 Jiangwen Li
Jiangwen Li Bangyin He1
Bangyin He1- 1Shaanxi Key Laboratory of Chinese Jujube, College of Life Sciences, Yan’an University, Yan’an, China
- 2Key Laboratory of Grassland Resources of the Ministry of Education of China, Key Laboratory of Forage Cultivation, Processing and High Efficient Utilization of the Ministry of Agriculture and Rural Affairs of China, Key Laboratory of Inner Mongolia Autonomous Region for Grassland Management and Utilization, College of Grassland, Resources and Environment, Inner Mongolia Agricultural University, Hohhot, China
- 3The New Zealand Institute for Plant and Food Research Limited, Auckland, New Zealand
Excessive grazing causes a decrease in plant diversity of grassland and also leads to changes in the functional traits of grassland plants. Based on the relationship between plant diversity and the functional traits of constructive species, the patterns of change in plant diversity can be predicted based on change in plant functional traits under different stocking rates. For the present study, Stipa breviflora desert steppe in Inner Mongolia was studied to characterize the plant community and population characteristics and plant functional traits of S. breviflora in grazing areas with different stocking rates [without grazing, light grazing (LG, 0.93 sheep unit hm–2half yr–1), moderate grazing (MG, 1.82 sheep unit hm–2half yr–1), heavy grazing (HG, 2.71 sheep unit hm–2 half yr–1)]. The results showed that: (1) LG significantly weakened the competitive advantage of the constructive species (S. breviflora) (P < 0.05), while HG significantly strengthened its competitive advantage in the community (P < 0.05); (2) Changes in plant diversity were generally significantly related to changes in S. breviflora root traits. The competitive advantage of S. breviflora in the community and the change in root traits could be used to predict the change in plant diversity in the desert steppe under different stocking rates. This research can provide a theoretical basis for maintaining plant diversity and sustainability in the desert steppe.
Introduction
Long-term over grazing causes continued degradation and decrease in the productivity of grassland. In a desert steppe ecosystem with extremely low-productivity, plant diversity is a key indirect factor determining community productivity after grazing disturbance (Wu et al., 2019). For example, decrease in the plant diversity of Stipa breviflora desert steppe directly leads to a decline in community productivity (Zhang et al., 2018). Over grazing can significantly change species composition of plant communities and cause significant decreases in plant diversity (Herrero-Jáuregui and Oesterheld, 2018). The negative response of plant diversity to stocking rates is greater in arid grassland ecosystems with low productivity than in semi-humid and humid ecosystems. Moreover, drought intensifies the impact of grazing (Eldridge et al., 2016). For instance, Plant diversity under high stocking rates can be nearly 50% lower than under low stocking rates. Increasing intensity of grazing may result in a significant decrease in species richness (Fetzel et al., 2017). In water and nutrient-restricted environments, increasing grazing intensity tends to cause the death of plants, and ultimately reduces species richness, negative effects that have been widely found in grasslands (Li et al., 2016; Shen et al., 2022a). How to predict changes in plant diversity is a key focus of ecological research. Studies have shown that grazing can exhibit both positive (the intermediate disturbance hypothesis, Fox, 1979) and negative (grazing reversal hypothesis, Proulx and Mazumder, 1998) effects on species richness. Intermediate disturbance hypothesis can be used for predicting the relationships between grazing and plant diversity, however, this relation is not certain. Especially, the tolerance of plants might be altered by changing grazing degree in arid or semiarid conditions, leading to uncertainty of the predicted results of the plant diversity under different grazing degree (Milchunas et al., 1988). Change in plant diversity is not only affected by inter-species competition, but also by the characteristics of plants (plant functional traits). However, existing studies have mainly explained changes in plant diversity from the perspective of plant communities (Jäschke et al., 2020). Some studies analyzed the changes in plant functional diversity based on functional traits (Díaz et al., 2007), while few of them predicted the changes in plant diversity from the perspective of cluster and tiller.
Plant functional traits can be effective indicators reflecting the response of plants to biotic and abiotic stresses (Li et al., 2021). The differences in the sensitivities of plant leaves, roots, structure and other traits to grazing disturbance result in different phenotypic traits. Functional traits of plant individuals exhibit variable response patterns to different environmental disturbances (Luo et al., 2019). For instance, over grazing can lead to a significant degeneration in the root traits of grassland plants (Wei et al., 2019). Increasing stocking rate leads to decreases in traits such as plant leaf area and leaf dry weight (Wang et al., 2016), while plant individuals exhibit stuntedness (Li et al., 2014). Plant functional traits show a regular pattern of change under the effect of different stocking rates, and could be used as indicators reflecting alterations in the plants’ internal environment as well as useful indices linking environmental driving factors with plant diversity (Bruelheide et al., 2018). Hence, exploring the response of grassland plant diversity to grazing based on changes in functional traits might be an effective way to predict patterns of change in plant diversity in the desert steppe.
The desert steppe is arid and vegetation is sparse, which makes it extremely sensitive to the external environment and vulnerable to degradation (Ren et al., 2018). The ability to predict changes in plant diversity based on plant functional traits is of profound significance for maintaining species diversity in desert steppes (Jin et al., 2021). Based on the foregoing analysis, we hypothesized that the plant diversity of the desert steppe would increase under light grazing (LG), while it would decrease under heavy grazing (HG). In addition, changes in plant diversity might be closely related to the aboveground and underground traits of the constructive species. To verify the above hypotheses, a representative S. breviflora desert steppe was chosen to investigate functional traits of the constructive species (S. breviflora) and the population and community characteristics in grazing areas with different stocking rates. Based on the data collected, the relationship between plant functional traits, diversity and grazing under different stocking rates was analyzed to explore the variation in S. breviflora functional traits under the disturbance of different stocking rates and its effects on plant diversity of the whole community. The aim was to provide a new approach to predict patterns of change in plant diversity in desert steppe under long-term sheep grazing and thereby provide a theoretical basis for maintaining plant diversity and sustainability in the desert steppe.
Materials and methods
Study site
The study site is located in Siziwang Banner, Inner Mongolia Autonomous Region, China (41°47′17″ N, 111°53′46″ E; elevation: 1450 m). The area has a typical temperate continental monsoon climate, with a mean annual precipitation of 221.7 mm, a mean annual temperature of 3.7°C with a range from –22.07°C (January) to 26.23°C (July), an annual accumulated temperature (≥10°C) of 2200–2500°C, a mean annual sunshine duration of 3117.7 h and an average frost-free period of 108 d. Rainfall mainly occurs from June to September, and the lowest and highest temperatures occur in January and July, respectively. The studied S. breviflora desert steppe is dominated by S. breviflora, Artemisia frigida, and Cleistogenes songorica with Convolvulus ammannii, Heteropappus altaicu, Artemisia pectinata, and Kochia prostrate as co-dominants. The main soil type is light chestnut soil.
Experimental design
The area of the study site is about 50 hm2, and the vegetation and soil types are relatively homogenous. The long-term grazing study site was established in 2004. Twelve ∼4.2 hm2 paddocks were constructed using a completely randomized block design, with four stocking rate treatments: control (CK, without grazing), light grazing (LG, 0.91 sheep unit hm–2 half year–1), moderate grazing (MG, 1.82 sheep unit⋅hm–2⋅half year–1), and heavy grazing (HG, 2.71 sheep unit⋅hm–2⋅half year–1), and each treatment had three replicates. Stocking rates in this study were calculated using the methods described by Wang and Ma (1994) and Wei (2000), and the degree of MG in this study was closer to the actual grazing status. The grazing period was from June to December in every year. Before the start of grazing, ten movable enclosures with a size of 1.5 m × 1.5 m were set up in each grazing plot (moved once per year, and the location was not repeated), and a 1 m × 1 m sized quadrat was set in each enclosure.
Data collection
Indicators of community and population characteristics, including biomass, average height, total coverage, and density, were measured in the enclosed plot during the biomass peak period in August 2017. In addition, at least six well grown and disease-free S. breviflora plants were selected in the same enclosure, and the functional traits of these clusters and their tillers were measured. The determination method was based on the plant functional trait manuals compiled by Perez-Harguindeguy et al. (2016) and Wigley et al. (2020), and used indicators shown in Table 1.

Table 1. Plant community characteristics and Stipa breviflora functional traits measured.
Community survey data were used to calculate the summed dominance ratio (SDR), which reflects the dominance of the species in the community:
where C is the relative coverage of each species, F is their relative frequency, H is their relative height, D is their relative density, and W is their relative biomass. The species were ranked based on the SDR. One-way analysis of variance (ANOVA) was used to detect differences among grazing treatments in the SDR of S. breviflora and community species richness, and the least significant difference (LSD) method was used for the post hoc test. The community characteristics and S. breviflora functional traits that closely related to plant richness were screened by stepwise regression analysis, and the selected traits were again analyzed by one-way ANOVA. All analyses were performed using R 3.6.3.
Results
The response of plant diversity and summed dominance ratio to stocking rate
The results in Figure 1 showed that the SDR of S. breviflora was obviously higher than that of other species under all stocking rates. Most other species exhibited higher SDR under the LG treatment, and generally exhibited the lowest SDR under the HG treatment, or even disappeared in heavily grazed areas. That is, LG weakened the competitive advantage of S. breviflora in the community, while HG increased its competitiveness. In addition, the LG treatment areas had significantly higher species richness (P < 0.05), and species richness decreased significantly with increasing stocking rate (Figure 1A, P < 0.05). In general, relative to community species richness, the SDR of S. breviflora exhibited the opposite trend (Figure 1B). In addition, there was significant negative relationship between species richness and SDR of S. breviflora (Figure 2).
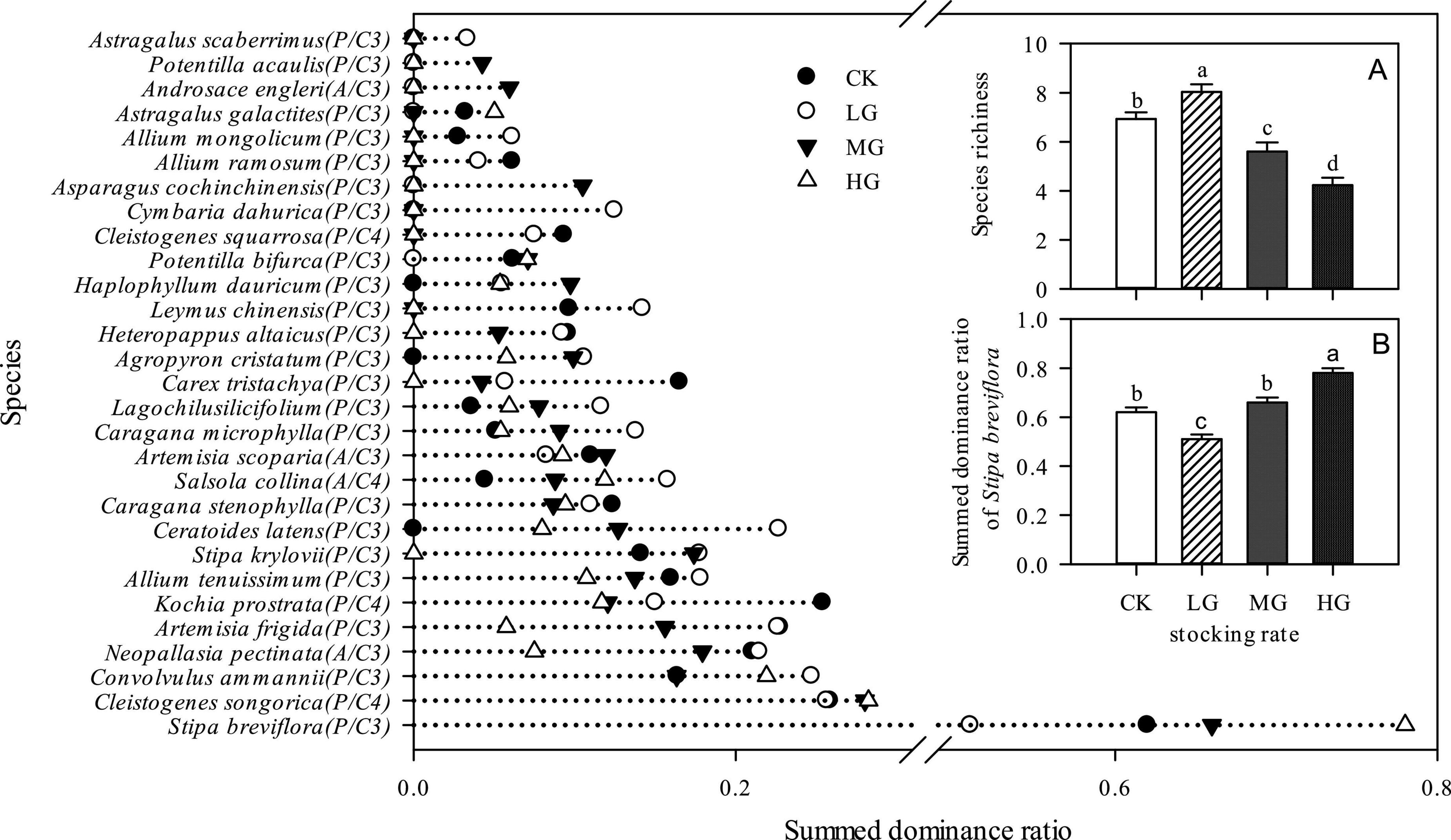
Figure 1. Ranking of species by summed dominance ratio (SDR), species richness (A) and SDR of Stipa breviflora (B) in areas with different stocking rates. Different letters above bars in panels (A,B) indicate significant differences among treatments (P < 0.05). P represents perennial species, A represents annual species, and C3/C4 represent C3 or C4 plants.
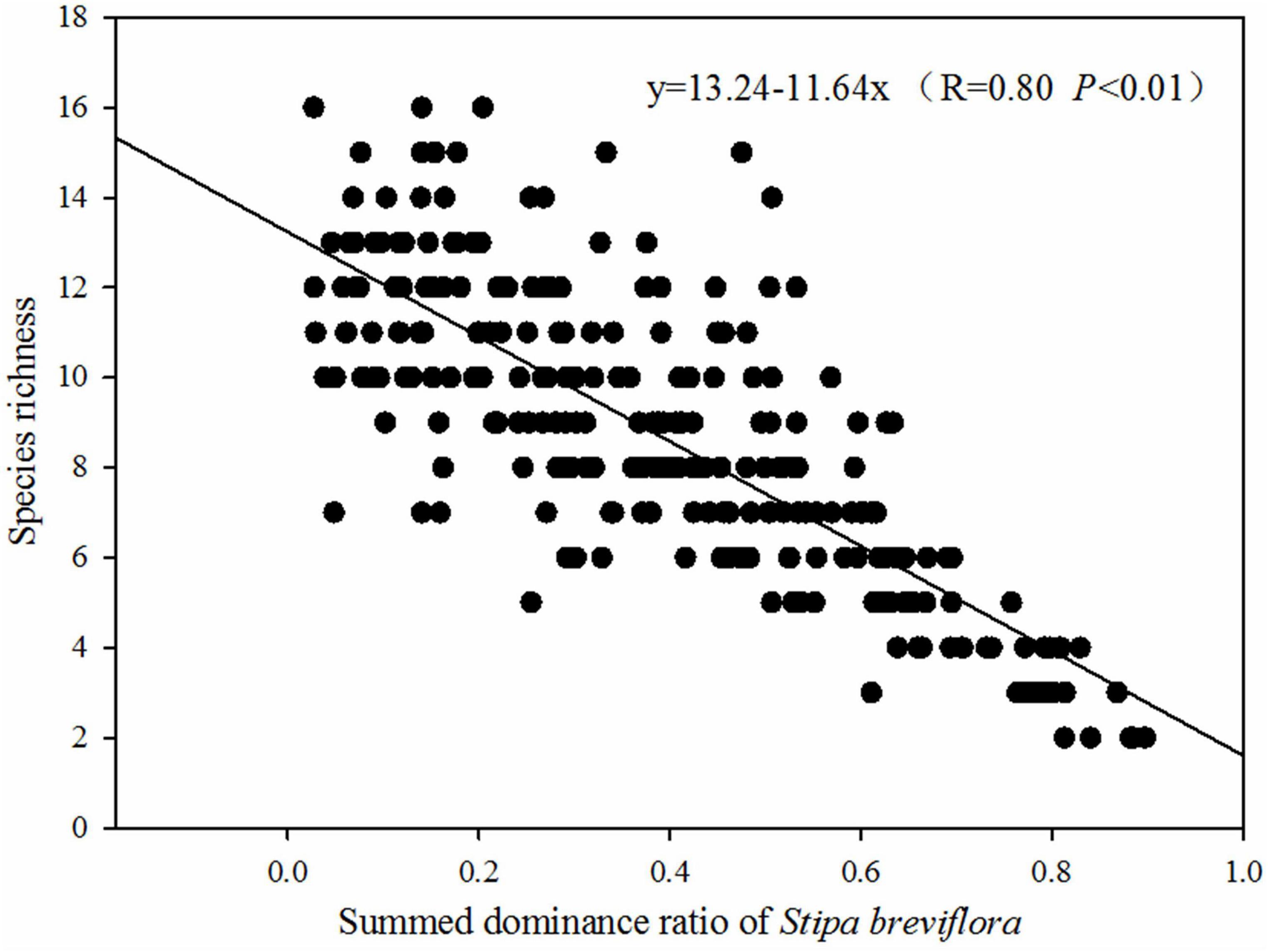
Figure 2. Regression fitting analysis of species richness and summed dominance ratio (SDR) of Stipa breviflora.
Correlation among plant diversity and functional traits
Ten S. breviflora functional traits and community characteristics that closely related to plant diversity were screened by stepwise regression analysis (Table 2). Seven of them were related to S. breviflora root traits (i.e., root surface area of cluster, cluster diameter of stem, root-leaf ratio of tiller, cluster root depth, tiller specific root length, root-shoot ratio of cluster, and tiller root tip number), and the remaining indicators were related to plant leaf traits (i.e., leaf number of tiller), and community and population characteristics (i.e., aboveground net primary productivity and SDR). Among these indicators, the SDR of species, cluster root depth, and the tiller root tip number were significantly and negatively correlated with plant diversity, and the other indicators were all positively correlated with plant diversity. In general, the root traits of S. breviflora exhibited the closest correlations with the plant diversity of the community.
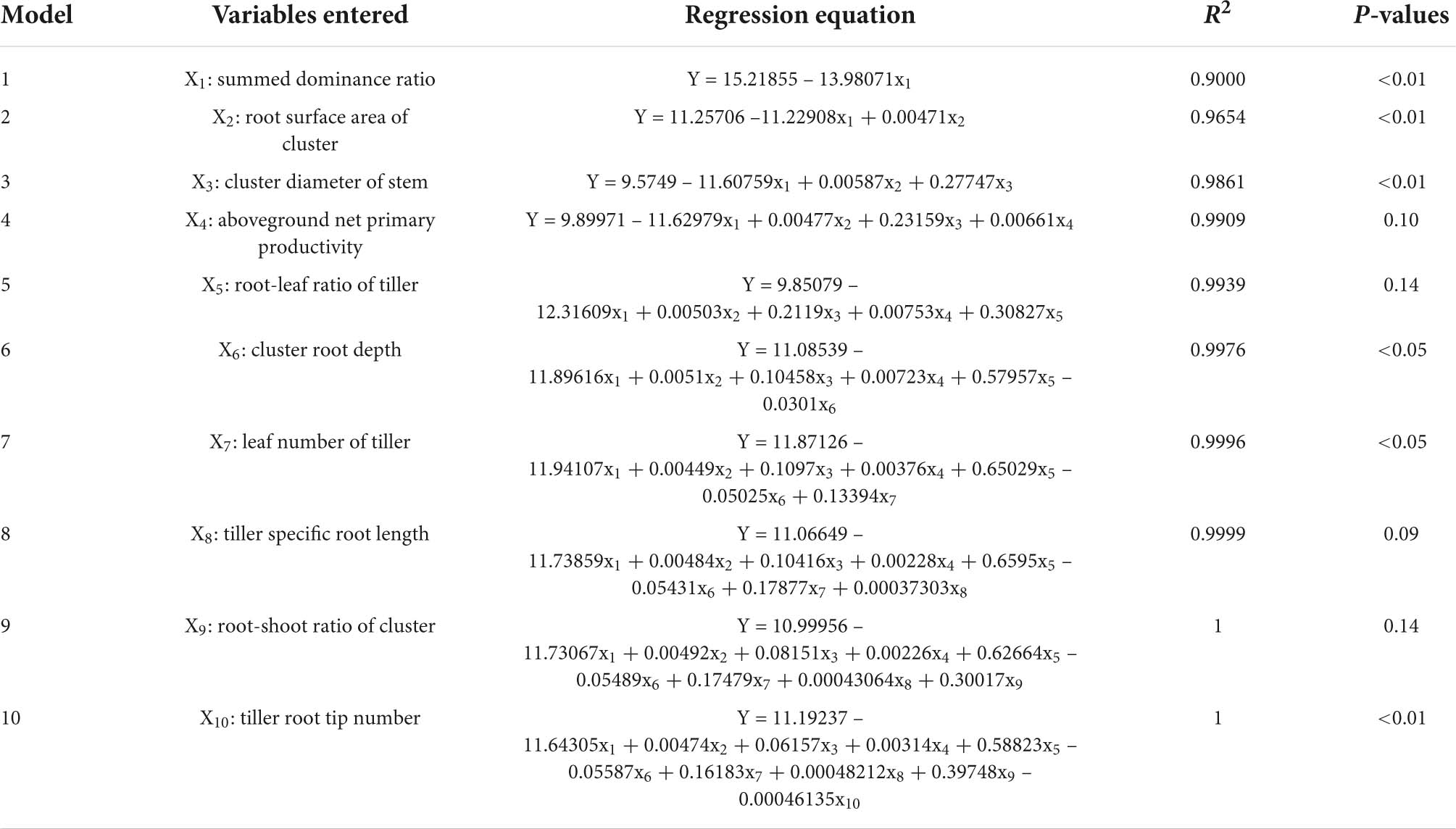
Table 2. Results of stepwise regression screening of plant diversity (Y) and Stipa breviflora functional traits.
Response of plant functional traits to stocking rate
One-way ANOVA for the ten mentioned functional traits of S. breviflora (Figure 3, except for the SDR) indicated that the root surface area of clusters decreased significantly with increasing grazing intensity (P < 0.05). LG and HG treatments significantly increased stem cluster diameter of S. breviflora, while MG and HG treatments significantly decreased aboveground net primary productivity (P < 0.05). HG treatment significantly increased the root-leaf ratio of tiller and root-shoot ratio of clusters of S. breviflora (P < 0.05), and all treatments increased its tiller specific root length and tiller root tip number (P < 0.05). In addition, only the LG treatment significantly decreased cluster root depth of S. breviflora, while both LG and HG significantly decreased the leaf number of tillers (P < 0.05).
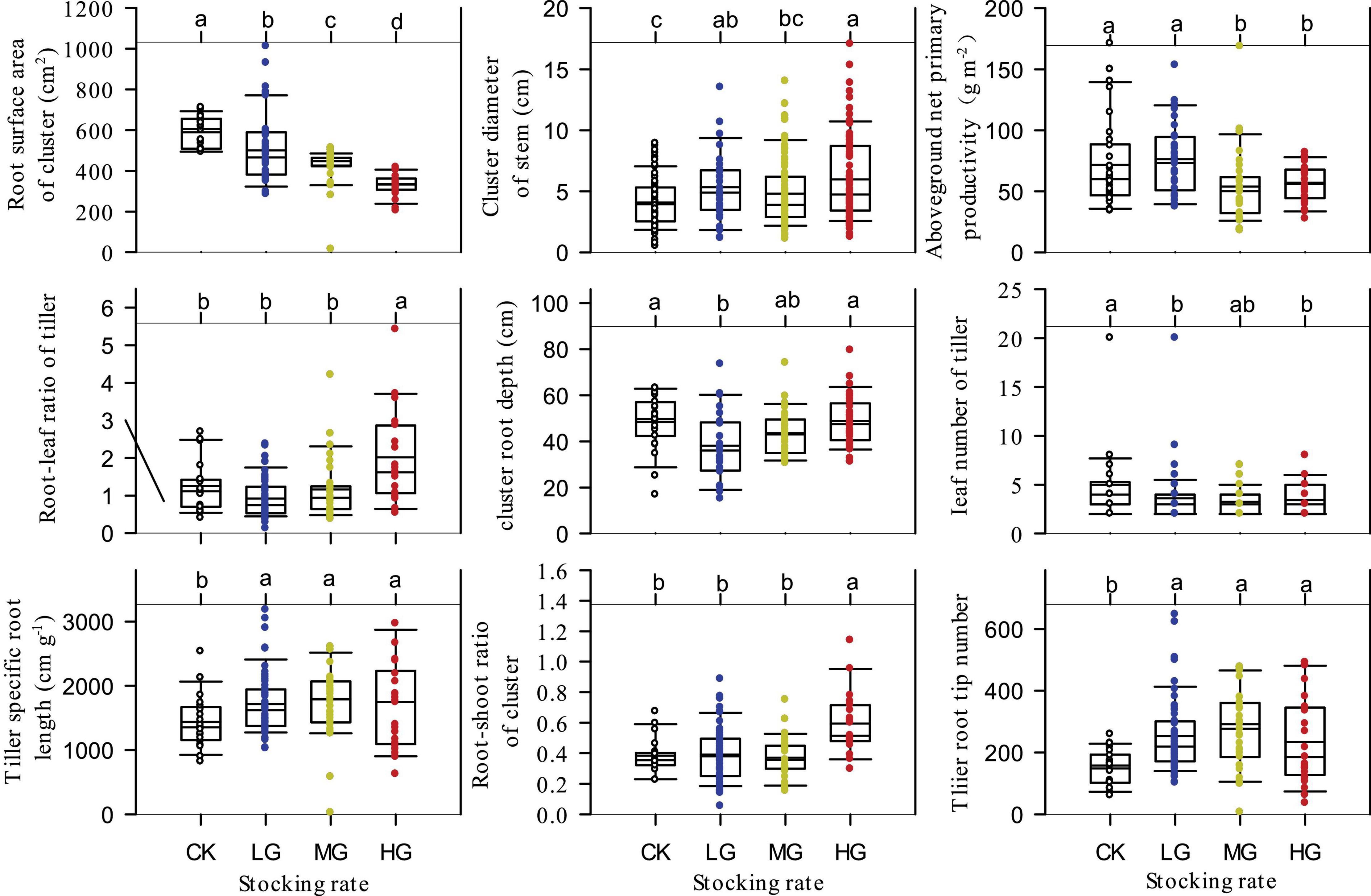
Figure 3. Analysis of variance (ANOVA) for the functional traits of Stipa breviflora under different stocking rates.
Discussion
Response of plant community composition to stocking rate
Grazing can cause changes in the species composition of plant communities due to the selective ingestion of non-gramineous plants with better palatability and nutritional value by livestock, which leads to decreases in the relative abundance of palatable species and increases in the proportion of unpalatable and grazing-resistant species (Liang et al., 2018). For instance, Liu et al. (2016) found that herbivores usually exhibit grazing preferences, and plant diversity has a significant negative correlation with herbivore preferences. Furthermore, different plant species or functional groups have their unique tolerance and escape strategies (Su et al., 2017). For example, perennial weeds are more sensitive to grazing, the average coverage of gramineous plants is sensitive to HG, and the height of perennial forage is sensitive to grazing intensity (Dorina et al., 2016). Grazing may lead to an increase or decrease in the proportion of the dominant species or functional groups, e.g., gramineous and non-gramineous species (Gornish and Ambrozio dos Santos, 2016). LG tends to maintain the relative coverage of erect and prostrate species, while moderate and HG lead to increases in the abundance of prostrate and non-gramineous species (Pizzio et al., 2016). Changes in the species composition of plant communities caused by grazing will directly result in changes in plant diversity. Hence, the biological and non-biological environment of new communities formed by the surviving species will be changed, and the biological environment formed by the surviving species will become one of the main factors affecting species diversity. Consequently, the effects of the functional traits of each species should be taken into consideration when investigating changes in species diversity.
The dominant position of Stipa breviflora in areas with different stocking rates
The results of the SDR analysis of S. breviflora showed that LG weakened the competitive advantage of S. breviflora in the community, and HG exhibited the opposite effect. This might be firstly attributed to the resistance to drought and grazing of S. breviflora (Zhang et al., 2010). For example, among the 9 plant functional traits of S. breviflora selected through stepwise regression in this study, root surface area of cluster, cluster root depth and leaf number of tillers of S. breviflora were significantly reduced under the LG treatment, while the cluster diameter of stem, root-leaf ratio of tillers, tiller specific root length, root-shoot ratio of cluster and tiller root tip number were significantly increased in HG areas (Yu et al., 2020). The weakening of the competitive advantage of S. breviflora in the community in LG treated areas might have resulted from decreases in these leaf traits, while the strengthening of its competitive advantage in HG treated areas might be caused by increases in its root traits. In addition, the HG treatment led to stuntedness of the plant individuals and the death of their tillers, which would result in the decreasing numbers of tillers and increases in the density of individuals, consequently causing an increase in the dominant position of S. breviflora (Wang et al., 2016; Song et al., 2021). HG led to a significant decrease in aboveground net primary productivity of the plant community, and indirectly provided more living space for S. breviflora. On the other hand, the non-dominant forages usually exhibit limited resistance to grazing and thus cannot adapt to the stress caused by HG. LG would firstly affect the growth of dominant species and thus provide more living space for non-dominant species, which also contributed to the changes in the competitive advantage of S. breviflora. In addition, grazing significantly altered the biotic (community) and abiotic (soil) environment (Jiang et al., 2020; Yin et al., 2020; Zhang et al., 2022), and the alterations caused by long-term grazing would lead to changes in plant diversity (Shinoda et al., 2021).
Notably, we found that all treatments did not totally alter the dominance of S. breviflora. Actually, the grazing intensity in the present study has reached the allowed lowest and highest boundaries of the stocking rates of the S. breviflora desert steppe, which indicated that this species indeed has more favorable adaptability than other species in this environment. Chen et al. (2002) reported that HG in early summer lead to a dominance of this C4 grass, such as Cleistogenes spp. However, similar trend was not observed in the present study. This might be caused by the significant increase in the SDR of S. breviflora, which is obviously higher than C. songorica. Hence, its increases in SDR might inhibit the photosynthesis, and consequently, the growth of C. songorica (Supplementary Figure 2). However, we did find that some shrub species, such as K. prostrata, exhibited an increasing trend. In addition, its SDR exhibited positive relationships with that of S. breviflora (Supplementary Figure 2). Hence, grazing might cause shrub-encroaching of community (Graw et al., 2016; Shen et al., 2022b). Certainly, this phenomenon should be studied in the following investigations.
Prediction of plant diversity changes based on Stipa breviflora root traits
The results of stepwise regression showed that ten functional straits of S. breviflora exhibited significant correlations with the plant diversity of the community, while seven of them reflected root traits. Our results indicated that among the selected seven traits, the root surface area of clusters decreased significantly with increasing grazing intensity, while the other six root traits (including cluster diameter of stem, root-leaf ratio of tiller, cluster root depth, tiller specific root length, root-shoot ratio of cluster, and tiller root tip number) increased significantly under the HG treatment. Previous studies have shown that grazing has inhibitory effects on roots. For instance, HG decreases the root biomass and belowground net primary productivity (Gao et al., 2008; Wei et al., 2019), which subsequently lead to a negative response of the root surface area of clusters to increasing stocking rate. However, other investigations have also found that long-term continuous grazing, especially HG, can disturb the functional balance between roots and aboveground plants parts (Wang et al., 2019), and lead to increases in plant root biomass and belowground net primary productivity (Gao et al., 2008). Root biomass is strongly affected by the diameter of stems and the root depth of clusters, and these indicators exhibited significant increases under the HG treatment. However, when the tillers were observed, it was found that a proportion of their root traits exhibited a positive response to HG treatment. For instance, the tiller root tip number gradually increased with increasing grazing intensity. This could be seen as a manifestation of the grazing avoidance mechanism of plants (Luo et al., 2019). The main function of the root tip of the plant is to absorb water, and an increase in its number can increase water use efficiency of the vegetation and the survival rate of plants. In arid areas, grazing will cause damage to the aboveground structure of individual plants, thus the water utilization efficiency of plants plays an even more important role in their growth and development. This might force the plants to allocate more resources to root growth, which could lead to deeper growth of roots. The tiller specific root length refers to the root length per unit weight of fine roots, which represents the relationship between root revenue and cost. When revenue exceeds cost, it is conducive to the accumulation of root biomass, so the reduction of specific root length is conducive to the accumulation of biomass. In the present study, all grazing treatments caused increases in the tiller specific root length of S. breviflora, which might be another factor causing the decrease in root biomass.
Although changes in the root biomass of constructive species caused by HG is still controversial, in the present study the allocation of aboveground and belowground biomass showed some regularities (Ma and Wang, 2020). For instance, HG led to increases in the root-shoot ratio of cluster and root-leaf ratio of tillers. Comprehensively considering all related indicators, grazing might cause gradual decreases in the aboveground and belowground biomass of S. breviflora. However, the aboveground part exhibited more obvious decreases. Furthermore, the aboveground part only exhibited decreases in the leaf number of tillers and aboveground net primary productivity, while the belowground root system was affected in several aspects, such as reductions in the surface area of clusters and increases in cluster root depth, cluster diameter of stem and the tiller root tip number. In general, we found that if (1) the aboveground net primary productivity of the community decreased and (2) the SDR of the constructive species increased, while its roots became thinner and deeper and with more root tips after grazing, then (3) the plant diversity of the community would exhibit a decreasing tendency.
This study mainly took desert steppe as an object, and predicting the plant diversity under different stocking rates based on functional traits of constructive species (S. breviflora). Moreover, the dominant species of grassland did not change at present. Hence, the change of plant diversity still needs long-term monitoring and verification in the future. At the same time, this study only addressed the regular of plant diversity change in desert steppe. To predict the regular of plant diversity changes in other various grassland types (e.g., typical steppe, meadow steppe, etc.) in northern China based on plant functional traits, it is still necessary to conduct long-term located monitoring experiments of different grassland types.
Conclusion
1) Light grazing weakened the competitive advantage of S. breviflora in the community, while HG strengthened its competitive advantage.
2) The competitive advantage of constructive species in the community and changes in its root traits could be used to predict the pattern of variation in plant diversity in a desert steppe under different stocking rates.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Author contributions
GH and JL designed the study and processed the data. All authors contributed to the results, related discussions, manuscript writing, contributed to the article, and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (31770500, 31760143), the Innovative Team of Grassland Resources from the Ministry of Education of China (IRT_17R59), the Natural Science Basic Research Plan in Shaanxi Province of China (2021JQ-623), and the Specialized Research Fund for the Doctoral Program of Yan’an University (YDBK2019-18).
Conflict of interest
SZ was employed by The New Zealand Institute for Plant and Food Research Limited.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This work was supported by Siziwang Banner Scientific Research Base, Comprehensive Experimental Demonstration Center, Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences. We thank Jie Qin, Mengqi Han, Yuxi Jin, Fengyuan Yu, and Jing Kang for their valuable comments on this manuscript.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.865703/full#supplementary-material
References
Bruelheide, H., Dengler, J., Purschke, O., Lenoir, J., Jiménez-Alfaro, B., Hennekens, S. M., et al. (2018). Global trait–environment relationships of plant communities. Nat. Ecol. Evol. 2, 1906–1917. doi: 10.1038/s41559-018-0699-8
Chen, L., Michalk, D. L., and Millar, G. D. (2002). The ecology and growth patterns of Cleistogenes species in degraded grasslands of eastern Inner Mongolia, China. J. Appl. Ecol. 39, 584–594. doi: 10.1046/j.1365-2664.2002.00735.x
Díaz, S., Lavorel, S., De Bello, F., Quétier, F., Grigulis, K., and Robson, T. M. (2007). Incorporating plant functional diversity effects in ecosystem service assessments. PNAS 104, 20684–20689. doi: 10.1073/pnas.0704716104
Dorina, Song, Y. T., Wu, Y. N., Huo, G. W., Wang, X. M., and Xu, Z. C. (2016). Response of plant leaf traits to grazing intensity in Stipa krylovii steppe. J. Appl. Ecol. 27, 2231–2238. doi: 10.13287/j.1001-9332.201607.034
Eldridge, D. J., Poore, A. G., Ruiz-Colmenero, M., Letnic, M., and Soliveres, S. (2016). Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecol. Appl. 26, 1273–1283. doi: 10.1890/15-1234
Fetzel, T., Havlik, P., Herrero, M., and Erb, K. H. (2017). Seasonality constraints to livestock grazing intensity. Global Change Biol. 23, 1636–1647. doi: 10.1111/gcb.13591
Fox, J. F. (1979). Intermediate-disturbance hypothesis. Science 204, 1344–1345. doi: 10.1126/science.204.4399.1344
Gao, Y. Z., Giese, M., Lin, S., Sattelmacher, B., Zhao, Y., and Brueck, H. (2008). Belowground net primary productivity and biomass allocation of a grassland in Inner Mongolia is affected by grazing intensity. Plant Soil 307, 41–50. doi: 10.1007/s11104-008-9579-3
Gornish, E. S., and Ambrozio dos Santos, P. (2016). Invasive species cover, soil type, and grazing interact to predict long-term grassland restoration success. Restor. Ecol. 24, 222–229. doi: 10.1111/rec.12308
Graw, V., Oldenburg, C., and Dubovyk, O. (2016). “Bush Encroachment Mapping for Africa: Multi-scale analysis with remote sensing and GIS,” in ZEF-Center for Development Research University of Bonn, Discussion Paper No 218, (Germany: Center for Development Research ZEF). doi: 10.2139/ssrn.2807811
Herrero-Jáuregui, C., and Oesterheld, M. (2018). Effects of grazing intensity on plant richness and diversity: a meta-analysis. Oikos 127, 757–766. doi: 10.1111/oik.04893
Jäschke, Y., Heberling, G., and Wesche, K. (2020). Environmental controls override grazing effects on plant functional traits in Tibetan rangelands. Funct. Ecol. 34, 747–760. doi: 10.1111/1365-2435.13492
Jiang, Z. Y., Hu, Z. M., Lai, D. Y., Han, D. R., Wang, M., Liu, M., et al. (2020). Light grazing facilitates carbon accumulation in subsoil in Chinese grasslands: A meta-analysis. Global Change Biol. 26, 7186–7197. doi: 10.1111/gcb.15326
Jin, C., Jiang, B., Ding, Y., Yang, S., Xu, Y., Jiao, J., et al. (2021). Functional traits change but species diversity is not influenced by edge effects in an urban forest of Eastern China. Urban For. Urban Green. 64:127245. doi: 10.1016/j.ufug.2021.127245
Li, J., Zheng, Z., Zhao, N., and Gao, Y. (2016). Relationship between ecosystem multifuntionality and species diversity in grassland ecosystems under land-use types of clipping, enclosure and grazing. Chin. J. Plant Ecol. 40, 735–747. doi: 10.17521/cjpe.2015.0457
Li, X., Hou, X., and Wu, X. (2014). Plastic responses of stem and leaf functional traits in Leymus chinensis to long-term grazing in a meadow steppe. Chin. J. Plant Ecol. 38, 440–451. doi: 10.3724/SP.J.1258.2014.00040
Li, X., Png, G. K., Li, Y., Jimoh, S. O., Ding, Y., Li, F., et al. (2021). Leaf plasticity contributes to plant anti-herbivore defenses and indicates selective foraging: Implications for sustainable grazing. Ecol. Indicat. 122:107273. doi: 10.1016/j.ecolind.2020.107273
Liang, M., Chen, J., Gornish, E. S., Bai, X., Li, Z., and Liang, C. (2018). Grazing effect on grasslands escalated by abnormal precipitations in Inner Mongolia. Ecol. Evol. 8, 8187–8196. doi: 10.1002/ece3.4331
Liu, W., Wei, Z., Lv, S., Sun, S., Jia, L., Zhang, S., et al. (2016). Response mechanism of plant diversity to herbivore foraging in desert grassland. Chin. J. Plant Ecol. 40, 564–573. doi: 10.1016/j.ecoleng.2016.06.106
Luo, W., Zuo, X., Griffin-Nolan, R. J., Xu, C., Ma, W., Song, L., et al. (2019). Long term experimental drought alters community plant trait variation, not trait means, across three semiarid grasslands. Plant Soil 442, 343–353. doi: 10.1007/s11104-019-04176-w
Ma, X., and Wang, X. (2020). Aboveground and belowground biomass and its’ allometry for Salsola passerina shrub in degraded steppe desert in Northwestern China. Land Degrad. Dev. 32, 714–722. doi: 10.1002/ldr.3772
Milchunas, D. G., Sala, O. E., and Lauenroth, W. K. (1988). A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Natural. 132, 87–106. doi: 10.1086/284839
Perez-Harguindeguy, N., Diaz, S., Garnier, E., Lavorel, S., Poorter, H., Jaureguiberry, P., et al. (2016). Corrigendum to: new handbook for standardised measurement of plant functional traits worldwide. Austr. J. Bot. 64, 715–716. doi: 10.1071/BT12225_CO
Pizzio, R., Herrero-Jáuregui, C., Pizzio, M., and Oesterheld, M. (2016). Impact of stocking rate on species diversity and composition of a subtropical grassland in Argentina. Appl. Veget. Sci. 19, 454–461. doi: 10.1111/avsc.12229
Proulx, M., and Mazumder, A. (1998). Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems. Ecology 79, 2581–2592. doi: 10.1890/0012-9658(1998)079[2581:ROGIOP]2.0.CO;2
Ren, H., Qin, J., and Yan, B. (2018). Mass loss and nutrient dynamics during litter decomposition in response to warming and nitrogen addition in a desert steppe. Front. Agric. Sci. Eng. 5, 64–70. doi: 10.15302/J-FASE-2017194
Shen, X., Liu, B., Henderson, M., Wang, L., Jiang, M., and Lu, X. (2022a). Vegetation greening, extended growing seasons, and temperature feedbacks in warming temperate grasslands of China. J. Climate 35, 1–51. doi: 10.1175/JCLI-D-21-0325.1
Shen, X., Liu, Y., Liu, B., Zhang, J., Wang, L., Lu, X., et al. (2022b). Effect of shrub encroachment on land surface temperature in semi-arid areas of temperate regions of the Northern Hemisphere. Agr. Forest Meteorol. 320:108943. doi: 10.1016/j.agrformet.2022.108943
Shinoda, Y., Uchida, K., Koyama, A., and Akasaka, M. (2021). Deer grazing changes seed traits and functions of grazing-intolerant plants. Plant Spec. Biol. 36, 126–132. doi: 10.1111/1442-1984.12300
Song, Y., Liu, T., Han, X., Lu, Y., Xu, X., Wang, L., et al. (2021). Adaptive traits of three dominant desert-steppe species under grazing-related degradation: Morphology, structure, and function. Global Ecol. Conserv. 28:e01647. doi: 10.1016/j.gecco.2021.e01647
Su, R., Cheng, J., Chen, D., Bai, Y., Jin, H., Chao, L., et al. (2017). Effects of grazing on spatiotemporal variations in community structure and ecosystem function on the grasslands of Inner Mongolia, China. Sci. Rep. 7:40. doi: 10.1038/s41598-017-00105-y
Wang, J., Zhong, M., Wu, R., Dong, Q., Wang, K., and Shao, X. (2016). Response of plant functional traits to grazing for three dominant species in alpine steppe habitat of the Qinghai-Tibet Plateau, China. Ecol. Res. 31, 515–524. doi: 10.1007/s11284-016-1360-0
Wang, M., and Ma, C. (1994). A Study on methods of estimating the carrying capacity of grassland. Grassland China 5, 19–22.
Wang, Z., Jin, J., Zhang, Y., Liu, X., Jin, Y., Wang, C., et al. (2019). Impacts of mixed-grazing on root biomass and belowground net primary production in a temperate desert steppe. Roy. Soc. Open Sci. 6:180890. doi: 10.1098/rsos.180890
Wei, X., Zhong, M., Liu, Y., Wu, R., and Shao, X. (2019). Covariation in root traits of Leymus chinensis in response to grazing in steppe rangeland. Rangeland J. 41, 313–322. doi: 10.1071/RJ18099
Wei, Z. (2000). The Response of Stipa breviflora community to stocking rate. Grassland China 6, 2–6.
Wigley, B., Charles-Dominique, T., Hempson, G., Stevens, N., Tebeest, M., Archibald, S., et al. (2020). A handbook for the standardised sampling of plant functional traits in disturbance-prone ecosystems, with a focus on open ecosystems. Austr. J. Bot. 68, 473–531. doi: 10.1071/BT20048
Wu, J., Li, M., Fiedler, S., Ma, W., Wang, X., Zhang, X., et al. (2019). Impacts of grazing exclusion on productivity partitioning along regional plant diversity and climatic gradients in Tibetan alpine grasslands. J. Environ. Manag. 231, 635–645. doi: 10.1016/j.jenvman.2018.10.097
Yin, M., Gao, X., Tenuta, M., Li, L., Gui, D., Li, X., et al. (2020). Enhancement of N2O emissions by grazing is related to soil physicochemical characteristics rather than nitrifier and denitrifier abundances in alpine grassland. Geoderma 375:114511. doi: 10.1016/j.geoderma.2020.114511
Yu, R., Zhang, W., Yu, Y., Yu, S., Lambers, H., and Li, L. (2020). Linking shifts in species composition induced by grazing with root traits for phosphorus acquisition in a typical steppe in Inner Mongolia. Sci. Total Environ. 712:136495. doi: 10.1016/j.scitotenv.2020.136495
Zhang, Q., Niu, J., Ding, Y., Kang, S., and Dong, J. (2010). Current research advances and future prospects of biology and ecology of Stipa breviflora. Chin. J. Grassland 32, 93–101.
Zhang, R., Wang, Z., Han, G., Schellenberg, M. P., Wu, Q., and Gu, C. (2018). Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the Desert Steppe, Northern China. Agric. Ecosys. Environ. 265, 73–83. doi: 10.1016/j.agee.2018.05.014
Keywords: plant functional traits, plant diversity, desert steppe, Stipa breviflora, grazing
Citation: Li J, He B, Zhou S, Zhang X, Li C and Han G (2022) Prediction of plant diversity under different stocking rates based on functional traits of constructive species in a desert steppe, northern China. Front. Ecol. Evol. 10:865703. doi: 10.3389/fevo.2022.865703
Received: 30 January 2022; Accepted: 01 July 2022;
Published: 25 July 2022.
Edited by:
Pavel Kindlmann, Charles University, CzechiaReviewed by:
Guogang Zhang, Tianjin Normal University, ChinaXiangjin Shen, Northeast Institute of Geography and Agroecology (CAS), China
Yang Yang, Northwest A&F University, China
Copyright © 2022 Li, He, Zhou, Zhang, Li and Han. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiangwen Li, bGlqaWFuZ3dlbkB5YXUuZWR1LmNu; Guodong Han, aGFuZ3VvZG9uZ0BpbWF1LmVkdS5jbg==