 Jacob E. Lucero
Jacob E. Lucero Akasha M. Faist
Akasha M. Faist Christopher J. Lortie2
Christopher J. Lortie2 Ragan M. Callaway
Ragan M. Callaway
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 09 May 2022
Sec. Population, Community, and Ecosystem Dynamics
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.886690
This article is part of the Research TopicBiological Invaders: Always the Bad Guys?View all 6 articles
Positive interactions can drive the assembly of desert plant communities, but we know little about the species-specificity of positive associations between native shrubs and invasive annual species along aridity gradients. These measures are essential for explaining, predicting, and managing community-level responses to plant invasions and environmental change. Here, we measured the intensity of spatial associations among native shrubs and the annual plant community—including multiple invasive species and their native neighbors—along an aridity gradient across the Mojave and San Joaquin Deserts, United States. Along the gradient, we sampled the abundance and species richness of invasive and native annual species using 180 pairs of shrub and open microsites. Across the gradient, the invasive annuals Bromus madritensis ssp. rubens (B. rubens), B. tectorum, B. diandrus, Hordeum murinum, and Brassica tournefortii were consistently more abundant under shrubs than away from shrubs, suggesting positive effects of shrubs on these species. In contrast, abundance of the invasive annual Schismus spp. was greater away from shrubs than under shrubs, suggesting negative effects of shrubs on this species. Similarly, native annual abundance (pooled) and native species richness were greater away from shrubs than under shrubs. Shrub-annual associations were not influenced by shrub size or aridity. Interestingly, we found correlative evidence that B. rubens reduced native abundance (pooled), native species richness, and exotic abundance (pooled) under, but not away from shrubs. We conclude that native shrubs have considerable potential to directly (by increasing invader abundance) and indirectly (by increasing negative impacts of invaders on neighbors) facilitate plant invasions along broad environmental gradients, but these effects may depend more upon invader identity than environmental severity.
Positive interactions among species (facilitation) enhance biodiversity and ecosystem functioning (Wright et al., 2017, 2021) while buffering communities against environmental change (Bulleri et al., 2018). Facilitation occurs when a foundation species alters the biotic or abiotic environment in a way that enhances the performance, survival, or fitness of beneficiary species (Bronstein, 2009; mechanisms reviewed by Filazzola and Lortie, 2014). Importantly, positive interactions can be species-specific because some foundation species are better facilitators than others, and not all beneficiary species are facilitated equally (Callaway, 1998). The intensity of facilitation also depends upon environmental severity. The stress-gradient hypothesis (SGH) predicts that the frequency (Bertness and Callaway, 1994) and intensity (le Roux and McGeoch, 2010) of positive interactions increase as environmental stress increases. Hundreds of studies have examined the SGH, and meta-analyses have found broad support for its basic predictions (e.g., Lortie and Callaway, 2006; He et al., 2013; Romero et al., 2015; Adams et al., 2022).
Positive interactions can also influence the trajectory of biological invasions by exotic plant species (Cavieres, 2021). Exotic species commonly facilitate each other (Braga et al., 2018), but native species also facilitate plant invasions (Northfield et al., 2018). Native facilitators can promote invasions directly by increasing the local abundance, performance, and fitness of invasive species; or indirectly by increasing the negative impacts of invasive species on neighbors. For instance, Lucero et al. (2021) showed that the invasive annual Bromus tectorum achieved much greater abundance, biomass, and fitness near native shrubs than away from shrubs; and Lortie et al. (2021) showed that invasive annual species had stronger competitive effects on native neighbors when they were facilitated than when they were not. Environmental context can mediate the strength of positive interactions experienced by invasive species (Cavieres et al., 2008; Saccone et al., 2010; Llambi et al., 2018; Filazzola et al., 2020), but we still know very little about the species-specificity of facilitated invasions.
Facilitation by desert shrubs is a useful framework for addressing these issues. Arid and semi-arid ecosystems cover over 40% of the earth’s land surface, comprise 44% of the world’s arable land, support more than 50% of the world’s livestock, and contribute substantially to global biodiversity (Millennium Ecosystem Assessment, 2005; Davies et al., 2021). However, increasing aridity associated with anthropogenic climate change and accelerating biological invasions by exotic plant species are degrading the biodiversity and functioning of desert ecosystems at an alarming rate (DiTomaso et al., 2010; Huang et al., 2016; Phillips et al., 2022). Shrubs can facilitate desert plant communities by creating favorable microsites underneath their canopies that ameliorate environmental extremes (Callaway, 2007; Michalet and Pugnaire, 2016). For instance, shade from shrub canopies can buffer understory plants against solar radiation and heat (Valiente-Benuet and Ezcurra, 1991), and soil nutrients (Schlesinger et al., 1996) and water (Segoli et al., 2012) are often more concentrated under shrub canopies than away from shrubs. Many exotic invaders capitalize on positive interactions in deserts (e.g., Butterfield et al., 2010; Badano et al., 2016; López et al., 2016; Filazzola et al., 2020; Slate et al., 2020), and sometimes to a greater extent than native competitors (Lucero et al., 2019). The SGH predicts that shrub-mediated interactions should be most positive in the most arid environments, but there remains considerable opportunity to test this relationship with multiple invasive species simultaneously (but see Holzapfel and Mahall, 1999; Brooks, 2003; Filazzola et al., 2020). Understanding of the species-specificity of facilitated invasions along aridity gradients—including indirect effects on native competitors—is necessary for explaining, predicting, and ultimately managing the ecological consequences of concomitant exotic plant invasion and environmental change in desert systems.
The objective of this study was to evaluate the potential for native shrubs to directly (by increasing the local abundance of invasive species) and indirectly (by increasing the negative impacts of invasive species on neighbors) facilitate annual plant invasions along an aridity gradient across the Mojave and San Joaquin Deserts, United States. We tested two interrelated hypotheses. First, we tested the hypothesis that the intensity of shrub-invasive annual associations depends upon invasive species identity and relative aridity. Specifically, we predicted that: (i) the intensity of shrub-invasive annual associations varies by species; and (ii) the intensity of shrub-invasive annual associations becomes more positive as relative aridity increases, as predicted by the SGH. Second, we hypothesized that association with native shrubs alters the outcome of biotic interactions (inferred via spatial relationships) between invasive annuals and herbaceous neighbors, including native and exotic species. Specifically, we predicted that abundance relationships between invasive annuals and herbaceous neighbors are more negative (i.e., more competitive) near shrubs than away from shrubs (after Lortie et al., 2021).
We surveyed annual plant communities at peak flowering in April 2020 at nine sites that spanned an aridity gradient (see Supplementary Table 1 for site names, locations, and aridity values) across the Mojave (n = 6) and San Joaquin (n = 3) Deserts, United States. Sites in the Mojave Desert were located near Cal-Nev-Ari, NV; the Heart of the Mojave ORV Trail, CA; Mesquite, NV; Mojave, CA; Las Vegas, NV; and Yucca Grove, CA. Sites in the San Joaquin Desert were located near Carrizo Plain National Monument, CA; Cuyama, CA; and Panoche Hills, CA. We selected sites in climax native shrub communities that had not recently experienced a major disturbance (e.g., fire). One site (Carrizo) was grazed by cattle during the study year, but all others were not grazed. For each site, we calculated the de Martonne aridity index (AdM) (de Martonne, 1920) during the study year as follows:
where P was the total precipitation (mm) from May 1, 2019, to April 30, 2020, and T was the mean daily temperature (°C) during the same interval. Thus, low AdM values indicated high aridity. We also calculated a 30-year AdM value for each site by averaging yearly AdM values from 1991 to 2020. Importantly, our statistical analyses used the AdM of the study year (2019–2020), not the 30-year average, because current climatic trends are more relevant to the performance of annual plant species than long-term trends. We chose this aridity index because weather stations nearly always report its components, which allows for direct calculations of AdM at spatiotemporal scales relevant to biotic interactions in local communities.
Sites in the Mojave Desert were dominated by the native shrub Larrea tridentata, and sites in the San Joaquin Desert were dominated by the native shrub Ephedra californica. Other shrub species were present at relatively low densities, but no one shrub species was present at all sites. We focused on the potential for native shrubs to act as facilitators because they are the dominant physiognomic class across our study area and are common targets for land management actions (Eldridge et al., 2011).
We identified exotic annuals to species at each site. Bromus madritensis ssp. rubens (B. rubens hereafter), Schismus spp. (S. arabicus and S. barbatus are difficult to reliably differentiate in the field and were thus combined as a taxa), and Erodium cicutarium were present at all sites. Bromus tectorum was present at Cuyama, Mesquite, Mojave, Las Vegas, and Yucca Grove. Bromus diandrus was present at Cuyama and Panoche Hills. Hordeum murinum was present at Carrizo and Cuyama. Brassica tournefortii was present at Cal-Nev-Ari and Mesquite. Each of these exotic species (1) was translocated at some point from a distant native range (Eurasia/Northern Africa) to North America, (2) has established self-sustaining populations in North America, (3) is expanding its range in North America, and (4) can negatively impact biodiversity and ecosystem functioning in North American plant communities. Thus, each of these species meets the criteria of Blackburn et al. (2011) for being “invasive,” and each is considered invasive by the California Invasive Plant Council (CAL-IPC, 2022).
During the study year, annual precipitation, mean annual temperature, and AdM at the study sites ranged from 137 to 287 mm, 16.32–21.00°C, and 4.41–10.89 (AdM is unitless), respectively. Thirty-year AdM values ranged from 3.44 to 24.04, a much broader range than the study year. For six of our nine sites, the AdM for the study year fell within the 95% CI of the 30-year AdM, but Carrizo, Heart of the Mojave, and Panoche Hills were more arid than usual (Supplementary Table 1).
We sampled the annual plant community using a paired shrub-open microsite contrast with a 0.5 × 0.5 m quadrat (Pescador et al., 2014). Shrub microsites were defined as the area immediately beneath the canopy of a focal shrub, and open microsites were defined as interstitial spaces at least 1 m from any shrub canopy but <5 m from the focal shrub. We did not sample areas more than 5 m away from shrubs. For shrub microsites, sampling quadrats were placed midway between the shrub center and dripline. A total of 180 pairs of shrub and open microsites were sampled (n = 20 shrub-open pairs at each study site), and for each shrub-open pair, we noted the species, radius (r; m), and height (h; m) of the shrub. We used the formula for an oblate spheroid:
to estimate the volume (m3) of each shrub. We used this formula to estimate shrub size because it is more nuanced than shrub height or width alone (Lortie et al., 2018; Lucero et al., 2021). Shrub-open pairs for sampling were chosen randomly by facing a random compass bearing and then blindly tossing a tennis ball backward in the opposite direction. The shrub-open pair nearest to the ball was sampled. In the Mojave, focal shrubs were L. tridentata (n = 74), Ambrosia dumosa (n = 24), Coleogyne ramossissima (n = 12), and Lycium andersonii (n = 10). In the San Joaquin, E. californica was the focal shrub for all shrub-open pairs (n = 60).
In sampling quadrats, we recorded the abundance (no. plants rooted inside the quadrat) of each exotic annual species, exotic annual species combined, native annual species combined, and the total annual plant community (all annual species combined); as well as the richness of native annual species, exotic annual species, and the total annual plant community (all annual species combined). Relationships among these particular measures are used to describe the invasiveness and impacts of exotic species in non-native communities (Pearson et al., 2016). We did not encounter species that reproduced asexually or vegetative mats suggestive of extensive tillering.
Relative interaction indices (RIIs; Armas et al., 2004) were used as a simple effect size measure to estimate the intensity of shrub-annual associations. We calculated RII as follows:
where M_s was a vegetation measure (e.g., B. rubens abundance, native species richness, etc.) in a shrub microsite and M_o was the same measure in the paired open microsite. RII values range from −1 to +1. Negative RII values indicate negative (competitive) shrub-annual associations, positive values indicate positive (facilitative) associations, and a value of 0 indicates no (neutral) association.
We used t-tests and linear mixed-effects models to characterize relationships among shrubs and annual plant species. To evaluate the direction and intensity of shrub-annual associations at each site and across all sites, we performed one-sample t-tests with RII as the response variable. T-tests across all sites used RIIs averaged at the site level. We evaluated the influence of aridity and shrub size on the intensity of shrub-annual associations (i.e., RIIs) using linear mixed-effects models with RII as the response variable; AdM, shrub volume, and their interaction as covariates; and shrub species nested within study site as a random effect. We inferred biotic interactions among neighboring annuals in shrub and open microsites using linear mixed-effects models with annual plant abundance or species richness (log-transformed) as the response variable; exotic annual abundance (species-specific or combined) as a covariate (log-transformed); and shrub species nested within study site as a random effect. For these analyses, negative line slopes (P < 0.05) suggest negative interactions among neighbors, positive line slopes (P < 0.05) suggest positive interactions, and line slopes with P > 0.05 suggest neutral interactions. We performed species-specific analyses only for exotic annual taxa present at more than two study sites.
All analyses were performed in R, version 4.0.2 (R Development Core Team, 2020). We did t-tests using the t-test function (R Development Core Team, 2020). Linear mixed-effects models used the lmer function of the “lmerTest” package (Kuznetsova et al., 2017). R2-values were extracted using the r2 function (R Development Core Team, 2020).
Invasive annuals formed species-specific associations with native shrubs (Figure 1 and Supplementary Table 2). Across sites, mean RIIs for B. rubens, B. tectorum, B. diandrus, H. murinum, and B. tournefortii ranged from 0.829 (0.043 SE; P < 0.001) to 1.000, indicating that each of these invasive species formed remarkably intense positive associations with native shrubs across our study area. In contrast, mean RII across sites for invasive annual grass Schismus spp. was −0.435 (0.195 SE; P = 0.032), indicating a negative association with shrubs across our study area. Mean RII across sites for E. cicutarium was −0.374 (0.147 SE; P = 0.061), indicating no strong associations with shrubs. Interestingly, at the site level, RIIs for B. rubens, B. tectorum, B. diandrus, H. murinum, and B. tournefortii were always positive and never fell below 0.545 (0.059 SE; P < 0.001), suggesting consistent positive associations with native shrubs at each site. At the site level, RIIs for Schismus spp. and E. cicutarium varied from −1.000 to 0.645 (1.06 SE), suggesting that their associations with shrubs were not only negative net neutral or, but also highly variable. Across sites, mean RIIs for native abundance and native species richness (all native species combined) were −0.358 (0.153 SE; P = 0.024) and −0.321 (0.132 SE; P = 0.020), respectively, suggesting that the native annual community was negatively associated with shrubs across our study area. Across sites, mean RIIs for exotic species richness (all exotic species combined) and total species richness (all species combined) were 0.014 (0.046 SE; P = 0.740) and −0.121 (0.074 SE; P = 0.101), respectively, suggesting no meaningful effect of shrubs on the species richness of the exotic or total annual community across our study area.
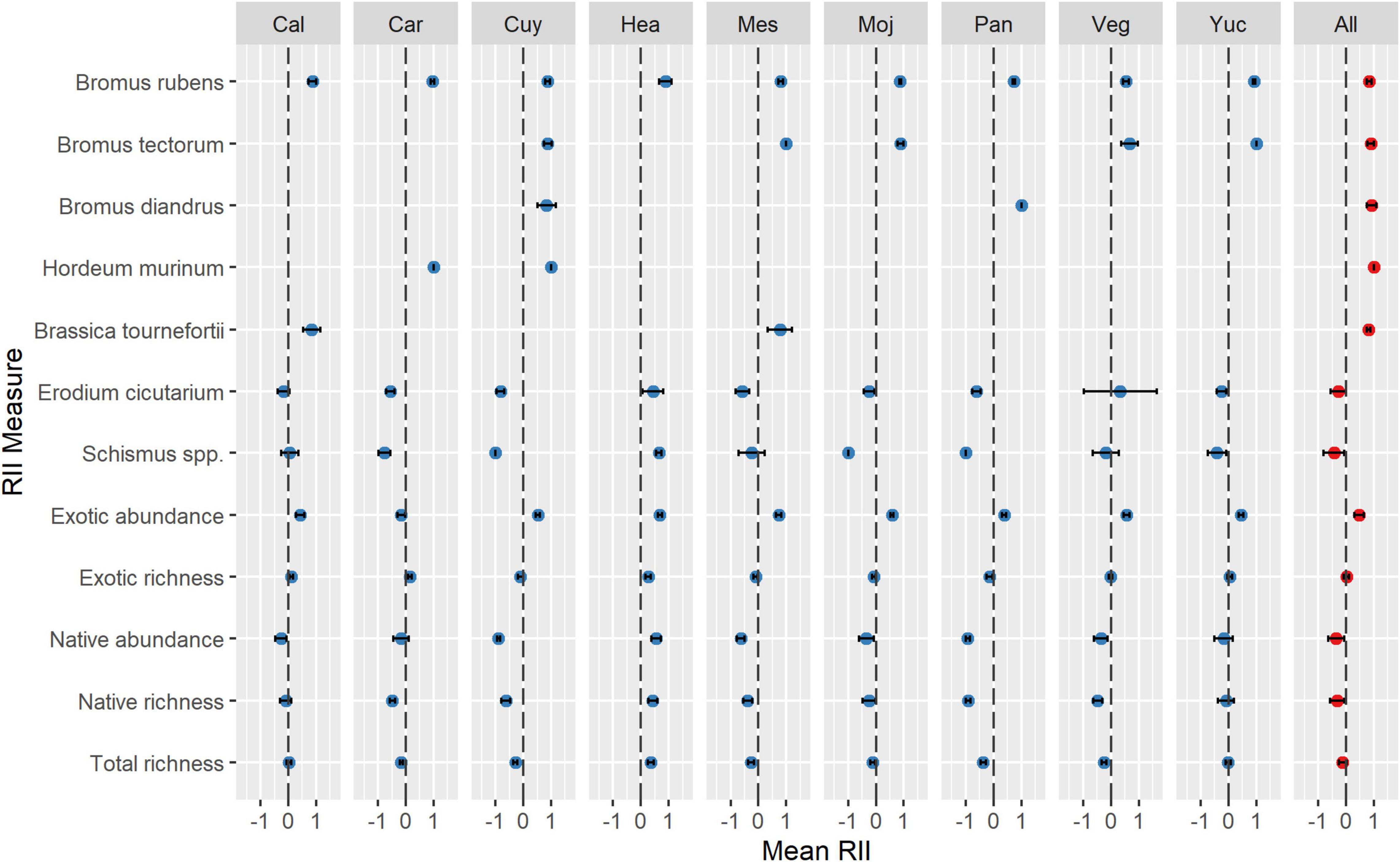
Figure 1. Mean intensity (RII ± 95% CI) of shrub-herbaceous plant associations at each study site and averaged across all sites (“All”; shown in red). RIIs > 0 suggests positive (i.e., facilitative) shrub-annual associations; RIIs < 0 suggests negative (i.e., antagonistic) associations. Site names are abbreviated by their first three letters (see Supplementary Table 1 for full names).
The intensity of shrub-annual associations was not influenced by aridity (i.e., AdM) or shrub size (Table 1). As aridity and shrub size increased, RIIs for all vegetation measures remained constant. Thus, we found no support for the SGH. In addition, we found no evidence for aridity × shrub size interactions (Table 1).
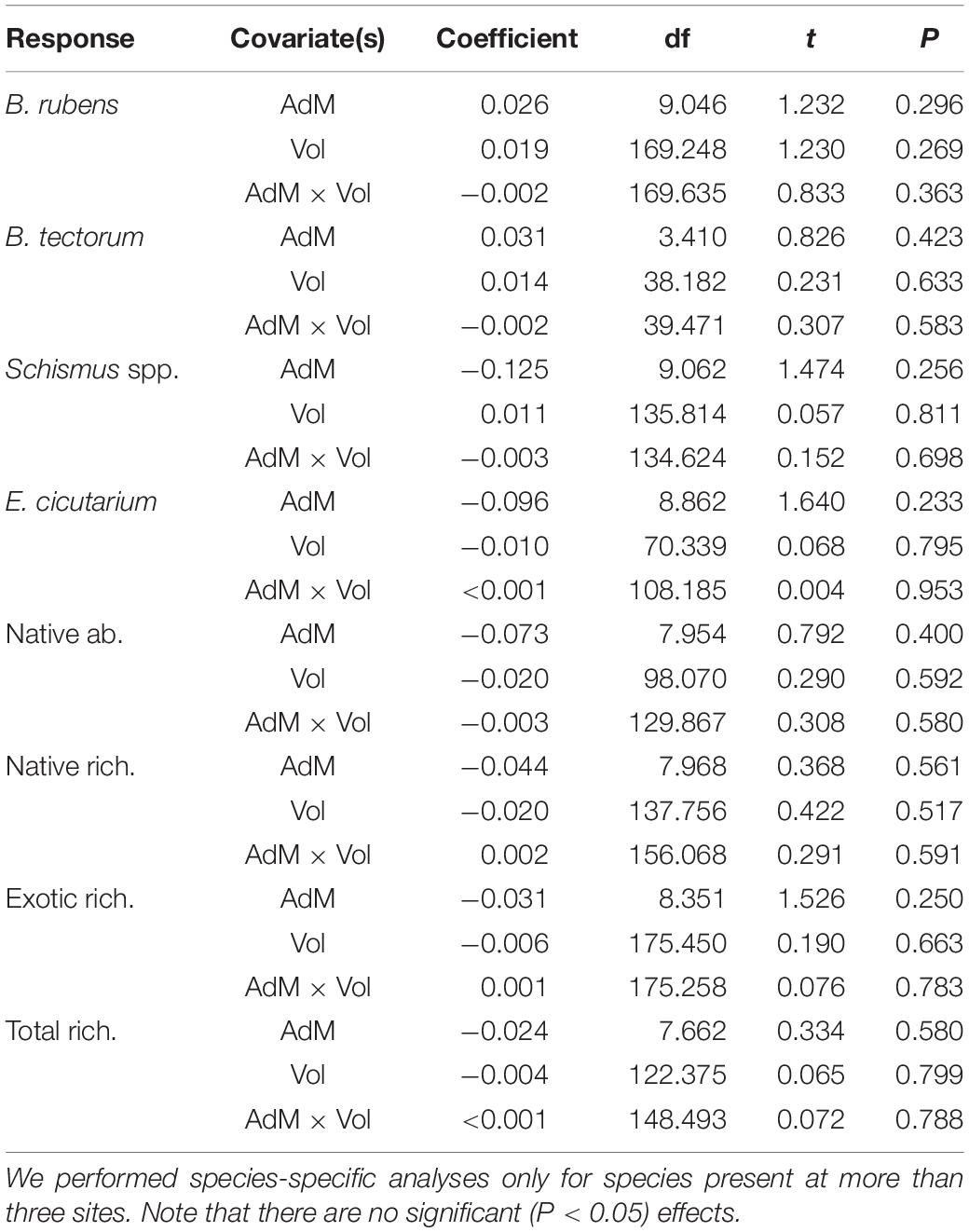
Table 1. Results of linear mixed-effects models with RII as the response variable; de Martonne aridity score (Adm), shrub volume (Vol), and their interaction (AdM × Vol) as covariates; and shrub species nested within site as a random effect (not shown).
Native shrubs altered abundance relationships between the invasive annual B. rubens and the native annual community (Figure 2 and Table 2). In shrub microsites, increasing B. rubens abundance corresponded to decreasing native abundance and native species richness, suggesting competitive interactions between B. rubens and native annuals under shrubs. In contrast, in open microsites, we found no significant relationships between B. rubens abundance and native abundance or native species richness, suggesting relatively weak interactions in the open. Increasing exotic abundance (all exotic species combined) corresponded to decreasing native abundance and native species richness in shrub microsites, but again, not in open microsites (Figure 3). These negative relationships under shrubs were apparently driven primarily by B. rubens, because removing B. rubens from the exotic abundance data eliminated all significant exotic-native relationships (Figure 4).
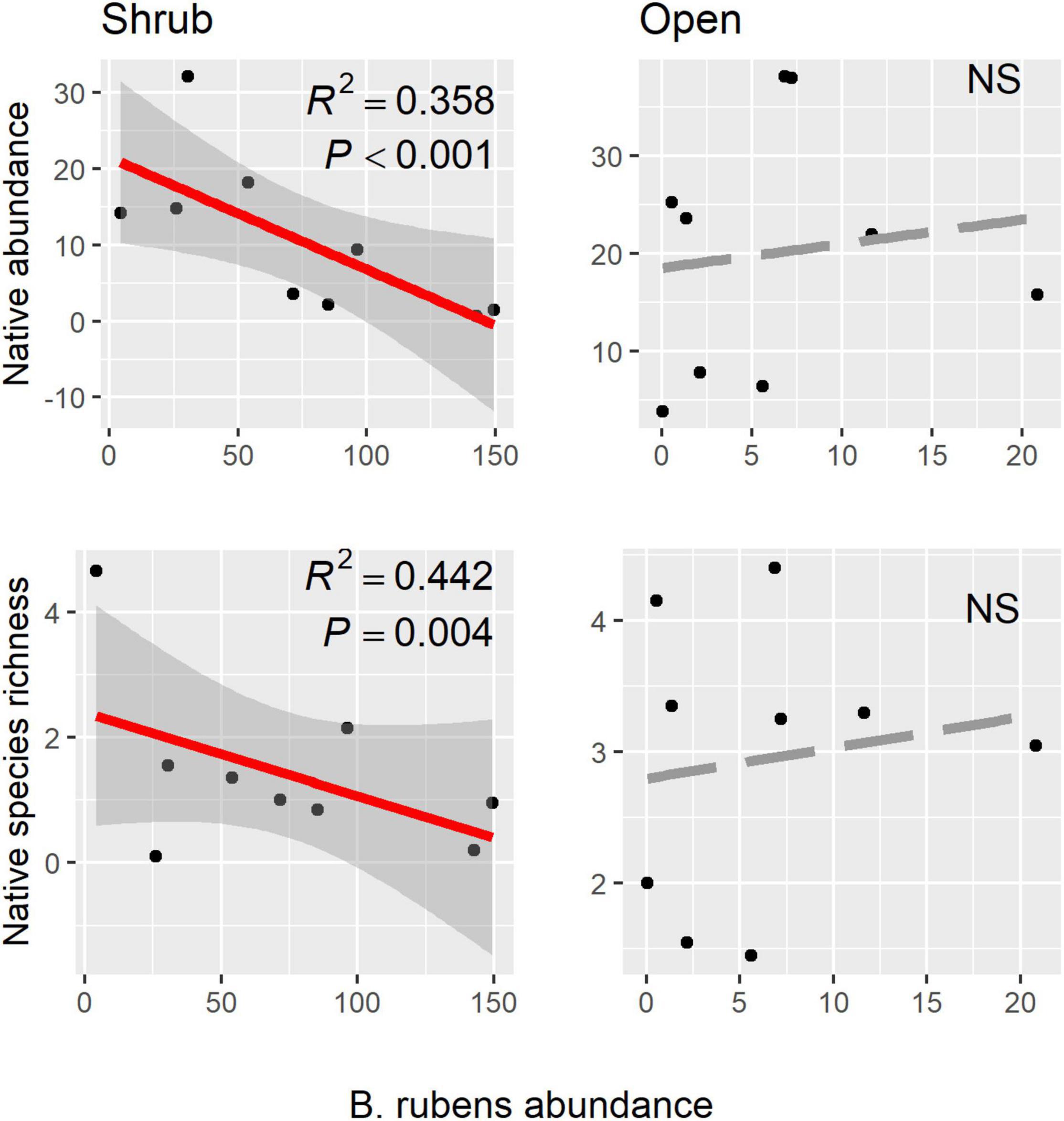
Figure 2. Native abundance and native species richness (all native species combined) plotted against B. rubens abundance in shrub and open microsites. Red regression lines (± 95% CI) indicate significant native-B. rubens relationships.
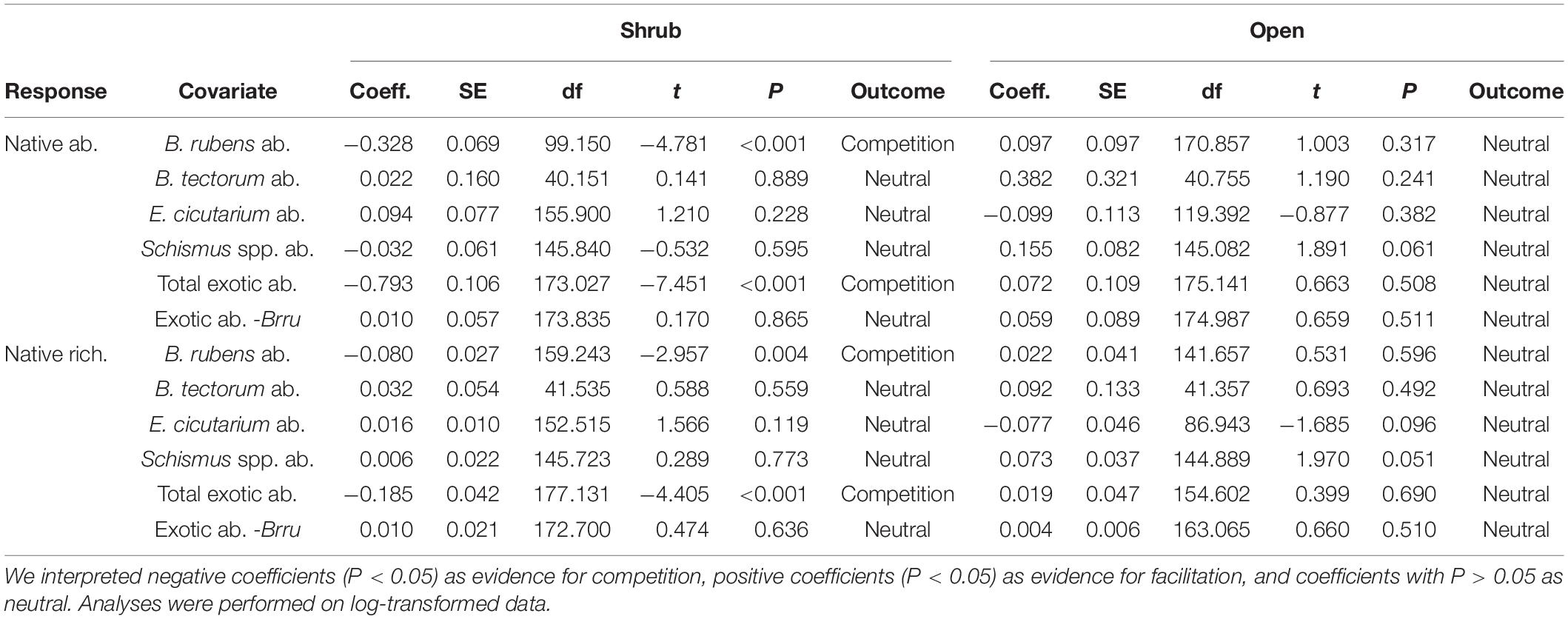
Table 2. Results of linear mixed-effects models replicated for shrub and open microsites with native abundance or native species richness (all native species combined) as the response variable; abundance of B. rubens, B. tectorum, E. cicutarium, Schismus spp., all exotic species combined (Total exotic ab.), or exotic species except for B. rubens (Exotic ab. -Brru) as the covariate; and shrub species nested within study site as a random effect (not shown).
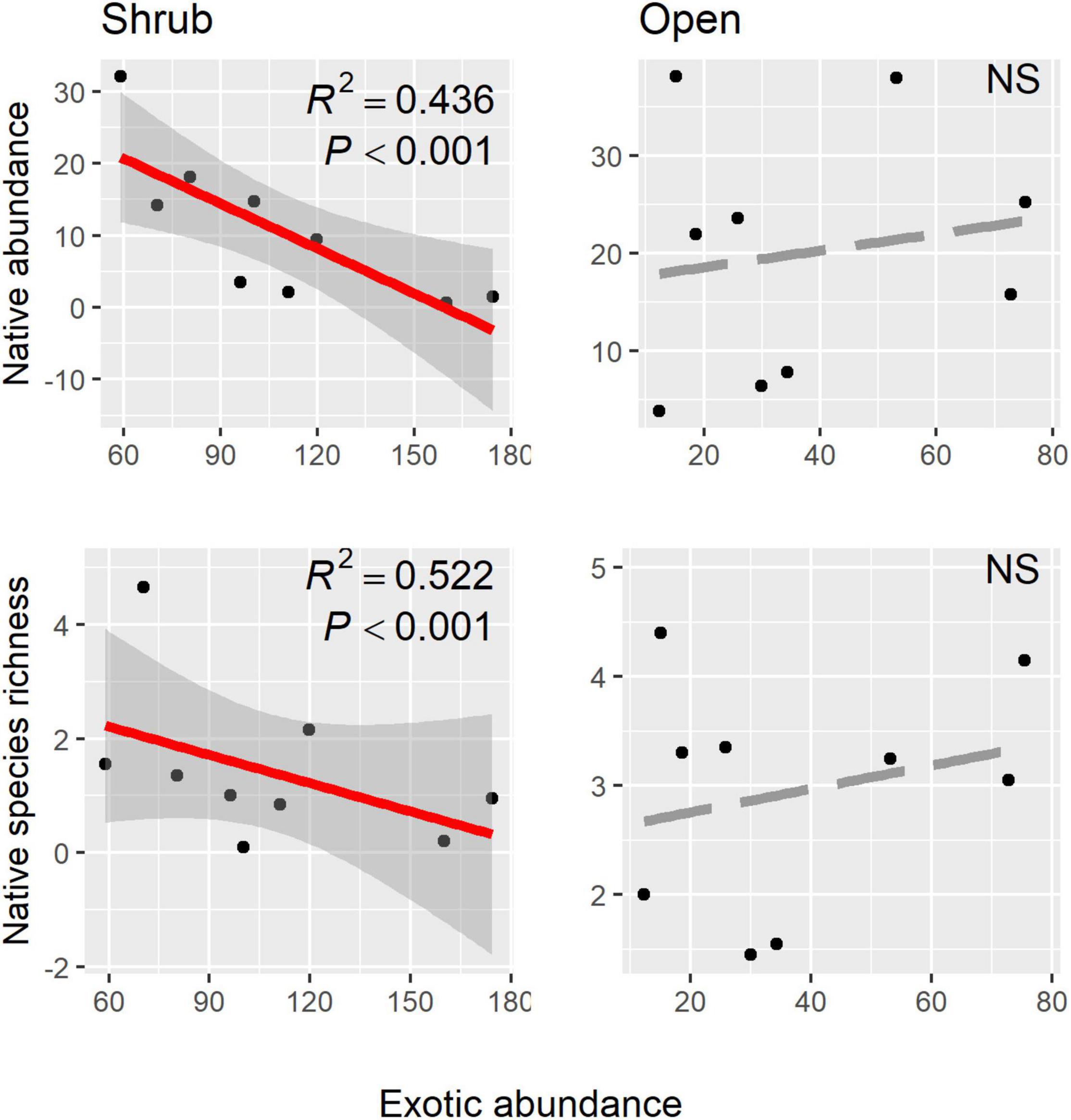
Figure 3. Native abundance and native species richness (all native species combined) plotted against exotic abundance (all exotic species combined) in shrub and open microsites. Red regression lines (± 95% CI) indicate significant native-exotic relationships.
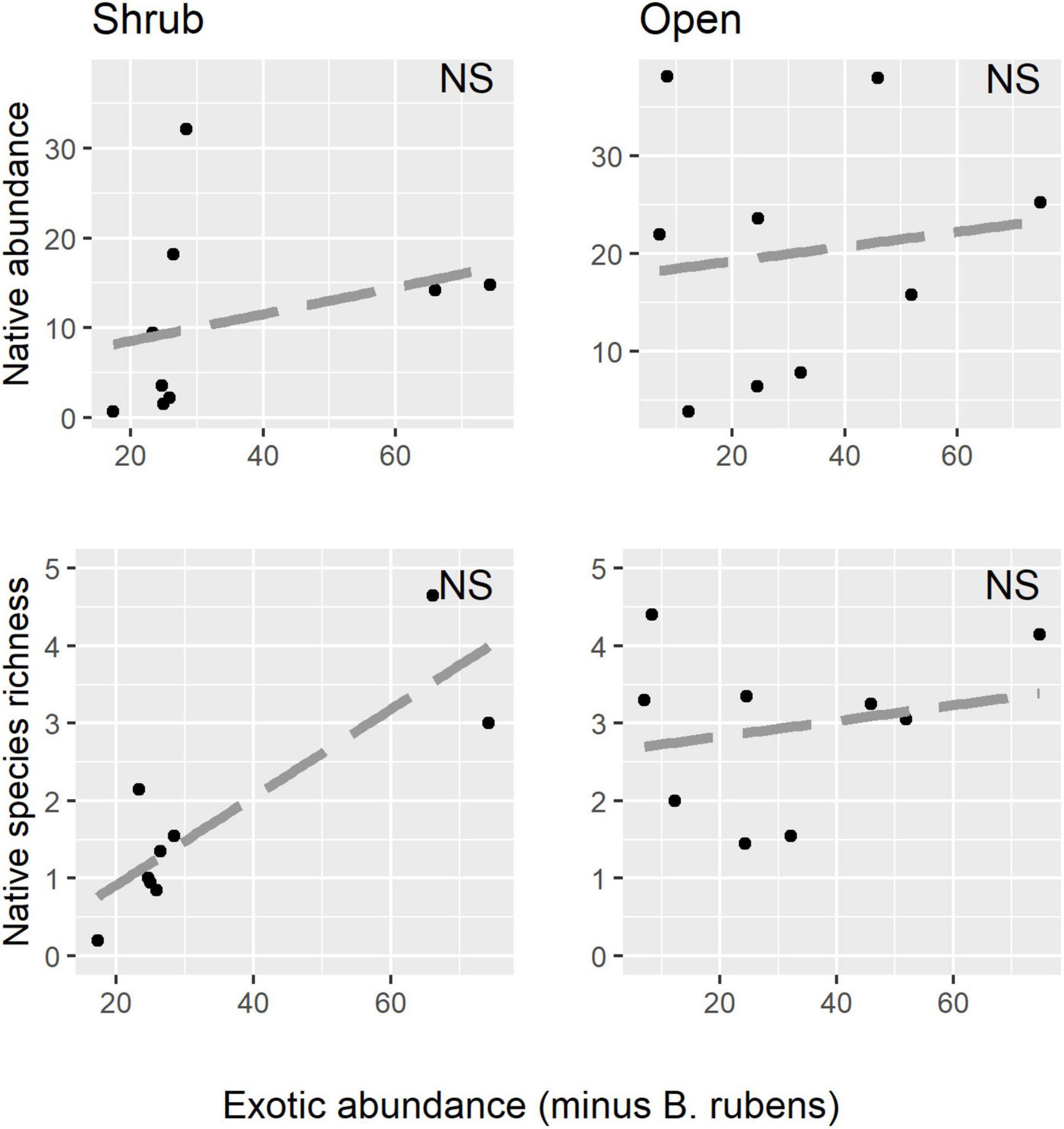
Figure 4. Native abundance and native species richness (all native species combined) plotted against exotic abundance with B. rubens omitted from the data in shrub and open microsites. Note that there are no significant native-exotic relationships when B. rubens is omitted from the data.
Shrubs also altered abundance relationships between B. rubens and the broader exotic annual community (all exotic species combined except for B. rubens) (Supplementary Figure 2 and Supplementary Table 3). In shrub microsites, increasing B. rubens abundance was negatively related to exotic abundance but had no relationship to exotic species richness. In open microsites, increasing B. rubens abundance had no relationship to exotic abundance but was positively related to exotic species richness.
Our study enriches our understanding of the species-specificity of facilitated invasions. We have long known that not all species are facilitated equally (Callaway, 1998). However, few studies have evaluated the potential for positive interactions to affect multiple invasive species simultaneously along aridity gradients. We found that spatial association with native shrubs consistently increased the abundance of some invasive annual species (B. rubens, B. tectorum, B. diandrus, H. murinum, and B. tournefortii), decreased the abundance of other invasive annuals (Schismus spp.), and had no consistent effects on another invasive annual (E. cicutarium). These findings support our hypothesis that the intensity of shrub-invasive annual associations depends strongly upon invasive species identity. Interestingly, the direction and intensity of these associations were unaffected by shrub size or relative aridity across our observed gradient. Thus, we found no support for the hypothesis that the intensity of shrub-invasive annual associations depends upon relative aridity. Additionally, we found correlative evidence (via abundance relationships) that native shrubs exacerbated the negative impacts of B. rubens on neighboring annuals. This supports the hypothesis that spatial association with native shrubs can alter the outcome of biotic interactions between invasive annuals and herbaceous neighbors. Taken together, our main findings suggest that native shrubs have considerable potential to facilitate exotic plant invasions across a broad spatial scale, but this potential may depend more upon invader identity than environmental severity. This accords with studies showing that species-specific factors can be more important than environmental conditions as drivers of nurse plant effects in drylands (Soliveres et al., 2012; Filazzola et al., 2020).
Our main findings corroborate that desert shrubs can facilitate heterospecific neighbors, including invasive annuals. Positive effects of desert shrubs on annual species include increased abundance, performance, and population growth (Callaway, 2007; Griffith, 2010). Desert shrubs and their interactions can bolster the biodiversity (Bashirzadeh et al., 2022) and functioning (Khosravi Mashizi and Sharafatmandrad, 2019) of desert ecosystems (but see Báez and Collins, 2008), but desert shrubs do not facilitate all taxa equally (Filazzola et al., 2020). In this context, our findings coincide nicely with the literature review of Abella and Smith (2013), who found that across the Mojave Desert, exotic annuals were generally more abundant near native perennials than in interspaces, suggesting overall facilitation of exotic annuals by native perennials. In particular, they found that B. rubens formed stronger positive associations with native perennials than Schismus spp. or E. cicutarium. Our findings also coincide with reports that exotic annuals respond more strongly to positive interactions than native annuals at the provenance level. In a southern California rangeland, Lucero et al. (2019) found that exotic annuals as a group were facilitated over 2.75 times stronger in abundance than native annuals, and across the Mojave and San Joaquin Deserts, Lucero et al. (2020) found that positive associations were intense and important for invasive B. rubens but not the native annual community. By promoting plant invasions (Lortie et al., 2021), intense facilitation of invasive species can indirectly erode biodiversity and ecosystem functioning (Northfield et al., 2018). Thus, any community-level benefits of shrub facilitation may backfire when invasive species are strongly facilitated. Our work extends this literature to multiple invasive taxa interacting with native shrubs across a broad geographical scale that spans substantial environmental variability.
Our study highlights the precocious ability of invasive Bromus species to capitalize on native facilitators. Native shrubs, trees, and grasses across the Great Basin, Mojave, San Joaquin, and Colorado Plateau Deserts can increase the abundance, biomass, fitness, and population growth of B. tectorum (Belnap and Sherrod, 2008; Griffith, 2010; Kane et al., 2011; Lucero et al., 2021). Across the Mojave and San Joaquin Deserts, native perennials often strongly facilitate B. rubens (reviewed by Abella and Smith, 2013). In central California, invasive B. diandrus and B. mollis can be facilitated by native Quercus douglasii (Callaway et al., 1991). In this study, each invasive Bromus species we encountered (B. rubens, B. tectorum, B. diandrus) formed intense positive associations with native shrubs, regardless of environmental context. Taken together, this literature suggests that invasive Bromus species are broadly facilitated by native plants from a variety of functional groups and environments. We suggest that disrupting positive interactions from native facilitators may be a useful strategy for controlling Bromus invasions across western North America. For instance, herbicide applications to reduce invasive Bromus densities could be most cost-effective when focused under shrub canopies. Importantly, we acknowledge that invasive Bromus species do not necessarily need native facilitators to establish (Holzapfel and Mahall, 1999; Brooks and Berry, 2006; Lucero et al., 2021) or spread (Reid et al., 2006). Rather, our findings suggest that native shrubs can aggravate Bromus invasions by increasing their local densities and impacts.
Biotic interactions do not always become more positive as aridity increases. Meta-analyses across taxa and biomes have returned broad support for the prediction derived from the SGH that positive interactions become more important as environmental severity increases (Bertness and Callaway, 1994; Lortie and Callaway, 2006; He et al., 2013; Romero et al., 2015; Adams et al., 2022). However, with respect to aridity gradients in drylands, and especially with exotic species, support for the SGH has varied (Butterfield et al., 2016; Bashirzadeh et al., 2022). For instance, Lucero et al. (2021) showed that invasive B. tectorum formed strong positive associations with native shrubs along a substantial aridity gradient across the Great Basin, Mojave, and San Joaquin Deserts, but relative aridity did not predict the intensity of these associations. Similarly, our analyses found no support for the SGH for any vegetation measure along our aridity gradient. It is possible that our gradient was too shallow to detect significant aridity-RII relationships, but our findings over a single growing season coincide with similar studies across much steeper aridity gradients (Lucero et al., 2021). We hypothesize that the intensity of shrub-invasive annual associations may not closely track aridity if invasive annuals are less limited by water than other resources, such as nutrients (Rao and Allen, 2010; Suazo et al., 2012; Bansal et al., 2014; Gill et al., 2018; see discussion below). In this context, an exciting next step would be to experimentally evaluate shrub facilitation of invasive species along multiple, interacting stress gradients.
Our findings indicate that desert shrubs can alter biotic interactions between invasive plants and their neighbors. Invasive species in deserts can inhibit native species in both shrub and open microsites (Brooks, 2000; Bishop et al., 2019), and many studies have investigated the potential for environmental factors to shift competitive outcomes (Hart and Marshall, 2013). Our study reinforces this literature by highlighting the potential for native shrubs to indirectly facilitate plant invasions by increasing the competitive impacts of invasive species (Lortie et al., 2021). We hypothesize that negative B. rubens-herbaceous neighbor relationships (evidence for competition) arose near but not away from shrubs due to relatively fertile soils near shrubs (Schlesinger et al., 1996) that ameliorated nutrient limitation and favored the performance of B. rubens over neighbors. Across the literature, nutrient enrichment almost always promotes the local dominance of invasive species (Brooks, 2003; Levine et al., 2004; Davis and Pelsor, 2008; Besaw et al., 2011). Of course, we do not discount the possibility that enhanced soil water near shrubs relative to open areas (Segoli et al., 2012) also bolstered the competitive impacts of B. rubens because water and nutrient availability act synergistically to affect plant performance (Rao and Allen, 2010; Suazo et al., 2012; Plett et al., 2020). These possibilities underscore the need to experimentally manipulate shrub-invasive annual interactions along mixed water and nutrient gradients. Importantly, this pattern arose only for B. rubens, suggesting that the risk of facilitated invasion (both directly via increased abundance and indirectly via increased impacts) is particularly high for this species across our study area. All of the invasive species we examined can negatively impact natives, but in our system, B. rubens appears to be the invader most responsible for displacing native annuals under shrubs, though experimentation is needed to confirm this.
This observational study did not test for mechanisms of species-specific facilitation, but the functional traits of our focal species may provide some clues (Funk et al., 2017). Facilitation in plant communities can arise via a number of pathways, including abiotic stress amelioration, herbivore protection, pollinator attraction, substrate modification, seed trapping, soil moisture retention, and soil nutrient enhancement (Flores and Jurado, 2003; Filazzola and Lortie, 2014). Not all shrub species are equal facilitators because not all shrub species have equal traits (McAuliffe, 1988). For instance, Larrea tridentata, the dominant shrub species across our Mojave Desert sites, can be a relatively poor facilitator compared to other shrub species due to strong competitive effects and allelopathy (reviewed by Callaway, 2007), though we found limited evidence for this. A key mechanism that determines whether shrub-neighbor interactions are positive or negative is the match between what the shrub offers and what the beneficiary species requires (Paterno et al., 2016). Importantly, the offerings and requirements of species depend upon their functional traits (Funk et al., 2017). Thus, species-specific patterns of shrub-annual associations in our study likely reflect the unique functional traits of our focal species. In this context, our three most commonly encountered invasive annual species—B. rubens, Schismus spp., and E. cicutarium—have distinct physiological characteristics. Relative to other Mojave Desert species, B. rubens uses water very rapidly, has high N uptake, and high net assimilation; Schismus spp. has thin leaves, low leaf mass per area, and low leaf N content; and E. cicutarium may be less physiologically specialized than B. rubens or Schismus spp. (DeFalco et al., 2003; Steers et al., 2011). These trait sets suggest that B. rubens can rapidly capitalize on water and nutrient additions, Schismus spp. has relatively low N requirements, and E. cicutarium may have less specialized physiological requirements than B. rubens or Schismus spp. (DeFalco et al., 2003; Steers et al., 2011). These functional trait syndromes coincide nicely with our species-specific findings: B. rubens associated strongly and consistently with shrubs, perhaps due to its ability to rapidly capitalize on shrub-mediated resource additions; Schismus spp. consistently formed negative associations with shrubs, perhaps due to its relatively low requirements for shrub-mediated nutrient additions; and E. cicutarium varied in its spatial association with shrubs, perhaps due to its intermediate or facultative requirements for shrub-mediated resources. These possibilities present exciting avenues for future experimentation.
We found a pattern consistent with invasional meltdown in open microsites. Invasional meltdown, or positive interactions among exotic species (Simberloff and Von Holle, 1999), can accelerate the transformation of communities by exotic invaders (Braga et al., 2018). In this context, we found positive relationships between B. rubens abundance and exotic species richness in open microsites across the study area. This finding closely coincides with that of Lucero et al. (2021), which reported a positive relationship between B. tectorum abundance and exotic species richness in open microsites across the Great Basin Desert. Because our findings were based on abundance relationships, it is unclear if B. rubens directly facilitated exotic species richness, or if B. rubens and the broader exotic community capitalized on similar advantageous microsites away from shrubs. Experimentation is necessary to disentangle these possibilities and explain why these patterns emerged in open, not shrub, microsites. Again, we emphasize that we only detected this pattern for B. rubens.
Along an aridity gradient across the Mojave and San Joaquin Deserts, invasive annual species formed highly species-specific associations with native shrubs that varied from positive to negative. Interestingly, the direction and intensity of these associations were unaffected by shrub size or relative aridity, and we found correlative evidence that native shrubs exacerbated the negative impacts of B. rubens (but no other invader) on neighboring annuals. We conclude that native shrubs have considerable potential to increase the abundance and impacts of invasive plant species across broad spatial scales, but this potential may depend more upon invader identity than environmental severity.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
JL, CL, RC, and AF conceived the idea. JL collected the data, analyzed the data, and wrote the manuscript. RC, CL, and AF contributed to editorial input. All authors contributed substantially to the development of this manuscript.
We acknowledge financial support from the New Mexico State University Agricultural Experiment Station, a Bureau of Land Management Cooperative Agreement with NMSU, a York Science Fellowship award, a Natural Sciences and Engineering Research Council of Canada Discovery Grant, and National Science Foundation EPSCoR Cooperative Agreement OIA-1757351.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
JL thanks G.W. Gong for fantastic advice. As always, N.B. Harris provided exceptional logistical support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.886690/full#supplementary-material
Abella, S. R., and Smith, S. D. (2013). Annual-perennial plant relationships and species selection for desert restoration. J. Arid Land 5, 298–309. doi: 10.1007/s40333-013-0172-0
Adams, A. E., Besozzi, E. M., Shahrokhi, G., and Patten, M. A. (2022). A case for associational resistance: apparent support for the stress gradient hypothesis varies with study system. Ecol. Lett. 25, 202–217. doi: 10.1111/ele.13917
Armas, C., Ordinales, R., and Pugnaire, F. I. (2004). Measuring plant interactions: a new comparative index. Ecology 85, 2682–2686. doi: 10.1890/03-0650
Badano, E. I., Samour-Nieva, O. R., Flores, J., Flores-Flores, J. L., Flores-Cano, J. A., and Rodas-Ortíz, J. P. (2016). Facilitation by nurse plants contributes to vegetation recovery in human-disturbed desert ecosystems. J. Plant Ecol. 9, 485–497. doi: 10.1093/jpe/rtw002
Báez, S., and Collins, S. L. (2008). Shrub invasion decreases diversity and alters community stability in northern Chihuahuan Desert plant communities. PLoS One 3:e2332. doi: 10.1371/journal.pone.0002332
Bansal, S., James, J. J., and Sheley, R. L. (2014). The effects of precipitation and soil type on three invasive annual grasses in the western United States. J. Arid Environ. 104, 38–42. doi: 10.1016/j.jaridenv.2014.01.010
Bashirzadeh, M., Soliveres, S., Farzam, M., and Ejtehadi, H. (2022). Plant-plant interactions determine taxonomic, functional and diversity in severe ecosystems. Glob. Ecol. Biogeogr. 31, 649–662. doi: 10.1111/geb.13451
Belnap, J., and Sherrod, S. K. (2008). “Soil amendment effects on the exotic annual grass Bromus tectorum L. and facilitation of its growth by the native perennial grass Hilaria jamesii (Torr.) Benth,” in Herbaceous Plant Ecology, ed. A. G. Van der Valk (Dordrecht: Springer), 709–721. doi: 10.1007/s11258-008-9463-5
Bertness, M. D., and Callaway, R. M. (1994). Positive interactions in communities. Trends Ecol. Evol. 9, 191–193. doi: 10.1016/0169-5347(94)90088-4
Besaw, L. M., Thelen, G. C., Sutherland, S., Metlen, K., and Callaway, R. M. (2011). Disturbance, resource pulses and invasion: short-term shifts in competitive effects, not growth responses, favour exotic annuals. J. Appl. Ecol. 48, 998–1006. doi: 10.1111/j.1365-2664.2011.01988.x
Bishop, T. B., Gill, R. A., McMillan, B. R., and St. Clair, S. B. (2019). Fire, rodent herbivory, and plant competition: implications for invasion and altered fire regimes in the Mojave Desert. Oecologia 192, 155–167. doi: 10.1007/s00442-019-04562-2
Blackburn, T. M., Pyšek, P., Bacher, S., Carlton, J. T., Duncan, R. P., Jarošík, V., et al. (2011). A proposed unified framework for biological invasions. Trends Ecol. Evol. 26, 333–339. doi: 10.1016/j.tree.2011.03.023
Braga, R. R., G—́omez-Aparicio, L., Heger, T., Vitule, J. R. S., and Jeschke, J. M. (2018). Structuring evidence for invasional meltdown: broad support but with biases and gaps. Biol. Invasions 20, 923–936. doi: 10.1007/s10530-017-1582-2
Bronstein, J. L. (2009). The evolution of facilitation and mutualism. J. Ecol. 97, 1160–1170. doi: 10.1111/j.1365-2745.2009.01566.x
Brooks, M. L. (2000). Competition between alien annual grasses and native annual plants in the Mojave Desert. Am. Midland Nat. 144, 92–108. doi: 10.1674/0003-0031(2000)144[0092:cbaaga]2.0.co;2
Brooks, M. L. (2003). Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. J. Appl. Ecol. 40, 344–353. doi: 10.1046/j.1365-2664.2003.00789.x
Brooks, M. L., and Berry, K. H. (2006). Dominance and environmental correlates of alien annual plants in the Mojave Desert, USA. J. Arid Environ. 67, 100–124. doi: 10.1016/j.jaridenv.2006.09.021
Bulleri, F., Eriksson, B. K., Queirós, A., Airoldi, L., Arenas, F., Arvanitidis, C., et al. (2018). Harnessing positive species interactions as a tool against climate-driven loss of coastal biodiversity. PLoS Biol. 16:e2006852. doi: 10.1371/journal.pbio.2006852
Butterfield, B. J., Betancourt, J. L., Turner, R. M., and Briggs, J. M. (2010). Facilitation drives 65 years of vegetation change in the Sonoran Desert. Ecology 91, 1132–1139. doi: 10.1890/09-0145.1
Butterfield, B. J., Bradford, J. B., Armas, C., Prieto, I., and Pugnaire, F. I. (2016). Does the stress-gradient hypothesis hold water? Disentangling spatial and temporal variation in plant effects on soil moisture in dryland systems. Funct. Ecol. 30, 10–19. doi: 10.1111/1365-2435.12592
CAL-IPC (2022). The California Invasive Plant Council. Available online at: https://www.cal-ipc.org/plants [Accessed Feb 22, 2022].
Callaway, R. M. (1998). Are positive interactions species-specific? Oikos 82, 202–207. doi: 10.2307/3546931
Callaway, R. M. (2007). Positive Interactions And Interdependence In Plant Communities. Dordrecht: Springer.
Callaway, R. M., Nadkarni, N. M., and Mahall, B. E. (1991). Facilitation and interference of Quercus douglasii on understory productivity in central California. Ecology 72, 1484–1499. doi: 10.2307/1941122
Cavieres, L. A. (2021). Facilitation and the invasibility of plant communities. J. Ecol. 109, 2019–2028. doi: 10.1111/1365-2745.13627
Cavieres, L. A., Quiroz, C. L., and Molina-Montenegro, M. A. (2008). Facilitation of the non-native Taraxacum officinale by native nurse cushion species in the high Andes of central Chile: are there differences between nurses? Funct. Ecol. 22, 148–156.
Davies, J., Poulsen, L., Schulte-Herbrüggen, B., Mackinnon, K., Crawhall, N., Henwood, W. D., et al. (2021). Conserving Dryland Biodiversity. Nairobi: IUCN.
Davis, M. A., and Pelsor, M. (2008). Experimental support for a resource-based mechanistic model of invasibility. Ecol. Lett. 4, 421–428. doi: 10.1046/j.1461-0248.2001.00246.x
DeFalco, L. A., Bryla, D. R., Smith-Longozo, V., and Nowak, R. S. (2003). Are Mojave Desert annual species equal? Resource acquisition and allocation for the invasive grass Bromus madritensis subsp. rubens (Poaceae) and two native species. Am. J. Bot. 90, 1045–1053. doi: 10.3732/ajb.90.7.1045
DiTomaso, J. M., Masters, R. A., and Peterson, V. F. (2010). Rangeland invasive plant management. Rangelands 32, 43–47.
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722. doi: 10.1111/j.1461-0248.2011.01630.x
Filazzola, A., and Lortie, C. J. (2014). A systematic review and conceptual framework for the mechanistic pathways of nurse plants. Glob. Ecol. Biodivers. 23, 1335–1345. doi: 10.1111/geb.12202
Filazzola, A., Lortie, C. J., Westphal, M. F., and Michalet, R. (2020). Species specificity challenges the predictability of facilitation along a regional desert gradient. J. Veg. Sci. 31, 887–898. doi: 10.1111/jvs.12909
Flores, J., and Jurado, E. (2003). Are nurse–protégé interactions more common among plants from arid environments? J. Veg. Sci. 14, 911–916. doi: 10.1111/j.1654-1103.2003.tb02225.x
Funk, J. L., Larson, J. E., Ames, G. M., Butterfield, B. J., Cavender-Bares, J., Firn, J., et al. (2017). Revisiting the Holy Grail: using plant functional traits to understand ecological processes. Biol. Rev. 92, 1156–1173. doi: 10.1111/brv.12275
Gill, R. A., O’Connor, R. C., Rhodes, A., Bishop, T. B., Laughlin, D. C., and St. Clair, S. B. (2018). Niche opportunities for invasive annual plants in dryland ecosystems are controlled by disturbance, trophic interactions, and rainfall. Oecologia 187, 755–765. doi: 10.1007/s00442-018-4137-z
Griffith, A. B. (2010). Positive effects of native shrubs on Bromus tectorum demography. Ecology 91, 141–154. doi: 10.1890/08-1446.1
Hart, S. P., and Marshall, D. J. (2013). Environmental stress, facilitation, competition, and coexistence. Ecology 94, 2719–2731. doi: 10.1890/12-0804.1
He, Q., Bertness, M. D., and Altieri, A. H. (2013). Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 16, 695–706. doi: 10.1111/ele.12080
Holzapfel, C., and Mahall, B. (1999). Facilitation and interference between shrubs and annuals in the Mojave Desert. Ecology 80, 1747–1761. doi: 10.1890/0012-9658(1999)080[1747:bfaibs]2.0.co;2
Huang, J., Yu, H., Guan, X., Wang, G., and Guo, R. (2016). Accelerated dryland expansion under climate change. Nat. Clim. Change 6, 166–171. doi: 10.1038/nclimate2837
Kane, J. M., Meinhardt, K. A., Chang, T., Cardall, B. L., and Whitham, T. G. (2011). Drought-induced mortality of a foundation species (Juniperus monosperma) promotes positive afterlife effects in understory vegetation. Plant Ecol. 212, 733–741. doi: 10.1007/s11258-010-9859-x
Khosravi Mashizi, A., and Sharafatmandrad, M. (2019). Assessing the effects of shrubs on ecosystem functions in arid sand dune ecosystems. Arid Land Res. Manage. 34, 171–187. doi: 10.1080/15324982.2019.1634655
Kuznetsova, A., Brockhoff, P. B., and Christensen, R. H. B. (2017). lmertest package: tests in linear mixed effects models. J. Stat. Softw. 82, 1–26.
le Roux, P. C., and McGeoch, M. A. (2010). Interaction intensity and importance along two stress gradients: adding shape to the stress-gradient hypothesis. Oecologia 162, 733–745. doi: 10.1007/s00442-009-1484-9
Levine, J. M., Adler, P. B., and Yelenik, S. G. (2004). A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 7, 975–989. doi: 10.1111/j.1461-0248.2004.00657.x
Llambi, L. D., Hupp, N., Saez, A., and Callaway, R. M. (2018). Reciprocal interactions between a facilitator, natives, and exotics in tropical alpine plant communities. Perspect. Plant Ecol. Evol. Syst. 30, 82–88. doi: 10.1016/j.ppees.2017.05.002
López, R. P., Squeo, F. A., Armas, C., Kelt, D. A., and Gutierrez, J. R. (2016). Enhanced facilitation at the extreme end of the aridity gradient in the Atacama Desert: a community-level approach. Ecology 97, 1593–1604. doi: 10.1890/15-1152.1
Lortie, C. J., and Callaway, R. M. (2006). Re-analysis of meta-analysis: support of the stress-gradient hypothesis. J. Ecol. 94, 7–16. doi: 10.1111/j.1365-2745.2005.01066.x
Lortie, C. J., Filazzola, A., Brown, C., Lucero, J. E., Zuliani, M., Ghazian, N., et al. (2021). Facilitation enables plant invasions and indirect negative interactions. Oikos 130, 1056–1061. doi: 10.1002/ece3.1483
Lortie, C. J., Gruber, E., Filazzola, A., Noble, T., and Westphal, M. (2018). The Groot effect: plant facilitation and desert shrub regrowth following extensive damage. Ecol. Evol. 8, 706–715. doi: 10.1002/ece3.3671
Lucero, J. E., Callaway, R. M., Faist, A. M., and Lortie, C. J. (2021). An unfortunate alliance: native shrubs increase the abundance, performance, and apparent impacts of Bromus tectorum across a regional aridity gradient. Basic Appl. Ecol. 57, 41–53. doi: 10.1016/j.baae.2021.09.001
Lucero, J. E., Noble, T., Haas, S., Westphal, M., Butterfield, H. S., and Lortie, C. J. (2019). The dark side of facilitation: native shrubs facilitate exotic annuals more strongly than native annuals. NeoBiota 44, 75–93. doi: 10.3897/neobiota.44.33771
Lucero, J. E., Seifan, M., Callaway, R. M., and Lortie, C. J. (2020). Positive associations with native shrubs are intense and important for an exotic invader but not the native annual community across an aridity gradient. Divers. Distrib. 26, 1177–1197. doi: 10.1111/ddi.13111
McAuliffe, J. R. (1988). Markovian dynamics of simple and complex desert plant communities. Am. Nat. 131, 459–490. doi: 10.1086/284802
Michalet, R., and Pugnaire, F. I. (2016). Facilitation in communities: underlying mechanisms, community and ecosystem implications. Funct. Ecol. 30, 3–9. doi: 10.1111/1365-2435.12602
Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-Being. Washington, DC: Island Press.
Northfield, T. D., Laurance, S. G. W., Mayfield, M. M., Paini, D. R., Snyder, W. E., Stouffer, D. B., et al. (2018). Native turncoats and indirect facilitation of species invasions. Proc. R. Soc. B 285:20171936. doi: 10.1098/rspb.2017.1936
Paterno, G. B., Filho, J. A. S., and Ganade, G. (2016). Species-specific facilitation, ontogenetic shifts and consequences for plant community succession. J. Veg. Sci. 27, 606–615. doi: 10.1111/jvs.12382
Pearson, D. E., Ortega, Y. E., Eren, Ö, and Hierro, J. (2016). Quantifying “apparent” impact and distinguishing impact from invasiveness in multispecies plant invasions. Ecol. Appl. 26, 162–173. doi: 10.1890/14-2345
Pescador, D. S., Chacon-Labella, J., de la Cruz, M., and Escudero, A. (2014). Maintaining distances with the engineer: patterns of coexistence in plant communities beyond the patch-bare dichotomy. New Phytol. 204, 140–148. doi: 10.1111/nph.12899
Phillips, M. L., McNellis, B. E., Howell, A., Lauria, C. M., Belnap, J., and Reed, S. C. (2022). Biocrusts mediate a new mechanism for land degradation under a changing climate. Nat. Clim. Change 12, 71–76. doi: 10.1038/s41558-021-01249-6
Plett, D. C., Ranathunge, K., Melino, V. J., Kuya, N., Uga, Y., and Kronzuker, H. J. (2020). The intersection of nitrogen nutrition and water use in plants: new paths toward improved crop productivity. J. Exp. Bot. 71, 4452–4468. doi: 10.1093/jxb/eraa049
R Development Core Team (2020). R: A Language And Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rao, L. E., and Allen, E. B. (2010). Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 162, 1035–1046. doi: 10.1007/s00442-009-1516-5
Reid, C. R., Goodrich, S., and Bowns, J. E. (2006). “Cheatgrass and red brome: history and biology of two invaders,” in Proceedings–Shrublands Under Fire: Disturbance and Recovery in a Changing World. Fort Collins, CO: US Department of Agriculture, Forest Service, Rocky Mountain Research Station, 27–32.
Romero, G. Q., Goncalves-Souza, T., Vieira, C., and Koricheva, J. (2015). Ecosystem engineering effects on species diversity across ecosystems: a meta-analysis. Biol. Rev. 90, 877–890. doi: 10.1111/brv.12138
Saccone, P., Pages, J. P., Griel, J., and Michalet, R. (2010). Acer negundo invasion along a successional gradient: early direct facilitation by native pioneers and late indirect facilitation by conspecifics. New Phytol. 187, 831–842. doi: 10.1111/j.1469-8137.2010.03289.x
Schlesinger, W. H., Raikes, J. A., Hartley, A. E., and Cross, A. E. (1996). On the spatial pattern of soil nutrients in desert ecosystems. Ecology 77, 364–374. doi: 10.2307/2265615
Segoli, M., Ungar, E. D., Giladi, I., Arnon, A., and Shachak, M. (2012). Untangling the positive and negative effects of shrubs on herbaceous vegetation in drylands. Landsc. Ecol. 27, 899–910. doi: 10.1007/s10980-012-9736-1
Simberloff, D., and Von Holle, B. (1999). Positive interactions of nonindigenous species: invasional meltdown? Biol. Invasions 1, 21–32.
Slate, M. L., Tsombou, F. M., Callaway, R. M., Majumdar, S., and Inderjit El-Keblawy, A. A. (2020). Exotic Prosopis suppresses understory diversity but promotes agricultural weeds more than a native congener. Plant Ecol. 221, 659–669. doi: 10.1007/s11258-020-01040-1
Soliveres, S., Eldridge, D. J., Hemmings, F., and Maestre, F. (2012). Nurse plant effects on plant species richness in drylands: The role of grazing, rainfall, and species-specificity. Perspect. Plant Ecol. Evol. Syst. 14, 402–410. doi: 10.1016/j.ppees.2012.09.003
Steers, R. J., Funk, J. L., and Allen, E. B. (2011). Can resource-use traits predict native vs. exotic plant success in carbon amended soils? Ecol. Appl. 21, 1211–1224. doi: 10.1890/09-2345.1
Suazo, A. A., Spencer, J. E., Engel, E. C., and Abella, S. R. (2012). Responses of native and non-native Mojave Desert winter annuals to soil disturbance and water additions. Biol. Invasions 14, 215–227. doi: 10.1007/s10530-011-9998-6
Valiente-Benuet, A., and Ezcurra, E. (1991). Shade as a cause of the association between the cactus Neobuxbaumia tetezo and the nurse plant Mimosa luisana in the Tehuacán Valley, Mexico. J. Ecol. 79, 961–971. doi: 10.2307/2261091
Wright, A. J., Barry, K. E., Lortie, C. J., and Callaway, R. M. (2021). Biodiversity and ecosystem functioning: have our experiments and indices been underestimating the role of facilitation? J. Ecol. 109, 1962–1968. doi: 10.1111/1365-2745.13665
Keywords: desert, facilitated invasion, facilitation, invasive species, positive interactions, shrubs, species-specificity, stress-gradient hypothesis
Citation: Lucero JE, Faist AM, Lortie CJ and Callaway RM (2022) Risk of Facilitated Invasion Depends Upon Invader Identity, Not Environmental Severity, Along an Aridity Gradient. Front. Ecol. Evol. 10:886690. doi: 10.3389/fevo.2022.886690
Received: 28 February 2022; Accepted: 19 April 2022;
Published: 09 May 2022.
Edited by:
Bruce Arthur Osborne, University College Dublin, IrelandReviewed by:
Richard A. Gill, Brigham Young University, United StatesCopyright © 2022 Lucero, Faist, Lortie and Callaway. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jacob E. Lucero, amVsdWNlcm9Abm1zdS5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.