 Helina S. Woldekiros
Helina S. Woldekiros A. Catherine D’Andrea
A. Catherine D’Andrea- 1Department of Anthropology, Washington University in St. Louis, St. Louis, MO, United States
- 2Department of Archaeology, Simon Fraser University, Burnaby, BC, Canada
The earliest settlements and states in the Horn of Africa were founded in mid to high-elevation areas by farmers and herders who were pioneers in agriculture and herding. Even today, places between mid- and high-elevation remain densely populated. The ancient Pre-Aksumites and Aksumites (1600 cal BCE–800 cal CE) of the north Ethiopian and Eritrean highlands established one of the most powerful states in the Horn of Africa in these high elevation areas through control of long-distance trade and intensive and extensive agriculture. However, despite the fact that agriculture was a significant source of wealth and subsistence for these ancient polities, there has been little research into the agricultural strategies of Pre-Aksumite and Aksumite societies. Using archaeological and faunal data collected from the site of Mezber dating from 1600 cal BCE to 400 cal CE, as well as prevsiously published data, this article provides zooarchaeological evidence for the earliest farming practices in the Horn of Africa. The research demonstrates a resilient highland agricultural strategy based on multispecies animal and plant resources, similar to most tropical agricultural systems today. A second important strategy of Pre-Aksumite farmers was the incorporation of both indigenous and exogenous plants and animals into their subsistance strategies. The Mezber site also offers one of the most thoroughly collected data to support multispecies farming practice in the north Ethiopian and Eritrean highlands.
Introduction
The transition from hunting and gathering to an agricultural way of life was a crucial time in human history. Agriculture was an especially important economic strategy for many ancient complex societies ranging from China to Mesoamerica (Blench and MacDonald, 2000; Ascalone and Frongia, 2007; Arbuckle, 2014; Denham et al., 2016). It helped people occupy new environments and create new economic niches. This process contributed to the rise of powerful states, polities, cities, and urban centers globally (Zeder, 1991; Zeder et al., 2006; Denham et al., 2016). Agropastoral products also fueled major ancient long- and short-distance global trade networks such as the silk and spice routes (Sidebotham, 2011; Frankopan, 2015; Frachetti et al., 2017; Spengler, 2019). Plants and animals transported thousands of miles across land and maritime trade routes were key in the formation of early agricultural systems. These early agriculture complexes, however, are less studied than domestication processes due to a scarcity of data and greater focus on studies in areas of the world where plant and animal domestication took place in situ (Zeder et al., 2006; Harrower et al., 2010; Denham et al., 2016). Understanding early agricultural systems provides not only a unique insight into the ways humans managed plants and animals in the past but also provides information on the technologies developed by early farmers to maintain and integrate the newly introduced domestic species to local environments and farming practices, as well as to maximize production and preserve biodiversity.
Case studies of prehistoric and historic agricultural strategies in the archaeological record—where there is evidence of successful implementation of local farming technologies for an extended period—provide information on the ways early farmers were instrumental in the creation and maintenance of resilient farming systems. A society dependent on an agricultural economy must develop a farming technology that is suitable for local environmental challenges. These environmental challenges could range from unpredictable seasonal rainfall to altitude, disease, drought, and war. Zooarchaeological studies globally, for example, show how and why early farmers developed cattle, sheep, goat, and chicken breeds that were meant to thrive in diverse natural and cultural environments. For example, dwarf or small-sized cattle, sheep, and goat breed types were developed in West Africa starting around 2000 BCE to deal with humidity-related diseases such as trypanosomiasis, a fly-borne disease (MacDonald and MacDonald, 2000; Gifford-Gonzalez and Hanotte, 2011). The diversity of domesticated maize (Zea mays ssp. mays) found widely distributed in Central, North, and Mesoamerica, on the other hand, is a good example of breed selection that is motivated by increased production and cuisine preference (Wang et al., 2017; Yang et al., 2019). This biodiversity is often created and maintained through continuous selection by local farmers over many generations (Wang et al., 2017). Not all early agricultural systems were, however, successful. We see this in the failure and multiple reintroductions of domestic animals in places such as Cyprus (Vigne et al., 2011).
Livestock played a significant role in the agricultural strategy of early settled farmers and mobile pastoralists in the Horn of Africa. Over the past few decades, research has increased our understanding of the origins and development of livestock keeping in the Horn prior to the Aksumite period (Shaw, 1995; Marshall, 2000; Marshall and Hildebrand, 2002; Gifford-Gonzalez and Hanotte, 2011; Lesur et al., 2014). Zooarchaeological and genetic studies demonstrate that major domesticates, such as cattle (Bos taurus), sheep (Ovis aries), and goats (Caprus hircus) have been central to the economic history of Africa, affecting mobile pastoralists, settled farmers, and urban dwellers (Mitchell, 2005; Pérez-Pardal et al., 2018). Domestic donkeys (Equus asinus) (circa [ca.] 6000 Before Present [BP]) (Marshall, 2007; Rossel et al., 2008; Kimura et al., 2013) and possibly cattle (ca. 9500 BP) (Wendorf and Schild, 1980; Stock and Gifford-Gonzalez, 2013) were the only animals domesticated in Africa. Mobile herders spread taurine cattle, donkeys, and ancient Asian sheep and goat breeds throughout arid northeastern Africa after 7000 BP. Cattle, sheep, and goats first appear in the Sudan ca. 6000 BP and northern Kenya ca. 5000–4000 BP. By 4700–3500 BP, sheep at Kerma in Sudan were long-legged and hairy breeds without fat tails (Chaix and Grant, 1987). By 3000–2000 BP in eastern Africa, distinctive large cattle appear, which have been linked to the introduction of humped cattle to Africa (Marshall, 1989). To date, evidence for herding in the rugged Ethiopian highlands is documented only at ca. 3500 BP, with pastoralist contexts suggesting similar processes of pastoralist migration and faunal transfer to those documented elsewhere in northeastern Africa, with the possibility of mountain barriers to early introduction (Marshall, 2000; Negash, 2001; Lesur, 2007; Lesur et al., 2014; Negash and Marshall, 2021).
In this article, we discuss the agricultural practices of the Pre-Aksumite and Aksumite polities, which controlled the northern Ethiopian and Eritrean highlands between 1600 cal BCE and 800 cal CE. Large faunal assemblages have been published from two sites in the Aksumite capital (Cain, 1999, 2000; Chaix, 2013) but beyond this little is known about the appearance or earliest husbandry of domestic animals in the Ethiopian highlands. Here we present new zooarchaeological data from the site of Mezber, a Pre-Aksumite highland elite-rural farming settlement in eastern Tigray, Ethiopia (Figure 1). The Mezber site was occupied continuously for at least 1,600 years (D’Andrea et al., in review)1. In this article, we identify the evolution of farming in the Horn of Africa by analyzing and discussing the way Pre-Aksumite rural farmers structured livestock keeping strategies. We compare data from Mezber with datasets from previously studied sites such as the D and K sites at Aksum (Cain, 1999, 2000), and ON and OAZ at Bieta Giyorgis (Chaix, 2013; Supplementary Table 1), to assess the farming technology of the Pre-Aksumites and Aksumites living in highland regions. These findings reflect on how an agricultural strategy that combined both indigenous and exogenous farming technology helped Pre-Aksumite and Aksumite elites, leaders, and farmers establish one of the most successful precolonial African states. It has been argued that the ancient Pre-Aksumites and Aksumites gained power and prestige through control of long-distance trade and prosperous agriculture. However, there has been very little study of the agricultural strategies prior to or during the Aksumite period that allowed the Aksumites to create such a powerful state and adapt to a highland environment.
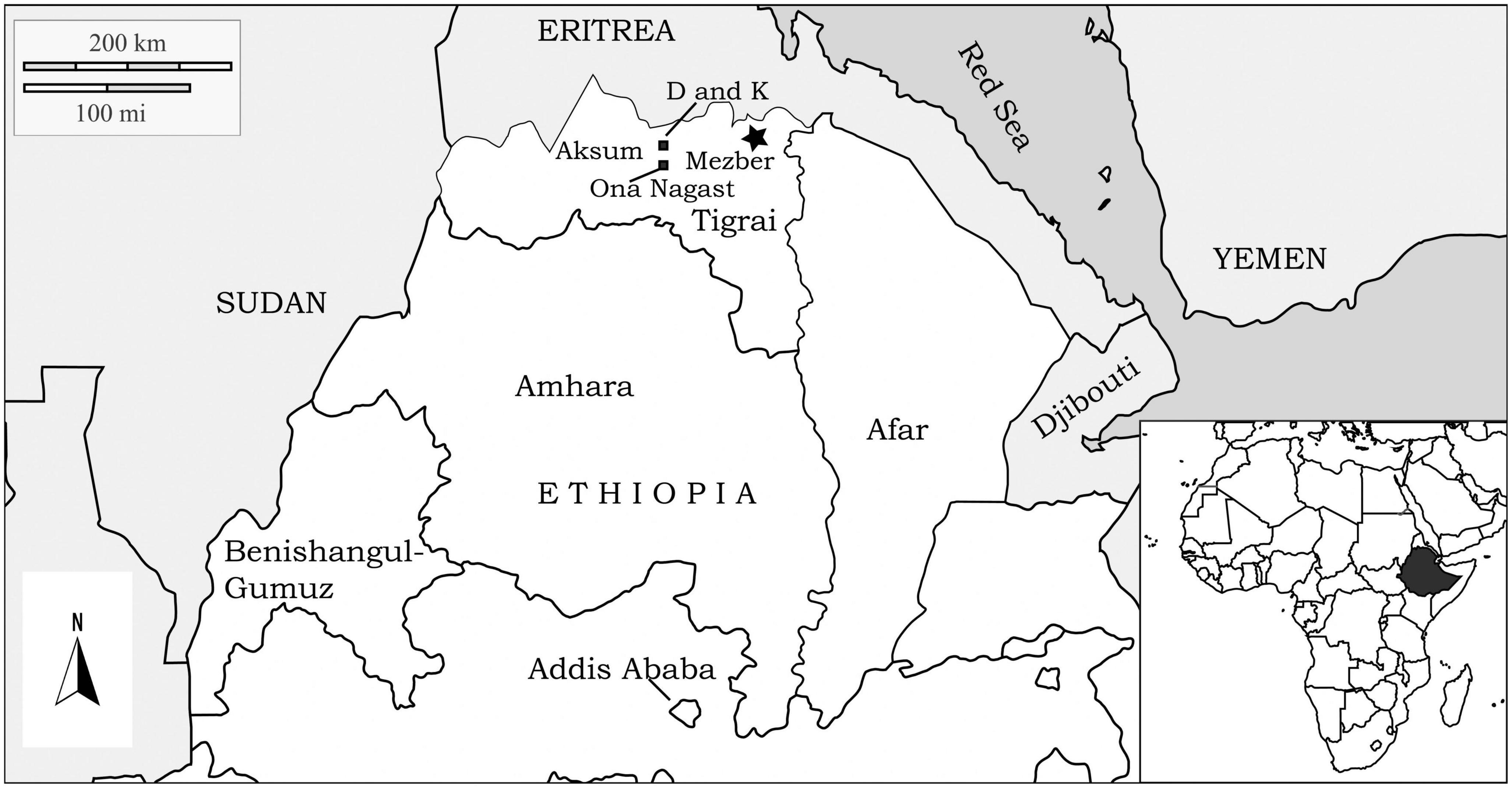
Figure 1. The location of the Pre-Aksumite site Mezber and Aksumite sties mentioned in the text (sites D, K, and Ona Nagast).
The earliest cattle, sheep, and goat remains in Ethiopia date to ca. 3500–3900 BP and were found at the site of Lake Besaka in central Ethiopia (c. 3500 BP) (Brandt, 1982), Danei Kawlos (3358 ± 47 BP, A-0214) (Negash and Marshall, 2021), at Gobedra (c. 3000 BP) (Phillipson, 1977) in northern Ethiopia, and at the site of Laga Oda in eastern Ethiopia (c. 3500 BP) (Clark and Prince, 1978). Although these dates signify a trajectory of movement of domestic cattle, sheep, and goats from North to South in Africa, there is still a lacuna in our understanding of the timing and the possible pathways for the spread of these domestic species. The goal of this study from Mezber is to fill these gaps and offer future research trajectories.
Pre-Aksumite and Aksumite societies, animals, and external relations
Pre-Aksumite (1600–450 cal BCE) and Aksumite (450 cal BCE–800 cal CE) peoples of the North Ethiopian and Eritrean highlands were well known for their extensive global trade relations from at least the early first millennium BCE to the late first millennium CE. Highland cities such as Aksum, Matara, and Yeha were occupied by elites that had contacts in the Arabian Peninsula, and from the second century onward had strong ties to Christian communities in West Asia, ancient Rome and the Nile Valley (Harrower et al., 2020). Trade routes linked these highland cities to the coastal port of Adulis, from which they conducted both direct and indirect trade across the Red Sea and Indian Ocean and inland north to the Nile Valley. Ancient epigraphic sources support that the Aksumites were importing prestige goods such as Roman glass, beads, wine, and olive oil and exporting a wide range of goods, including exotic animals such as civets and monkeys, and animal products including ivory, rhinoceros horn, hippopotamus hide and teeth, and tortoiseshell (Kobishchanov, 1979; Cain, 2000). There is also abundant archaeological evidence for imported ceramics and glass, but little for animal imports or exports. These trade interactions are best known from studies of South Arabian and Mediterranean ceramics as well as art and ancient documents and inscriptions (Fattovich, 1990, 1977; Manzo, 2009).
More recently, locally-based agriculture has been seen as the basis for the wealth, subsistence, and cultural complexity of the Aksumites (450 cal BCE–800 cal CE) (D’Andrea et al., 2008b; Harrower et al., 2010). Phillipson (2012) emphasizes the significance of African cattle to the Aksumite economy. However, the role of introduced crops in Aksumite agriculture has also long been recognised. Studies of the nature of domestic African cereals and the introduction and integration of West Asian cereals in ancient economies of the Horn of Africa have shown an intricate relationship between the incorporation of exotic crops and local innovation, which is still not fully understood. West Asian domesticates including wheat, barley, flax, and lentils and African domesticates such as t’ef and sorghum were both utilised in the Horn and adjacent regions by the early- to mid-first millennium BCE (Phillipson, 2000; D’Andrea et al., 2008a, 2011; Schmidt et al., 2008; Beldados and Costantini, 2011).
Even less is known about the timing of the introduction and process of integration of Asian animals such as humped cattle (Bos indicus), sheep (Ovis aries) and goat (Caprus hircus) breeds, camels (Camelus dromedarius), and chickens (Gallus gallus domesticus L.) into early farming systems and agricultural practices of the Horn. Faunal evidence for the earliest humped cattle in Ethiopian farming systems has proven inconclusive, with small numbers of bifid vertebrae suggestive of humped cattle at Aksum (Cain, 2000). Microsatellite genetic analyses of contemporary African cattle demonstrate that humped cattle were introduced to the rest of Africa from the Horn (Hanotte et al., 2002). Several Aksumite ceramic basins with figurines depict yoked cattle pulling plow (Phillipson, 2012). The practice of plowing is also distinctive of Ethiopian farming and is thought to have been introduced to the Horn, probably during the Pre-Aksumite period.
Evidence for early camels has so far proven elusive, with direct dates impossible to obtain from Gobedra where camel bones were found in a rock shelter context (Phillipson, 1977). While chickens have not been widely considered a component of the Aksumite agricultural complex, recent faunal data from Mezber and Aksum suggests the possibility that they were present in the Ethiopian highlands earlier than previously assumed (c. 3000 BP) (Woldekiros and D’Andrea, 2017). It has been further suggested that the Horn of Africa was a possible route for the introduction of chickens to Africa (MacDonald, 1992; Gifford-Gonzalez and Hanotte, 2011).
In general, floral and faunal exchanges in the Horn are widely thought to have been related to Aksumite trading activities in the Red Sea region (Marshall, 2000; Hanotte et al., 2002). Recent research suggests that efficient seafaring and small-scale opportunistic entrepreneurs on the coast played a significant role in changing the configuration of early farming systems in East Africa and the Horn through their interactions with inland societies (Fuller et al., 2011). Harrower et al. (2010), however, suggest that general models that consider early farming communities like the Pre-Aksumites to have partially received their agricultural life-ways from elsewhere give insufficient consideration to the complexity of these processes.
Archaeological context
The site of Mezber is a Pre-Aksumite rural settlement located in eastern Tigray, in the northern Ethiopian highlands (Figure 1). The site is 0.83 hectares, located in a valley bottom along a stream at 2,242 m above sea level, and surrounded by plateaus and mountains. Mezber was discovered in 2005 by the Eastern Tigrai Archaeological Project (ETAP) (D’Andrea et al., 2008b; Harrower and D’Andrea, 2014). The site was excavated from 2007 to 2012 and proceeded in five units or Fields (A-E). Fields A-E were further subdivided into areas (e.g., A1, A2, etc.) following Harrison et al., 2006; (Figure 2). Excavation has revealed an early and long sequence of material culture dating from the mid-second millennium to the end of the first millennium BCE (D’Andrea et al., 2011; D’Andrea et al., in review see text footnote 1). Our research here discusses one of the most significant large farming faunal assemblages known for the region in the context of four major phases identified for Mezber. D’Andrea et al’s (D’Andrea et al., in review see text footnote 1; D’Andrea and Welton, in preparation)2 forthcoming chronology based on 40 accelerator mass spectrometry dates (AMS) collected from well-preserved bones, plants, and charcoal (Table 1) stratigraphic data and Bayesian modelling indicates that the site was continuously occupied for at least 1,600 years with four clear occupational phases. The dates for these phases, ranging from the earliest to the latest, include ca. calibrated (cal) 1600–900 BCE (Initial), ca. cal 850–750 BCE (Early), ca. cal 500–400 BCE (Middle), and ca. cal 400 BCE–25 cal CE (Late) Pre-Aksumite phases (D’Andrea et al., in review see text footnote 1; Table 2). Full publication of detailed results from all the calibrated AMS dates and associated archaeological features is in progress (D’Andrea et al., in review see text footnote 1).
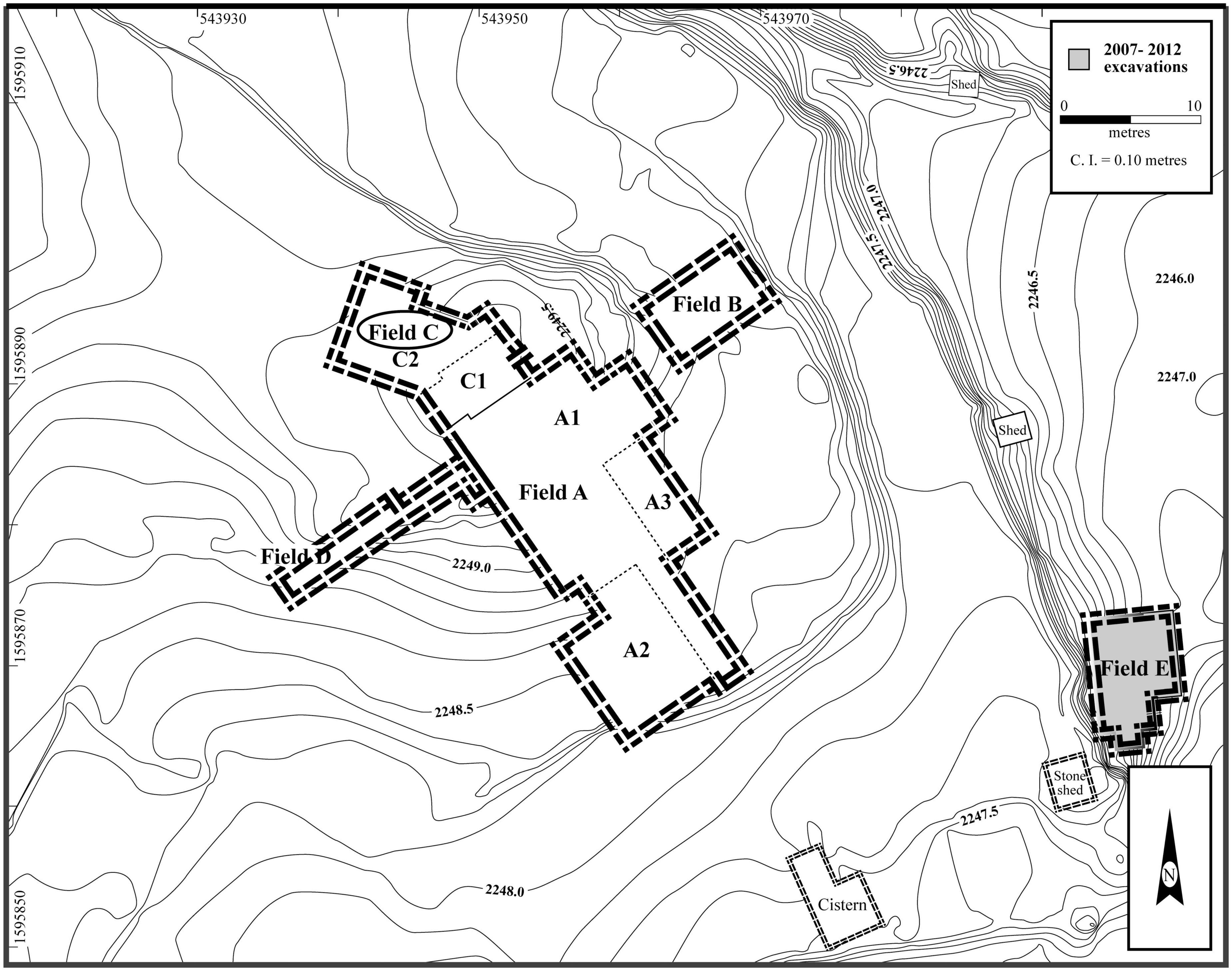
Figure 2. Fields/contexts with faunal remains excavated at Mezber. Map by Lynn Welton.
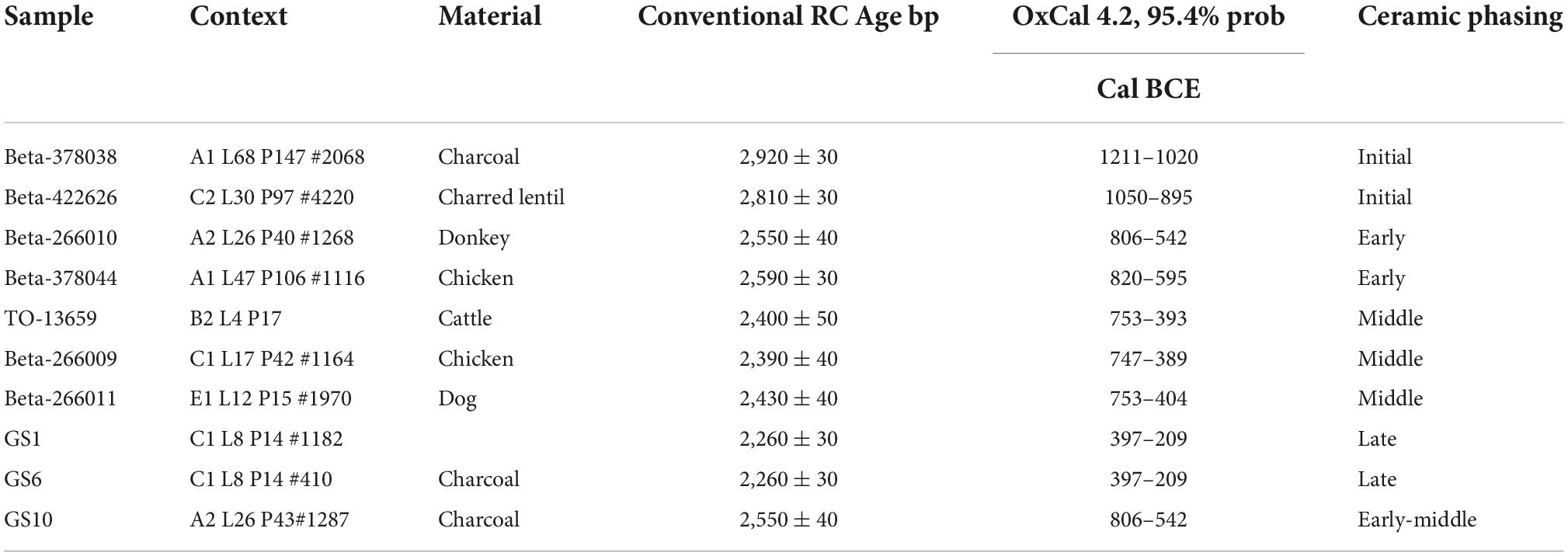
Table 1. Accelerator mass spectrometry radiocarbon dates of fauna from household context at Mezber (D’Andrea et al., 2011; Woldekiros and D’Andrea, 2017; Mercuri et al., 2018).

Table 2. Mezber modelled phases (from D’Andrea et al., in review see text footnote 1).
Mezber is a rural farming settlement with possible elite groups that was probably owned by one or more extended family units. The status of the site as an elite settlement is exhibited by the scale of the architecture, which includes one large building and several rectangular stone-walled buildings and room structures sometimes separated by stone-pavements. The architecture is marked by at least two distinctive building styles associated with the Early (ca. cal 850–750 BCE) and Middle (ca. cal 500–400 BCE) Phases respectively. Functions of identified spaces vary from living areas to cooking spaces to possible grain threshing areas. Unlike the Early and Middle Phases, no architectural features or buildings have been associated with the Initial Phase. The archaeological features associated with the Initial Phase may indicate utilisation of the site by semi-mobile pastoralists.
To date, excavations with systematic retrieval of biological data that bear on trade and exchange and early agricultural strategies have been conducted at only a handful of urban settlements. These include the elite areas of Aksum, Beita Georgis (Bard et al., 1997), the D site (Phillipson, 2000), and Asmara Plateau (Schmidt et al., 2008). Mezber is the first rural settlement to have been excavated and yields the most substantial faunal samples for the region.
Materials and methods
Zooarchaeological specimens were recovered from seven different contexts with established proveniences and AMS dates: Field A.1 (one complete room and a cobbled surface); A.2 (walls and domestic space); A.3 (rooms and a unique hearth-like feature); B (compare [cf.] outdoor crop processing area); C (room fill); D (room fill); and E (room fill and trash disposal area) (Figure 2). Each field included deep soundings excavated to bedrock. In order to make a meaningful analysis, the samples were grouped into the four main cultural phases, and interpretations of animal remains were based on stratigraphic information available from deep sounding in Field C (stratigraphic trench), that provided a continuous representation of the cultural sequences found at the site (D’Andrea et al., in review see text footnote 1). In addition to the AMS dates and stratigraphic information from deep sounding C and other fields, chronology for the faunal analysis was also based on direct AMS dates from, chicken, cattle, domestic dog, and donkey bones (Table 1). Sample size data for each excavation area, for all the field seasons, are summarised in Table 3 and Figure 3.
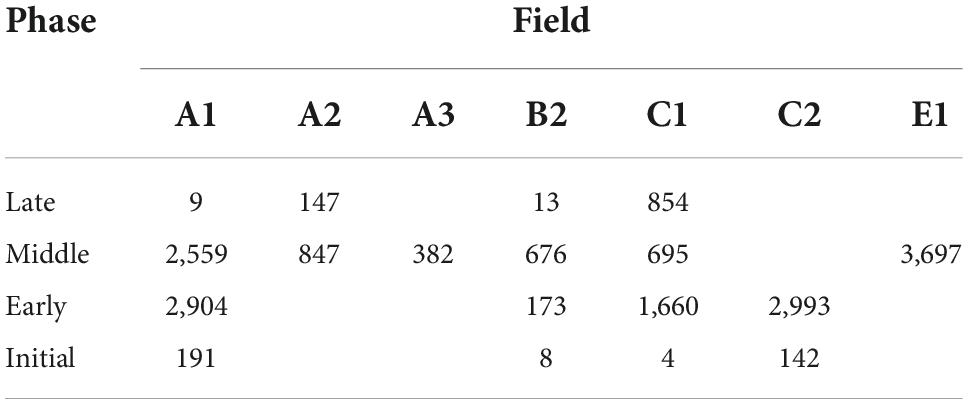
Table 3. Horizontal distribution of animal remains at Mezber.
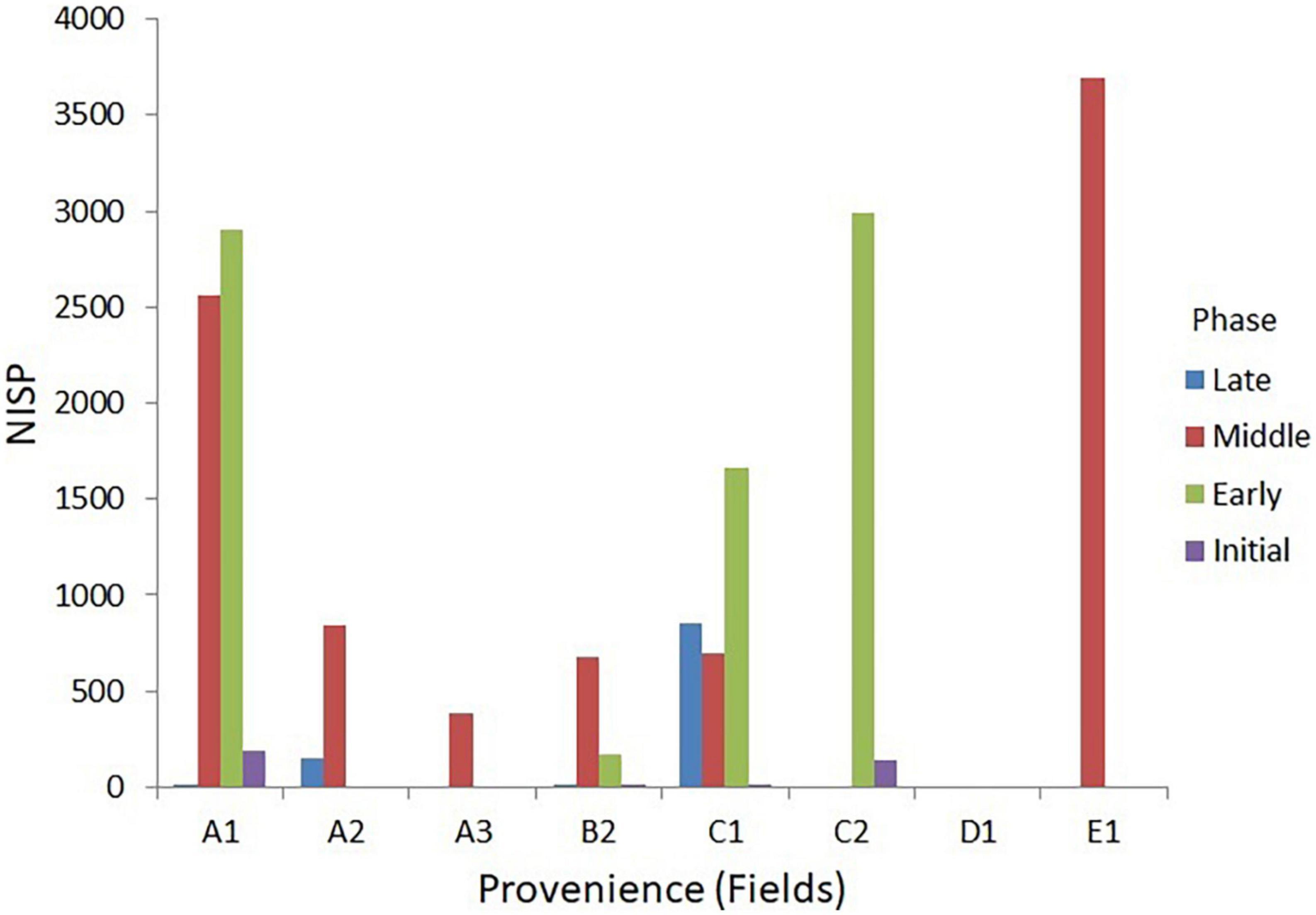
Figure 3. Sample size by phase and field.
Zooarchaeological identifications were made using ETAP reference collections and by following African zooarchaeological protocols for recording animal bones (Gifford et al., 1980; Walker, 1985; Marshall, 1990). Avian identifications were made following MacDonald (1992) and Bochenski and Tomek (2009). Modern comparative specimens collected near the site and housed at the History of Veterinary Medicine and Institute of Paleoanatomy in Munich, Germany, were also used. Identification of sheep and goats was carried out using Boessneck et al. (1964) and Zeder and Lapham’s (2010) methodology. Methods used to record age, or where possible sex cattle, sheep, and goats, were those of Silver (1969), Payne (1973), Noddle (1974), Grigson (1982), and Halstead (1985). Aging was attempted only on bones with clear epiphyseal fusion patterns. Most of the bones were highly processed and did not provide complete tooth remains for aging based on dentition. This is very common for most African faunal remains from archaeological sites.
For estimation of taxonomic abundance and diversity, numbers of identified specimens (NISPs) were counted following Klein and Cruz-Uribe (1984) and Lyman (2008). NISPs were preferred because they present the best opportunity for inter- and intrasite comparison, and because NISP values were favoured to account for the high degree of fragmentation at the site (Gautier, 1984). The diversity of species included in the economy of Pre-Aksumite rural farmers through time was calculated using Simpson’s diversity index (Faith and Du, 2018). We preferred to use Simpson’s diversity index because it accounts for both richness (species abundance) and evenness (how individuals are distributed among species) at the same time (Lyman, 2008; Faith and Du, 2018). The equation for Simpson’s Diversity Index is as follows:
In this formula, ni represents the number of specimens (NISP) of each taxon and N represents total NISP of all taxa. The Simpson’s diversity index (1-D’) value ranges between 1 and 0; 0 is no diversity and 1 represents infinite diversity. The larger the value of this index, the more evenly distributed specimens are across taxa. Greater richness indicates a multispecies dependent economy, whereas lesser evenness indicates the domination of few species in the economy. In order to avoid counting the same species twice, only taxa identified to family or genus level were included in this calculation. Fauna categorised as small, medium, and large bovid, and mammal were not included (Lyman, 2008).
The general anatomical categorisation of specimens included: cranial, axial, long bone, and NID (not-identifiable). Animal bones that were not identifiable to family or subfamily were categorised to bovid size class following Brain (1981), Bunn et al. (1986), Marshall (1986), and Gifford-Gonzalez (2005). These categories included: small bovid (<23 kg), medium bovid (23–84 kg), and large bovid or mammal (84–296 kg). All complete bones were measured following the methods described by Von den Driesch (1976).
Results and analysis
Taxonomic abundance and diversity
Numerous species of plants and animals were recovered from Mezber. Here we focus only on analysis of animal remains. A total of 22,514 animal bones were recovered from the ETAP excavation at Mezber. Of these, 7,747 number of identified specimens (NISP) were identifiable to the level of class, family, subfamily, genus, or species (Table 4). Taxonomic richness and evenness varied through time. Taxonomic richness was greater in the Early and Middle Phases and lesser in the Initial and Late Phases (Table 5). This indicates that the Middle and Early Phases were more taxonomically heterogeneous compared to the Initial and Late Phases. The ancient occupants of Mezber utilised both domestic and non-domestic animal resources. The majority of the specimens, however, are from domestic livestock. The most commonly identified domestic animals were cattle (Bos taurus/Bos indicus or hybrids), sheep (Ovis aries), goats (Capra hircus), and in addition chickens (Gallus gallus Domesticus L.), donkeys (Equus asinus africanus L.), and dogs (Canis familiaris L.) were identified. We used criteria presented by Boessneck et al. (1964) and Zeder (2006), as well as locally collected comparative material and published osteological atlases to discriminate between domestic sheep/goat and wild Ethiopian bovids and walia ibex (Capra walie Rüppell, 1852). Characteristics of the mandible and size indicated that the Canis sp. discovered at Mezber closely resembled the domestic dog than the wild canids found in the Ethiopian Highlands.
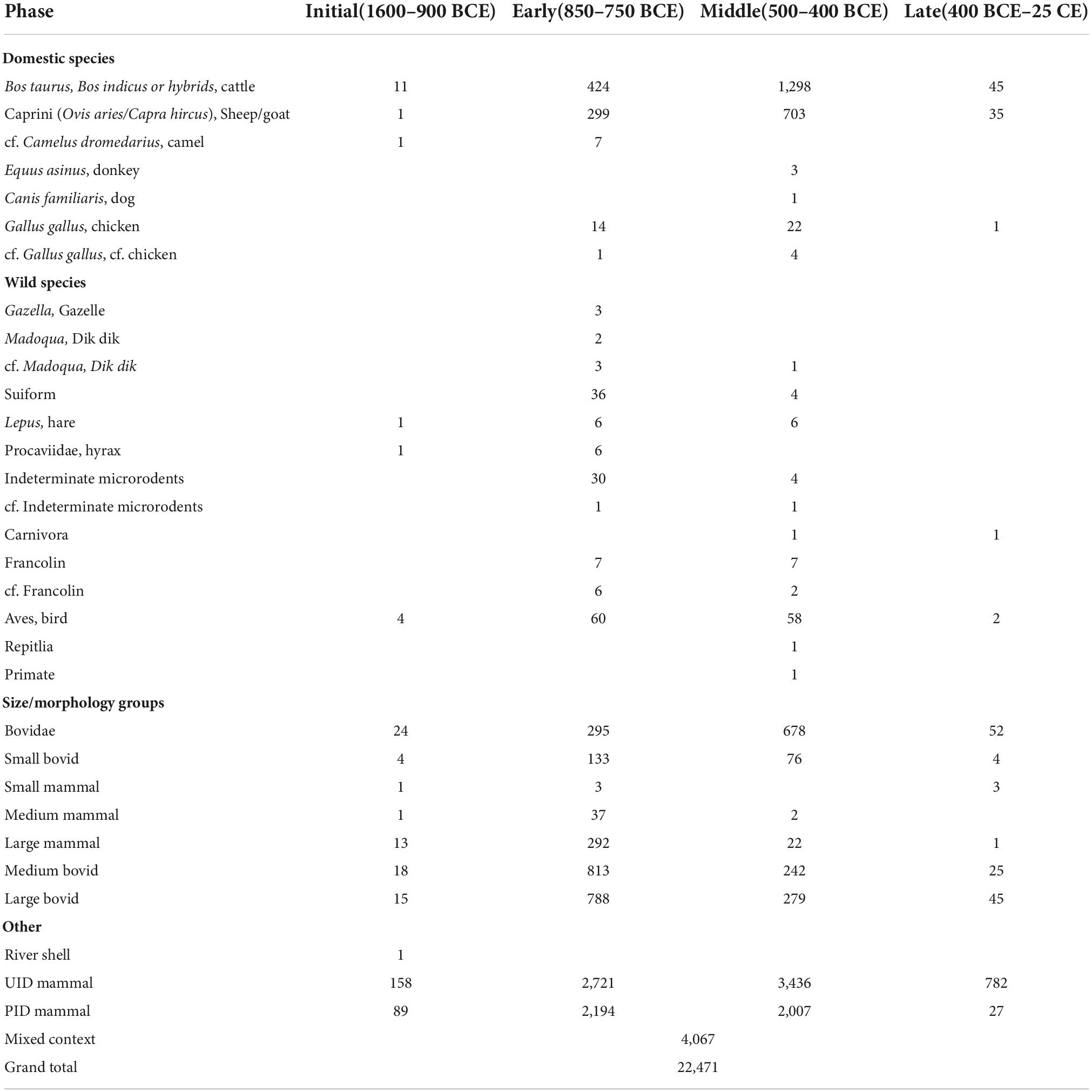
Table 4. Summary of livestock and wild animal remains recovered from Mezber by NISP and by phase.

Table 5. Simpsons diversity indices at Mezber calculated using NISP.
The overall percentage of domestic animal remains present at Mezber for the entire assemblage included: cattle (62%), sheep/goats (35%), chickens (1.5%), donkeys (0.11%), and cf. camels (0.28%). The wild fauna made up only 6% of the assemblage and included wild fowl (cf. Numidinae and Francolinus sp.), gazelle (Gazella), dik-dik (Madoqua), and cf. kudu (Tragelaphus) (Table 4 and Figure 3). Other taxa such as primates, rodents, hyrax (Procaviidae), carnivores, and galliform bird eggshells were also present in minimal quantities (8%) (Table 4). Although less numerous than domestic fauna, the presence of wild animals reflects variability in animal resource utilisation at Mezber. Evenness indices for Mezber fauna also showed variation in the percentage representation of domestic fauna through time with evenness indices for the Middle Phase (0.13) and Early Phase (0.19) lower than those of the Initial Phase (0.47) and Late Phase (0.44) (Table 5). This is mainly because of the domination of cattle and caprines compared to other species. During the Initial Phase, cattle were by far the most common domestic animal, accounting for 92% of the total of domestic animal assemblage. Cattle represented 61% of the identifiable fauna in the Middle Phase and 47% in the Early Phase. By the same token, caprines, the second most important domestic animals represented 33% of the domestic animal assemblage in both the Middle and Early Phases (Figure 4).
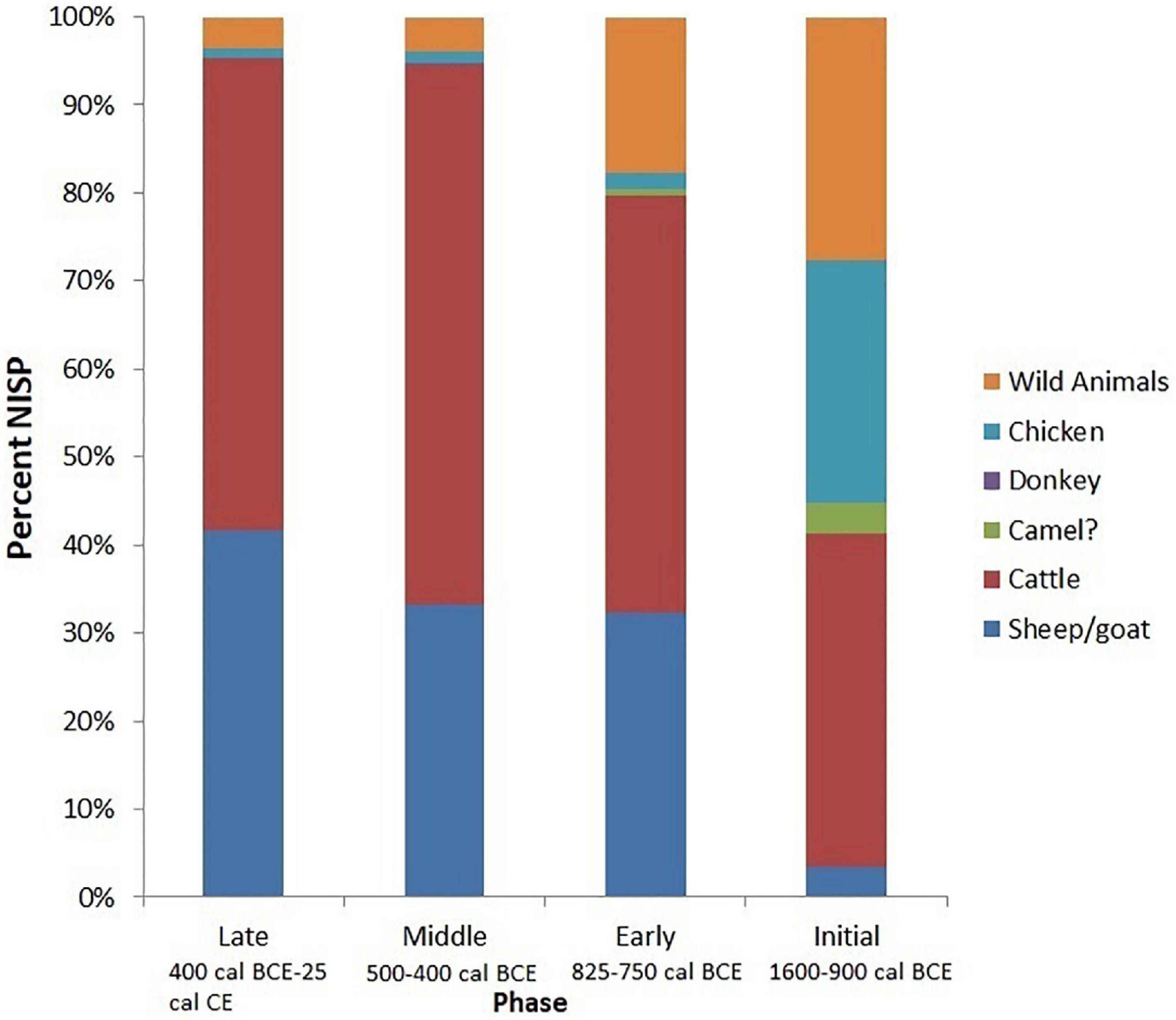
Figure 4. Summary of taxa identified at Mezber, using number of identified specimens (NISP).
It is clear that cattle were the most abundant taxon identified in the Mezber assemblage (Table 4). The stratigraphic context of fauna and direct AMS date at Mezber from the deep sounding Field C demonstrates that cattle were present at the site in the Initial Phase by 1600–900 cal BCE, before domestic caprines (sheep/goats). Well preserved cattle bones were found associated with a directly dated charred lentil dating to 1050–895 cal BCE (Beta-422626, C2 L30 P97 #4220) and charcoal dating to 1211–1020 cal BCE (Beta-378038, A1 L68 P147 #2068) in the Initial Phase (Table 1 and Supplementary Table 4). Directly dated cattle bone from the Middle Phase also showed that cattle keeping was well established at Mezber by 753–393 cal BCE (B2 L4 P17, TO-13659) (Table 1). This is slightly later than the earliest appearance of osteological data for the presence of cattle in the northern Ethiopian highlands which were recorded at Danei Kawlos rock shelter by 3358 ± 47 BP (Marshall and Negash, 2002; Negash and Marshall, 2021). Using paleoclimatic data from diatoms, isotopes, pollen, and change in the Rift Valley lakes of Ethiopia, scholars have shown that there was increasing aridity in the Horn of Africa starting from ca. 6000 BP, with a marked arid phase at around 3500 BP (Gasse, 1977; Gasse and Street, 1978; Negash and Marshall, 2021). Negash and Marshall (2021) argue that Danei Kawlos was possibly occupied by mobile pastoralists during this marked arid phase around 3500 BP. This date also coincides with the Initial Phase at Mezber which dates to ca. 1600–900 cal BCE. Mobile pastoralists might have been attracted to the Mezber site and surrounding highlands, because of the cool highland environment and available spring water for their cattle at a time when most of the Horn of Africa was experiencing an arid phase (Terwilliger et al., 2011, 2014).
Prior to Pre-Aksumite data from Mezber, zooarchaeological data on cattle utilisation for the Pre-Aksumite period was limited to the D and K sites in Aksum (Cain, 2000; Phillipson, 2000; Figure 1). Similar to Pre-Aksumite sites, zooarchaeological analysis of fauna from Aksumite period sites is also limited (Cain, 1999, 2000; Chaix, 2013; Supplementary Table 1). Outside the Ethiopian highlands, cattle dating to the Pre-Aksumite period have also been recovered from ancient Ona sites in Eritrea (Shoshani et al., 2008).
In order to compare the size of cattle at Mezber with those at other contemporaneous sites in northeast and East Africa and to gain perspective on ancient cattle breeds, measurements of the length (GLpe = mm) and breadth (BP = mm) of first phalanges were taken and compared with a published mean of a standard animal from Lidar Höyük, an early Bronze Age to Middle ages site in Turkey (Kussinger, 1988 in Hongo, 1997; Figure 5). We also compared the size of cattle at Mezber to published measurements from Ngamuriak, a Kenyan Neolithic site dating to 3000 BP (Marshall, 2000), a Sudanese Middle Kerma site dating to 2000–1800 BCE (Chaix, 2007), and K-7-41, an Egyptian Predynastic site dating to 4850–4250 BCE (Linseele et al., 2014; Figure 5). As is typical for African sites, few measurable specimens were preserved at Mezber (N = 10). Despite the small sample size, it can be seen from the result of this analysis that large cattle breeds were present in all four Pre-Aksumite phases (Figure 5). Most of the samples used for this test with GLpe range of 56–61 fall above the standard mean of GL-57 (Hongo, 1997; Figure 5), but are smaller than cattle from the pastoral site of Ngamuriak in Kenya (Glpe = 60–68; Supplementary Table 3; Marshall, 2000), and from Middle Kerma sites in Sudan (Glpe = 54–72) (Chaix, 2007; Figure 5). When Mezber cattle sizes were compared to those identified at previously published Pre-Aksumite and Aksumite sites, the Mezber cattle considerably overlap or fall in the intermediate range with animals found at sites D and K (GLpe = 53–70) and Ona Nagast (GLpe = 55–60) (Supplementary Table 3 and Figure 5; Cain, 2000; Chaix, 2013).
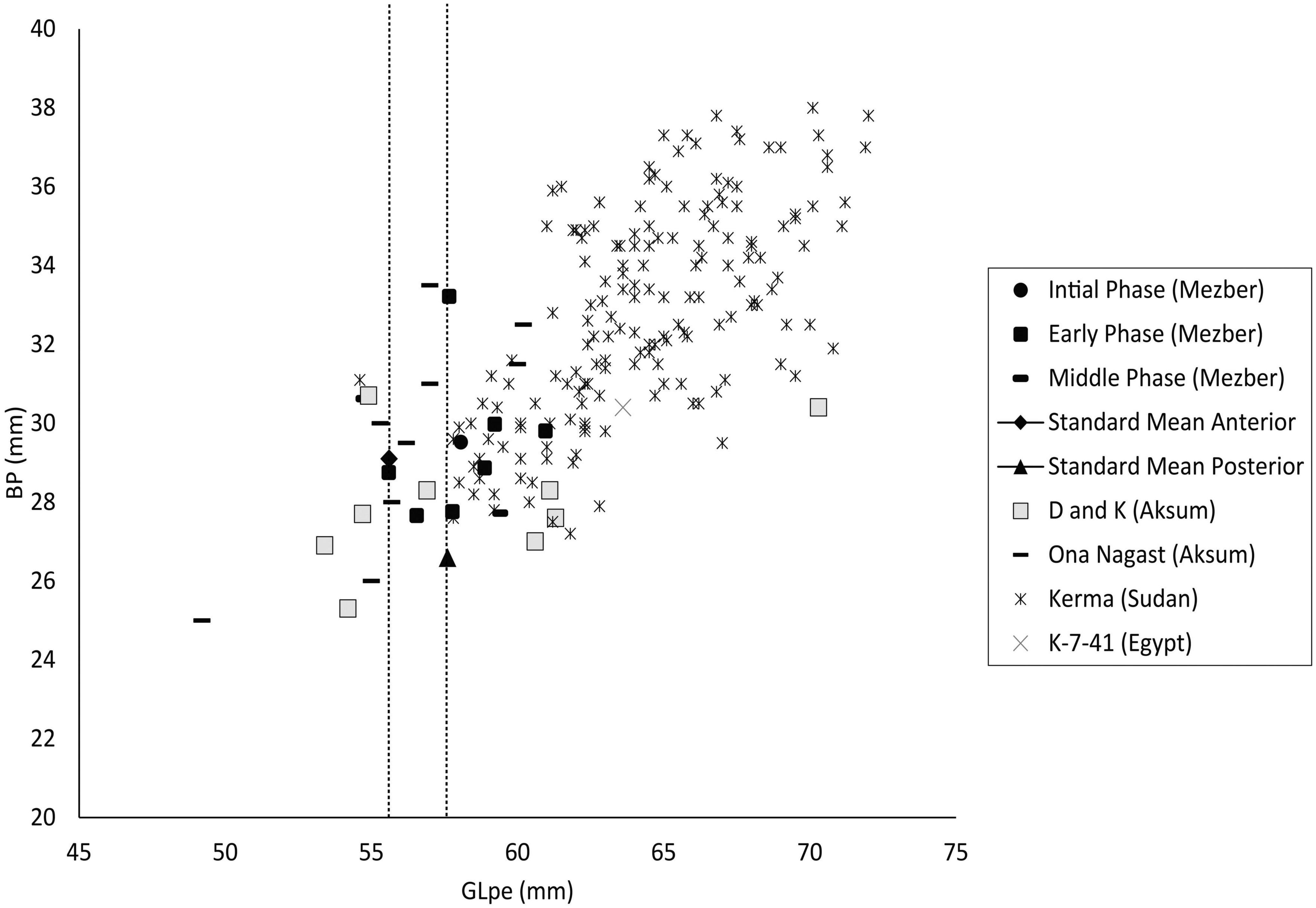
Figure 5. Length and breadth measurements (GLpe and Bp, in mm) of first phalanges from all contexts (fields) and phases at Mezber compared with cattle from Ona Nagast and sites D and K (Aksum), Kerma (Sudan), K-7-41 (Egypt) (Data from Cain, 2000; Chaix, 2007, 2013; Linseele et al., 2014). All measurments are taken following Von den Driesch (1976).
The second most abundant domestic fauna present at Mezber was sheep/goat (caprines) (Table 4). Both goat (Capra hircus) and sheep (Ovis aries) were present. Differentiation of sheep and goat would have allowed examination of their differing roles but in this highly fragmented domestic assemblage there were few skeletal parts preserved that could be used to distinguish between sheep and goats. As a result, specimens were combined as a general category sheep/goat (caprines) for quantitatively based discussion of medium-sized domestic stock.
Even though cattle appear earlier at the site, once sheep and goats (caprines) were introduced we see a constant increase in their importance (Table 4). Ratios that compare domestic animal species to one another help identify farmers’ agricultural practices that are geared toward diversity and integration of different species rather than simplification (Vandermeer et al., 1998). At Mezber, cattle outnumber sheep/goats by 1.7 to 1. However, cattle outnumber sheep/goats by 9 to 1 at urban Aksumite sites (Cain, 2000; Supplementary Table 1). In farming communities such as Mezber that relied heavily on both cereals and meat for subsistence, keeping large quantities of both cattle and sheep/goats was advantageous. This relying on rapidly reproducing small stock allowed farmers to use low investment animals for stock for regular meat consumption. It is also possible that cattle were supplied to the Aksumite capital region by rural settlements such as Mezber. Sheep and goat also diversify landscape and dietary needs of livestock herds and resistance to drought and disease (Dahl and Hjort, 1976). From this perspective, increasing caprine use through time reflects an increasingly diversified economy with higher tolerance to risk. High proportions of small stock are common in modern and ancient pastoral societies among more resource poor herders and those recovering from drought (Dahl and Hjort, 1976; Marshall, 1990; Janzen, 2015).
Chickens were the third most common domestic species at Mezber. A unique feature of the Mezber site is that it contained the earliest directly-dated chicken (820–595 cal BCE, Beta-378044, A1 L47 P106) in East Africa and the Horn to be found in a non-ritual and non-ceremonial context (Woldekiros and D’Andrea, 2017). Chicken remains, when compared to the other abundant domestic stock, are notably scarce at Mezber throughout the stratigraphic sequences. Domestic donkeys, though in small quantity, were also present at Mezber (Table 4). There is no historical or archaeological evidence that domestic donkeys were consumed as food during the Pre-Aksumite period, therefore disposal was unlikely in settlement middens. As a result, the presence of few skeletal remains of donkeys does not indicate that they were not widely used for trade or transport in the area (see also Marshall, 2007).
The role of cereals and grains amid the animal husbandry practice of farmers at Mezber was conspicuous. A significant quantity of plant data has been recorded from the site (Beldados et al., in review3). Limited study of stable isotopic evidence collected from the faunal remains, including chickens, cattle, sheep, and goats, shows that farm animals were not only allowed to graze but were also given fodder from domestic cereals. The animals were given both C3 (which could include straw chaff from wheat and barley) and C4 (t’ef, finger millet, and sorghum) cereals (D’Andrea et al., 2011). The C4 species: t’ef, finger millet, and sorghum are all indigenous African plants. The ratio of C3 to C4 plants found in each species varied. The stable isotope data also indicates a distinctive husbandry practice or food preferences for the three main farm animals. A 100% C4-plants value is indicated for the cattle and donkey, and a 50% mix of C3 and C4 plants is indicated for the sheep, goats, and chickens (D’Andrea et al., 2011). The straw of C4 plants like t’ef is still preserved for cattle today in Ethiopia and Eritrea, indicating this foddering practice might also have been present in ancient times.
Element representation
The study of body part representation at Mezber provided insights into management of the three dominant domesticates: cattle, sheep, and goats. Patterns of element representation at Mezber for cattle and caprines are presented in Supplementary Table 2 and a summary in Table 6. These data showed that body part representation is very similar for both cattle and sheep/goats. However, element frequencies for cattle and caprines varied throughout the four phases at Mezber.
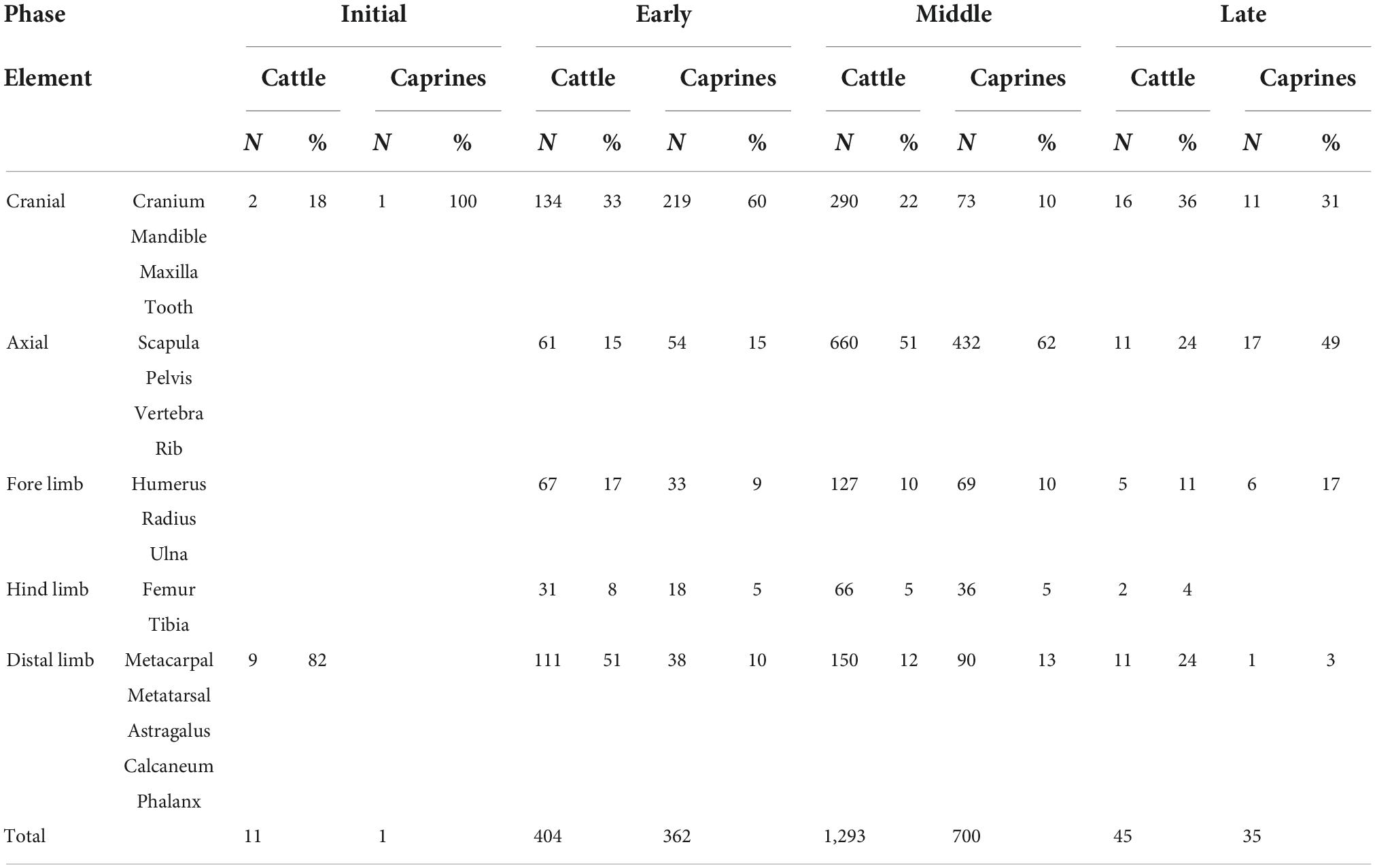
Table 6. Skeletal element representation.
During the Initial Phase, cattle distal limbs (92%) were present in higher proportions than cranial (9%) and axial (9%) elements. However, in the Early Phase, cattle and caprine cranial elements were over represented, followed by caprine and cattle axial elements. Similarly in the Middle Phase, there was a high proportion of cattle axial elements (51%), followed by cranial (22%), distal limb (10%), forelimb (10%), and hindlimb (5%). A similar pattern is seen in caprine axial elements (62%) distal limb (13%), cranial (10%), forelimb (10%), and hindlimb (5%). The dominance of caprine axial elements continues into the Late Phase (axial 49%), followed by cranium (31%) and forelimb (17%) bones. There were a small number of caprine distal limbs (3%) and no hind limb bones in the Late Phase. In the Late Phase cattle, however, crania dominated with cranium (36%), axial (24%), and distal limb (24%) elements. Though in small numbers, cattle fore limb (11%) and hind limb (4%) were represented.
Some of the variability in element representation through time could be due to bone density. In the case of Mezber however, there is a high representation of low-density axial bones, especially in the Middle and Late Phases, indicating cultural factors at play (Woldekiros and D’Andrea, 2017). Some of these include cuisine, cut preference and cooking style. Cattle and sheep/goat skeletal part representation at Mezber with high proportions of axial elements and varied cuts, shows that animals were raised and slaughtered at the site. Heavy processing of bones is also evident, and the most meaty hind and forelimb fragements are present at low frequency. The similarity in body part processing among cattle, sheep, and goats also implies similarity in their utilisation. The lack of high utility elements could indicate distribution elsewhere. Use-wear analyses of stone scrapers also suggests that a hide-proceessing craft industry was present at Mezber (Peterson, 2017). It is possible, therefore, that the high percentage of low-utility skeletal parts at Mezber such as metapoidals and phalanges was associated with leather production. How this fits with evidence for large scale leather trade from Aksum is unknown but fauna from this site was part of a larger regional system of agriculture and commerce.
Age and pathologies
The results for epiphyseal fusion data for cattle and caprines are summarised in Tables 7, 8. The tables give the relative age range in which cattle and caprines were slaughtered during the different time phases. However, there were small sample sizes with very few complete bones from the Initial and the Late Phase, therefore reporting focuses on the Early and Middle phases. In the Early Phase, it appears that the majority of the cattle, about 58% were slaughtered when they were between the ages of 24–36 months old. Very few cattle were butchered after 36 months or between 12 and 18 months. During the Middle Phase there is a later age range for cattle slaughtering at the site with the majority dying between 36 and 48 months. Though in small percentage, cattle aged 24–36 months as well as 12–18 months old were also butchered, indicating that adult cattle above the age of 36 months were butchered at the site more than younger cattle. The presence of older animals may indicate keeping of cattle for both meat and secondary products, including traction. The Late Phase like the Initial Phase did not yield large quantity of bone specimens that provided age data. The result for this phase, however, does not indicate animals that were slaughtered young between 12 and 18 months or old between 36 and 48 months.
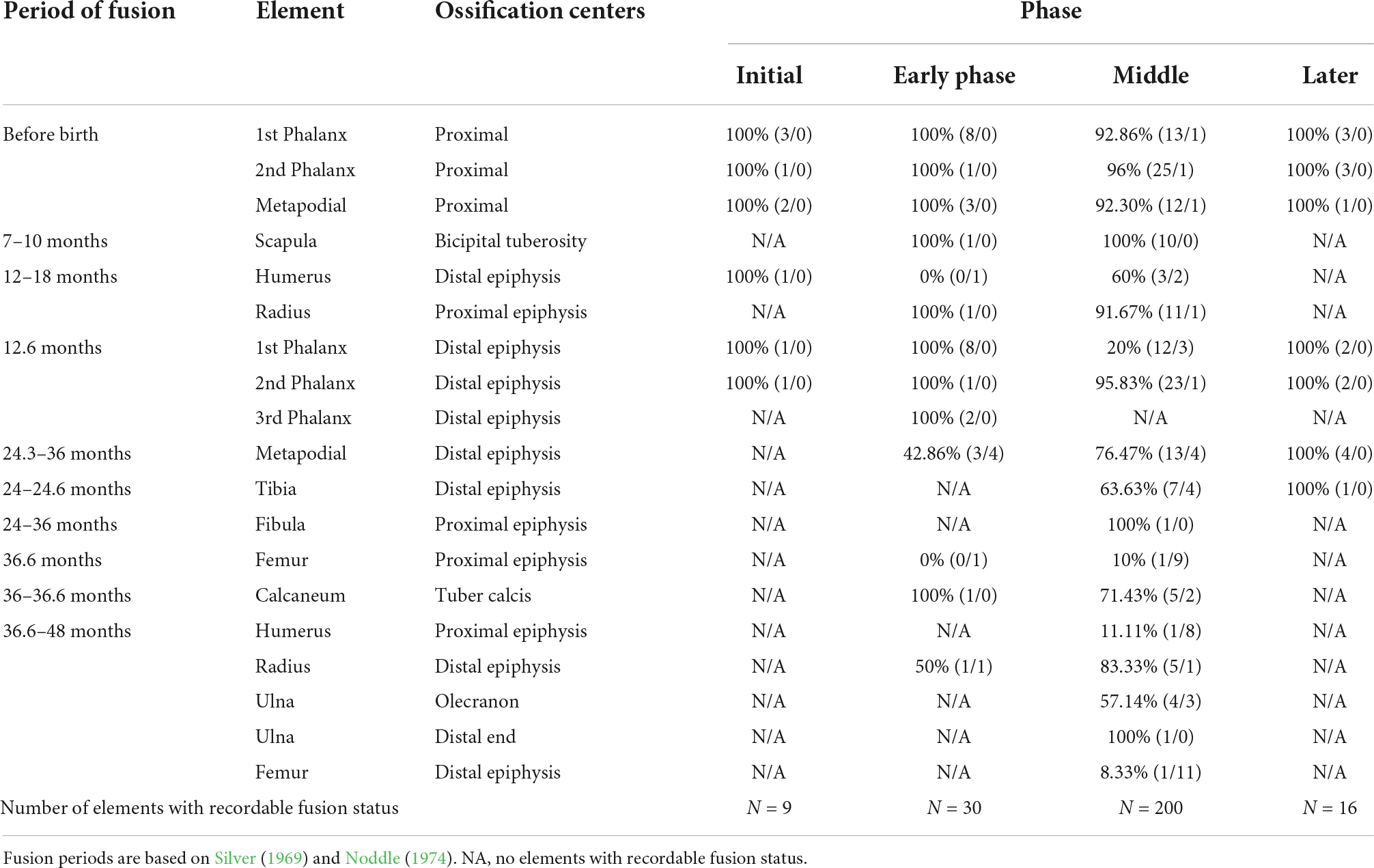
Table 7. Epiphyseal fusion by context for cattle and large sized bovids [% of NISP fused (#fragments fused/#fragments unfused)].
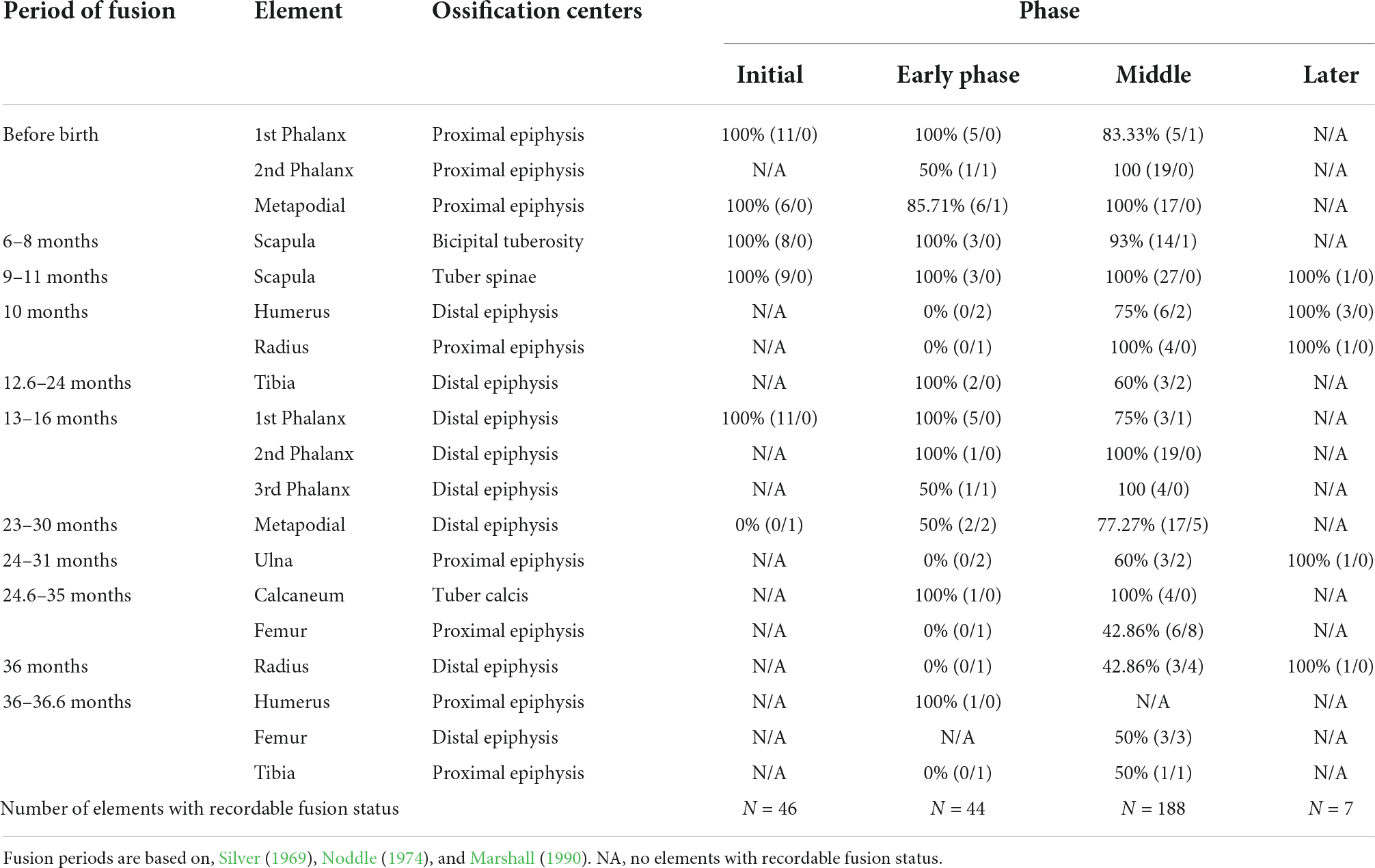
Table 8. Epiphyseal fusion by context for sheep/goats and medium sized bovids [% of NISP fused (#fragments fused/#fragments unfused)].
The age profile for caprines at Mezber shows a more widely spread age pattern during the Early and Middle Phases, where caprines of all ages, adult to juvenile were represented at the site. A large quantity of bones that normally fuse between the ages of 23–36 months were observed to be unfused. The Late Phase provided the least number of bones that can be used for age profile. The implication of the age structure of cattle and caprines is discussed later.
Evidence for traction can also be explored through pathologies. Although the numbers were too small for quantitative analyses, pathologies consistent with traction (Bartosiewicz and Gal, 2013) were present on at least three cattle phalanges recovered from Field E1. Use of cattle to pull ploughs would be consistent with Aksumite foot washers that depict yoked oxen (Phillipson, 2000).
Spatial and temporal distribution
Horizontal and vertical distribution of fauna at the site shows that during the Initial Phase sections of Fields A1 and C2 were utilised to process and consume animal products. In the Early Phase there was a clear expansion and use of the site as indicated by the construction of buildings with large stone walls and inclusion of Field B2 and C1, with the highest concentration of fauna at Field C2 and A1 (Figure 3). Agricultural intensification reaches its highest in the Middle Phase as indicated by the large number of fauna and broader site use with the addition of Fields A2, E1, and A3. A drop in agricultural intensification at the site during the Late Phase is indicated by the reduction of horizontal distribution of fauna, which was limited to Field A2 and C1.
Discussion: Integrated, multispecies agricultural strategy of the Pre-Aksumites and Aksumites
Analysis of animal remains from the site of Mezber indicates an agricultural strategy that was focused on multispecies with an emphasis on cattle, sheep, goats, donkeys, and chickens. Three agricultural strategies practiced by the Pre-Aksumite farmers at Mezber were identified: (1) a complex (multispecies) agroeconomy, (2) multipurpose livestock keeping for food (meat, milk and milk products, eggs), other secondary products (e.g., leather, food trade), and work animals (traction and pack animals, cattle, donkeys), and (3) an agricultural strategy that effectively integrated African and imported plant and animals domesticates in local farming and regional networks of trade and exchange. The success of these strategies is evidenced by the continued presence of these species in the archaeological record through the Pre-Aksumite period to the present day.
By keeping multispecies farm animals such as cattle, sheep, goats, and chicken, Pre-Aksumite farmers were able to not only increase production but also mitigate risks during times of conflict, epidemic, drought, or low rainfall. The Initial Phase occupation of Mezber during a time when the Horn of Africa was experiencing a marked arid phase around 3500 BP is a good example of the inception of this type of agricultural strategy in the highlands and uplands of Ethiopia and Eritrea. Small-scale farmers living in tropical regions still practice multispecies agriculture with the livelihood and culture of the family dependent on a diverse and well-integrated farming practice (Vandermeer et al., 1998). Sheep and goats were likely also critical risk-mitigating counterparts to cattle since they are easier to keep and maintain even during times of drought (Dahl and Hjort, 1976). The nature of the multispecies farming strategy of the early phase of agriculture at Mezber differs, however, from that of ancient pastoralist sites (such as Danei Kowlos in the region, or East African sites) (Janzen, 2015; Negash and Marshall, 2021; Prendergast et al., 2021) where chickens were not incorporated into the animal repertoire, cattle were not used for traction, possibilities for feeding animals on waste from fields were lower, but local or long distance family mobility and communal access to pasture was an important strategy. In contrast, the dominant form of mobility integrated into highland farming systems was local and long-distance trade. The Mezber site was used by Pre-Aksumite farmers who not only subsisted using their domestic herds but were also part of a chain of rural settlements that provisioned Pre-Aksumite towns, urban centers and trade routes with primary and secondary animal products in the form of meat as well as, milk and hide products. It is significant that reliance on diverse animals in highland farming systems was interdigitated with local crop production and processing, and political and economic relationships and trade among rural settlements and urban economies. Cattle, sheep and goat hides may have provided a valuable trade commodity and source of wealth for the local farming community. A local leather processing industry at Mezber is indicated by high proportions of low utility cattle, sheep and goat lower limb elements with the prevalence of stone scrapers associated with mass hide processing through use-wear analysis (Peterson, 2017). Field E at Mezber preserves a workshop dedicated to making the scrapers (D’Andrea et al., in review see text footnote 1). The concentration of scrapers found at Mezber is the largest from any Pre-Aksumite site.
The role of traction in highland agricultural systems
Cattle and donkeys played an important role in traction as animals used in field preparation through ploughing and in transport at Mezber. Mezber provides the first direct dates (806–542 cal BCE) for the presence of donkeys in the Horn of Africa in a context where local transport and trade were important. The sustained importance of cattle at Mezber and other Pre-Aksumite and Aksumite sites also makes it clear that the role of this animal in Pre-Aksumite and Aksumite economies was more complex than providing food, raw materials or even trade. Age profiles and pathology, with representations in Aksumite ceramics, indicate that by the Initial or Early Phases at Mezber (1600–900 cal BCE or 825–750 cal BCE), cattle were likely an important work animal. This represents the earliest integrated osteological and material evidence for ploughs in the Ethiopian highlands. Ox-ploughs were especially advantageous for Pre-Aksumite, Aksumite, and post-Aksumite societies because the highland diet heavily relied on cereals and pulses that were planted in rotation including cereals: t’ef, finger millet, sorghum, wheat, and barley, pulses: chickpeas, lentils, peas, along with oils: linseed and noog (D’Andrea, 2008; D’Andrea et al., 2018). Ploughs altered the dynamics of farming labor allowing rapid deeper preparation of varied soils with a range of gradients and elevations (1,500–3,000 m a.s.l). McCann (1995) argues that ox-plough cultivation in the Ethiopian highlands represented an “ecological revolution.” Use of plough expanded farming technology, altered agricultural landscapes, represented conscious environmental management, and was a key part of the establishment of a diverse but intensive agricultural system in the Ethiopian highlands and is still important to small-holder farmers today (McCann, 1995).
The timing of introduction of exotic domesticates and coming together of multispecies systems
The timing of introduction of animals from Eurasia and the ways that complex agricultural and trade systems came together influenced the trajectory of agriculture in the Ethiopian Highlands and regional agrobiodiversity today. To date, zooarchaeological data provide information regarding the sequence of introduction and adoption of cattle, sheep, goat, donkeys or chickens but little information on breeds. Mezber provides new direct dates for the presence of cattle in the Horn of Africa and information on cattle size. Direct dates for cattle in the Ethiopian highlands have been obtained from the pastoral site of Danei Kowlos (Negash, 2001; Marshall and Negash, 2002) where they date to ∼3700–3900 BP and from Mezber where they date to ∼1600–900 cal BCE (Table 1). African taurine breeds were early in Africa, with humped cattle thought to be introduced to East Africa in the second-century cal BP in eastern Africa (Marshall, 2000). Humped cattle were introduced to Egypt during the first half of the second millennium BCE (Payne, 1964; Epstein, 1971; Marshall, 2000, 1989). At least 28 cattle breeds exist in Ethiopia today including large and small East African Zebu, Sanga, Zenga, and the taurine populations (Assefa and Hailu, 2018). Some of these have adapted to varied altitudes and fly-borne diseases (Assefa and Hailu, 2018). Highland Zebu are smaller in size than Sanga but are well adapted to a confined pasture (McCann, 1995). Measurable elements from the site of Mezber indicate that cattle in all phases were smaller than those from Middle Kerma in Sudan and Ngamuriak in Kenya tentatively associated with early zebu, but larger than the standard animal from Turkey. The stature of the Mezber cattle was also not outside the range of previously reported cattle from other Pre-Aksumite and Aksumite sites. There is, therefore, no evidence for pronounced changes in animal size through time. It is possible that more complete specimens or biomolecular approaches will provide new insights into changing cattle breeds and selection in the Horn of Africa through time.
Sheep and goats were economically significant at Mezber by 825–750 cal BCE and increased in importance through time. Rock art and data from other regions provides useful insights into breeds that may have been introduced. Long-legged, thin-tailed sheep were present in Egypt by 6800 BP and replaced by fat-tailed sheep (Gautier, 1980; Chaix and Grant, 1987; Blench, 2006; Gautier and Van Neer, 2006). Sheep appear in East Africa and the Horn by ca. 4500–3700 BP but little is known about the origin of the fat-tailed sheep, which appears in rock art in the Horn of Africa (Clark and Williams, 1978; Marshall, 2000). Dwarf goats were present in the Middle Niger region of West Africa by 3000 BP, but the origin of goat breeds in the Horn is unknown (Gifford-Gonzalez and Hanotte, 2011). A study of recent goats shows considerable diversity of goat breeds in the Horn, especially in Ethiopia (Ayalew et al., 2006). This suggests long term use of goats in the Horn with adaptation and breed development by farmers to different ecological niches.
Another animal kept as a work animal at Mezber was the domestic donkey. We obtained a direct accelerator mass spectrometry date (806–542 calBCE, Beta-266010) for a donkey at Mezber (Table 1), the first direct date from the Horn. Ethnoarchaeological research (Woldekiros, 2019) in the Afar and Tigrai region demonstrates the likely role of pack animals in trade and as a source of wealth for the Aksumites. The fact that very few faunal remains belonging to pack animals (donkeys and camels) were recovered from Mezber, even though the present-day concentration of donkeys and camels in northern Ethiopia is the largest in the world (Gebreab et al., 2005) supports Cain (2000) and Marshall’s (2007) arguments regarding a faunal bias against these transport animals because they were rarely eaten or kept in settlements. It is likely that donkeys played a significant role in transport at the settlement and local and regional trade from Mezber.
The inception of adoptive agricultural strategies in novel environments?
The ease or difficulty with which exotic species were incorporated into the early African farming system could be attributed to social factors. For example, prior to the introduction of domestic chickens, we have evidence that early farming communities in Africa were exploiting wild fowls belonging to the Phasianidae family, to which domestic chickens also belong. Marshall (2000) argues that the reason the domestic camel is a late comer to the African herding system is that there was no prior experience with the wild ancestor of Camelidae in Africa. Other species including cattle, sheep, and goats that have attributes similar to the wild African antelope may have been more readily incorporated into African herding systems.
Even though domestic fauna dominates the assemblage at Mezber, they were sometimes supplemented by wild taxa. Wild species such as Francolinus sp., gazelle (Gazella), dik-dik (Madoqua), and cf. kudu (Tragelaphus) were the most common. The wild taxa at Mezber and Aksumite sites indicate a difference between wild taxa consumed locally and those that were acquired for trade or exchange. Ancient texts such as the Periplus of the Erythraean Sea (Huntingford, 1980) describe the trade of animals and animal products that are more ornamental than edible including rhinoceros horn, elephant ivory, tortoiseshell, monkeys, civet, and giraffe.
To summarize, cultural change in the north Ethiopian and Eritrean highlands did not disrupt an agricultural strategy that was over 3,000 years old. Although variability in seasonal rain and drought was a risk in the highlands during the Pre-Aksumite and Aksumite periods, it did not deter farmers from keeping cattle as a major farm animal. Inhabitants of Mezber used multispecies agricultural strategies to settle and adapt to their highland environment for at least 1,600 years. Comparison of data from D and K sites at Aksum and OA and OAZ sites at Bieta Giyorgis show that the same general approach to agriculture was adopted in the highlands.
Conclusion
At Mezber, we see early farming systems in Africa in a settled village context with evidence for long-term resilient African farming systems that depended on biodiversity and a multispecies agroeconomy. We see at Mezber an economic system that depended on both African and non-local animals—an economy that combined domestic species with wild animals at times. This economic system is in line with a strategy that was probably developed during the Holocene by early African herders as part of a risk management system in an ever-changing environment (see Marshall et al., 2011).
In addition to demonstrating a pattern of risk management, this study also indicates that social complexity and political stability at Mezber allowed for a settled way of life with a rich, multispecies economic system that included non-local animals acquired through exchange and trade. This study integrates faunal data from Mezber into a wider regional context aimed at understanding long-term human and animal interaction patterns in the Horn of Africa. Further work at sites with evidence for Pre-Aksumite and Aksumite influence will provide additional insights into regional differentiation in agricultural practices across space and time.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
AD’A was the PI of the project, she initiated and designed the project, undertook field data collection and laboratory analyses (plants), and other artifacts. HW undertook data analyses (all animal remains), including statistics, and produced faunal data images. HW wrote the initial draft of this manuscript with AD’A. Both authors contributed to the article and approved the submitted version.
Funding
Zooarchaeological research at Mezber was financially supported by grants from the Social Sciences and Humanities Research Council of Canada (SSHRC) including Standard Research Grant (No. 410-2007-2472), Standard Research Grant (No. 410-2011-1646), and Insight Grant (No. 435-2014-0182).
Acknowledgments
We are grateful to the team members of ETAP from 2008 to 2012, especially Stephen Batiuk (University of Toronto), Shannon Wood (Simon Fraser University), Lynn Welton (Durham University), and Steven Brandt (University of Florida). Permission for Mezber excavations was granted by the Authority for Research and Conservation of Cultural Heritage (ARCCH), Addis Ababa. We are grateful for the support of Tigrai Culture and Tourism Agency, Mekelle. We thank our assigned ARCCH representatives, including Habtamu Mekonnen and Ato Abebe Menguistu. We thank the communities of Tsahwa, Aby Adi, and Adigrat for their hospitality during our fieldwork and, in particular, Waizaro Medhin Abate and her family. We also thank Fiona Marshall who commented on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.901446/full#supplementary-material
Footnotes
- ^ D’Andrea, A. C., Welton, L., Manzo, A., Woldekiros, H. S., Brandt, S. A., Beldados, A., et al. (in review). The pre-aksumite period: indigenous origins and development in the horn of africa.
- ^ D’Andrea, A. C., and Welton, L. (in preparation). Excavation at mezber, ethiopia.
- ^ Beldados, A., Ruiz-Giralt, A., Lancelotti, C., Meresa, Y., and D’Andrea, A. C. (in review). Pre-aksumite plant husbandry in the horn of africa.
References
Arbuckle, B. S. (2014). The rise of cattle cultures in bronze age Anatolia. J. East. Mediterr. Archaeol. Herit. Stud. 2, 277–297. doi: 10.5325/jeasmedarcherstu.2.4.0277
Ascalone, E., and Frongia, R. M. G. (2007). Mesopotamia: Assyrians, Sumerians, Babylonians. Berkeley, CA: University of California press.
Assefa, A., and Hailu, A. (2018). Ethiopian indigenous cattle breed’s diversity, distribution, purpose of keeping, and their potential threats. J. Bio Innov. 7, 770–789.
Ayalew, W., Peacock, C., Alemayehu, N., Reda, A., and Rey, B. (2006). “The characterization of indigenous goat types of Ethiopia and Eritrea,” in The origins and development of African livestock: Archaeology, genetics, linguistics and ethnography, eds R. M. Blench and K. C. MacDonald (London: UCL Press), 280–289.
Bard, K. A., Fattovich, R., Manzo, A., and Perlingieri, C. (1997). Archaeological investigations at Bieta Giyorgis (Aksum), Ethiopia: 1993–1995 field seasons. J. Field Archaeol. 24, 387–403. doi: 10.1179/jfa.1997.24.4.387
Bartosiewicz, L., and Gal, E. (2013). Shuffling nags, lame ducks: The archaeology of animal disease. Oxford: Oxbow books. doi: 10.2307/j.ctvh1djdq
Beldados, A., and Costantini, L. (2011). Sorghum exploitation at Kassala and its environs, Northeastern Sudan in the second and first millennium BC. Nyame Akuma 75, 33–39.
Blench, R., and MacDonald, K. (eds) (2000). The origins and development of African livestock: Archaeology, genetics, linguistics and ethnography. London: UCL Press.
Bochenski, Z. M., and Tomek, T. (2009). A key for the identification of domestic bird bones in Europe. Krakow: Polish Academy of Sciences.
Boessneck, J., Müller, H.-H., and Teichert, M. (1964). Osteologische unterscheidungsmerkmale zwischen Schaf (Ovis aries Linné) und Ziege (Capra hircus Linné). Berlin: Akademie-Verlag.
Brain, C. K. (1981). The hunters or the hunted? An introduction to African cave taphonomy. Chicago, IL: University of Chicago Press.
Brandt, S. A. (1982). A late quaternary cultural/environmental sequence from Lake Besaka, southern Afar, Ethiopia. Ph.D. dissertation. Berkeley, CA: University of California.
Bunn, H. T., Kroll, E. M., Ambrose, S. H., Behrensmeyer, A. K., Binford, L. R., Blumenschine, R. J., et al. (1986). Systematic butchery by plio/pleistocene hominids at Olduvai Gorge, Tanzania [and comments and reply]. Curr. Anthropol. 27, 431–452. doi: 10.1086/203467
Cain, C. (1999). Results from zooarchaeological analysis at Axum, Ethiopia. Archaeozoologia 10, 27–45.
Cain, C̃ (2000). Animals at Axum: Initial zooarchaeological research in the later prehistory of the northern Ethiopian Highlands. Ph.D. dissertation. St. Louis, MO: Washington University in St. Louis.
Chaix, L. (2007). “Contribution to the knowledge of domestic cattle in Africa: The osteometry of fossil Bos taurus L. from Kerma, Sudan (2050–1750 BC),” in Skeletal series and their socio-economic context, eds G. Grupe and J. Peters (Rahden: Documenta archaeobiologiae), 120–249.
Chaix, L. (2013). The Fauna from the UNO/BU excavations at Bieta Giyorgis (Aksum) in Tigray, Northern Ethiopia: Campaigns 1995–2003; pre-aksumite, 700–400 BC to late aksumite, AD 800–1200. J. Afr. Archaeol. 11, 211–241. doi: 10.3213/2191-5784-10244
Chaix, L., and Grant, A. (1987). A study of a prehistoric population of sheep (Ovis aries L.) from Kerma (Sudan). Archaeozoological and archaeological implications. Archaeozoologia 1, 77–92.
Clark, J. D., and Prince, G. (1978). Use-wear on later stone age microliths from Laga Oda, Haraghi, Ethiopia and possible functional interpretations. Azania J. Br. Inst. East. Afr. 13, 101–110. doi: 10.1080/00672707809511633
Clark, J. D., and Williams, M. (1978). Recent archaeological research in southeastern Ethiopia. 1974-1975. Present. Ann. Ethiopie Editions Table Ronde 11, 19–44. doi: 10.3406/ethio.1978.902
D’Andrea, A. C., Manzo, A., Harrower, M. J., and Hawkins, A. L. (2008b). The Pre-Aksumite and Aksumite settlement of NE Tigrai. Ethiopia. J. Field Archaeol. 33, 151–176. doi: 10.1179/009346908791071268
D’Andrea, A. C., Schmidt, P., and Curtis, M. (2008a). “Palaeoethnobotanical analysis and agricultural economy in early 1st millennium BCE sites around Asmara,” in The archaeology of ancient Eritrea, eds P. R. Schmidt, M. C. Curtis, and Z. Teka (Trenton, NJ: Red Sea Press, Inc.), 207–216.
D’Andrea, A. C. (2008). T’ef (Eragrostis tef) in ancient agricultural systems of highland Ethiopia. Econ. Bot. 62, 547–566. doi: 10.1007/s12231-008-9053-4
D’Andrea, A. C., Perry, L., Nixon-Darcus, L., Fahmy, A. G., and Attia, E. A. (2018). “A pre-aksumite culinary practice at the mezber site, northern Ethiopia,” in Plants and people in the African past, eds A. M. Mercuri, A. C. D’Andrea, R. Fornaciari, and A. Höhn (Switzerland: Springer), 453–478. doi: 10.1007/978-3-319-89839-1_20
D’Andrea, A. C., Richards, M. P., Pavlish, L. A., Wood, S., Manzo, A., and Wolde-Kiros, H. (2011). Stable isotopic analysis of human and animal diets from two pre-Aksumite/Proto-Aksumite archaeological sites in northern Ethiopia. J. Archaeol. Sci. 38, 367–374. doi: 10.1016/j.jas.2010.09.015
Dahl, G., and Hjort, A. (1976). Having herds: Pastoral herd growth and household economy. Stockholm: Department of Social Anthropology, University of Stockholm.
Denham, T. P., Iriarte, J., and Vrydaghs, L. (2016). Rethinking agriculture: Archaeological and ethnoarchaeological perspectives. New York, NY: Routledge. doi: 10.4324/9781315421018
Epstein, H. (1971). The origin of the domestic animals of Africa. New York, NY: Africana publishing corporation.
Faith, J. T., and Du, A. (2018). The measurement of taxonomic evenness in zooarchaeology. Archaeol. Anthropol. Sci. 10, 1419–1428. doi: 10.1007/s12520-017-0467-8
Fattovich, R. (1977). “Pre-Aksumite civilization of Ethiopia: A provisional review,” in Proceedings of the seminar for Arabian studies (Oxford: JSTOR), 73–78.
Fattovich, R. (1990). Remarks on the Pre-Aksumite period in northern Ethiopia. J. Ethiop. Stud. 23, 1–33.
Frachetti, M. D., Smith, C. E., Traub, C. M., and Williams, T. (2017). Nomadic ecology shaped the highland geography of Asia’s silk roads. Nature 543, 193–198. doi: 10.1038/nature21696
Fuller, D. Q., Boivin, N., Hoogervorst, T., and Allaby, R. (2011). Across the Indian ocean: The prehistoric movement of plants and animals. Antiquity 85, 544–558. doi: 10.1017/S0003598X00067934
Gasse, E., and Street, F. (1978). Late quaternary lake-level fluctuations and environments of the northern Rift Valley and Afar region (Ethiopia and Djibouti). Palaeogeogr. Palaeoclimatol. Palaeoecol. 24, 279–325. doi: 10.1016/0031-0182(78)90011-1
Gasse, F. (1977). Evolution of Lake Abhé (Ethiopia and TFAI), from 70,000 bp. Nature 265, 42–45. doi: 10.1038/265042a0
Gautier, A. (1980). “Contributions to the archaeozoology of Egypt,” in Prehistory of the Eastern Sahara, Vol. 4, eds F. Wendorf and R. Schild (New York, NY: Academic Press), 317–340.
Gautier, A. (1984). How do I count you? Let me count the ways. Problems in archaeozoological quantification. Anim. Archaeol. 4, 237–251.
Gautier, A., and Van Neer, W. (2006). Animal remains from mahal Teglinos (Kassala, Sudan) and the arrival of pastoralism in the southern Atbai. J. Afr. Archaeol. 4, 223–233. doi: 10.3213/1612-1651-10073
Gebreab, F., Wold, A. G., Kelemu, F., Ibro, A., and Yilma, K. (2005). “Donkey utilization and management in Ethiopia,” in Donkeys, people and development: A resource book of the animal traction network for Eastern and Southern Africa (ATNESA), eds D. Fielding and P. Starkey (Netherlands: Wageningen), 46–52.
Gifford, D. P., Isaac, G. L., and Nelson, C. M. (1980). Evidence for predation and pastoralism at prolonged drift: A pastoral neolithic site in Kenya. AZANIA J. Br. Inst. East. Afr. 15, 57–108. doi: 10.1080/00672708009511277
Gifford-Gonzalez, D. (2005). “Pastoralism and its consequences,” in African archaeology: A critical introduction, ed. A. B. Stahl (Malden, MA: Blackwell Pub.), 187–224.
Gifford-Gonzalez, D., and Hanotte, O. (2011). Domesticating animals in Africa: Implications of genetic and archaeological findings. J. World Prehist. 24, 1–23. doi: 10.1007/s10963-010-9042-2
Grigson, C. (1982). “Sex and age determination of some bones and teeth of domestic cattle: A review of the literature,” in Ageing and sexing animal bones from archaeological sites, Vol. 109, eds B. Wilson, C. Grigson, and S. Payne (Oxford: B.A.R), 7–24.
Halstead, P. (1985). A study of mandibular teeth from Romano-British contexts at Maxey. Archaeol. Environ. Low. Welland Val. 1, 219–224.
Hanotte, O., Bradley, D. G., Ochieng, J. W., Verjee, Y., Hill, E. W., and Rege, J. E. O. (2002). African pastoralism: Genetic imprints of origins and migrations. Science 296, 336–339. doi: 10.1126/science.1069878
Harrison, T., Snow, H., Batiuk, S., Unruh, J., and Welton, L. (2006). Tayinat archaeological project: Field manual.
Harrower, M. J., and D’Andrea, A. C. (2014). Landscapes of state formation: Geospatial analysis of Aksumite settlement patterns (Ethiopia). Afr. Archaeol. Rev. 31, 513–541. doi: 10.1007/s10437-014-9165-4
Harrower, M. J., Dumitru, I. A., Perlingieri, C., Nathan, S., Zerue, K., Lamont, J. L., et al. (2020). Beta samati: Discovery and excavation of an Aksumite town–Corrigendum. Antiquity 94, 289–289. doi: 10.15184/aqy.2019.84
Harrower, M. J., McCorriston, J., and D’Andrea, A. C. (2010). General/specific, local/global: Comparing the beginnings of agriculture in the Horn of Africa (Ethiopia/Eritrea) and southwest Arabia (Yemen). Am. Antiq. 75, 452–472.
Hongo, H. (1997). Patterns of animal husbandry, environment, and ethnicity in central Anatolia in the Ottoman Empire period: Faunal remains from Islamic layers at Kaman-Kalehöyük. Nichibunken Jpn. Rev. 8, 275–307.
Janzen, A. (2015). Mobility and herd management strategies of early pastoralists in South-Central Kenya, 3000-1200 BP. Ph.D. thesis. Santa Cruz, CA: UC Santa Cruz.
Kimura, B., Marshall, F., Beja-Pereira, A., and Mulligan, C. (2013). Donkey domestication. Afr. Archaeol. Rev. 30, 83–95. doi: 10.1007/s10437-012-9126-8
Klein, R. G., and Cruz-Uribe, K. (1984). The analysis of animal bones from archeological sites. Chicago, IL: University of Chicago press.
Kussinger, S. (1988). Tierknochenfunde vom lidar höyük (südostanatolien). Unpublished Ph.D. thesis. München: University of Munich.
Lesur, J. (2007). Chasse et élevage dans la Corne de l’Afrique entre le Néolithique et les temps historiques. Oxford: Archaeopress (British Archaeological Reports Ltd). doi: 10.30861/9781407300191
Lesur, J., Hildebrand, E. A., Abawa, G., and Gutherz, X. (2014). The advent of herding in the Horn of Africa: New data from Ethiopia, Djibouti and Somaliland. Quat. Int. 343, 148–158. doi: 10.1016/j.quaint.2013.11.024
Linseele, V., Van Neer, W., Thys, S., Phillipps, R., Cappers, R., Wendrich, W., et al. (2014). New archaeozoological data from the Fayum “Neolithic” with a critical assessment of the evidence for early stock keeping in Egypt. PLoS One 9:e108517. doi: 10.1371/journal.pone.0108517
Lyman, R. L. (2008). Quantitative paleozoology. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511813863
MacDonald, K. C. (1992). The domestic chicken (Gallus gallus) in sub-Saharan Africa: A background to its introduction and its osteological differentiation from indigenous fowls (Numidinae and Francolinus sp.). J. Archaeol. Sci. 19, 303–318. doi: 10.1016/0305-4403(92)90019-Y
MacDonald, K. C., and MacDonald, R. H. (2000). The origins and development of domesticated animals in arid West Africa. London: UCL Press.
Manzo, A. (2009). Capra nubiana in berbere sauce? Afr. Archaeol. Rev. 26, 291–303. doi: 10.1007/s10437-009-9066-0
Marshall, F. (1986). Implications of bone modification in a Neolithic faunal assemblage for the study of early hominid butchery and subsistence practices. J. Hum. Evol. 15, 661–672. doi: 10.1016/S0047-2484(86)80003-3
Marshall, F. (1989). Rethinking the role of bos indicus in sub-Saharan Africa. Curr. Anthropol. 30, 235–240. doi: 10.1086/203737
Marshall, F. (1990). Origins of specialized pastoral production in East Africa. Am. Anthropol. 92, 873–894. doi: 10.1525/aa.1990.92.4.02a00020
Marshall, F. (2000). “The origins and spread of domestic animals in East Africa,” in The origins and development of African livestock: Archaeology, genetics, linguistics and ethnography, eds R. Blench and K. C. MacDonald (Padstow: T. J International Ltd.), 191–221.
Marshall, F., and Hildebrand, E. (2002). Cattle before crops: The beginnings of food production in Africa. J. World Prehist. 16, 99–143. doi: 10.1023/A:1019954903395
Marshall, F., and Negash, A. (2002). “Early hunters and herders of northern Ethiopia: The fauna from Danei Kawlos and Baati Ataro rockshelters,” in Paper Presented at the society for Africanist archaeologists meeting, Tucson, Arizona, (Tucson, AZ), 18–21.
Marshall, F. (2007). “African pastoral perspectives on domestication of the donkey: A first synthesis,” in Rethinking agriculture: Archaeological and ethnoarchaeological perspectives, eds T. P. Denham, J. Iriarte, and L. Vrydaghs (Walnut Creek, CA: Left Coast Press), 371–407.
Marshall, F., Grillo, K., and Arco, L. (2011). “Prehistoric pastoralists and social responses to climatic risk in East Africa,” in Sustainable lifeways: Cultural persistence in an ever-changing environment, eds N. F. Miller, K. M. Moore, and K. Ryan (Philadelphia, PA: UPenn Museum of Archaeology), 39–74. doi: 10.9783/9781934536322.39
McCann, J. C. (1995). People of the plow: An agricultural history of Ethiopia, 1800–1990. Madison, WI: Univ of Wisconsin press.
Mercuri, A. M., D’Andrea, A. C., Fornaciari, R., and Höhn, A. (2018). Plants and people in the African past: Progress in African archaeobotany. Cham: Springer. doi: 10.1007/978-3-319-89839-1
Mitchell, P. (2005). African connections: Archaeological perspectives on Africa and the wider world. Lanham, MD: AltaMira Press.
Negash, A. (2001). The holocene prehistoric archaeology of the Temben region, Northern Ethiopia. Ph.D. dissertation. Gainesville, FL: University of Florida.
Negash, A., and Marshall, F. (2021). Early hunters and herders of northern Ethiopia: The fauna from Danei Kawlos. SINET Ethiop. J. Sci. 44, 251–222. doi: 10.4314/sinet.v44i2.8
Noddle, B. (1974). Ages of epiphyseal closure in feral and domestic goats and ages of dental eruption. J. Archaeol. Sci. 1, 195–204.
Payne, S. (1973). Kill-off patterns in sheep and goats: The mandibles from Aşvan Kale. Anatol. Stud. 23, 281–303. doi: 10.2307/3642547
Pérez-Pardal, L., Sánchez-Gracia, A., Álvarez, I., Traoré, A., Ferraz, J. B. S., Fernández, I., et al. (2018). Legacies of domestication, trade and herder mobility shape extant male zebu cattle diversity in South Asia and Africa. Sci. Rep. 8, 1–8. doi: 10.1038/s41598-018-36444-7
Peterson, E. (2017). Development of craft specialisation during the Pre-Aksumite period in Eastern Tigrai, Ethiopia: A study of hideworking traditions. Ph.D. dissertation. Vancouver: Simon Fraser University.
Phillipson, D. W. (1977). The excavation of Gobedra rock-shelter, Axum: An early occurrence of cultivated finger millet in northern Ethiopia. AZANIA J. Br. Inst. East. Afr. 12, 53–82. doi: 10.1080/00672707709511248
Phillipson, D. W. (2000). Archaeology at Aksum, Ethiopia, 1993-97. London: British Institute in Eastern Africa and Society of Antiquaries.
Phillipson, D. W. (2012). Foundations of an African Civilisation: Aksum and the northern Horn, 1000 BC-AD 1300. Woodbridge: James Currey Publishers.
Prendergast, M. E., Grillo, K. M., Gidna, A. O., and Mabulla, A. Z. P. (2021). Grinding-stone features from the pastoral neolithic at Luxmanda, Tanzania. Antiquity 95, 1–9. doi: 10.15184/aqy.2021.13
Rossel, S., Marshall, F., Peters, J., Pilgram, T., Adams, M. D., and O’Connor, D. (2008). Domestication of the donkey: Timing, processes, and indicators. Proc. Natl. Acad. Sci. U.A.S. 105, 3715–3720. doi: 10.1073/pnas.0709692105
Schmidt, P. R., Curtis, M. C., and Teka, Z. (2008). The archaeology of ancient Eritrea. Trenton, NJ: Red Sea Press, Inc.
Shoshani, J., Ghebregiorgis, M., and Schmidt, P. (2008). “Interpretations of faunal remains from archaeological sites on the Asmara plateau of Eritrea,” in The archaeology of ancient Eritrea, eds P. R. Schmidt, M. C. Curtis, and Z. Teka (Trenton, NJ: Red Sea Press, Inc.), 217–233.
Sidebotham, S. E. (2011). Berenike and the ancient maritime spice route. Berkeley, CA: Univ of California Press. doi: 10.1525/california/9780520244306.001.0001
Silver, I. A. (1969). “The ageing of domestic animals,” in Science in archaeology, eds D. Brothwell and E. Higgs (New York, NY: Praeger Publishers), 283–302.
Spengler, R. N. III (2019). Fruit from the sands: The silk road origins of the foods we eat. Berkeley, CA: Univ of California Press. doi: 10.1525/9780520972780
Stock, F., and Gifford-Gonzalez, D. (2013). Genetics and African cattle domestication. Afr. Archaeol. Rev. 30, 51–72. doi: 10.1007/s10437-013-9131-6
Terwilliger, V. J., Eshetu, Z., Huang, Y., Alexandre, M., Umer, M., and Gebru, T. (2011). Local variation in climate and land use during the time of the major kingdoms of the Tigray Plateau in Ethiopia and Eritrea. Catena 85, 130–143. doi: 10.1016/j.catena.2010.08.003
Terwilliger, V. T., Eshetu, Z., Adderley, P. W., Jacob, J., Fogel, M. L., and Kassa, T. G. (2014). “Palaeoenvironmental change and the rise and fall D’MT and Aksum in Northern Ethiopia: How an unambiguous proxy for rainfall can improve interpretations of micromorphological and botanical data,” in Paper presented at the open PAGES focus 4 workshop, Louvain.
Vandermeer, J., van Noordwijk, M., Anderson, J., Ong, C., and Perfecto, I. (1998). Global change and multi-species agroecosystems: Concepts and issues. Agric. Ecosyst. Environ. 67, 1–22. doi: 10.1016/S0167-8809(97)00150-3
Vigne, J.-D., Carrere, I., Briois, F., and Guilaine, J. (2011). The early process of mammal domestication in the near east: New evidence from the pre-neolithic and pre-pottery neolithic in cyprus. Curr. Anthropol. 52, S255–S271. doi: 10.1086/659306
Von den Driesch, A. (1976). A guide to the measurement of animal bones from archaeological sites, Vol. 1. Cambridge, MA: Peabody Museum Press.
Walker, R. (1985). A guide to post-cranial bones of East African animals: Mrs Walker’s bone book. England: Hylochoerus Press.
Wang, L., Beissinger, T. M., Lorant, A., Ross-Ibarra, C., Ross-Ibarra, J., and Hufford, M. B. (2017). The interplay of demography and selection during maize domestication and expansion. Genome Biol. 18:215. doi: 10.1186/s13059-017-1346-4
Wendorf, F., and Schild, R. (1980). Prehistory of the eastern Sahara. Cambridge, MA: Academic Press Inc.
Woldekiros, H. S. (2019). The route most traveled: The afar salt trail. North Ethiopia. Chungara 51, 95–110. doi: 10.4067/S0717-73562019005000502
Woldekiros, H., and D’Andrea, A. (2017). Early evidence for domestic chickens (Gallus gallus domesticus) in the Horn of Africa. Int. J. Osteoarchaeol. 27, 329–341. doi: 10.1002/oa.2540
Yang, C. J., Samayoa, L. F., Bradbury, P. J., Olukolu, B. A., Xue, W., York, A. M., et al. (2019). The genetic architecture of teosinte catalyzed and constrained maize domestication. Proc. Natl. Acad. Sci. U.S.A. 116, 5643–5652. doi: 10.1073/pnas.1820997116
Zeder, M. A. (1991). Feeding cities: Specialized animal economy in the ancient near east. Washington, DC: Smithsonian Inst Pr.
Zeder, M. A. (2006). “Reconciling rates of long bone fusion and tooth eruption and wear in sheep (OVIS) and goat (CAPRA),” in Ageing and sexing animals from archaeological site (Oxford: Oxbow books), 87–118.
Zeder, M. A., and Lapham, H. A. (2010). Assessing the reliability of criteria used to identify postcranial bones in sheep, Ovis, and goats, Capra. J. Archaeol. Sci. 37, 2887–2905. doi: 10.1016/j.jas.2010.06.032
Keywords: zooarchaeology, livestock, highland agriculture, multispecies farming, Horn of Africa
Citation: Woldekiros HS and D’Andrea AC (2022) Complex (multispecies) livestock keeping: Highland agricultural strategy in the northern Horn of Africa during the Pre-Aksumite (1600 BCE–400 BCE) and Aksumite (400 BCE–CE 800) periods. Front. Ecol. Evol. 10:901446. doi: 10.3389/fevo.2022.901446
Received: 22 March 2022; Accepted: 31 October 2022;
Published: 29 November 2022.
Edited by:
Giedre Motuzaite Matuzeviciute, Vilnius University, LithuaniaReviewed by:
Mica Jones, School of Archaeology, Social Sciences Division, University of Oxford, United KingdomTaylor Hermes, Max Planck Institute for Evolutionary Anthropology, Germany
Copyright © 2022 Woldekiros and D’Andrea. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Helina S. Woldekiros, aHN3b2xkZWtAd3VzdGwuZWR1