
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 29 July 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.918773
This article is part of the Research TopicSex and Gender Effects on Power, Status, Dominance, and Leadership – An Interdisciplinary Look at Human and Other Mammalian SocietiesView all 22 articles
 Peter M. Kappeler1,2*†
Peter M. Kappeler1,2*† Elise Huchard3*†
Elise Huchard3*† Alice Baniel4
Alice Baniel4 Charlotte Canteloup5,6
Charlotte Canteloup5,6 Marie J. E. Charpentier3
Marie J. E. Charpentier3 Leveda Cheng7,8
Leveda Cheng7,8 Eve Davidian9
Eve Davidian9 Julie Duboscq10,11
Julie Duboscq10,11 Claudia Fichtel1
Claudia Fichtel1 Charlotte K. Hemelrijk12
Charlotte K. Hemelrijk12 Oliver P. Höner9
Oliver P. Höner9 Lee Koren13Jérôme Micheletta14Lea Prox1,11
Lee Koren13Jérôme Micheletta14Lea Prox1,11 Tommaso Saccà12
Tommaso Saccà12 Lauren Seex12
Lauren Seex12 Nikolaos Smit3Martin Surbeck7,8
Nikolaos Smit3Martin Surbeck7,8 Erica van de Waal5,6Cédric Girard-Buttoz15,16
Erica van de Waal5,6Cédric Girard-Buttoz15,16The causes and consequences of being in a particular dominance position have been illuminated in various animal species, and new methods to assess dominance relationships and to describe the structure of dominance hierarchies have been developed in recent years. Most research has focused on same-sex relationships, however, so that intersexual dominance relationships and hierarchies including both sexes have remained much less studied. In particular, different methods continue to be employed to rank males and females along a dominance hierarchy, and sex biases in dominance are still widely regarded as simple byproducts of sexual size dimorphism. However, males and females regularly compete over similar resources when living in the same group, and sexual conflict takes a variety of forms across societies. These processes affect the fitness of both sexes, and are mitigated by intersexual hierarchies. In this study, we draw on data from free-ranging populations of nine species of mammals that vary in the degree to which members of one sex dominate members of the other sex to explore the consequences of using different criteria and procedures for describing intra- and intersexual dominance relationships in these societies. Our analyses confirmed a continuum in patterns of intersexual dominance, from strictly male-dominated species to strictly female-dominated species. All indices of the degree of female dominance were well correlated with each other. The rank order among same-sex individuals was highly correlated between the intra- and intersexual hierarchies, and such correlation was not affected by the degree of female dominance. The relative prevalence of aggression and submission was sensitive to variation in the degree of female dominance across species, with more submissive signals and fewer aggressive acts being used in societies where female dominance prevails. Thus, this study provides important insights and key methodological tools to study intersexual dominance relationships in mammals.
When the Norwegian zoologist Schjelderup-Ebbe (1922) published his dissertation on the social psychology of chickens a century ago, he was the first to report that the directionality and consistency with which hen peck at each other during feeding and resting yields a stable arrangement among individuals that he called a pecking order. As in other species, dominance relationships among chicken are established and maintained through agonistic interactions whereby one individual may exhibit aggressive behavior, whereas the partner either displays submissive behavior or avoids confrontation with the aggressor altogether (Rowell, 1974). Other, structurally more complex types of hierarchies have since been described for various animal societies in which individuals recognize individual conspecifics and remember the outcome of previous agonistic interactions with co-residents (Reddon et al., 2021). Whereas solitary or unfamiliar animals can also establish a dominance relationship without prior interaction, for example, based on mutual assessment of body size, ornaments or other intrinsic cues of fighting ability, we are here primarily interested in this aspect of the social structure of species that live permanently in groups that contain both sexes because social dominance represents a prominent feature of many animal societies.
Dominance relationships have various determinants that are not mutually exclusive and can feed back upon each other in determining the outcome of an agonistic interaction (Dehnen et al., 2022). First, in some species, a given dyadic dominance relationship reflects differential social support received by each member of the dyad (Clutton-Brock and Huchard, 2013a,b), either in the form of parental (Holekamp and Smale, 1991) or non-parental support (Schülke et al., 2010). Second, memories of previous interactions with known individuals can promote a learning effect that leads individuals to exhibit submissive behavior toward certain other conspecifics (Guhl, 1968). The effect of this dyadic interaction-outcome history is further reinforced by the winner-loser effect (Chase et al., 1994), according to which winning increases the probability to be victorious again and losing makes it more likely that the victim loses again in the next fight against any other opponent (Rutte et al., 2006; Franz et al., 2015). Finally, dominance relationships are often based upon an asymmetry in agonistic power grounded on intrinsic attributes, such as physical superiority, fighting ability, motivation, or leverage that are often age-based (Hand, 1986; Lewis, 2002; Jonart et al., 2007; Dunham, 2008; Clutton-Brock and Huchard, 2013a,b; Dehnen et al., 2022; Tibbetts et al., 2022). In species with pronounced sexual size dimorphism and/or elaborate weapons, these intrinsic attributes are linked to sex, with members of the larger and/or better armed sex often using their attributes to establish and stabilize dominance relationships with members of the opposite sex (Kappeler, 1993; Lewis, 2018), just as size and strength are important determinants of within-sex dominance. However, recent evidence highlighted that sex differences in physical attributes may often fail to predict who is dominant in an intersexual relationship, as in bonobos (Pan paniscus) and spotted hyenas (Crocuta crocuta) (Surbeck and Hohmann, 2013; Vullioud et al., 2019).
In species in which groups consist of permanently associated males and females, past research has often focused on intrasexual dominance relations because they are thought to mediate access to fitness-limiting factors, such as mates and resources. As a result, most previous studies of dominance hierarchies have been framed in separate theoretical frameworks. Studies of female dominance relations focused on the ecological drivers of interspecific variation (Sterck et al., 1997; Clutton-Brock and Janson, 2012). Studies of male dominance relationships, in contrast, focused on their functional outcomes for mating access and reproductive skew (Cowlishaw and Dunbar, 1991; Alberts, 2012). However, such a binary approach overlooks the potential importance of intersexual agonism for the fitness of both sexes. First, when males and females live in the same group, competition can occur both within and between the sexes for access to various resources, including – but not necessarily limited to – the feeding context (Valé et al., 2020; Koenig et al., 2022). Second, sexual conflict is nearly ubiquitous, and takes various forms that all generate intersexual strife (Chapman et al., 2003; Davidian et al., 2022), which affects the fitness of both sexes and may be mitigated by intersexual hierarchies. Thus, while it may make sense to use separate intrasexual hierarchies for species with pronounced sexual size dimorphism, with sex differences in the steepness, linearity and stability of the within-sex hierarchies, and in which the mechanisms of rank acquisition differ between the sexes, this approach may not be appropriate for other species.
A few studies indicate that sex is one key variable for scrutinizing dominance hierarchies in more detail. First, in some species individuals are distributed within a group’s hierarchy as a function of their sex, with either all or most males outranking all females or – more rarely – vice versa (Kappeler, 1993; Smuts and Smuts, 1993; Stevens et al., 2007; Surbeck and Hohmann, 2013; Izar et al., 2021). Such sex clusters likely reflect sex differences in morphology, physiology or life-history that underlie agonistic power and therefore offer an opportunity to identify sex-specific determinants of dominance. Second, males and females compete for different resources, and may therefore employ different strategies to establish and maintain their dominance rank among their same-sex peers, with unknown consequences for the establishment of intersexual hierarchies. Notably, the steepness, linearity and stability of the male and female hierarchies often differ. In chimpanzees and gorillas, for example, males have conspicuous and relatively despotic dominance hierarchies, whereas dominance relations among females are more elusive and are based on age-based queuing conventions (Stevens et al., 2007; Foerster et al., 2016). In Barbary macaques, the opposite pattern has been found, with males being more egalitarian and females more despotic in dominance style (Preuschoft et al., 1998; Hemelrijk and Gygax, 2004). Contrasting hierarchies across dyads of same- and opposite-sex members can therefore reveal informative subtleties on sex differences in social competition beyond species differences. Finally, the mechanisms of rank acquisition may differ between the sexes, with males typically relying on intrinsic attributes to compete for high rank, whereas female ranks tend to depend more often on social support (Holekamp and Smale, 1991; Clutton-Brock and Huchard, 2013b).
Nonetheless, dominance relationships between the sexes have not received the same theoretical and empirical attention as same-sex dominance relationships. Because male-biased sexual dimorphism is widespread among mammals (Lindenfors et al., 2007) and represents a default expectation based on conventional sex roles, the ability of males of these species to dominate females was typically considered an unavoidable side-effect of physical superiority and greater aggressiveness, rather than as an adaptive trait per se (Kappeler, 1993; Smuts and Smuts, 1993; Lewis, 2018). In contrast, the rare cases of female dominance often required special explanation and generated several hypotheses that typically invoke taxon-specific factors to explain the evolution of this sex role “reversal” (Jolly, 1984; Richard and Dewar, 1991; van Schaik and Kappeler, 1996; Wright, 1999; Dunham, 2008; Kappeler and Fichtel, 2015). These few taxa or studies were not deemed of enough general interest, however, to prompt a general synthesis of intersexual relationships.
The dichotomous classification of species as either male- or female-dominated has been challenged by more recent studies indicating that these patterns only represent the endpoints of a continuum (Hemelrijk et al., 2008, 2020; Davidian et al., 2022; Kappeler et al., 2022). It is now more widely appreciated that there are taxa where members of one sex only win a proportion of all agonistic interactions with the members of the other sex or where they dominate only some, but not all, opposite-sex individuals (Surbeck and Hohmann, 2013, Young et al., 2017; Vullioud et al., 2019; Hemelrijk et al., 2020). In addition, different methods continue to be employed to rank males and females along a dominance hierarchy (Pereira and Kappeler, 1997; Hemelrijk et al., 2008), potentially obscuring interesting biological patterns. In order to analyze this variation in a comparative fashion, comparable data on intersexual dominance relationships are required. However, existing studies have used various methods for recording details of agonistic interactions, for inferring dominance relationships, and for determining dominance hierarchies, and different species use different types and variable proportions of acts and signals to establish dominance relationships, thereby hampering comparative studies on this topic. We are therefore only beginning to explore whether different methods to analyze agonistic interactions are equivalent or whether some methods should be preferred or discouraged (Sánchez-Tójar et al., 2018; Vilette et al., 2020).
The general aim of this study is, therefore, to systematically determine the consequences of using different behavioral data, criteria and methods for inferring patterns of intersexual dominance relationships. Specifically, we aim to make recommendations about standardized methods of data collection and analyses for future studies of intersexual dominance relationships that go beyond taxon-specific idiosyncrasies. We are also interested in possible similarities across species in the nature and pattern of intersexual dominance relationships. For instance, it remains unknown whether an individual’s position in the intrasexual hierarchy relates to its position in the intersexual hierarchy and, hence, whether intersexual dominance patterns are an emergent property of intrasexual ones.
To this end, we have collated datasets from nine mammalian species which were chosen because they differ in the degree to which members of one sex dominate the members of the other sex, ranging from complete male dominance to complete female dominance. Our aim was to explore the consequences of using different criteria and procedures for describing intra- and intersexual dominance relations in this sample of animal societies and not a comparative study across a wide range of taxa and social systems. Most of these datasets are based on observations spanning at least a full year for at least two different groups, thereby accounting for seasonal variation and group idiosyncrasies. With this dataset, we (1) calculated and compared different indices of the degree of intersexual dominance in a group, (2) examined whether an individual’s rank in the same-sex hierarchy predicts its position in the intersexual hierarchy, i.e., whether high-ranking females in the female hierarchy are more likely to dominate males, for example, and if the degree of correlation between intra- and intersexual hierarchy changes along the intersexual dominance spectrum, and (3) explored whether within- and between-sex conflicts differ in nature and intensity in terms of their reliance on aggressive and submissive behavior along the intersexual dominance spectrum. Taken together, our study provides a first set of recommendations and predictions for future studies aimed at explaining interspecific variation in intersexual dominance.
Data on agonistic interactions were collected from nine different mammalian species: spotted hyenas (Crocuta crocuta), rock hyraxes (Procavia capensis) and seven primates including two lemurs: Verreaux’s sifakas (Propithecus verreauxi) and redfronted lemurs (Eulemur rufifrons), four Old World monkeys: chacma baboons (Papio ursinus), crested macaques (Macaca nigra), mandrills (Mandrillus sphinx), and vervet monkeys (Chlorocebus pygerythrus) and one ape: bonobos (Pan paniscus), as part of nine long-term individual-based field studies, including the Ngorongoro Hyena Project, Ein Gedi Rock Hyrax Sociality Project, Kirindy Forest Lemur Project, Tsaobis Baboon Project, Macaca Nigra Project, Mandrillus Project, Inkawu Vervet Project and the Kokolopori Bonobo Research Project. All studies were approved and authorized by the respective national authorities. The localization and ecology of each field site, as well as the demographic, life-history and behavioral monitoring of each study population are described in the Supplementary Materials, alongside the species-specific agonistic behavioral repertoire.
Data on agonistic interactions were recorded as detailed in the Supplementary Materials and grouped by year for each species and study group, yielding 24 group-years of data (range 1–4 per species). An agonistic interaction was defined by the expression of at least one species-specific agonistic behavior (see Hausfater, 1975). Following Deag (1977), agonistic behavioral elements were functionally characterized as either aggressive or submissive (see also Walters, 1980). Structurally, we distinguished between acts, which involve physical contact or locomotion, such as lunging or fleeing, and visual or vocal signals, such as non-physical threats or grimacing (see Pereira and Kappeler, 1997). We considered only dyadic interactions between adult individuals.
Group size ranged between 4 and 72 adult individuals, including 1–62 females and 2–26 males (Table 1). For each dataset, the corresponding co-author(s) entered the data into a standardized template. In this template, we entered only dyadic interactions and each agonistic interaction constituted a row. For each agonistic interaction, we entered the following information: (1) the identity of the two participants, (2) the sex of each participant, (3) whether the conflict was decided (i.e., only one opponent exhibited submissive behavior) or not, (4) the identity of the winner of a decided conflict, and (5) whether one or the other or both individuals displayed an aggressive act, an aggressive signal, a submissive act and/or a submissive signal (see Supplementary Figure 1).
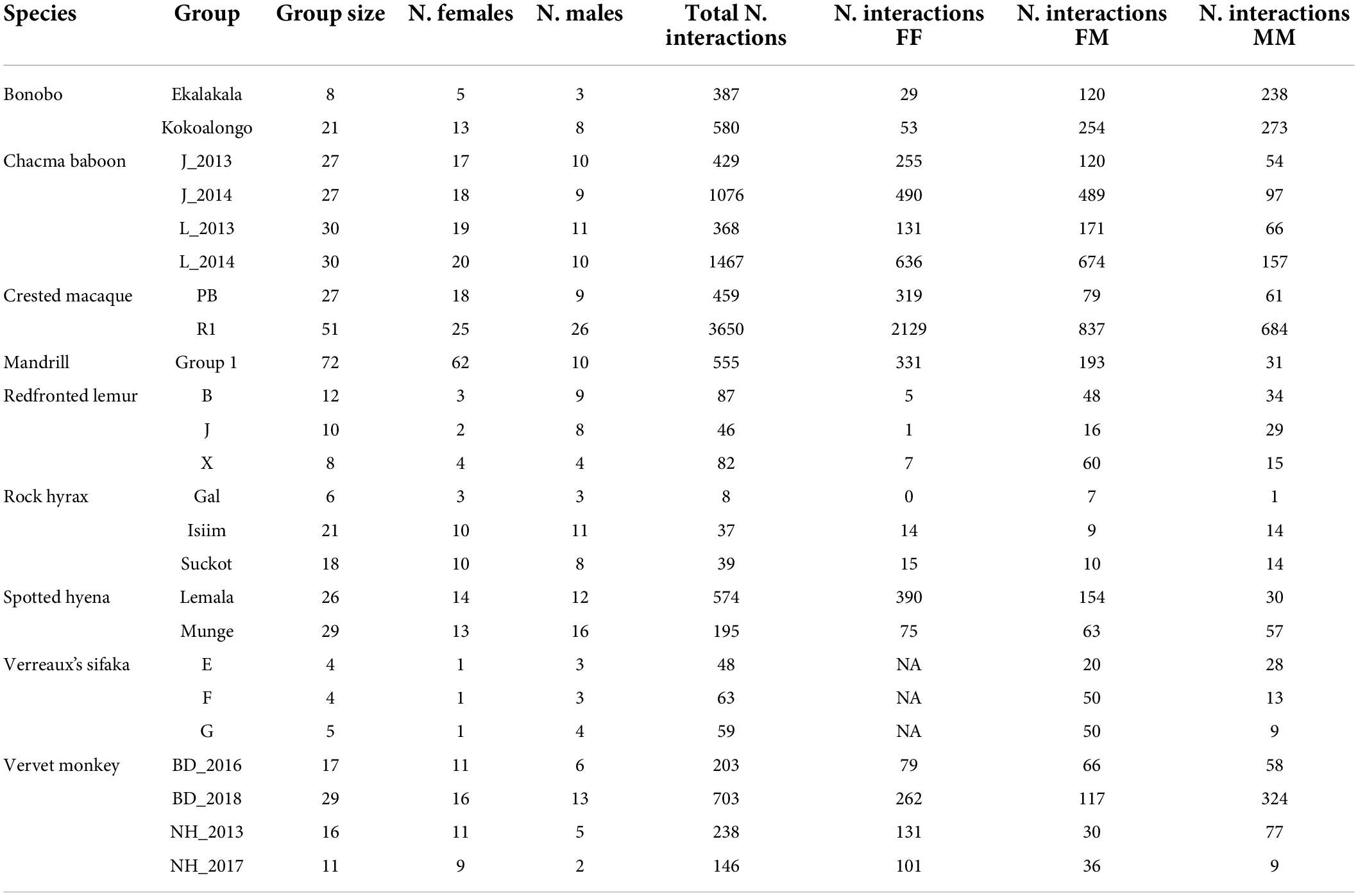
Table 1. Summary of the data collated for this study.
The final dataset comprised a total of 11,499 agonistic interactions, including 5,453 interactions between females, 2,373 interactions between males, and 3,673 interactions between males and females. The majority of all agonistic interactions (10,005 or 87%) were decided. The total number of interactions per group ranged from 8 to 3,650 (mean ± SD = 479.1 ± 764.9, Table 1).
We computed hierarchies based on David’s scores and I&SI, using the function “DS” and “ISI” of the R package “EloRating” (Neumann et al., 2011). David’s score is calculated based on power relationships between individuals, and the score of each individual is calculated based on the proportion of conflicts won and lost with other individuals in the group. For this metric, the number of conflicts is of importance because David’s score does not simply provide an ordinal rank but yields power differences between individuals. However, for the purpose of our study, we only used David’s score to establish individual rank order (with the highest score receiving rank 1, the second highest rank 2, etc.). One drawback of David’s score is that it is relatively sensitive to the percentage of missing dyads in the matrix and to differences in observation time between individuals (Neumann et al., 2011). I&SI, on the other hand, is based on multiple iterations of randomization of the interaction matrix, until reaching the most parsimonious rank order. The advantage of this method is that it is designed to provide ordinal rank order and is less sensitive to missing data. One drawback, however, is that it may provide several equally likely rank order solutions with large datasets and in particular when interactions are lacking for a large proportion of dyads, potentially complicating the interpretation and limiting replicability. In fact, re-running the algorithm several times on the same matrices does not provide exactly the same result.
For each hierarchy, we then calculated the percentage of males dominated by each female (“the degree of female dominance“) and averaged this percentage across all females to obtain two indices.
Index 1: average percentage of males in a group dominated by each female (Hemelrijk et al., 2008, 2020) using hierarchies based on male-female interactions only. This index was calculated twice, once using David’s score (de Vries, 1998) to calculate the hierarchy (Index 1a) and once using hierarchies based on I&SI (de Vries, 1998, Index 1b).
Index 2: the female dominance index, FDI (Hemelrijk et al., 2020), calculated as the average percentage of males in a group dominated by each female using hierarchies based on all agonistic interactions (i.e., inter- and intrasexual conflicts). We also computed this index twice, using hierarchies based on either David’s score (Index 2a) or I&SI (Index 2b).
To evaluate whether alternative estimates of the degree of female dominance or methodological differences in calculation may alter assessment of intersexual dominance relationships, we calculated three additional indices as follows:
Index 3: percentage of intersexual conflicts won by females, calculated for each female and averaged across all females.
Index 4: percentage of intersexual conflicts won by females in each intersexual dyad averaged across all intersex or something of that nature dyads.
Index 5: total percentage of intersexual conflicts won by females overall.
We then compared the correspondence among these indices by calculating pair-wise Spearman correlation coefficients for each species.
Next, we examined whether the assignment of individual rank is sensitive to the type of data used to calculate dominance hierarchies. We constructed three separate hierarchies: a female-only hierarchy based on female-female interactions only, a male-only hierarchy based on male-male interactions only, and an intersexual hierarchy based on all decided agonistic interactions. We constructed these hierarchies using David’s scores since they allow for reproducibility of the analyses (i.e., the same hierarchy is obtained from the same interaction matrix each time, which is not the case with I&SI method since it is based on matrix randomization). We then extracted the ordinal dominance rank of each female (i.e., between 1 and N, N being the number of females in the group) among all other females in the intersexual hierarchy. We then calculated the Spearman rank correlation coefficient between female ordinal ranks from the intrasexual versus intersexual hierarchies. We repeated this process for males, yielding two correlation coefficients per group-year: one for female-only hierarchies and one for male-only hierarchies.
For all three (female-only, male-only, and intersexual) hierarchies of each group, we compiled an index of triangular transitivity, using the function “transitivity” in the R package “EloRating” (Neumann et al., 2011), which provides an index of hierarchical linearity based on the proportion of triads in the hierarchy that have transitive dominance relationships (Shizuka and McDonald, 2012). Indices below 0.75 indicate that hierarchies are less transitive than random, and an index of 1 reflects a completely transitive hierarchy. We resorted to using triangular transitivity since it is less sensitive to missing data than alternative measures of linearity (h and h′; Shizuka and McDonald, 2012; Neumann et al., 2018).
We used two separate generalized linear mixed models (GLMMs) to assess whether the degree of female dominance in a group is sensitive to differences in individual rank as a result of using either intra- or intersexual hierarchies for both, females (Model 1a) and males (Model 1b). In Model 1a, each correlation coefficient of the females’ ranks between the intra and the inter-sexual hierarchy for each group-year constituted a data point; the same applies to correlation coefficients for males in Model 1b. We used separate GLMMs with beta error structure because the response in each model was bound between 0 and 1. We transformed the response using the following formula, recommended for models using a beta error distribution because it transforms zeros (which cannot be handled by beta models) into very small non-zero values (Smithson and Verkuilen, 2006):
where x′ is the transformed value of x (here the correlation coefficient) and N the sample size.
In each model we used the degree of female dominance in the group (as expressed by Index 1a) as test predictor. Note that all five indices of intersexual dominance were found to be highly correlated (see section “Results”). We added as control predictor the percentage of male-female dyads that did not interact since this can influence David’s scores (see above) and, therefore, the difference in ranks in the intra- versus intersexual dominance hierarchy. We could not include other control predictors, such as group size or adult sex ratio (e.g., Hemelrijk et al., 2008; Kappeler, 2017), because of our small sample size (only 21 data points in Models 1a and 24 data points in Model 1b) to avoid overparameterization. In addition to the fixed effects we added species as a random effect to account for the non-independent repeated measures on several groups of the same species.
Finally, we assessed whether the degree of female dominance in a group covaries with variation in the form of agonistic interactions within and between the sexes. More specifically, we used four GLMMs to test whether the degree of female dominance influenced the likelihood of at least one of the two opponents exhibiting an aggressive act (Model 2a), an aggressive signal (Model 2b), a submissive act (Model 2c), or a submissive signal (Model 2d) in an agonistic interaction. For each model, we counted both decided and undecided agonistic interactions in which it was clear whether one or both individuals exhibited an agonistic act or signal (N = 11492 interactions). We used GLMMs with binomial error structure to model whether an act or signal was produced (Y/N) during each agonistic interaction. As test predictors, we included the degree of female dominance as quantified by index 1a, the dyad type (FF, MM, FM) and their interaction. We fitted this interaction term to test whether the effect of the degree of female dominance on the likelihood to engage in certain acts or signals differed across the three dyad types. Group size and adult sex ratio were included as control predictors in each model because they can influence social dynamics (Hemelrijk et al., 2008; Kappeler, 2017). To account for the non-independence of repeated observations involving the same individuals, dyads and species, we added the four following random effects in each model: species, identity of the actor in the interaction, identity of the recipient and identity of the dyad. For Model 2d, the incorporation of group size as a control variable created some instability in the model preventing us from obtaining meaningful estimates for the effect of the test predictors. We therefore reran the model without group size as a variable and present the results of this second model.
While the behaviors coded as submissive acts and signals and aggressive signals were broadly similar across all the study species (see Supplementary Materials), the behaviors coded as aggressive acts differed for some datasets. In particular, displacement was considered an aggressive act in all datasets except two, the crested macaque and spotted hyena datasets. In the latter, displacement was only considered an aggressive act if the approaching individual expressed an aggressive signal. This divergence may reflect meaningful differences in the context and function of displacement behavior and variation in aggressiveness. To account for this different coding and to assess whether it had any impact on the conclusions drawn, we re-ran Model 2a (Model 2a bis) using a reduced dataset excluding crested macaque and spotted hyena datasets.
We ran all statistical models in R 4.1.2 (R Core Team, 2021), using the function “glmmTMB” from the package glmmTMB (Brooks et al., 2017) for Models 1a and 1b, and the function “glmer” from the package lme4 (Bates et al., 2011) for Models 2a–2d and 2a bis. For mixed-effects models, we included the maximum random slope structure between each fixed predictor (test and control) and each random effect (Baayen et al., 2008; Barr et al., 2013). In each model, we tested for the overall significance of the test predictors by comparing the full model to a reduced model comprising all control predictors, all the random effects and random slopes, but without any test predictor, using a likelihood ratio test (LRT, Dobson, 2002). We then assessed the significance of each predictor using a LRT between the full model and a reduced model comprising all predictors except the one to evaluate. We repeated this process across all variables using the drop1 function. For each model, we tested for collinearity issues between our predictor variables using the function vif from the package “car” (Fox and Weisberg, 2011). Collinearity was not an issue (all vifs < 3). We also assessed model stability removing one level of each random effect at a time and recalculating the estimates of the different predictors that revealed no stability issue (except for model 2d, see above). Finally, we tested for overdispersion in Models 1a and 1b, which was not an issue (all dispersion parameters < 1.08). For Models 2a-d and Model 2a bis, we calculated the marginal R2 (i.e., the variance explained by the fixed effects) and the conditional R2 (i.e., the variance explained by the entire model including both fixed and random effects) using the function r.squaredGLMM of the package “MuMin” (Barton, 2020). For Models 1a and 1b we could not compile the R2 due to negative model’s distribution-specific variance.
The interaction matrices compiled for this study were highly heterogeneous in terms of missing dyads, from being almost full, with most dyads interacting with each other, as in the Ekalakala bonobo group, to being largely empty, with interactions absent for > 85% of dyads, as in the mandrill or rock hyrax groups (Table 2 and Supplementary Tables 1–3). Most inter- and intrasexual hierarchies were highly transitive (transitivity index ≥ 0.97, Tables 2 and Supplementary Tables 1–3), but a few hierarchies had a transitivity index close to the random level of transitivity set at 0.75 as in redfronted lemurs (group X) or well below this random level as in rock hyraxes (group Isiim) (Table 2). There were no strong correlations between the degree of female dominance and the degree of transitivity (correlation coefficient = 0.07, Supplementary Figure 2).
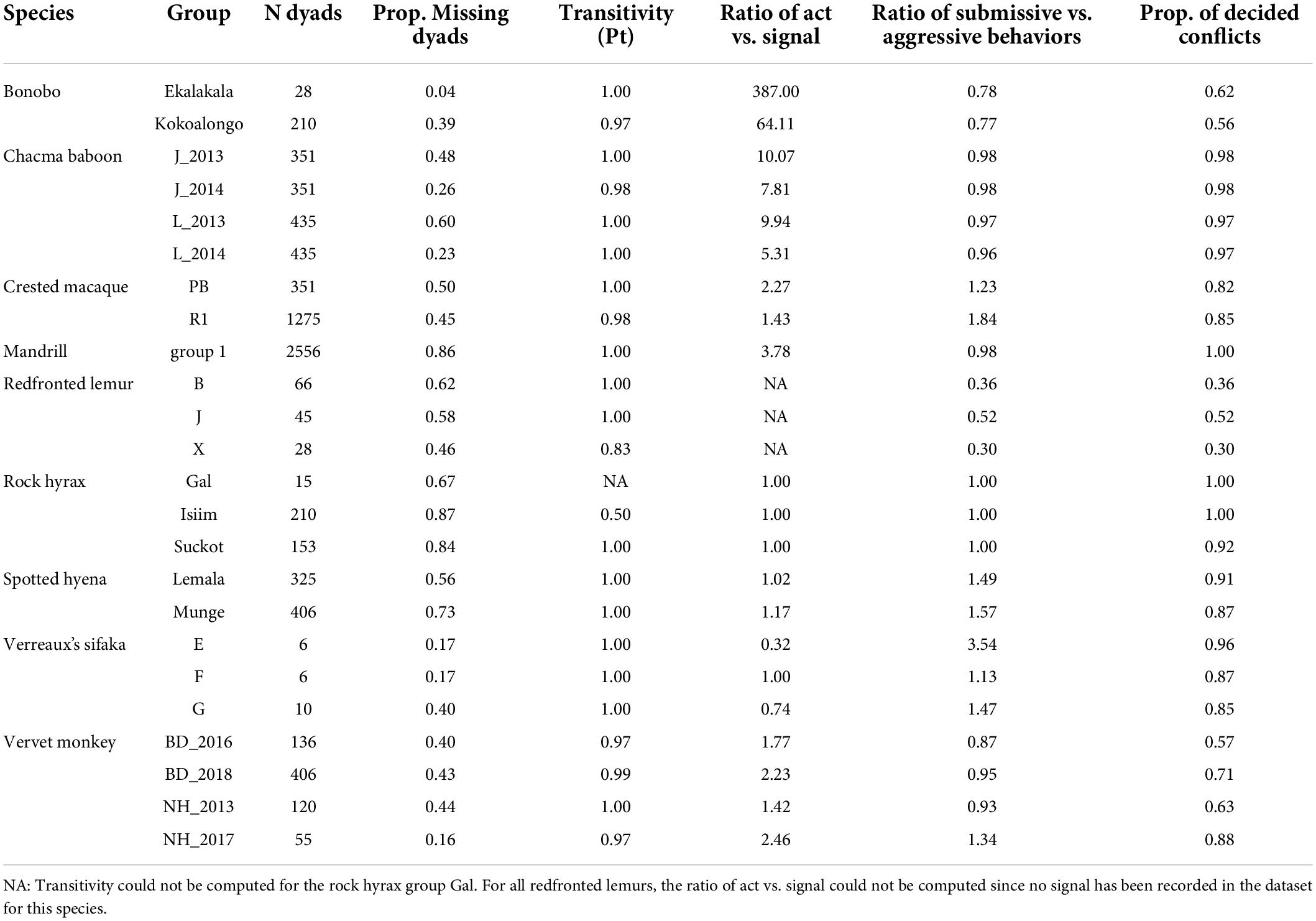
Table 2. Characteristics of the intersexual hierarchies and conflict types used in this study.
The species included in this comparative study represent a continuum in terms of the degree of intersexual dominance, from strictly male-dominated species as in the two chacma baboon groups (all indices ≤ 0.09% group males dominated by each female), to strictly female-dominated species as in the three Verreaux’s sifaka groups (all indices ≥ 0.94, Figures 1, 2 and Supplementary Table 4). Importantly, our dataset does not comprise only species at the extreme ends of the spectrum of intersexual dominance but also several species where dominance is not very biased toward one sex (e.g., some redfronted lemur, bonobo or vervet monkey groups, Figures 1, 2 and Supplementary Table 4).
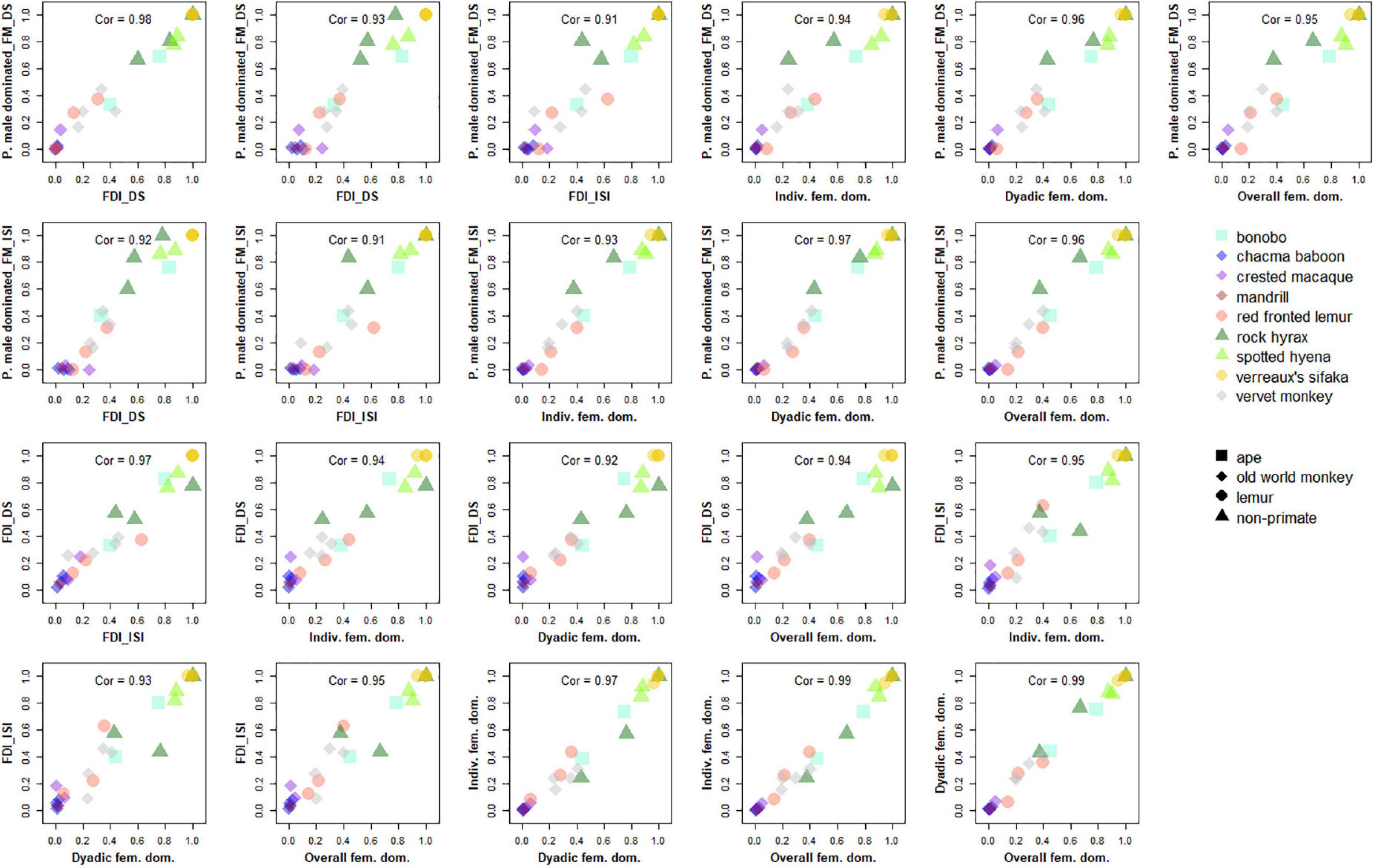
Figure 1. Correlations among the five female dominance indices. All pairwise comparisons among the five indices are shown, with each point representing one social group of the nine species. The 7 indices are abbreviated on the axis as follow: P. male dominated._FM_DS: percentage of males dominated by each female in the hierarchy compiled using David’s score and based on heterosexual conflicts only (Index 1a); P. male dominated._FM_ISI: percentage of males dominated by each female in the hierarchy compiled using I&SI and based on heterosexual conflicts only (Index 1b); FDI_DS: female dominance index based on hierarchies compiled using David’s score and based on all conflicts (Index 2a); FDI_ISI: female dominance index based on hierarchies compiled using I&SI and based on all conflicts (Index 2b); Indiv. fem. dom.: Individual female dominance, percentage of intersexual conflicts won by females calculated for each female and averaged across all females (Index 3); Dyadic fem. dom.: Dyadic female dominance, percentage of intersexual conflicts won by females in each male-female dyad and averaged across all heterosexual dyads (Index 4); Overall fem. dom.: Individual female dominance, percentage of intersexual conflicts won by females over males overall across all heterosexual conflicts (Index 5). “Cor”: correlation coefficient calculated using Spearman rank correlation tests.
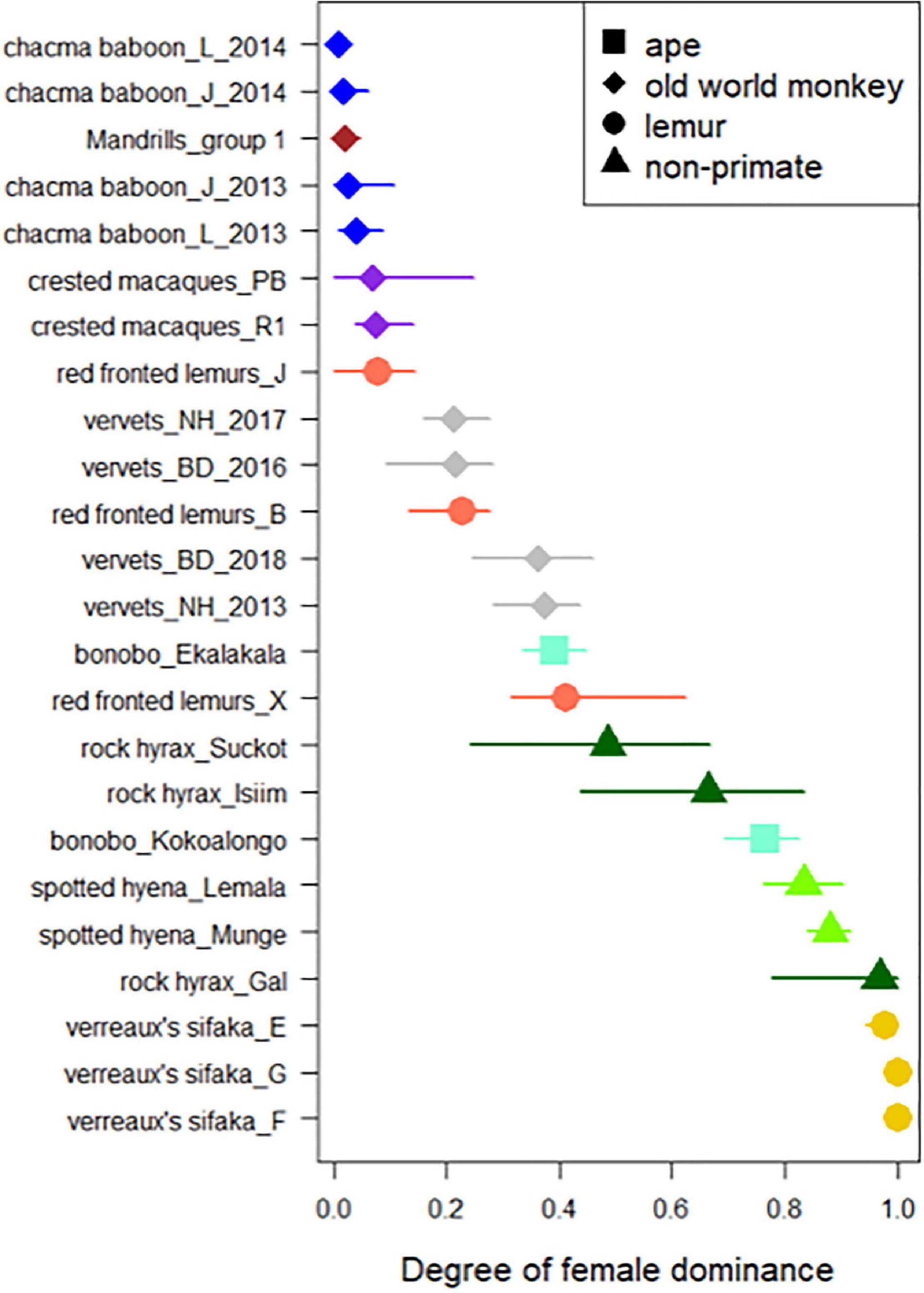
Figure 2. Distribution of the female dominance indices in the 24 study groups. Each line represents a study group during a given year. The symbol depicts the mean across all seven female dominance indices and the line the range of these indices. Each species is depicted with a different color. Apes, old world monkeys, lemurs and non-primates are depicted using squares, diamonds, circles and triangles, respectively.
All five female dominance indices correlated strongly with each other (Spearman R2 range: 0.92–0.98, Figures 1, 2), indicating that different indices of female dominance can be used interchangeably, as they do not influence the position of each group/species on the spectrum of the extent of female dominance. Please note however that for datasets with a large proportion of missing dyads the different indices provided more variable values (Figure 2 and Supplementary Tables 1–3).
In models 1a and 1b, the full model was not significantly different from the null model (Model 1a: N = 21 group_years, df = 1, χ2 = 0.096, P = 0.756; Model 1b: N = 24 group_years, df = 1, χ2 = 1.613, P = 0.204) indicating that variable degrees of intersexual dominance did not significantly impact the differences in dominance rank observed between the intra- and the intersexual dominance hierarchy for both males and females (Supplementary Table 5 and Figure 3). These changes were rather influenced by the structure of the datasets, at least for the female hierarchies in which correlation between the intra- and the intersexual dominance hierarchies were influenced by the percentage of missing dyads (P = 0.051, Supplementary Table 5). Nevertheless, overall, intra- and intersexual hierarchies were highly correlated, across variable degrees of intersexual dominance, especially for females (Figure 3).

Figure 3. Degree of female dominance and correlation between intra- and intersexual hierarchies for females (left panel) and males (right panel). Each species is depicted using a different color. Apes, old world monkeys, lemurs and non-primates are depicted using squares, diamonds, circles and triangles, respectively. Each dot represents one study group in a given year. The black lines indicate the model’s predictive lines for the effect of the degree of female dominance on the correlation between individual ranks from the intra- and from the intersexual hierarchies for females (Model 1a, left panel) and for males (Model 1b, left panel). The lines are only indicative since the p-values for these effects were non-significant in both models. Verreaux’s sifakas do not appear in the left panel since there was only one female in each of the three groups and correlations between female ranks in the intra and intersexual hierarchies could thus not be calculated.
Individuals in most study species used acts more than signals in agonistic interactions, with some species, like redfronted lemurs and bonobos, using acts almost exclusively (Table 2). Verreaux’s sifakas diverge from this general trend: they used acts and signals equally in one group and signals more often than acts in the two other groups. Together with mandrills, Verreaux’s sifakas are also the study species using the highest ratio of submissive to aggressive behaviors.
The full model was significantly different from the null model in three out of four models testing the effect of the degree of female dominance and of the dyad type (M-F, F-F, M-M) on the probability of using aggressive and submissive acts or signals (Ninteractions = 11.492, Ndyads = 2908, Nindividuals = 508, LRT, df = 5, Model 2a: χ2 = 34.97, P < 0.001; Model 2b: χ2 = 57.25, P < 0.001; Model 2c: χ2 = 8.30, P = 0.141, and Model 2d: χ2 = 37.15, P < 0.001). In Model 2a investigating the probability of using aggressive acts, the interaction between the degree of female dominance and dyad type was marginally non-significant (P = 0.070, Table 3). For FM and FF dyads, the probability to use aggressive acts decreased slightly with increasing female dominance, and such decrease was steeper for MM dyads (Figure 4A and Supplementary Figure 4). We found similar results in Model 2a bis with a reduced dataset excluding crested macaques and spotted hyenas (Supplementary Figure 5) with the exception that the interaction between the degree of female dominance and dyad type was significant (P = 0.001, Supplementary Table 6) and that the probability to use aggressive acts did not decrease with increasing female dominance for FF dyads. In Model 2b, we found a significant interaction between the degree of female dominance and the probability of expressing aggressive signals (P = 0.033, Table 3). For FM and FF dyads, the probability for aggressive signals to occur in agonistic interactions decreased with increasing degree of female dominance but remained stable for MM dyads (Figure 4B and Supplementary Figure 4). In Model 2c, the degree of female dominance and dyad type did not significantly affect the probability for submissive acts to occur during agonistic interactions (Figure 4C and Supplementary Figure 4). Finally, Model 2d revealed a significant interaction between the degree of female dominance and dyad type on the probability for submissive signals to occur (P = 0.004, Table 3). While the probability for submissive signals to occur during agonistic interactions increased steadily with the degree of female dominance, this positive relationship was less pronounced for FM dyads compared to MM and FF dyads (Figure 4D and Supplementary Figure 4). The marginal R2 were 0.142, 0.202, 0.050, 0.057, and 0.190 for Models 1a,1a bis, 1b,1c, and 1d, respectively. The conditional R2 were 0.819, 0.608, 0.864, 0.531, and 0.907 for Models 1a, 1a bis, 1b, 1c, and 1d, respectively.
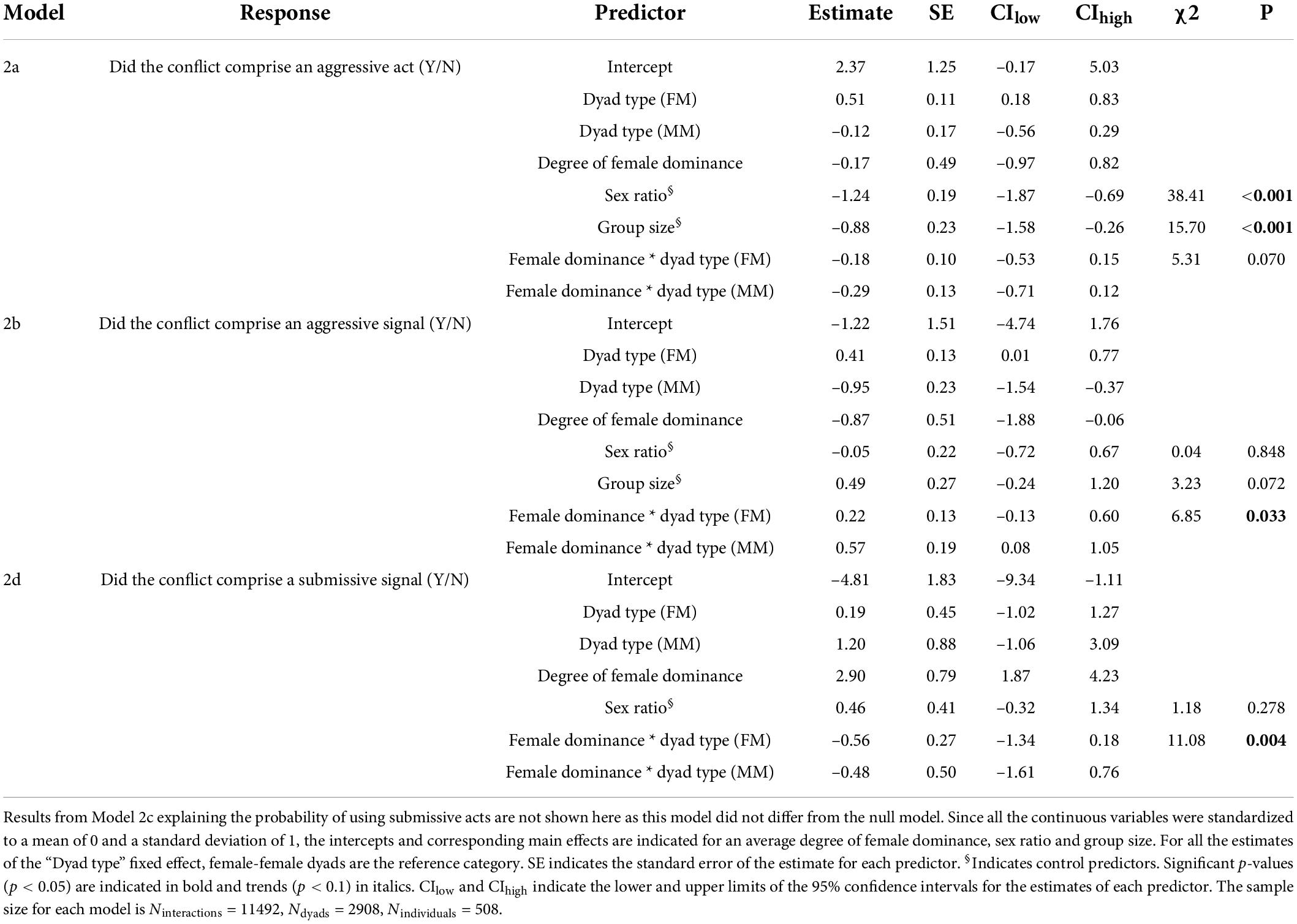
Table 3. Effect of the degree of female dominance and dyad type (FM, FF, MM) on the probability of using aggressive acts (Model 2a), aggressive signals (Model 2b), and submissive signals (Model 2d).
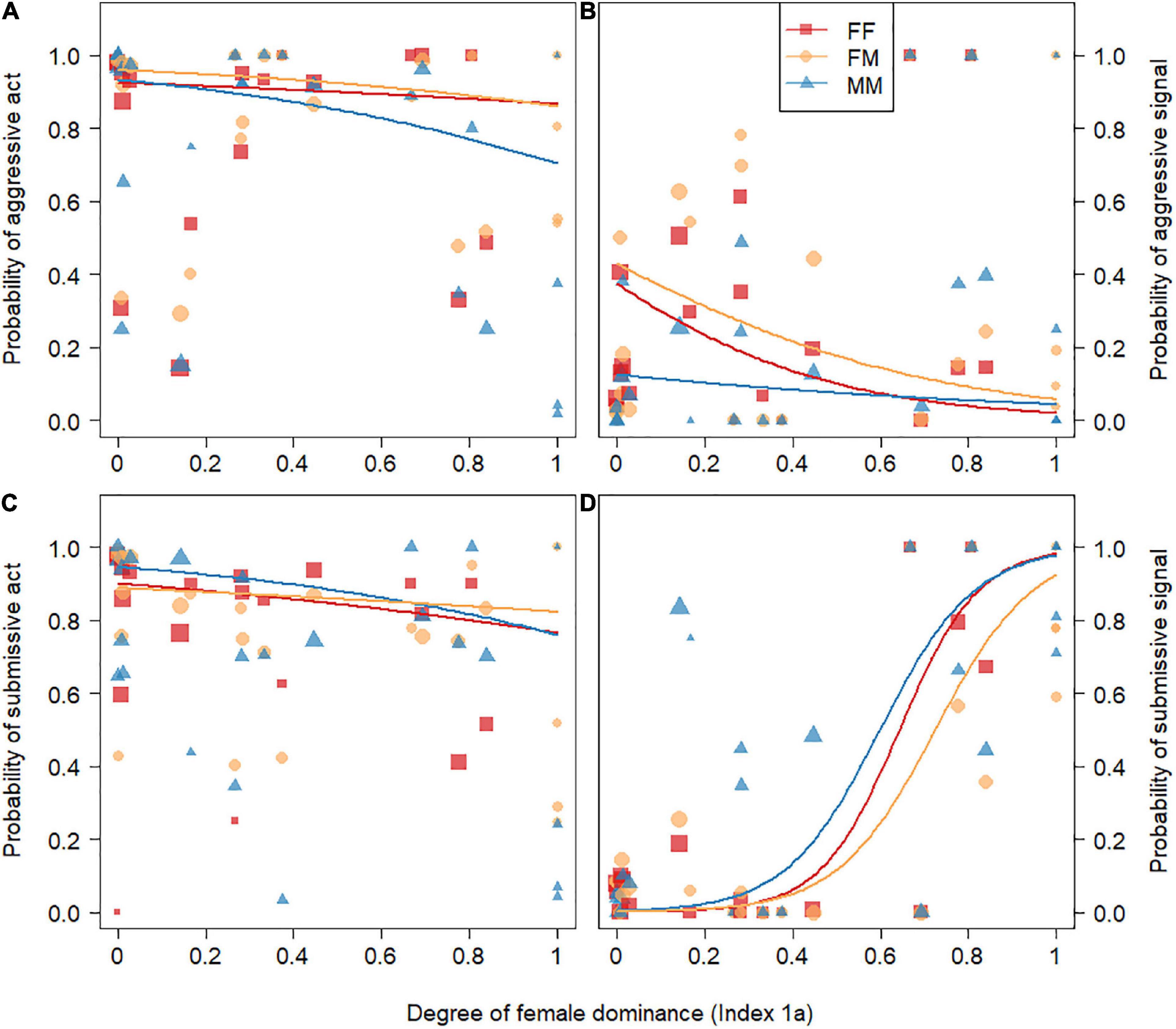
Figure 4. Influence of the degree of female dominance (Index 1a) and dyad type (FM, FF, MM) on the likelihood for conflicts to comprise aggressive acts (A), aggressive signals (B), submissive acts (C), or submissive signals (D). Each dyad type is depicted using a different color and symbol: Red square: female-female dyads (FF), orange circle: female-male dyads (FM) and blue triangle: male-male dyads (MM). Each dot represents one dyad type of a study group on a given year and dot size is proportional to the log number of dyads. Log scale was chosen here because the number of dyads in each dyad type in the largest groups were orders of magnitude larger than in smallest groups. The red, orange and blue lines depict the predicted relationship between the degree of female dominance and the likelihood of each act or signal to occur for FF, FM, and MM dyads respectively. These lines are derived from Models 2a (panel A), 2b (panel B), 2c (panel C), and 2d (panel D). Please note that for Model 2c the full model did not significantly differ from the null model and the corresponding lines do not represent meaningful statistical relationships but are used for illustration.
Our study confirmed that intersexual dominance varies along a continuum from strict female dominance to strict male dominance independent of the measure used. All indices of the degree of female dominance were well correlated with each other, and the rank order among same-sex individuals was highly correlated between the intrasexual and intersexual hierarchies, and such correlation was not significantly affected by the degree of female dominance in the group. In most study groups, within sex ranks were highly correlated between the intra- and the intersexual hierarchies and variations in this correlation were function of the dataset property (i.e., % of missing dyads in Model 1a). Interestingly, the relative prevalence of each type of agonistic behavior was sensitive to variation in the degree of female dominance, with more submissive signals and fewer aggressive acts being used by both sexes and across all dyad types in societies where female dominance prevails. Below, we discuss each of these main results in light of the current knowledge of heterosexual relationships across mammalian societies.
One of the novelties of the present study was the standardized application of several methods to calculate intersexual dominance across a range of mammalian species with different social systems. We found the degree of female dominance to vary continuously from strict male dominance to strict female dominance, adding to a growing number of studies (Hemelrijk et al., 2008, 2020; Rina Evasoa et al., 2019; Davidian et al., 2022; Kappeler et al., 2022) breaking with traditional binary categorizations into female-dominant vs. male-dominant species. Clearly, binary categories are insufficient to capture the variation in intersexual dominance relationships occurring both across and within species. The latter insight is illustrated by species for which we had data from more than one group, such as bonobos, redfronted lemurs and vervet monkeys, confirming results of several recent studies (Surbeck and Hohmann, 2013; Vullioud et al., 2019; Hemelrijk et al., 2020; Izar et al., 2021). These patterns open the way for future investigations of the sources of inter- and intraspecific variation in intersexual dominance patterns.
Our study will facilitate future comparative studies by demonstrating strong positive correlations among the five indices that measure the degree of female dominance. Some indices, namely 1 and 2, required the construction of intersexual hierarchies while others, namely 3–5, simply quantified the proportion of dyadic interactions won by one sex. Their high correlation suggests that they capture the same behavioral phenomenon, and validate the use of dominance hierarchies to measure asymmetries between the sexes in the propensity to win intersexual agonistic interactions. Special care should be taken for datasets with high uncertainty induced by missing interactions for a large number of dyads, as in redfronted lemurs and rock hyraxes (Supplementary Figure 3) for which female dominance indices varied greatly. However, even with such a variation, the study groups were positioned in the same area of the spectrum characterizing the degree of female dominance, regardless of the index chosen (Figure 2). Overall these indices are thus robust and consistent. This is further confirmed by the similar values generated by David’s score or I&SI methods. These results indicate that all five indices capture meaningful aspects of dominance relationships between males and females, ensuring the comparability of past, present and future studies using one or several of these indices.
By positioning each study group along the intersexual dominance gradient using a standardized approach, we generated a fine-grained picture of their relative order. Such positions were largely consistent with previous descriptions of intersexual dominance for most of these species; some were already described as predominantly male dominant, like mandrills (Setchell et al., 2001), chacma baboons (Kalbitzer et al., 2015), and crested macaques (Duboscq et al., 2013; Tyrrell et al., 2020), or predominantly female dominant, like Verreaux’s sifaka (Richard and Heimbuch, 1975), spotted hyenas (Vullioud et al., 2019), bonobos (Parish, 1996; Parish et al., 2000), or rock hyraxes (Koren et al., 2006; Koren and Geffen, 2009). Similarly, redfronted lemurs exhibited no sex-biased or slightly male-biased intersexual dominance as previously studied in captive and wild populations (Pereira et al., 1990; Ostner and Kappeler, 1999). The relatively balanced and flexible intersexual dominance characterizing vervet monkeys (Young et al., 2017; Hemelrijk et al., 2020) was also reported by recent studies of capuchin monkeys (Izar et al., 2021). For other groups or species, indices of female dominance contrasted with previous empirical evidence. For example, this study revealed that female dominance in bonobos is far from strict, since one group appears predominantly male dominant (Table 3). Generally, intersexual dominance relationships have been particularly well studied in the set of species selected for this study, but much remains to be learned about the generality of the patterns reported here from future studies of many other species and populations where social hierarchies have so far been examined separately for males and females.
One cannot simply assume that an individual’s position in the intrasexual hierarchy is directly predictive of its position in the intersexual hierarchy given the functional and structural differences between male and female intrasexual hierarchies (Clutton-Brock and Huchard, 2013b). Males and females often – but not always (Vullioud et al., 2019) – show distinct mechanisms of rank acquisition (de Waal, 1984; Holekamp and Smale, 1991; Foerster et al., 2016), as well as different hierarchical properties, such as steepness (Stevens et al., 2007) or stability (Holekamp and Smale, 1991). We found that in most cases, individuals maintained their same-sex dominance ordering in the intersexual hierarchy. For example, if four males are ranked A-B-C-D in the intrasexual hierarchy, it is very likely that the rank order of these males will be maintained in the intersexual dominance hierarchy, even if some females are ranked in between or above those males. This result clarifies an important aspect of the structure of intersexual hierarchies. Like intrasexual hierarchies in most species included in this study, intersexual hierarchies are mostly highly transitive (with the exception of one group of rock hyrax and one group of redfronted lemurs, Supplementary Figure 2). Intersexual ranks are simply obtained by merging both intrasexual hierarchies, respecting their initial order, but at variable levels, i.e., from a full entanglement, sometimes referred to as “codominance” (Lewis, 2018), to strict male or female dominance, where all members of one sex outrank all members of the other sex. The fact that the degree of female dominance was not found to influence these patterns suggests that this effect persists across the whole gradient of intersexual dominance. Overall, individuals of both sexes can thus be ordered together in a common, meaningful intersexual hierarchy, according to their competitive abilities, whatever they might be based on. While models suggest that an individual’s experiences with the self-reinforcing effects of winning and losing fights may concern interactions with both males and females (Hemelrijk et al., 2008), this is not necessarily so, as indicated by evidence of a sex difference in the winner-loser effect in hamsters (Solomon et al., 2007).
A few social groups in our dataset showed a relatively low correlation between an individual’s position in the same-sex vs. intersexual hierarchy, as in males of one chacma baboon group, or in females of one bonobo, one crested macaque and one rock hyrax group (Figure 3). It is possible that the number of dyads for which no interaction was recorded may affect the robustness of hierarchies. Yet, this is unlikely to explain our results given that the percentage of missing dyads was included as a control factor in our models and did not yield a significant effect on the correlation between the two hierarchies. In addition, there are species and sex differences in the dynamics of hierarchies that are not accounted for here, and which may influence hierarchical properties, especially transitivity. Specifically, the methods used here to build hierarchies ignore temporal changes in hierarchies, whereas other methods continuously adjust and update a hierarchy over time when it is characterized by high temporal dynamics (e.g., use of the dynamic Elo-ranking methods: Neumann et al., 2011). Static approaches may be suitable in some cases, such as the stable, linear and heritable hierarchies of female cercopithecines, but less so for unstable male hierarchies in seasonal breeding species where most males are not permanent group members, as in mandrills (Brockmeyer et al., 2015) and rock hyraxes (Barocas et al., 2011), or species where males move frequently between groups, as in crested macaques (Neumann et al., 2011; Marty et al., 2016). The moderate correspondence between intra- and intersexual hierarchies may thus reflect the social dynamics in these species, which may occur over the course of a year (i.e., the time frame used here to derive hierarchies).
We finally investigated whether the relative importance of agonistic acts vs. signals and of aggressive vs. submissive behavior varies across the spectrum of interspecific variation in intersexual dominance. This analysis highlighted at least two important results. First, the relative frequencies with which different agonistic behaviors were used followed the same pattern in relation to the degree of female dominance for male-male, female-female, and male-female dyads. Second, and most noticeably, an increase in the degree of female dominance was related to a steep increase in submissive signals during contests and a modest decrease in the use of aggressive acts. This pattern suggests that in societies where dominance is biased toward females, signals are particularly important for structuring social life and likely limit the use of direct aggression during conflicts, compared to societies dominated by males. Nonetheless, the intensity and frequency of agonistic acts or signals can vary even between species that are positioned alongside on intersexual dominance spectrum. For example, across macaque societies, which are all largely male-dominated, there are well-described differences between species in the ratio of contact aggression and non-contact aggression, which are typically related to a species’ “dominance style,” which characterizes dominance relationships from most egalitarian to most despotic (Thierry, 2007).
The relationship between the degree of female dominance and the differential use of aggression and submission might partly reflect a reduction in male aggressiveness across the female dominance gradient, along with a decreasing magnitude of sexual dimorphism. Indeed, in several species where females dominate males, their body size or levels of aggressiveness and androgens resemble those of males – as in many lemurs (Jolly, 1984; Petty and Drea, 2015; Grebe et al., 2019), spotted hyenas (McCormick et al., 2021), rock hyraxes (Koren et al., 2006), and meerkats (Davies et al., 2016), and ongoing research confirms that the degree of sexual dimorphism is a strong predictor of the outcome of intersexual agonistic interactions across primates (Huchard et al., unpubl. data). In addition, species-specific patterns of agonistic interactions (Supplementary Figure 2) suggest that the general, interspecific relationship is largely driven by spotted hyenas and sifakas, which stand out by using fewer aggressive acts than other female-dominant species. In contrast, the increased use of submissive signals in female-dominant species is robust and involves most populations where dominance is largely female-biased, except for bonobos. Overall, female-biased dominance appears associated with a higher ratio of submissive signals to aggressive acts, a result that needs verification with a larger number of species, as well as using analyses that control for phylogenetic proximity between species. Controlling for phylogeny was here complicated by the low number of species involved, coupled with a heterogeneous phylogenetic coverage comprising a disproportionate number of primates as well as a couple of other, distantly related species.
The generality of this phenomenon across all dyad types may be compatible with the idea that agonistic interactions are ruled by convention-based norms that are shared by group members across age-sex-classes. These norms might possibly be socially transmitted, or flexibly influenced by prior experience, such as winner-loser effects (Tibbetts et al., 2022). Regardless of the underlying mechanism, these results suggest that female-dominated societies may rely less on aggression than male-dominated ones. This constitutes an important finding to identify the consequences of intersexual power for individual health, welfare and fitness, as well as for social dynamics and population demography. Our findings may also have ramifications for a better understanding of variation in the level of intra- and intersexual violence across past and contemporary human societies (Gómez et al., 2016; Glowacki et al., 2020).
Other patterns are more discrete. The use of aggressive signals also covaried with the degree of female dominance in female-female and intersexual dyads, with fewer threats observed in societies where ritualized submissive signals limit the use of direct aggression; threats may therefore lose their intimidating function when they are not reinforced by direct aggression. Alternatively, it is possible that either aggressive or submissive signals are needed to maintain a dominance hierarchy (Tibbetts et al., 2022), but that both are functionally redundant and rarely co-exist. Male-male dyads, in contrast, hardly use any threats across the continuum, possibly because it is risky to threaten a rival in male-dominant societies where male-male competition is often intense and contest-based, while threats are uncommon in female-dominant societies for the reasons discussed above. Finally, the slight (non-significant) decline of submissive acts along the female dominance gradient likely reflects the decline of aggressive acts, probably because submissive acts represent responses to aggressive acts. In contrast, aggressive acts are not systematically followed by a submissive act, which may explain why the decline is less pronounced for submissive than aggressive acts.
This study provides important insights and key methodological tools to study intersexual dominance relationships in mammals, and perhaps in other vertebrates. First, we show that several distinct quantitative indices of intersexual dominance are equally successful at ordering groups from several populations and species along an intersexual gradient ranging from strict male to strict female dominance. Second, we show that intersexual hierarchies are meaningful emergent properties of interactions occurring within and between the sexes. These hierarchies arise from merging male and female hierarchies, where individuals retain their intrasexual rank, but can be outranked by a variable number of opposite-sex group members. Third, we found continuous variation in patterns of agonistic interactions across species, characterized by less direct aggression and more ritualized submissions in female-dominant societies, which have apparently developed potent mechanisms of conflict mitigation that promote peaceful interactions and inhibit aggression. This work provides important foundations for future studies of intersexual dominance across mammals to uncover determinants and consequences of variation in intersexual dominance comprehensively, using standardized, quantitative measures within and across societies.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: Data deposited in Figshare: https://figshare.com/articles/dataset/Dataset_for_Sex_and_Dominance_How_to_Assess_and_Interpret_Intersexual_Dominance_Relationships_in_Mammalian_Societies_/19583215.
This study uses data from nine different species coming from countries with different regulations. All details regarding permits and authorizations have been provided in the relevant section of the manuscript.
The manuscript was conceived collectively based on shared discussions among co-authors during a workshop that took place in 2020 in Leiden (Netherlands), co-organized by CH and PK. PK, EH, and CG-B drafted the manuscript. CG-B performed the data analyses. All authors contributed critically to writing, either collected, curated, and/or analyzed the behavioral data (from their respective study population).
EH was funded by the ANR ERS-17-CE02-0008, 2018-2021 during the write-up of Tsaobis Baboon Project. Kokolopori Bonobo Research Project is funded by the Harvard University, Max-Planck Society. Project Mandrillus is funded by the SEEG Lekedi and LIA (INEE-CNRS) to MC, Agence Nationale de la Recherche (ANR SLEEP 17-CE02-0002 to MC and ANR ERS-17-CE02-0008, 2018-2021 to EH), Deutsche Forschungsgemeinschaft (DFG, KA 1082-20-1) to PK and MC, as well as by the State Scholarships Foundation (IKY) under the scholarship program from the proceeds of the “Nic. D. Chrysovergis” bequest to NS. Inkawu vervet project is funded by the Fyssen Foundation (granted to CC) and the Swiss National Science Foundation (31003A_159587 and PP00P3_170624) along with the Branco Weiss Fellowship—Society in Science (granted to EW). Macaca nigra Project is funded by the German Research Council within the Emmy-Noether programme, the Federal Ministry for Economic Cooperation and Development and the German Academic Exchange Service (to Antje Engelhardt), by the Volkswagen Stiftung (to JD) and the University of Portsmouth (to JM). Ein Gedi rock hyrax sociality project was funded by the Israel Science Foundation and United States-Israel Binational Science Foundation (2015088). Ngorongoro Hyena Project is funded by the Leibniz Institute for Zoo and Wildlife Research; Werner Dessauer Stiftung.
We would like to thank colleagues who contributed ideas or suggestions during the various discussions and exchanges that led to the setup of the study, as well as two referees for their constructive comments, and Christof Neumann for fruitful discussions. The Kokolopori Bonobo Research Project thanks the ICCN, Bonobo Conservation Initiative and Vie Sauvage for their support of the study, the Ministry of Scientific Research and Technology in the Democratic Republic of the Congo for the permission to work in the DRC, the people of the villages of Bolamba, Yete, Yomboli, and Yasalakose for granting access to their forest, all local and international field assistants for their support in the field, as well as Harvard University and the Max-Planck Society for funding the project. The Tsaobis Baboon Project is grateful to the volunteers from 2013 and 2014 for collecting behavioral data. This research was carried out with the permission of the Ministry of Environment and Tourism, the Ministry of Land Reform, and the National Commission on Research, Science, and Technology. We further thank the Tsaobis beneficiaries for permission to work at Tsaobis, the Gobabeb Namib Research Institute and Training Centre for affiliation, and Johan Venter and the Snyman and Wittreich families for permission to work on their land. This article is n° ISEM 2022-144. The Mandrillus Project is grateful to the past and present field assistants who collected behavioral data and to the SODEPAL-COMILOG society (ERAMET group) for their long-term logistical support. This is a Project Mandrillus publication number 27. This study was approved by an authorization from the CENAREST institute (permit number, AR0060/18/MESRS/CENAREST/CG/CST/CSAR). The Inkawu Vervet Project (IVP) thanks the van der Walt family for permission to conduct the study on their land and the whole IVP team for their help and support in the field. The Macaca Nigra Project in Tangkoko thank all team members, especially Antje Engelhardt and Jan-Boje Pfeifer. We thank Joko Pamungkas from the Primate Research Centre, Muhammad Agil and Perwitasari Farajallah from the Institute Pertanian Bogor University for supporting our stay in Indonesia. We gratefully acknowledge the permission of the Indonesian Institute of Sciences, the Directorate General of Forest Protection and Nature Conservation and the Department for the Conservation of Natural Resource, North Sulawesi, particularly Pak Domingus and Pak Yunus, to conduct research in the Tangkoko Reserve. The Kirindy Forest Lemur Project thanks the Malagasy Ministry of the Environment, the University of Antananarivo and the CNFEREF Morondava for authorizing our work, and the Kirindy field assistants for help with data collection. The Ein Gedi rock hyrax sociality project is grateful for the hospitality and logistic support from the Ein Gedi Field School and the Ein Gedi Nature Reserve. The Ngorongoro Hyena Project thanks the Tanzania Commission for Science and Technology, the Tanzania Wildlife Research Institute, and the Ngorongoro Conservation Area Authority for permission to conduct the study, and Bettina Wachter and Philemon Naman for long-term behavioral and demographic monitoring of the eight spotted hyena clans.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.918773/full#supplementary-material
Abernethy, K. A., White, L. J. T., and Wickings, E. J. (2002). Hordes of mandrills (Mandrillus sphinx): extreme group size and seasonal male presence. J. Zool. 258, 131–137. doi: 10.1017/S0952836902001267
Alberts, S. C. (2012). “Magnitude and sources of variation in male reproductive performance,” in The Evolution of Primate Societies, eds J. C. Mitani, J. Call, P. M. Kappeler, R. A. Palombit, and J. B. Silk (Chicago, IL: University of Chicago Press), 412–431.
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–267. doi: 10.1163/156853974x00534
Andelman, S. J. (1987). Evolution of concealed ovulation in vervet monkeys (Cercopithecus aethiops). Am. Nat. 129, 785–799. doi: 10.1086/284675
Andelman, S. J., Else, J. G., Hearn, J. P., and Hodges, J. K. (1985). The non-invasive monitoring of reproductive events in wild vervet monkeys (Cercopithecus aethiops) using urinary pregnanediol-3α-glucuronide and its correlation with behavioural observations. J. Zool. 205, 467–477. doi: 10.1111/j.1469-7998.1985.tb03538.x
Anderson, G. D., and Herlocker, D. J. (1973). Soil factors affecting the distribution of the vegetation types and their utilization by wild animals in Ngorongoro Crater, Tanzania. J. Ecol. 61, 627–651. doi: 10.2307/2258640
Arseneau-Robar, T. J. M., Taucher, A. L., Schnider, A. B., van Schaik, C. P., and Willems, E. P. (2017). Intra- and interindividual differences in the costs and benefits of intergroup aggression in female vervet monkeys. Anim. Behav. 123, 129–137. doi: 10.1016/j.anbehav.2016.10.034
Baayen, R. H., Davidson, D. J., and Bates, D. M. (2008). Mixed-effects modeling with crossed random effects for subjects and items. J. Mem. Lang. 59, 390–412. doi: 10.1016/j.jml.2007.12.005
Baniel, A., Cowlishaw, G., and Huchard, E. (2017). Male violence and sexual intimidation in a wild primate society. Curr. Biol. 27, 2163.e3–2168.e3. doi: 10.1016/j.cub.2017.06.013
Baniel, A., Cowlishaw, G., and Huchard, E. (2018). Jealous females? Female competition and reproductive suppression in a wild promiscuous primate. Proc. R. Soc. Lond. B 285:20181332. doi: 10.1098/rspb.2018.1332
Barocas, A., Ilany, A., Koren, L., Kam, M., and Geffen, E. (2011). Variance in centrality within rock hyrax social networks predicts adult longevity. PLoS One 6:e22375. doi: 10.1371/journal.pone.0022375
Barr, D. J., Levy, R., Scheepers, C., and Tily, H. J. (2013). Random effects structure for confirmatory hypothesis testing: keep it maximal. J. Mem. Lang. 68, 255–278. doi: 10.1016/j.jml.2012.11.001
Bates, D., Maechler, M., and Dai, B. (2011). lme4: Linear Mixed-Effects Models Using S4 Classes. Available online at: http://lme4.r-forge.r-project.org/ (accessed April 19, 2022).
Brockmeyer, T., Kappeler, P. M., Willaume, E., Benoit, L., Mboumba, S., and Charpentier, M. J. E. (2015). Social organization and space use of a wild mandrill (Mandrillus sphinx) group. Am. J. Primatol. 77, 1036–1048. doi: 10.1002/ajp.22439
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Canteloup, C., Puga-Gonzalez, I., Sueur, C., and van de Waal, E. (2021). The consistency of individual centrality across time and networks in wild vervet monkeys. Am. J. Primatol. 83:e23232. doi: 10.1002/ajp.23232
Chapman, T., Arnqvist, G., Bangham, J., and Rowe, L. (2003). Sexual conflict. Trends Evol. Evol. 18, 41–47. doi: 10.1016/S0169-5347(02)00004-6
Charpentier, M. J. E., Peignot, P., Hossaert-McKey, M., Gimenez, O., Setchell, J. M., and Wickings, E. J. (2005). Constraints on control: factors influencing reproductive success in male mandrills (Mandrillus sphinx). Behav. Ecol. 16, 614–623. doi: 10.1093/beheco/ari034
Charpentier, M. J. E., Peignot, P., Hossaert-McKey, M., and Wickings, E. J. (2007). Kin discrimination in juvenile mandrills, Mandrillus sphinx. Anim. Behav. 73, 37–45.
Charpentier, M. J. E., Harté, M., Poirotte, C., de Bellefon, J. M., Laubi, B., Kappeler, P. M., et al. (2020). Same father, same face: deep learning reveals selection for signaling kinship in a wild primate. Sci. Adv. 6:eaba3274.
Chase, I. D., Bartolomeo, C., and Dugatkin, L. A. (1994). Aggressive interactions and inter-contest interval: how long do winners keep winning? Anim. Behav. 48, 393–400. doi: 10.1006/anbe.1994.1253
Cheney, D. L., and Seyfarth, R. M. (1990). How Monkeys See the World. Chicago, IL: University of Chicago Press.
Cheney, D. L., Seyfarth, R. M., Fischer, J., Beehner, J. C., Bergman, T. J., Johnson, S. E., et al. (2006). “Reproduction, mortality, and female reproductive success in chacma baboons of the Okavango Delta, Botswana,” in Reproduction and Fitness in Baboons: Behavioral, Ecological, and Life History Perspectives, eds L. Swedell and S. R. Leigh (Boston, MA: Springer), 147–176.
Clark, P. R., Waller, B. M., Burrows, A. M., Julle-Danière, E., Agil, M., Engelhardt, A., et al. (2020). Morphological variants of silent bared-teeth displays have different social interaction outcomes in crested macaques (Macaca nigra). Am. J. Phys. Anthropol. 173, 411–422. doi: 10.1002/ajpa.24129
Clutton-Brock, T. H., and Huchard, E. (2013a). Social competition and its consequences in female mammals. J. Zool. 289, 151–171. doi: 10.1111/jzo.12023
Clutton-Brock, T. H., and Huchard, E. (2013b). Social competition and selection in males and females. Philos. Trans. R. Soc. Lond. B 368:20130074. doi: 10.1098/rstb.2013.0074
Clutton-Brock, T. H., and Janson, C. H. (2012). Primate socioecology at the crossroads: past, present, and future. Evol. Anthropol. 21, 136–150. doi: 10.1002/evan.21316
Cowlishaw, G. (2013). “Papio ursinus Chacma baboon,” in Mammals of Africa. Volume II: Primates, eds T. Butynski, J. Kingdon, and J. Kalina (London: Bloomsbury), 225–228.
Cowlishaw, G., and Dunbar, R. I. M. (1991). Dominance rank and mating success in primates. Anim. Behav. 41, 1045–1056. doi: 10.1016/S0003-3472(05)80642-6
Davidian, E., Courtiol, A., Wachter, B., Hofer, H., and Höner, O. P. (2016). Why do some males choose to breed at home when most other males disperse? Sci. Adv. 2:e1501236. doi: 10.1126/sciadv.1501236
Davidian, E., Surbeck, M., Lukas, D., Kappeler, P. M., and Huchard, E. (2022). The eco-evolutionary landscape of power relationships between males and females. Trends Evol. Evol. 37, 706–718. doi: 10.1016/j.tree.2022.04.004
Davidian, E., Wachter, B., Heckmann, I., Dehnhard, M., Hofer, H., and Höner, O. P. (2021). The interplay between social rank, physiological constraints and investment in courtship in male spotted hyenas. Funct. Ecol. 35, 635–649. doi: 10.1111/1365-2435.13733
Davies, C. S., Smyth, K. N., Greene, L. K., Walsh, D. A., Mitchell, J., Clutton-Brock, T. H., et al. (2016). Exceptional endocrine profiles characterise the meerkat: sex, status, and reproductive patterns. Sci. Rep. 6:35492. doi: 10.1038/srep35492
de Vries, H. (1998). Finding a dominance order most consistent with a linear hierarchy: a new procedure and review. Anim. Behav. 55, 827–843. doi: 10.1006/anbe.1997.0708
de Waal, F. B. M. (1984). Sex differences in the formation of coalitions among chimpanzees. Ethol. Sociobiol. 5, 239–255. doi: 10.1016/0162-3095(84)90004-9
Deag, J. M. (1977). Aggression and submission in monkey societies. Anim. Behav. 25, 465–474. doi: 10.1016/0003-3472(77)90021-5
Dehnen, T., Arbon, J. J., Farine, D. R., and Boogert, N. J. (2022). How feedback and feed-forward mechanisms link determinants of social dominance. Biol. Rev. 97, 1210–1230. doi: 10.1111/brv.12838
Dezeure, J., Charpentier, M. J. E., and Huchard, E. (2022). Fitness effects of seasonal birth timing in a long-lived social primate living in the equatorial forest. Anim. Behav. 185, 113–126. doi: 10.1016/j.anbehav.2022.01.002
Dobson, A. J. (2002). An Introduction to the Generalized Linear Models, 2nd Edn. Boca Raton, FL: Chapman and Hall, CRC Press.
Douglas, P. H., Hohmann, G., Murtagh, R., Thiessen-Bock, R., and Deschner, T. (2016). Mixed messages: wild female bonobos show high variability in the timing of ovulation in relation to sexual swelling patterns. BMC Evol. Biol. 16:140. doi: 10.1186/s12862-016-0691-3
Drea, C. M. (2020). Design, delivery and perception of condition-dependent chemical signals in strepsirrhine primates: implications for human olfactory communication. Philos. Trans. R. Soc. Lond. B 375:20190264. doi: 10.1098/rstb.2019.0264
Duboscq, J., Micheletta, J., Agil, M., Hodges, K., Thierry, B., and Engelhardt, A. (2013). Social tolerance in wild female crested macaques (Macaca nigra) in Tangkoko-Batuangus Nature Reserve, Sulawesi, Indonesia. Am. J. Primatol. 75, 361–375. doi: 10.1002/ajp.22114
Duboscq, J., Neumann, C., Agil, M., Perwitasari-Farajallah, D., Thierry, B., and Engelhardt, A. (2017). Degrees of freedom in social bonds of crested macaque females. Anim. Behav. 123, 411–426. doi: 10.1016/j.anbehav.2016.11.010
Dunham, A. E. (2008). Battle of the sexes: cost asymmetry explains female dominance in lemurs. Anim. Behav. 76, 1435–1439. doi: 10.1016/j.anbehav.2008.06.018
East, M. L., Höner, O. P., Wachter, B., Wilhelm, K., Burke, T., and Hofer, H. (2009). Maternal effects on offspring social status in spotted hyenas. Behav. Ecol. 20, 478–483. doi: 10.1093/beheco/arp020
Fedigan, L., and Fedigan, L. M. (1988). “Cercopithecus aethiops: a review of field studies,” in A Primate Radiation: Evolutionary Biology of the African Guenons, eds A. Gautier-Hion, F. Bourliere, J.-P. Gautier, and J. Kingdon (Cambridge, MA: Cambridge University Press), 389–411.
Fichtel, C., Schnoell, A. V., and Kappeler, P. M. (2018). Measuring social tolerance: an experimental approach in two lemurid primates. Ethology 124, 65–73. doi: 10.1111/eth.12706
Fischer, J., Higham, J. P., Alberts, S. C., Barrett, L., Beehner, J. C., Bergman, T. J., et al. (2019). Insights into the evolution of social systems and species from baboon studies. eLife 8:e50989. doi: 10.7554/eLife.50989
Foerster, S., Franz, M., Murray, C. M., Gilby, I. C., Feldblum, J. T., Walker, K. K., et al. (2016). Chimpanzee females queue but males compete for social status. Sci. Rep. 6:35404. doi: 10.1038/srep35404
Fox, J., and Weisberg, S. (2011). An {R} Companion to Applied Regression, 2nd Edn. Thousand Oaks, CA: Sage.
Franz, M., McLean, E., Tung, J., Altmann, J., and Alberts, S. C. (2015). Self-organizing dominance hierarchies in a wild primate population. Proc. R. Soc. Lond. B 282:20151512. doi: 10.1098/rspb.2015.1512
Glowacki, L., Wilson, M. L., and Wrangham, R. W. (2020). The evolutionary anthropology of war. J. Econ. Behav. Organ. 178, 963–982. doi: 10.1016/j.jebo.2017.09.014
Gómez, J. M., Verdú, M., González-Megías, A., and Méndez, M. (2016). The phylogenetic roots of human lethal violence. Nature 538, 233–237. doi: 10.1038/nature19758
Goodall, J. (1968). The behaviour of free-living chimpanzees in the Gombe Stream Reserve. Anim. Behav. Monogr. 1, 161–311. doi: 10.1016/S0066-1856(68)80003-2
Grebe, N. M., Fitzpatrick, C., Sharrock, K., Starling, A., and Drea, C. M. (2019). Organizational and activational androgens, lemur social play, and the ontogeny of female dominance. Horm. Behav. 115:104554. doi: 10.1016/j.yhbeh.2019.07.002
Gruber, T., and Clay, Z. (2016). A comparison between bonobos and chimpanzees: a review and update. Evol. Anthropol. 25, 239–252. doi: 10.1002/evan.21501
Guhl, A. M. (1968). Social inertia and social stability in chickens. Anim. Behav. 16, 219–232. doi: 10.1016/0003-3472(68)90003-1
Hand, J. L. (1986). Resolution of social conflicts: dominance, egalitarianism, spheres of dominance, and game theory. Q. Rev. Biol. 61, 201–220. doi: 10.1086/414899
Hausfater, G. (1975). Dominance and reproduction in baboons (Papio cynocephalus). Contrib. Primatol. 7, 1–150.
Hemelrijk, C. K., and Gygax, L. (2004). Dominance style, differences between the sexes and species: an agent-based model. Interact. Stud. 5, 131–146. doi: 10.1075/is.5.1.07hem
Hemelrijk, C. K., Wantia, J., and Isler, K. (2008). Female dominance over males in primates: self-organisation and sexual dimorphism. PLoS One 3:e2678. doi: 10.1371/journal.pone.0002678
Hemelrijk, C. K., Wubs, M., Gort, G., Botting, J., and van de Waal, E. (2020). Dynamics of intersexual dominance and adult sex-ratio in wild vervet monkeys. Front. Psychol. 11:839. doi: 10.3389/fpsyg.2020.00839
Higham, J. P., Heistermann, M., Agil, M., Perwitasari-Farajallah, D., Widdig, A., and Engelhardt, A. (2021). Female fertile phase synchrony, and male mating and reproductive skew, in the crested macaque. Sci. Rep. 11:4251. doi: 10.1038/s41598-021-81163-1
Hofer, H., and East, M. L. (2003). Behavioral processes and costs of co-existence in female spotted hyenas: a life history perspective. Evol. Ecol. 17, 315–331. doi: 10.1023/A:1027352517231
Holekamp, K. E., and Smale, L. (1991). Dominance acquisition during mammalian social development: the “inheritance” of maternal rank. Am. Zool. 31, 306–317. doi: 10.1093/icb/31.2.306
Höner, O. P., Wachter, B., East, M. L., Runyoro, V. A., and Hofer, H. (2005). The effect of prey abundance and foraging tactics on the population dynamics of a social, territorial carnivore, the spotted hyena. Oikos 108, 544–554. doi: 10.1111/j.0030-1299.2005.13533.x
Höner, O. P., Wachter, B., East, M. L., Streich, W. J., Wilhelm, K., Burke, T., et al. (2007). Female mate-choice drives the evolution of male-biased dispersal in a social mammal. Nature 448, 798–801. doi: 10.1038/nature06040
Höner, O. P., Wachter, B., Hofer, H., Wilhelm, K., Thierer, D., Trillmich, F., et al. (2010). The fitness of dispersing spotted hyaena sons is influenced by maternal social status. Nat. Commun. 1:60. doi: 10.1038/ncomms1059
Huchard, E., Alvergne, A., Fejan, D., Knapp, L. A., Cowlishaw, G., and Raymond, M. (2010). More than friends? Behavioural and genetic aspects of heterosexual associations in wild chacma baboons. Behav. Ecol. Sociobiol. 64, 769–781. doi: 10.1007/s00265-009-0894-3
Huchard, E., Charpentier, M. J. E., Marshall, H., King, A. J., Knapp, L. A., and Cowlishaw, G. (2013). Paternal effects on access to resources in a promiscuous primate society. Behav. Ecol. 24, 229–236. doi: 10.1093/beheco/ars158
Huchard, E., Courtiol, A., Benavides, J. A., Knapp, L. A., Raymond, M., and Cowlishaw, G. (2009). Can fertility signals lead to quality signals? Insights from the evolution of primate sexual swellings. Proc. R. Soc. Lond. B 276, 1889–1897. doi: 10.1098/rspb.2008.1923
Ilany, A., Barocas, A., Koren, L., Kam, M., and Geffen, E. (2011). Do singing rock hyraxes exploit conspecific calls to gain attention? PLoS One 6:e28612. doi: 10.1371/journal.pone.0028612
Ilany, A., Barocas, A., Koren, L., Kam, M., and Geffen, E. (2013). Structural balance in the social networks of a wild mammal. Anim. Behav. 85, 1397–1405. doi: 10.1016/j.anbehav.2013.03.032
Izar, P., Fernández-Bolaños, M., Seex, L., Gort, G., Suscke, P., Tokuda, M., et al. (2021). Female emancipation in a male dominant, sexually dimorphic primate under natural conditions. PLoS One 16:e0249039. doi: 10.1371/journal.pone.0249039
Jaeggi, A. V., Boose, K. J., White, F. J., and Gurven, M. (2016). Obstacles and catalysts of cooperation in humans, bonobos, and chimpanzees: behavioural reaction norms can help explain variation in sex roles, inequality, war and peace. Behaviour 153, 1015–1051. doi: 10.1163/1568539X-00003347
Jolly, A. (1984). “The puzzle of female feeding priority,” in Female Primates: Studies by Women Primatologists, ed. M. Small (New York, NY: Alan R. Liss), 197–215.
Jonart, L. M., Hill, G. E., and Badyaev, A. V. (2007). Fighting ability and motivation: determinants of dominance and contest strategies in females of a passerine bird. Anim. Behav. 74, 1675–1681. doi: 10.1016/j.anbehav.2007.03.012
Kalbitzer, U., Heistermann, M., Cheney, D., Seyfarth, R., and Fischer, J. (2015). Social behavior and patterns of testosterone and glucocorticoid levels differ between male chacma and Guinea baboons. Horm. Behav. 75, 100–110. doi: 10.1016/j.yhbeh.2015.08.013
Kappeler, P. M. (1993). “Female dominance in primates and other mammals,” in Perspectives in Ethology 10, eds P. P. G. Bateson, P. H. Klopfer, and N. S. Thompson (New York, NY: Plenum), 143–158.
Kappeler, P. M. (2017). Sex roles and adult sex ratios: insights from mammalian biology and consequences for primate behaviour. Philos. Trans. R. Soc. Lond. B 372:20160321. doi: 10.1098/rstb.2016.0321
Kappeler, P. M., and Fichtel, C. (2012a). “A 15-year perspective on the social organization and life history of sifaka in Kirindy Forest,” in Long-Term Field Studies of Primates, eds P. M. Kappeler and D. P. Watts (Heidelberg: Springer), 101–121.
Kappeler, P. M., and Fichtel, C. (2012b). Female reproductive competition in Eulemur rufifrons: eviction and reproductive restraint in a plurally breeding Malagasy primate. Mol. Ecol. 21, 685–698. doi: 10.1111/j.1365-294X.2011.05255.x
Kappeler, P. M., and Fichtel, C. (2015). Eco-evo-devo of the lemur syndrome: did adaptive behavioral plasticity get canalized in a large primate radiation? Front. Zool. 12(Suppl. 1):S15. doi: 10.1186/1742-9994-12-S1-S15
Kappeler, P. M., Fichtel, C., and Radespiel, U. (2022). The island of female power? Intersexual dominance relationships in the lemurs of Madagascar. Front. Ecol. Evol. 10:858859. doi: 10.3389/fevo.2022.858859
Kappeler, P. M., and Port, M. (2008). Mutual tolerance or reproductive competition? Patterns of reproductive skew among male redfronted lemurs (Eulemur fulvus rufus). Behav. Ecol. Sociobiol. 62, 1477–1488. doi: 10.1007/s00265-008-0577-5
Kappeler, P. M., and Schäffler, L. (2008). The lemur syndrome unresolved: extreme male reproductive skew in sifakas (Propithecus verreauxi), a sexually monomorphic primate with female dominance. Behav. Ecol. Sociobiol. 62, 1007–1015. doi: 10.1007/s00265-007-0528-6
Kerhoas, D., Perwitasari-Farajallah, D., Agil, M., Widdig, A., and Engelhardt, A. (2014). Social and ecological factors influencing offspring survival in wild macaques. Behav. Ecol. 25, 1164–1172. doi: 10.1093/beheco/aru099
Kitchen, D. M., Cheney, D. L., and Seyfarth, R. M. (2005). Contextual factors meditating contests between male chacma baboons in Botswana: effects of food, friends and females. Int. J. Primatol. 26, 105–125. doi: 10.1007/s10764-005-0725-y
Koch, F., Ganzhorn, J. U., Rothman, J. M., Chapman, C. A., and Fichtel, C. (2017). Sex and seasonal differences in diet and nutrient intake in Verreaux’s sifakas (Propithecus verreauxi). Am. J. Primatol. 79:e22595. doi: 10.1002/ajp.22595
Koch, F., Signer, J., Kappeler, P. M., and Fichtel, C. (2016). Intergroup encounters in Verreaux’s sifkas (Propithecus verreauxi): who fight and why? Behav. Ecol. Sociobiol. 70, 797–808. doi: 10.1007/s00265-016-2105-3
Koenig, A., Miles, A., Riaz, D.-E.-A., and Borries, C. (2022). Intersexual agonism in gray langurs reflects male dominance and feeding competition. Front. Ecol. Evol. 10:860437. doi: 10.3389/fevo.2022.860437
Koren, L., and Geffen, E. (2009). Androgens and social status in female rock hyraxes. Anim. Behav. 77, 233–238. doi: 10.1016/j.anbehav.2008.09.031
Koren, L., and Geffen, E. (2011). Individual identity is communicated through multiple pathways in male rock hyrax (Procavia capensis) songs. Behav. Ecol. Sociobiol. 65, 675–684. doi: 10.1007/s00265-010-1069-y
Koren, L., Mokady, O., and Geffen, E. (2006). Elevated testosterone levels and social ranks in female rock hyrax. Horm. Behav. 49, 470–477. doi: 10.1016/j.yhbeh.2005.10.004
Koren, L., Mokady, O., and Geffen, E. (2008). Social status and cortisol levels in singing rock hyraxes. Horm. Behav. 54, 212–216. doi: 10.1016/j.yhbeh.2008.02.020
Koren, L., Weissman, Y., Schnitzer, I., Beukeboom, R., Bar Ziv, E., Demartsev, V., et al. (2019). Sexually opposite effects of testosterone on mating success in wild rock hyrax. Behav. Ecol. 30, 1611–1617. doi: 10.1093/beheco/arz125
Leigh, S. R., Setchell, J. M., Charpentier, M. J. E., Knapp, L. A., and Wickings, E. J. (2008). Canine tooth size and fitness in male mandrills (Mandrillus sphinx). J. Hum. Evol. 55, 75–85. doi: 10.1016/j.jhevol.2008.01.001
Lewis, R. J. (2002). Beyond dominance: the importance of leverage. Q. Rev. Biol. 77, 149–164. doi: 10.1086/343899
Lewis, R. J. (2018). Female power in primates and the phenomenon of female dominance. Annu. Rev. Anthropol. 47, 533–551. doi: 10.1146/annurev-anthro-102317-045958
Lewis, R. J., and van Schaik, C. P. (2007). Bimorphism in male Verreaux’s sifaka in the Kirindy Forest of Madagascar. Int. J. Primatol. 28, 159–182. doi: 10.1007/s10764-006-9107-3
Lindenfors, P., Gittleman, J. L., and Jones, K. E. (2007). “Sexual size dimorphism in mammals,” in Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism, eds D. J. Fairbairn, W. U. Blanckenhorn, and T. Szekely (Oxford: Oxford University Press), 16–26.
Lucchesi, S., Cheng, L., Janmaat, K., Mundry, R., Pisor, A., and Surbeck, M. (2020). Beyond the group: how food, mates, and group size influence intergroup encounters in wild bonobos. Behav. Ecol. 31, 519–532. doi: 10.1093/beheco/arz214
Malalaharivony, H. S., Fichtel, C., Heistermann, M., and Kappeler, P. M. (2021). Maternal stress effects on infant development in wild Verreaux’s sifaka (Propithecus verreauxi). Behav. Ecol. Sociobiol. 75:143. doi: 10.1007/s00265-021-03085-2
Martin, P., and Bateson, P. (2021). Measuring Behaviour: An Introductory Guide, 4th Edn. Cambridge, MA: Cambridge University Press.
Martinez Inigo, L. (2018). Intergroup Interactions in Crested Macaques (Macaca nigra): Factors Affecting Intergroup Encounter Outcome and Intensity. Ph.D. thesis. Lincoln: University of Lincoln.
Marty, P. R. (2015). Male Migration and Alpha Male Takeovers in Crested Macaques, Macaca nigra. Ph.D. thesis. Göttingen: University of Göttingen.
Marty, P. R., Hodges, K., Agil, M., and Engelhardt, A. (2016). Determinants of immigration strategies in male crested macaques (Macaca nigra). Sci. Rep. 6:32028. doi: 10.1038/srep32028
Mass, V., Heistermann, M., and Kappeler, P. M. (2009). Mate-guarding as a male reproductive tactic in Propithecus verreauxi. Int. J. Primatol. 30, 389–409. doi: 10.1007/s10764-009-9345-2
McCormick, S. K., Holekamp, K. E., Smale, L., Weldele, M. L., Glickman, S. E., and Place, N. J. (2021). Sex differences in spotted hyenas. Cold Spring Harb. Perspect. Biol. 14:a039180. doi: 10.1101/cshperspect.a039180
Micheletta, J., Waller, B. M., Panggur, M. R., Neumann, C., Duboscq, J., Agil, M., et al. (2012). Social bonds affect anti-predator behaviour in a tolerant species of macaque, Macaca nigra. Proc. R. Soc. Lond. B 279, 4042–4050. doi: 10.1098/rspb.2012.1470
Neumann, C., Assahad, G., Hammerschmidt, K., Perwitasari-Farajallah, D., and Engelhardt, A. (2010). Loud calls in male crested macaques, Macaca nigra: a signal of dominance in a tolerant species. Anim. Behav. 79, 187–193. doi: 10.1016/j.anbehav.2009.10.026
Neumann, C., Duboscq, J., Dubuc, C., Ginting, A., Irwan, A. M., Agil, M., et al. (2011). Assessing dominance hierarchies: validation and advantages of progressive evaluation with Elo-rating. Anim. Behav. 82, 911–921. doi: 10.1016/j.anbehav.2011.07.016
Neumann, C., McDonald, D. B., and Shizuka, D. (2018). Dominance ranks, dominance ratings and linear hierarchies: a critique. Anim. Behav. 144, e1–e16. doi: 10.1016/j.anbehav.2018.07.012
Norscia, I., and Palagi, E. (2015). The socio-matrix reloaded: from hierarchy to dominance profile in wild lemurs. PeerJ 3:e729. doi: 10.7717/peerj.729
Nsi Akoue, G., Mbading-Mbading, W., Willaume, E., Souza, A., Mbatchi, B., and Charpentier, M. J. E. (2017). Seasonal and individual predictors of diet in a free-ranging population of mandrills. Ethology 123, 600–613. doi: 10.1111/eth.12633
O’Brien, T. G., and Kinnaird, M. F. (1997). Behavior, diet, and movements of the Sulawesi crested black macaque (Macaca nigra). Int. J. Primatol. 18, 321–351. doi: 10.1023/A:1026330332061
Ostner, J., and Heistermann, M. (2003). Endocrine characterization of female reproductive status in wild redfronted lemurs (Eulemur fulvus rufus). Gen. Comp. Endocrinol. 131, 274–283. doi: 10.1016/s0016-6480(03)00013-3
Ostner, J., and Kappeler, P. M. (1999). Central males instead of multiple pairs in redfronted lemurs, Eulemur fulvus rufus (Primates, Lemuridae)? Anim. Behav. 58, 1069–1078. doi: 10.1006/anbe.1999.1222
Ostner, J., Kappeler, P. M., and Heistermann, M. (2002). Seasonal variation and social correlates of androgen excretion in male redfronted lemurs (Eulemur fulvus rufus). Behav. Ecol. Sociobiol. 52, 485–495. doi: 10.1007/s00265-002-0532-9
Overdorff, D. J. (1993). Similarities, differences, and seasonal patterns in the diets of Eulemur rubriventer and Eulemur fulvus rufus in the Ranomafana National Park, Madagascar. Int. J. Primatol. 14, 721–753. doi: 10.1007/BF02192188
Parish, A. R., De Waal, F. B. M., and Haig, D. (2000). The other “closest living relative”: how bonobos (Pan paniscus) challenge traditional assumptions about females, dominance, intra-and intersexual interactions, and hominid evolution. Ann. N. Y. Acad. Sci. 907, 97–113.
Peignot, P., Charpentier, M. J. E., Bout, N., Bourry, O., Massima, U., Dosimont, O., et al. (2008). Learning from the first release project of captive-bred mandrills Mandrillus sphinx in Gabon. Oryx 42, 122–131. doi: 10.1017/S0030605308000136
Pereira, A. E., Kaufman, R., Kappeler, P. M., and Overdorff, D. J. (1990). Female dominance does not characterize all of the Lemuridae. Folia Primatol. 55, 96–103. doi: 10.1159/000156505
Pereira, M. E., and Kappeler, P. M. (1997). Divergent systems of agonistic behaviour in lemurid primates. Behaviour 134, 225–274. doi: 10.1163/156853997X00467
Petty, J. M. A., and Drea, C. M. (2015). Female rule in lemurs is ancestral and hormonally mediated. Sci. Rep. 5:9631. doi: 10.1038/srep09631
Port, M., Johnstone, R. A., and Kappeler, P. M. (2010). Costs and benefits of multi-male associations in redfronted lemurs (Eulemur fulvus rufus). Biol. Lett. 6, 620–622. doi: 10.1098/rsbl.2010.0091
Preuschoft, S., Paul, A., and Kuester, J. (1998). Dominance styles of female and male Barbary macaques (Macaca sylvanus). Behaviour 135, 731–755. doi: 10.1163/156853998792640477
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reddon, A. R., Ruberto, T., and Reader, S. M. (2021). Submission signals in animal groups. Behaviour 159, 1–20. doi: 10.1163/1568539X-bja10125
Richard, A. F., and Dewar, R. E. (1991). Lemur ecology. Annu. Rev. Ecol. Evol. Syst. 22, 145–175. doi: 10.1146/annurev.es.22.110191.001045
Richard, A. F., Dewar, R. E., Schwartz, M., and Ratsirarson, J. (2002). Life in the slow lane? Demography and life histories of male and female sifaka (Propithecus verreauxi verreauxi). J. Zool. 256, 421–436. doi: 10.1017/S0952836902000468
Richard, A. F., and Heimbuch, R. (1975). “An analysis of the social behavior of three groups of Propithecus verreauxi,” in Lemur Biology, eds I. Tattersall and R. W. Sussman (Boston, MA: Springer), 313–333.
Rina Evasoa, M., Zimmermann, E., Hasiniaina, A. F., Rasoloharijaona, S., Randrianambinina, B., and Radespiel, U. (2019). Sources of variation in social tolerance in mouse lemurs (Microcebus spp.). BMC Ecol. 19:20. doi: 10.1186/s12898-019-0236-x
Rowe, N., and Myers, M. (2017). “All the world’s primates,” in The International Encyclopedia of Primatology, ed. A. Fuentes (Charlestown, RI: Primate Conservation, Inc).
Rowell, T. E. (1974). The concept of social dominance. Behav. Biol. 11, 131–154. doi: 10.1016/S0091-6773(74)90289-2
Rutte, C., Taborsky, M., and Brinkhof, M. W. G. (2006). What sets the odds of winning and losing? Trends Ecol. Evol. 21, 16–21. doi: 10.1016/j.tree.2005.10.014
Sánchez-Tójar, A., Schroeder, J., and Farine, D. R. (2018). A practical guide for inferring reliable dominance hierarchies and estimating their uncertainty. J. Anim. Ecol. 87, 594–608. doi: 10.1111/1365-2656.12776
Schjelderup-Ebbe, T. (1922). Beiträge zur Sozialpsychologie des Haushuhns. [Observation on the social psychology of domestic fowls]. Z. Psychol. Physiol. Sinnesorg. Abt. Z. Psychol. 88, 225–252.
Scholz, F., and Kappeler, P. M. (2004). Effects of seasonal water scarcity on the ranging behavior of Eulemur fulvus rufus. Int. J. Primatol. 25, 599–613. doi: 10.1023/B:IJOP.0000023577.32587.0b
Schülke, O., Bhagavatula, J., Vigilant, L., and Ostner, J. (2010). Social bonds enhance reproductive success in male macaques. Curr. Biol. 20, 2207–2210. doi: 10.1016/j.cub.2010.10.058
Setchell, J. M., Charpentier, M. J. E., and Wickings, E. J. (2005). Mate guarding and paternity in mandrills: factors influencing alpha male monopoly. Anim. Behav. 70, 1105–1120. doi: 10.1016/j.anbehav.2005.02.021
Setchell, J. M., Lee, P. C., Wickings, E. J., and Dixson, A. F. (2001). Growth and ontogeny of sexual size dimorphism in the mandrill (Mandrillus sphinx). Am. J. Phys. Anthropol. 115, 349–360. doi: 10.1002/ajpa.1091
Setchell, J. M., and Wickings, E. J. (2005). Dominance, status signals and coloration in male mandrills (Mandrillus sphinx). Ethology 111, 25–50. doi: 10.1111/j.1439-0310.2004.01054.x
Seyfarth, R. M., Cheney, D. L., and Marler, P. (1980). Monkey responses to three different alarm calls: evidence of predator classification and semantic communication. Science 210, 801–803. doi: 10.1126/science.7433999
Shizuka, D., and McDonald, D. B. (2012). A social network perspective on measurements of dominance hierarchies. Anim. Behav. 83, 925–934. doi: 10.1016/j.anbehav.2012.01.011
Silk, J. B., Beehner, J. C., Bergman, T. J., Crockford, C., Engh, A. L., Moscovice, L. R., et al. (2009). The benefits of social capital: close social bonds among female baboons enhance offspring survival. Proc. R. Soc. Lond. B 276, 3099–3104. doi: 10.1098/rspb.2009.0681
Smith, J. E., Van Horn, R. C., Powning, K. S., Cole, A. R., Graham, K. E., Memenis, S. K., et al. (2010). Evolutionary forces favoring intragroup coalitions among spotted hyenas and other animals. Behav. Ecol. 21, 284–303. doi: 10.1093/beheco/arp181
Smithson, M., and Verkuilen, J. (2006). A better lemon squeezer? Maximum-likelihood regression with beta-distributed dependent variables. Psychol. Methods 11, 54–71. doi: 10.1037/1082-989X.11.1.54
Smuts, B. B., and Smuts, R. W. (1993). Male aggression and sexual coercion of females in nonhuman primates and other mammals: evidence and theoretical implications. Adv. Study Behav. 22, 1–63. doi: 10.1016/S0065-3454(08)60404-0
Solomon, M. B., Karom, M. C., and Huhman, K. L. (2007). Sex and estrous cycle differences in the display of conditioned defeat in Syrian hamsters. Horm. Behav. 52, 211–219. doi: 10.1016/j.yhbeh.2007.04.007
Sterck, E. H. M., Watts, D. P., and van Schaik, C. P. (1997). The evolution of female social relationships in nonhuman primates. Behav. Ecol. Sociobiol. 41, 291–309.
Stevens, J. M., Vervaecke, H., De Vries, H., and van Elsacker, L. (2007). Sex differences in the steepness of dominance hierarchies in captive bonobo groups. Int. J. Primatol. 28, 1417–1430. doi: 10.1007/S10764-007-9186-9
Surbeck, M., Boesch, C., Crockford, C., Thompson, M. E., Furuichi, T., Fruth, B., et al. (2019). Males with a mother living in their group have higher paternity success in bonobos but not chimpanzees. Curr. Biol. 29, R354–R355. doi: 10.1016/j.cub.2019.03.040
Surbeck, M., and Hohmann, G. (2013). Intersexual dominance relationships and the influence of leverage on the outcome of conflicts in wild bonobos (Pan paniscus). Behav. Ecol. Sociobiol. 67, 1767–1780. doi: 10.1007/s00265-013-1584-8
Surbeck, M., Langergraber, K. E., Fruth, B., Vigilant, L., and Hohmann, G. (2017). Male reproductive skew is higher in bonobos than chimpanzees. Curr. Biol. 27, R640–R641. doi: 10.1016/j.cub.2017.05.039
Tecot, S. R., Baden, A. L., Romine, N., and Kamilar, J. M. (2013). “Reproductive strategies and infant care in the Malagasy primates,” in Building Babies: Proximate and Ultimate Perspectives of Primate Developmental Trajectories, eds K. B. H. Clancy, K. Hinde, and J. N. Rutherford (New York, NY: Springer), 321–359. doi: 10.1093/ije/dyaa233
Thierry, B. (2007). Unity in diversity: lessons from macaque societies. Evol. Anthropol. 16, 224–238. doi: 10.1002/evan.20147
Thierry, B. (2000). “Covariaton of conflict management patterns across macaque species,” in Natural Conflict Resolution, eds F. Aureli and F. B. M. De Waal (Berkeley: University of California Press), 106–128.
Tibbetts, E. A., Pardo-Sanchez, J., and Weise, C. (2022). The establishment and maintenance of dominance hierarchies. Philos. Trans. R. Soc. Lond. B 377:20200450. doi: 10.1098/rstb.2020.0450
Tokuyama, N., and Furuichi, T. (2016). Do friends help each other? Patterns of female coalition formation in wild bonobos at Wamba. Anim. Behav. 119, 27–35. doi: 10.1016/j.anbehav.2016.06.021
Turner, T. R., Schmitt, C. A., Cramer, J. D., Lorenz, J., Grobler, J. B., Jolly, C. J., et al. (2018). Morphological variation in the genus Chlorocebus: ecogeographic and anthropogenically mediated variation in body mass, postcranial morphology, and growth. Am. J. Phys. Anthropol. 166, 682–707. doi: 10.1002/ajpa.23459
Tyrrell, M., Berman, C. M., Duboscq, J., Agil, M., Sutrisno, T., and Engelhardt, A. (2020). Avoidant social style among wild crested macaque males (Macaca nigra) in Tangkoko Nature Reserve, Sulawesi, Indonesia. Behaviour 157, 451–491. doi: 10.1163/1568539X-bja10009
Valé, P. D., Béné, J.-C. K., N’Guessan, A. K., Crockford, C., Deschner, T., Koné, I., et al. (2020). Energetic management in wild chimpanzees (Pan troglodytes verus) in Taï National Park, Côte d’Ivoire. Behav. Ecol. Sociobiol. 75:1. doi: 10.1007/s00265-020-02935-9
van Schaik, C. P., and Kappeler, P. M. (1996). The social systems of gregarious lemurs: lack of convergence with anthropoids due to evolutionary disequilibrium? Ethology 102, 915–941. doi: 10.1111/j.1439-0310.1996.tb01171.x
Vilette, C., Bonnell, T., Henzi, P., and Barrett, L. (2020). Comparing dominance hierarchy methods using a data-splitting approach with real-world data. Behav. Ecol. 31, 1379–1390. doi: 10.1093/beheco/araa095
Vullioud, C., Davidian, E., Wachter, B., Rousset, F., Courtiol, A., and Höner, O. P. (2019). Social support drives female dominance in the spotted hyaena. Nat. Ecol. Evol. 3, 71–76. doi: 10.1038/s41559-018-0718-9
Walters, J. (1980). Interventions and the development of dominance relationships in female baboons. Folia Primatol. 34, 61–89. doi: 10.1159/000155948
Weingrill, T., Lycett, J. E., and Henzi, S. P. (2000). Consortship and mating success in chacma baboons (Papio cynocephalus ursinus). Ethology 106, 1033–1044. doi: 10.1046/j.1439-0310.2000.00616.x
Wright, P. C. (1999). Lemur traits and Madagascar ecology: coping with an island environment. Am. J. Phys. Anthropol. 110, 31–72. doi: 10.1002/(sici)1096-8644(1999)110:29+<31::aid-ajpa3>3.0.co;2-0
Yapuncich, G. S., Bowie, A., Belais, R., Churchill, S. E., and Walker, C. S. (2020). Predicting body mass of bonobos (Pan paniscus) with human-based morphometric equations. Am. J. Primatol. 82:e23088. doi: 10.1002/ajp.23088
Young, C., McFarland, R., Barrett, L., and Henzi, S. P. (2017). Formidable females and the power trajectories of socially integrated male vervet monkeys. Anim. Behav. 125, 61–67. doi: 10.1016/j.anbehav.2017.01.006
Keywords: dominance, sex, hierarchy, mammals, methodology
Citation: Kappeler PM, Huchard E, Baniel A, Canteloup C, Charpentier MJE, Cheng L, Davidian E, Duboscq J, Fichtel C, Hemelrijk CK, Höner OP, Koren L, Micheletta J, Prox L, Saccà T, Seex L, Smit N, Surbeck M, van de Waal E and Girard-Buttoz C (2022) Sex and dominance: How to assess and interpret intersexual dominance relationships in mammalian societies. Front. Ecol. Evol. 10:918773. doi: 10.3389/fevo.2022.918773
Received: 12 April 2022; Accepted: 06 July 2022;
Published: 29 July 2022.
Edited by:
Geoffrey M. While, University of Tasmania, AustraliaReviewed by:
Arrilton Araujo, Federal University of Rio Grande do Norte, BrazilCopyright © 2022 Kappeler, Huchard, Baniel, Canteloup, Charpentier, Cheng, Davidian, Duboscq, Fichtel, Hemelrijk, Höner, Koren, Micheletta, Prox, Saccà, Seex, Smit, Surbeck, van de Waal and Girard-Buttoz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter M. Kappeler, cGthcHBlbEBnd2RnLmRl; Elise Huchard, ZWxpc2UuaHVjaGFyZEB1bW9udHBlbGxpZXIuZnI=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.