 Ismael G. Espinoza1*
Ismael G. Espinoza1* Felipe Franco-Gaviria2
Felipe Franco-Gaviria2 Ivonne Castañeda3Charlotte Robinson2Alex Room4Juan Carlos Berrío5
Ivonne Castañeda3Charlotte Robinson2Alex Room4Juan Carlos Berrío5 Dolors Armenteras1
Dolors Armenteras1 Dunia H. Urrego2*
Dunia H. Urrego2*- 1Laboratorio de Ecología del Paisaje y Modelación de Ecosistemas — ECOLMOD, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá, Colombia
- 2Geography, Global Systems Institute, College of Life and Environmental Sciences, University of Exeter, Exeter, United Kingdom
- 3Independent Research Consultant, Medellín, Colombia
- 4School of Mathematics, Cardiff University, Cardiff, United Kingdom
- 5School of Geography, Geology and the Environment, University of Leicester, Leicester, United Kingdom
Rapid climate changes and the increasing presence of humans define the Holocene Epoch (11.6 calibrated kiloyears before present – hereafter kyr BP), when biological systems have faced the most recent and abrupt environmental changes. Understanding how biodiversity responds to extrinsic factors requires determining the effects of varying climatic conditions, changes in disturbance regimes, and increasing anthropogenic impacts. Despite being one center for biodiversity, the potential synergies of long-term anthropogenic and climate changes in shaping areas of high Andean biodiversity have yet to be explored fully. Here we present new pollen and charcoal records from the Pantano de Monquentiva (hereafter Monquentiva) on the highlands of the eastern flank of the Colombian Cordillera Oriental (CCO) to document relationships between climate, vegetation, and fire through the Holocene. We found compositional transitions at 8.7, 6.1, and 4.1 kyr BP at Monquentiva resulting from the interaction of climate, fire, and human occupation. Reduced moisture and temperature caused a compositional shift in Páramo vegetation from ca. 8.7 kyr BP. Fire activity was recorded throughout the Holocene and increased slightly during the Mid-Holocene when regional and local fire decoupling suggested human activities as the source of ignition. Mid-Holocene fires had a large effect on the vegetation composition at Monquentiva which recorded a rapid shift at ca. 6.8 kyr BP. Fire activity increased sharply from 4.1 kyr BP, promoting the reorganization of plant communities at 3.8 kyr BP. This shift in fire activity was likely related to more severe ENSO events and subsequently intensified by human activities after 3.8 kyr BP. Although high climatic sensitivity explains most Holocene vegetation changes in the eastern flank of the CCO, our study highlights the relevance of fire activity, uneven distribution of climatic variables, and human intervention to the composition of the vegetation we see today.
Introduction
The Colombian Andes are one of the most biologically and culturally diverse regions in the world. Colombian Andean ecosystems host ca. 28,000 plant species, highlighting their importance as a biodiversity hotspot (Bernal et al., 2015; Rangel-Ch, 2015). Since pre-Hispanic times, human groups have drawn on Andean natural resources, promoting the development of diverse societies (Aceituno et al., 2013; Archila et al., 2021; Cano et al., 2021). Despite the relevance of Colombian Andean ecosystems, expansion of human activities jeopardizes their continuity in the coming decades. Current land-use changes (Etter et al., 2008; Rodríguez Eraso et al., 2013), and uncontrolled fires (Armenteras et al., 2019) currently threaten Colombian Andean biodiversity. Novel climates are expected to completely restructure ecosystems by 2,100 (Williams et al., 2007), and in the tropical Andes, biodiversity is especially vulnerable because many species occupy narrow climatic and geographic ranges (Bush, 2002). Recent species range shifts in the tropical Andes have been documented (Feeley et al., 2011; Duque et al., 2015) and emphasize the urgency of understanding the coupled effects of climatic shifts and human intervention on Andean biodiversity.
The sensitivity of Andean biodiversity to long-term environmental variation has important effects on species migration and local extinctions because of unstable climatic conditions, responsible for rapid forest compositional shifts during the Holocene (Urrego et al., 2010, 2016). In the Colombian Andes, Hooghiemstra and van der Hammen (1993) recorded altitudinal species migrations of up to 1,400 m as a response to post-glacial warming in forests of the western flank of the Colombian Cordillera Oriental (CCO). Marchant et al. (2001) reported the spread of xerophytic vegetation over the Northern Andes, indicative of persistent dry conditions during the Middle Holocene. Despite recent efforts to understand how environmental drivers have shaped Andean biodiversity (Etter and van Wyngaarden, 2000; Boom et al., 2002; Gómez et al., 2007; Groot et al., 2011; van Boxel et al., 2014; Bird et al., 2017), the degree to which past environmental change, human presence and fire activity have produced ecologically novel communities in the tropical Andes remains understudied (but see Valencia et al., 2010; Sarmiento, 2012).
Despite being a driving force of forest composition and structural change, a key question remains open to debate: the source of ignition of fires in the Andes and whether these are solely the result of anthropogenic activities. In the long term, arid phases over the past ∼4.8 kyr BP (Bird et al., 2017) and multi-decadal regional climate forcing (Román-Cuesta et al., 2014) are thought to largely control fire occurrence in the Tropical Andes. However, the human influence on fire regimes in tropical contexts has also been acknowledged (Keating, 2007; Aguilera et al., 2009), with the fingerprint of anthropogenic fires on northern Andean ecosystems (Vélez et al., 2020) and the development of agricultural activities (Gómez et al., 2007) starting in the Late Holocene. Archeological research has found fires to be rare in grass páramo settings without human activity as igniting factor (White, 2013) and over the last century the prevalence of fire in the Colombian Andean region is thought to result from complex interactions between climate, vegetation types and human activities (e.g., Armenteras-pascual et al., 2011). Overall, the lack of long-term records of fire has hindered so far, the identification of fire activity baselines for the tropical Andes and the main sources of ignition.
Here we present a new sedimentary record from Monquentiva, located in the eastern flank of the CCO to explore the main paleoenvironmental drivers that have shaped the tropical high Andean ecosystems. We present pollen and charcoal records from Monquentiva spanning the Holocene to investigate the ecological baseline of vegetation and fire activity in this part of the Andes. We use novel statistical methods to identify regime shifts in vegetation and long-term ecological changes and explore to what degree the vegetation we find in Monquentiva today is novel to the site and results from enhanced Late-Holocene anthropogenic activity.
Study Site, Materials and Methods
Monquentiva (4°54′ N; 73°44′ W) is a peatbog inside a paramo that bears the same name (Páramo de Monquentiva). Monquentiva lies at ∼3,000 masl on the eastern flank of the CCO (Figure 1) and forms part of the Vista Hermosa Regional Natural Park, in the Guatavita municipality, ca. 50 km north of Bogotá. The bog has a single outlet, the Lagunero River, which drains into the Monquentiva River. Both the Lagunero and Monquentiva rivers flow eastward as they form part of the Gachetá River drainage basin. Despite hosting fluvial activity, our field observations show that the Monquentiva peatbog is a flat, ovoidal area of nearly 370 ha, which contains lacustrine and fluvioglacial deposits infilled during the Quaternary. The shape and position of these deposits are controlled structurally by the Río Blanco-Machetá Anticline and the Machetá Fault, which folded and faulted the sedimentary Cretaceous rocks of the Arenisca Dura Formation (Corredor and Terraza, 2015).
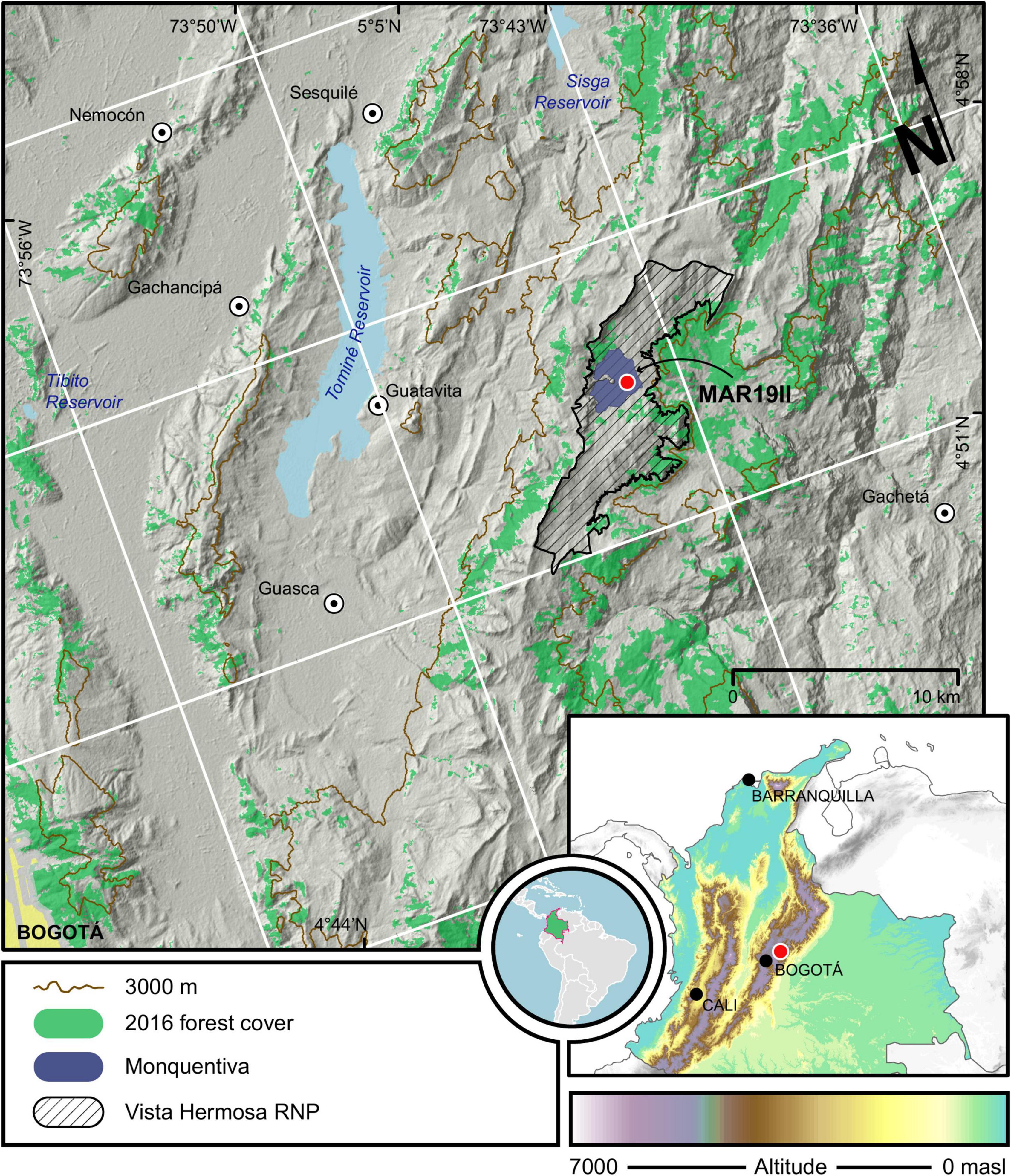
Figure 1. Map showing the location of Monquentiva on the eastern flank of the Colombian Cordillera Oriental (CCO) and within the Vista Hermosa Regional Natural Park (RNP). The red dot indicates the location of coring sites MAR19-II and MAR19-G1 collected next to each other from the center of the peatbog. Green patches show the current distribution of the High Andean Forest (IDEAM, 2016).
The mean annual precipitation in Monquentiva is 1,425 mm, with a unimodal rainfall distribution and the rainiest period between April–November (CAR, 2007). This unimodal distribution of precipitation in Monquentiva is controlled primarily by the annual migration of the inter-tropical convergence zone (ITCZ) and modified by the topography and the influence of southeast trade winds bringing moisture from the Amazon and the Atlantic Ocean (Poveda et al., 2006; Saylor et al., 2009). The annual temperature in the high CCO varies little over the year, with mean annual temperatures around 11°C in Monquentiva (CAR, 2007). El Niño-Southern Oscillation (ENSO) also influences the interannual precipitation variability in the Colombian Andes, with El Niño phase characterized by prolonged dry seasons and La Niña phase associated with increased precipitation (Poveda et al., 2006).
Vegetation in Monquentiva includes a mosaic of high Andean ecosystems coexisting within a narrow altitudinal gradient. High Andean Forest, Subpáramo, and Páramo are the most abundant vegetation groups. The High Andean Forests are typically dominated by Weinmannia, Hesperomeles, Clethra, Escallonia, Miconia with presence of Drimys, Diplostephium, Symplocos, and Vallea stipularis (Rangel-Ch, 2000). The Subpáramo follows in order of representation, composed by Pentacalia, Hypericum, Vaccinium, Pernettya, Buchetia, and Gaultheria (Avella-M et al., 2014). The Páramo, composed mostly of Poaceae and hosts Calamagrostis and Chusquea (Rangel-Ch, 2000). Near the coring site, stand wetland and peat-forming vegetation with cushions and ground layers, dominated by Geranium, Plantago, Hydrocotyle, Sphagnum, Blechnum, Azorella, and Distichia (Avella-M et al., 2014).
Core Collection, Description, and Chronology
We collected two sedimentary cores in the center of Monquentiva peatbog, using a modified Livingstone (Colinvaux et al., 1999) to extract the deepest sediments (MAR19-II; 4°54.49′ N, 73° 44.51′ W; length: 365 cm) and a UWITEC gravity corer (Melles et al., 1994) for the surface sediment (MAR19-GI, 4° 54.56′ N, 73° 44.06′ W; length: 30 cm). The total length of the core MAR19-II was collected in four segments (Drives 1–4), whereas MAR19-GI was a continuous short core. Both cores were transported to the laboratory at the University of Exeter, United Kingdom, split lengthwise, visually described, and stored at 4°C before analyses. The sediment description followed the modified Troels-Smith system (Kershaw, 1997), accounting for the physical characteristics, composition, and level of humification of the sediments. This study focused on the upper 77 cm (Drive 1) of the sediment core MAR19-II and complemented with MAR19-GI as the coupling effects of human disturbance and climate fluctuations on vegetation was of our highest interest. We collected volumetric samples of 0.5 and 1 cm3 every 2 cm for pollen and charcoal analysis, respectively. Seven bulk sediment samples for MAR19-II (from Drives 1 and 2) and three samples for MAR19-GI were collected for accelerator mass spectrometry radiocarbon analysis (AMS 14C) at the CHRONO laboratory of the Queen’s University Belfast. We constructed an age-depth model for each core, using the rbacon package (Blaauw and Christeny, 2011) and the IntCal20 radiocarbon calibration curve (Reimer et al., 2020). Ages are reported as the mean probability of calibrated ages.
Palynology
Samples were processed using a modified version from Faegri and Iversen (1964). One tablet with 20,848 of Lycopodium clavatum spores was added to each sample to estimate pollen concentrations (number of pollen grains/cm3). Identification and counting of pollen and spores were carried out under a microscope Leica DM500 at 400 and 1,000× magnifications. A minimum of 300 pollen grains were quantified, where possible. Aquatic taxa (Myriophyllum and Isoëtes) were quantified but excluded from the pollen sum. The pollen identification followed available taxonomic keys (e.g., Hooghiemstra, 1984; Velásquez, 1999), online databases (e.g., Bush and Weng, 2007) and reference materials at the Laboratory of Paleoecology and Palynology of the Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. The ecological grouping (Supplementary Table 1) followed published literature from vegetation surveys conducted in the CCO (Cuatrecasas, 1958; Hooghiemstra and van der Hammen, 1993; Rangel-Ch et al., 1997; Rangel-Ch, 2000; Gómez et al., 2007; Avella-M et al., 2014). We produced stratigraphic diagrams using the calibrated ages, pollen, and charcoal counts in the C2 program version 1.7.7 (Juggins, 2007). We used a CONISS analysis (Grimm, 1987) to define local pollen zones and delimit the most relevant ecological periods based on vegetation compositional changes from taxa belonging to the pollen sum.
Charcoal
Sedimentary charcoal particles were identified and quantified in petri dishes using a Zeiss Stemi stereo microscope at 200× magnification. Intervals for charcoal particle sizes were defined following the methods proposed by Clark et al. (1998) and Whitlock and Larsen (2001). Charcoal fragments smaller than 64 μm were not quantified to reduce the effect of particles that may have broken during chemical processing. Charcoal particle fractions between 64 and 102 μm, and >180 μm were quantified to reconstruct the regional and local fire signals, respectively (Whitlock and Larsen, 2001). We did not quantify the 64–102 μm fraction to restrict overlapping between local and regional signals. As charcoal morphology has proven useful to track fuel sources (Prince et al., 2018; Dussol et al., 2021), particles larger than 180 μm were classified according to seven fuel-source categories and 27 well-defined morphological subclasses drawing upon particle geometry, texture, and luster (Mustaphi and Pisaric, 2014).
We performed charcoal time-series analysis using CharAnalysis (Higuera et al., 2009). We produced equally spaced intervals and calculated charcoal accumulation rates (CHAR, particles cm–2 yr–1) by interpolating charcoal counts to the median sample resolution (ca. 200 years). We identified charcoal peaks as the positive residuals exceeding the 95th percentile threshold of a locally fitted Gaussian mixture CHAR background model (smoothed to 1,000 years). Peaks that resulted from statistically insignificant variations in CHAR were also identifiable in CharAnalysis. If the maximum count in a CHAR peak had a >5% chance of coming from the same Poisson-distributed population as the minimum charcoal count of the preceding 75 years, then the peak was rejected (Higuera et al., 2010).
Detrended Correspondence Analysis
A detrended correspondence analysis (DCA) was used to summarize ecological changes in the pollen record. The DCA is relatively easy to interpret as its scores are given in units of standard deviations (SD). The resulting scores can be interpreted directly in terms of ecological turnover, either between contiguous samples (to identify periods of ecological instability from the Euclidean distance between them) or by assigning a reference point (to compare moments with similar ecological conditions) (Correa-Metrio et al., 2014). The DCA was used to infer temporal relationships between vegetation and environmental changes. For instance, one taxon can assemble itself with any other taxon with the closest environmental response at a specific time, even if such a relationship does not exist today (Urrego et al., 2016). Because the resulting DCA ecological affinities rely only on the variability within the dataset (Urrego et al., 2016), DCA analysis is also a convenient method for understanding ecological novelty over time. We performed DCA analysis and distance calculations using the most representative pollen taxa of MAR 19-II (i.e., those present at >10% of the pollen sum), employing the package vegan (Oksanen et al., 2018) developed for the statistical software R (R Core Team, 2020).
Regime Shift Detection
We performed a Sequential t-test Analysis of Regime Shifts (STARS) (Rodionov, 2004) using the Euclidean distances between contiguous samples that resulted from the DCA analysis, to determine the timing of changes that produced a new and persistent ecological phase. In the STARS method, a Regime Shift Index (RSI) is calculated to confirm or reject the existence of a regime change in each of the observations within a time series. STARS tests whether each observation is statistically different from the current regime, and then checks the result by performing the test on subsequent observations (Rodionov, 2004; Rodionov and Overland, 2005). Two parameters, the cut-off length, and the significant level p, would impact the results and the minimum duration of a regime. Considering the relatively long-term series of SD and referring to the discussion of Rodionov (2004), we chose the cut-off length of 9 and a p-value of 0.05. We conducted the regime shift analysis in R, using the package rshift (Room et al., 2022).
Results
Chronology
We established age control from seven bulk sediment samples for the upper 116 cm of the core MAR19-II (Drives 1 and 2). The basal age for Drive 1 of core MAR-19II was ca. 10.6 kyrBP (Supplementary Table 1), spanning most of the Holocene Epoch. Radiocarbon dating at the top of the core revealed that the most recent 1,500 years were missing. This likely resulted from the coring operation which started in the least perturbed parts of the sediments. Alternatively, this sedimentary inconsistency in the uppermost centimeters could be the result of stumping cattle grazing activities developed in Monquentiva until the 20th century (CAR, 2007). This gap was covered and complemented by the core MAR19-GI with a basal age of ca. 2.8 kyr BP. The pollen record from the most recent sample in MAR19-GI reflects the vegetation we observed in Monquentiva during fieldwork in 2019. The resulting age-depth model (Figure 2; Blaauw and Christeny, 2011) was created assuming a probability with a gamma distribution for calibrated ages within a 95% confidence interval. The core MAR19-II has an average temporal resolution between samples of ca. 240 yr (min: 80 yr, max: 450 yr), whereas MAR19-GI has an average resolution of ca. 200 yr (min: 75 yr, max: 290).
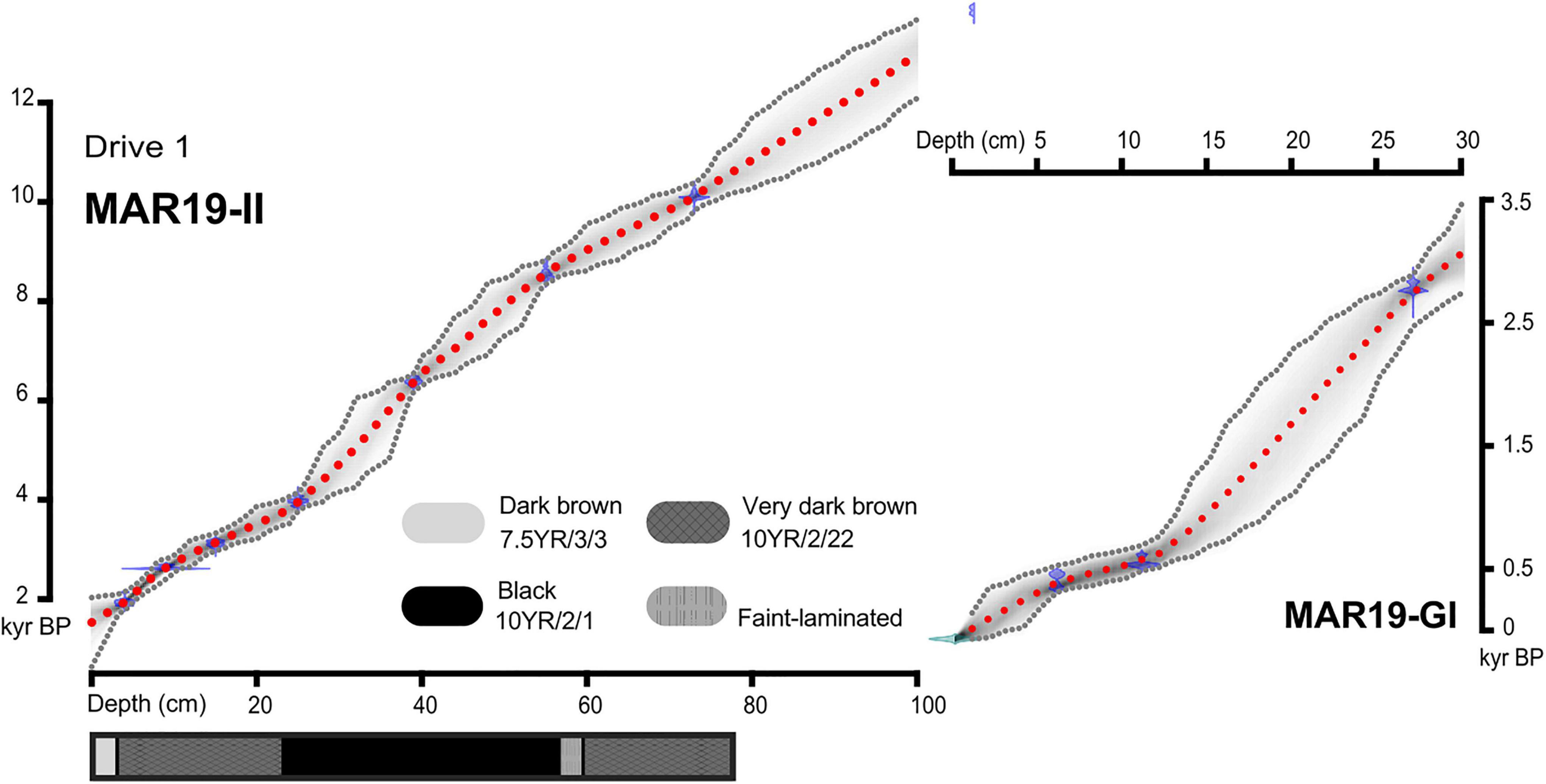
Figure 2. Age-depth model for cores MAR19-II and MAR19-G1 from Monquentiva calculated using rbacon (Blaauw and Christeny, 2011) and showing: calibrated radiocarbon ages (blue bows), all likely age-depth models (gray gradient); 95% confidence intervals (gray dotted lines), and the accepted model (red dotted lines).
Palynology
The pollen record was composed of 83 pollen types, nine pteridophyte spore types, five algae and four fungi spore types. The average count was 609 (minimum 376; maximum 2393) and the average pollen sum was 384 (min. 258; max. 612). Poaceae, Asteraceae, Weinmannia, Hedyosmum, Alnus, Apiaceae, Miconia and Morella were the most abundant pollen taxa. Among pteridophytes, Cyathea and Polypodium had remarkable high values. Other taxa like Plantago, Valeriana, and Isoëtes were also abundant. Less common, but relevant elements in Andean forests were Quercus humboldtii, Clusia, and Galium. Pollen and spore taxa present at above 10% of the pollen sum are shown in Figure 3. The pollen record was divided into four local pollen zones (ML1–ML4) following the grouping from CONISS (Grimm, 1987).
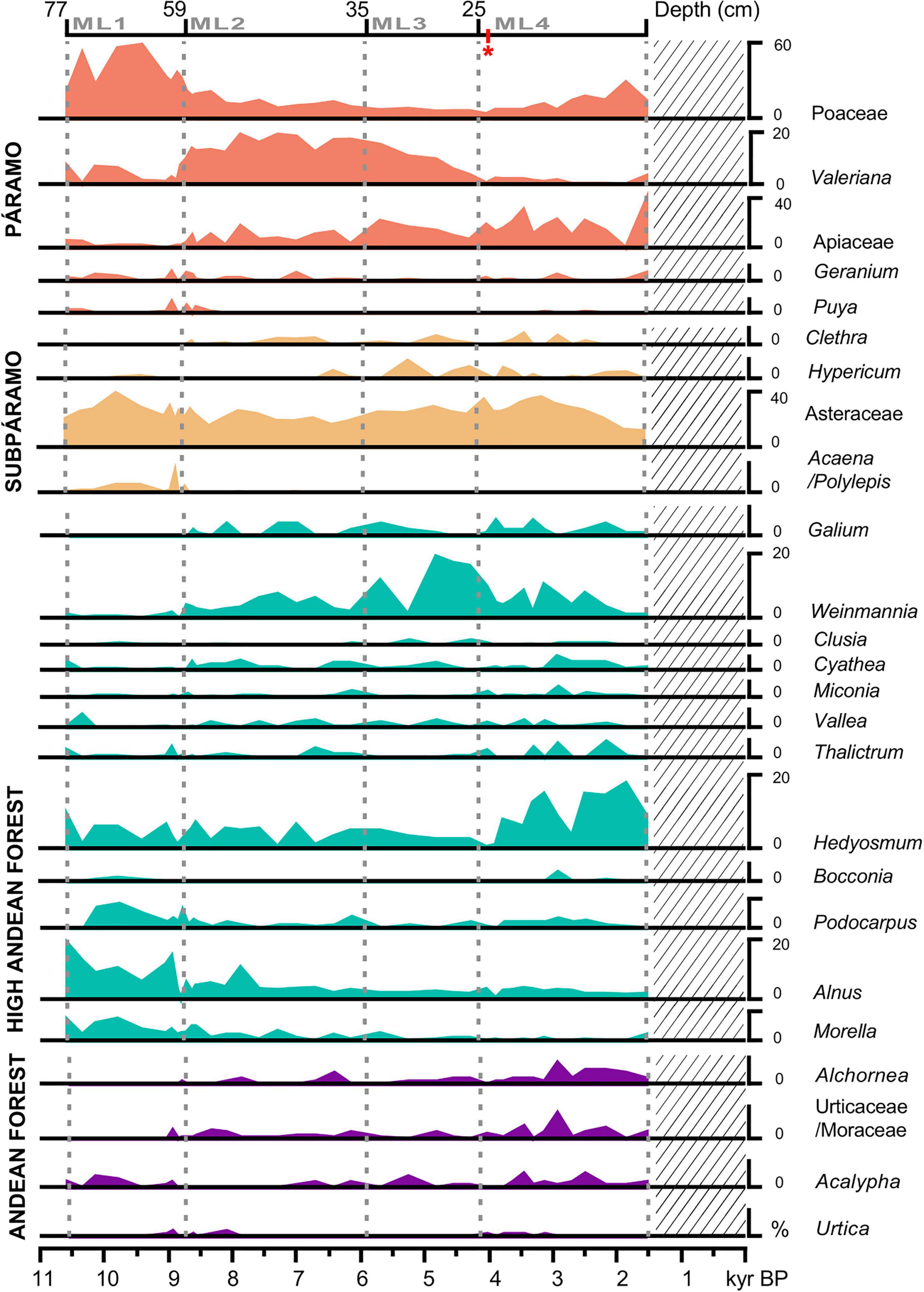
Figure 3. Summary pollen diagram of MAR19-II core from Monquentiva showing main regional taxa (>10%) organized by vegetation groups. Dashed gray lines indicate the limits for Monquentiva local pollen zones ML1 to ML4 from the CONISS zonation. The lattice denotes a gap in the MAR19-II sedimentary record. The red * indicates one sample where the pollen count was below 300 grains and reached 258 grains.
Zone ML1 (77–60 cm; 10.6–8.7 kyr BP)
Within this zone, pollen concentration fluctuated between 90,000 and 410,000 grains/cm3, reaching the lowest point at ca. 8.7 kyr BP (Supplementary Figure 1). This zone had the highest abundance values in the entire record for Poaceae (59%), Acaena/Polylepis (6%), Puya (2%), Gaultheria (3%), Alnus (20%), and Podocarpus (4%). Aquatic plants peaked at the interval 64–65 cm (ca. 9 kyr BP), while pteridophytes were almost absent.
Zone ML2 (60–36 cm; 8.7–6.1 kyr BP)
Zone ML2 was characterized by the fluctuation of the pollen concentration between nearly 118,000 and ∼321,000 grains/cm3. Poaceae steadily decreased (from 47 to 8%), while Valeriana (10 to 26%) and Clethra increased (0 to 3.3%) (Figure 3). Starting at 7.6 kyr BP, Alnus concentration values dropped to 3%, a value which was maintained through the rest of the sequence. Taxa such as Weinmannia and Hedyosmum fluctuated between 2 and 11%, and <1 and 4%, respectively (Figure 3). Also, Isoëtes abruptly decreased to 4% while Plantago surged from 3% at 8.7 kyr BP to 13% ca. 6.5 kyr BP (Figure 4).
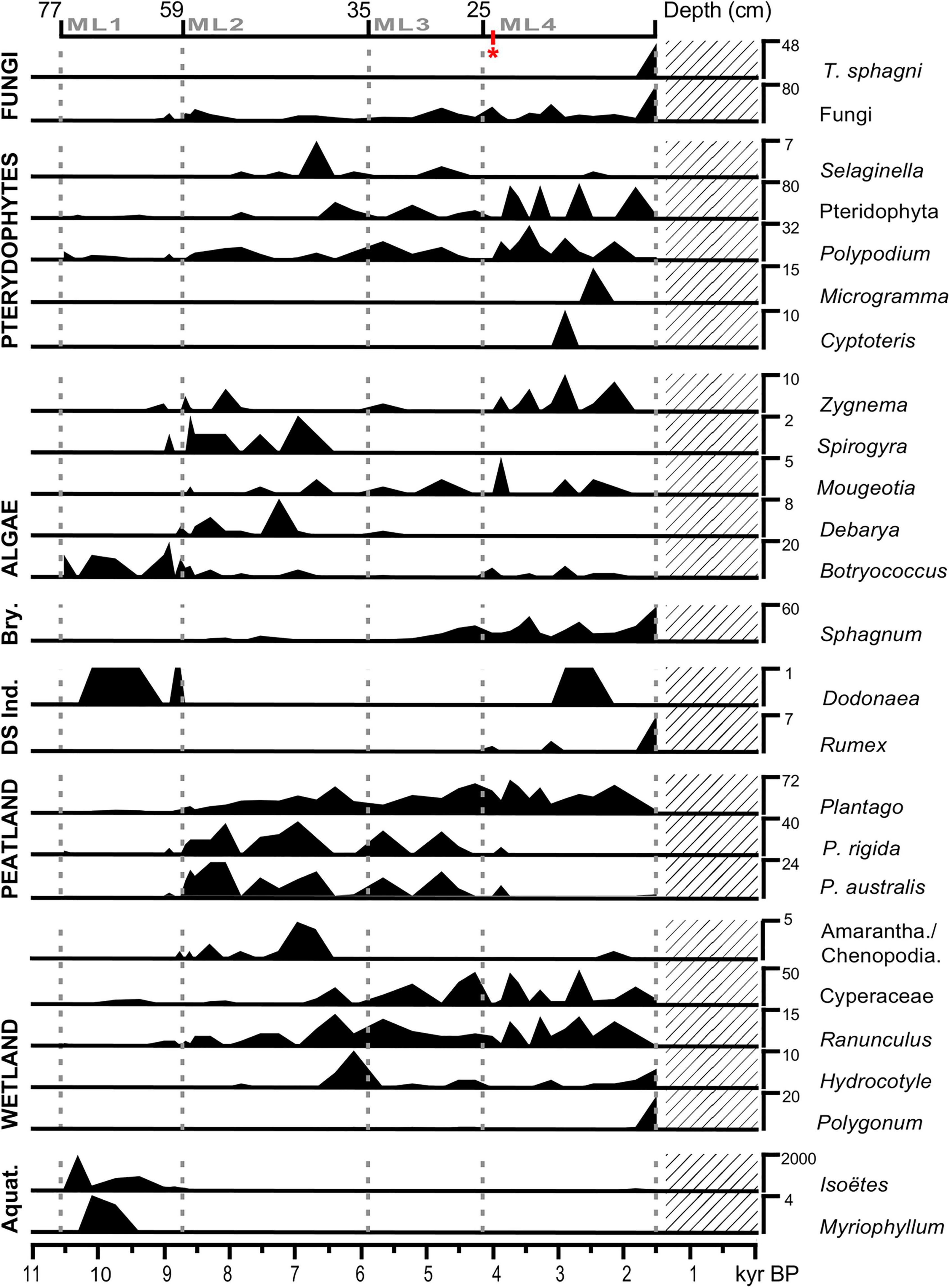
Figure 4. Summary pollen diagram of MAR19-II core from Monquentiva showing local taxa organized by vegetation groups and presented as counts. Dashed gray lines indicate the limits for local pollen zones ML1 to ML4 from the CONISS zonation. The lattice denotes a gap in the MAR19-II sedimentary record. The red * indicates one sample where the pollen count was below 300 grains and reached 258 grains. Bry., Bryophites; DS Ind., Disturbance Indicator; Aquat., Aquatic plants.
Zone ML3 (36–26 cm; 6.1–4.1 kyr BP)
Pollen concentration fluctuated between ∼144,000 at and 362,000 grains/cm3. Within the zone ML3, Valeriana declined from 18 to 1%, Hypericum fell from 8 to 2% and Poaceae hit a low point of 5.5%. Conversely, Weinmannia peaked at 17% and Asteraceae rose to 37% at ca. 4.1 kyr BP and a surge of Sphagnum from 1 to 19%, and Polypodium up to 8.5%, took place (Figure 4).
Zone ML4 (26–0 cm; 4.1–1.5 kyr BP)
Pollen concentration reached its maximum of 552,000 grains/cm3 at ca. 3.1 kyr BP. At the intersection between ML3 and ML4, Rumex was recorded for the first time. Hedyosmum and Weinmannia oscillated between 1 and 12%, Poaceae recovered from 5% to almost 40% and Asteraceae remained relatively stable with values around 29%. In this zone, fungi reached their highest values, along with a peak of Polypodium (22%) and other ferns (Figure 4).
Core MAR19-GI (27–0 cm; 2.6–0 kyr BP)
Pollen taxa such as Weinmannia and Hedyomum remained low (<10%) throughout the zone, but decreased considerably after 0.5 kyr BP. Contrarily, Poaceae reached values of ca. 30% and Rumex was recorded through the entire zone with values exceeding 5% from 0.5 kyr BP onwards (Supplementary Figure 2).
Charcoal Record
Charcoal concentrations of both particle fractions showed two main stages: one of low and stable concentrations from 10.6 to ca. 4.1 kyr BP followed by another of high and variable concentrations from 4.1 kyr BP to 1.5 kyr BP. Through the first stage, CharAnalysis identified significant microcharcoal (64–102 μm) peaks every ∼600 years (Figure 5), whereas significant macrocharcoal (>180 μm) peaks occurred at 8.6 and 6.4 kyr BP. The second stage was characterized by seven charcoal peaks detected through CharAnalysis for the macrocharcoal, with three significant peaks at 3.8, 3.1, and 2.9 kyr BP, coinciding with a period when macrocharcoal concentrations grew gradually (Figure 5).
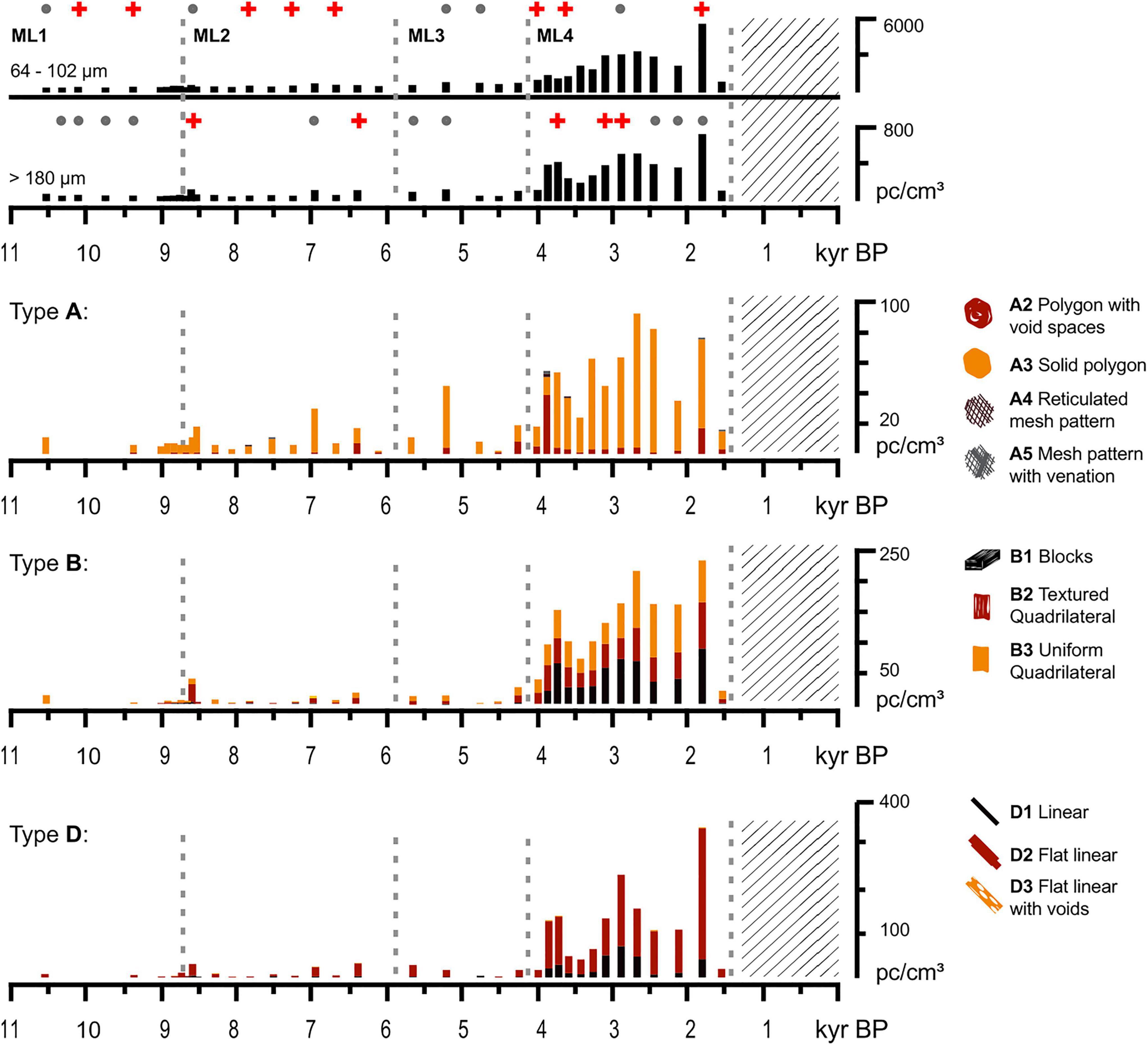
Figure 5. Charcoal concentration and morphotypes found in MAR19-II core from Monquentiva. The top two panels show total charcoal concentration for micro (64–102 μm) and macrocharcoal (>180 μm). Red crosses and gray dots represent significant and non-significant fire peaks from CharAnalysis (Higuera et al., 2009), respectively. The bottom three panels show most abundant charcoal morphotypes from MAR19-II core presented as concentration. Dashed gray lines indicate the limits for Monquentiva local pollen zones ML1 to ML4 from the CONISS zonation. The lattice denotes a gap in the MAR19-II sedimentary record.
Twenty-three charcoal morphotypes were observed, being polygonal, rectangular, and elongate geometries (classes A, B, and D) the most abundant (Figure 5). All the samples studied had some amount of charcoal, but morphotypes from class F (Mustaphi and Pisaric, 2014) were not observed (Supplementary Figure 3). Before 8.7 kyr BP, charcoal counts were the lowest of the record and did not exhibit major changes. Apart from microcharcoal peaks at ca. 10 and 9.4 kyr BP, charcoal amounts did not show a distinct signal (Figure 5). Throughout the Mid-Holocene micro and macrocharcoal concentrations increased considerably three times: ca. 7.0, 6.4, and 5.4 kyr BP being A3, B2, and D2 (Figure 5) the most relevant types during those periods. From ca. 4.1 kyr BP onwards, microcharcoal concentration increased, reaching its maximum value at 1.8 kyr BP, and the most prominent macrocharcoal types were A3, B1, B2, B3, D1, and D2 (Figure 5). Finally, after 1.8 kyr BP, macro and microcharcoal concentration experienced a 10-fold decrease, but types D2, B1, B2, B3, and A3 remained abundant. Charcoal concentrations recorded in MAR19-GI fluctuated but increased considerably after 0.5 kyr BP (Supplementary Figure 2).
Multivariate Analysis
The DCA analysis of the fossil pollen record grouped taxa along Axis 1 (eigenvalue 0.22, axis length 1.74) and Axis 2 (eigenvalue 0.15, axis length 1.42). The pollen taxa with the highest scores in Axis 1 were Acaena/Polylepis, Puya, Poaceae and Styloceras at the positive end, and Weinmannia, Miconia and Symplocos at the negative end (Figure 6). Along Axis 2, Plantago rigida and Valeriana attained the highest positive scores, whereas Rumex, Polygonum, Bocconia, and Cyperaceae were distributed over the negative extreme of the Axis (Figure 6). Valeriana, Geranium, and Asteraceae gravitated around the center of the DCA plot. The DCA ordination displayed a clear separation along Axis 1, between samples from the Early Holocene (i.e., positive end) and the Late Holocene (i.e., negative end) (Figure 6).
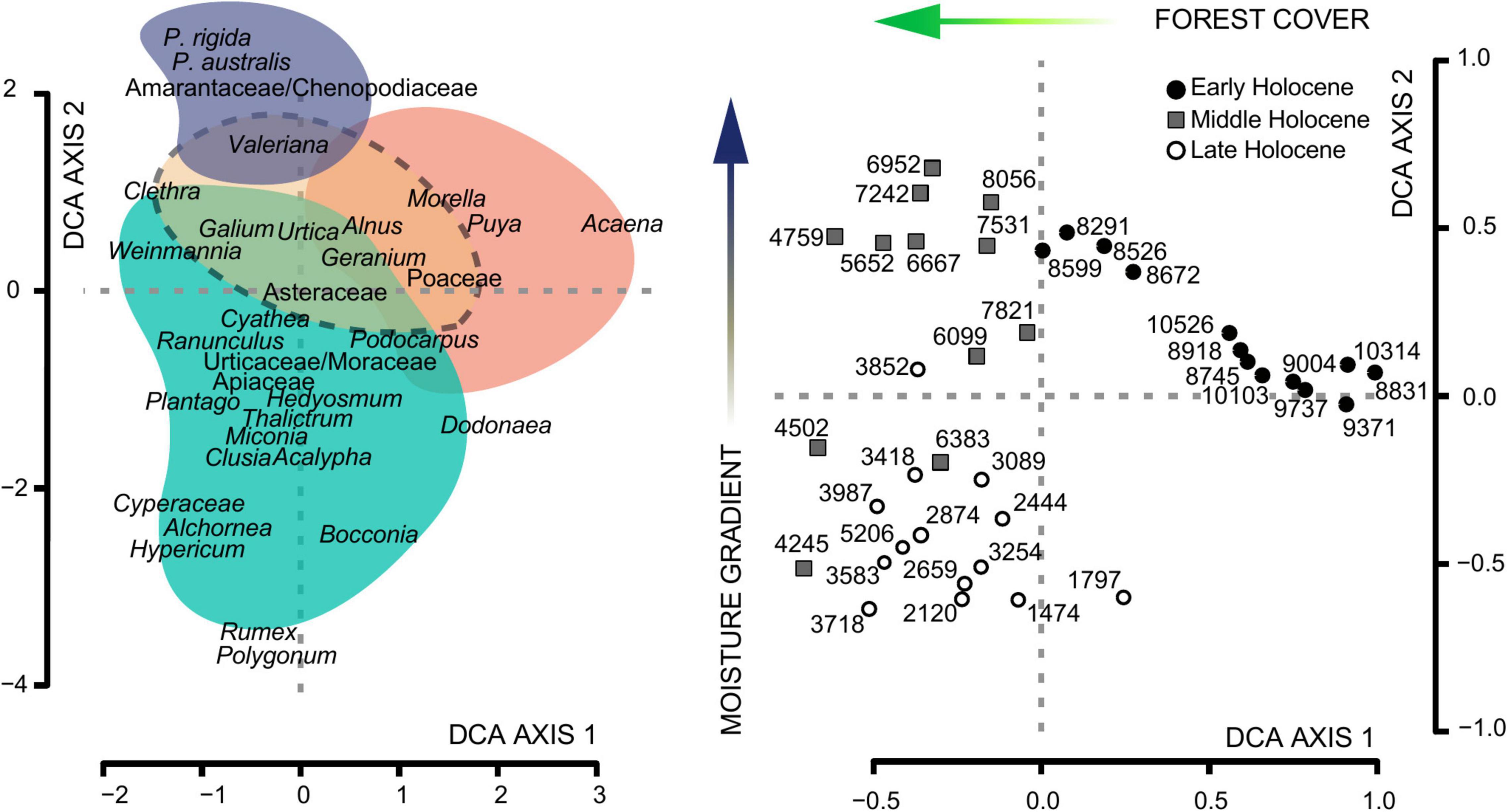
Figure 6. Detrended Correspondence Analysis (DCA) of the pollen record from MAR19-II core from Monquentiva. The left panel shows the DCA species scores for pollen taxa with shaded areas representing main vegetation groups: Páramo (orange); Subpáramo (coral pink); High Andean Forest (cyan); Peatland (purple). The right panel shows DCA sample scores with ages in years before present (BP). Arrows indicate the direction of interpreted environmental variables.
The DCA sample scores plotted against age showed positive values in Axis 1 until ca. 8.4 kyr BP. After this time, scores maintained negative values hitting a low point at ca. 4.4 kyr BP (Figure 7). In contrast, Axis 2 exhibited a fluctuating pattern which started having positive DCA score values after ca. 6.5 kyr BP (Figure 7). The Euclidean distance between contiguous samples showed highly variable scores between ca. 6.8 and 3.8 kyr BP with values up to 0.9 standard deviations (SD). Such timing coincided with the two regime shifts identified by the STARS algorithm (Figure 7). Apart from this period, SD values remained below 0.6. The distance between the oldest (10.6 kyr BP) and the most recent sample (1.5 kyr BP) was 1.2 SD. A turning point was recorded ca. 6.5 kyr BP along with a sustained difference lower than 1.0 SD from 4.6 kyr BP. Finally, after 3.8 kyr BP SD values decreased progressively up to the present (Figure 7).
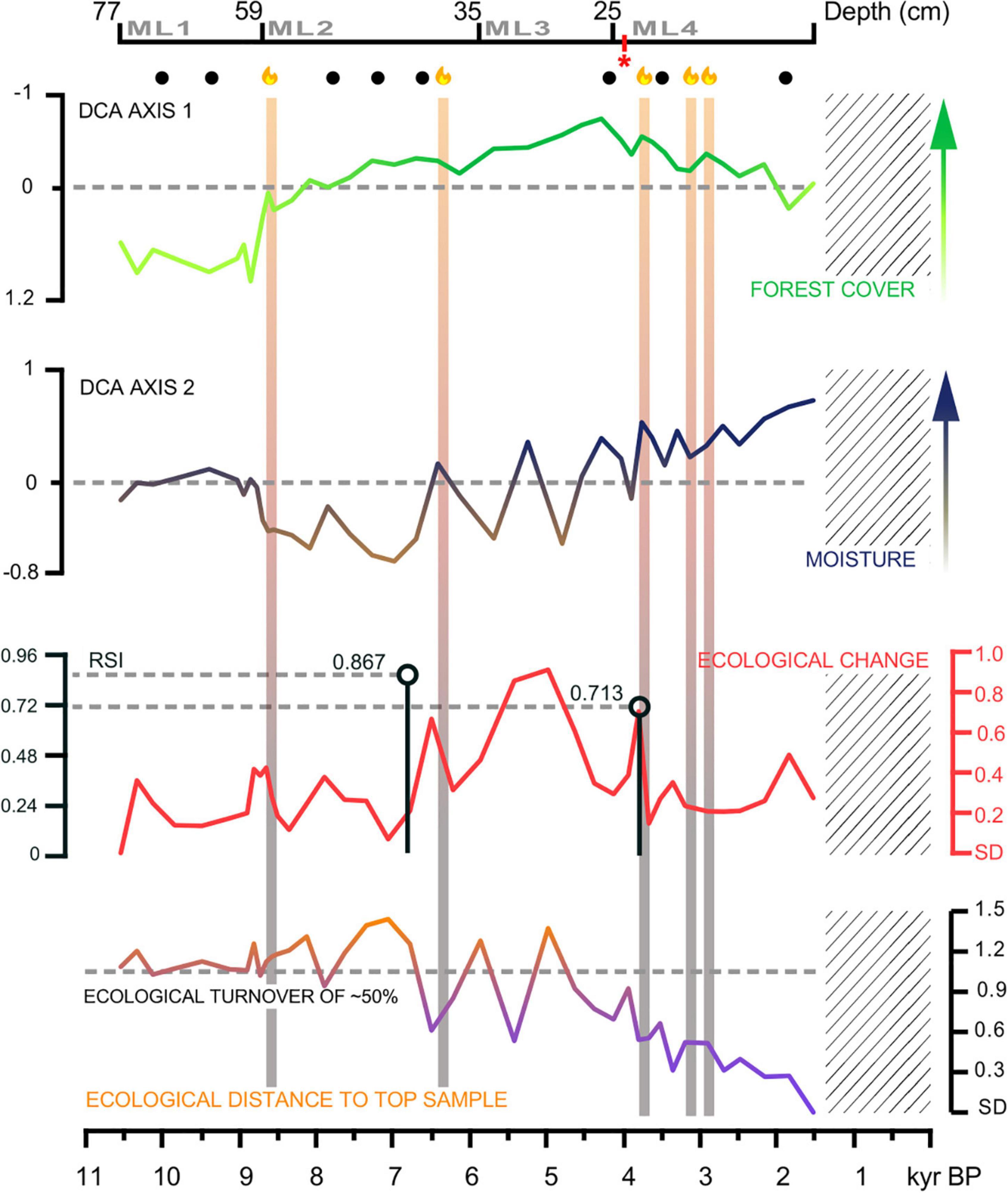
Figure 7. DCA scores and ecological change metrics presented as number of Standard Deviations (SD) and plotted against time for MAR19-II record from Monquentiva. The two top panels show DCA ordination axis scores for Axis 1 and Axis 2. The arrows indicate interpreted environmental variables. The third panel shows Euclidean distance calculated between consecutive samples and used as a metric for ecological change. The black lines indicate significant shifts identified with the Regime Shift Index (RSI) derived from the STARS method (Rodionov, 2004). The bottom panel shows Euclidean distance calculated between the top sample in MAR19-II and all other samples, used as a metric for ecological similarity. The dashed line represents SD = 1 and indicates an ecological turnover of 50%. The color intensity represents the relative magnitude of ecological turnover, with purple indicating the most novel plant communities. The vertical bars highlight fire peaks from CharAnalysis (Higuera et al., 2009), with fire flames indicating local fires and black dots indicating regional fire peaks. The lattice denotes a gap in the MAR19-II sedimentary record. The red * indicates one sample where the pollen count was below 300 grains and reached 258 grains.
Discussion
Regional and Local Vegetation Changes in Monquentiva Through the Holocene
Monquentiva was dominated by Páramo vegetation between ca. 10.6 and 8.7 kyr BP, with a high abundance of Poaceae, Puya, Geranium and other open vegetation (Figure 3). The high presence of Alnus before ca. 8.7 kyr BP might indicate poorly drained soils (van’t Veer et al., 2000) or be the result of over-representation as Alnus is known to be a generous pollen producer (Weng et al., 2004). We did not observe Alnus in the Monquentiva peatbog, suggesting Alnus does not represent a successional hydrarch stage at this elevation. However, Alnus occurrence along with a low abundance of other Andean Forest elements (i.e., Acalypha and Urticaceae/Moraceae) probably suggests open areas where pollen from forests at lower altitudes could be transported through rising air masses (Weng et al., 2004; Hagemans et al., 2019). Although Páramo vegetation around Monquentiva is important today (Avella-M et al., 2014), it occupies a smaller area than it did at the start of the Holocene. The dominance of Páramo pollen taxa during the Early Holocene is likely related to decreased temperatures at the latter stages of the El Abra Stadial (11 to 10 kyr BP); equivalent to the Younger Dryas Stadial (van der Hammen and Hooghiemstra, 1995). Such an event is known for being a cooling phase of up to 4°C compared to the temperature of the preceding Guantiva Interstadial in northern South America (Kuhry et al., 1993). Reduced temperatures are supported by the predominance of Poaceae and Asteraceae in the Monquentiva record (Figure 3). Although the high abundance of these taxa might also reflect increased aridity (Markgraf, 1993), the presence of Acaena/Polylepis supports the occurrence of a cold phase (van’t Veer et al., 2000).
Monquentiva and the eastern flank of the CCO appear to have been wet before 8.7 kyr BP, unlike the CCO western flank. The persistence of aquatic vegetation such as Isoëtes and Myriophyllum, and the presence of Botryococcus suggested the presence of water bodies in Monquentiva. High water levels at Monquentiva contrast with low water levels recorded in Lake Fúquene, located ca. 1,000 m downslope from Monquentiva on the western flank of the CCO (Vélez et al., 2006). Reduced lake levels at Fúquene have been linked to a dry phase in the central part of the CCO (van’t Veer et al., 2000; Vélez et al., 2006) and were not recorded in Monquentiva. These differences in climatic histories highlight the variability in moisture distribution in the Northern Andes. Uneven Andean moisture distribution in the CCO can be explained by the orographic control of the cordillera on regional atmospheric circulation and precipitation distribution (Saylor et al., 2009). Today, the position of the cordillera leads to the discharge of moisture-laden winds on its eastern flank and a moisture deficit on its highest and central parts (Mora et al., 2008). The persistence of this hydroclimatic pattern in the past has been suggested to explain the antiphase behavior between the central part of the CCO and its eastern flank during the Early and Late Holocene (Martin et al., 1997; van’t Veer et al., 2000; Bird et al., 2017). The orographic effect in the CCO is also suggested to impose its influence on today’s local precipitation over the migration patterns of the ITCZ (van’t Veer et al., 2000).
Between 8.7 and 6.1 kyr BP the history of Monquentiva was marked by a compositional shift in the Páramo and the expansion of High Andean Forest vegetation. Arboreal taxa such as Weinmannia, Hedyosmum, Vallea, and Galium were the most prominent elements in the forests surrounding Monquentiva. Since Weinmannia has a poor dispersion outside forested areas (Olivera-Moscol et al., 2009) its pollen abundance indicates a local presence and marks the development and ascent of the High Andean Forest on the surrounding slopes. The climatic conditions resulted in a smaller water body. Plantago pads were established around seasonally flooded patches and shallow waters with Cyperaceae, Ranunculus and Hydrocotyle. The flat areas, seasonally flooded, restricted the distribution of Poaceae but favored the expansion of Valeriana and the development of wetland vegetation for about 4,500 years (Figure 4). The onset of Mid-Holocene warmer temperatures and reduced precipitation (Marchant et al., 2001), enhanced the accumulation of organic matter evidenced by the consolidated peat layer found in the sedimentary record of Monquentiva starting at ca. 8.6 kyr BP (depth 56 cm).
Between 6.1 and ca. 4.1 kyr BP the sustained drop of Valeriana along with the rise of Cyperaceae suggests a contraction of the water body and the subsequent allocation of water in small local patches (van’t Veer et al., 2000). The surge of Sphagnum further supports this inference as it indicates the colonization of waterlogged soils around small water puddles, similar to those found in the study site today (Avella-M et al., 2014). By 5.9 kyr BP, the expansion of Weinmannia indicated warmer temperatures. Weinmannia-dominated forests surrounded Subpáramo grasses and shrubs, as indicated by the spread of Hypericum and Asteraceae (Rangel-Ch et al., 1997). The prevalence of warmer and drier conditions during the latter stages of the Mid-Holocene has also been recorded regionally in the Colombian highlands and lowlands (Marchant et al., 2001). Temperature fluctuations during this time are thought to be little in comparison to full glacial conditions (Marchant et al., 2001). Consequently, moisture fluctuations and regional atmospheric circulation probably exerted a critical role in the contrasting responses of the vegetation in the Northern Andes through the Mid-Holocene.
The Late Holocene was marked by the prevalence of High Andean forests, which were dominated by Hedyosmum, Podocarpus, and Weinmannia. The presence of these three taxa indicated forest advancing upslope (Bogotá-A, 2011) and developing in the surrounding slopes of Monquentiva. The gradual recovery of Páramo vegetation coincided with the drop of Supáramo vegetation and the attenuation of warm and dry conditions. This was supported by the increasing trend of Botryococcus and Isoëtes. The prevalence of wetter conditions ca. 3.2 kyr BP coincided with recorded variations in moisture contents at the eastern slopes of the CCO from ca. 3.5 kyr BP [i.e., Ubaque lake; (Bird et al., 2017)] but differs to those at the western parts of the Andes [see González-Carranza et al. (2012)].
The pollen record also suggested the influence of human activities in the vegetation of Monquentiva during the Late Holocene. The presence of Rumex from ca. 4.1 kyr BP in the study site could indicate the presence of degraded soils due to anthropic intervention (Bakker et al., 2008). The development of crops and the increase in plant consumption between 4.0 and 3.5 kyr BP by early Andean occupants (van der Hammen et al., 1990) could partially explain the rise of Apiaceae in the pollen record of Moquentiva during the Late Holocene. Regionally, the increase in charcoal concentration and the presence of Zea mays in the Pantano de Vargas (Gómez et al., 2007) suggested pre-Hispanic societies established crops and used fire regularly in the CCO after 3.8 kyr BP. No maize pollen grains were found in Monquentiva, and extended counts were not performed because a high-elevation large peatbog like Monquentiva is unlikely to have been a suitable site for corn cultivation. However, the absence of corn cannot be used to indicate the lack of human activity in this part of the CCO. Instead, the absence of maize pollen might be the result of the poor stratigraphical representation derived from short pollen dispersal distances of corn (Taylor et al., 2013). No maize grains could also exemplify the mobile nature of early human groups (Rothhammer and Dillehay, 2009), who likely had to reach lower elevations with better climatic conditions for maize farming (Langebaek, 2019). Holocene agricultural practices by early people in the Colombian Andes is beyond the scope of this research, but future paleoecological work on cultigens adapted to cold conditions could improve our knowledge of early human occupation in the high Andes.
The occurrence of Rumex over the last 1,500 years, pointed to the development of human activities as a feature of the Monquentiva landscape (Supplementary Figure 2). However, the invariable and low percentages of Poaceae, besides the steady presence of Weinmannia and Hedyosmum, indicated reduced forest intervention between 1.5 and 0.5 kyr BP, when compared to the onset of the Late Holocene. Such limited anthropogenic imprint around this time might reflect the fact that Monquentiva was visited occasionally rather than being a place of permanent residency. This interpretation is consistent with Muisca groups reunited around large, urbanized centers as those described from nearby sites in the CCO by this time (Langebaek, 2019). Contrastingly, after 0.5 kyr BP, Poaceae expansion was concomitant with forest clearing, as depicted by a Weinmannia, Clusia, and Hedyosmum decline, suggesting more intense human disruption.
Fire History in Monquentiva Through the Holocene
The sedimentary charcoal record from Monquentiva showed sustained fire activity throughout the Holocene. We observed coupled micro and macrocharcoal signals indicating that local and regional fire activity covaried and signal a non-human source of ignition during the Early Holocene. Charcoal concentration during the Early and Middle Holocene suggested a moderate fire history with sizable activity ca. 8.6, 6.6, and 6.4 kyr BP (Figure 5). Around 8.6 kyr BP, fire activity coincided with the minimum values of Isoëtes and an increase of Plantago suggesting that such fire increment was related to warm and drier conditions post the El Abra Stadial (Hooghiemstra and van der Hammen, 1993). These fires acted locally on grasslands and shrublands, as suggested by the low frequency of woody materials and the dominance of morphotypes D2 and B2, derived from monocotyledonous leaves (Figure 5) (Mustaphi and Pisaric, 2014). Through the Mid-Holocene, significant fires occurred with a frequency of up to ∼600 years (Figure 5) and a high concentration of types A3 (punky wood and leaves). This increase of monocotyledonous leaves-derived particles during the Middle Holocene suggested the source of the burning biomass remained outside forest areas.
Did Humans Ignite Fires in Monquentiva?
The use of fire by humans in the CCO appears to have been limited before ca. 4.1 kyr BP (Figure 5), despite their arrival at ca. 12 kyr BP in the northern Andes (Correal, 1989; Gnecco and Mora, 1997; Gnecco, 2003). Archeological evidence during the Early and Middle Holocene suggests food foraging was the primary subsistence strategy (Delgado, 2018), which potentially limited burning practices. Mid-Holocene human presence was likely restricted as adverse climatic conditions caused demographic contractions and population displacements of hunter-gatherer groups (Delgado, 2012). However, as permanent residents have been recorded in the CCO from ca. 7.5 kyr BP (Archila et al., 2021), it is probable that if humans were using fire, this was at a limited scale in the surrounding areas of Monquentiva during the Mid-Holocene. This is consistent with the decoupling of local and regional charcoal signals ca. 6.5 kyr BP suggesting Mid-Holocene low-scale anthropogenic fires around Monquentiva.
Both regional and local fire activity rose during the Late Holocene at Monquentiva, indicated by increased micro and macrocharcoal concentrations. By ca. 4.1 kyr BP, the rise in morphotypes A2, B1, B3, and C4 indicated a shift in the fuel source to woody materials, which along with a drop in Weinmannia, suggest selective wood extraction and burning by the early Late Holocene. Similarly, Poaceae gradually replacing Weinmannia, a known timber species, might be a sign of downslope Páramo expansion as a response to human disturbance. Charcoal peaks, a statistically robust proxy for local fire episodes (Higuera et al., 2010), were recorded at ca. 3.8 and 2.6 kyr BP followed by a subtle decline toward the present until ca. 1.8 kyr BP when fires escalated. Although the two charcoal fractions followed similar increasing trends, a clearer decoupling of their signals between 3.8 and 3.2 kyr BP suggested that human activities were indeed an effective ignition source. The rise of wood-derived morphotypes pointed to these fires burning the arboreal taxa from surrounding High Andean Forests and the shrubby Subpáramo in Monquentiva, aside from acting over the open Páramo vegetation. Although the extent to which humans inhabited Monquentiva through the Holocene needs further research, the demand for wood in the area and the associated forest transformation became evident during the Late Holocene. Such a timber demand and forest clearing by fire seemed especially important during the last 0.5 kyr BP, after the arrival of European explorations (Martin, 2005) and the resulting demographic and land-use changes (Etter and van Wyngaarden, 2000).
Did Climate Induce Fires in Monquentiva?
The increased fire activity in the Middle and Late Holocene in Monquentiva might reflect a marked influence of ENSO. The effect of Holocene ENSO on hydroclimatic patterns at the Northern Andes have already been detected in other parts of the eastern flank of the CCO, and it is believed to be a substantial control on fire activity over long time scales (Bird et al., 2017). The general trend of ENSO during the Holocene are severe events at ca. 1,500-year intervals starting at 7.0 kyr BP, followed by an increase in ENSO activity between ca. 4.2 kyr BP and ∼1.2 kyr BP, and a subsequent reduction toward the present (Moy et al., 2002; Toth et al., 2012; Hagemans et al., 2021). This coincides with fire events at higher frequencies from 7.0 kyr BP, their surge at 4.1 kyr BP, and two notable peaks at 3.8 and 2.6 kyr BP in the Monquentiva record and suggest a climate control on fire activity through the Holocene.
The increase in Mid-Holocene fire activity probably resulted from human activities as a source of ignition and was facilitated by climatic conditions. Mid-Holocene dry conditions reduced water levels at Monquentiva, allowing the expansion of Subpáramo vegetation after ca. 6.1 kyr BP (Figure 3). Reduced moisture drove plant compositional changes, favoring the development of the open vegetation that today lies in Monquentiva. This open vegetation is better adapted to dry conditions and is known today to be fire-prone (Horn and Kappelle, 2009; Ramsay, 2014). Thus, fuel availability likely increased around Monquentiva due to changes in the vegetation composition. As such, the current vegetation of Monquentiva could be seen as a shifted baseline (Sylvester et al., 2017). Climatic conditioning continued long after ca. 4.1 kyr BP when increased ENSO activity (Haug et al., 2001; Moy et al., 2002; Gomez et al., 2004) enhanced drought and ultimately fire activity. Furthermore, the human influence on fires likely started over the Mid-Holocene and escalated through the Late Holocene, reaching high points at ca. 1.8 and 0.5 kyr BP, when its effects were imposed over climate conditions. Thus, increased drought due to intensification of ENSO and human intervention seem to have acted as coupled factors driving fires in Monquentiva, which combined served as local fire amplifiers after 4.1 kyr BP.
Ecological Novelty in Monquentiva
The ecological change index calculated between contiguous samples indicated that the largest change in the Holocene vegetation of Monquentiva came after ca. 6.8 kyr BP (Figure 7). The ecological distances of up to ∼0.9 standard deviations (SD) from ca. 5.4 kyr BP indicates the temporary establishment of plant communities with a species turnover close to 50%. These highly variable communities are likely the result of individualistic responses of pollen taxa to the Mid-Holocene changing environmental conditions. As climate shifted and fire occurred more frequently, rearrangement of species promoted their co-occurrence at different proportions as novel communities (Williams and Jackson, 2007). The Mid-Holocene novel communities of Monquentiva were characterized by a high abundance of Asteraceae and Weinmannia, and the lowest values of Poaceae and Valeriana (Figure 3). This episode represented well regional Mid-Holocene rapid rates of change in the forest composition in the southern tropical Andes (Urrego et al., 2010) and extreme dry phases in the Peruvian-Bolivian Altiplano (Baker et al., 2001).
The most pronounced ecological changes in Monquentiva were identified as two prominent regime shifts recorded at 6.8 and 3.8 kyr BP and coincided with fire peaks (Figure 7). Biomass burning is therefore likely to have been a major driver of ecological change in Monquentiva at a time when humans become a likely source of ignition. The timing of increased sedentism of early human groups in the CCO at 7.5 kyr BP (Archila et al., 2021) also supports our interpretation. Climatically, increased ENSO frequency from 7.0 kyr BP (Moy et al., 2002) also facilitated the ignition of the fires that had such a large impact on the Monquentiva vegetation. The second ecological regime shift identified at 3.8 kyr coincides with human-induced forest clearance and the agricultural land-use changes reported in the CCO (Gomez et al., 2004; Vélez et al., 2020). Such regime shifts in Monquentiva seem to replicate a continental trend of environmental disruption. Our results follow the continental tendency of accelerated rates of change (Urrego et al., 2009; Mottl et al., 2021) and suggest an escalation in the Holocene anthropic imprint on montane Andean ecosystems (Delgado, 2012).
The ecological distance between the most recent sample and the rest of the record suggests that plant communities that existed at 1.5 kyr BP in Monquentiva were compositionally novel when compared to Middle and Early Holocene vegetation (Figure 7). These novel plant communities formed in Monquentiva ca. 4.6 kyr BP after a 2,000-year period of ecological instability when the ecological distance to present showed large fluctuations. Such ecological instability likely represented the occurrence of the shifting conditions needed for the emergence of current ecological communities and a prelude to what could be called an “alternative stable state” (Scheffer et al., 2001). The Late Holocene is marked by the return of relatively stable ecological conditions, as indicated by low variation in ecological distances between consecutive pollen assemblages (Figure 7). This reduced variability in ecological change suggests that the novel plant communities would have been adapted to more frequent fires and human presence.
Conclusion
The Holocene vegetation history of Monquentiva highlights its sensitivity to fluctuating climates, human intervention, and fire activity. Fires occurred in the area throughout the Holocene Epoch and were initially induced by climate during the Early Holocene and later induced by human activities and facilitated by ENSO. Whether ignited by people or climate, fires have played a fundamental role in shaping Monquentiva plant communities. Fire activity resulted in two ecological regime shifts at 6.8 and 3.8 kyr BP at Monquentiva. Our results underline the importance of fire activity on high Andean landscapes and highlight its role in shaping the vegetation we find in the CCO today.
Vegetation responses to fire, climate, and human activities in Monquentiva resulted in compositionally novel plant communities that were established ca. 4,000 years ago. Under current climate destabilization, long-term ecological responses to external changes such as those described for the Monquentiva area demand urgent actions. The increased likelihood of a more frequent and intense ecological turnover in the coming decades is imminent as uncontrolled land-use changes are widespread in regions of high biological diversity and complexity like the CCO. Forest fragmentation, uncontrolled fires and other alarming dynamics will surely prompt instability as has been registered for the Middle Holocene if no concrete management actions are taken. Management efforts must therefore include explicit strategies (e.g., fire control and prevention, ecosystem services, etc.) that consider the complexities of Andean ecosystems, thus promoting the survival of their biological and cultural heritage.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation. Any queries should be directed towards the corresponding author(s).
Author Contributions
IE, FF-G, DA, and DU conceived the research and approach, conducted the analyses, and led the manuscript writing. DU acquired the funding. IE, IC, and CR collected and curated the data. AR, FF-G, and DU developed the Regime Shift Index code for palaeoecological analyses. JB contributed to site selection and research design. All authors contributed to the writing of the manuscript and approved it for publication.
Funding
Support for this research came from the UK Natural Environment Research Council (NERC) grant number NE/R017980/1. AR was supported by a NERC GW4+ Undergraduate Research Experience Placement.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This manuscript is a product of the research project “BioResilience: Biodiversity resilience and ecosystem services in post-conflict socio-ecological systems in Colombia” (https://blogs.exeter.ac.uk/bioresilience/). The data presented here represent an exceptional effort on the part of researchers, field and lab assistants in Colombia and the United Kingdom. We particularly thank Don Elias Romero, Wilmar Romero, and the community of Monquentiva for receiving us and supporting us in different phases of the research. We thank the Alcaldía de Guatavita and CAR Cundinamarca for their help. Jack Oughton and Thya van den Berg are thanked for their assistance in the field. We are grateful to the personnel at the University of Exeter, especially to Ang Elliot, Joana Zaragoza-Castells, and Charlotte Durden for their assistance in the lab. We thank the ECOLMOD research group at the Universidad Nacional de Colombia and the Tropical Palaeoecology Lab at the University of Exeter for their support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.895152/full#supplementary-material
References
Aceituno, F. J., Loaiza, N., Delgado-Burbano, M. E., and Barrientos, G. (2013). The initial human settlement of Northwest South America during the Pleistocene/Holocene transition: synthesis and perspectives. Quat. Int. 301, 23–33. doi: 10.1016/j.quaint.2012.05.017
Aguilera, C. F., Armenteras, D., and González-Alonso, F. (2009). Geographic and temporal distribution of fire in Colombia using thermal anomalies data | Distribución geográfica y temporal de incendios en Colombia utilizando datos de anomalías térmicas. Caldasia 31, 303–318.
Archila, S., Groot, A. M., Ospina, J. P., Mejía, M., and Zorro, C. (2021). Dwelling the hill: traces of increasing sedentism in hunter-gatherers societies at Checua site, Colombia (9500-5052 cal BP). Quat. Int. 578, 102–119. doi: 10.1016/j.quaint.2020.07.040
Armenteras, D., Schneider, L., and Dávalos, L. M. (2019). Fires in protected areas reveal unforeseen costs of Colombian peace. Nat. Ecol. Evol. 3, 20–23. doi: 10.1038/s41559-018-0727-8
Armenteras-pascual, D., Retana-alumbreros, J., Molowny-horas, R., Roman-cuesta, R. M., Gonzalez-alonso, F., and Morales-rivas, M. (2011). Characterising fire spatial pattern interactions with climate and vegetation in Colombia. Agric. For. Meteorol. 151, 279–289. doi: 10.1016/j.agrformet.2010.11.002
Avella-M, A., Torres-R, S., Gómez-A, W., and Pardo-P, M. (2014). Los páramos y bosques altoandinos del pantano de Monquentiva o pantano de Martos (Guatavita, Cundinamarca, Colombia): caracterización ecológica y estado de conservación. Biota Colomb. 15, 3–39.
Baker, P. A., Seltzer, G. O., Fritz, S. C., Dunbar, R., Grove, M., Tapia, P. M., et al. (2001). The history of South American tropical precipitation for the past 25,000 years. Science 291, 640–643. doi: 10.1126/science.291.5504.640
Bakker, J., Olivera-Moscol, M., and Hooghiemstra, H. (2008). Holocene environmental change at the upper forest line in northern Ecuador. Holocene 18, 877–893. doi: 10.1177/0959683608093525
Bernal, R., Gradstein, S. R., and Celis, M. (2015). Catálogo de Plantas y Líquenes de Colombia. Available online at: http://catalogoplantasdecolombia.unal.edu.co/en/ (accessed January 3, 2019).
Bird, B. W., Rudloff, O., Escobar, J., Gilhooly, W. P., Correa-Metrio, A., Vélez, M., et al. (2017). Paleoclimate support for a persistent dry island effect in the Colombian Andes during the last 4700 years. Holocene 28, 217–228. doi: 10.1177/0959683617721324
Blaauw, M., and Christeny, J. A. (2011). Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474. doi: 10.1214/11-BA618
Bogotá-A, R. G. (2011). Pleistocene Centennial-Scale Vegetation, Environmental and Climatic Change in the Colombian Andes: Based on Biotic and Abiotic Analyss from Lake Fúquene Sediments. Ph.D. thesis. Amsterdam: Universiteit van Amsterdam.
Boom, A., Marchant, R., Hooghiemstra, H., and Sinninghe Damsté, J. S. (2002). CO2- and temperature-controlled altitudinal shifts of C4- and C3-dominated grasslands allow reconstruction of palaeoatmospheric pCO2. Palaeogeogr. Palaeoclimatol. Palaeoecol. 177, 151–168. doi: 10.1016/S0031-0182(01)00357-1
Bush, M. B. (2002). Distributional change and conservation on the Andean flank: a palaeoecological perspective. Glob. Ecol. Biogeogr. 11, 463–473. doi: 10.1046/j.1466-822X.2002.00305.x
Bush, M. B., and Weng, C. (2007). Introducing a new (freeware) tool for palynology. J. Biogeogr. 34, 377–380. doi: 10.1111/j.1365-2699.2006.01645.x
Cano, M. C., López, C. E., and Méndez, R. (2021). Archaeological evidences on early peopling in the fluvio-volcanic Pereira-Armenia fan (Colombia): volcanic activity influence on cultural adaptation and depopulation events. Quat. Int. 578, 131–138. doi: 10.1016/j.quaint.2020.08.037
CAR (2007). Elaboración del Diagnostico, Prospectiva y Formulación Cuenca Hidrográfica del Río Gacheta Subcuenca Río Monquentiva. Bogotá: CAR.
Clark, J. S., Lynch, J., Stocks, B. J., and Goldammer, J. G. (1998). Relationships between charcoal particles in air and sediments in west-central Siberia. Holocene 8, 19–29. doi: 10.1191/095968398672501165
Colinvaux, P., De Oliveira, P. E., and Moreno Patiño, J. E. (1999). Amazon Pollen Manual and Atlas, 1st Edn. Amsterdam: Taylor & Francis.
Correal, G. (1989). Aguazuque: Evidencias de Cazadores, Recolectores y Plantadores en la Altiplanicie de la Cordillera Oriental. Bogotaì: Fundacion de Investigaciones Arqueologicas Nacionales. Banco de la República, 271.
Correa-Metrio, A., Dechnik, Y., Lozano-García, S., and Caballero, M. (2014). Detrended correspondence analysis: a useful tool to quantify ecological changes from fossil data sets. Bol. Soc. Geol. Mex. 66, 135–143. doi: 10.18268/BSGM2014v66n1a10
Corredor, V. E., and Terraza, R. (2015). Geología de la Plancha 228 Bogotá Noreste, Memoria Técnica. Bogotá: Servicio Geológico Colombiano.
Cuatrecasas, J. (1958). Aspectos de la vegetación natural de Colombia. Parte I. Rev. Acad. Colomb. Cien. Exactas Fís. Nat. 40, 221–264. doi: 10.18257/raccefyn.570
Delgado, M. (2012). Mid and Late Holocene population changes at the Sabana de Bogotá (Northern South America) inferred from skeletal morphology and radiocarbon chronology. Quat. Int. 256, 2–11. doi: 10.1016/j.quaint.2011.10.035
Delgado, M. (2018). Stable isotope evidence for dietary and cultural change over the Holocene at the Sabana de Bogotá region, Northern South America. Archaeol. Anthropol. Sci. 10, 817–832. doi: 10.1007/s12520-016-0403-3
Duque, A., Stevenson, P. R., and Feeley, K. J. (2015). Thermophilization of adult and juvenile tree communities in the northern tropical Andes. Proc. Natl. Acad. Sci. U.S.A. 112, 10744–10749. doi: 10.1073/pnas.1506570112
Dussol, L., Vannière, B., Purdue, L., Orange, F., Testé, M., Castanet, C., et al. (2021). How to highlight slash-and-burn agriculture in ancient soils? A modern baseline of agrarian fire imprint in the Guatemalan lowlands using charcoal particle analysis. J. Archaeol. Sci. Rep. 35:102725. doi: 10.1016/j.jasrep.2020.102725
Etter, A., and van Wyngaarden, W. (2000). Patterns of landscape transformation in Colombia, with emphasis in the Andean region. AMBIO J. Hum. Environ. 29, 432–439. doi: 10.1579/0044-7447-29.7.432
Etter, A., McAlpine, C., and Possingham, H. (2008). Historical patterns and drivers of landscape change in Colombia since 1500: a regionalized spatial approach. Ann. Assoc. Am. Geogr. 98, 2–23. doi: 10.1080/00045600701733911
Faegri, K., and Iversen, J. (1964). Textbook of Pollen Analysis, Segunda Edn. Copenhague: Hafner Publishing Co.
Feeley, K. J., Silman, M. R., Bush, M. B., Farfan, W., Cabrera, K. G., Malhi, Y., et al. (2011). Upslope migration of Andean trees. J. Biogeogr. 38, 783–791. doi: 10.1111/j.1365-2699.2010.02444.x
Gnecco, C. (2003). Against ecological reductionism: late Pleistocene hunter-gatherers in the tropical forests of northern South America. Quat. Int. 109–110, 13–21. doi: 10.1016/S1040-6182(02)00199-4
Gnecco, C., and Mora, S. (1997). Late Pleistocene/early Holocene tropical forest occupations at San Isidro and Peña Roja, Colombia. Antiquity 71, 683–690. doi: 10.1017/S0003598X00085409
Gómez, A., Berrío, J. C., Hooghiemstra, H., Becerra, M., and Marchant, R. (2007). A Holocene pollen record of vegetation change and human impact from Pantano de Vargas, an intra-Andean basin of Duitama, Colombia. Rev. Palaeobot. Palynol. 145, 143–157. doi: 10.1016/j.revpalbo.2006.10.002
Gomez, B., Carter, L., Trustrum, N. A., Palmer, A. S., and Roberts, A. P. (2004). El Nino-Southern Oscillation signal associated with middle Holocene climate change in intercorrelated terrestrial and marine sediment cores, North Island, New Zealand. Geology 32, 653–656. doi: 10.1130/G20720.1
González-Carranza, Z., Hooghiemstra, H., and Vélez, M. I. (2012). Major altitudinal shifts in Andean vegetation on the Amazonian flank show temporary loss of biota in the Holocene. Holocene 22, 1227–1241. doi: 10.1177/0959683612451183
Grimm, E. C. (1987). CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares *. Comput. Geosci. 13, 13–35. doi: 10.1016/0098-3004(87)90022-7
Groot, M. H. M., Bogotá, R. G., Bogotá, R. G., Lourens, L. J., Hooghiemstra, H., Vriend, M., et al. (2011). Ultra-high resolution pollen record from the northern Andes reveals rapid shifts in montane climates within the last two glacial cycles. Clim. Past 7, 299–316. doi: 10.5194/cp-7-299-2011
Hagemans, K., Nooren, K., de Haas, T., Córdova, M., Hennekam, R., Stekelenburg, M. C. A., et al. (2021). Patterns of alluvial deposition in Andean lake consistent with ENSO trigger. Quat. Sci. Rev. 259:106900. doi: 10.1016/j.quascirev.2021.106900
Hagemans, K., Tóth, C. D., Ormaza, M., Gosling, W. D., Urrego, D. H., León-Yánez, S., et al. (2019). Modern pollen-vegetation relationships along a steep temperature gradient in the tropical Andes of Ecuador. Quat. Res. (U. S.) 92, 1–13. doi: 10.1017/qua.2019.4
Haug, G. H., Hughen, K. A., Sigman, D. M., Peterson, L. C., and Röhl, U. (2001). Southward migration of the intertropical convergence zone through the Holocene. Science 293, 1304–1308. doi: 10.1126/science.1059725
Higuera, P. E., Brubaker, L. B., Anderson, P. M., Hu, F. S., and Brown, T. A. (2009). Vegetation mediated the impacts of postglacial climate change on fire regimes in the south-central Brooks Range, Alaska. Ecol. Monogr. 79, 201–219. doi: 10.1890/07-2019.1
Higuera, P. E., Gavin, D. G., Bartlein, P. J., and Hallett, D. J. (2010). Peak detection in sediment-charcoal records: impacts of alternative data analysis methods on fire-history interpretations. Int. J. Wildl. Fire 19, 996–1014. doi: 10.1071/WF09134
Hooghiemstra, H. (1984). Vegetation and Climatic History of the High Plain of Bogotá: A Continuous Record of the Last 3.5 Million Years. Dissertationes Botanicae, Vol. 79. Amsterdam: J. Cramer Vaduz.
Hooghiemstra, H., and van der Hammen, T. (1993). Late quaternary vegetation history and paleoecology of Laguna Pedro Palo (subandean forest belt, Eastern Cordillera, Colombia). Rev. Palaeobot. Palynol. 77, 235–262. doi: 10.1016/0034-6667(93)90006-G
Horn, S. P., and Kappelle, M. (2009). “Fire in the Páramo ecosystems of Central and South America,” in Tropical Fire Ecology, 1st Edn, ed. M. A. Cochrane (Berlin: Springer Berlin Heidelberg), 505–539. doi: 10.1007/978-3-540-77381-8_18
IDEAM (2016). Suerficie de Bosque no Bosque año 2016. Available online at: http://www.ideam.gov.co/capas-geo (accessed June 6, 2019).
Juggins, S. (2007). C2 User Guide: Software for Ecological and Palaeoecological Data Analysis and Visualization. Newcastle upon Tyne: University of Newcastle, 1–73.
Keating, P. L. (2007). Fire ecology and conservation in the high tropical Andes: observations from northern Ecuador. J. Latin Am. Geogr. 6, 43–62. doi: 10.1353/lag.2007.0003
Kershaw, A. (1997). A modification of the Troels-Smith system of sediment description and portrayal. Quat. Australas. 15, 63–68.
Kuhry, P., Hooghiemstra, H., van Geel, B., and van der Hammen, T. (1993). The El Abra stadlal in the Eastern Cordillera of Colombia (South America). Quat. Sci. Rev. 12, 333–343. doi: 10.1016/0277-3791(93)90041-J
Langebaek, C. H. (2019). Los Muiscas: la Historia Milenaria de Un Pueblo Chibcha. Primera. Bogotá: Penguin Random House Grupo Editorial S.A.S.
Marchant, R., Behling, H., Berrio, J. C., Cleef, A., Duivenvoorden, J., Hooghiemstra, H., et al. (2001). Mid- to Late-Holocene pollen-based biome reconstructions for Colombia. Quat. Sci. Rev. 20, 1289–1308. doi: 10.1016/S0277-3791(00)00182-7
Markgraf, V. (1993). Younger Dryas in southernmost south America — An update. Quat. Sci. Rev. 12, 351–355. doi: 10.1016/0277-3791(93)90043-L
Martin, G. J. (2005). All Possible Worlds. The Age of Exploration, 4th Edn. London: Oxford University Press, 537.
Martin, L., Bertaux, J., Corrège, T., Ledru, M. P., Mourguiart, P., Sifeddine, A., et al. (1997). Astronomical forcing of contrasting rainfall changes in tropical South America between 12,400 and 8800 cal yr B.P. Quat. Res. 47, 117–122. doi: 10.1006/qres.1996.1866
Melles, M., Kulbe, T., Overduin, P. P., and Verkulich, S. (1994). “The expedition Bunger Oasis 1993/94 of the AWI Reearch Unit Potsdam,” in The Expeditions NORILSK/TAYMYR 1993 and Bunger Oasis 1993/94 of the AWI Reearch Unit Potsdam, ed. M. Melles (Bremerhaven: Alfred Wegener Institute for Polar and Marine Research), 29–80.
Mora, A., Parra, M., Strecker, M. R., Sobel, E. R., Hooghiemstra, H., Torres, V., et al. (2008). Climatic forcing of asymmetric orogenic evolution in the Eastern Cordillera of Colombia. Bull. Geol. Soc. Am. 120, 930–949. doi: 10.1130/B26186.1
Mottl, O., Flantua, S. G. A., Bhatta, K. P., Felde, V. A., Giesecke, T., Goring, S., et al. (2021). Global acceleration in rates of vegetation change over the past 18,000 years. Science 372, 860–864. doi: 10.1126/science.abg1685
Moy, C. M., Seltzer, G. O., Rodbell, D. T., and Anderson, D. M. (2002). Variability of El Niño/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature 420, 162–165. doi: 10.1038/nature01194
Mustaphi, C. J. C., and Pisaric, M. F. J. (2014). A classification for macroscopic charcoal morphologies found in Holocene lacustrine sediments. Prog. Phys. Geogr. 38, 734–754. doi: 10.1177/0309133314548886
Oksanen, A. J., Blanchet, F. G., Kindt, R., Legen-, P., Minchin, P. R., Hara, R. B. O., et al. (2018). Vegan: Community Ecology Package. Available online at: https://cran.r-project.org/package=vegan (accessed November 24, 2021).
Olivera-Moscol, M., Duivenvoorden, J. F., and Hooghiemstra, H. (2009). Pollen rain and pollen representation across a forest-páramo ecotone in northern Ecuador. Rev. Palaeobot. Palynol. 157, 285–300. doi: 10.1016/j.revpalbo.2009.05.008
Poveda, G., Waylen, P. R., and Pulwarty, R. S. (2006). Annual and inter-annual variability of the present climate in northern South America and southern Mesoamerica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 234, 3–27. doi: 10.1016/j.palaeo.2005.10.031
Prince, T. J., Pisaric, M. F. J., and Turner, K. W. (2018). Postglacial reconstruction of fire history using Sedimentary charcoal and pollen from a small lake in southwest Yukon Territory, Canada. Front. Ecol. Evol. 6:209. doi: 10.3389/fevo.2018.00209
R Core Team (2020). A Language and Environment for Statistical Computing. Vienna: R Statistical Foundation for Statistical Computing.
Ramsay, P. M. (2014). Giant rosette plant morphology as an indicator of recent fire history in Andean páramo grasslands. Ecol. Indic. 45, 37–44. doi: 10.1016/j.ecolind.2014.03.003
Rangel-Ch, J. O. (2015). La biodiversidad de Colombia: significado y distribución regional. Rev. Acad. Colomb. Cien. Exactas Fís. Nat. 39, 176. doi: 10.18257/raccefyn.136
Rangel-Ch, J. O., Lowy-C, P. D., and Aguilar-P M. (1997). “Distribucion de los tipos de Vegetación en las regiones,” in Colombia Diversidad Biotica II Tipos de vegetación en Colombia, 1st Edn. (Bogotá, DC: Instituto de Ciencias Naturales, Universidad Nacional de Colombia), 383–402.
Rangel-Ch, O. (2000). “La región paramuna y franja aledaña en Colombia,” in Colombia Diversidad Biótica III. La región de vida paramuna, ed. O. Rangel-Ch (Bogotá: Instituto de Ciencias Naturales – Instituto Alexander von Humboldt), 1–23.
Reimer, P. J., Austin, W. E., Bard, E., Bayliss, A., Blackwell, P. G., Bronk Ramsey, C., et al. (2020). The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0-55 cal kBP). Radiocarbon 62, 725–757. doi: 10.1017/RDC.2020.41
Rodionov, S. N. (2004). A sequential algorithm for testing climate regime shifts. Geophys. Res. Lett. 31, 2–5. doi: 10.1029/2004GL019448
Rodionov, S., and Overland, J. E. (2005). Application of a sequential regime shift detection method to the Bering Sea ecosystem. ICES J. Mar. Sci. 62, 328–332. doi: 10.1016/j.icesjms.2005.01.013
Rodríguez Eraso, N., Armenteras-Pascual, D., and Alumbreros, J. R. (2013). Land use and land cover change in the Colombian Andes: dynamics and future scenarios. J. Land Use Sci. 8, 154–174. doi: 10.1080/1747423X.2011.650228
Román-Cuesta, R. M., Carmona-Moreno, C., Lizcano, G., New, M., Silman, M., Knoke, T., et al. (2014). Synchronous fire activity in the tropical high Andes: an indication of regional climate forcing. Glob. Change Biol. 20, 1929–1942. doi: 10.1111/gcb.12538
Room, A. H., Franco-Gaviria, F., and Urrego, D. H. (2022). rshift: Paleoecology Functions for Regime Shift Analysis. R package version 2.1.1. Available online at: https://cran.r-project.org/web/packages/rshift/rshift.pdf (accessed January 31, 2022).
Rothhammer, F., and Dillehay, T. D. (2009). The late pleistocene colonization of South America: an interdisciplinary perspective. Ann. Hum. Genet. 73, 540–549. doi: 10.1111/j.1469-1809.2009.00537.x
Sarmiento, F. O. (2012). Contesting Paìramo: Critical Biogeography of the Northern Andean Highlands. Charlotte, NC: Kona Pub. and Media Group.
Saylor, J. E., Mora, A., Horton, B. K., and Nie, J. (2009). Controls on the isotopic composition of surface water and precipitation in the Northern Andes, Colombian Eastern Cordillera. Geochim. Cosmochim. Acta 73, 6999–7018. doi: 10.1016/j.gca.2009.08.030
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., and Walker, B. (2001). Catastrophic shifts in ecosystems. Nature 413, 591–596. doi: 10.1038/35098000
Sylvester, S. P., Heitkamp, F., Sylvester, M. D. P. V., Jungkunst, H. F., Sipman, H. J. M., Toivonen, J. M., et al. (2017). Relict high-Andean ecosystems challenge our concepts of naturalness and human impact. Sci. Rep. 7:3334. doi: 10.1038/s41598-017-03500-7
Taylor, Z., Horn, S., and Finkelstein, D. (2013). Maize pollen concentrations in Neotropical lake sediments as an indicator of the scale of prehistoric agriculture. Holocene 23, 78–84. doi: 10.1177/0959683612450201
Toth, L. T., Aronson, R. B., Vollmer, S. V., Hobbs, J. W., Urrego, D. H., Cheng, H., et al. (2012). ENSO drove 2500-year collapse of Eastern Pacific coral reefs. Science 336, 81–84. doi: 10.1126/science.1221168
Urrego, D. H., Bush, M. B., and Silman, M. R. (2010). A long history of cloud and forest migration from Lake Consuelo, Peru. Quat. Res. 73, 364–373. doi: 10.1016/j.yqres.2009.10.005
Urrego, D. H., Bush, M. B., Silman, M. R., Correa-metrio, A., Ledru, M., Mayle, F. E., et al. (2009). “Millennial-scale ecological changes in tropical South America since the last glacial maximum,” in Past Climate Variability in South America and Surrounding Regions, eds F. Vimeux, F. Sylvestre, and M. Khodri (Dordrecht: Springer), 283–300. doi: 10.1007/978-90-481-2672-9_12
Urrego, D. H., Hooghiemstra, H., Rama-Corredor, O., Martrat, B., Grimalt, J. O., Thompson, L., et al. (2016). Millennial-scale vegetation changes in the tropical Andes using ecological grouping and ordination methods. Clim. Past 12, 697–711. doi: 10.5194/cp-12-697-2016
Valencia, B. G., Urrego, D. H., Silman, M. R., and Bush, M. B. (2010). From ice age to modern: a record of landscape change in an Andean cloud forest. J. Biogeogr. 37, 1637–1647. doi: 10.1111/j.1365-2699.2010.02318.x
van Boxel, J. H., González-Carranza, Z., Hooghiemstra, H., Bierkens, M., and Vélez, M. I. (2014). Reconstructing past precipitation from lake levels and inverse modelling for Andean Lake La Cocha. J. Paleolimnol. 51, 63–77. doi: 10.1007/s10933-013-9755-1
van der Hammen, T., and Hooghiemstra, H. (1995). The El Abra stadial, a younger dryas equivalent in Colombia. Quat. Sci. Rev. 14, 841–851. doi: 10.1016/0277-3791(95)00066-6
van der Hammen, T., Correal, G., and van Klinken, G. J. (1990). Isotopos estables y dieta del hombre prehistórico en la Sabana de Bogotá (un estudio inicial). Bol. Arqueol. 5, 1–10.
van’t Veer, R., Islebe, G. A., and Hooghiemstra, H. (2000). Climatic change during the Younger Dryas chron in northern South America: a test of the evidence. Quat. Sci. Rev. 19, 1821–1835. doi: 10.1016/S0277-3791(00)00093-7
Velásquez, C. (1999). Atlas Palinologico de la Flora Vascular Paramuna de Colombia: Angiospermae. Medelliìn: Universidad Nacional de Colombia, Sede Melelliìn, Facultad de Ciencias, 1–2.
Vélez, M. I, MacKenzie, K., Boom, A., Bremond, L., Gonzalez, N., Carr, A. S., et al. (2020). Lacustrine responses to middle and late Holocene anthropogenic activities in the northern tropical Andes. J. Paleolimnol. 65, 123–136. doi: 10.1007/s10933-020-00152-y
Vélez, M. I., Hooghiemstra, H., Metcalfe, S., Wille, M., and Berrío, J. C. (2006). Late Glacial and Holocene environmental and climatic changes from a limnological transect through Colombia, northern South America. Palaeogeogr. Palaeoclimatol. Palaeoecol. 234, 81–96. doi: 10.1016/j.palaeo.2005.10.020
Weng, C., Bush, M. B., and Chepstow-Lusty, A. J. (2004). Holocene changes of Andean alder (Alnus acuminata) in highland Ecuador and Peru. J. Quat. Sci. 19, 685–691. doi: 10.1002/jqs.882
White, S. (2013). Grass páramo as hunter-gatherer landscape. Holocene 23, 898–915. doi: 10.1177/0959683612471987
Whitlock, C., and Larsen, C. (2001). “Charcoal as a fire proxy,” in Tracking Environmental Change Using Lake Sediments. Terrestrial, Algal, and Siliceous Indicators, Vol. 3, eds J. P. Smol, H. J. B. Birks, and W. M. Last (Dordrecht: Kluwer Academic Publishers), 75–97. doi: 10.1007/0-306-47668-1_5
Williams, J. W., and Jackson, S. T. (2007). Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 5:475–482. doi: 10.1890/070037
Keywords: Colombian Andes, Holocene, paleoecology, ecological novelty, fire, charcoal, pollen
Citation: Espinoza IG, Franco-Gaviria F, Castañeda I, Robinson C, Room A, Berrío JC, Armenteras D and Urrego DH (2022) Holocene Fires and Ecological Novelty in the High Colombian Cordillera Oriental. Front. Ecol. Evol. 10:895152. doi: 10.3389/fevo.2022.895152
Received: 13 March 2022; Accepted: 02 May 2022;
Published: 09 June 2022.
Edited by:
Angelica Feurdean, Goethe University Frankfurt, GermanyReviewed by:
Encarni Montoya, University of Liverpool, United KingdomMark Bush, Florida Institute of Technology, United States
Copyright © 2022 Espinoza, Franco-Gaviria, Castañeda, Robinson, Room, Berrío, Armenteras and Urrego. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ismael G. Espinoza, igarciae@unal.edu.co; Dunia H. Urrego, D.Urrego@exeter.ac.uk