 Daniel Aguilera-Olivares1*†
Daniel Aguilera-Olivares1*† Bárbara Torres-Guzmán2
Bárbara Torres-Guzmán2 Alberto Arab3†
Alberto Arab3† Luis Flores-Prado4†
Luis Flores-Prado4† Hermann M. Niemeyer5*†
Hermann M. Niemeyer5*†- 1Departamento de Silvicultura, Universidad de Concepción, Concepción, Chile
- 2Departamento de Ciencias Ecológicas, Universidad de Chile, Santiago, Chile
- 3Centro de Ciências Naturais e Humanas, Universidade Federal do ABC (UFABC), Santo André, Brazil
- 4Instituto de Entomología, Universidad Metropolitana de Ciencias de la Educación, Santiago, Chile
- 5Departamento de Química, Universidad de Chile, Santiago, Chile
Single-piece nesting termites live and forage in the same piece of wood throughout their life, which limit their colony size. In certain species, more than one colony thrive in a given piece of wood (multicolonial substrate) and intraspecific competition become important in this limited resource, as has been reported in Zootermopsis nevadensis (Hagen, 1858) and Neotermes chilensis (Blattodea: Kalotermitidae) (Blanchard, 1851). The effects of such competition have been described mainly at population and colony levels rather than at the individual level. In eusocial insects such as termites, intraspecific competition constitutes a stress factor imposed to a colony as a whole and should also cause developmental instability in soldiers produced under such conditions. Investment in the production of soldiers involves a trade-off between colony maintenance costs and defense benefits. Hence, we hypothesize that body size and fluctuating asymmetry, two indicators of developmental instability, will increase when two or more colonies of N. chilensis share a piece of wood (high intraspecific competition scenario). Our results showed that soldiers developing in multicolonial substrates were indeed larger and more asymmetric than soldiers developing in unicolonial substrates. The large body size in a soldier could improve its chance to win a physical contest with a non-nestmate opponent; thus, despite the high cost to produce large soldiers in small colonies, larger soldier production could be an adaptative strategy to avoid being outcompeted. However, the effects of deviations from perfect symmetry on soldier performance are not clear.
Introduction
Intraspecific competition studies in social insects have focused mainly at the colony level (Passera et al., 1996; Thomas et al., 2005; Sanada-Morimura et al., 2006; Cronin et al., 2012; Lloyd and Poulin, 2014; Blight et al., 2016) and at the population level (Holway et al., 1998; Adams and Tschinkel, 2001; Korb and Linsenmair, 2001; Boulay et al., 2007; Grohmann et al., 2010; Bourguignon et al., 2011; Perdereau et al., 2011; Tsuji, 2013; Pringle and Tarnita, 2017); however, the effects of intraspecific competition at the individual level are poorly studied.
Termites are eusocial species that live in colonies organized into three castes (Thorne, 1996; Korb and Hartfelder, 2008; Eggleton, 2011): (i) primary reproductives involved in reproduction, (ii) soldiers involved in defense against intruders, and (iii) true workers (or pseudergates) involved in nest keeping and taking care of juveniles and other castes and also, together with soldiers, in defense. Mandibles of termite soldiers are morphologically adapted to defense (Scholtz et al., 2008); hence, soldiers are unable to feed themselves and they depend on pseudergates or workers for survival (Haverty, 1977; Henderson, 1998). Consequently, investment in the production of soldiers involves a trade-off between colony maintenance costs and defense benefits (Noirot, 1989; Chouvenc et al., 2015). Pseudergates are undifferentiated and totipotent individuals who may develop into other castes such as soldiers or primary reproductives, or they may spend their whole life as pseudergates, depending on environmental conditions (Ogino et al., 1993; Roisin and Korb, 2011). Thus, environmental factors such as colony conditions (colony size, reproductive status, caste ratio, resource availability, etc.), temperature and seasonality significantly affect hormone and gene expressions (Scharf et al., 2007; Miura and Scharf, 2011) which trigger the differentiation from pseudergate to presoldier instar. This process may also be enhanced by ecological factors such as inter and/or intraspecific competition as a form of induced defense at the colony level (Passera et al., 1996; Aguilera-Olivares et al., 2017).
Termite species may be classified according to their nesting behavior in two main categories: separate-piece nesting termites, where foraging substrates and nesting substrate are different; and single-piece nesting termites, who spent their whole life in the same piece of wood (Abe, 1991; Shellman-Reeve, 1997). The colonies of separate-piece nesting termites are composed commonly by true workers and their size ranges from thousands to millions of individuals who can forage and move for thousands of square meters. On the other hand, colonies of single-piece nesting termites are characterized by the presence of pseudergates and colony size (usually no larger than a few thousand individuals) is limited by the volume of the piece of wood where they thrive (Abe, 1991; Shellman-Reeve, 1997; Korb and Hartfelder, 2008; Mizumoto and Bourguignon, 2021). If several colonies of single-piece nesting termites share the same substrate, inter and/or intra-specific competition is expected to occur, particularly when nesting resources become limited (Thorne et al., 2003; Ripa and Luppichini, 2004; Amarillo-Suárez et al., 2011).
Neotermes chilensis (Blattodea: Kalortermitidae) (Blanchard, 1851) is a single-piece nesting termite (i.e., it completes its life cycle within a finite resource) which uses dry scapes (stems of inflorescences) of the bromeliads Puya alpestris ssp. zoellneri [ex P. berteroniana—Zizka et al. (2013)] and P. chilensis (Molina, 1782) among its hosts (Aguilera-Olivares et al., 2015). In previous work at the same study site (Aguilera-Olivares et al., 2017), we reported that about half of the scapes were occupied by a single colony (unicolonial substrates) and the other half contained between 2 and 9 colonies (multicolonial substrates). Additionally, we observed galleries interconnecting two colonies within a scape which had been blocked with sawdust, probably following an agonistic interaction. Under this intraspecific competition scenario, the soldiers/non-soldiers ratio significantly increased. This implies that colonies exposed to competition have an inherently higher cost due to the increased production and maintenance of soldiers, and since resources are finite in this termite, an increased developmental stress is imposed on these soldiers. N. chilensis stands as an interesting species to study the effect of intraspecific competition in an eusocial insect because: 1) it lives on a limited resource; 2) it is not able to move to another resource, so it must compete when the nesting substrate is shared with other colonies; 3) due to low colony sizes (no more than 500 individuals) it is easier to study than other termite species with typical colony sizes in the thousands to millions of individuals; and 4) about 90% of individuals inside a colony are pseudergates, i.e. totipotential individuals which could develop as soldiers, alates or remain as pseudergates their whole life according environmental signals. Moreover, the soldier caste of N. chilensis appears as particularly relevant in terms of intraspecific competition at the individual level, especially the consequences of stress during development.
Developmental stability is the ability of an organism to produce its developmentally programed phenotype despite epigenetic perturbations; contrastingly, developmental instability occurs when an organism is unable to buffer those perturbations (Markow, 1995). In species with symmetric morphological traits, the developmentally programed phenotype is one with perfect symmetry of such traits; and any random deviations from this symmetry on individuals inside a population leads to fluctuating asymmetry (Palmer, 1994; Graham et al., 2010). These deviations may be associated with genetic stress such as hybridization (Graham and Felley, 1985; Handy et al., 2004), and with environmental stress such as food limitation (Swaddle and Witter, 1994), heat shock (Hosken et al., 2000), and intraspecific competition (Witter and Swaddle, 1994). In insects, there is evidence that high levels of larval density constitute an environmental stressor inducing fluctuating asymmetry (Clarke and McKenzie, 1992; Gibbs and Breuker, 2006; Beasley et al., 2013), which may be caused by the effects of intraspecific competition due to a reduction of food availability (Hunt and Allen, 2000). Nevertheless, the effect on fluctuating asymmetry of another scenario of intraspecific competition caused by a restriction in the use of a nesting substrate, as occurs when two or more insect colonies share the same nest, has not been assessed.
On the other hand, body size is a life-history trait which has been used in insects as an indicator of developmental stress (Warren et al., 2006; Couret and Benedict, 2014). Thus, the effect of factors such as altitude (Cushman et al., 1993; Smith et al., 2007; Hoiss et al., 2012), temperature (Bochdanovits and De Jong, 2003), and competition (Heinrich, 1993; Warren et al., 2006; Amarillo-Suárez et al., 2011; Wills et al., 2014; Korallo-Vinarskaya et al., 2015) during insect development are important predictors of body size outcome. When interference competition occurs, body size generally determines who will dominate the resources; thus, larger individuals are frequently more successful than smaller ones (Heinrich and Bartholomew, 1979; Otronen, 1988; Heinrich, 1993; Zobel and Paxton, 2007; Bespalova and Helms, 2014). In termites, significantly higher fluctuating asymmetry has been detected in soldiers of Coptotermes formosanus Shiraki, which develop within incipient colonies (instable and stressful scenario) in comparison with soldiers which develop within mature colonies (more stable environment) (Chouvenc et al., 2014a). Furthermore, studies in three Reticulitermes (R. speratus, R. virginicus, and R. flavipes) and two Cryptotermes (C. secundus and C. domesticus) termite species have shown increased female-to-male ratio because female soldiers are larger than male soldiers and thus have a better chance to win in a physical encounter (Matsuura, 2006; Muller and Korb, 2008). In summary, intraspecific competition is a stressful scenario for termite colonies which could affect body size and symmetry properties of developing individuals and hence their performance.
The aim of this work was to study the effects of intraspecific competition at the individual level in an eusocial insect. If competition in the context of finite resources induces colonies to invest in larger and/or more numerous soldiers, it would directly impose a developmental stress on these individuals. We hypothesized that soldiers of N. chilensis developing in colonies that share a nesting resource with other colonies (high intraspecific competition scenario) would show significantly higher values of fluctuating asymmetry and they would be larger than soldiers whose colonies occur singly within a nesting resource (low intraspecific competition scenario).
Materials and Methods
Species and Study Area
N. chilensis is an endemic termite from Chile distributed between 26 and 33.5°S (Ripa and Luppichini, 2004). The study area (Las Chilcas: 32°52′S; 70°52′W) is located within the sclerophyllous shrub community of central Chile (Gajardo, 1994) whose predominant species are Adesmia arborea Bert. ex Savi (Fabaceae), Colliguaya odorifera Mol. (Euphorbiaceae), Echinopsis chiloensis (Colla) H. Friedrich and G.D. Rowley (Cactaceae), Puya chilensis and P. alpestris ssp. zoellneri (Bromeliaceae). In the study area, N. chilensis builds its nests inside the dry scapes of P. alpestris ssp. zoellneri, with colonies containing up to 500 individuals and it is the only termite species present. Scapes were severed from the rest of the plant, brought to the laboratory in Santiago after being enclosed within a mesh to avoid loss of individuals, and maintained in a breeding room at 16±1°C under darkness for a maximum of 2 weeks until they were dissected.
Collection of Samples
A total of sixteen scapes of P. alpestris ssp. zoellneri were dissected and the number of colonies and of soldiers and non-soldiers were recorded. Eighteen colonies and fifty-three individuals in total were used in this study. Individuals collected were stored in 70% v/v alcohol. Multicolonial soldiers: five scapes had 2 or 3 colonies (mean ± sem: 2.75 ± 0.25 colonies per scape) some of which had soldiers (6.14 ± 1.62 soldiers per colony). A total of seven colonies from multicolonial substrates with 29–178 individuals (104.71 ± 23.3 individuals per colony) were used in the study. The analysis included 22 multicolonial soldiers (2–5 soldiers were extracted from each colony, 3.14 ± 0.46). Unicolonial soldiers: 11 scapes had a unique colony. In these scapes, colony size ranged from 48 to 492 individuals (136.1 ± 38.4 individuals per colony) and all had soldiers (7.91 ± 2.07 soldiers per colony). All the 11 colonies in unicolonial substrates were used in the study. The analysis included 31 unicolonial soldiers (2–4 soldiers were extracted from each colony, 2.82 ± 0.18).
Measurement of Traits
Since termites have a non-quitinized abdomen, body size changes dramatically with hydration level. Hence, only quitinized traits were used (17 in total) to estimate body size (Muller and Korb, 2008; Johnson et al., 2011): six mandibular, seven head and four thoracic traits (described in Table 1 and Figures 1A–E). On the other hand, the left and the right side of four traits were used to assess developmental instability through fluctuating asymmetry (Table 1 and Figure 1C). One photograph was taken of each trait for body size estimation and one photograph was taken of each side of traits selected for the fluctuating asymmetry study. An Olympus® Trinocular Stereo Zoom Microscope Model SZ61 was used with an integrated MSHOT 30 camera; the zoom used for each trait is given in Table 1. Using the software M-shot Digital Imaging System (Micro-shot Technology Co, 2010), the traits were measured in triplicate in three different days to avoid bias and reduce errors.
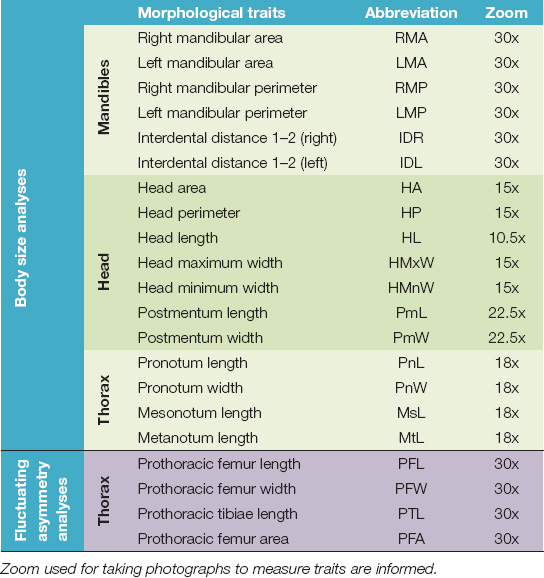
Table 1. Morphological traits used to evaluate body size and fluctuating asymmetry in soldiers of N. chilensis from unicolonial and multicolonial substrates.
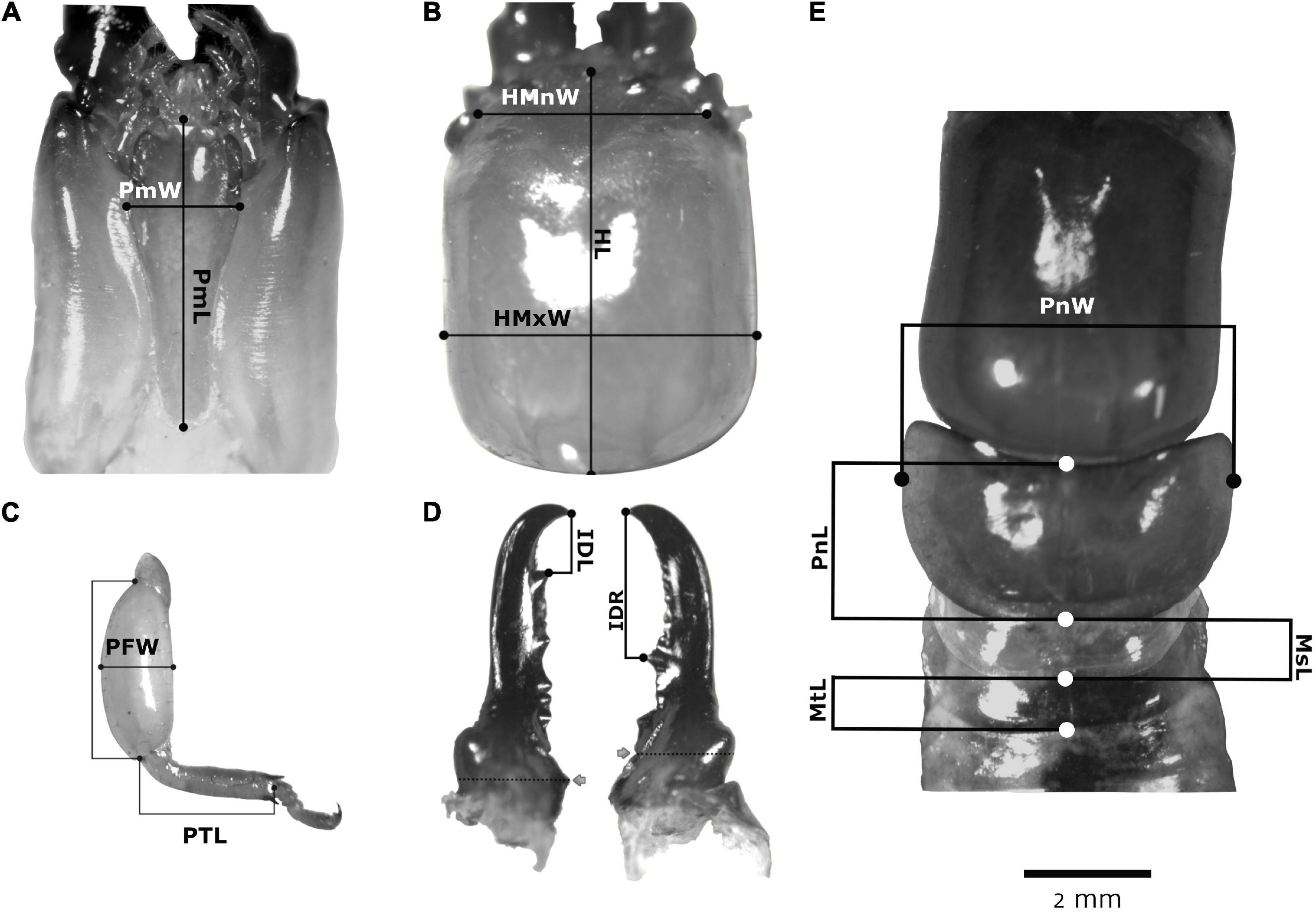
Figure 1. Morphological traits used to evaluate body size and fluctuating asymmetry in soldiers of N. chilensis from unicolonial and multicolonial substrates. (A) Ventral view of the head; (B) dorsal view of the head; (C) prothoracic legs; (D) mandibles and (E) dorsal view of the thorax. Abbreviations used are described in Table 1.
Statistical Analyses
All the analyses and graphics were performed and generated using R version 3.4.4 (R Core Team, 2019).
Body Size
A Principal Component Analyses (PCA) was performed with the 17 traits measured using the FactoMineR package (Le et al., 2008) to retrieve a set of principal component variables that explained major differences in soldier morphology. The total contribution of each trait was calculated using the function fviz.contrib, the most contributed traits were selected as explained in Supplementary Figure 1. The principal components that most explained variation in soldier morphology were then used as predictor variables in Generalized Mixed Models (GLMM), in order to test significant differences in body size between soldiers from unicolonial and multicolonial substrates using the glmmTMB package (Magnusson et al., 2017). Coloniality (unicolonial and multicolonial substrates) was the fixed effect and colony size, i.e., the total number of individuals per colony, was the covariate. In order to control the indirect effect of the colony of origin, this parameter was used as the random factor in the models. The significance of the random effect was tested in reduced models. For each model, we assessed residual distribution using the DHARMa package in R (Hartig, 2021).
Fluctuating Asymmetry
The difference between the mean right side and the mean left side of each trait (R-L) was calculated. Following Palmer (1994), the following analyses were performed: (i) normality of the (R-L) distribution was assessed using the Kolmogorov-Smirnov test with Lilliefors correction to discard significant antisymmetry; (ii) significant deviation from zero was tested using a one-sample t-test to discard significant directional asymmetry; (iii) a regression was performed between R-L and trait size to discard size dependence; and (iv) a two-way ANOVA was performed with soldiers and side (triplicate data for each side) as factors to discard measurement error.
In order to assess deviations from perfect symmetry, two fluctuating asymmetry indexes were chosen (Palmer, 1994):
1) FA1= mean | R-L |, calculated for each four of traits in Table 1. We performed a PCA with the four FA1 index values obtained. We performed a GLMM to test significant differences in fluctuating asymmetry between soldiers from unicolonial and multicolonial substrates with PC1 and PC2 as predictor variables, similar to body size analyses described above.
2) FA10 = σ2, which gives an estimate of the variance between right and left sides after removing the effects of measurement errors and possible directional asymmetry. Thus, from the result of the two-way ANOVA mentioned above, the mean square of the interaction (MSint), mean square of the error (MSerror), number of replicates (Nr), number of soldiers (Ni), and number of sides (Ns) were used to calculate the variance () for each trait (Equation 1) and their approximate degrees of freedom (Equation 2):
Results
Body Size Analysis
Our preliminary PCA showed that the variation in morphology between soldiers from unicolonial and multicolonial substrates was mainly explained by 10 traits (RMA, RMP, IDR, PnW, PnL, LMA, HP, HA, HMxW, and HL) (Supplementary Figure 1A and Supplementary Table 1). These traits were selected and a second PCA was run, in which the first two principal components explained 77% of total variance of soldier morphology (Supplementary Figure 1B and Supplementary Table 2). The first principal component (PC1) was named “head scores” because head variables (head area, head perimeter, head length, and head maximum width) were the most contributory variables (51.6%) to this component (Supplementary Figure 1B and Supplementary Table 2). These variables showed strong positive correlations with this component (correlation coefficients from 0.81 to 0.90—Supplementary Table 2). High values of PC1 represent soldiers with larger heads. The second axis (PC2) was the “mandible and head scores” because mandible traits (right mandible perimeter, right mandible area, interdental distance 1–2 right) and a head trait (head length) were the most contributory variables (67.11%) to this component (Supplementary Figure 1B and Supplementary Table 2). These variables showed positive correlation with mandibular traits (correlation coefficients from 0.45 to 0.70 for RMP—Supplementary Table 2) and a negative correlation with the head trait (correlation coefficient −0.44—Supplementary Table 2). The GLMM on PC1 showed that soldiers from multicolonial substrates had larger heads than soldiers from unicolonial substrates (z = −2.59; p = 0.009) (Figure 2 and Table 2). It was also found that colony size had a significant positive effect on head traits (z = 3.49; p < 0.002) but did not have a significant effect on the second principal component (Table 2). Similar results were obtained when the random effect was excluded from the models (Table 2). No significant deviations from model assumptions were detected (Supplementary Figure 3A).
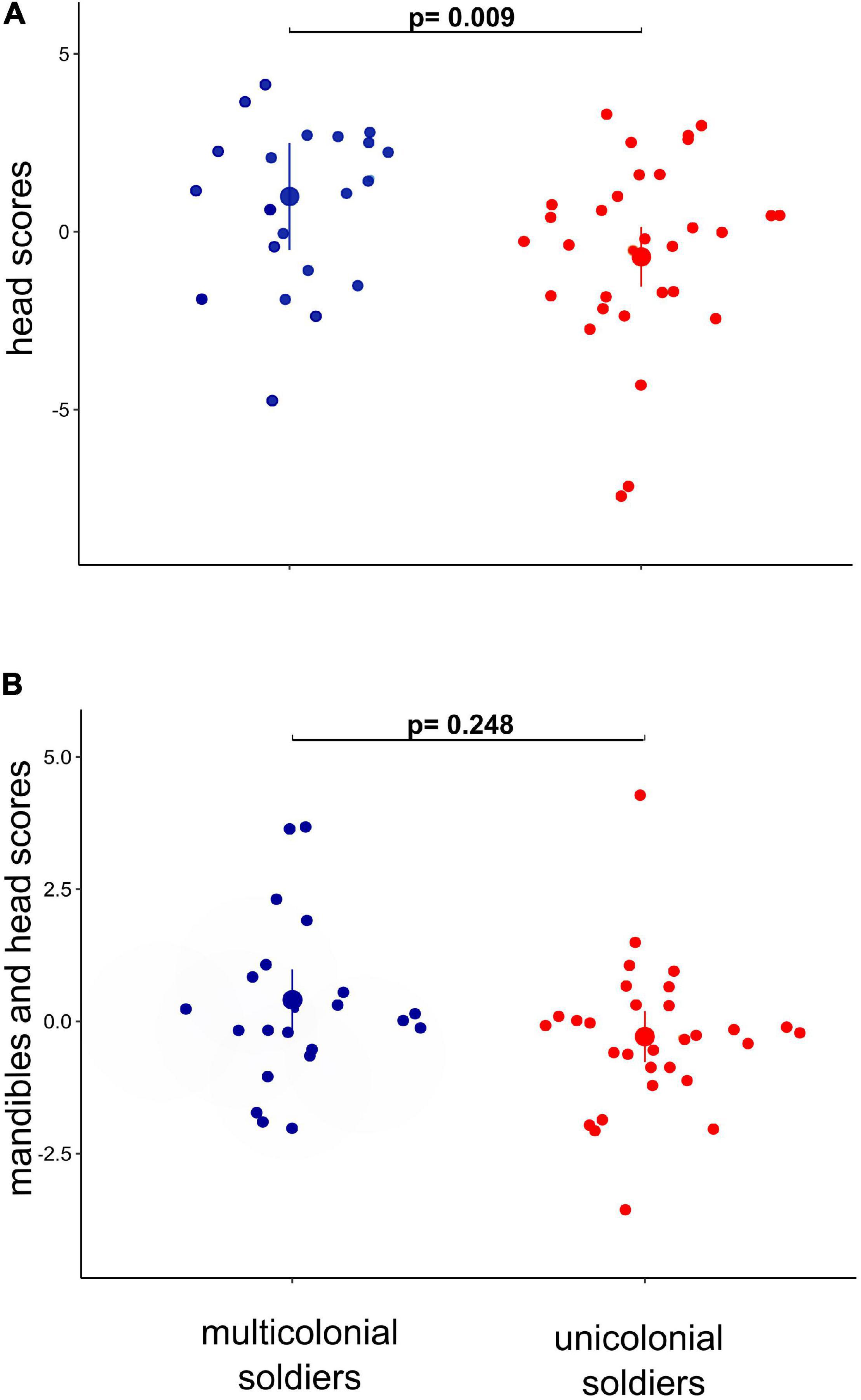
Figure 2. Soldiers’ head (A) and mandible/head (B) components among soldiers from multicolonial and unicolonial substrates. In the plot, the modeled means are represented by large circles, bars are the modeled 95% confidence intervals, and the model-adjusted individual response values are represented by the small dots. Metric scores were obtained from PCA of body size metrics. P-values indicate significance at p < 0.05.
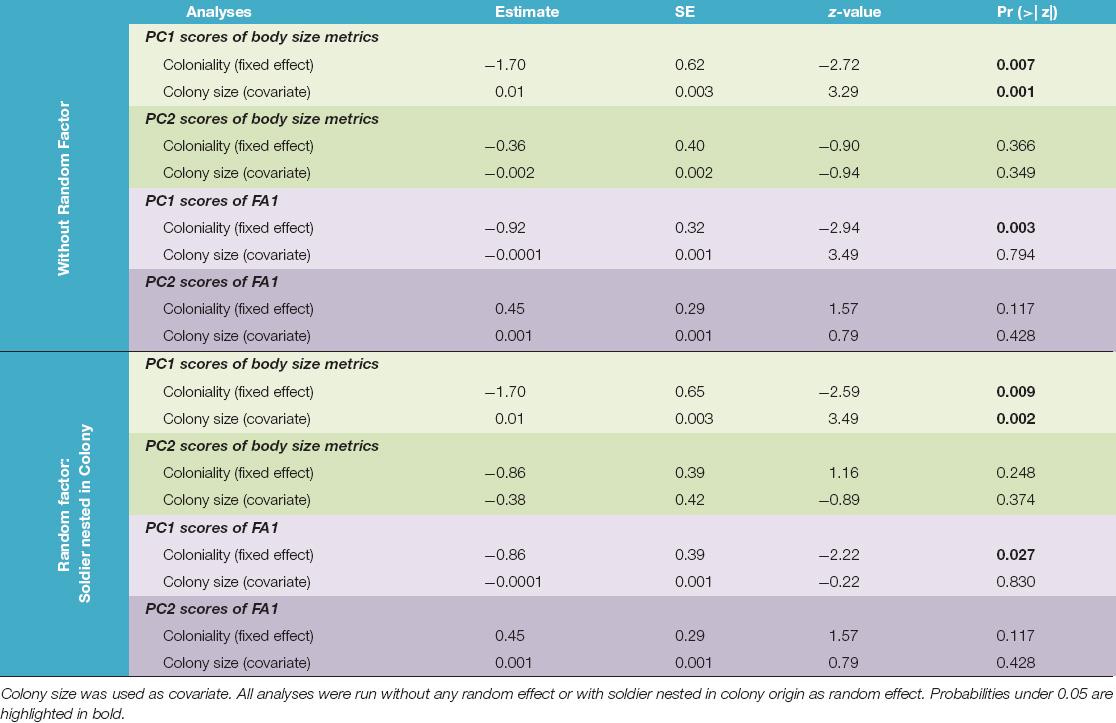
Table 2. Fixed effects of the GLMM analyses for body size metrics (green shadow) and fluctuating asymmetric indexes 1 (FA1) (purple shadow) of soldiers of N. chilensis from multicolonial and unicolonial substrates.
Fluctuating Asymmetry Analysis
Preliminary tests showed non-significant antisymmetry (Supplementary Table 3), non-significant directional asymmetry (Supplementary Table 4), non-significant dependence of the trait on size (Supplementary Table 4); and non-significant measurement errors (Supplementary Table 5) in the four traits analyzed.
The first principal component (PC1) explained the 36.2% of variation among soldiers. Since prothoracic femur area, prothoracic femur length and prothoracic femur width FA1 index values had the highest factor scores for this principal component (Supplementary Table 6), we named it “prothoracic femur” component (Supplementary Figure 2). The second principal component (PC2) explained the 27.7% of variation among soldiers and prothoracic tibia length FA1 index values had the highest factor score (Supplementary Table 6); hence, this principal component was named “prothoracic tibia” (Supplementary Figure 2). GLMM showed that soldiers from multicolonial substrates had significantly higher fluctuating asymmetry indexes than soldiers from unicolonial substrates in prothoracic femur component (z = −2.22; p = 0.027); however, non-significant differences (z = 1.57; p = 0.117) were found in the prothoracic tibia component (Table 2 and Figure 3). Colony size had non-significant effect on both components (Table 2). Similar results were obtained when no random factors were included in the model (Table 2). No significant deviations from model assumptions were detected (Supplementary Figure 3B).
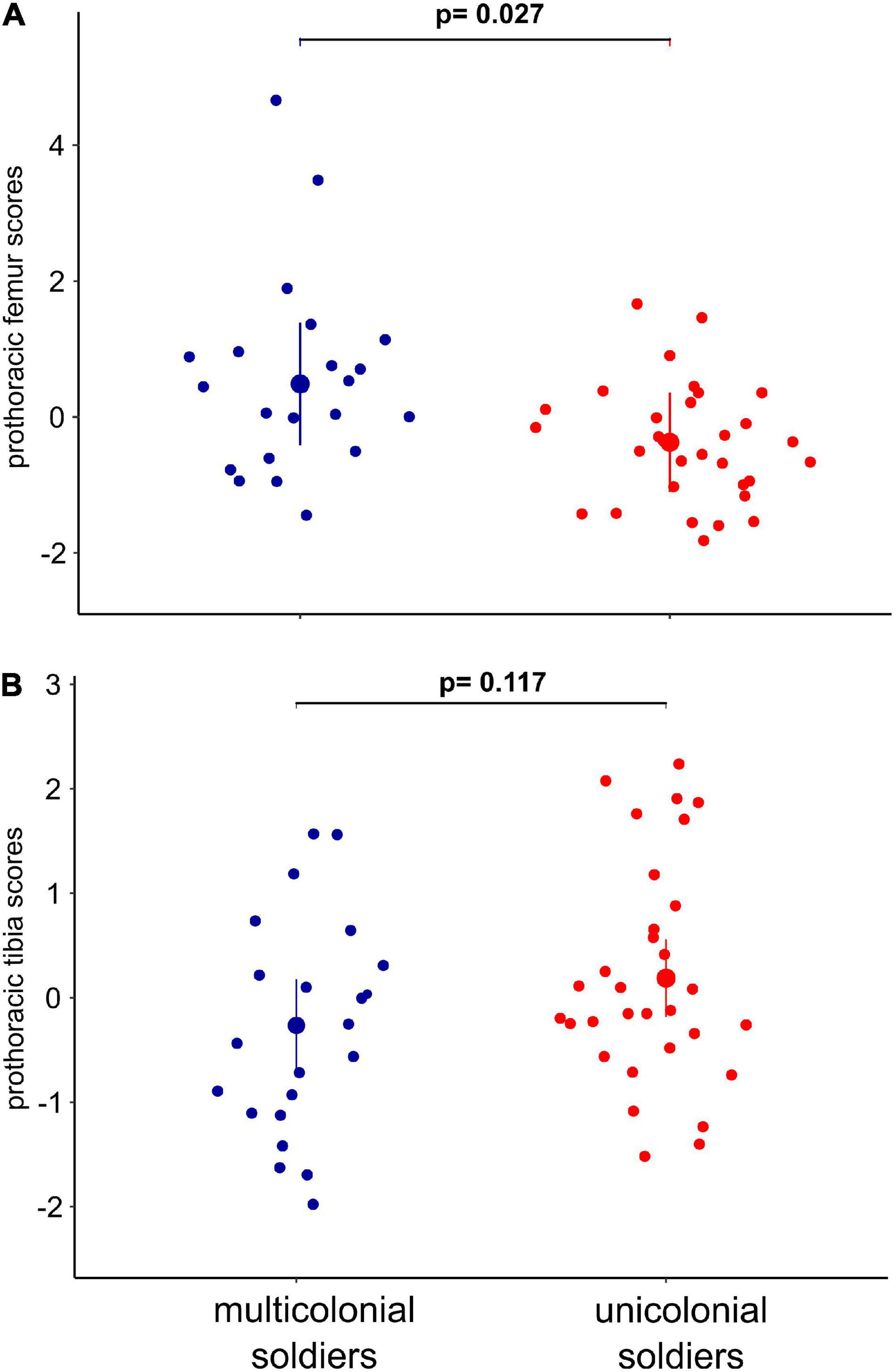
Figure 3. Soldiers’ prothoracic femur (A) and prothoracic tibia (B) principal components from FA1 indexes among soldiers from multicolonial and unicolonial substrates. In the plot, the modeled means are represented by large circles, bars are the modeled 95% confidence intervals of each mean, and the model-adjusted individual response values are represented by the small dots. Scores were obtained from PCA of fluctuating asymmetry indexes (FA1). P-values indicate significance at p < 0.05.
Finally, the FA10 index showed a significantly more fluctuating asymmetry in soldiers from multicolonial substrates than in soldiers from unicolonial substrates in all femur traits (Table 3).
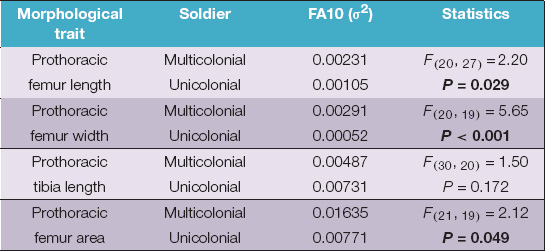
Table 3. Fluctuating asymmetry index 10 (FA10) of four morphological traits of soldiers of N. chilensis from multicolonial and unicolonial substrates.
Discussion
Many abiotic and biotic factors may affect the normal ontogenetic development of insects (Palmer, 1994; Markow, 1995; Graham et al., 2010; Couret and Benedict, 2014). In this work, termite colonies were collected from a common site; hence, those present in unicolonial and multicolonial substrates were exposed to the same abiotic factors. Additionally, N. chilensis was the only termite species present in the study site and N. chilensis termites were the only insects found inside the scapes. Hence, intraspecific competition operating on a termite colony as a whole is expected to be the most important stress factor that may differentially affect unicolonial and multicolonial soldier individual development. All the indexes and analyses used in this work showed significantly higher levels of fluctuating asymmetry in soldiers developed in multicolonial substrates than soldiers developed in unicolonial substrates. Thus, our results showed that the presence of neighboring colonies generated developmental instability in soldier differentiation from pseudergates.
When two or more colonies share a nesting substrate, several scenarios could follow, e.g., their galleries could meet and colonies could fuse, they could fight against each other with the possible consequence of the elimination of one of them, or they could avoid each other. In exotic invasive species, nestmate recognition mechanisms generally fail because genetic diversity in the population is low and relatedness among nestmates is high. This may lead to colony fusion, as has been shown in termites (Perdereau et al., 2010, 2011; Lee et al., 2019) and ants (Holway et al., 1998; Eyer et al., 2018). On the other hand, in native species, where genetic diversity in the population is high and relatedness between colonies is low, colony fusion is rare (Shelton and Grace, 1996; Deheer and Vargo, 2004; Smith et al., 2012). In the population of the native species N. chilensis under study, the mean relatedness of nestmate pairs was shown to be 0.465 ± 0.0085 (mean ± sem), corresponding closely to that expected for full siblings (r = 0.50) and the relatedness between non-nestmate pairs was close to zero (Aguilera-Olivares et al., 2015). This suggests that if two or more colonies meet within the nesting substrate, colony fusion would be rare or inexistent. In fact, nestmate recognition was reported in N. chilensis, soldiers being more aggressive toward non-nestmates than toward nestmates from different castes (Aguilera-Olivares et al., 2016a,b). These observations suggest that individuals from two colonies of N. chilensis within a scape either fight and eliminate opponent colonies or avoid them. Additionally, the colony of origin had a very slight if any effect when its inclusion or exclusion as random factor was considered in the models; thus, despite the fact that individuals inside a colony of N. chilensis are closely related, there is enough variation among soldiers to be considered independent samples, suggesting that the response to intraspecific competition in our analyses is independent of their degree of relatedness.
Body size is a life-history trait that has been used in insects as an indicator of developmental stress (Warren et al., 2006; Couret and Benedict, 2014). Intraspecific competition as a stress factor has been described in literature (Heinrich, 1993; Warren et al., 2006; Amarillo-Suárez et al., 2011; Wills et al., 2014; Korallo-Vinarskaya et al., 2015). In territorial species where interference competition occurs with physical encounters displayed, larger individuals have better chances to win a fight and monopolize resources than smaller individuals (Price et al., 2011; Bespalova and Helms, 2014; Holland et al., 2021). In our study, we demonstrated that soldiers from multicolonial substrates had significantly larger heads than those from unicolonial substrates, strongly suggesting that in a competitive scenario, colonies produce larger soldiers which increase the chance to win fights and monopolize resources. Furthermore, Muller and Korb (2008) showed that females of Zootermopsis nevadensis (Hagen, 1858), a single-piece nesting termite, develop more often into soldiers than males because they are significantly larger, which is an advantage when blocking holes/galleries or fighting with other individuals (Zobel and Paxton, 2007). In our work, quitinized traits of N. chilensis that showed differences in size between uni- and multicolonial substrates are closely related to defensive tasks. Thus, head traits (head area, head perimeter, head length, and head maximum width) could be related to the blocking of galleries with the head and to more developed muscles to produce stronger bites (Bespalova and Helms, 2014). When the scape used by two or more colonies is depleted and the tunnels dug by members of different colonies finally meet, all of these traits could improve the chance to win a fight in a hypothetical scenario of interference intraspecific competition (Heinrich and Bartholomew, 1979; Otronen, 1988; Heinrich, 1993; Zobel and Paxton, 2007; Bespalova and Helms, 2014).
Soldiers totally depend on workers or pseudergates for survival (Haverty, 1977; Henderson, 1998); thus, the soldier production represent a trade-off at colony level between benefits (colony defense) and costs (investment in soldier nurturing) (Noirot, 1989; Chouvenc et al., 2015). Additionally, in incipient colonies some larvae must develops into soldiers instead of workers in order to maintain a relatively stable soldier proportion, which could have consequences in colony growth as a result of a reduction of worker number (Chouvenc et al., 2015). In Coptotermes species, a subterranean and separate-piece nesting termite, incipient colonies with low numbers of workers and hence a reduced nurturing capacity produce smaller and more asymmetric soldiers (i.e., low quality soldiers produced at low cost) as an strategy to keep a stable caste ratio which produces a delay on the growth of colonies (Chouvenc and Su, 2014b; Chouvenc et al., 2015, 2017); while soldiers from mature colonies which contain high number of workers and hence exhibit high nurturing capacity, are larger and symmetric (high quality soldiers produced at high cost) (Chouvenc and Su, 2014b; Chouvenc et al., 2017). In single-piece nesting termites, such as N. chilensis, colony survival is directly related with its capacity to get and defend resources; thus, despite the high cost to produce large soldiers in small colonies, larger soldier production could be an adaptative strategy to avoid being outcompeted.
Conclusion
This study shows for first time the effect of intraspecific competition at the individual level in an eusocial insect. Soldiers of N. chilensis which developed from pseudergates in a high intraspecific competition scenario (multicolonial substrates) were larger and exhibited more asymmetric traits than soldiers which developed in a low intraspecific competition scenario (unicolonial substrates). While the large body size in a soldier could improve its chance to win a physical contest with a non-nestmate opponent, the effects of deviations from perfect symmetry on soldier performance are not clear. In single-piece nesting termites, such as N. chilensis, colony survival is directly related with its capacity to get and defend resources; thus, despite the high cost to produce large soldiers in small colonies, larger soldier production could be an adaptative strategy to avoid being outcompeted. Having shown the effects of intraspecific competition on body size and asymmetry of soldiers, it would be interesting to test the fitness consequences of larger body size and fluctuating asymmetry in soldiers of N. chilensis and its consequences for colony life. Additionally, the effects could be tested in other single-piece nesting termites such as the worldwide distributed (except in Asia) Cryptotermes brevis (Walker, 1853) and in Z. nevadensis, where multicolonial substrates have been reported.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
DA-O, LF-P, BT-G, and HN: study conception and design. BT-G and DA-O: material preparation and data collection. AA, BT-G, and DA-O: statistical analyses. DA-O: writing first draft of the manuscript. All authors commented on previous versions of the manuscript and read and approved the final manuscript.
Funding
This work was financially supported by FONDECYT grant 1120210 to HN. DA-O was grateful for FONDECYT postdoctoral project 3190381. AA was supported by the São Paulo Research Foundation (FAPESP), grant #2018/22839-6.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the two reviewers for helpful comments and suggestions to improve the previous version of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.882357/full#supplementary-material
References
Abe, T. (1991). Ecological factors associated with the evolution of worker and soldier castes in termites. Ann. Entomol. 9, 101–107.
Adams, E. S., and Tschinkel, W. R. (2001). Mechanisms of population regulation in the fire ant Solenopsis invicta: an experimental study. J. Anim. Ecol. 70, 355–369. doi: 10.1046/j.1365-2656.2001.00501.x
Aguilera-Olivares, D., Burgos-Lefimil, C., Melendez, W., Flores-Prado, L., and Niemeyer, H. M. (2016a). Chemical basis of nestmate recognition in a defense context in a one-piece nesting termite. Chemoecology 26, 163–172. doi: 10.1007/s00049-016-0217-x
Aguilera-Olivares, D., Rizo, J. F., Burgos-Lefimil, C., Flores-Prado, L., and Niemeyer, H. M. (2016b). Nestmate recognition in defense against nest invasion by conspecifics during swarming in a one-piece nesting termite. Rev. Chil. Hist. Nat. 89:11. doi: 10.1186/s40693-016-0063-9
Aguilera-Olivares, D., Flores-Prado, L., Véliz, D., and Niemeyer, H. M. (2015). Mechanisms of inbreeding avoidance in the one-piece drywood termite Neotermes chilensis. Insectes Soc. 62, 237–245. doi: 10.1007/s00040-015-0399-1
Aguilera-Olivares, D., Palma-Onetto, V., Flores-Prado, L., and Niemeyer, H. M. (2017). X-ray computed tomography reveals that intraspecific competition promotes soldier differentiation in a one-piece nesting termite. Entomol. Exp. Appl. 163, 26–34. doi: 10.1111/eea.12557
Amarillo-Suárez, A. R., Stillwell, R. C., and Fox, C. W. (2011). Natural selection on body size is mediated by multiple interacting factors: a comparison of beetle populations varying naturally and experimentally in body size. Ecol. Evol. 1, 1–14. doi: 10.1002/ece3.1
Beasley, D. A. E., Bonisoli-Alquati, A., and Mousseau, T. A. (2013). The use of fluctuating asymmetry as a measure of environmentally induced developmental instability: a meta-analysis. Ecol. Indic. 30, 218–226. doi: 10.1016/j.ecolind.2013.02.024
Bespalova, I., and Helms, K. R. (2014). Social selection and the evolution of a female weapon in queens of the ant Messor pergandei (Hymenoptera: Formicidae). Biol. J. Linn. Soc. 113, 1011–1020. doi: 10.1111/bij.12376
Blight, O., Díaz-Mariblanca, A. G., Cerdá, X., and Boulay, R. (2016). A proactive-reactive syndrome affects group success in an ant species. Behav. Ecol. 27, 118–125. doi: 10.1093/beheco/arv127
Bochdanovits, Z., and De Jong, G. (2003). Temperature dependent larval resource allocation shaping adult body size in Drosophila melanogaster. J. Evol. Biol. 16, 1159–1167. doi: 10.1046/j.1420-9101.2003.00621.x
Boulay, R., Cerdá, X., Simon, T., Roldan, M., and Hefetz, A. (2007). Intraspecific competition in the ant Camponotus cruentatus: should we expect the “dear enemy” effect? Anim. Behav. 74, 985–993. doi: 10.1016/j.anbehav.2007.02.013
Bourguignon, T., Leponce, M., and Roisin, Y. (2011). Are the spatio-temporal dynamics of soil-feeding termite colonies shaped by intra-specific competition? Ecol. Entomol. 36, 776–785. doi: 10.1111/j.1365-2311.2011.01328.x
Chouvenc, T., Basille, M., Li, H., and Su, N.-Y. (2014a). Developmental instability in incipient colonies of social insects. PLoS One 9:e113949. doi: 10.1371/journal.pone.0113949
Chouvenc, T., Basille, M., and Su, N.-Y. (2015). The production of soldiers and the maintenance of caste proportions delay the growth of termite incipient colonies. Insect. Soc. 62, 23–29. doi: 10.1007/s00040-014-0369-z
Chouvenc, T., Basille, M., and Su, N.-Y. (2017). Role of accelerated developmental pathway and limited nurturing capacity on soldier developmental instability in subterranean termite incipient colonies. Insect. Soc. 64, 477–483. doi: 10.1007/s00040-017-0566-7
Chouvenc, T., and Su, N.-Y. (2014b). Colony age-dependent pathway in caste development of Coptotermes formosanus Shiraki. Insect. Soc. 61, 171–182. doi: 10.1007/s00040-014-0343-9
Clarke, G. M., and McKenzie, L. J. (1992). Fluctuating asymmetry as a quality control indicator for insect mass rearing processes. J. Econ. Entomol. 85, 2045–2050. doi: 10.1093/jee/85.6.2045
Couret, J., and Benedict, M. Q. (2014). A meta-analysis of the factors influencing development rate variation in Aedes aegypti (Diptera: Culicidae). BMC Ecol. 14:3. doi: 10.1186/1472-6785-14-3
Cronin, A. L., Fédérici, P., Doums, C., and Monnin, T. (2012). The influence of intraspecific competition on resource allocation during dependent colony foundation in a social insect. Oecologia 168, 361–369. doi: 10.1007/s00442-011-2098-6
Cushman, J. H., Lawton, J. H., and Manly, B. F. J. (1993). Latitudinal patterns in European ant assemblages: variation in species richness and body size. Oecologia 95, 30–37. doi: 10.1007/BF00649503
Deheer, C. J., and Vargo, E. L. (2004). Colony genetic organization and colony fusion in the termite Reticulitermes flavipes as revealed by foraging patterns over time and space. Mol. Ecol. 13, 431–441. doi: 10.1046/j.1365-294X.2003.2065.x
Eggleton, P. (2011). “An introduction to termites: biology, taxonomy and functional morphology,” in Biology of Termites: a Modern Synthesis, eds D. E. Bignell, Y. Roisin, and N. Lo (Netherlands: Springer), 1–26.
Eyer, P. A., McDowell, B., Johnson, L. N. L., Calcaterra, L. A., Fernandez, M. B., Shoemaker, D., et al. (2018). Supercolonial structure of invasive populations of the tawny crazy ant Nylanderia fulva in the US. BMC Evol. Biol. 18:209. doi: 10.1186/s12862-018-1336-5
Gajardo, R. (1994). La Vegetación Natural de Chile. Clasificación y Distribución. Santiago: Editorial Universitaria.
Gibbs, M., and Breuker, C. (2006). Effect of larval-rearing density on adult life-history traits and developmental stability of the dorsal eyespot pattern in the speckled wood butterfly, Pararge aegeria. Entomol. Exp. Appl. 118, 41–47. doi: 10.1111/j.1570-7458.2006.00361.x
Graham, J. H., and Felley, J. D. (1985). Genomic coadaptation and developmental stability within introgressed populations of Enneacanthus gloriosus and E. obesus (Pisces. Centrarchidae). Evolution 39:104. doi: 10.2307/2408520
Graham, J. H., Raz, S., Hel-Or, H., and Nevo, E. (2010). Fluctuating asymmetry: methods, theory, and applications. Symmetry 2, 466–540. doi: 10.3390/sym2020466
Grohmann, C., Oldeland, J., Stoyan, D., and Linsenmair, K. E. (2010). Multi-scale pattern analysis of a mound-building termite species. Insect. Soc. 57, 477–486. doi: 10.1007/s00040010-0107-0
Handy, S. M., McBreen, K., and Cruzan, M. B. (2004). Patterns of fitness and fluctuating asymmetry across a broad hybrid zone. Int. J. Plant Sci. 165, 973–981. doi: 10.1086/423872
Hartig, F. (2021). DHARMa | Diagnostics for HierArchical Regession Models. R Package Version 0.4.4. http://florianhartig.github.io/DHARMa/
Haverty, M. I. (1977). The proportion of soldiers in termite colonies: a list and a bibliography (Isoptera). Sociobiology 2, 199–216.
Heinrich, B. (1993). “Beetles large and small”, in The Hot-Blooded Insects, (Heidelberg: Springer Berlin Heidelberg), 191–226.
Heinrich, B., and Bartholomew, G. A. (1979). Roles of endothermy and size in inter- and intraspecific competition for elephant dung in an african dung beetle, Scarabaeus laevistriatus. Physiol. Zool. 52, 484–496. doi: 10.1086/physzool.52.4.30155939
Henderson, G. (1998). “Primer pheromones and possible soldier caste influence on the evolution of sociality in lower termites,” in Pheromone Communication in Social Insects: Ants, Wasps, Bees, and Termites, eds R. K. Vander Meer, M. D. Breed, M. L. Winston, and K. E. Espelie (Boulder, CO: Westview Press), 314–330.
Hoiss, B., Krauss, J., Potts, S. G., Roberts, S., and Steffan-Dewenter, I. (2012). Altitude acts as an environmental filter on phylogenetic composition, traits and diversity in bee communities. Proc. R Soc. B Biol. Sci. 279, 4447–4456. doi: 10.1098/rspb.2012.1581
Holland, J. B., Nakayama, S., Porfiri, M., Nov, O., and Bloch, G. (2021). Body size and behavioural plasticity interact to influence the performance of free-foraging bumble bee colonies. Insects 12:236. doi: 10.3390/insects12030236
Holway, D. A., Suarez, A. V., and Case, T. J. (1998). Loss of intraspecific aggression in the success of a widespread invasive social insect. Science 282, 949–952. doi: 10.1126/science.282.5390.949
Hosken, D. J., Blanckenhorn, W. U., and Ward, P. I. (2000). Developmental stability in yellow dung flies (Scathophaga stercoraria): fluctuating asymmetry, heterozygosity and environmental stress. J. Evol. Biol. 13, 919–926. doi: 10.1046/j.1420-9101.2000.00239.x
Hunt, J., and Allen, G. R. (2000). Larval density and developmental instability in the acoustically orienting parasitoid Homotrixa alleni. Acta Ethol. 2, 129–132. doi: 10.1007/s102110000018
Johnson, S. E., Breisch, N. L., Momen, B., and Thorne, B. L. (2011). Morphology and gonad development of normal soldiers and reproductive soldiers of the termite Zootermopsis nevadensis nevadensis (Isoptera, Archotermopsidae). Zookeys 148, 15–30. doi: 10.3897/zookeys.148.1672
Korallo-Vinarskaya, N. P., Vinarski, M. V., Khokhlova, I. S., Shenbrot, G. I., and Krasnov, B. R. (2015). Intraspecific variation of body size in a gamasid mite Laelaps clethrionomydis: environment, geography and host dependence. Parasitol. Res. 114, 3767–3774. doi: 10.1007/s00436-015-4606-9
Korb, J., and Hartfelder, K. (2008). Life history and development - A framework for understanding developmental plasticity in lower termites. Biol. Rev. 83, 295–313. doi: 10.1111/j.1469-185X.2008.00044.x
Korb, J., and Linsenmair, K. E. (2001). The causes of spatial patterning of mounds of a fungus-cultivating termite: results from nearest-neighbour analysis and ecological studies. Oecologia 127, 324–333. doi: 10.1007/s004420000597
Le, S., Josse, J., and Husson, F. (2008). FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 25, 1–18. doi: 10.18637/jss.v025.i01
Lee, S.-B., Mullins, A., Aguilera-Olivares, D., Chouvenc, T., and Su, N.-Y. (2019). Fused colonies of the Formosan Subterranean Termite (Blattodea: Rhinotermitidae) for laboratory experiments. J. Econ. Entomol. 112, 2311–2315. doi: 10.1093/jee/toz154
Lloyd, M. M., and Poulin, R. (2014). Geographic variation in caste ratio of trematode colonies with a division of labour reflect local adaptation. Parasitol. Res. 113, 2593–2602. doi: 10.1007/s00436-014-3913-x
Markow, T. A. (1995). Evolutionary ecology and developmental instability. Annu. Rev. Entomol. 40, 105–120. doi: 10.1146/annurev.en.40.010195.000541
Matsuura, K. (2006). A novel hypothesis for the origin of the sexual division of labor in termites: which sex should be soldiers? Evol. Ecol. 20, 565–574. doi: 10.1007/s10682-006-9117-9
Micro-shot Technology Co (2010). M-shot Digital Imaging System. Guangzhou: Micro-shot Technology Co., Ltd.
Miura, T., and Scharf, M. E. (2011). “Molecular basis underlying caste differentiation in termites,” in Biology of termites: a modern synthesis, eds D. E. Bignell, Y. Roisin, and N. Lo (Netherlands: Springer), 211–253.
Mizumoto, N., and Bourguignon, T. (2021). The evolution of body size in termites. Proc. R Soc. B 288:20211458. doi: 10.1098/rspb.2021.1458
Muller, H., and Korb, J. (2008). Male or female soldiers? An evaluation of several factors which may influence soldier sex ratio in lower termites. Insectes Soc. 55, 213–219. doi: 10.1007/s00040-008-0996-3
Noirot, C. H. (1989). Social structure in termite societies. Ethol. Ecol. Evol. 1, 1–17. doi: 10.1080/08927014.1989.9525528
Ogino, K., Hirono, Y., Matsumoto, T., and Ishikawa, H. (1993). Juvenile hormone analogue, S-31183, causes a high level induction of presoldier differentiation in the japanese damp-wood termite. Zoolog. Sci. 10, 361–366.
Otronen, M. (1988). The effect of body size on the outcome of fights in burying beetles (Nicrophorus). Ann. Zool. Fennici 25, 191–201. doi: 10.1086/687392
Palmer, A. R. (1994). “Fluctuating asymmetry analyses: a primer,” in Developmental Instability: Its Origins and Evolutionary Implications, ed. T. A. Markow (Dordrecht: Kluwer Academic Publishers), 335–364.
Passera, L., Roncin, E., Kaufmann, B., and Keller, L. (1996). Increased soldier production in ant colonies exposed to intraspecific competition. Nature 379, 630–631. doi: 10.1038/379630a0
Perdereau, E., Bagnères, A.-G., Dupont, S., and Dedeine, F. (2010). High occurrence of colony fusion in a European population of the American termite Reticulitermes flavipes. Insectes Soc. 57, 393–402. doi: 10.1007/s00040-010-0096-z
Perdereau, E., Dedeine, F., Christidès, J. P., Dupont, S., and Bagnères, A.-G. (2011). Competition between invasive and indigenous species: an insular case study of subterranean termites. Biol. Invasions 13, 1457–1470. doi: 10.1007/s10530-010-9906-5
Price, P. W., Denno, R. F., Eubanks, M. D., Finke, D. L., and Kaplan, I. (2011). Insect Ecology: Behavior, Populations and Communities. Cambridge: Cambridge University Press.
Pringle, R. M., and Tarnita, C. E. (2017). Spatial self-organization of ecosystems: integrating multiple mechanisms of regular-pattern formation. Annu. Rev. Entomol. 62, 359–377. doi: 10.1146/annurev-ento-031616-035413
R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Ripa, R., and Luppichini, P. (2004). “Especies de termitas de importancia económica presentes en Chile,” in Termitas y otros insectos xilófagos en Chile: especies, biología y manejo, eds R. Ripa and P. Luppichini (Quillota: Instituto de Investigaciones Agropecuarias), 73–105.
Roisin, Y., and Korb, J. (2011). “Social organisation and the status of workers in termites,” in Biology of Termites: a Modern Synthesis, eds D. E. Bignell, Y. Roisin, and N. Lo (Dordrecht: Springer), 133–164. doi: 10.1111/brv.12330
Sanada-Morimura, S., Satoh, T., and Obara, Y. (2006). Territorial behavior and temperature preference for nesting sites in a pavement ant Tetramorium tsushimae. Insectes Soc. 53, 141–148. doi: 10.1007/s00040-005-0849-2
Scharf, M. E., Buckspan, C. E., Grzymala, T. L., and Zhou, X. (2007). Regulation of polyphenic caste differentiation in the termite Reticulitermes flavipes by interaction of intrinsic and extrinsic factors. J. Exp. Biol. 210, 4390–4398. doi: 10.1242/jeb.010876
Scholtz, O. I., Macleod, N., and Eggleton, P. (2008). Termite soldier defence strategies: a reassessment of Prestwich’s classification and an examination of the evolution of defence morphology using extended eigenshape analyses of head morphology. Zool. J. Linn. Soc. 153, 631–650. doi: 10.1111/j.1096-3642.2008.00396.x
Shellman-Reeve, J. S. (1997). “The spectrum of eusociality in termites,” in The Evolution of Social Behavior in Insects and Arachnids, eds J. C. Choe and B. J. Crespi (Cambridge: Cambridge University Press), 52–93.
Shelton, T. G., and Grace, J. K. (1996). Review of agonistic behaviors in the Isoptera. Sociobiology 28, 155–176.
Smith, K. F., Stefaniak, L., Saito, Y., Gemmill, C. E. C., Cary, S. C., and Fidler, A. E. (2012). Increased inter-colony fusion rates are associated with reduced COI haplotype diversity in an invasive colonial ascidian Didemnum vexillum. PLoS One 7:e30473. doi: 10.1371/journal.pone.0030473
Smith, R. J., Hines, A., Richmond, S., Merrick, M., Drew, A., and Fargo, R. (2007). Altitudinal variation in body size and population density of Nicrophorus investigator (Coleoptera: Silphidae). Environ. Entomol. 29, 290–298. doi: 10.1093/ee/29.2.290
Swaddle, J. P., and Witter, M. S. (1994). Food, feathers and fluctuating asymmetries. Proc. R Soc. Lond. B 255, 147–152. doi: 10.1098/rspb.1994.0021
Thomas, M. L., Tsutsui, N. D., and Holway, D. A. (2005). Intraspecific competition influences the symmetry and intensity of aggression in the Argentine ant. Behav. Ecol. 16, 472–481. doi: 10.1093/beheco/ari014
Thorne, B. L. (1996). Termite Terminology. Sociobiology 28, 253–263. doi: 10.1111/j.1469-185x.2008.00044.x
Thorne, B. L., Breisch, N. L., and Muscedere, M. L. (2003). Evolution of eusociality and the soldier caste in termites: influence of intraspecific competition and accelerated inheritance. Proc. Natl. Acad. Sci. U. S. A. 100, 12808–12813. doi: 10.1073/pnas.2133530100
Tsuji, K. (2013). Kin selection, species richness and community. Biol. Lett. 9, 11–13. doi: 10.1098/rsbl.2013.0491
Warren, M., McGeoch, M. A., Nicolson, S. W., and Chown, S. L. (2006). Body size patterns in Drosophila inhabiting a mesocosm: interactive effects of spatial variation in temperature and abundance. Oecologia 149, 245–255. doi: 10.1007/s00442-006-0434-z
Wills, B. D., Moreau, C. S., Wray, B. D., Hoffmann, B. D., and Suarez, A. V. (2014). Body size variation and caste ratios in geographically distinct populations of the invasive big-headed ant, Pheidole megacephala (Hymenoptera: Formicidae). Biol. J. Linn. Soc. 113, 423–438. doi: 10.1111/bij.12386
Witter, M. S., and Swaddle, J. P. (1994). Fluctuating asymmetries, competition and dominance. Proc. R Soc. Lond. B 256, 299–303. doi: 10.1098/rspb.1994.0084
Zizka, G., Schneider, J. V., Schulte, K., and Novoa, P. (2013). Taxonomic revision of the Chilean Puya species (Puyoideae, Bromeliaceae), with special notes on the Puya alpestris-Puya berteroniana species complex. Brittonia 65, 387–407. doi: 10.1007/s12228-012-9290-9
Keywords: developmental instability, drywood termite, fluctuating asymmetry, Kalotermitidae, Neotermes chilensis, stress, Isoptera, Blattodea
Citation: Aguilera-Olivares D, Torres-Guzmán B, Arab A, Flores-Prado L and Niemeyer HM (2022) Body Size and Symmetry Properties of Termite Soldiers Under Two Intraspecific Competition Scenarios. Front. Ecol. Evol. 10:882357. doi: 10.3389/fevo.2022.882357
Received: 23 February 2022; Accepted: 18 May 2022;
Published: 01 July 2022.
Edited by:
David Andrew Gray, California State University, Los Angeles, United StatesReviewed by:
Thomas Chouvenc, University of Florida, United StatesJoël Meunier, UMR 7261 Institut de Recherche sur la Biologie de l’Insecte (IRBI), France
Copyright © 2022 Aguilera-Olivares, Torres-Guzmán, Arab, Flores-Prado and Niemeyer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel Aguilera-Olivares, ZGFndWlsZXJhQHVkZWMuY2w=; Hermann M. Niemeyer, bmllbWV5ZXJAYWJ1bGFmaWEuY2llbmNpYXMudWNoaWxlLmNs
†ORCID: Daniel Aguilera-Olivares, orcid.org/0000-0003-0776-0275; Alberto Arab, orcid.org/0000-0003-0009-6658; Luis Flores-Prado, orcid.org/0000-0002-3373-8154; Hermann M. Niemeyer, orcid.org/0000-0003-2273-1608