
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 08 June 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.904576
This article is part of the Research Topic Nesting in Reptiles: Natural and Anthropogenic Threats and Evolutionary Responses View all 18 articles
 Frank J. Mazzotti1*
Frank J. Mazzotti1* Sergio A. Balaguera-Reina1
Sergio A. Balaguera-Reina1 Laura A. Brandt2
Laura A. Brandt2 Venetia Briggs-González1
Venetia Briggs-González1 Mike Cherkiss3
Mike Cherkiss3 Seth Farris1†
Seth Farris1† Avishka Godahewa1
Avishka Godahewa1Nesting ecology of American crocodiles (Crocodylus acutus) in Florida has been both positively and negatively influenced by anthropogenic and natural factors since the species was placed on the federally endangered species list in 1975. This includes a shift in nesting sites and an expansion of nesting to anthropogenic habitat. Using a 50-year record of monitoring data (1970-2020), we assessed factors influencing nesting ecology (number of nests, nest morphology, success rate, and habitat use) from a total of 3,013 nests recorded across South Florida. We detected a change in nesting success rate, increasing from 61% in the 1970’s to near 90% since 2010. Our hot spot analysis illustrates that nesting sites in northeastern Florida Bay and Flamingo/Cape Sable (Everglades National Park) were important for American crocodiles. Anthropogenic habitats, such as canals provided vital habitat nesting in areas such as Flamingo/Cape Sable (Everglades National Park), Turkey Point Power Plant, and Crocodile Lake National Wildlife Refuge for the current Florida population. Environmental parameters suspected to affect nesting success have shown an increasing trend over the past 50 years and minimum temperature and rainfall, during the summer season, are correlated with increased nesting success and temporal variation across South Florida. The adaptive capacity that American crocodiles exhibited in Florida gave the species advantages to face changes in climate and landscape over the last 50 years, however, it does not imply that the adaptive capacity of the species to face these changes (evolutionary potential) cannot reach a limit if changes continue. Here, we document C. acutus nesting ecology population responses to ecosystem restoration efforts in Florida; and further demonstrate the value of protecting and restoring habitat to support recovery of listed species.
Successful nesting is a quantifiable metric of how well a species is doing in its environment (Ugarte et al., 2013). For crocodylians, development of the egg in the nest is a vulnerable period in life, for example, increased predation risk due to exposure in less secure, elevated, open areas such as beaches or increased risk of flooding due to nesting in wetlands (Platt et al., 2008; Balaguera-Reina et al., 2015) and therefore one of the best researched population traits (see: Webb et al., 1977; Ogden, 1978; Hall and Johnson, 1987; Mazzotti, 1989; Platt and Thorbjarnarson, 2000; Ugarte et al., 2013; Combrink et al., 2017).
The American crocodile (Crocodylus acutus) is the most widely distributed species of western hemisphere crocodiles and occurs at its northernmost distribution in South Florida, then across coastal Mexico, down into South America and along the Caribbean and Pacific coasts including insular areas (Cuba, Jamaica, and Hispaniola; Thorbjarnarson et al., 2006). The species has experienced severe declines due to overexploitation and habitat loss for nesting throughout its historical distribution (Thorbjarnarson, 2010). It is presently classified as Vulnerable across its range by the International Union for Conservation of Nature (IUCN) Red List (Rainwater et al., 2021) and locally in the United States as threatened by U.S. Federal Endangered Species List (USFWS, 2007). It is also listed on Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora except for Mexico, Cuba, and Cispata Bay in Colombia (Convention on International Trade in Endangered Species of Wild Fauna and Flora [CITES], 2021).
In South Florida, nesting of American crocodiles was restricted to a small area of Northeastern Florida Bay (NEFB) in Everglades National Park (ENP) and northern Key Largo (NKL) by the early 1970s (Kushlan and Mazzotti, 1989a; Figure 1). In 1975, the species was placed on the Federal Endangered Species List (Federal Register 40). In 1978, the first nest at Turkey Point Power Plant (TP; Homestead, FL) was recorded and the first hatchlings captured, becoming since then a source of nesting for American crocodiles (Gaby et al., 1985). Expansion of crocodile nesting to the Flamingo/Cape Sable area (ENP) in the mid 1980’s, concurrent with a local restoration project that plugged both Buttonwood and East Cape canals in those areas, provided a new location for nesting (Mazzotti et al., 2007a). Critical monitoring and management efforts of the Florida population of American crocodiles (see Brandt et al., 1995; Mazzotti et al., 2007b) provided valuable information to reclassify the species from endangered to threatened in 2007 (Federal Register 72; USFWS, 2007).
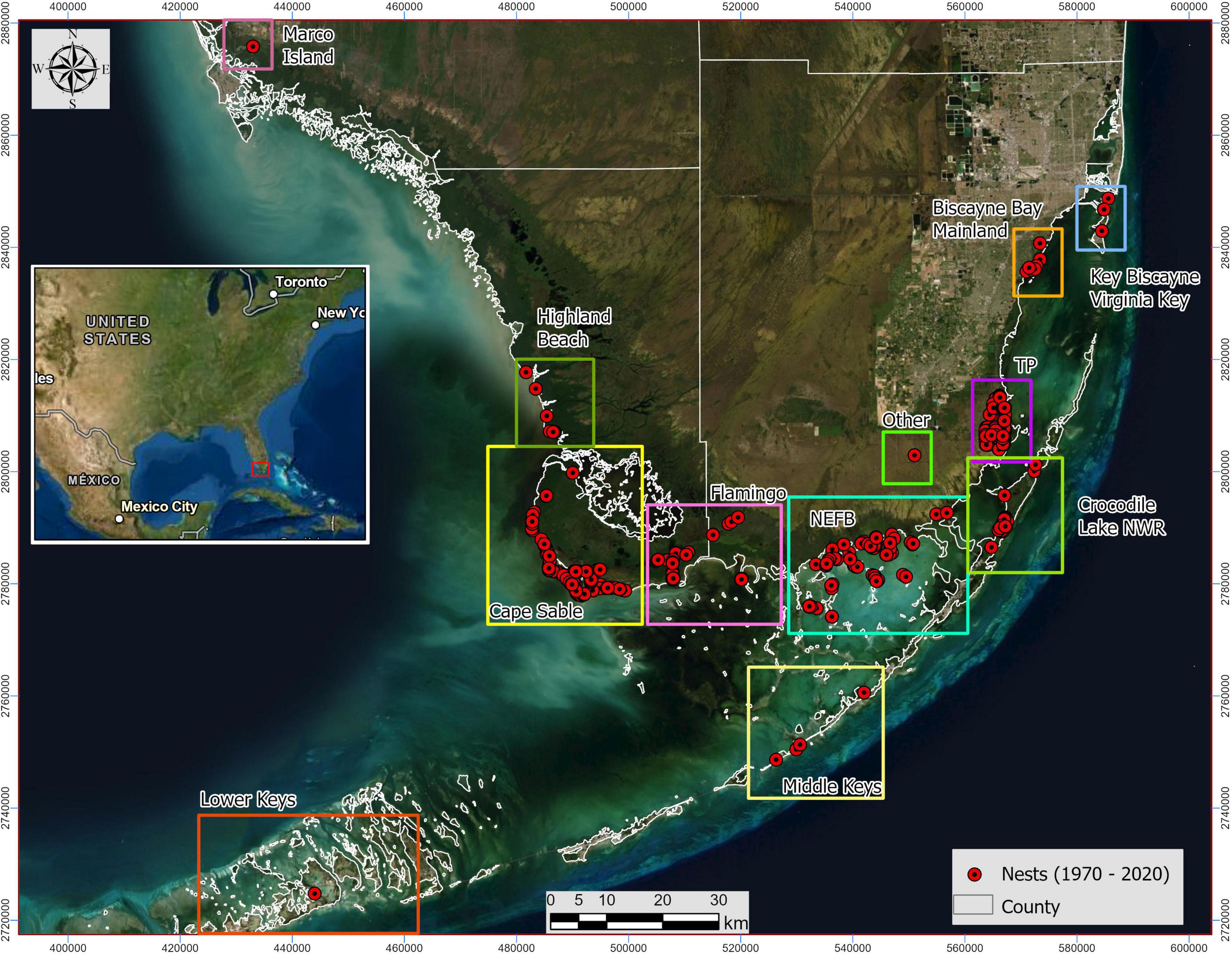
Figure 1. Main nesting areas of American crocodiles (Crocodylus acutus) identified from 1970 through 2020 across South Florida, United States. The “Other” area refers to C-110 and MM113 Overseas Highway Southbound. Notice the most historically representative areas are within the Everglades National Park (ENP; Cape Sable, Flamingo, and Northeast Florida Bay –NEFB), Crocodile Lake National Wildlife Refuge (NWR), and Turkey Point Power Plant (TP)). However, areas around the Keys, the western coast (Marco Island and Broad River) and the eastern part of South Florida (Biscayne Bay and Virginia Key) are also nesting areas used by American crocodiles across this 50-year study.
Nesting by American crocodiles in Florida historically has been along coastal areas where nearby freshwater or brackish habitats with low salinity and low wind and wave action provided critical nursery conditions for hatchlings (Mazzotti, 1983). In Florida, courtship is likely to occur in late January into February (Kushlan and Mazzotti, 1989b) and nesting season is from March to August, including nest preparation from March to May and egg laying in April and May (Mazzotti, 1989). The incubation period is around 90 days and is temperature dependent (Mazzotti, 1989; Thorbjarnarson, 1989). Hatching generally occurs in July and August (Mazzotti, 1989) though hatchlings have also been found in September (see Briggs-Gonzalez et al., 2021). More recently, Cherkiss et al. (2020), found that hatching is occurring earlier in Florida (late June) over a 37-year nesting period illustrating that increasing temperature could be influencing earlier hatching.
American crocodiles in Florida make two types of nests: a mound nest created when a crocodile first creates a mound and then digs a hole on top of the mound to deposit eggs, and a hole nest when a crocodile deposits eggs in a chamber below ground-level (Mazzotti, 1989). The former nests are most frequently associated with sandy beach sites and the latter with nests made on marl creek banks or on artificial surfaces such as canal banks (Mazzotti, 1989). Crocodiles nest in available substrate, for example sand, marl, or peat, and do not incorporate other nesting materials such as vegetation, into their nests, except incidentally (Mazzotti, 1989).
Mazzotti (1989) suggested that six major factors can adversely affect nesting success (percentage of nests that produce at least one hatchling) and hatching success (percentage of eggs in successful nests that produce hatchlings) of crocodylians: fertility, predation, extreme temperatures, moisture conditions, erosion of nest sites, and human disturbance to nest sites or breeding animals. The success of these two parameters (nesting and hatching) depends on when, where, and how females deposit eggs into nests and the environmental conditions present during egg incubation (Mazzotti et al., 1988; Mazzotti, 1989).
The aim of this study was to assess natural and anthropogenic factors influencing the nesting ecology (number of nests, nest morphology, success rate, and habitat use) of American crocodiles in Florida using long-term monitoring data (1970-2020). We focused on understanding the natural history of nesting across space and time as well as the ecological and evolutionary responses of C. acutus to anthropogenic loss and creation of habitat, and to changes in temperature and precipitation across South Florida. We used the theory of orientation ecology (Jander, 1975) to predict that spatial and temporal orientation of nesting behavior will maximize distance from stress sources and minimize distance from resources in both space and time, and importantly we recognized that tradeoffs and bet-hedging (Hairston and Fox, 2013; Rajon et al., 2014) may occur. We illustrate species adaptative capacity (Angeler et al., 2019) and population responses to ecosystem restoration efforts in Florida with nest numbers and nesting conditions.
The study was conducted at the southern tip of mainland Florida from northern Biscayne Bay/Virginia Key, NEFB, and Cape Sable northwest through Marco Island and south into the Florida Keys, covering a total of 12 main areas (Figure 1 and Table 1). We conducted surveys for crocodile nests by motorboat, jon boat, canoe, helicopter, and on foot of known and potential nesting habitat during April and May (egg laying period) and monitored historical (known) and potential nesting habitat from June through August (hatching period). We noted nesting activity (tail drags, digging, or scraping), the presence of eggs or hatchlings, and the number and causes of egg failure in both successful and failed nests whenever possible. We categorized nests as hole or mound construction (Mazzotti, 1989) and either isolated, near other nests, or within the same mound (see Cherkiss et al., 2020). Hatched nests were identified by the presence of an open hole, evidence of digging, or hatched shells. Shells of hatched eggs or hatchlings located at an open nest were considered evidence of successful nests and a nest was considered successful if at least one hatchling was produced. Hatching success could not be determined because some females moved some or all of the eggshells (hatched and unhatched) and hatchlings from the nest site. Once a nest had completed hatching, the egg chamber was inspected for any remaining unhatched eggs which were then counted; this process ensured that multiple nests in proximity that hatch in succession were not confused with each other. We defined failed nests when the nest passed hatching time (August) and no hatchlings emerged from it, and a depredated nest when we found any indication that at least one egg had been depredated. We also classified nest as “not determined” when no information could be recovered from the nest that allowed us to assign it to any of the categories described above. Hatch dates for all known nests were determined when possible. We recorded date, time, location (on a map or measured by global positioning system, GPS), and habitat type (categories = shoreline beach, pond, lagoon, creek, cove, roadside, and canal) of nests.
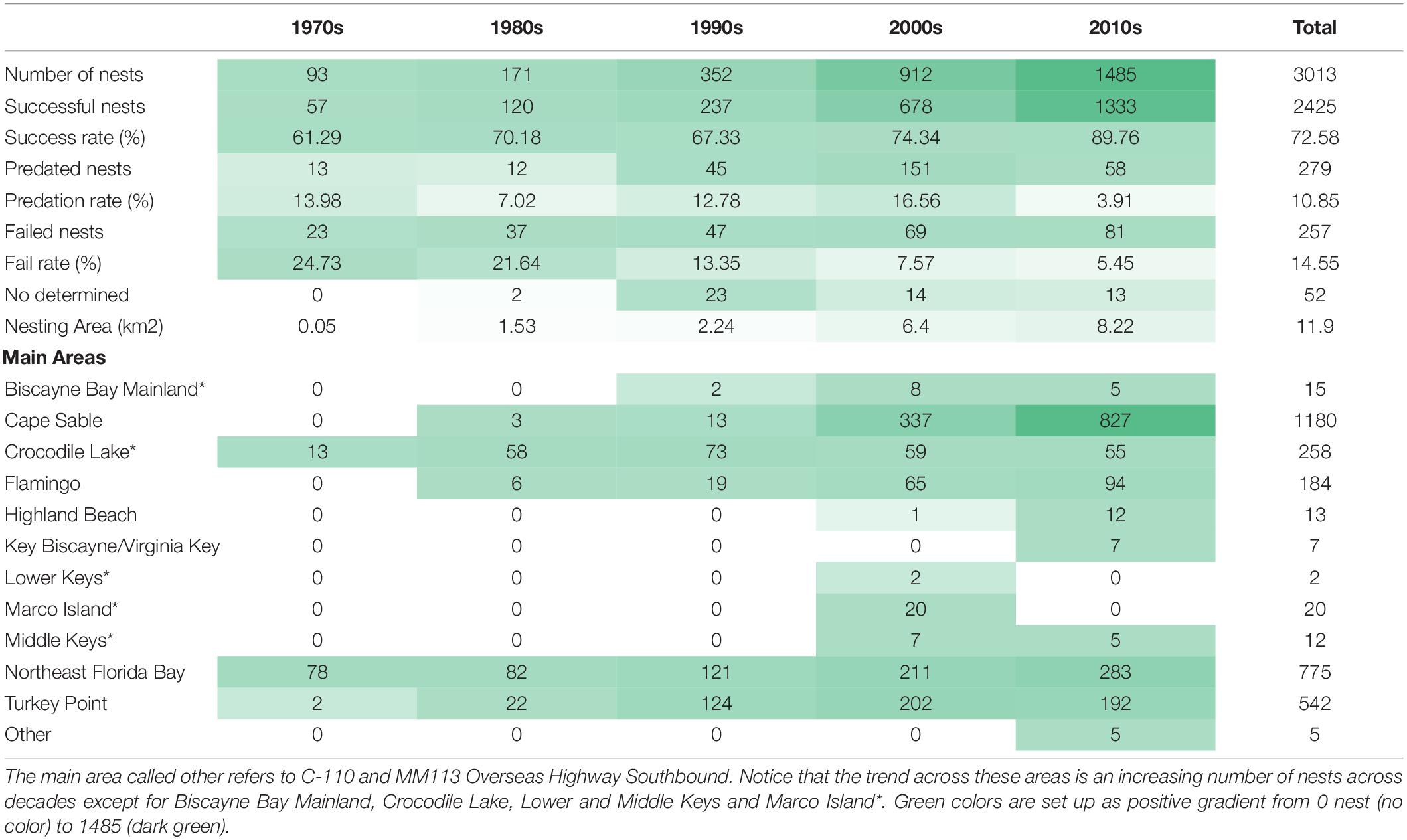
Table 1. American crocodile (Crocodylus acutus) nesting summary recorded across the 12 main areas evaluated in South Florida from 1970 to 2020 analyzed by decades.
We estimated the area used by American crocodiles for nesting in Florida based on a 100-m buffer calculated around nest geolocation (incident point data- IPD) and dissolved by year via ArcGIS Pro 2.9.1 (ESRI, 2021). We also assessed habitat type of nesting areas based on both first-hand observation from the field and the cooperative land cover map version 3.4 compiled by Florida Fish and Wildlife Conservation Commission (FWC) and Florida Natural Areas Inventory (FNAI, Kawula and Redner, 2018). We calculated success, predation, and failure rates of nest based on the number of successful, predated, or failed nests divided by the total number of nest per decade.
We performed an optimized hot spot analysis (OHSA) based on IPD by decade and the whole dataset (50-year record) classifying areas based on statistical evidence [strong (p-value < 0.01), moderate (< 0.05), weak (< 0.10) or little-to-no evidence (> 0.10; Muff et al., 2022)] supporting high and low nest aggregations (hot and cold spots) using the Getis-Ord Gi statistic. For this analysis, IPD were aggregated and counted (weighted) based on the buffer layer previously calculated by decade, estimating the average nearest neighbor, and identifying via incremental spatial autocorrelation –Global Moran’s I statistic method, the appropriate scale of analysis. Finally, we defined statistical evidence for IPD weighted based on the false discovery rate correction for multiple testing and spatial dependence (ESRI, 2021).
We analyzed minimum (min temp) and maximum (max temp) temperature as well as precipitation changes across nesting areas from 1970 through 2018 (due to environmental data availability) and the potential effect of these parameters in the number of nests found by year based on historical monthly weather data developed by Harris et al. (2014) downscaled to 2.5 min (∼ 21 km2) with WorldClim 2.1 (Fick and Hijmans, 2017). Environmental values were extracted monthly using coordinates from nest per year via ArcGIS Pro and analyzed in R version 4.0.3 (R Core Team, 2020) to assess trends across time. We included in this analysis data from spring (March, April, and May) and summer (June, July, and August) as the period where the nesting process occurs (from laying eggs to hatching) in South Florida (Mazzotti, 1989). We assessed relationships between environmental parameters and years and environmental parameters and number of nests reported by year via linear regression models using the lm function in R reporting 2.5 and 97.5 % confidence intervals (CI).
A total of 3,013 American crocodile nests were recorded in Florida from 1970 to 2020 with an overall increasing trend in number of nests recorded by decade (from 93 up to 1,485) as well as area used (100-m buffer around nest dissolved changed from 0.05 km2 up to 11.9 km2; Table 1 and Figure 1). Nest success rate also showed an increasing trend from 61.3% in the 1970s up to 89.8% in the 2010s with an exception in the 1990’s (67.3%). Most unsuccessful nests were either depredated (N = 279) or failed by unknown causes (N = 257). Failure rate showed a defined pattern across time with higher values in the 1970s and lower in the 2010s (24.7 and 5.5%, respectively). In contrast, predation rate showed a more variable pattern with the highest in the 2000s (16.6%) and the lowest in the 2010s (3.9%).
Nests recorded from the early 1970’s came from NEFB (Figures 1, 2). New nesting areas were recorded in early 1970’s [Crocodile Lake National Wildlife Refuge (Crocodile Lake NWR)], 1978 TP], 1986 (Flamingo and Cape Sable), 1997 (Biscayne Bay Mainland), 2000 (Marco Island), 2002 (Middle Keys), 2006 (Highland Beach), 2008 (Lower Keys), 2012 (referred as other in the main areas: C-110 and MM113 Overseas Highway Southbound), and 2015 (Key Biscayne/Virginia Key). From those, Cape Sable and NEFB are the areas with the largest number of nests reported (1,180 and 775, respectively) followed by TP, Crocodile Lake NWR, and Flamingo (542, 258, and 184, respectively). Seven out of the 12 areas assessed showed an increasing trend in the number of nests reported by decade whereas areas such as Biscayne Bay Mainland, Crocodile Lake NWR, Lower and Middle Keys, and Marco Island showed a decreasing trend in the last two decades (Table 1). Most nests recorded in South Florida where nest type was described were hole nests (N = 1,008) but at least one third of the total historically recorded nests with a known type have been mound nests (N = 573). Northeastern Florida Bay is the area with the greatest number of mound nests historically recorded (N = 394) and Flamingo/Cape Sable and Crocodile Lake NWR are the areas with the greatest number of hole nests (N = 591 and 246, respectively). Mound nests predominate along the sandy beaches found in northeastern Florida Bay, whereas hole nests predominate on peat and marl anthropogenic substrates found along canals at Crocodile Lake NWR or in the Flamingo/Cape Sable area.
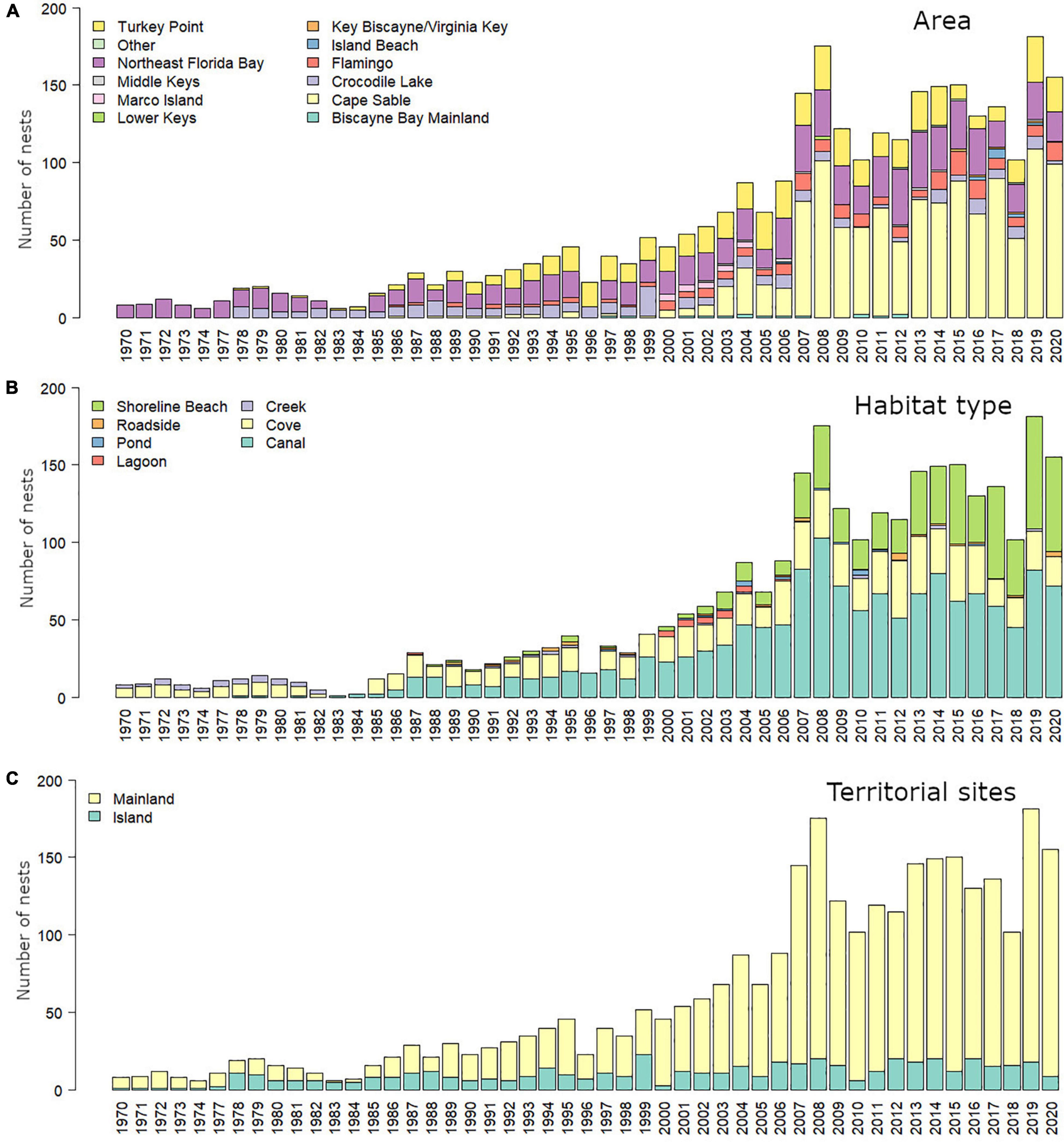
Figure 2. Number of American crocodile (Crocodylus acutus) nests reported in South Florida, United States, from1970 to 2020. Nesting has been increasing across main areas (A) and habitat types (B) including manmade habitats. There has also been a greater increase in nest on mainland sites than island sites (C) likely related with space availability (see discussion).
Nest have been consistently found in natural habitats such as coves, creeks, shoreline beaches, lagoons, and ponds across South Florida (N = 1,459; Figure 2). However, anthropogenic habitats such as canals have also played an important role in American crocodile nesting steadily increasing since recorded in 1978 at Crocodile Lake NWR and TP, representing almost half of the total nests reported across the last four decades (N = 1,406; Figure 2). Most nests have been reported in mainland (N = 2,510) rather than insular (N = 503) areas except in 1983 and 1984 where we reported larger number of nests on insular areas. Nests have been found across 22 types of habitats based on the Florida land cover classification system (Kawula and Redner, 2018), from which mangrove swamp (33.85%), cultural – estuarine (estuarine ditch/channel; 28.58%), marine (17.46%), and estuarine (4.50%) habitats were the most common used for nesting by American crocodiles across South Florida. Assuming no major land cover changes on the nesting areas where we found nests in the last 50 years, we observed an increasing number of habitat types being used by American crocodiles from the 1970s [mangrove swamp, marine, cultural – lacustrine, salt marsh, and tidal flat) to the 2010s (22 total, Table 2)]. Natural habitats (mangrove swamp – 48.53% and marine – 31.98%) were the most relevant in the 1970s. By the 1980s, manmade habitats such as cultural – lacustrine (27.57%; artificial impoundment/reservoir and industrial cooling pond) peak as important areas for nesting and by the 1990s this habitat turned out to be the most used for nesting (42.87%). After 2000, mangrove swamp (2000s – 32.72%, 2010s – 36.14%) and cultural – lacustrine (2000s – 30.98, 2010s – 21.21%) have been the most relevant habitats for nesting across South Florida.
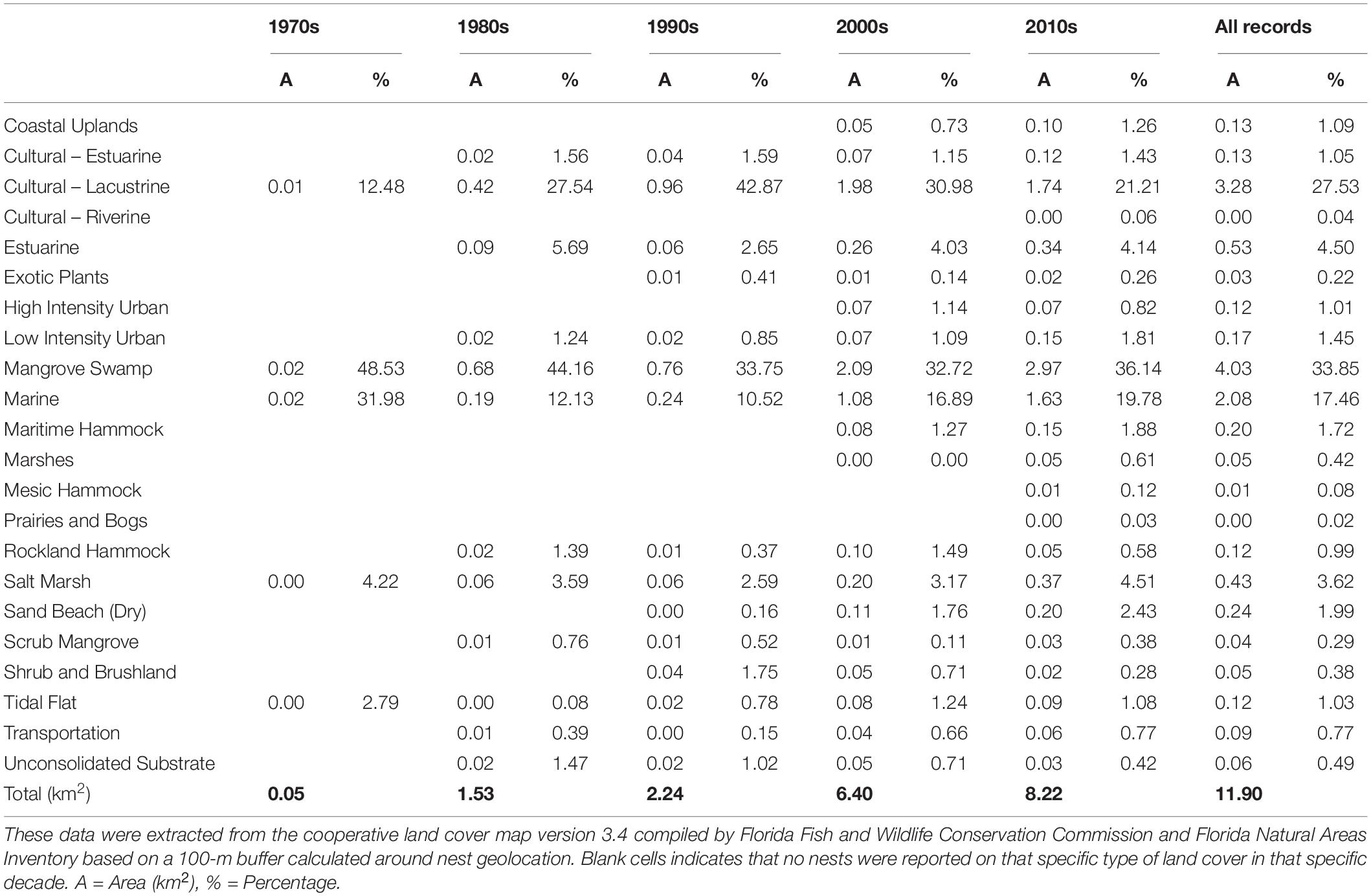
Table 2. Types of habitats found across American crocodile (Crocodylus acutus) nesting areas in South Florida based on the Florida land cover classification system (Kawula and Redner, 2018).
Optimized hot spot analysis defined a total of 2,887 valid input features (nests with valid coordinates) based on the IPD and the buffer layer, from which 2,882 weighted IPD showed strong evidence of no spatial dependence (Figure 3). Incremental spatial autocorrelation analysis based on a peak clustering model defined an appropriate scale (distance band) for spatial analysis that ranges between 1.46 km for 2010s and 12.55 km for 1990s (Table 3). Northeastern Florida Bay and TP showed strong evidence for high and low values of nest spatial clusters (hot and cold spots, respectively) in the 1970s and 1980s due to a high number of nests and little variation in the former area, and lower but constant number of nests in the latter area. Crocodile Lake NWR in the 1980s and Cape Sable and Flamingo in the 1990s showed also strong evidence of nest spatial clusters although in low numbers (cold spots). By the 2000s, Cape Sable showed strong statistical evidence of high values of nest spatial clusters (hot spot) along with NEFB, with constant presence of reproductively active American crocodiles and a higher number of nests. Areas such as Biscayne Bay Mainland, Highland Beach, Middle keys, Marco Island, and Key Biscayne/Virginia Key, showed strong evidence of low values of nest spatial clusters after 2000s and are currently defined as cold spot nesting areas (Figure 3). In contrast, we found little to no evidence for hot/cold spot areas such as the lower Keys (i.e., Gi Z-score = -1.05, p-value = 0.29), such that no patterns could be identified in the number of nests registered and could be considered sporadic nesting areas.
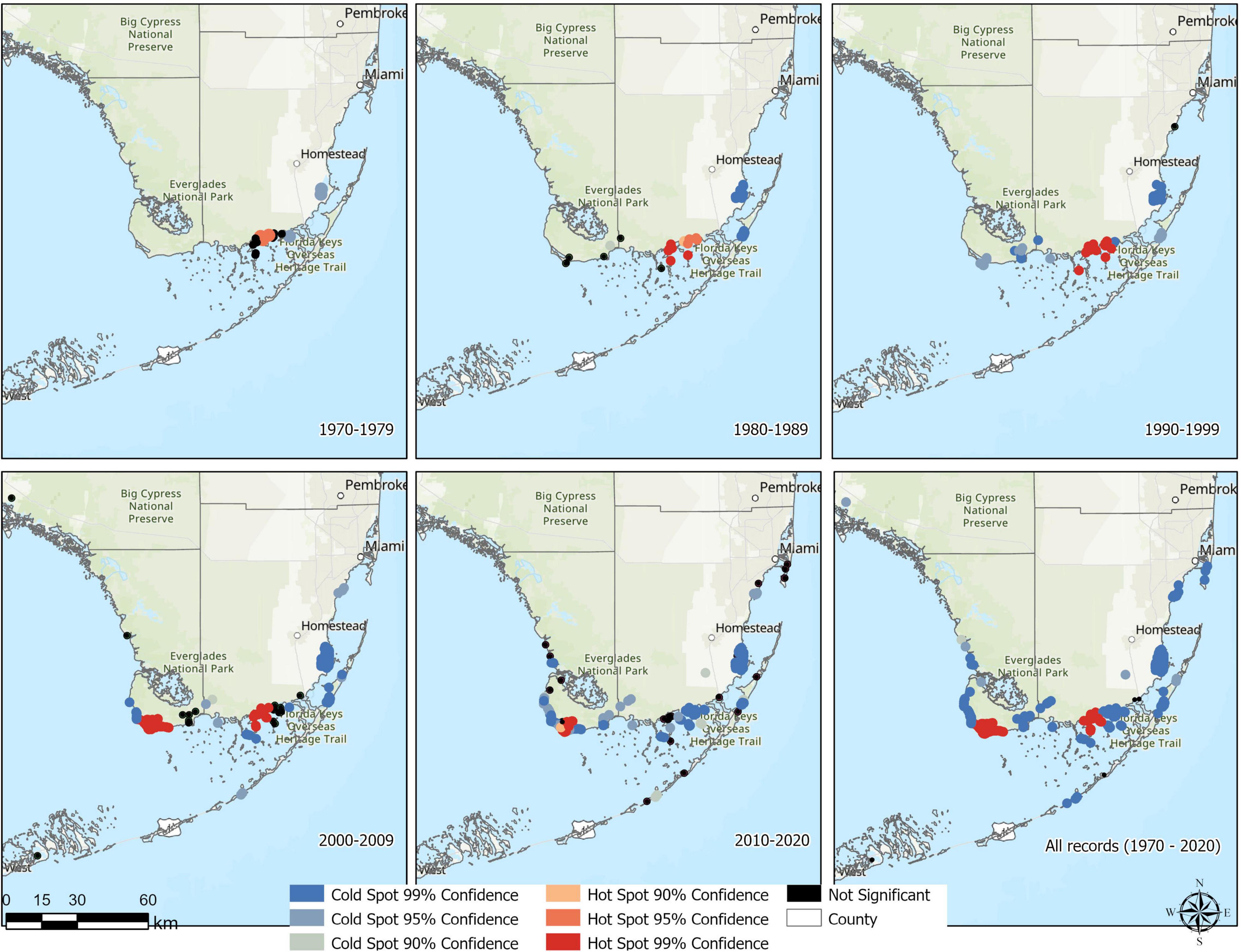
Figure 3. Optimized hot spot analysis of American crocodile (Crocodylus acutus) nests registered across South Florida, United States from 1970 to 2020. Red dots represent hot spot areas with different levels of evidence (strong ≤ 99 %, moderate ≤ 95 %, and weak ≤ 90 %) where high incident counts (high and repeated number of nests) cluster based on a specific distance band (see Table 3 for details). In contrast, blue-grayish dots represent cold areas with different levels of evidence where low incident counts (low but repeated number of nests) cluster based on a specific distance band.
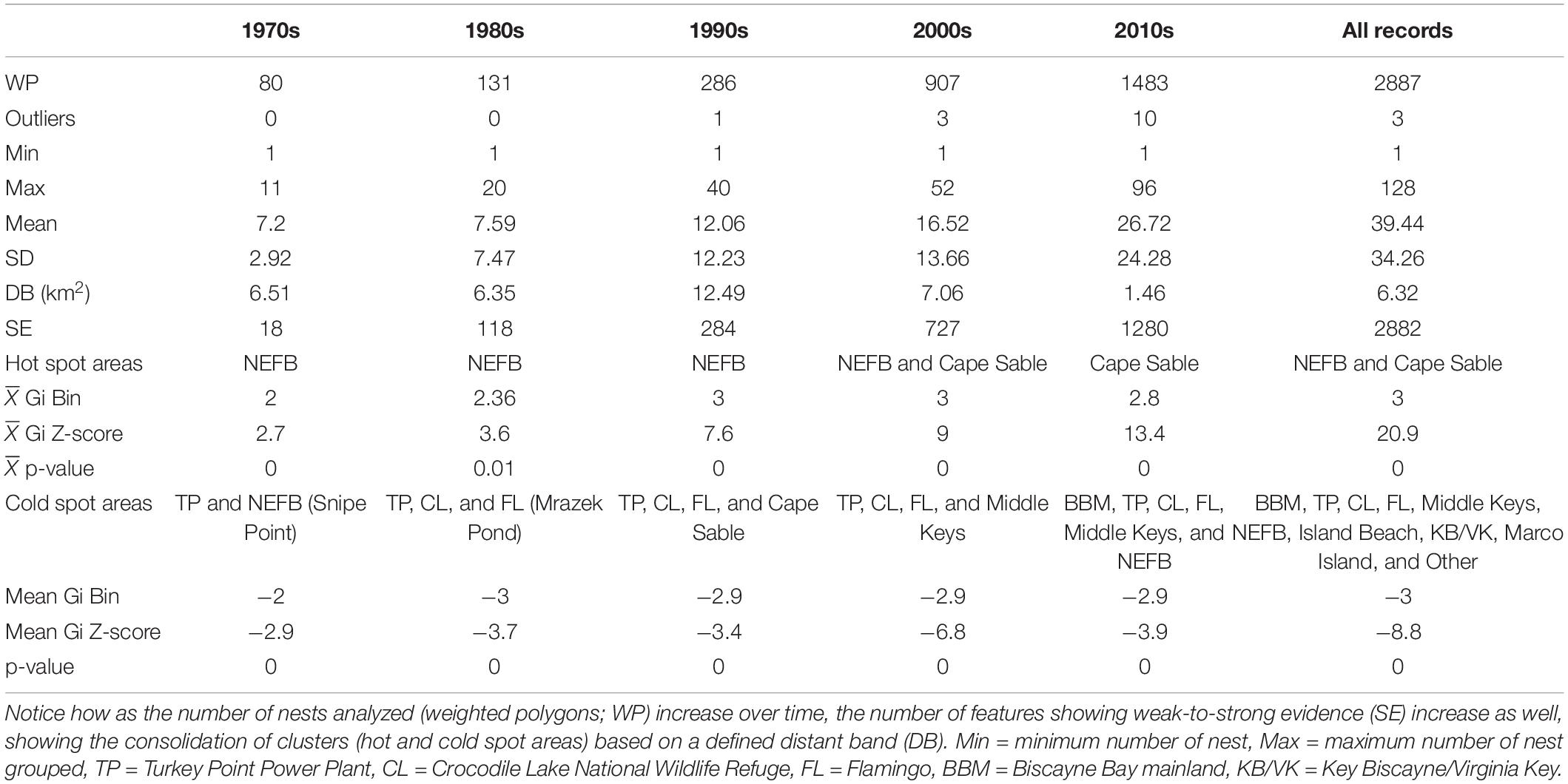
Table 3. Results of the optimized hot spot analysis (OHSA) of American crocodile (Crocodylus acutus) nests identified across South Florida from 1970 to 2020.
Environmental parameters in nesting areas across South Florida ranged between 42.70 ± 23.21 mm (March) and 228.0 ± 74.0 mm (June) for precipitation, 17.9 ± 1.51°C (March) and 25.0 ± 0.45°C (August) for min temp, and 26.4 ± 1.27°C (March) and 32.6 ± 0.43°C (August) for max temp from 1970 through 2018. Data revealed weak-to-strong statistical evidence that these environmental parameters are positively associate with time in South Florida (Figure 4) except for mean spring (March, April, and May) precipitation where no evidence for a linear relationship was found (p-value = 0.79). Min and max temp in summer (June, July, and August) were the most relevant variables explaining at least 38% and 24% of the total variation of the number of nests. This trend was also identified when assessing the relationship between environmental parameters individually and number of nests reported, showing weak-to-strong evidence for a positive effect of environmental parameters on the number of nests over time and by season (spring and summer) except for mean precipitation in spring where we found little-to-no evidence (p-value = 0.86; Figure 5). Again, min and max temp in summer were the most relevant variables explaining at least 35% and 16% of the total variation of the number of nests reported in South Florida. However, a multiple linear regression analysis including all parameters showed that min temp (slope = 39.04, CI = 3.56 – 64.52, p-value = 0.004) in combination with precipitation (slope = 0.20, CI = -0.04 – 0.43, p-value = 0.094) in summer are the most relevant variables to describe the variation in the number of nests reported across South Florida (F = 6.926, df = 40, p-value < 0.001, adjusted r2 = 0.44).
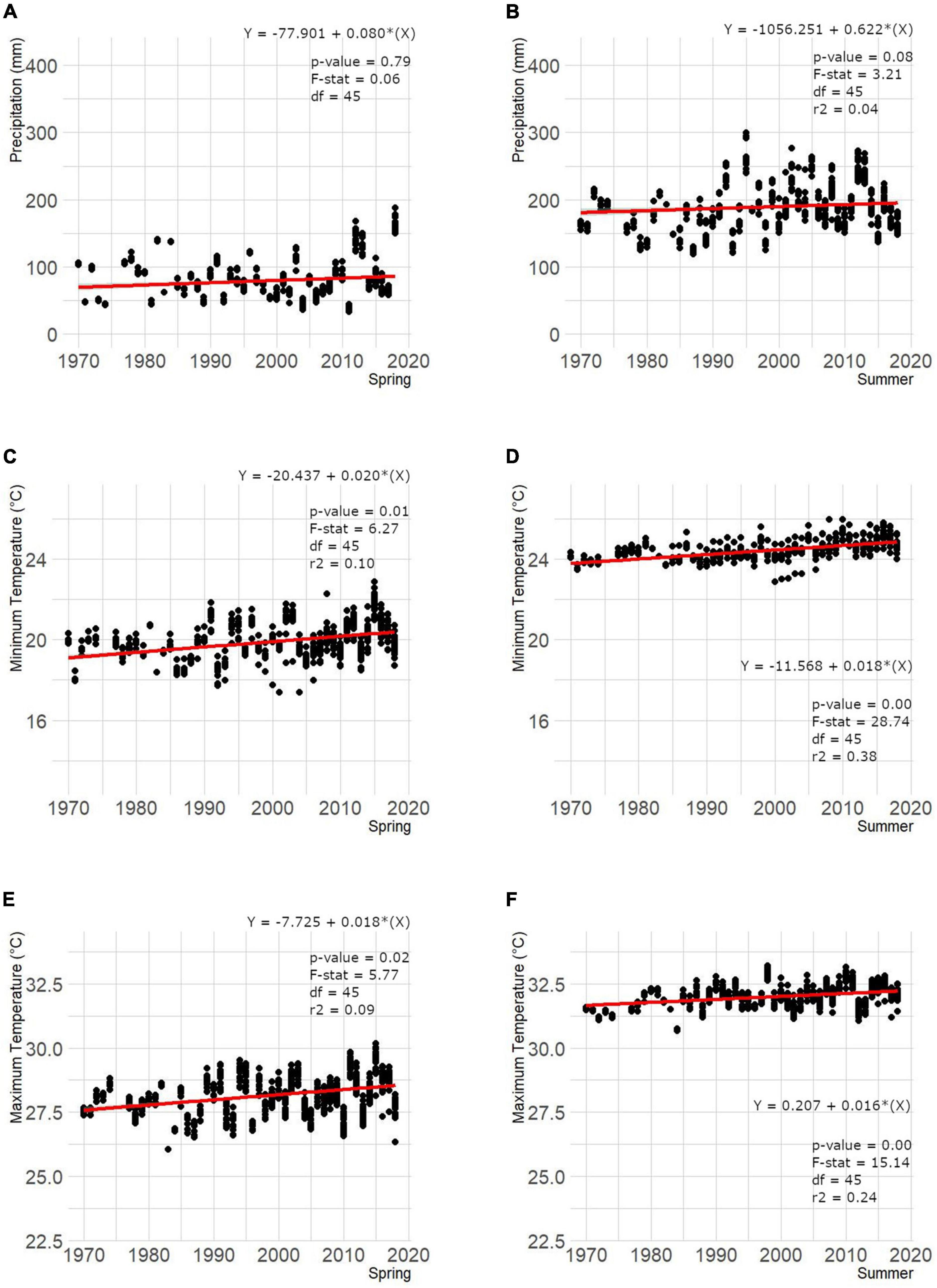
Figure 4. Mean precipitation (A,B), minimum (C,D) and maximum (E,F) temperature by season [spring (March, April, and May) and summer (June, July, and August)] from 1970 through 2018 reported across American crocodile (Crocodylus acutus) nesting areas in South Florida, United States. There is overall weak-to-strong evidence for a positive trend in all variables across time except for mean spring precipitation.
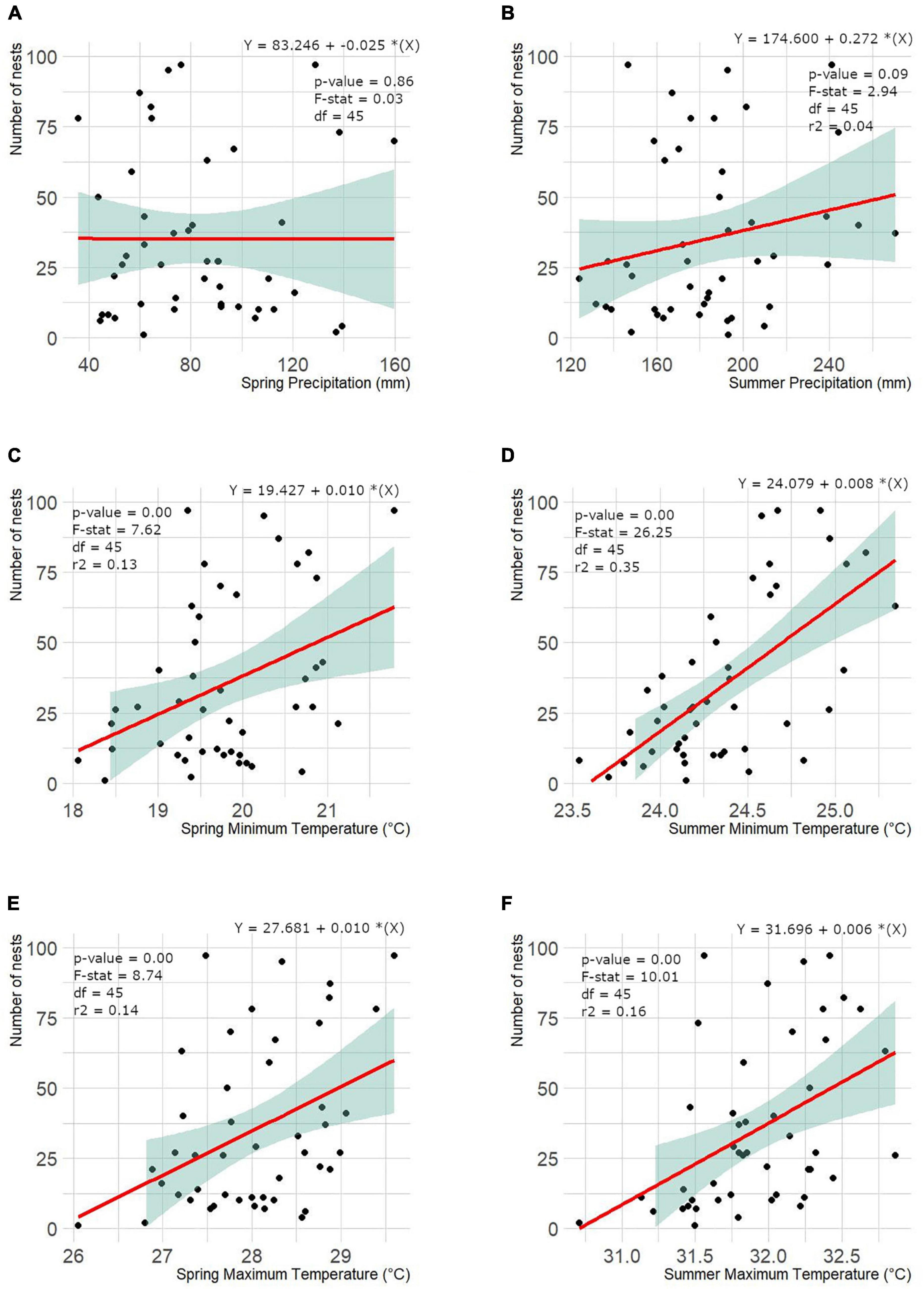
Figure 5. Linear relationship between the number of nests found by year and the mean precipitation (A,B), minimum (C,D) and maximum (E,F) temperature reported across South Florida from 1970 through 2018. Solid red line represents the linear model predicted and the shaded area represents the standard error of the model. All relationships showed weak-to-strong evidence for positive effect of environmental parameters on number of nests except for spring precipitation and number of nests.
The increase in numbers and success of American crocodile nests in South Florida is attributable to recovery of a once endangered species (Mazzotti et al., 2007b), successful adaptation to a seasonable and variable environment (Mazzotti, 1989; Cherkiss et al., 2014), and underscores the value of providing, protecting, and restoring habitat to support recovery of listed species. In addition to protected crocodile nesting habitat that was provided in Everglades National Park, the U.S. Fish and Wildlife Service purchased Crocodile Lake National Wildlife Refuge to protect crocodile nesting habitat that had been created as a result of an illegal dredge and fill operation, and Florida Power and Light Co committed to managing American crocodiles at their Turkey Point Power Plant site in southern Miami-Dade County as part of its permitting and licensing process (Brandt et al., 1995). The rapid increase in nesting of American crocodiles during 1986 to 2008 is correlated with the plugging of canals in the Cape Sable area (Mazzotti et al., 2007b). We have no explanation, or even a good speculation, for why the increase in nesting leveled off during 2010 to 2020 as it does not appear that nesting or juvenile habitat had become saturated in the Cape Sable area.
Cape Sable has become a nesting hot spot (Figure 3) and the most important location for successful crocodile nests in Florida. The occurrence of new nests in a new area was primary impetus for reclassifying the American crocodiles from endangered to threatened (Mazzotti et al., 2007b). Increases in both numbers of nests and areas where nesting occurs were associated with crocodiles nesting on artificial substrates, especially along levees and berms of canals and ditches, which were dug for drainage, navigation, or cooling purposes (Mazzotti et al., 2007a; Figure 2). For instance, the expansion of crocodile nesting to the Flamingo/Cape Sable area in the mid 1980’s concurrent with plugging of Buttonwood and East Cape canals, provided new locations as well as lower salinities in interior wetlands leading to increased growth and survival of hatchling crocodiles (Mazzotti et al., 2007a,2009; Briggs-Gonzalez et al., 2021). That crocodiles found and nested in these new locations and habitats illustrates the dispersal and wandering instincts of this species (Kushlan and Mazzotti, 1989b; Cherkiss et al., 2014) and crocodylians in general (Read et al., 2007; Campbell et al., 2013; Calverley and Downs, 2015) as well as the willingness to nest in any diggable substrate at suitable elevation.
More crocodiles nesting in more places at least partially accounts for increase in habitats where crocodile nests occur (Table 2) and demonstrates that different substrates (e.g., marl, peat, sand) can provide suitable conditions for incubation of eggs and nest success (Swanepoel et al., 2000). However, impacts of different nesting substrates on temperature and moisture conditions of nest environments on sex, growth, and survival of hatchling American crocodiles remains unknown (Charruau, 2012).
Although substrate type varied, all nest sites were on elevated surfaces. It is common for crocodylian species to nest on drier, elevated surfaces (Metzen, 1977; Thorbjarnarson and Hernández, 1993; Platt and Thorbjarnarson, 2000; Platt et al., 2008) to avoid flooding conditions. However, nests at elevated sites, especially linear ones such as riverbanks, berms, levees, and beaches are at increased risk of predation (Metzen, 1977; Magnusson, 1982; Hall and Johnson, 1987; Platt et al., 2008). That American crocodiles can quickly find and use elevated sites is best illustrated by that a crocodile nested on a canal plug within hours of construction completion (Mazzotti et al., 2007a).
Nesting habitat on artificial substrate (peat deposited from dredging canals) at Crocodile Lake NWR has diminished in quality due to erosion and subsidence, and that the artificial peat substrate does not regenerate naturally (Mazzotti per. obs.). This is likely why Crocodile Lake NWR has exhibited a cooling trend for number of nests in the last decades (Figure 3). Follow up research on these aspects could help to elucidate the overall effect of artificial substrates on American crocodile nesting and the long-term effect on populations.
There are two components to the increasing trend in successful crocodile nests that require further consideration: that both the rates of failure of nests and rate of predation decreased (Table 1). Failure of American crocodile nests in Florida is primarily caused by flooding or desiccation which occurs in very wet or very dry years (Mazzotti et al., 1988; Mazzotti, 1989). Both flooding and desiccation are cryptic factors in American crocodile nests, and unless you know a nest was there, it would not be counted as a nest at all. We hypothesize that the decrease in failed nests could be a result of an increase in failure to find failed nests in the last decades, possibly due to the high number and spatial spread of nests checked every year, rather than a decrease in the rate of nests failing. An alternative hypothesis is that the success rate of nests is increasing concurrent with increasing summer rainfall and minimum summer temperature. In this study, the combination of summer rainfall and minimum temperature explained most of the variation in number of successful nests found and as described below, crocodile nesting behavior results in crocodiles avoiding cool, dry spring conditions. Similarly, nesting of alligators in the Everglades (Kushlan and Jacobsen, 1990) and Morelet’s crocodile in Belize (Platt et al., 2008) is timed to avoid dry conditions during nesting.
Predation rate was variable through the 2000s and then declined in the 2010s, which interestingly relates with the diminished numbers of raccoons (Procyon lotor) across ENP and the concurrent increase in Burmese pythons (Python bivittatus; Dorcas et al., 2012). Raccoons have been documented as the primary predator of crocodile nests as well as other beach-nesting species such as marine turtles (Deitz and Hines, 1980; Mazzotti, 1989; Garmestani and Percival, 2005; Hénaut and Charruau, 2012; Wilson, 2017). We hypothesize that a decreasing population of raccoons influenced the predation rate historically recorded. This relationship (decreasing nest predation rate with increasing number of pythons) has been documented for turtles in southern Florida (Wilson, 2017).
American crocodiles have a suite of adaptations for orientation of nesting in time and space that help explain the rate of success of crocodile nests. These adaptations provide opportunities for successful nesting in an environment that is both seasonal and variable (Mazzotti and Brandt, 1994). For instance, South Florida is characterized by a warm wet season (May to October) followed by a cool dry season (November to April). However, conditions within a season can be variable (wet seasons can be extremely dry causing desiccating conditions in nests, or extremely wet, causing flooding). Both, desiccation and flooding, cause mortality of eggs, delay incubation, or result in deformed hatchlings (Mazzotti et al., 1988; Mazzotti, 1989). When and where crocodiles nest maximizes the distance in space and time from these sources of stress and minimizes the distance from resources such as suitable temperature and moisture conditions. Tradeoffs occur since not all resources or stress sources occur in the same place or at the same time and that sometimes a stress source and resource can occur together.
This adaptive capacity can be seen in NEFB where crocodile nests are located on sandy beaches surrounded by shoreline vegetation comprised primarily of red, black, and white mangrove (Rhizophora mangle, Avicennia germinans, and Laguncularia racemosa) interspersed with tropical hardwoods such as mahogany (Swietenia mahagoni) and gumbo limbo (Bursera simaruba; Olmsted et al., 1981), providing the well-drained elevated sites preferred by female crocodiles for their nest (Mazzotti, 1989; Mazzotti et al., 2007a). However, beach sites are in areas of higher salinity (> 30 psu) and greater exposure to wind and wave action which are stressful to crocodiles especially for hatchlings. Beach sites can also be further from nursery habitat and the need to travel further to protected areas could be an additional stress (Mazzotti, 1983, 1989; Mazzotti et al., 1986). In this case female crocodiles tradeoff putting eggs at a location with good conditions for incubation but stressful conditions for adult and hatchling crocodiles with little parental care of hatchlings evident, resulting in quick dispersal from nest sites.
Nesting behavior of American crocodiles in Florida exhibits a bet-hedging strategy which was defined by Hairston and Fox (2013) “as a trait of an organism, living in a variable environment, that provides greater net fitness over a range of environmental conditions than would a trait specialized for any single environment.” In this case, the variable environment for American crocodiles is the variation in moisture and temperature conditions during incubation. In natural habitats, American crocodiles hedge their nesting bets by nesting in sand mound nests along beaches that are resistant to flooding but vulnerable to desiccation, and by nesting in marl hole nests which are resistant to desiccation but prone to flooding (Mazzotti et al., 1988; Mazzotti, 1989). This potentially guarantees that some nests will hatch even under extreme conditions. Climate change and sea level rise have the potential to impact crocodile nesting by changing the temperature during nesting and by flooding nests on low lying areas. However, manmade areas can play a role in securing some nest sites. Thus, whereas relatively low-lying nests on beaches and creek banks are vulnerable to flooding from sea level rise, nests on higher elevated sites along canals should prove less affected.
Timing of nesting is such that incubation is bracketed by cool-dry spring conditions not conducive to development of eggs and warm-wet conditions of late summer that can cause hatchling deformation and mortality of eggs (Mazzotti, 1989). As a result of cool spring temperatures, egg laying for American crocodiles in Florida occurs later than in other parts of its range (Charruau, 2012; Balaguera-Reina et al., 2015). The predictable nature of spring temperatures as a cue for crocodile nesting leads to a pulse nesting pattern (nests hatch within a few weeks of each other) rather than an extended nesting period as exemplified by other crocodylian species (Grigg and Kirshner, 2015). We hypothesize that pulse versus extended nesting periods is an adaptation to the predictability of seasonal patterns of temperature and rainfall. We predict that pulse nesting should occur when seasonal patterns of temperature and rainfall are predictable as for example in alligators (Kushlan and Jacobsen, 1990; Hall, 1991) and Morelet’s crocodiles (Crocodylus moreletii) (Platt et al., 2008). Why hedge your bet when you have a sure thing? In contrast, we predict that extended nesting should occur when suitable conditions for nesting are not predictable temporally as for example for Crocodylus porosus (Magnusson, 1982) or for Crocodylus novaeguineae (Hall, 1991). In support of this, increased flooding of alligator nests in Everglades National Park was associated with changes in the predictable natural hydro-pattern caused by water management (Kushlan and Jacobsen, 1990).
Location of nests plays an important role in determination of incubation temperature (Maciejewski, 2006) and incubation temperature plays an important role in sex determination and survival of embryos (Charruau, 2012). Crocodiles have some plasticity in terms of where they deposit their nests with respect to distance from water, depth of the clutch, and whether the nest is in sun or shade (Platt and Thorbjarnarson, 2000; Charruau, 2012; Balaguera-Reina et al., 2015). However, this behavioral plasticity may not be enough to mitigate increases in temperature brought about by climate change (Murray et al., 2016). The only study done on nest temperatures in Florida was performed on sand beach nests and marl creek nests in northeastern Florida Bay by Lutz and Dunbar-Cooper (1984). Nothing is known about nest temperatures in anthropogenic substrates or about how nest temperatures may have changed over time. Since we cannot determine sex of hatchlings reliably, we also have no information on any changes in sex determination that may have occurred.
Cherkiss et al. (2020) found that the timing of nesting of American crocodiles in Florida has shifted in response to increasing temperatures which could have potential implications for the timing pattern displayed until now, showing that the variety of behaviors in timing and location of nests can mitigate some of the adverse effects of climate change. Nonetheless, we hypothesize that there is a limit to how much American crocodiles can shift the incubation period as conditions earlier in the season may be too dry to support incubation of eggs.
Overall, nesting ecology of American crocodiles in Florida has proven to be both positively and negatively influenced by anthropogenic factors, from the unintentional construction of nesting habitats through canals and ditches, and the introduction of invasive species that potentially reduced predation pressure on nests and hatchlings, to the unknown potential effects these events can have on the future of the community (i.e., diversity loss, bottom-up trophic effects). The bet-hedging strategy American crocodiles exhibit in Florida gives the species potential advantages to face the rapid (in evolutionary terms) changes in climate and landscape in the last 50 years, which can be seen in the increasing number of nests (and adults) reported in the last decades. However, it does not imply that the adaptive capacity of the species to face these changes (evolutionary potential) cannot reach a limit if changes continue. Careful consideration of the effects of climate change and sea level rise at local scales (i.e., temperature and moisture variation across nests, landscape modification, and prey availability) in the long term can help us further understand impacts on nesting ecology.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
The animal study was reviewed and approved by University of Florida IACUC # 202109072.
FM: initiated project, designed study, acquisition of funding, conceptualized, and contributed to manuscript. SB-R: statistical analysis, conceptualized, and contributed to manuscript. LB: conceptualized project, acquisition of funding, and manuscript review. VB-G: reviewed manuscript. MC, SF, and AG: data collection and management, reviewed manuscript. All authors contributed to the article and approved the submitted version.
Funding and support for nest monitoring has been provided to the authors by the National Park Service, U.S. Army Corps of Engineers (REstoration COordination and VERification Monitoring and Assessment Plan), U.S. Geological Survey (Priority Ecosystem Science Program), U.S. Fish and Wildlife Service, Florida Power and Light Co., Florida Fish and Wildlife Conservation Commission, and the University of Florida. Federal funding was obtained through the South Florida and Caribbean Cooperative Ecosystems Studies Unit. Many permits and approvals have been obtained for crocodile nest monitoring. Currently we operate under IACUC # 202109072, Everglades National Park permit # EVER-SCI-2020-0032 and USFWS # TE077258-5. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Many individuals have contributed to monitoring American crocodile nesting in Florida. These include John Ogden who monitored crocodile nesting in the early 1970’s, William B. Robertson Jr. and James A. Kushlan who initiated nest monitoring in Everglades National Park, Paul Moler, who started nest monitoring on North Key Largo before it was Crocodile Lake National Wildlife Refuge, and Steve Klett and Jeremy Dixon who continued monitoring at Crocodile Lake NWR. Jim Lindsay, Mike Dorazio, Joe Wasilewski, Mario Aldecoa, and Mike Lloret supervised crocodile nest monitoring at the Turkey Point Power Plant site. Paige Patty, Jeff Beauchamp, Brian Jeffery, and Rafael Crespo supervised nest monitoring in ENP. We thank the many Croc Docs (http://crocdoc.ifas.ufl.edu/) and volunteers who helped us feed mosquitoes during crocodile nest hatching.
Angeler, D. G., Fried-Petersen, H. B., Allen, C. R., Garmestani, A., Twidwell, D., Chuang, W. C., et al. (2019). “Adaptive capacity in ecosystems,” in Advances in Ecological Research, In D. A. Bohan and A. J. Dumbrell (eds) (London: Academic Press) 60, 1–24. doi: 10.1016/bs.aecr.2019.02.001
Balaguera-Reina, S. A., Venegas-Anaya, M., Sanjur, O. I., Lessios, H. A., and Densmore, L. D. (2015). Reproductive ecology and hatchling growth rates of the American Crocodile (Crocodylus acutus) on Coiba Island. Panama. South Am. J. Herpetol. 10, 10–22. doi: 10.2994/SAJH-D-14-00024.1
Brandt, L. A., Mazzotti, F. J., Wilcox, J. R., Barker, P. D. Jr., Hasty, G. L. Jr., and Wasilewski, J. (1995). Status of the American crocodile (Crocodylus acutus) at a power plant site in Florida, USA. Herpetol. Nat. Hist. 3, 29–36.
Briggs-Gonzalez, V. S., Basille, M., Cherkiss, M. S., and Mazzotti, F. J. (2021). American crocodiles (Crocodylus acutus) as restoration bioindicators in the Florida Everglades. PLoS One 16:e0250510. doi: 10.1371/journal.pone.0250510
Calverley, P. M., and Downs, C. T. (2015). Movement and home range of Nile crocodiles in Ndumo game reserve, South Africa. Koedoe 57, 1–13. doi: 10.4102/koedoe.v57i1.1234
Campbell, H. A., Dwyer, R. G., Irwin, T. R., and Franklin, C. E. (2013). Home range utilization and long-range movement of estuarine crocodiles during the breeding and nesting season. PLoS One 8:e62127. doi: 10.1371/journal.pone.0062127
Charruau, P. (2012). Microclimate of American crocodile nests in Banco Chinchorro biosphere reserve, Mexico: effect on incubation length, embryos survival and hatchlings sex. J. Therm. Biol. 37, 6–14. doi: 10.1016/j.jtherbio.2011.10.010
Cherkiss, M. S., Mazzotti, F. J., Hord, L., and Aldecoa, M. (2014). Remarkable movements of an American crocodile (Crocodylus acutus) in Florida. Southeast. Nat. 13, 52–56. doi: 10.1656/058.013.0407
Cherkiss, M. S., Watling, J. I., Brandt, L. A., Mazzotti, F. J., Lindsay, J., Beauchamp, J. S., et al. (2020). Shifts in hatching date of American crocodile (Crocodylus acutus) in southern Florida. J. Therm. Biol. 88:102521. doi: 10.1016/j.jtherbio.2020.102521
Combrink, X., Warner, J. K., and Downs, C. T. (2017). Nest-site selection, nesting behaviour and spatial ecology of female Nile crocodiles (Crocodylus niloticus) in South Africa. Behav. Process. 135, 101–112. doi: 10.1016/j.beproc.2016.12.006
Convention on International Trade in Endangered Species of Wild Fauna and Flora [CITES] (2021). Apendices I, II and III. Maison internationale de l’environnement. Chemin des Anemones. Chate-laine. Geneva.
Deitz, D. C., and Hines, T. C. (1980). Alligator nesting in north-central Florida. Copeia 1980, 249–258. doi: 10.2307/1444001
Dorcas, M. E., Willson, J. D., Reed, R. N., Snow, R. W., Rochford, M. R., Miller, M. A., et al. (2012). Severe mammal declines coincide with proliferation of invasive Burmese pythons in Everglades National Park. Proc. Natl. Acad. Sci. U.S.A 109, 2418–2422. doi: 10.1073/pnas.1115226109
ESRI (2021). ESRI [Environmental Systems Research Institute] ArcGIS Pro 2.9.1 2021. Redlands, CA: ESRI.
Fick, S. E., and Hijmans, R. J. (2017). WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Gaby, R., McMahon, M. P., Mazzotti, F. J., Gillies, W. N., and Wilcox, J. R. (1985). Ecology of a population of Crocodylus acutus at a power plant site in Florida. J. Herpetol. 19, 189–198. doi: 10.2307/1564172
Garmestani, A. S., and Percival, H. F. (2005). Raccoon removal reduces sea turtle nest depredation in the ten thousand islands of Florida. Southeast. Nat. 4, 469–472.
Grigg, G., and Kirshner, D. (2015). Biology and Evolution of Crocodylians. Clayton, MO: CSIRO Publishing. doi: 10.1071/9781486300679
Hairston, N. G. Jr., and Fox, J. (2013). Dormancy and Diapause.in Encyclopedia of Biodiversity. 2nd Edn. Amsterdam: Elsevier. 665–669. doi: 10.1016/B978-0-12-384719-5.00210-0
Hall, P. M. (1991). Estimation of nesting female crocodilian size from clutch characteristics: correlates of reproductive mode, and harvest implications. J. Herpetol. 25, 133–141.
Hall, P. M., and Johnson, D. R. (1987). Nesting biology of Crocodylus novaeguinae in lake Murray. Herpetologica 42, 249–258.
Harris, I., Jones, P. D., Osborn, T. J., and Lister, D. H. (2014). Updated high-resolution grids of monthly climatic observations – the CRU TS3.10 Dataset. Int. J. Climatol. 34, 623–642. doi: 10.1002/joc.3711
Hénaut, Y., and Charruau, P. (2012). Nest attendance and hatchling care in wild American crocodiles (Crocodylus acutus) in Quintana Roo, Mexico. Anim. Biol. 62, 29–51. doi: 10.1163/157075511X597629
Jander, R. (1975). Ecological aspects of spatial orientation. Annu. Rev. Ecol. Evol. Syst. 6, 171–188. doi: 10.1146/annurev.es.06.110175.001131
Kawula, R., and Redner, J. (2018). Florida Land Cover Classification System. Tallahassee: Florida Fish and Wildlife Conservation Commission.
Kushlan, J. A., and Jacobsen, T. (1990). Environmental variability and the reproductive success of Everglades alligators. J. Herpetol. 24, 176–184. doi: 10.2307/1564225
Kushlan, J. A., and Mazzotti, F. J. (1989a). Historic and present distribution of the American crocodile in Florida. J. Herpetol. 23, 1–7. doi: 10.2307/1564309
Kushlan, J. A., and Mazzotti, F. J. (1989b). Population biology of the American crocodile. J. Herpetol. 23, 7–21. doi: 10.2307/1564310
Lutz, P. L., and Dunbar-Cooper, A. (1984). The nest environment of the American crocodile (Crocodylus acutus). Copeia, 153–161.
Maciejewski, K. (2006). “Temperature-dependent sex determination in the Nile crocodile, Crocodylus niloticus,” in the Okavango River, Botswana, and the effect of global climate change Doctoral dissertation (Stellenbosch: Stellenbosch University).
Magnusson, W. E. (1982). Mortality of eggs of the crocodile Crocodylus porosus in northern Australia. J. Herpetol. 16, 121–130. doi: 10.2307/1563804
Mazzotti, F. J. (1983). “The ecology of Crocodylus acutus,” in Florida – A Ph. D. thesis in ecology (State College, PA: The Pennsylvania State University).
Mazzotti, F. J. (1989). Factors affecting the nesting success of the American crocodile, Crocodylus acutus, in Florida Bay. Bull. Mar. Sci. 44, 220–228.
Mazzotti, F. J., Best, G. R., Brandt, L. A., Cherkiss, M. S., Jeffery, B. M., and Rice, K. G. (2009). Alligators and crocodiles as indicators for restoration of Everglades ecosystems. Ecol. Indic. 9, S137–S149. doi: 10.1016/j.ecolind.2008.06.008
Mazzotti, F. J., Bohnsack, B., McMahon, M. P., and Wilcox, J. R. (1986). Field and laboratory observations on the effects of high temperature and salinity on hatchling Crocodylus acutus. Herpetologica 42, 191–196.
Mazzotti, F. J., and Brandt, L. A. (1994). “Ecology of the American alligator in a seasonally fluctuating environment,” in Everglades: the ecosystem and its restoration, eds S. Davis and J. C. Ogden 485–505.
Mazzotti, F. J., Cherkiss, M. S., Parry, M. W., and Rice, K. G. (2007a). Recent nesting of the American crocodile (Crocodylus acutus) in Everglades National Park, Florida, USA. Herpetol. Rev. 38, 285–289.
Mazzotti, F. J., Brandt, L. A., Moler, P., and Cherkiss, M. S. (2007b). American crocodile (Crocodylus acutus) in Florida: recommendations for endangered species recovery and ecosystem restoration. J. Herpetol. 41, 122–132.
Mazzotti, F. J., Kushlan, J. A., and Dunbar-Cooper, A. (1988). Desiccation and cryptic nest flooding as probable causes of egg mortality in the American crocodile, Crocodylus acutus, in Everglades National Park, Florida. Florida Sci. 51, 65–72.
Metzen, W. D. (1977). Nesting ecology of alligators on the Okefenokee National Wildlife Refuge. Proc. Ann. Conf. S. E. Assoc. Fish Wildl. Agencies 31, 29–32.
Muff, S., Nilsen, E. B., O’Hara, R. B., and Nater, C. R. (2022). Rewriting results sections in the language of evidence. Trends Ecol. 37, 203–210. doi: 10.1016/j.tree.2021.10.009
Murray, C. M., Easter, M., Padilla, S., Marin, M. S., and Guyer, C. (2016). Regional warming and the thermal regimes of American crocodile nests in the Tempisque Basin, Costa Rica. J. Therm. Biol. 60, 49–59. doi: 10.1016/j.jtherbio.2016.06.004
Ogden, J. (1978). Status and nesting biology of the American crocodile, Crocodylus acutus (Reptilia: Crocodilidae) in Florida. J. Herpetol. 12, 183–196. doi: 10.2307/1563406
Olmsted, I. C., Loope, L. L., and Russell, R. P. (1981). Vegetation of the southern coastal region of Everglades National Park between Flamingo and Joe Bay. National Park Service South Florida Research Center Report T-620. Homestead, FL: National Park Service. 18.
Platt, S. G., Rainwater, T. R., Thorbjarnarson, J. B., and McMurry, S. T. (2008). Reproductive dynamics of a tropical freshwater crocodilian: morelet’s crocodile in northern Belize. J. Zool. 275, 177–189. doi: 10.1111/j.1469-7998.2008.00426.x
Platt, S. G., and Thorbjarnarson, J. B. (2000). Nesting ecology of the American crocodile in the coastal zone of Belize. Copeia 3, 869–873. doi: 10.1098/rsos.150409
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rainwater, T. R., Platt, S. G., Charruau, P., Balaguera-Reina, S. A., Sigler, L., Cedeño-Vázquez, J. R., et al. (2021). Crocodylus acutus. The IUCN Red List of Threatened Species 2021: e.T5659A168712617. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T5659A168712617.en (accessed May 17, 2022).
Rajon, E., Desouhant, E., Chevalier, M., Débias, F., and Menu, F. (2014). The evolution of bet hedging in response to local ecological conditions. Am. Nat. 184, E1–E15. doi: 10.1086/676506
Read, M. A., Grigg, G. C., Irwin, S. R., Shanahan, D., and Franklin, C. E. (2007). Satellite tracking reveals long distance coastal travel and homing by translocated estuarine crocodiles, Crocodylus porosus. PLoS One 2:e949. doi: 10.1371/journal.pone.0000949
Swanepoel, D. G. J., Ferguson, N. S., and Perrin, M. R. (2000). Nesting ecology of Nile crocodiles (Crocodylus niloticus) in the Olifants River, Kruger National Park. Koedoe 43, 35–46. doi: 10.4102/koedoe.v43i2.197
Thorbjarnarson, J., Mazzotti, F., Sanderson, E., Buitrago, F., Lazcano, M., Minkowski, K., et al. (2006). Regional habitat conservation priorities for the American crocodile. Biol. Conserv. 128, 25–36. doi: 10.1890/06-0838.1
Thorbjarnarson, J. B. (1989). “Ecology of the American crocodile (Crocodylus acutus) in Crocodiles: their ecology, management, and conservation,” in Crocodiles: Proceedings of the 7th Working Meeting of the Crocodile Specialist Group of the Species Survival Commission of the International Union for Conservation of Nature and Natural Resources, I. U. C. N. Publication New Series: Gland 228–258.
Thorbjarnarson, J. B. (2010). “American crocodile Crocodylus acutus,” in Crocodiles. Status survey and conservation action plan, Vol. 3, (Gland: IUCN Publication New Series). 46–53.
Thorbjarnarson, J. B., and Hernández, G. (1993). Reproductive ecology of the Orinoco crocodile (Crocodylus intermedius) in Venezuela. II. Reproductive and social behavior. J. Herpetol. 27, 371–379.
Ugarte, C. A., Bass, O. L., Nuttle, W., Mazzotti, F. J., Rice, K. G., Fujisaki, I., et al. (2013). The influence of regional hydrology on nesting behavior and nest fate of the American alligator. J. Wildl. Manag. 77, 192–199. doi: 10.1002/jwmg.463
USFWS (2007). Endangered and threatened wildlife and plants; reclassification of the American crocodile distinct population segment in Florida from endangered to threatened. Fed. Regist. 72, 13027–13041.
Webb, G. J., Messel, W. H., and Magnusson, W. (1977). The nesting of Crocodylus porosus in Arnhem Land, Northern Australia. Copeia 1977, 238–249. doi: 10.2307/1443905
Keywords: crocodylian, ecosystem restoration, nesting success, long-term monitoring, nesting ecology, Everglades
Citation: Mazzotti FJ, Balaguera-Reina SA, Brandt LA, Briggs-González V, Cherkiss M, Farris S and Godahewa A (2022) Natural and Anthropogenic Factors Influencing Nesting Ecology of the American Crocodile in Florida, United States. Front. Ecol. Evol. 10:904576. doi: 10.3389/fevo.2022.904576
Received: 25 March 2022; Accepted: 09 May 2022;
Published: 08 June 2022.
Edited by:
J. Sean Doody, University of South Florida, United StatesReviewed by:
Tom Langen, Clarkson University, United StatesCopyright © 2022 Mazzotti, Balaguera-Reina, Brandt, Briggs-González, Cherkiss, Farris and Godahewa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frank J. Mazzotti, ZmptYUB1ZmwuZWR1
†Present address: Seth Farris, U.S. Department of Agriculture, Agricultural Research Service, Invasive Plant Research Laboratory, Davie, FL, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.