 Li Zhang
Li Zhang Hai-Li Zhang
Hai-Li Zhang Yukai Chen
Yukai Chen Mir Muhammad Nizamani
Mir Muhammad Nizamani Qin Zhou
Qin Zhou Xiting Su
Xiting Su
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 20 September 2022
Sec. Urban Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.922829
This article is part of the Research Topic Distribution Patterns, Driving Mechanisms and Ecological Service Functions of Urban Plant Biodiversity View all 6 articles
Hopea hainanensis is a large evergreen tree belonging to the family Dipterocarpaceae. It is currently listed as an endangered species on the International Union for Conservation of Nature’s (IUCN) Red List of Threatened Species. It is found primarily on China’s Hainan Island and northern Vietnam. To protect the plant species with extremely small populations Hopea hainanensis and understand its community stability and the inter-specific associations between Hopea hainanensis and other occurring species, we set up 16 quadrats in the areas where wild Hopea hainanensis populations occurred on Hainan Island to investigate the composition of plant species in the quadrats. We used the M. Godron Community stability index to quantify community stability. Based on the two × two contingency tables, the statistic X2, association coefficient (AC), and Pearson correlation coefficient (PCC), percentage co-occurrence (PC), and OI index (OI) analyze the degree of association between Hopea hainanensis and the dominant species in each layer. The results showed that the relationship between Hopea hainanensis and the dominant species in the community was not significant. In addition, there were both positive and negative associations between Hopea hainanensis and other examined six tree species, positive associations with 13 shrub species, positive associations with two herbs, and negative associations with species of 7 species. The connection between Hopea hainanensis and herbs is more complex than that between trees, shrubs, and the dominant species. M. Godron’s stability analysis showed that the community where Hopea hainanensis occurred is relatively unstable (36.84/63.16). The ecology is somewhat fragile, indicating that the community is still at the early or middle stage of competing with other species for resource utilization. The stability of community structure and function is a suitable environment for Hopea hainanensis and finally achieve the purpose of adequate protection, we should choose species with strong positive associations for protecting and recovering Hopea hainanensis.
Hopea hainanensis is one of the Red List of Threatened Species of the International Union for Conservation of Nature (IUCN), mainly distributed in Hainan Island of China and northern Vietnam, and is one of the representative tree species of tropical rain forests in China (Satiraphan, 2012; Ly et al., 2018). It is mainly distributed in mountain valleys, streams, and hillsides with an altitude of 400–800 m in tropical rainforest areas such as Jianfengling, Bawangling, and Diaoluo Mountain in Hainan Province, China (Chen et al., 2015). Despite the high quality of its wood, it has been severely harvested by humans, resulting in a rapid decline in its population. Due to the excessive felling of mature trees on H. hainanensis, the population has decreased by 50–70% in recent decades (Ly et al., 2018). The slow recovery of the H. hainanensis population in the wild state has transformed it from a representative tree species of tropical rainforest to a rare species (Wang et al., 2007; Mehmood et al., 2018). Due to the narrow distribution area of H. hainanensis and the small number of individuals, the sensitivity to the external environment has attracted many scholars at home and abroad to study them.
Rare and endangered plant species typically have few individuals that live in specific communities and ecosystems. The mass ratio hypothesis suggests that the community and ecosystem processes and functions are influenced mainly by the dominant (or sub-dominant, or both) species other than endangered species and their traits (Grime, 1998; Sonkoly et al., 2019). Because the dominant species are more important than the endangered species and their traits (Lohbeck et al., 2015). Rare species also play essential roles in ecosystems, especially species with unique characteristics and intraspecific variations (Powell and Rillig, 2018). The spread of their propagules is necessary for the preservation of biodiversity. They are identifying and measuring functional traits of endangered species that show how they respond to changes in the environment and how the ecosystem functions could help us understand the way these species become endangered or threatened (Turner et al., 2017; Álvarez -Yépiz et al., 2019). They could help us better understand how these species become endangered or threatened, especially under the uncertainty of climate changes in the future (Di Musciano et al., 2020).
The interspecific association is an essential structural feature of plant communities. Interspecific connectivity is the basis for classifying plant communities, community evolution, and community structuring (Miyadokoro et al., 2003; Schnitzler and Closset, 2003). For instance, interspecific association analyses can investigate the changes in interspecific relationships during the stabilizing and succession of a forest community (Chen et al., 2021). Studying the interspecific connectivity and community stability is essential for the evaluation of the coexistence mechanisms in the communities where the rare and endangered plants occur and thereby can provide the theoretical basis for conservation strategies (Zhang et al., 2022).
Currently, studies on the population characteristics of H. hainanensis mainly focus on the geographical distribution patterns, population development characteristics, habitat survey and prediction, the relationship between seedling distribution and parent tree, seedling classification, chloroplast genome characteristics, and phylogeny (Song et al., 2020; Mehmood et al., 2022). Likewise, research regarding the conservation of H. hainanensis mainly focuses on the ex-situ conservation and seedling breeding, etc. This study focuses on the distribution and habitat characteristics, population structure types, wild seed dispersal, seedling growth, and ex-situ conservation of H. hainanensis in Hainan. Environmental selection and seedling cultivation, and the phylogenetic relationship of Dipterocarpaceae. Ecological selection and seedling cultivation for ground protection, the phylogenetic relationship of Dipterocarpaceae plants, etc. However, few studies at the community level, inter-specific association, and community stability of H. hainanensis.
To protect the endangered species H. hainanensis, and understand its community stability and inter-specific associations, we set up 16 quadrats (20 m × 20 m) in the areas where wild H. hainanensis population occurs on Hainan Island to investigate the composition of plant species in the quadrats. Then we used the analysis method combining population connectivity and community stability to explore the connectivity of H. hainanensis and the associated species and their status in the community, to predict changes in community structure and succession trends, and to provide scientific information for the restoration and reconstruction of H. hainanensis.
Hainan (E108°37i-111°031n N18°101-20°101n is a tropical and continental island off the southern coast of the Chinese mainland (Zhang et al., 2022). It is the largest island city (Figure 1), covering 35,400 km2.1 Hainan Island has a tropical monsoon climate with a mild climate, long summers, and short winters, with an average annual temperature of 22–27°C and abundant rainfall. It is rich in forest resources and a vital distribution area of China’s monsoon and tropical rain forests. The main forest vegetation types of Hainan Island include tropical rain forests, mangroves, tropical monsoon forests, evergreen broad-leaved forests, coniferous forests, artificial forests, etc. Plant components mostly have tropical, subtropical, and temperate properties (Zhang et al., 2022). Biodiversity is relatively rich in the central and southern mountainous areas, with more rare and endangered wild plants. At the same time, the northern and coastal regions have relatively low biodiversity and fewer rare and endangered species due to the greater intensity of human disturbance (Guo et al., 2006).
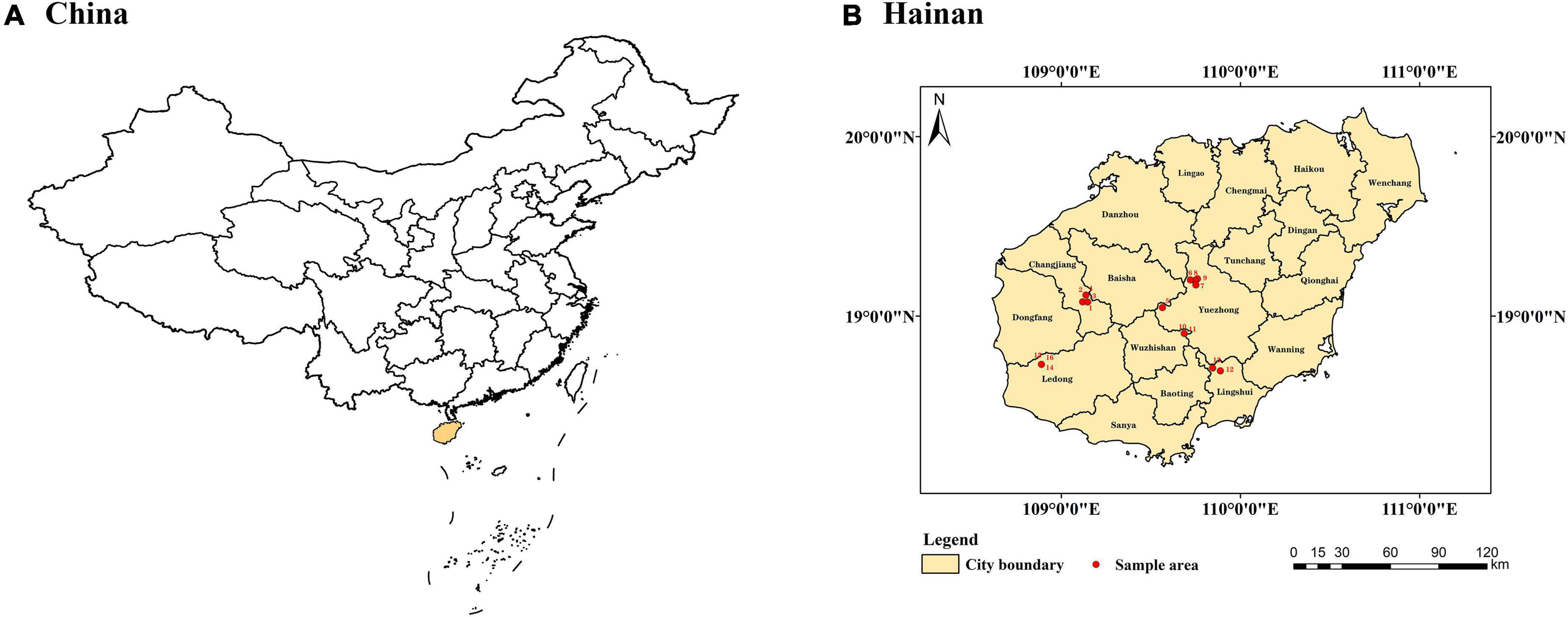
Figure 1. (A) Hainan Island is located in the yellow region of China. (B) Basic status of Hopea hainanensis populations in Hainan Island, China.
A total of 16 quadrats (20 m × 20 m) were set up in the area of wild H. hainanensis and distributed in Hainan (Figure 1 and Table 1). The adjacent grid method is then divided into 16 quadrats (5 m × 5 m). Each quadrat is 400 m2, and the total of 16-quadrat area is 6,400 m2. We investigate the plants in the quadrats by tree layer, shrub layer, and herb layer, respectively. Among them, the species name, height, diameter at breast height, crown width, etc., these species are recorded in the tree layer and the shrub layer, respectively. A small quadrat of (l m × l m) in the herb layer is selected at the four corners of each quadrat, and the species name, height, coverage, number of plants, etc. At the same time, the GPS coordinates, altitude, slope, slope aspect, slope position, natural regeneration, stand growth, threatened status, and degree of human interference of each plot were recorded. Species within the plot were identified using the “An Illustrated Flora of Hainan” (Yang, 2015). In the survey, we found 366 species of 228 genera in 92 families.
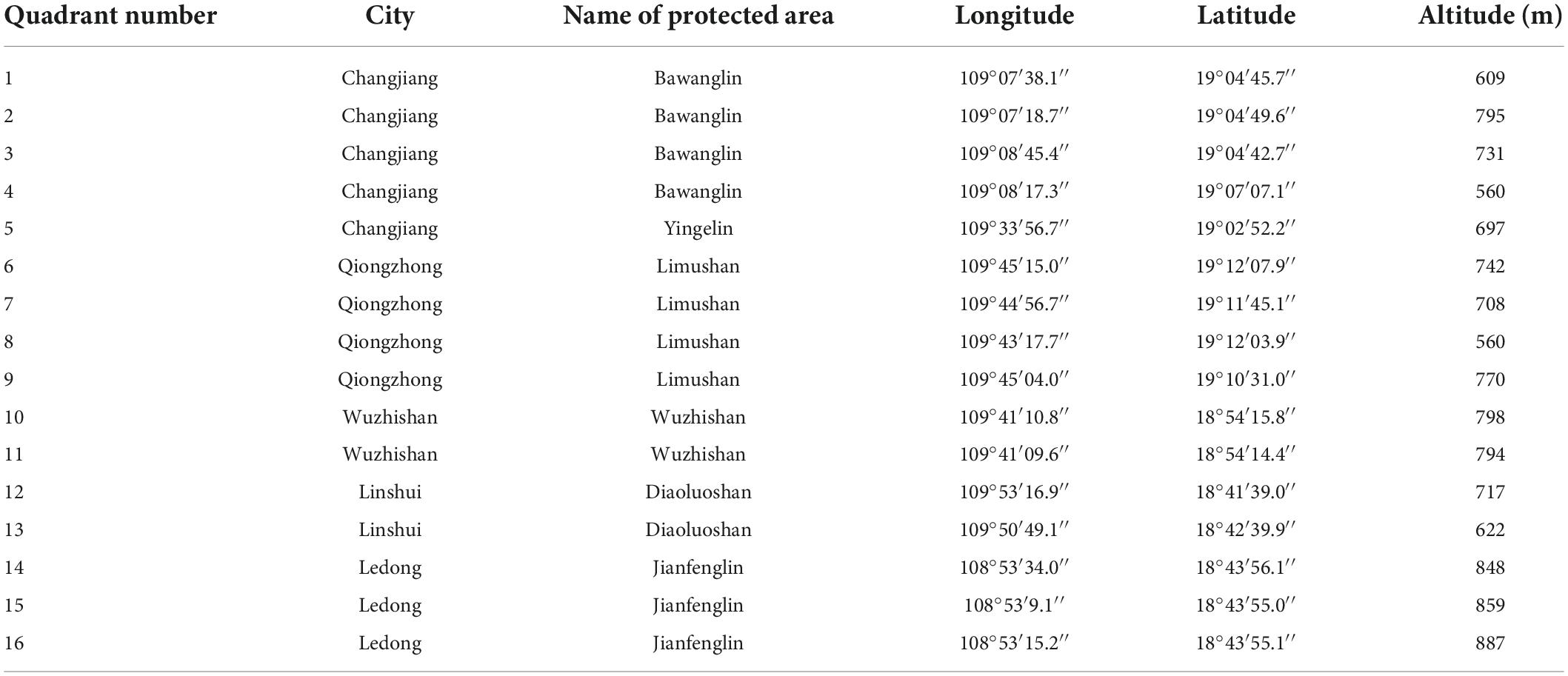
Table 1. Basic status of Hopea hainanensis populations in Hainan Island.
Importance value (IV) = [(relative density (RA) + relative frequency (RF) + relative significance (RP)]/3.
The overall correlation between species reflects whether there is a significant correlation between multiple species that appear in a particular place, and the overall correlation is determined by the variance ratio method (VR). First, make the null hypothesis, that is, there is no significant correlation between the study species, and use the following formula to calculate the test statistic (Wang et al., 2021) as follows (1)–(3):
In the formula, S is the total number of species, N is the total number of the quadrat, Tj is the total number of species appearing in quadrat j, ni is the quadrat number of species i occurring, and t is the average number of populations in the quadrat. The VR value is the overall connection index between plant species in the community. VR = 1, that is, there is no correlation among all species; VR>1 means that the overall performance of the species is positive, and VR<1 means that the overall performance of the species is negative.
The statistic W is used to test the significant degree of VR value deviation by 1, W = N × VR. If the interspecies correlation is not significant, the probability of W falling within the limit of X20.95(N)<W<X20.05(N) has a 90% probability.
The X2 statistic is often used to measure the deviation between the observed and expected values on a probabilistic basis. The connectivity of paired species was determined by X2 according to a 2 × 2 contingency table, and calculated by using Yates’s continuity correction formula as follows (4):
N represents the total number of squares, a is the number of squares with both species present. b is the number of squares with species B, but no species A. c is the number of squares with species A but no species B. d is no quadratic number for both species (the same below). If X2<3.841 (P > 0.05), it means that the species-pair connection is not significant; if 3.841 < X2 < 6.635 (0.01 < P < 0.05), it means that the species-pair connection is substantial; if X2> 6.635 (P < 0.01), then indicates that species pair linkages are incredibly significant. When ad > bc, it is determined to be a positive connection. Otherwise, it is a negative connection.
The X2 test can more accurately reflect whether the association between species is significant, but species pairs that are not significantly associated still have a particular association (Huang et al., 2015). To further determine the degree of association, point correlation coefficient (PCC), association coefficient (AC), Ochiai index (OI), and percent co-occurrence (PC) were used for analysis. The formula is as follows (5)–(9):
When calculating the connection coefficient AC, to avoid a, b, or d being 0 and causing AC = 1 or AC = –1 to be unable to compare the degree of correlation, weight a, b, c, d all 0–1.
The M. Godron stability measurement method uses the relationship between the relative frequency of each species in the plot and the number of species to determine stability. The relative frequencies of different species in the quadrat are gradually accumulated and added in order from large to small. Then we match them with the reciprocal percentage of species and establish a fuzzy scatter smooth curve model (11), and finally, the coordinates of the curve and straight-line intersection (12) are calculated) (Zhang et al., 2022). The equation is as follows: (11)–(14):
Smooth Curve Simulation Model,
Straight line equation,
The two equations are brought into the solution to obtain the following formula,
All populations of H. hainanensis showed a net positive association. The degree of deviation of the VR value is tested by the statistic W. The value of W falls between X2(0.95, N) and X2(0.05, N), which means that there is 90% reliability that the deviation of VR from 1 is not significant. The community shows no significant positive associations, overall, reflecting the consistency of the populations in the community on habitat requirements. Still, the community succession was at the early stage with poor community maturity and stability. Therefore, the overall association degree of the community can also reflect the stability of the community (Table 2).

Table 2. Overall inter specific associations among dominant populations of Hopea hainanensis community.
Based on the overall association test of the community, based on the 2 × 2 contingency table, the statistic X2, association coefficient (AC) and Pearson correlation coefficient (PCC), percentage co-occurrence (PC), and OI index (OI), we analyze the degree of association between H. hainanensis and dominant species in each layer.
The association results between H. hainanensis and the twelve dominant species with tree layer importance value > 1 (Table 3). H. hainanensis and tree species pair statistics X2t<3.841 (P>0.05). The results show that H. hainanensis is not significantly associated with all dominant tree species. From the results of AC and PCC, six species are positively linked to H. hainanensis, and six species are negatively correlated. there are five species with │AC│ < 0.1, and these species accounted for 41.77% of the total number of arbor plants. They are Cyclobalanopsis patelliformis, Sarcosperma laurinum, Gironniera subaequalis, Hancea hookeriana, and Vatica mangachapoi, which are positively connected with H. hainanensis. The OI index and PC value of the Sarcosperma laurinum are the largest, indicating H. hainanensis and Sarcosperma laurinum have a higher probability of co-occurring, and the connection between the Sarcosperma laurinum stronger among the 5 species pairs (Table 4). There were seven species with 0.1<│AC│<0.3, accounting for 58.33% of the total number of arbor plants and H. hainanensis showed a positive association with Sarcosperma laurinum and showed a negative association with Nephelium topengii, Polyalthia laui, Walsura robusta, Engelhardia roxburghiana, Cryptocarya chinensis, and Polyosma cambodiana, and revealing that they have a low correlation coefficient.
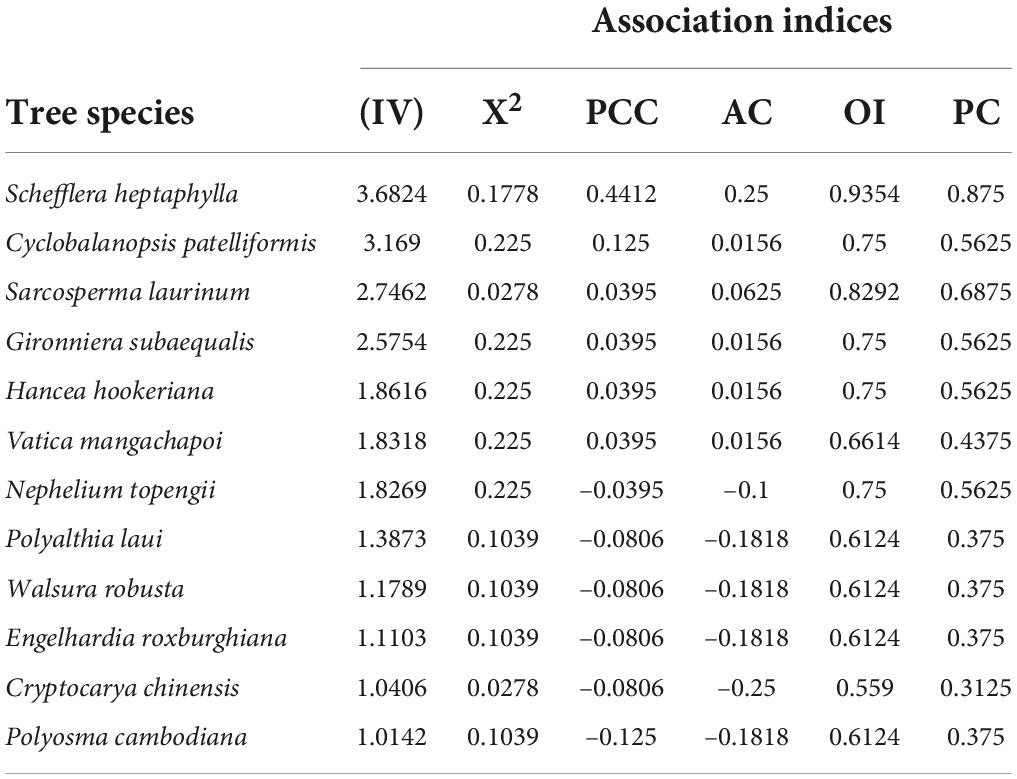
Table 3. Indices of inter-specific association between Hopea hainanensis and tree species.
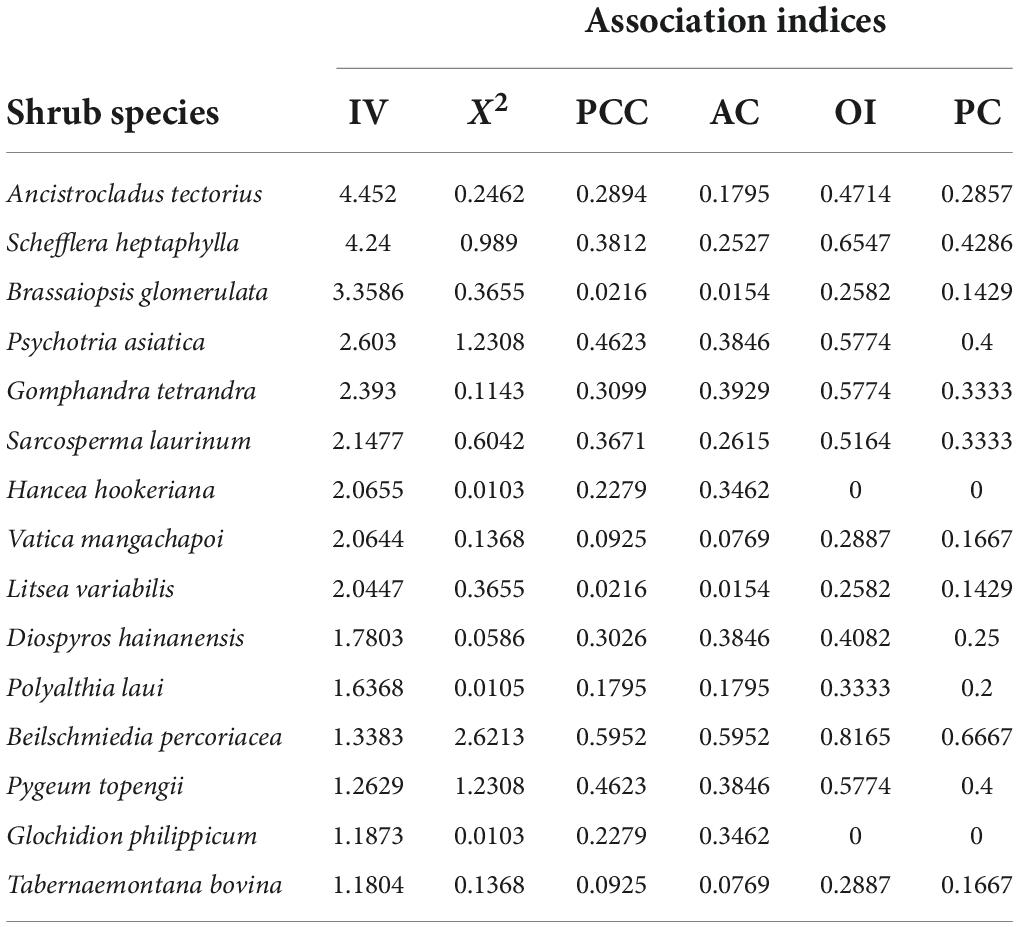
Table 4. Indices of inter-specific association between Hopea hainanensis and shrub species.
The association results between H. hainanensis and the dominant species in the shrub layer (Table 4). H. hainanensis and shrub species pair statistics X2t < 3.841 (P > 0.05), indicate that H. hainanensis is not significantly associated with all shrub’s dominant species. From the results of AC and PCC, H. hainanensis was positively associated with all shrub’s dominant species. There is only one species with │AC│ > 0.5, which is Beilschmiedia percoriacea, and its PCC, AC, OI, and PC value are all ranked first. There are six species with 0.3<│AC│<0.5, accounting for 40% of the total number of shrubs. The PC value of the Psychotria asiatica and Pygeum topengii is higher, and the positive connection with the H. hainanensis is closer. There are four species with 0.1 <│AC│<0.3: Ancistrocladus tectorius, Schefflera heptaphylla, Sarcosperma laurinum, and Polyalthia laui. There are four species with │AC│<0.1: Brassaiopsis glomerulata, Vatica mangachapoi, Litsea variabilis, Tabernaemontana bovina. The OI and PC value of Hancea hookeriana and Glochidion philippicum are both 0. The results show that H. hainanensis is utterly independent of the species of Hancea hookeriana and Glochidion philippicum.
The association results of the top 14 herb dominant species with importance value (IV) of H. hainanensis and herb layer. The statistic of H. hainanensis and herb species pair X2t < 3.841 (P > 0.05), indicates that the association between H. hainanensis and dominant herb species is not significant (Table 5). From the results of AC and PCC, two species are positively linked to H. hainanensis, and 12 species are negatively correlated. There are two species with 0.3<│AC│pe<0.5, Diplazium donianum and Selaginella doederleinii. However, the OI and PC value of these two species are both 0, indicating that there is no correlation with H. hainanensis, and the species are entirely independent. There are eight species with 0.1<│AC│pe<0.3, accounting for 57.14% of the total number of herbs. Among them, the OI and PC value of Licuala fordiana, Allantodia calogramma, and Daemonorops jankinsianus are 0, and there is no correlation with H. hainanensis. The OI and PC value of Pronephrium simplex are ranked first and they have similar ecological habits and environmental requirements to H. hainanensis. There are four species with │AC│ < 0.1: Dinochloa utilis, Cayratia japonica, Smilax lanceifolia, and Alpinia galanga. Among them, the OI and PC value of Cayratia japonica are relatively large, and their connection is more closely than the other three species. Based on AC, PCC, OI, and PC, two species are positively associated with H. hainanensis and dominant herbaceous species, seven are negatively associated, and five are completely independent.
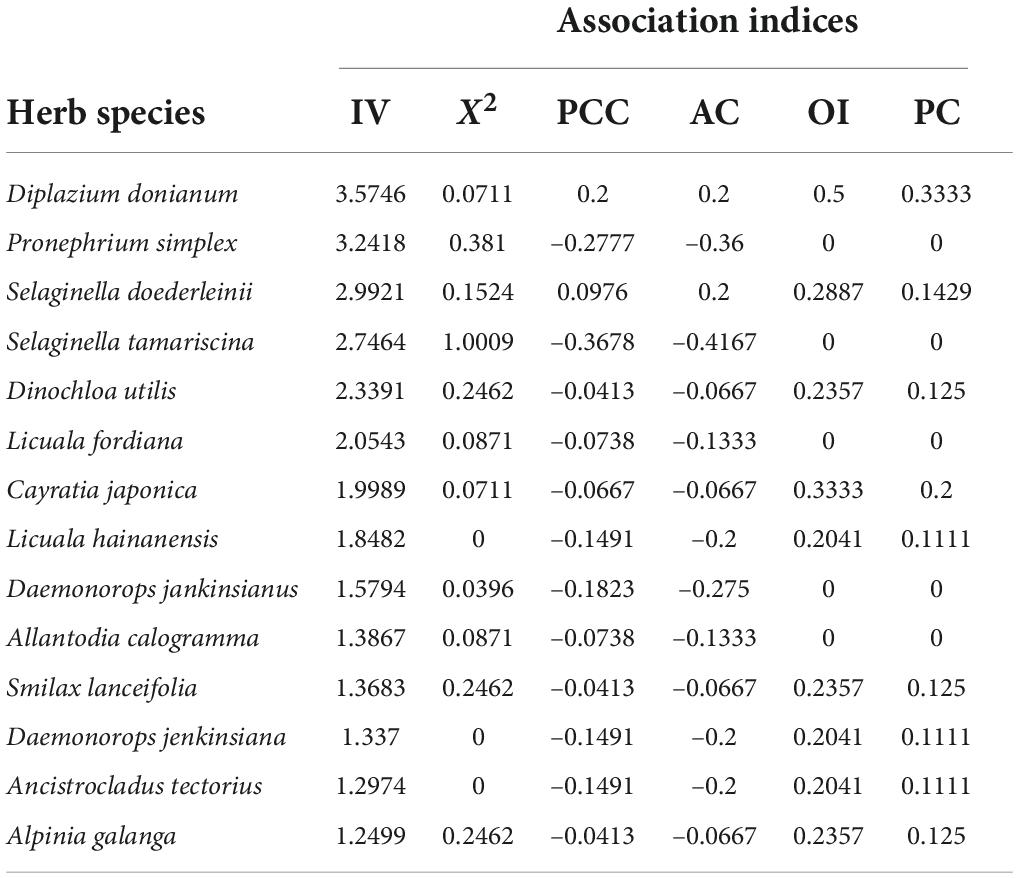
Table 5. Indices of inter-specific association between Hopea hainanensis and herb species.
The relative frequencies of all plants in the community were accumulated and plotted by using the M. Godron stability assay. The stability diagram of the community in which H. hainanensis is located was obtained (Figure 2). The results showed that the stable point of the community was 36.84/63.16, far from the stable point of 20/80, indicating that the community where H. hainanensis was unstable (Table 6). To judge the stability of each level of the community accurately, the maps were drawn according to the tree layer, the shrub layer, and the herb layer (Figure 2). With a coefficient of determination (R2 > 0.99), the coordinates of the intersection points of each layer were 44.94/55.06 for the tree layer, 41.76/58.24 for the shrub layer, and 42.69/57.31 for the herb layer. It shows the stability of the community where H. hainanensis is located, and the community, the tree layer, the shrub layer, and the herb layer are all in an unstable state. The stability of each layer is shrub layer > herb layer > tree layer.
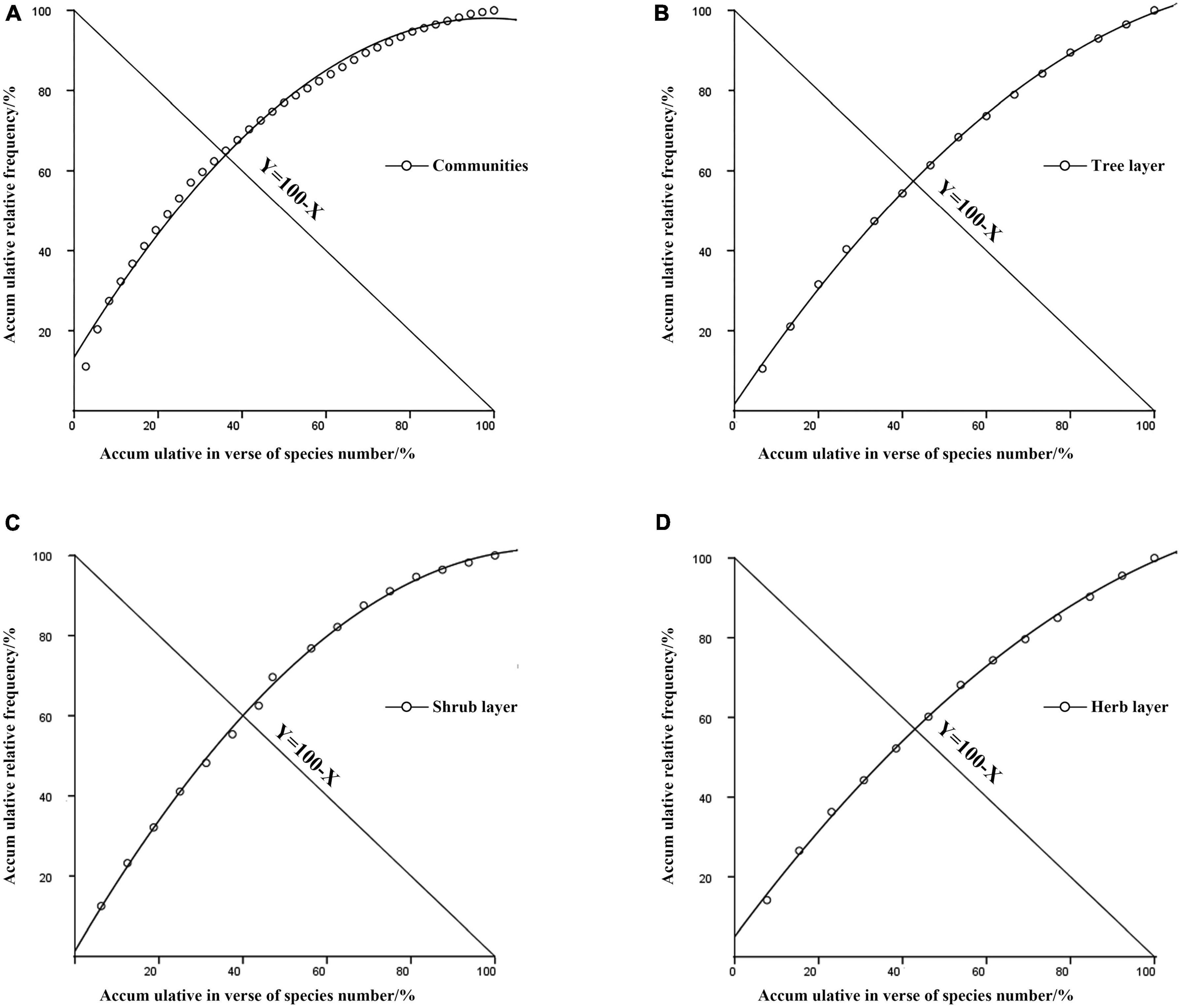
Figure 2. Community stability graph of Hopea hainanensis. (A) Communities accum ultative in verse of species number. (B) Tree layer accum ultative in verse of species number. (C) Shrub layer accum ultative in verse of species number. (D) Herb layer accum ultative in verse of species number.

Table 6. Results of community stability (M. Godron’s method).
Interspecies connectivity can reflect community stability and succession status (Haak et al., 2019). The magnitude of interspecies connectivity reflects the interactions between populations and community dynamics. The progress of community succession, the interspecific and intraspecific competition of species, community structure, and community species composition will continue to be stable. The interspecific relationship will also be positively correlated to achieve a steady equilibrium state between different populations (Hu and Ding, 2000; Wang et al., 2021). The positive and negative association between species implies the function of species in the community and affects the succession of the community. Analyzing the community’s interspecific association can help to understand better the direction of the community’s succession (Jin et al., 2013). The Chi-square test showed that the community where H. hainanensis was located showed a positive correlation (VR > 1) but did not reach a significant level [W < X20.05(N)]. The interspecific connection of the community where H. hainanensis was relatively loose, followed by the ratio of species reciprocal percentage to cumulative relative frequency was 36.84/63.16, far from the stable point of 20/80. The community where the H. hainanensis was located was unstable, and the ecology was relatively fragile. The community where H. hainanensis is located is still in the early or middle stage of succession. The community structure and function are still unstable, the community species are in dynamic regulation, and a complex and stable community with multi-species coexistence has not been formed. H. hainanensis and other species in the community exist to utilize resources competition.
The association between the H. hainanensis population and the dominant species of trees, shrubs, and herbs in the community did not reach a significant level. The correlation coefficient of interspecific association between H. hainanensis and most species was low, suggesting the vertical structure of H. hainanensis and the interspecific association of the dominant species in each community layer. The connection relationship was relatively loose and tended to be independent of each other, with great randomness. There was no direct connection between H. hainanensis and the dominant species in the community and this was similar to the research examining the associations between rare and endangered plants. It is similar to the results of the endangered species Berchemiella wilsonii, Sinojackia rehderiana, Phyllitis scolopendrium, and their associated species in their communities (Hu et al., 2005; Huang et al., 2015; Zhou et al., 2017). We argue that three possible reasons may help account for this. First, the biological characteristics of the H. hainanensis itself may lead to a loose interspecific connection loss. Besides, there is a low altitude that can be easily affected by human beings, so the community where it is located has suffered different degrees of damage for years. In addition, H. hainanensis is an indicator species of tropical rainforest, in which the species diversity is exceptionally high, and the community composition is relatively complex. This environment results in a low probability of encounters between species. Therefore, the phenomenon of weak connection, zero connection, or negative connection between H. hainanensis and other species appears, further confirming that tropical rain forests’ constituent populations are characterized by relatively independent distribution.
The association between H. hainanensis populations and herbs was more complex than that between trees and shrubs, and it was more closely associated with the dominant tree species. The connection coefficient and point correlation coefficient analysis showed six positively and negatively related species of H. hainanensis and arbor species. In the tree layer, the PCC, AC, OI index, and PC value of Schefflera heptaphylla were the highest. Although they did not reach a significant level, the connection between Schefflera heptaphylla and H. hainanensis was relatively close, and the ecological habits and demands on the environment of the two species had specific differences. The IV of Schefflera heptaphylla was 3.68%. It ranked first in the arbor layer and was in the upper layer of the community. It has a relatively strong ability to utilize resources. The population and distribution range of Schefflera heptaphylla is relatively large. The probability of encountering H. hainanensis is higher than in other species. Thirteen shrub species were positively related to H. hainanensis, accounting for 86.67% of the total number of shrub species. Still, the correlation coefficients were relatively low, and two species (Hancea hookeriana and Glochidion philippicum) were not related to H. hainanensis. The number of H. hainanensis saplings is scarce, and the interspecific relationship with other species tends to be positively correlated to promote coexistence. The low correlation coefficient of H. hainanensis also maintained a certain degree of independence in the community, reducing intraspecific competition, which was beneficial for the development of H. hainanensis saplings in the shrub layer. Two species were positively associated with herbs, seven were negatively associated, and five were completely independent. This behavior could be due to the high mortality rate of H. hainanensis seedlings. There is competition and interference between H. hainanensis seedlings and the dominant species in the herb layer, which is not conducive to their development. A larger importance value of a species typically indicates a larger niche breadth of the species and this will lead to more significant niche overlap and thereby a tighter interspecific association of the species with other species (Li et al., 2015; Chen et al., 2019).
H. hainanensis represented a major component both in the tree and the herb layer, and its importance value was in the order of herb layer (23.58) > arbor layer (3.30) > shrub layer (1.07). These results suggest that the resources in the community where H. hainanensis present are not fully utilized. It is still in an active development state, as indicated by the lack of a consistent relationship between H. hainanensis’s critical value and its tight association with other species. H. hainanensis is more dominant in the herb layer and arbor layer. The number of adults of H. hainanensis in the community was relatively small, but its diameter at breast height is significant and its height was high. The number of seedlings in the herb layer is more, and the coverage is high. It initially shows a declining trend in the wild H. hainanensis population, which was consistent with the study examining the population structure of H. hainanensis in the Bawangling Nature Reserve (Zhang et al., 2022). It has been challenging to protect and reintroduce rare and endangered plants in minimal numbers (Godefroid et al., 2011). However, such practices of introducing H. hainanensis have been successful in three experimental sites of Puwen, Xishuangbanna, and Xunwang in Yunnan. Still, compared with plantations that typically have simple community structures, natural forests can better support biodiversity conservation and achieve ecosystem services such as surface carbon storage, soil conservation, and water conservation (Hua et al., 2022). Therefore, the restoration of wild H. hainanensis populations is essential. The interspecific relationship between the H. hainanensis population and herbs is more complex than trees and shrubs. The overall positive connection ratio between the H. hainanensis and other dominant species is lower, and the negative connection between the H. hainanensis seedlings in the herb layer is higher than Shrub layer and tree layer. The competition for resources among H. hainanensis and other species is fierce, and the mortality of H. hainanensis seedlings is high. Zhang et al. (2019) conducted a 6-month shading experiment on semi-annual H. hainanensis seedlings to study the effect of light intensity on H. hainanensis seedlings and found that light is an important environmental factor affecting H. hainanensis seedlings. H. hainanensis seedlings lower 7–8 months grew better under certain shading conditions, and the seedlings after 7–8 months are suitable for growing under insufficient light. However, many factors could cause the high mortality rate of seedlings. Further studies examining the causes of the high mortality rate of seedlings on H. hainanensis are warranted. The problem that seedlings cannot be transit to saplings should be solved and we should construct strategies for restoring wild H. hainanensis populations.
The community where H. hainanensis is found is unstable, and the ecology is also relatively fragile. The community where H. hainanensis is located is still in the early or middle stage of succession. The community structure and function are still unstable, the community species are in a dynamic adjustment, a complex and stable community with multi-species coexistence has not been formed, and there are still competitions for resources between H. hainanensis and other species in the community to exist. The correlation between H. hainanensis and the dominant species in the community is not significant. The connection between the H. hainanensis population and herbs is more complex than that of trees and shrubs, and it is more closely connected with the dominant species of trees. To conserve the wild H. hainanensis population, future studies should conduct further research on the high mortality of seedlings.
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
LZ, H-LZ, MN, and YC: conceptualization. YC and LZ: methodology. LZ: software, data curation, formal analysis, and investigation. LZ and H-LZ: validation. LZ, H-LZ, MN, QZ, and XS: writing—original draft preparation and writing—review and editing. YC: project administration. All authors have read and agreed to the published version of the manuscript.
This study was supported by the National Science Foundation of China (31760119) and the National Science Foundation of Hainan Province (320MS038).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer SB declared a shared affiliation with the authors H-LZ, MN, and QZ to the handling editor at the time of review.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Álvarez -Yépiz, J. C., Búrquez, A., Martínez-Yrízar, A., and Dovciak, M. (2019). A trait-based approach to the conservation of threatened plant species. Oryx 53, 429–435. doi: 10.1017/S003060531800087X
Chen, H. X., Huang, C. T., He, F., Zheng, W., and Feng, J. P. (2015). Review on research progress of Hopea hainanensis. Trop. Fores. 43, 4–6.
Chen, L., Xin, J. N., Su, Y., Li, Y. F., Song, N. P., Wang, L., et al. (2019). Effects of heterogeneous habitats on community composition and niche characteristics of different plant populations in the desert steppe of China. Acta Ecol. Sin. 39, 6187–6205. doi: 10.5846/stxb201810182255
Chen, Y. K., Yang, Y., Jin, Y. H., Yao, J., Wang, Y., and Zhang, Y. (2021). Genomic characteristics and phylogenic analysis of chloroplast of the endangered plant Hopea hainanensis. Genom. Appl. Biol. 40, 365–374.
Di Musciano, M., Di Cecco, V., Bartolucci, F., Conti, F., Frattaroli, A. R., and Di Martino, L. (2020). Dispersal ability of threatened species affects future distributions. Plant Ecol. 221, 265–281. doi: 10.1007/s11258-020-01009-0
Godefroid, S., Piazza, C., Rossi, G., Buord, S., Stevens, A. D., Aguraiuja, R., et al. (2011). How successful are plant species reintroductions? Biol. Conservat. 144, 672–682. doi: 10.1016/j.biocon.2010.10.003
Grime, J. P. (1998). Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 86, 902–910. doi: 10.1046/j.1365-2745.1998.00306.x
Guo, Z., Zhang, Y., Deegen, P., and Uibrig, H. (2006). Economic analyses of rubber and tea plantations and rubber-tea intercropping in hainan, china. Agroforest. Syst. 66, 117–127. doi: 10.1007/s10457-005-4676-2
Haak, C., Hui, F. K. C., and Cowles, G. W. (2019). Positive inter- specific associations consistent with social information use shape juvenile fish assemblages. Ecology 101:e02920. doi: 10.1002/ecy.2920
Hu, L. L., Jiang, M. X., Dang, H. S., and Huang, H. D. (2005). Community studies on the status of the endangered plant, berchemiella wilsonii var. pubipetiolata, using interspecific association analysis. Acta Phytoecol. Sin. 29, 258–265. doi: 10.17521/cjpe.2005.0033
Hu, Y. J., and Ding, X. Q. (2000). A study on the plant species diversity of tropical natural forest in bawangling, hainan Island. Bio. Sci. 8, 370–377. doi: 10.17520/biods.2000052
Hua, F., Bruijnzeel, L. A., Meli, P., Martin, P. A., Zhang, J., Nakagawa, S., et al. (2022). The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 2022:4649. doi: 10.1126/science.abl4649
Huang, X. T., Yin, H., Huang, Q. J., Cui, K. F., Zhao, Y., Jin, H., et al. (2015). Characteristics of population structure and community species diversity of an extremely small population of protected Phyllitis scolopendrium. Acta Ecol. Sin. 38, 2481–2492. doi: 10.5846/stxb201704190706
Jin, J. Y., Qin, W. G., Tan, W. N., Luo, L. J., and Qin, G. L. (2013). Interspecific association among dominant populations in endangered plant Kmeria septentrionalis communities. J. West China Fores. Sci. 42, 86–94.
Li, G. Y., Chen, F. F., and Yang, Z. L. (2015). Studies on seedling cultural techniques of Hopea hainanensis. Trop. Fores. 43, 7–9.
Lohbeck, M., Lebrija-Trejos, E., Martínez-Ramos, M., Meave, J. A., Poorter, L., and Bongers, F. (2015). Functional trait strategies of trees in dry and wet tropical forests are similar but differ in their consequences for succession. PLoS One 10:e0123741. doi: 10.1371/journal.pone.0123741
Ly, V., Nanthavong, K., Pooma, R., Hoang, V. S., Khou, E., and Newman, M. F. (2018). Hopea hainanensis the IUCN red list of threatened species2018: e.T32357A2816074. doi: 10.2305/IUCN.UK.2018-1.RLTS.T32357A2816074.en
Mehmood, S., Ahmed, W., Alatalo, J. M., Mahmood, M., Imtiaz, M., Ditta, A., et al. (2022). Herbal plants- and rice straw-derived biochars reduced metal mobilization in fishpond sediments and improved their potential as fertilizers. Sci. Total Environ. 826:154043. doi: 10.1016/j.scitotenv.2022.154043
Mehmood, S., Saeed, D. A., Rizwan, M., Khan, M. N., Aziz, O., Bashir, S., et al. (2018). Impact of different amendments on biochemical responses of sesame (Sesamum indicum L.) plants grown in lead-cadmium contaminated soil. Plant Physiol. Biochem. 132, 345–355. doi: 10.1016/j.plaphy.2018.09.019
Miyadokoro, T., Nishimura, N., and Yamamoto, S. (2003). Population structure and spatial patterns of major trees in a subalpine old-growth coniferous forest, central Japan. Forest. Ecol. Manage. 182, 259–272. doi: 10.1016/S0378-1127(03)00045-8
Powell, J. R., and Rillig, M. C. (2018). Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 220, 1059–1075.
Satiraphan, M. (2012). Lupane triterpenes from the leaves of the tropical rain forest tree hopea odorata roxb.and their cytotoxic activities, supplementary information. Biochem. Syst. Ecol. 44, 407–412. doi: 10.1016/j.bse.2012.05.014
Schnitzler, A., and Closset, D. (2003). Forest dynamics in unexploited birch (Betula pendula) stands in the vosges (France): structure, architecture and light patterns. Forest. Ecol. Manage. 183, 0–220. doi: 10.1016/S0378-1127(03)00118-X
Song, Y. B., Shen-Tu, X. L., and Dong, M. (2020). Intraspecific variation of samara dispersal traits in the endangered tropical tree Hopea hainanensis (Dipterocarpaceae). Front. Plant Sci. 11:599764. doi: 10.3389/fpls.2020.599764
Sonkoly, J., Kelemen, A., Valkó, O., Deák, B., Kiss, R., Tóth, K., et al. (2019). Both mass ratio effects and community diversity drive biomass production in a grassland experiment. Sci. Rep. 9, 1–10. doi: 10.1038/s41598-018-37190-6
Turner, S. R., Lewandrowski, W., Elliott, C. P., Merino-Martín, L., Miller, B. P., Stevens, J. C., et al. (2017). Seed ecology informs restoration approaches for threatened species in water-limited environments: A case study on the short-range banded ironstone endemic ricinocarpos brevis (Euphorbiaceae). Austr. J. Bot. 65, 661–677. doi: 10.1071/BT17155
Wang, C., Ma, X., and Tang, L. (2021). Isolation and characterization of twelve polymorphic microsatellite markers in the endangered Hopea hainanensis (Dipterocarpaceae) (2020). Ecol. Evolu. 11, 4–10. doi: 10.1002/ece3.7077
Wang, F. G., Zhang, R. J., Xing, F. W., Ng, S. C., Ye, Y. S., and Chen, H. Q. (2007). Rare and endangered plants and conservation in yinggeling nature reserve, hainan. J. Wuhan Bot. Res. 25, 303–309.
Zhang, H. L., Cubino, J. P., Nizamani, M. M., Harris, A. J., Cheng, X. L., Da, L., et al. (2022). Wealth and land use drive the distribution of urban green space in the tropical coastal city of Haikou, China. Urban Forest. Urban Green. 2022:127554. doi: 10.1016/j.ufug.2022.127554
Zhang, L., Yang, X. B., Nong, S. Q., Li, D. H., Li, Y. L., and Song, J. Y. (2019). Comparative study on population development characteristics of Hopea hainanensis base in two different protection modes. Acta Ecol. Sin. 39, 3740–3748. doi: 10.5846/stxb201811012359
Keywords: Hopea hainanensis, population associativity, associated species, community stability, inter-specific association
Citation: Zhang L, Zhang H-L, Chen Y, Nizamani MM, Zhou Q and Su X (2022) Analyses of community stability and inter-specific associations between a plant species with extremely small populations (Hopea hainanensis) and its associated species. Front. Ecol. Evol. 10:922829. doi: 10.3389/fevo.2022.922829
Received: 18 April 2022; Accepted: 15 August 2022;
Published: 20 September 2022.
Edited by:
Salman Qureshi, Humboldt University of Berlin, GermanyReviewed by:
Saraj Bahadur, Hainan University, ChinaCopyright © 2022 Zhang, Zhang, Chen, Nizamani, Zhou and Su. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yukai Chen, Y2hlbnl1a2FpQGhhaW5udS5lZHUuY24=
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.