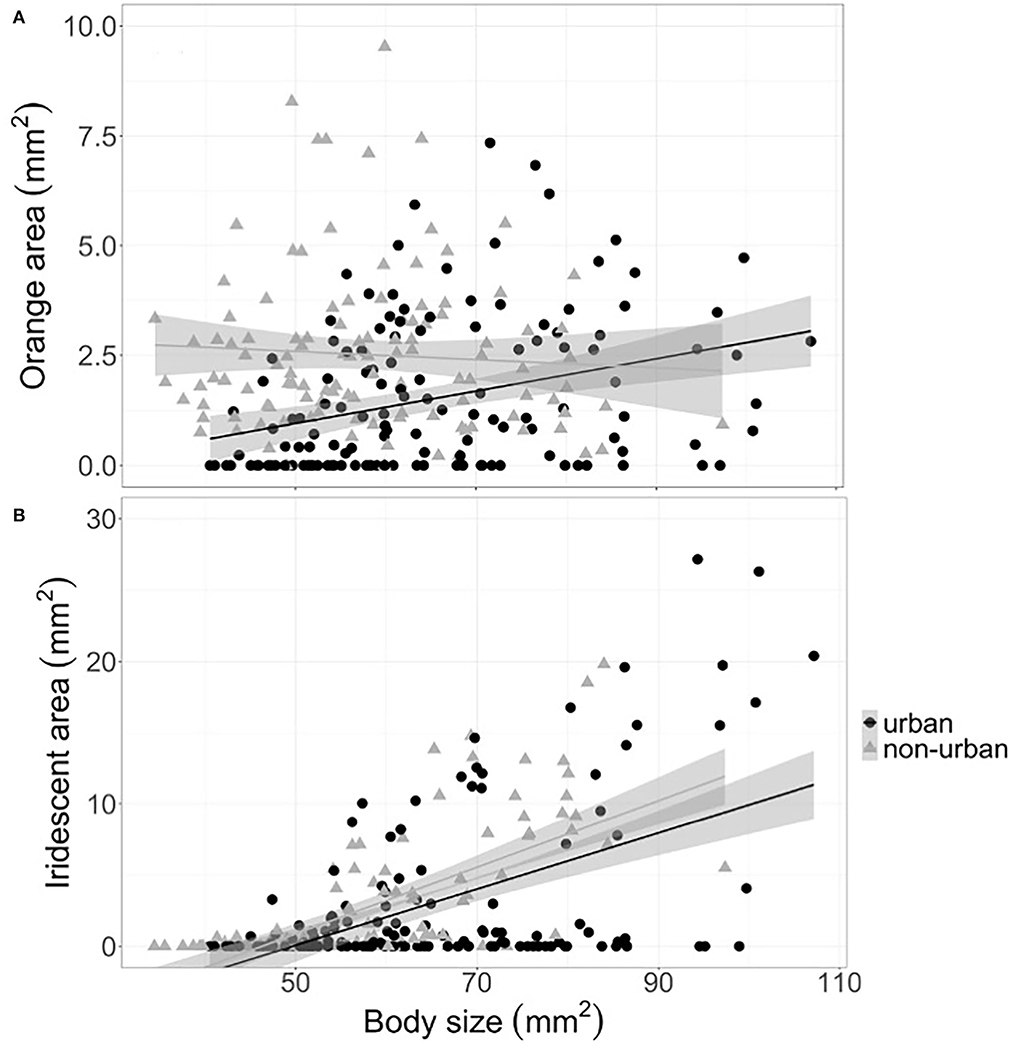
Figure 1. The relationship between (A) orange and (B) iridescent color areas and body size (area of the left side, mm2) for guppies in urban (black circles) and non-urban streams (gray triangles). The color area was calculated on the left side of each individual guppy. The color was defined based on the HSB color space (see Methods section for details). Symbols indicate individual guppy, solid lines show the linear tendency, and shaded areas are standard errors. Data shown combine all sampling years (2015, 2016, 2017, and 2021).
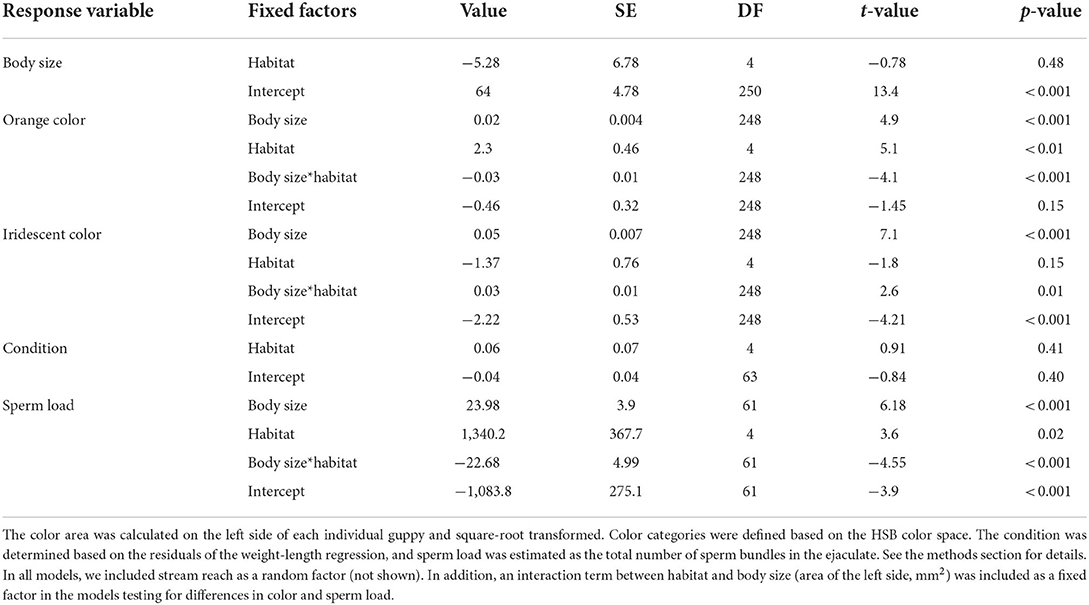
Table 1. The output of the linear mixed effects models was tested for differences in body size (area of the left side), colors (orange and iridescent), sperm load, and conditions between guppies occurring in urban and non-urban streams (habitat).
The sperm load was also determined by the interaction between habitat and body size (LMM t = −4.5, < 0.0001). This suggested that, at larger sizes, urban guppies had lower sperm load than non-urban guppies (Figure 2; Table 1). We found no difference in mean body size or condition between urban and non-urban guppies.
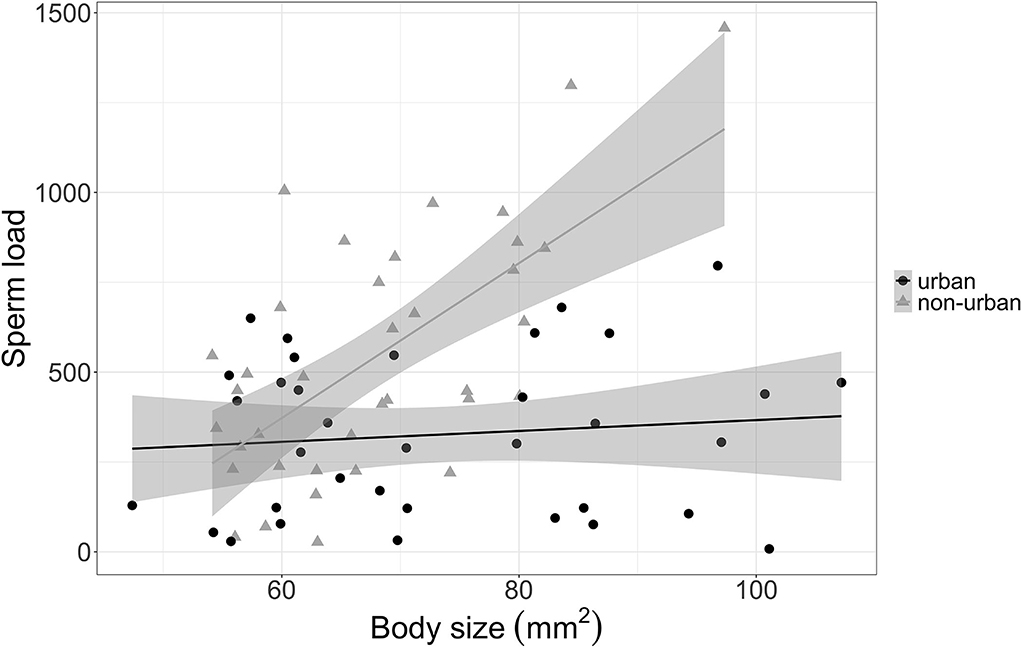
Figure 2. The relationship between sperm load and body size (area of the left side, mm2) for guppies in urban (black circles) and non-urban streams (gray triangles). Sperm load was estimated as the sum number of sperm bundles (spermatozeugmata) per individual. Symbols indicate individual guppy, solid lines show the linear tendency, and shaded areas are standard errors. Data shown include only guppies collected in the 2021 sampling year.
The relationship between individual body size, color, and sperm load varied between urban and non-urban guppies. We found that the orange color of urban guppies was positively related to sperm load (coef = 0.69) (LMM R2m = 0.23, R2c = 0.23), irrespective of body size (i.e., this factor was not included in the best models following model selection) (Table 2). In contrast, the iridescent color of urban guppies was negatively related to sperm load (coefavg = −0.46), with a positive effect on body size (coefavg = 4.87) (LMM R2m = 0.60, R2c = 0.68) (Table 2). The opposite pattern was found for non-urban guppies, for which orange color was negatively related to sperm load (coef = −0.51) (LMM R2m = 0.23, R2c = 0.25), irrespective of body size (Table 2). The iridescent color of non-urban guppies was positively related to sperm load (coefavg = 0.31) and body size (coefavg = 1.91) (LMM R2m = 0.18, R2c = 0.57) (Table 2). The sub models used to calculate coefavg can be found in Supplementary Table 1.
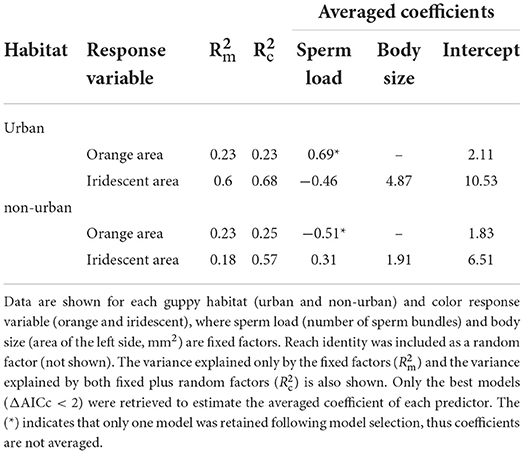
Table 2. Coefficients for each predictive linear mixed model.
Human disturbance can be a strong selective force for trait change with consequences for mate choice (Candolin et al., 2007). However, the extent to which urbanization affects sexual selection is largely unknown (Sepp et al., 2020). Here, we proved that urbanization does not change body size or condition, but it reduces the expression of orange color in smaller guppies and the expression of iridescent colors, especially in larger guppies. Urbanization also reduces the sperm load in large guppies. This suggests that sexual selection is weaker in urban than in non-urban systems. Urbanization also affects the relationship between such traits. The area of orange color is positively related to sperm load in urban guppies but negatively related to sperm load in non-urban guppies, and the opposite pattern is seen for iridescent color. Individual body size is not related to sperm load in urban guppies, but it has a positive effect on the sperm load of non-urban guppies.
Although the effect of urbanization on traits such as body size has been documented in a number of species, its role in sexual selection in urban populations has been poorly explored (Sepp et al., 2020). Evidence from non-urban systems suggests that a larger body size increases the expression of other sexual traits, which can confer fitness benefits (Skinner and Watt, 2007). For that reason, females tend to prefer mating with larger males (Cooper and Vitt, 1993; Marler and Ryan, 1997). However, our data show that the interaction between urbanization and body size reduces the expression of sexual traits. The extent to which this inverse effect of body size on sexual traits affects female preference and sexual selection is yet to be assessed. Laboratory tests on female preference (e.g., Hermann et al., 2015) are needed to further understand the role of body size in the mating of guppies and other urban animals.
Another pre-mating trait of interest is color. Increasing evidence suggests that urban animals decrease the expression of orange carotenoid-based coloration, which could accommodate the expression of more iridescent, structural colors than their non-urban counterparts (Biard et al., 2017; Giraudeau et al., 2018). However, our data show size-dependent reductions in both orange and iridescent colors in urban guppies (Figure 1). This overall decrease in color can be related to the discharge of wastewater into urban streams (Marques et al., 2020). Guppies respond to wastewater pollution by either increasing or decreasing the expression of orange and iridescent colors depending on the environmental context (Gomes-Silva et al., 2020). The specific mechanisms for such change are unclear. Pollution could be related to a low body condition that determines coloration (Peters et al., 2008; White, 2020). However, we found no change in conditions between urban and non-urban guppies. This raises the question of whether color signals remain condition-dependent in urban settings (Hutton and McGraw, 2016). Alternatively, changes to the expression of orange color can be related to altered carotenoid content of urban foods or the impairment of the metabolic conversion of dietary carotenoids caused by urban stressors (Hill, 1992; Giraudeau et al., 2018), but changes to the expression of iridescent colors can be related to passive, energetically inexpensive, changes to the self-assembly of the nanostructures of the integument (Maia et al., 2012). Exploring the drivers of color expression can provide important information on color signaling in guppies and other urban animals.
The effect of urbanization on post-mating traits remains unclear. In mating systems where females mate multiply (polyandry), such as in guppies, males often invest more into post-mating traits, such as sperm load, to improve sperm competition (Hämäläinen et al., 2018). Despite the high guppy density in urban streams, which should lead to increased male-male competition (Marques et al., 2020), we found that, among larger guppies, urban males had reduced sperm load when compared to non-urban ones. This can be related to specific responses to urban environmental stressors that could be affecting sperm production in guppies (Sepp et al., 2018). Empirical evidence is still needed to further understand investment toward post-mating traits in urban ecosystems (Cronin et al., 2022). Increased sample size associated with molecular tools to assess the links between body size, coloration, and paternity in replicated populations of urban and non-urban guppies can help estimate the contribution of pre- and post-mating traits to male sexual success (Devigili et al., 2015; Cattelan et al., 2020; Glavaschi et al., 2020). Common garden experiments can further help disentangle the genetic and environmental components driving the expression of matting traits in guppies and other urban animals (e. g. Reznick et al., 2019).
In addition to changing the expression of pre- and post-mating traits, urbanization affects the relationship between them. For many animals, the body size is related to sperm load suggesting that larger individuals have larger testis and higher sperm load than smaller individuals (Kodric-Brown et al., 2006; Emlen et al., 2012). While such prediction stands true for non-urban guppies in this study and elsewhere (Pitcher and Evans, 2001), our data show that body size is not related to sperm load in urban guppies (Figure 2). To the best of our knowledge, ours is the first study to show that urbanization can decouple the relationship between body size and sperm load. While the mechanisms for such disconnection are unclear, it is likely that the altered conditions of the urban environment force animals to invest more in traits that ensure survival, such as immune defense and parasite/pathogen resistance, rather than reproductive traits such as sperm load (Sepp et al., 2018).
Similarly, urbanization affects the relationship between color and sperm load. Evidence from non-urban systems suggests that male conspicuous color ornaments are positively correlated with fertility (e.g., sperm load) and are thus a good indicator of male quality because of the potential to maximize offspring production (Mautz et al., 2013; Cuthill et al., 2017; White, 2020). Ourdata suggest that such a relationship is context dependent. While orange color is positively related to sperm load in urban guppies, it is the iridescent color that has a positive relationship to sperm load in non-urban guppies. This suggests that males rely on different colors to signal fertility in urban vs. non-urban systems and implies that female preference for male coloration responds accordingly. This could be a response to the changes in the visual environment, mainly related to the high turbidity of urban streams (Paul and Meyer, 2001; Hutton et al., 2015). However, evidence is still lacking. In the future, laboratory tests on female preference under varying visual conditions can help understand the function of orange and iridescent colors in the mating of urban animals (e.g., Couldridge and Alexander, 2002).
Despite the long-standing interest in the interaction between pre- and post-mating episodes of sexual selection, few studies considered how environmental factors influence such relationships (Evans and Garcia-Gonzalez, 2016). Here, we showed that urbanization can profoundly change the interaction between pre- and post-mating traits, with potential consequences for adaptation. For instance, the reduced investment toward color expression associated with low sperm loads could lead us to conclude that urban guppies are striving to persist in their environment. However, guppies are very successful in urban streams, having population densities that are ~26 times higher in urban streams than in non-urban streams with no sign of decline through time (Marques et al., 2020). Such contradictions require further investigation because they can help us understand adaptation in the urban environment (Brady et al., 2019). For example, how the relationship between a male's color and fertility contributes to adaptation might depend on how strongly female fecundity is affected by differences in sperm load (Pilastro et al., 2008). Further understanding of how pre- and post-mating traits vary and interact in both sexes, in multiple species, is fundamental because it can expose the mechanisms through which sexual selection facilitates adaptation in urban environments (Hämäläinen et al., 2018; Brady et al., 2019).
By studying the nature of sexual selection in cities, we can advance evolutionary theory. As shown here, urbanization can challenge some of the classic assumptions about how sexual traits change and interact. This is likely because urbanization imposes novel selective pressures that can lead to unique responses in sexual traits (Sepp et al., 2020). Exposing specific drivers and the direction of sexual trait change in urban ecosystems can help develop key theoretical concepts toward understanding evolution through sexual selection (Cronin et al., 2022). Expanding that knowledge has the potential to provide important information to support biodiversity conservation in cities (Donihue and Lambert, 2015).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by the University of Victoria (2016-008), the State University of Rio de Janeiro (UERJ CEUA/005/2016) Animal Care Committees, and the Brazilian Ministry for the Environment (IBAMA 16152-1).
PM, EZ, RE-S, RM, and AP conceived the ideas for the study. PM and AP designed the experiments and contributed to the writing of the manuscript. PM, JA, YS, and LC collected and processed the data. PM carried out the statistical analyses. All authors contributed to editing drafts and gave final approval for publication.
This research was supported by the Science Without Borders Program (CAPES 1212/13-3 and CNPq 401258/2014-9) with the doctorate fellowship to PM and the visiting professor grant to RE-S. This work was also supported by the CAPES-PrInt Program (88887.369182/2019-00) with a postdoctoral fellowship to PM and a visiting professor grant to AP (88887.311746/2018-00). Support also came from the CNPq (PQ 1D−301463/2017-4 to RM and PQ2 – 308261/2017-8 to EZ) and FAPERJ (CNE–E-01/2016DSC and E-26/202.762/2018 to RM and JCNE–E-26/203.213/2017 and E-26/201.286/2021 to EZ) and Prociência UERJ to RM and EZ.
We thank the editor Dr. Brian Langerhans and two reviewers for their important contributions to the final version of this paper. We also thank Hugo Ricardo at the Universidade do Estado do Rio de Janeiro for processing export permits and the many individuals who helped us with the fieldwork.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.928277/full#supplementary-material
Alberti, M. (2008). Advances in Urban Ecology: Integrating Humans and Ecological Processes in Urban Ecosystems. New York, NY: Springer. doi: 10.1007/978-0-387-75510-6
Alberti, M., Correa, C., Marzluff, J. M., Hendry, A. P., Palkovacs, E. P., Gotanda, K. M., et al. (2017). Global urban signatures of phenotypic change in animal and plant populations. PNAS 114, 8951–8956. doi: 10.1073/pnas.1606034114
Biard, C., Brischoux, F., Meillère, A., Michaud, B., Nivière, M., Ruault, S., et al. (2017). Growing in cities: an urban penalty for wild birds? A study of phenotypic differences between urban and rural great tit chicks (parus major). Front. Ecol. Evol. 5, 79. doi: 10.3389/fevo.2017.00079
Brady, S. P., Bolnick, D. I., Barrett, R. D. H., Chapman, L., Crispo, E., Derry, A. M., et al. (2019). Understanding maladaptation by uniting ecological and evolutionary perspectives. Am. Nat. 194, 495–515. doi: 10.1086/705020
Brooks, R. C., and Postma, E. (2011). “Genetics of male guppy color patterns,” in Ecology and Evolution of Poeciliid Fishes, eds J. P. Evans, A. Pialstro, and I. Schlupp (Chicago: University of Chicago Press), 254–263.
Burnham, K. P., and Anderson, D. R. (2002). Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd Edn. New York, Ny: Springer-Verlag.
Candolin, U., Salesto, T., and Evers, M. (2007). Changed environmental conditions weaken sexual selection in sticklebacks. J. Evol. Biol. 20, 233–239. doi: 10.1111/j.1420-9101.2006.01207.x
Cattelan, S., Evans, J. P., Garcia-Gonzalez, F., Morbiato, E., and Pilastro, A. (2020). Dietary stress increases the total opportunity for sexual selection and modifies selection on condition-dependent traits. Ecol. Lett. 23, 447–456. doi: 10.1111/ele.13443
Cattelan, S., Evans, J. P., Pilastro, A., and Gasparini, C. (2016). The effect of sperm production and mate availability on patterns of alternative mating tactics in the guppy. Anim. Behav. 112, 105–110. doi: 10.1016/j.anbehav.2015.11.024
Cooper, W. E., and Vitt, L. J. (1993). Female mate choice of large male broad-headed skinks. Anim. Behav. 45, 683–693. doi: 10.1006/anbe.1993.1083
Cotton, S., Fowler, K., and Pomiankowski, A. (2004). Condition dependence of sexual ornament size and variation in the stalk-eyed fly Cyrtodiopsis Dalmanni (Diptera: Diopsidae). Evolution 58, 1038–1046. doi: 10.1111/j.0014-3820.2004.tb00437.x
Couldridge, V. C. K., and Alexander, G. J. (2002). Color patterns and species recognition in four closely related species of Lake Malawi cichlid. Behav. Ecol. 13, 59–64. doi: 10.1093/beheco/13.1.59
Cronin, A. D., Smit, J. A. H., Muñoz, M. I., Poirier, A., Moran, P. A., Jerem, P., et al. (2022). A comprehensive overview of the effects of urbanisation on sexual selection and sexual traits. Biol. Rev. 97, 1325–1345. doi: 10.1111/brv.12845
Cuthill, I. C., Allen, W. L., Arbuckle, K., Caspers, B., Chaplin, G., Hauber, M. E., et al. (2017). The biology of color. Science 357, eaan0221. doi: 10.1126/science.aan0221
Devigili, A., Evans, J. P., Di Nisio, A., and Pilastro, A. (2015). Multivariate selection drives concordant patterns of pre- and postcopulatory sexual selection in a livebearing fish. Nat. Commun. 6, 8291. doi: 10.1038/ncomms9291
Donihue, C. M., and Lambert, M. R. (2015). Adaptive evolution in urban ecosystems. AMBIO 44, 194–203. doi: 10.1007/s13280-014-0547-2
Doucet, S. M., and Meadows, M. G. (2009). Iridescence: a functional perspective. J. Royal Soc. Interface 6, S115–S132. doi: 10.1098/rsif.2008.0395.focus
Emlen, D. J., Warren, I. A., Johns, A., Dworkin, I., and Lavine, L. C. (2012). A mechanism of extreme growth and reliable signaling in sexually selected ornaments and weapons. Science 337, 860–864. doi: 10.1126/science.1224286
Endler, J. A. (1984). “Natural and sexual selection on color patterns in poeciliid fishes,” in Evolutionary Ecology of Neotropical Freshwater Fishes, ed T. M. Zaret (Dordrecht: Springer), 95–111. doi: 10.1007/978-94-015-7682-6_7
Evans, J. P., and Garcia-Gonzalez, F. (2016). The total opportunity for sexual selection and the integration of pre- and post-mating episodes of sexual selection in a complex world. J. Evol. Biol. 29, 2338–2361. doi: 10.1111/jeb.12960
Fox, J., and Weisberg, S. (2011). A R Companion to Applied Regression, 2nd Edn. Thousand Oaks CA: Sage.
Giraudeau, M., Toomey, M. B., Hutton, P., and McGraw, K. J. (2018). Expression of and choice for condition-dependent carotenoid-based color in an urbanizing context. Behav. Eco. 29, 1307–1315. doi: 10.1093/beheco/ary093
Glavaschi, A., Cattelan, S., Grapputo, A., and Pilastro, A. (2020). Imminent risk of predation reduces the relative strength of postcopulatory sexual selection in the guppy. Philos. Trans. Royal Soc. B Biol. Sci. 375, 20200076. doi: 10.1098/rstb.2020.0076
Gomes-Silva, G., Cyubahiro, E., Wronski, T., Riesch, R., Apio, A., and Plath, M. (2020). Water pollution affects fish community structure and alters evolutionary trajectories of invasive guppies (Poecilia reticulata). Sci. Total Environ. 730, 138912. doi: 10.1016/j.scitotenv.2020.138912
Grueber, C. E., Nakagawa, S., Laws, R. J., and Jamieson, I. G. (2011). Multimodel inference in ecology and evolution: challenges and solutions. J. Evol. Biol. 24, 699–711. doi: 10.1111/j.1420-9101.2010.02210.x
Hämäläinen, A., Immonen, E., Tarka, M., and Schuett, W. (2018). Evolution of sex-specific pace-of-life syndromes: causes and consequences. Behav. Ecol. Sociobiol. 72, 50. doi: 10.1007/s00265-018-2466-x
Hermann, C. M., Brudermann, V., Zimmermann, H., Vollmann, J., and Sefc, K. M. (2015). Female preferences for male traits and territory characteristics in the cichlid fish Tropheus moorii. Hydrobiologia 748, 61–74. doi: 10.1007/s10750-014-1892-7
Hill, G. E. (1992). Proximate basis of variation in carotenoid pigmentation in male house finches. Auk 109, 1–12. doi: 10.2307/4088262
Hutton, P., and McGraw, K. J. (2016). “Urban impacts on oxidative balance and animal signals,” Frontiers in Ecology and Evolution 4. Available online at: https://www.frontiersin.org/articles/10.3389/fevo.2016.00054 (accessed August 2, 2022).
Hutton, P., Seymoure, B. M., McGraw, K. J., Ligon, R. A., and Simpson, R. K. (2015). Dynamic color communication. Curr. Opin. Behav. Sci. 6, 41–49. doi: 10.1016/j.cobeha.2015.08.007
Janicke, T., Häderer, I. K., Lajeunesse, M. J., and Anthes, N. (2016). Darwinian sex roles confirmed across the animal kingdom. Sci. Adv. 2, e1500983. doi: 10.1126/sciadv.1500983
Kemp, D. J., Batistic, F.-K., and Reznick, D. N. (2018). Predictable adaptive trajectories of sexual coloration in the wild: evidence from replicate experimental guppy populations. Evolution 72, 2462–2477. doi: 10.1111/evo.13564
Kodric-Brown, A. (1989). Dietary carotenoids and male mating success in the guppy: an environmental component to female choice. Behav. Ecol. Sociobiol. 25, 393–401. doi: 10.1007/BF00300185
Kodric-Brown, A., Sibly, R. M., and Brown, J. H. (2006). The allometry of ornaments and weapons. Proc. Natl. Acad. Sci. U.S.A. 103, 8733–8738. doi: 10.1073/pnas.0602994103
Lambert, M. R., Brans, K. I., Des Roches, S., Donihue, C. M., and Diamond, S. E. (2021). Adaptive evolution in cities: progress and misconceptions. Trends Ecol. Evol. 36, 239–257. doi: 10.1016/j.tree.2020.11.002
Langerhans, B. R., and Kern, E. M. A. (2020). “Urbanization and evolution in aquatic environments,” in Urban Evolutionary Biology, eds M. Szulkin, J. Munshi-South, and A. Charmantier (New York: Oxford University Press), 157–174. Available online at: https://academic.oup.com/book/41750/chapter/354158772 (accessed August 5, 2022).
Lloret, J., Shulman, G., and Love, R. M. (2014). Condition and Health Indicators of Exploited Marine Fishes. New York, NY: John Wiley & Sons. doi: 10.1002/9781118752777
Long, K. D., and Houde, A. E. (1989). Orange spots as a visual cue for female mate choice in the guppy (Poecilia reticulata). Ethology 82, 316–324. doi: 10.1111/j.1439-0310.1989.tb00511.x
Maia, R., Macedo, R. H. F., and Shawkey, M. D. (2012). Nanostructural self-assembly of iridescent feather barbules through depletion attraction of melanosomes during keratinization. J. Royal Soc. Interface 9, 734–743. doi: 10.1098/rsif.2011.0456
Marler, C. A., and Ryan, M. J. (1997). Origin and maintenance of a female mating preference. Evolution 51, 1244–1248. doi: 10.1111/j.1558-5646.1997.tb03971.x
Marques, P. S., Manna, L. R., Frauendorf, T. C., Zandon,à, E., Mazzoni, R., and El-Sabaawi, R. (2020). Urbanization can increase the invasive potential of alien species. J. Anim. Ecol. 1:11. doi: 10.1111/1365-2656.13293
Marques, P. S., Manna, L. R., Mazzoni, R., and El-Sabaawi, R. (2019). Intraspecific trait variation in urban stream ecosystems: toward understanding the mechanisms shaping urban stream communities. Freshw. Sci. 38. doi: 10.1086/701652
Mautz, B. S., Møller, A. P., and Jennions, M. D. (2013). Do male secondary sexual characters signal ejaculate quality? A meta-analysis: secondary sexual characters and ejaculate quality. Biol. Rev. 88, 669–682. doi: 10.1111/brv.12022
Paul, M. J., and Meyer, J. L. (2001). Streams in the urban landscape. Ann. Rev. Ecol. Syst. 32:333–365. doi: 10.1146/annurev.ecolsys.32.081501.114040
Peters, A., Delhey, K., Andersson, S., Van Noordwijk, H., and Förschler, M. I. (2008). Condition-dependence of multiple carotenoid-based plumage traits: an experimental study. Funct. Ecol. 22, 831–839. doi: 10.1111/j.1365-2435.2008.01437.x
Pilastro, A., Gasparini, C., Boschetto, C., and Evans, J. P. (2008). Colorful male guppies do not provide females with fecundity benefits. Behav. Ecol.19, 374–381. doi: 10.1093/beheco/arm140
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., and R Core Team (2022). nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-159.
Pitcher, T. E., and Evans, J. P. (2001). Male phenotype and sperm number in the guppy (Poecilia reticulata). Cana. J. Zool. 79. doi: 10.1139/z01-142
R Core Team (2020). R: A Language and Environment for Statistical Computing. Available onlin at: https://www.r-project.org/ (accessed May 27, 2022).
Rahman, M. M., Kelley, J. L., and Evans, J. P. (2013). Condition-dependent expression of pre- and postcopulatory sexual traits in guppies. Ecol. Evol. 3, 2197–2213. doi: 10.1002/ece3.632
Reynolds, J. D., and Gross, M. R. (1992). Female mate preference enhances offspring growth and reproduction in a fish, Poecilia reticulata. Proceed. Royal Soc. Lond. Ser. B Biol. Sci. 250, 57–62. doi: 10.1098/rspb.1992.0130
Reznick, D. N., Bassar, R. D., Handelsman, C. A., Ghalambor, C. K., Arendt, J., Coulson, T., et al. (2019). Eco-evolutionary feedbacks predict the time course of rapid life-history evolution. Am. Nat. 194, 671–692. doi: 10.1086/705380
Rivkin, L. R., Santangelo, J. S., Alberti, M., Aronson, M. F. J., Keyzer, C. W., de Diamond, S. E., et al. (2019). A roadmap for urban evolutionary ecology. Evol. Appl. 12, 384–398. doi: 10.1111/eva.12734
Rogers, D. W., Denniff, M., Chapman, T., Fowler, K., and Pomiankowski, A. (2008). Male sexual ornament size is positively associated with reproductive morphology and enhanced fertility in the stalk-eyed fly Teleopsis dalmanni. BMC Evol Biol 8, 236. doi: 10.1186/1471-2148-8-236
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schmitz, O. (2017). Predator and prey functional traits: understanding the adaptive machinery driving predator–prey interactions. F1000Research 6, 1–10. doi: 10.12688/f1000research.11813.1
Sepp, T., McGraw, K. J., and Giraudeau, M. (2020). “Urban sexual selection,” in Urban Evolutionary Biology (Oxford: Oxford University Press). doi: 10.1093/oso/9780198836841.003.0014
Sepp, T., McGraw, K. J., Kaasik, A., and Giraudeau, M. (2018). A review of urban impacts on avian life-history evolution: Does city living lead to slower pace of life? Glob. Chang. Biol. 24, 1452–1469. doi: 10.1111/gcb.13969
Shochat, E., Lerman, S. B., Anderies, J. M., Warren, P. S., Faeth, S. H., and Nilon, C. H. (2010). Invasion, competition, and biodiversity loss in urban ecosystems. BioScience 60, 199–208. doi: 10.1525/bio.2010.60.3.6
Skinner, A. M. J., and Watt, P. J. (2007). Phenotypic correlates of spermatozoon quality in the guppy, Poecilia reticulata. Behav. Ecol. 18, 47–52. doi: 10.1093/beheco/arl049
Smith, C., Phillips, A., Polačik, M., and Reichard, M. (2014). Male coloration signals direct benefits in the European bitterling (Rhodeus amarus). Environ. Biol. Fish 97, 335–341. doi: 10.1007/s10641-013-0155-1
Snell-Rood, E., Cothran, R., Espeset, A., Jeyasingh, P., Hobbie, S., and Morehouse, N. I. (2015). Life-history evolution in the anthropocene: effects of increasing nutrients on traits and trade-offs. Evol. Appl. 8, 635–649. doi: 10.1111/eva.12272
Tüzün, N., Beeck, L. O., and de Stoks, R. (2017). Sexual selection reinforces a higher flight endurance in urban damselflies. Evol. Appl. 10, 694–703. doi: 10.1111/eva.12485
Walsh, C. J., Roy, A. H., Feminella, J. W., Cottingham, P. D., Groffman, P. M., and Morgan, R. P. (2005). The urban stream syndrome: current knowledge and the search for a cure. J. North Am. Benthol. Soc. 24, 706–723. doi: 10.1899/04-028.1
Keywords: intraspecific trait, urban evolution, urban stream syndrome, urban ecology, polyandry
Citation: Marques P, Zandonà E, Amaral J, Selhorst Y, El-Sabaawi R, Mazzoni R, Castro L and Pilastro A (2022) Using fish to understand how cities affect sexual selection before and after mating. Front. Ecol. Evol. 10:928277. doi: 10.3389/fevo.2022.928277
Received: 25 April 2022; Accepted: 09 September 2022;
Published: 28 September 2022.
Edited by:
R. Brian Langerhans, North Carolina State University, United StatesReviewed by:
Rudiger Riesch, University of London, United KingdomCopyright © 2022 Marques, Zandonà, Amaral, Selhorst, El-Sabaawi, Mazzoni, Castro and Pilastro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Piatã Marques, cGlhdGFfYmlvQHlhaG9vLmNvbS5icg==
†Present address: Piatã Marques, Department of Environment and Sustainability, University at Buffalo, Buffalo, NY, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.