
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 12 April 2022
Sec. Conservation and Restoration Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.861611
This article is part of the Research Topic Conserving Cultural Ecosystems: The Biodiversity and Ecosystem Service Provision of Semi-Natural Anthropogenic Ecosystems versus Natural Ecosystems View all 7 articles
 José Manuel Fernández-Guisuraga*
José Manuel Fernández-Guisuraga* Víctor Fernández-García
Víctor Fernández-García Reyes Tárrega
Reyes Tárrega Elena Marcos
Elena Marcos Luz Valbuena
Luz Valbuena Rayo PintoPaula MonteDavid BeltránSara Huerta
Rayo PintoPaula MonteDavid BeltránSara Huerta Leonor Calvo
Leonor Calvo
Understanding the effects of traditional livestock grazing abandonment on the ability of mountain grasslands to sustain multiple ecosystem functions (ecosystem multifunctionality; EMF) is crucial for implementing policies that promote grasslands conservation and the delivery of multiple ecosystem services. In this study, we evaluated the effect of short- and long-term transhumant sheep abandonment on EMF through a grazing exclusion experiment in a grassland of the Cantabrian Mountains range (NW Spain), where transhumant sheep flocks graze in summer. We considered four key ecosystem functions, derived from vegetation and soil functional indicators measured in the field: (A) biodiversity function, evaluated from total plant species evenness, diversity and richness indicators; (B) forage production function, evaluated from cover and richness of perennial and annual herbaceous species indicators; (C) carbon sequestration function, evaluated from woody species cover and soil organic carbon indicators; and (D) soil fertility function, evaluated from NH4+-N, NO3–-N, P and K content in the soil. The EMF index was calculated by integrating the four standardized ecosystem functions through an averaging approach. Based on linear mixed modeling we found that grazing exclusion induced significant shifts in the considered individual ecosystem functions and also on EMF. Long-term livestock exclusion significantly hindered biodiversity and forage production functions, but enhanced the carbon sequestration function. Conversely, the soil fertility function was negatively affected by both short- and long-term grazing exclusion. Altogether, grazing exclusion significantly decreased overall EMF, especially in long-term livestock exclusion areas, while the decline in EMF in short-term exclusions with respect to grazed areas was marginally significant. The results of this study support the sustainability of traditional transhumance livestock grazing for promoting the conservation of grasslands and their ecosystem function in mountain regions.
Grasslands represent approximately one third to half of the terrestrial land surface and support about a quarter of the Earth’s human population (Zhang et al., 2018; Jarque-Bascuñana et al., 2022), most of whom are largely dependent on the services provided by these ecosystems for their subsistence (Evans et al., 2017), as in the case of mountain regions (García-Ruiz et al., 2020). Crucial ecosystem services provided by mountain grasslands include (i) regulating services such as carbon sequestration for global warming mitigation (McSherry and Ritchie, 2013) and pollination (Bendel et al., 2019), (ii) provisioning services like food production for humans and livestock (Gennet et al., 2017), and (iii) cultural services such as aesthetic values and recreation (Bi et al., 2018). In addition, natural and semi-natural mountain grasslands are recognized globally for their high biodiversity (Bengtsson et al., 2019), sustaining a high number of endemic and endangered species (Ingty, 2021). However, approximately half of the Earth’s grasslands are being affected by unsustainable livestock grazing practices leading to land degradation (Chen et al., 2014), or by land use changes and land abandonment (Jarque-Bascuñana et al., 2022).
Livestock grazing is the dominant land use in mountain grassland ecosystems (Alkemade et al., 2013), and has played a crucial role in people’s livelihoods for millennia (Bengtsson et al., 2019), creating complex socio-ecological systems (Ingty, 2021). In general, the negative effects of overgrazing on the vegetation structure and ecosystem functioning of mountain grasslands worldwide are well documented (e.g., Hilker et al., 2014; Dlamini et al., 2016; Hao et al., 2018), even in productive ecosystems (Cingolani et al., 2014). However, traditional extensive livestock grazing in mountain regions, with low to moderate stocking densities and short seasonal use of grasslands, has been reported to support several ecosystem services, such as habitat biodiversity (Odriozola et al., 2017), primary production (Jarque-Bascuñana et al., 2022) and the conservation of cultural ecosystems associated with the traditions of local communities relying on pastoralism for their livelihood (Öllerer et al., 2019). Notwithstanding, a progressive abandonment of extensive livestock in European countries has been evidenced over the last decades (Cocca et al., 2012). This process is particularly relevant in mountain regions as a consequence of the low profitability and economic competitiveness compared to intensive lowland systems (Lasanta et al., 2006). Furthermore, the removal of European subsidies for marginal grazing land (Aldezabal et al., 2015) and the promotion of more productive livestock holdings at the expense of traditional practices by the Common Agricultural Policy (CAP) of the European Union (García-Llamas et al., 2019), has led to a significant reduction in the surface occupied by grassland ecosystems in Europe (Pe’er et al., 2014). Particularly meaningful is the decline, in many European countries, of the traditional livestock management that involves short-term transhumance (Román-Trufero et al., 2019), i.e., seasonal movements of extensive sheep flocks between different regions with complementary pasture productivity throughout the seasons, exploiting the natural vegetation growth in summer mountain pastures (García-Llamas et al., 2019). The gradual abandonment of this farming practice is particularly noticeable in the mountainous regions of northern Spain (Cantabrian Mountains and Pyrenees) since the middle of the last century (Velado-Alonso and Gómez-Sal, 2016).
The decline of sheep transhumance systems (Blanco et al., 2019), such as those on the Cantabrian Mountains range (Calvo et al., 2002; Morán-Ordóñez et al., 2013), involves the removal of important grazing processes such as defoliation and fertilization, and, therefore, changes in the grassland ecosystem structure and function can be expected (Aldezabal et al., 2015). Hence, understanding the effects of traditional grazing abandonment on the ability of mountain grassland ecosystems to deliver multiple functions (ecosystem multifunctionality; EMF) is crucial for implementing sustainable policies that promote ecosystem conservation and the delivery of multiple ecosystem services (Ren et al., 2018). In this context, complex trade-offs between ecosystem functions and services can arise as a consequence of land use changes and management priorities (Bengtsson et al., 2019). Therefore, an integrated approach for predicting land use change effects in the overall ecosystem function (i.e., EMF) through standardized indices is needed to avoid a biased perception of the ecosystem response (Odriozola et al., 2014; Lucas-Borja et al., 2021). In addition, most terrestrial ecosystems are valued primarily for the simultaneous provision of several ecosystem functions or services, instead of single functions (Hector and Bagchi, 2007; Maestre et al., 2012a), and the magnitude of land use change effects on ecosystem response may be stronger when multifunctionality is considered (Byrnes et al., 2014). Moreover, if the preservation of extensive livestock grazing is perceived as a necessary strategy for enhancing the provision of multiple ecosystem services in mountain regions (Öllerer et al., 2019), it is imperative to understand how grazing abandonment affects EMF (Maestre et al., 2012b).
Several straightforward methods have been recently developed to quantify EMF using experimental data, including the averaging approach (Maestre et al., 2012b) and the threshold approach (Zavaleta et al., 2010). Nevertheless, the effect of traditional livestock abandonment on the EMF of productive grasslands in mountainous regions remains unexplored, with only one study carried out by Ren et al. (2018) that evaluated the effects of grazing pressure on EMF in semi-arid grasslands. Furthermore, several studies related to traditional pastoralism on productive mountain grasslands focused on single functions or services such as ecosystem productivity (Jarque-Bascuñana et al., 2022), biodiversity (Ingty, 2021), carbon storage (Lu et al., 2015) or soil fertility (Semmartin et al., 2004). Conversely, an EMF approach will provide more integrated insights into the sustainability of traditional livestock grazing in relation to the conservation of biodiversity and global ecosystem function in mountain grasslands, which is largely unresolved and it can be particularly important to guide policies aimed at maintaining the sustainability of these areas.
In this study, we evaluated the effect of short- and long-term transhumant sheep grazing abandonment on EMF through a grazing exclusion experiment in a mountainous grassland of the Cantabrian Mountains range (NW Spain). We considered EMF by integrating four key ecosystem functions and their standardized indicators: (A) biodiversity, (B) forage production, (C) carbon sequestration and (D) soil fertility. First, we expected a decrease in the biodiversity and forage production functions since strong competitive effects exerted by encroaching shrub species are anticipated in the absence of moderate grazing in productive grasslands (Komac et al., 2013; Odriozola et al., 2017; Bi et al., 2018). Second, we expected a positive to neutral effect of grazing abandonment on carbon sequestration function as a consequence of woody plant encroachment (Álvarez-Martínez et al., 2016), and the lack of a clear relationship between grazing exclusion and soil carbon response (McSherry and Ritchie, 2013). Third, we expected that soil fertility function would be hindered under grazing abandonment since livestock contributes to accelerate nutrient cycling in productive grassland ecosystems through the modulation of the soil physical characteristics (Odriozola et al., 2014) and litter decomposition dynamics (Semmartin et al., 2008), among other processes. In general, we hypothesized that livestock abandonment from productive grasslands with long evolutionary history of grazing at low to moderate stocking rates would impact EMF, although the magnitude of the net effect on the global ecosystem function must be resolved.
The study site was located within the Cantabrian Mountains, a mountain range which covers an area of about 31,500 km2 along the northern border of the Iberian Peninsula on the boundary between the Atlantic-Eurosiberian and Mediterranean biogeographic regions (Rivas-Martínez et al., 1987). The vegetation of the Cantabrian Mountains is mainly comprised of a mosaic of deciduous forests dominated by oak species [e.g., Quercus petraea (Matt.) Liebl., Q. robur L. and Q. pyrenaica Willd.], beech (Fagus sylvatica L.) and birch (Betula spp. L.), closed and open shrublands and semi-natural grasslands (García-Llamas et al., 2019) managed in extensive grazing systems of transhumant sheep flocks, as well as cattle and horses (Morán-Ordóñez et al., 2011). Transhumance sheep activity has played a key role in shaping a characteristic large-scale mosaic of open shrublands and grasslands with high cultural value (Morán-Ordóñez et al., 2013). However, the decline of the transhumant activity since the last century (Velado-Alonso and Gómez-Sal, 2016) has caused a large increase in the area covered by later successional state woody vegetation encroaching semi-natural grasslands (Morán-Ordóñez et al., 2013). In fact, state-and-transition models (Jackson and Bartolome, 2002) applied to these semi-natural grasslands predict that traditional grazing abandonment triggers an alternate stable state of shrub-encroached systems that cannot be reverted unless intensive management practices are implemented (Targetti et al., 2010).
A mountain pass in the western area of the Cantabrian Mountains known as Las Pintas (municipality of Crémenes, León province; Figure 1) was chosen as the experimental site. Las Pintas covers an area of 389 ha and is characterized by a rugged topography with altitudes ranging between 1,060–1,989 m above sea level (ASL), dominated by limestone and sandstone lithology (GEODE, 2020). The average annual temperature is 6.5°C and the average annual rainfall is 1,350 mm (Ninyerola et al., 2005). The predominant habitats in Las Pintas pass are rock outcrops, beech forests, creeping juniper shrublands [Juniperus communis L. subsp. alpina (Suter) Čelak.], gorse shrublands (Genista hispanica subsp. occidentalis Rouy) and grasslands dominated by species with high pastoral value such as Festuca rubra L., Agrostis capillaris L., Sanguisorba minor Scop. and Trifolium repens L. Las Pintas mountain pass resumed the transhumant livestock use in the summer grasslands from July to September in 2015, with moderate stocking rates (0.2–0.4 LSU/ha; LSU = livestock unit) per season (Sebastià et al., 2008). Extensive sheep grazing in the mountain pass was the dominant land use until the early 2000s, whereas no anthropic use was registered at any location in the pass between 2005 and 2015. Large wildlife herbivores present in the experimental site include Pyrenean chamois (Rupicapra pyrenaica) and European roe deer (Capreolus capreolus).
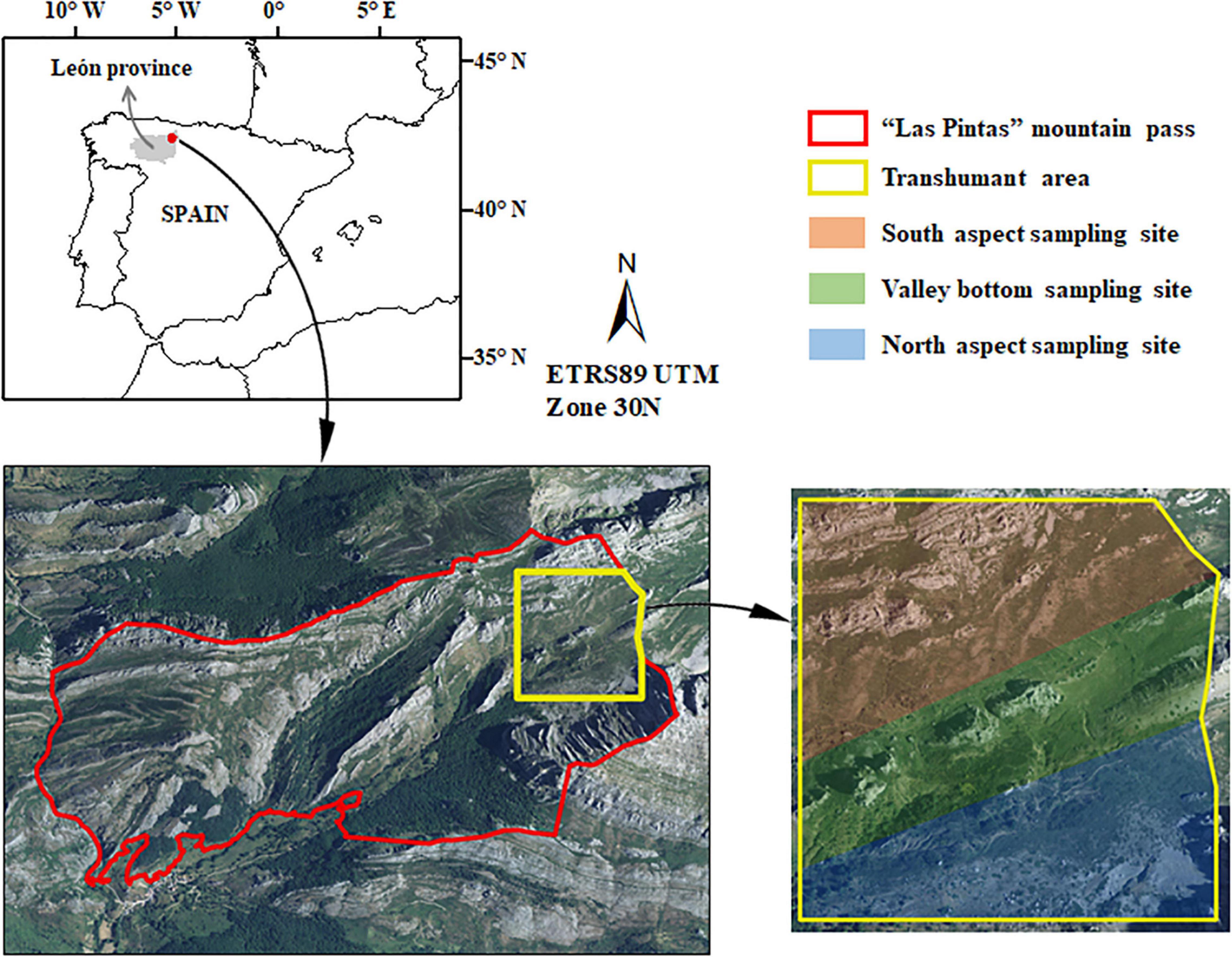
Figure 1. Location of Las Pintas mountain pass in León province (northwest Spain) and spatial distribution of the sampling sites within the transhumant area.
In June 2020, three sampling sites with different aspect were established in the grasslands within the eastern area of Las Pintas mountain pass, where transhumant sheep flocks concentrate: (i) flat valley area (12.9 ha), (ii) north aspect (13.8 ha) and (iii) south aspect (17.6 ha). The mean altitude of the three sites showed little variability (1,564–1,612 m ASL). Within each site, the experiment had a randomized complete block design, with five blocks of 50 m × 50 m per site. This experimental design was aimed at reducing the heterogeneity in abiotic environmental conditions (e.g., incident solar radiation or humidity) of the grasslands. The blocks were located within each sampling site to encompass both grazed and abandoned areas. Each block comprised three levels of grazing treatment (i.e., abandonment): (i) control, corresponding to current sheep grazing, (ii) short-term grazing exclusion −1 year-, and (iii) long-term grazing exclusion -between 15–20 years-. Grazing treatment had five replicates per sampling site, providing a total of 45 experimental quadrats or square plots (three sampling sites × three grazing treatment levels × five replicates), with a size of 1 m × 1 m. Grassland community composition was similar at each sampling site and grazing treatment areas (field observation). Short-term grazing exclusion plots, randomly located within each block, were fenced from June 2020 to June 2021 to prevent sheep access. Together with the fenced plots, we established within each block control grazed plots where sheep were allowed to graze continuously during the season (July to September), and long-term grazing exclusion plots in areas abandoned between 15–20 years based on interviews with local shepherds and city council technicians, as proposed by previous grazing exclusion research (De Bello et al., 2005; Sebastià et al., 2008; Aynekulu et al., 2017).
The vegetation survey was conducted in June 2021, during the peak aboveground biomass at the study site. The vascular plants present in each field plot were identified at the species level. Then, we measured species cover using a quadrat and a visual estimation method in steps of 5% (Anderson et al., 2005; Calvo et al., 2008). Plant species were classified into biotypes (woody species, and annual and perennial herbaceous species) for determining their relative cover and richness. We also calculated total species richness (S), species diversity (Shannon diversity index, H; Shannon and Weaver, 1949) and species evenness (Pielou evenness index, J; Pielou, 1966).
In addition, composite soil samples consisting of four soil samples (0–10 cm depth) were collected with a soil sampler tool in the cardinal points of each field plot for obtaining a representative sample (Fernández-García et al., 2021). Plant species, litter, and woody debris were removed before sampling collection. Soil samples were homogenized and sieved in the field (2 mm mesh) and separated into two fractions. One fraction was stored at 4°C in polyethylene bags and delivered to the laboratory, where it was frozen at −18°C until ammonium (NH4+-N) and nitrate (NO3–-N) content analyses. The second fraction was delivered to the laboratory, air dried for 1 week and stored in polyethylene bags until analysis of soil organic carbon (C), available phosphorus (P) and potassium (K). We determined organic C (%) by the Walkley-Black dichromate oxidation method (Nelson and Sommers, 1996). Available P (ppm) was determined through Olsen et al. (1954) method using 0.5 M NaHCO3 (pH: 8.5) extraction and determination at 882 nm on a UV Mini 1240 spectrophotometer (Shimadzu Corporation, Japan). We analyzed available K [cmol(+)/kg] using the 1 M NH4OAc (pH: 7) method (Warncke and Brown, 1998) and subsequent determination by an Agilent 5110 ICP-atomic emission spectrometer (Agilent Technologies, United States). NH4+-N (ppm) and NO3–-N (ppm) were determined using 2 M KCl extraction (Keeney and Nelson, 1982) and an automatic Kjeldahl analyzer (Büchi Labortechnik, Switzerland).
The vegetation and soil properties measured here were grouped into four ecosystem functions: (i) biodiversity, (ii) forage production, (iii) carbon sequestration and (iv) soil fertility (Table 1), all of which are relevant drivers of EMF in grassland ecosystems (Garland et al., 2021), and are consistent with previous studies (e.g., Zavaleta et al., 2010; Bradford et al., 2014; Jing et al., 2015; Valencia et al., 2015; Ren et al., 2018).
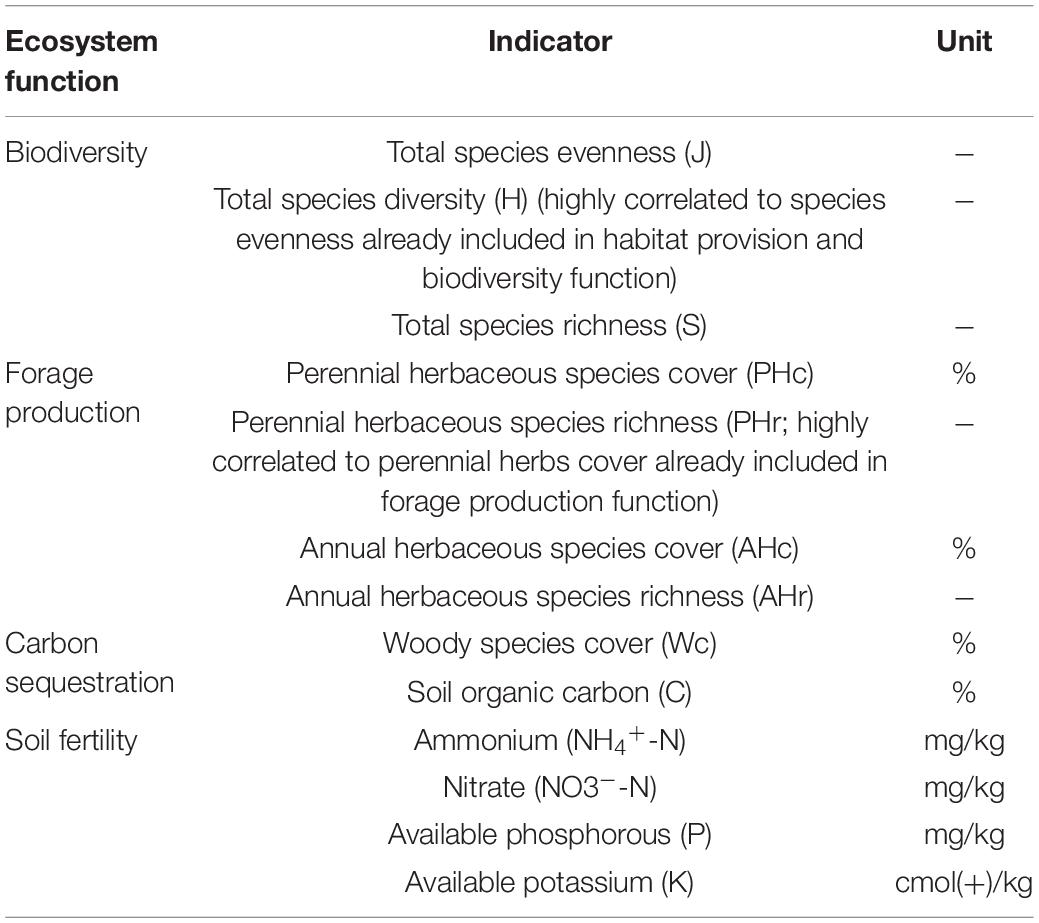
Table 1. Indicators of ecosystem functions considered in this study.
Total vascular plant species evenness, diversity and richness were used as a proxy of biodiversity function, which plays an important role in the promotion of ecosystem stability through multiple trophic levels (Haddad et al., 2011), pollination (Ebeling et al., 2011), biomass production (Reich et al., 2012), habitat complexity (Tews et al., 2004) and diversity of other functional groups (e.g., arthropods or birds; Kissling et al., 2008; Ebeling et al., 2014). Because of the importance of these processes, biodiversity has recently been considered as an ecosystem function on its own, and not only as a factor explaining EMF (Garland et al., 2021). Forage production function was assessed using the cover and richness of perennial and annual herbaceous species as proxies of aboveground biomass that can be consumed by livestock as a fraction of primary production in grasslands (Oñatibia et al., 2015; Moghli et al., 2022). Woody species cover and soil organic C, which are the main carbon pools in grassland ecosystems (Oñatibia et al., 2015), were measured as a proxy of carbon sequestration function, with substantial implications in climate regulation since grasslands store 20–25% of the global terrestrial C (Havstad et al., 2007). Finally, soil fertility function was derived from NH4+-N, NO3–-N, P and K content in the soil. NH4+-N is considered the preferred source of nitrogen (N) for soil microorganisms, and both NH4+-N and NO3–-N are the main source of N for vascular plants (Maestre et al., 2012b). Together with N, available P and K commonly limit plant growth in grassland ecosystems worldwide and are strongly associated with many metabolic processes in plants (Sardans and Peñuelas, 2015; Dong et al., 2019; Gao et al., 2019). Besides contributing to plant performance, the nutrients measured ultimately control many biogeochemical processes in terrestrial ecosystems (Maestre et al., 2012b).
We considered that the higher the values for the different indicators, the higher the ecosystem function following the recommendation of Maestre et al. (2012b). Highly correlated indicators within each ecosystem function (rpearson > 0.7) were removed to avoid redundant information (Moghli et al., 2022; Table 1). Raw indicators were first normalized using a square root transformation (Valencia et al., 2015), and then standardized using a z-score transformation (Bradford et al., 2014), which do not constrain the variability found in the raw ecosystem function indicators (Maestre et al., 2012b). The standardized indicators were grouped into the four ecosystem functions and then we averaged the functions to obtain the EMF index (Maestre et al., 2012b). This index is increasingly used in the literature (e.g., Maestre et al., 2012b; Bradford et al., 2014; Valencia et al., 2015; Mori et al., 2016; Huang et al., 2019; Lucas-Borja and Delgado-Baquerizo, 2019) because it provides a straightforward interpretation of ecosystem capacity to sustain multiple functions simultaneously (Byrnes et al., 2014) and is statically robust (Valencia et al., 2015).
The multivariate associations between all ecosystem function indicators (Table 1) and grazing treatment, as a proxy of grazing abandonment, were explored through a principal component analysis (PCA) and a permutational multivariate analysis of variance (PERMANOVA; 1000 permutations). These analyses were performed using R (R Core Team, 2021) and the “vegan” package (Oksanen et al., 2020). Differences in ecosystem function indicators among the levels of grazing treatment were tested with generalized linear mixed models (GLMMs) (Supplementary Material).
The effect of grazing treatment on both the four individual ecosystem functions and EMF index was evaluated using linear mixed models (LMMs). The response variables were: (i) biodiversity function, (ii) forage production function, (iii) carbon sequestration function, (iv) soil fertility function and (v) EMF, which were modeled following a Gaussian error distribution, using the identity link function. The predictor (fixed factor) in the models was the grazing treatment (control, short-term grazing exclusion and long-term grazing exclusion). The identity of the blocks nested within each sampling site (flat valley area, north aspect and south aspect) was included in the models as a nested random factor. The normality of model residuals was inspected through q-q plots. Predicted values of the response variable in the models were computed for each level of the grazing treatment, removing the uncertainty of the random effects (Taboada et al., 2018). We obtained the significance levels of the difference between short-term and long-term grazing exclusion, and the control treatment (allowed grazing) from the model output. The variance explained by fixed effects in the models was computed from the conditional R2 (Nakagawa and Schielzeth, 2013).
Linear mixed models were fitted using R (R Core Team, 2021) and the “lme4” package (Bates et al., 2015). Since lmer function does not provide p-values and R2, these parameters were computed using “lmerTest” (Kuznetsova et al., 2017) and “r2glmm” (Jaeger, 2017) packages, respectively.
Grazing abandonment induced significant shifts on the ecosystem functional indicators considered (PERMANOVA F = 9.53; p-value = 0.002). The samples of the three levels of grazing treatment (control, short-term and long-term grazing exclusion) were segregated by the first PCA axis, the long-term exclusion areas being more clearly separated and characterized by a high cover of woody species (Wc) (Figure 2). The remaining aboveground functional indicators (S, J, PHc, AHc, AHr), and more intensely NH4+-N and NO3–-N in relation to belowground indicators, tended to increase from the long-term exclusion areas to the grazed areas (control) (Figure 2). C, P and K soil content, highly correlated with the second PCA axis, was not associated with grazing treatment (Figures 1, 2 and Table 1 of the Supplementary Material). Control and short-term exclusion plots located at the sampling site with south aspect were clustered (Figure 2), which supports the inclusion of sampling site as a random factor in the LMMs for eliminating the associated variability.
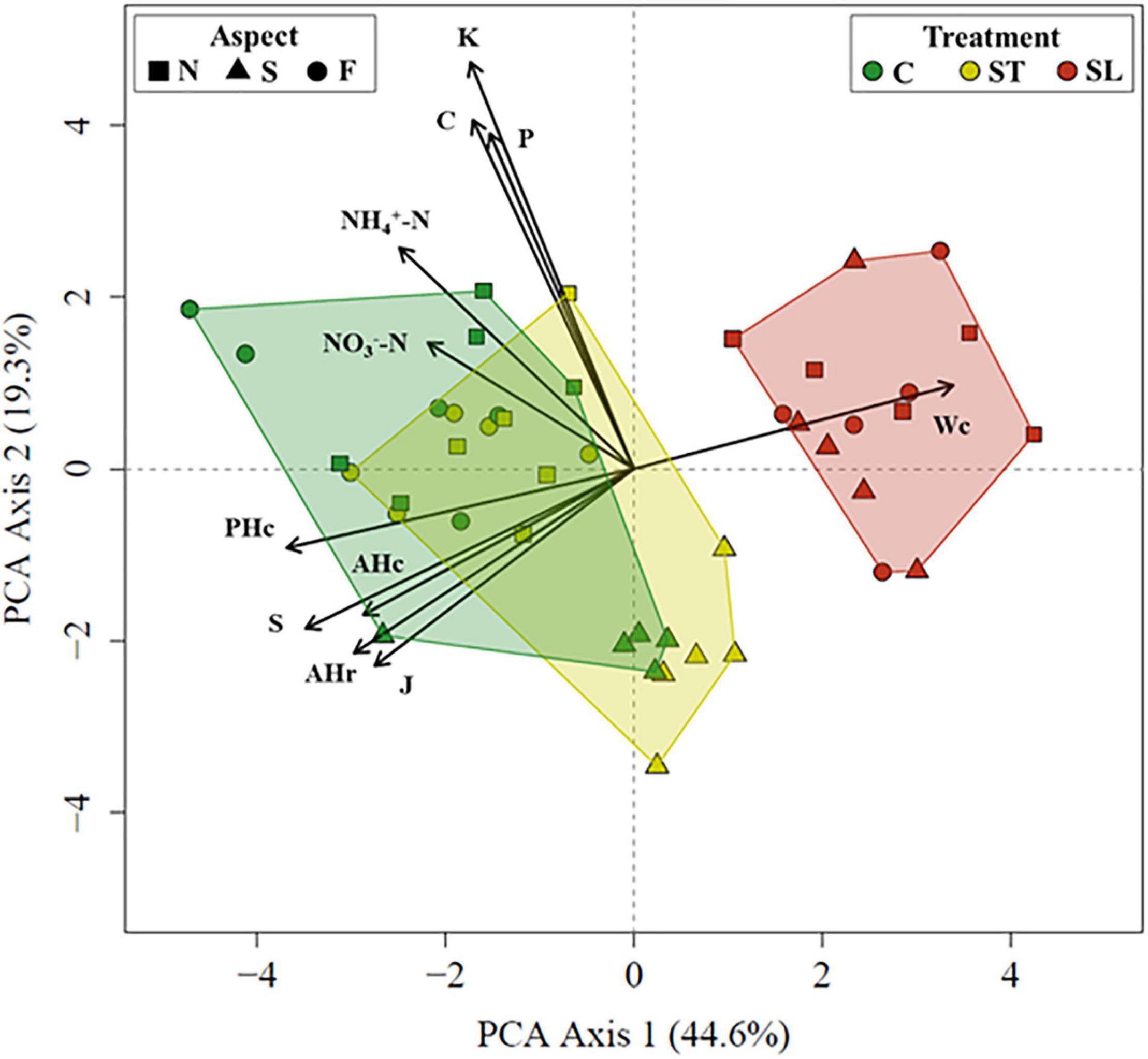
Figure 2. Principal component analysis (PCA) of the ecosystem function indicators. Variable codes correspond to those to those shown in Table 1. Convex hulls encompass all plots belonging to the same grazing treatment level (control -C- in green, short-term exclusion -ST- in yellow and long-term exclusion -LT- in red). Dot shape relates to the aspect (flat valley area - F-, north -N- and south -S-) of the sampling sites.
Individual ecosystem functions (i.e., biodiversity, forage production, carbon sequestration and soil fertility) and EMF varied with grazing treatment as a proxy of livestock abandonment, especially in relation to the long-term livestock exclusion. Biodiversity and forage production functions decreased significantly (p-value < 0.001) under long-term grazing exclusion relative to the control grazed areas (Table 2 and Figures 3A,B). Conversely, we found significantly higher values of the carbon sequestration function (p-value < 0.001) in long-term exclusion areas compared to the control (Table 2 and Figure 3C). The soil fertility function responded significantly and negatively to grazing exclusion in both the short (p-value = 0.018) and long term (p-value = 0.005) exclusions relative to the control situation (Table 2 and Figure 3D). Overall, grazing exclusion significantly impacted EMF, especially in long-term livestock exclusion areas (p-value < 0.001). The decline in EMF of short-term exclusion with respect to grazed areas was marginally significant (p-value = 0.059) (Table 2 and Figure 3E).
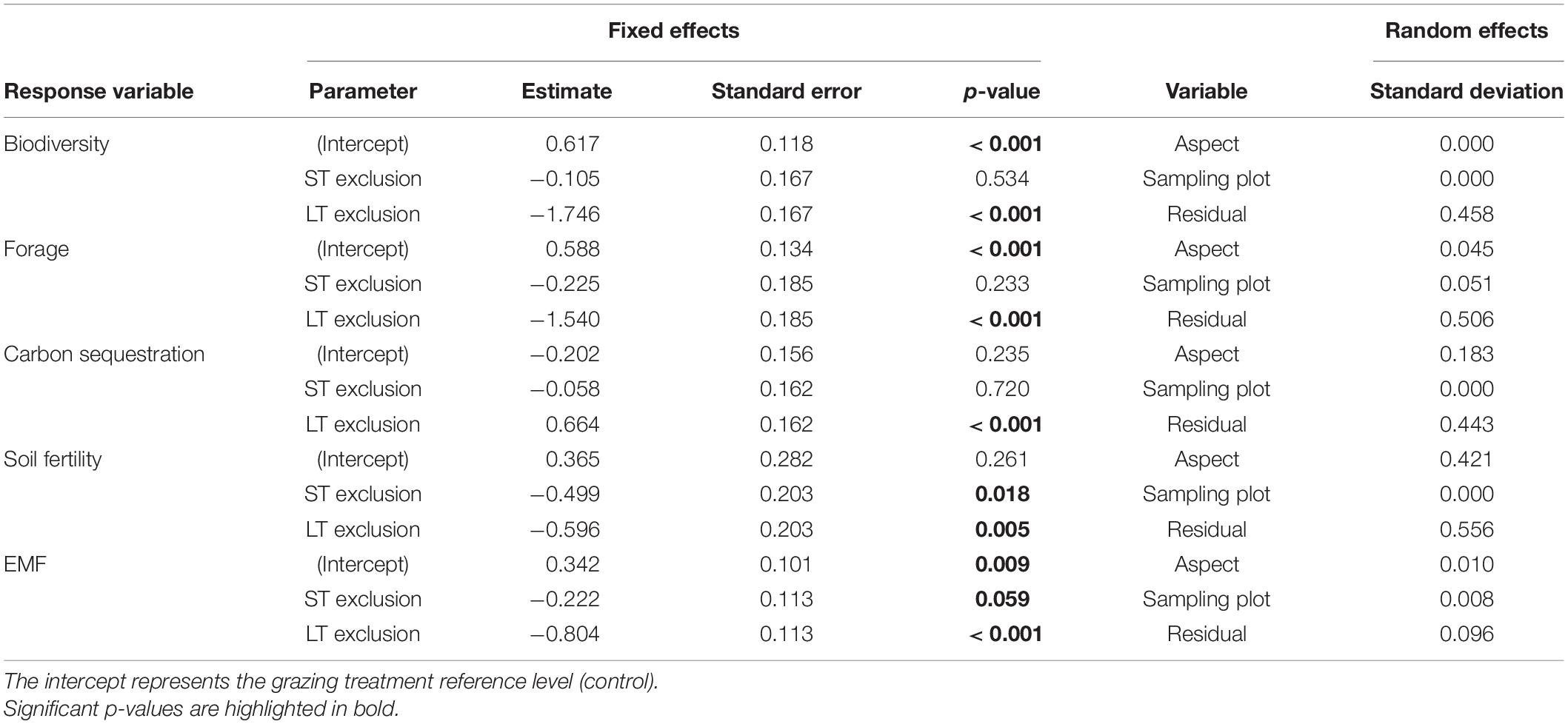
Table 2. Linear mixed model (LMM) results for the effects of grazing treatment (control, short-term -ST- and long-term -LT- grazing exclusion) on biodiversity, forage production, carbon sequestration and soil fertility ecosystem functions, as well as global ecosystem multifunctionality (EMF).
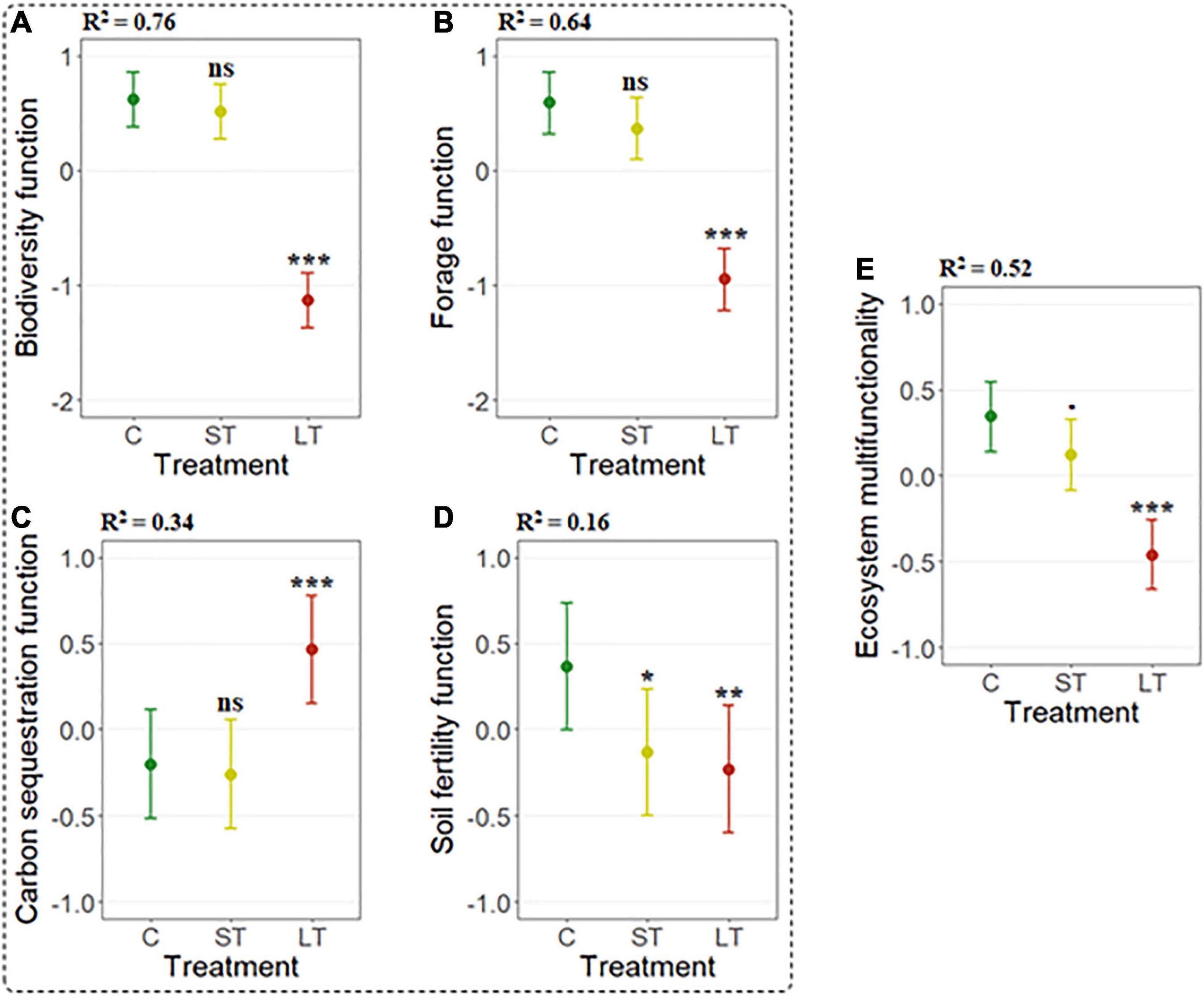
Figure 3. Linear mixed model (LMM) predicted effects (mean ± 95% confidence intervals) of grazing treatment (control - C-, short-term -ST- and long-term -LT- grazing exclusion) on individual ecosystem functions (A–D) and EMF (E). The significance of grazing treatment levels relative to the control is represented by ***(p-value < 0.001), **(p-value < 0.01), *(p-value < 0.05),⋅(p-value ≈ 0.05) and ns (p > 0.05). R2 indicates the standardized generalized variance explained by the fixed effects in the model.
Livestock herding is among the most widespread human activities and the dominant land use in grassland ecosystems (Alkemade et al., 2013; Yang et al., 2013), particularly in less-favored areas with harsh environmental conditions such as mountain regions (Erb et al., 2016). These areas exhibit a high vulnerability to marginalization and abandonment of traditional extensive grazing, such as the transhumance system (MacDonald et al., 2000). Accordingly, it is crucial to understand the effects of transhumant sheep grazing abandonment on the conservation of biodiversity and overall ecosystem function in mountain grasslands (Cingolani et al., 2014; Aldezabal et al., 2015). This study represents a first attempt to evaluate how short- and long-term abandonment affected EMF on mountain systems with long history of traditional grazing at moderate stocking rates. Our results provide evidence that this land use change induced negative impacts on most ecosystem functions, as well as on overall ecosystem functionality as we hypothesized. If the transhumant sheep grazing abandonment trend in mountainous regions of southern Europe continues, as expected (Aldezabal et al., 2015), the observed ecosystem function behavior would critically impair the ability of mountain grasslands to ensure ecosystem services delivery for the livelihoods of the inhabitants of these regions (Ren et al., 2018; García-Ruiz et al., 2020).
First, we found that long-term grazing exclusion significantly hampered biodiversity and forage production functions. In mesic grassland ecosystems, the relationships among plant diversity, productivity and herbivory are mostly modulated by light availability at the ground-level (Harpole and Tilman, 2007; Borer et al., 2014). Accordingly, the cessation of grazing in productive grasslands may prompt the exclusion of small-size plant species via a light competition mechanism (Odriozola et al., 2017) involving tall and upright herbaceous and woody species with strong competitive abilities (Bi et al., 2018), which dominated in the absence of transhumant sheep grazing in the long-term exclusions in this study. These competition mechanisms occur at neighborhood scale among individuals (∼1 m2 in grasslands; Borer et al., 2014), which is consistent with the scale of this study. Previous research also evidenced that woody species encroachment in semi-natural grasslands worldwide was a driver of biodiversity function decline (e.g., Mcadam et al., 2007; Báez and Collins, 2008; Eldridge et al., 2011; Komac et al., 2013; Koch et al., 2015). In addition, the lower biodiversity function found in long-term exclusions could be explained by the absence of transhumant sheep role as non-discriminant seed dispersers (Plue et al., 2019), and to the decrease in habitat spatial heterogeneity generated by patchiness in soil properties driven by livestock activity (Liu et al., 2016; Ingty, 2021). Along with biodiversity function decline, shrub encroachment has also been found to hinder forage availability as a consequence of competition (Zarovali et al., 2007; Rivest et al., 2011; Dahl et al., 2020), which is consistent with the lower forage production function found in plots under long-term transhumant sheep exclusion in this study. Also, grazing at moderate stocking rates can induce a production optimization process of herbaceous plants (Austrheim et al., 2014), that can be understood as a compensatory growth response induced by herbivory despite the herbaceous biomass consumption by domestic livestock (Oñatibia et al., 2015). This process can trigger herbaceous species cover increases (Angassa, 2014) and, therefore, forage production compared to areas where grazing is excluded.
Second, we found that carbon sequestration function (i.e., net balance of above and belowground carbon stocks) increased markedly in response to long-term transhumant sheep grazing exclusion, exhibiting a trade-off with biodiversity and forage production functions. It should be emphasized that woody species abundance was solely considered as the main aboveground carbon pool in terrestrial ecosystems (Moghli et al., 2022), since herbaceous species abundance was already used in this study as a proxy of forage production function. Nonetheless, woody species encroachment in traditionally managed grasslands leads to much greater gains in net aboveground carbon stocks than those associated with the absence of herbaceous biomass consumption by domestic livestock (Montané et al., 2007; Tanentzap and Coomes, 2012). Increased carbon sequestration function as a result of shrub species encroachment was not accompanied in this study with a significant response of soil organic carbon (Figure 1 and Table 1 of the Supplementary Material). Contradictory results regarding grazing exclusion effect in mountain grasslands were evidenced in previous research all over the world (e.g., Medina-Roldán et al., 2012; Shi et al., 2013; Speed et al., 2014; Lu et al., 2015; Bi et al., 2018). Here, the evidenced lack of soil carbon response could be related to the low pressure of transhumant sheep stocking rates (Shrestha and Stahl, 2008), and to the long-term nature of soil carbon accumulation mechanisms after grazing exclusion (more than 30 years; Medina-Roldán et al., 2012) regarding the time scale of this study.
Third, we found that both short and long-term grazing exclusion of transhumant sheep flocks undermined the soil fertility function. This behavior could be attributed to a slowing-down of ecosystem nutrient cycling caused either by (i) the promotion of plant functional traits related to high C/N ratios that hamper nutrient release in the short-term into the soil in the absence of grazing in productive grasslands (Semmartin et al., 2004; Odriozola et al., 2014; Aldezabal et al., 2015); and (ii) the lack of soil inputs of animal urine and feces, which are known to improve microbial activity (Medina-Roldán et al., 2012; Yang et al., 2013) and are an important pathway in nutrient cycling of grassland ecosystems (Lu et al., 2015).
The different patterns evidenced in the trends of the ecosystem functions considered (i.e., biodiversity, forage production, carbon sequestration and soil fertility) concerning grazing exclusion as a proxy of livestock abandonment conditions, suggest that grassland management policies may be biased when monitoring single ecosystem properties or functions exclusively (e.g., Schultz et al., 2011; Lu et al., 2015; Dlamini et al., 2016), instead of EMF (Ren et al., 2018). Overall, this study revealed a significant and gradual loss of ecosystem multifunctionality as time elapsed since livestock grazing exclusion. Therefore, transhumant sheep grazing with moderate stocking rates and short seasonal use is an activity that allows the conservation of the ecosystem multifunctionality in productive mountain grasslands with a long history of traditional livestock management. A straightforward implication of this result is that extensive livestock grazing is a key factor for conserving semi-natural mountain grassland ecosystems (Espunyes et al., 2019), since this land use is the only component that can be easily managed in mountain grasslands (Jarque-Bascuñana et al., 2022). Moreover, according to our findings, the marginally significant EMF loss over the short-term may imply that the overall function of these ecosystems could be feasibly recovered if transhumant sheep grazing use is reintroduced in case of recent loss (Pardo et al., 2015), as opposed to livestock abandonment over a long time period. Since the direct quantification of EMF could be challenging for the need of stakeholders (Luo et al., 2018), the identification of indirect EMF predictors such as those derived from remote sensing techniques (e.g., Arenas-Castro et al., 2019; LaRue et al., 2019) may be useful in future research for supporting sustainable management policies in grassland ecosystems with different environmental characteristics. Despite the impact of traditional livestock abandonment on EMF and the current European Union policies that promote the preservation of cultural landscapes (Corlett, 2016), we must emphasize that the cessation of human intervention for restoring original ecosystem processes by natural succession (i.e., rewilding; Perino et al., 2019) may be preferred when the goal is to promote specific ecosystem functions and services. For instance, woody species encroachment in semi-natural mountain grasslands can lay the foundations for carbon sequestration service, as evidenced in this study. In addition, indirect and non-use services, which are often disregarded in policy-making processes (Häyhä et al., 2015), can be substantially favored by rewilding (Pereira and Navarro, 2015). Finally, rewilding processes involving woody plant encroachment elicit complex changes in the trophic structure of micro and macrofauna (Thakur et al., 2020; Sepp et al., 2021), which are themselves strongly related to ecosystem functioning (Biederman and Boutton, 2009).
Our findings suggest that livestock grazing exclusion, as a proxy of traditional livestock use abandonment on mountain grasslands with long evolutionary history of grazing at moderate stocking rates, significantly hindered ecosystem multifunctionality, especially in areas subject to long-term grazing exclusion. Therefore, the transhumance system is crucial for the conservation of grassland ecosystems in mountainous regions. Since short- and long-term grazing exclusion had effects of varying direction and magnitude on the individual ecosystem functions considered (i.e., biodiversity, forage production, carbon sequestration and soil fertility), management policies should consider the overall ecosystem function for avoiding perception biases.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
JF-G, RT, EM, LV, and LC conceived and designed the experiment. JF-G, VF-G, RT, RP, PM, DB, and SH analyzed the data. JF-G wrote the first draft of the manuscript. VF-G, RT, EM, LV, RP, PM, DB, SH, and LC contributed to the writing. LC acquired the funding and coordinated the study. All authors read and approved the final manuscript.
This study was financially supported by the Spanish Ministry of Economy and Competitiveness, and the European Regional Development Fund (ERDF), in the framework of the FIRESEVES (AGL2017-86075-C2-1-R) project; by the Regional Government of Castilla and León in the framework of the WUIFIRECYL (LE005P20) project; and by a contract with CESEFOR (Fundación Centro de Servicios y Promoción Forestal y de su Industria de Castilla y León) in the framework of the project OVINNOVA (N° reg: 0190020007497), which was subsidized by the Spanish Ministry of Agriculture, Fisheries and Food.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.861611/full#supplementary-material
Aldezabal, A., Moragues, L., Odriozola, I., and Mijangos, I. (2015). Impact of grazing abandonment on plant and soil microbial communities in an Atlantic mountain grassland. Appl. Soil Ecol. 96, 251–260. doi: 10.1016/j.apsoil.2015.08.013
Alkemade, R., Reid, R. S., van den Berg, M., de Leeuw, J., and Jeuken, M. (2013). Assessing the impacts of livestock production on biodiversity in rangeland ecosystems. Proc. Natl. Acad. Sci. U.S.A. 110, 20900–20905. doi: 10.1073/pnas.1011013108
Álvarez-Martínez, J., Gómez-Villar, A., and Lasanta, T. (2016). The use of goats grazing to restore pastures invaded by shrubs and avoid desertification: a preliminary case study in the Spanish Cantabrian Mountains. Land Degrad. Dev. 27, 3–13. doi: 10.1002/ldr.2230
Anderson, S., Anderson, W., Hines, F., and Fountain, A. (2005). Determination of Field Sampling Methods for the Assessment of Curing Levels in Grasslands. Bushfire Cooperative Research Centre, Project A1.4 Report. East Melbourne: Bushfire Cooperative Research Centre.
Angassa, A. (2014). Effects of grazing intensity and bush encroachment on herbaceous species and rangeland condition in southern Ethiopia. Land Degrad. Dev. 25, 438–451. doi: 10.1002/ldr.2160
Arenas-Castro, S., Regos, A., Gonçalves, J. F., Alcaraz-Segura, D., and Honrado, J. (2019). Remotely sensed variables of ecosystem functioning support robust predictions of abundance patterns for Rare Species. Remote Sens. 11:2086. doi: 10.1371/journal.pone.0138995
Austrheim, G., Speed, J. D. M., Martinsen, V., Mulder, J., and Mysterud, A. (2014). Experimental effects of herbivore density on aboveground plant biomass in an Alpine grassland ecosystem. Arct. Antarct. Alp. Res. 46, 535–541. doi: 10.1657/1938-4246-46.3.535
Aynekulu, E., Mekuria, W., Tsegaye, D., Feyissa, K., Angassa, A., de Leeuw, J., et al. (2017). Long-term livestock exclosure did not affect soil carbon in southern Ethiopian rangelands. Geoderma 307, 1–7. doi: 10.1016/j.geoderma.2017.07.030
Báez, S., and Collins, S. L. (2008). Shrub Invasion Decreases Diversity and Alters Community Stability in Northern Chihuahuan Desert Plant Communities. PLoS One 3:e2332. doi: 10.1371/journal.pone.0002332
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48.
Bendel, C. R., Kral-O’Brien, K. C., Hovick, T. J., Limb, R. F., and Harmon, J. P. (2019). Plant–pollinator networks in grassland working landscapes reveal seasonal shifts in network structure and composition. Ecosphere 10:e02569.
Bengtsson, J., Bullock, J. M., Egoh, B., Everson, C., Everson, T., O’Connor, T., et al. (2019). Grasslands-more important for ecosystem services than you might think. Ecosphere 10:e02582. doi: 10.1016/j.scitotenv.2015.06.128
Bi, X., Li, B., Fu, Q., Fan, Y., Ma, L., Yang, Z., et al. (2018). Effects of grazing exclusion on the grassland ecosystems of mountain meadows and temperate typical steppe in a mountain-basin system in Central Asia’s arid regions, China. Sci. Total Environ. 630, 254–263. doi: 10.1016/j.scitotenv.2018.02.055
Biederman, L. A., and Boutton, T. W. (2009). Biodiversity and trophic structure of soil nematode communities are altered following woody plant invasion of grassland. Soil Biol. Biochem. 41, 1943–1950. doi: 10.1016/j.soilbio.2009.06.019
Blanco, G., Cortés-Avizanda, A., Frías, O., Arrondo, E., and Donázar, J. A. (2019). Livestock farming practices modulate vulture diet-disease interactions. Glob. Ecol. Conserv. 17:e00518. doi: 10.1016/j.gecco.2018.e00518
Borer, E. T., Seabloom, E. W., Gruner, D. S., Harpole, W. S., and Hillebrand, H. (2014). Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508, 517–520. doi: 10.1038/nature13144
Bradford, M. A., Wood, S. A., Bardgett, R. D., Black, H. I. J., Bonkowski, M., Eggers, T., et al. (2014). Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition. Proc. Natl. Acad. Sci. U.S.A. 111, 14478–14483. doi: 10.1073/pnas.1413707111
Byrnes, J. E. K., Gamfeldt, L., Isbell, F., Lefcheck, J. S., Griffin, J. N., Hector, A., et al. (2014). Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods Ecol. Evol. 5, 111–124. doi: 10.1111/2041-210x.12143
Calvo, L., Santalla, S., Valbuena, L., Marcos, E., Tárrega, R., and Luis-Calabuig, E. (2008). Post-fire natural regeneration of a Pinus pinaster forest in NW Spain. Plant Ecol. 197, 81–90. doi: 10.1007/s11258-007-9362-1
Calvo, L., Tarrega, R., and Luis, E. (2002). Regeneration patterns in a Calluna vulgaris heathland in the Cantabrian mountains (NW Spain): effects of burning, cutting and ploughing. Acta Oecol. 23, 81–90. doi: 10.1016/s1146-609x(02)01137-2
Chen, B., Zhang, X., Tao, J., Wu, J., Wang, J., Shi, P., et al. (2014). The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agric. For. Meteorol. 189, 11–18. doi: 10.1016/j.agrformet.2014.01.002
Cingolani, A. M., Vaieretti, M. V., Giorgis, M. A., Poca, M., Tecco, P. A., and Gurvich, D. E. (2014). Can livestock grazing maintain landscape diversity and stability in an ecosystem that evolved with wild herbivores? Perspect. Plant Ecol. Evol. Syst. 16, 143–153. doi: 10.1016/j.ppees.2014.04.002
Cocca, G., Sturaro, E., Gallo, L., and Ramanzin, M. (2012). Is the abandonment of traditional livestock farming systems the main driver of mountain landscape change in Alpine areas? Land Use Policy 29, 878–886. doi: 10.1016/j.landusepol.2012.01.005
Corlett, R. T. (2016). The role of rewilding in landscape design for conservation. Curr. Landsc. Ecol. Rep. 1, 127–133. doi: 10.1016/j.jenvman.2020.111392
Dahl, R., Dalgaard, T., and Bork, E. W. (2020). Shrub encroachment following wetland creation in mixedgrass prairie alters grassland vegetation and soil. Environ. Manage. 66, 1120–1132. doi: 10.1007/s00267-020-01386-2
De Bello, F., Lepš, J., and Sebastià, M.-T. (2005). Predictive value of plant traits to grazing along a climatic gradient in the Mediterranean. J. Appl. Ecol. 42, 824–833. doi: 10.1111/j.1365-2664.2005.01079.x
Dlamini, P., Chivenge, P., and Chaplot, V. (2016). Overgrazing decreases soil organic carbon stocks the most under dry climates and low soil pH: a meta-analysis shows. Agric. Ecosyst. Environ. 221, 258–269. doi: 10.1016/j.agee.2016.01.026
Dong, C., Wang, W., Liu, H., Xu, X., and Zeng, H. (2019). Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: evidence from soil extracellular enzyme stoichiometry. Ecol. Indic. 101, 453–464. doi: 10.1016/j.ecolind.2019.01.046
Ebeling, A., Klein, A. M., and Tscharntke, T. (2011). Plant–flower visitor interaction webs: temporal stability and pollinator specialization increases along an experimental plant diversity gradient. Basic Appl. Ecol. 12, 300–309. doi: 10.1016/j.baae.2011.04.005
Ebeling, A., Meyer, S. T., Abbas, M., Eisenhauer, N., Hillebrand, H., Lange, M., et al. (2014). Plant diversity impacts decomposition and herbivory via changes in aboveground arthropods. PLoS One 9:e106529. doi: 10.1371/journal.pone.0106529
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722. doi: 10.1111/j.1461-0248.2011.01630.x
Erb, K. H., Fetzel, T., Kastner, T., Kroisleitner, C., Lauk, C., Mayer, A., et al. (2016). “Livestock grazing, the neglected land use,” in Social Ecology. Human-Environment Interactions, eds H. Haberl, M. Fischer-Kowalski, F. Krausmann, and V. Winiwarter (Cham: Springer).
Espunyes, J., Lurgi, M., Büntgen, U., Bartolomé, J., Calleja, J. A., Gálvez-Cerón, A., et al. (2019). Different effects of alpine woody plant expansion on domestic and wild ungulates. Glob. Chang. Biol. 25, 1808–1819. doi: 10.1111/gcb.14587
Evans, R. D., Gill, R. A., Eviner, V. T., and Bailey, V. (2017). “Soil and belowground processes,” in Rangeland Systems: Processes, Management, and Challenges, ed. D. Briske (Cham: Springer), 131–168.
Fernández-García, V., Marcos, E., Huerta, S., and Calvo, L. (2021). Soil-vegetation relationships in Mediterranean forests after fire. For. Ecosyst. 8:18.
Gao, J., Meng, B., Liang, T., Feng, O., Ge, J., Yin, J., et al. (2019). Modeling alpine grassland forage phosphorus based on hyperspectral remote sensing and a multi-factor machine learning algorithm in the east of Tibetan Plateau, China. ISPRS J. Photogramm. Remote Sens. 147, 104–117. doi: 10.1016/j.isprsjprs.2018.11.015
García-Llamas, P., Geijzendorffer, I. R., García-Nieto, A. P., Calvo, L., Suárez-Seoane, S., and Cramer, W. (2019). Impact of land cover change on ecosystem service supply in mountain systems: a case study in the Cantabrian Mountains (NW of Spain). Reg. Environ. Change 19, 529–542. doi: 10.1007/s10113-018-1419-2
García-Ruiz, J. M., Tomás-Faci, G., Diarte-Blasco, P., Montes, L., Domingo, R., Sebastián, M., et al. (2020). Transhumance and long-term deforestation in the subalpine belt of the central Spanish Pyrenees: an interdisciplinary approach. CATENA 195:104744. doi: 10.1016/j.catena.2020.104744
Garland, G., Banerjee, S., Edlinger, A., Oliveira, E. M., Herzog, C., Wittwer, R., et al. (2021). A closer look at the functions behind ecosystem multifunctionality: a review. J. Ecol. 109, 600–613. doi: 10.1111/1365-2745.13511
Gennet, S., Spotswood, E., Hammond, M., and Bartolome, J. W. (2017). Livestock grazing supports native plants and songbirds in a California annual grassland. PLoS One 12:e0176367. doi: 10.1371/journal.pone.0176367
GEODE (2020). Cartografía Geológica Digital Continua a Escala 1:50.000. Madrid: Instituto Geológico y Minero de España.
Haddad, N. M., Crutsinger, G. M., Gross, K., Haarstad, J., and Tilman, D. (2011). Plant diversity and the stability of foodwebs. Ecol. Lett. 14, 42–46. doi: 10.1111/j.1461-0248.2010.01548.x
Hao, L., Pan, C., Fang, D., Zhang, X., Zhou, D., Liu, P., et al. (2018). Quantifying the effects of overgrazing on mountainous watershed vegetation dynamics under a changing climate. Sci. Total Environ. 639, 1408–1420. doi: 10.1016/j.scitotenv.2018.05.224
Harpole, W. S., and Tilman, D. (2007). Grassland species loss resulting from reduced niche dimension. Nature 446, 791–793. doi: 10.1038/nature05684
Havstad, K. M., Peters, D. P. C., Skaggs, R., Brown, J., Bestelmeyer, B., Fredrickson, E., et al. (2007). Ecological services to and from rangelands of the United States. Ecol. Econ. 64, 261–268. doi: 10.1016/j.ecolecon.2007.08.005
Häyhä, T., Franzese, P. P., Paletto, A., and Fath, B. D. (2015). Assessing, valuing, and mapping ecosystem services in Alpine forests. Ecosyst. Serv. 14, 12–23. doi: 10.1016/j.ecoser.2015.03.001
Hector, A., and Bagchi, R. (2007). Biodiversity and ecosystem multifunctionality. Nature 448, 188–190. doi: 10.1038/nature05947
Hilker, T., Natsagdorj, E., Waring, R. H., Lyapustin, A., and Wang, Y. (2014). Satellite observed widespread decline in Mongolian grasslands largely due to overgrazing. Glob. Chang. Biol. 20, 418–428. doi: 10.1111/gcb.12365
Huang, X., Su, J., Li, S., Liu, W., and Lang, X. (2019). Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Sci. Rep. 9:6979. doi: 10.1038/s41598-019-43475-1
Ingty, T. (2021). Pastoralism in the highest peaks: role of the traditional grazing systems in maintaining biodiversity and ecosystem function in the alpine Himalaya. PLoS One 16:e0245221. doi: 10.1371/journal.pone.0245221
Jackson, R. D., and Bartolome, J. W. (2002). A state-transition approach to understanding nonequilibrium plant community dynamics in Californian Grasslands. Plant Ecol. 162, 49–65.
Jaeger, B. (2017). r2glmm: Computes R Squared for Mixed (Multilevel) Models. R package version 0.1.2. Available online at: https://CRAN.R-project.org/package=r2glmm (accessed January 10, 2022).
Jarque-Bascuñana, L., Calleja, J. A., Ibañez, M., Bartolomé, J., Albanell, E., Espunyes, J., et al. (2022). Grazing influences biomass production and protein content of alpine meadows. Sci. Total Environ. 818:151771. doi: 10.1016/j.scitotenv.2021.151771
Jing, X., Sanders, N. J., Shi, Y., Chu, H., Classen, A. T., Zhao, K., et al. (2015). The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nat. Commun. 6:8159. doi: 10.1038/ncomms9159
Keeney, D. R., and Nelson, D. W. (1982). “Nitrogen in organic forms,” in Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties, eds A. L. Page, R. H. Miller, and D. R. Keeney (Madison, WI: American Society of Agronomy), 643–698.
Kissling, W. D., Field, R., and Böhning-Gaese, K. (2008). Spatial patterns of woody plant and bird diversity: functional relationships or environmental effects? Glob. Ecol. Biogeogr. 17, 327–339. doi: 10.1111/j.1466-8238.2007.00379.x
Koch, B., Edwards, P. J., Blanckenhorn, W. U., Walter, T., and Hofer, G. (2015). Shrub encroachment affects the diversity of plants, butterflies, and grasshoppers on two swiss subalpine pastures. Arct. Antarct. Alp. Res. 47, 345–357. doi: 10.1657/aaar0013-093
Komac, B., Kefi, S., Nuche, P., Escós, J., and Alados, C. L. (2013). Modeling shrub encroachment in subalpine grasslands under different environmental and management scenarios. J. Environ. Manage. 121, 160–169. doi: 10.1016/j.jenvman.2013.01.038
Kuznetsova, A., Brockhoff, P. B., and Christensen, R. H. B. (2017). lmerTest Package: tests in linear mixed effects models. J. Stat. Softw. 82, 1–26.
LaRue, E. A., Hardiman, B. S., Elliott, J. M., and Fei, S. (2019). Structural diversity as a predictor of ecosystem function. Environ. Res. Lett. 14:114011. doi: 10.1088/1748-9326/ab49bb
Lasanta, T., González-Hidalgo, J. C., Vicente-Serrano, S. M., and Sferi, E. (2006). Using landscape ecology to evaluate an alternative management scenario in abandoned Mediterranean mountain areas. Landsc. Urban Plan. 78, 101–114. doi: 10.1016/j.landurbplan.2005.06.003
Liu, C., Song, X., Wang, L., Wang, D., Zhou, X., Liu, J., et al. (2016). Effects of grazing on soil nitrogen spatial heterogeneity depend on herbivore assemblage and pre-grazing plant diversity. J. Appl. Ecol. 53, 242–250. doi: 10.1111/1365-2664.12537
Lu, X., Yan, Y., Sun, J., Zhang, X., Chen, Y., Wang, X., et al. (2015). Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: effects of grazing exclusion. Ecol. Evol. 5, 4492–4504. doi: 10.1002/ece3.1732
Lucas-Borja, M. E., and Delgado-Baquerizo, M. (2019). Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession. Sci. Total Environ. 697, 134204. doi: 10.1016/j.scitotenv.2019.134204
Lucas-Borja, M. E., Delgado-Baquerizo, M., Muñoz-Rojas, M., Plaza-Álvarez, P. A., Gómez-Sanchez, M. E., González-Romero, J., et al. (2021). Changes in ecosystem properties after post-fire management strategies in wildfire-affected Mediterranean forests. J. Appl. Ecol. 58, 836–846. doi: 10.1111/1365-2664.13819
Luo, G., Rensing, C., Chen, H., Liu, M., Wang, M., Guo, S., et al. (2018). Deciphering the associations between soil microbial diversity and ecosystem multifunctionality driven by long-term fertilization management. Funct. Ecol. 32, 1103–1116. doi: 10.1111/1365-2435.13039
MacDonald, D., Crabtree, J. R., Wiesinger, G., Dax, T., Stamou, N., Fleury, P., et al. (2000). Agricultural abandonment in mountain areas of Europe: environmental consequences and policy response. J. Environ. Manage. 59, 47–69. doi: 10.1006/jema.1999.0335
Maestre, F. T., Castillo-Monroy, A. P., Bowker, M. A., and Ochoa-Hueso, R. (2012a). Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J. Ecol. 100, 317–330. doi: 10.1111/j.1365-2745.2011.01918.x
Maestre, F. T., Quero, J. L., Gotelli, N. J., Escudero, A., Ochoa, V., Delgado-Baquerizo, M., et al. (2012b). Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 335, 214–218.
Mcadam, J. H., Sibbald, A. R., Teklehaimanot, Z., and Eason, W. R. (2007). Developing silvopastoral systems and their effects on diversity of fauna. Agrofor. Syst. 70, 81–89. doi: 10.1007/s10457-007-9047-8
McSherry, M. E., and Ritchie, M. E. (2013). Effects of grazing on grassland soil carbon: a global review. Glob. Chang. Biol. 19, 1347–1357. doi: 10.1111/gcb.12144
Medina-Roldán, E., Paz-Ferreiro, J., and Bardgett, R. D. (2012). Grazing exclusion affects soil and plant communities, but has no impact on soil carbon storage in an upland grassland. Agric. Ecosyst. Environ. 149, 118–123. doi: 10.1016/j.agee.2011.12.012
Moghli, A., Santana, V. M., Soliveres, S., and Baeza, M. J. (2022). Thinning and plantation of resprouting species redirect overstocked pine stands towards more functional communities in the Mediterranean basin. Sci. Total Environ. 806:150715. doi: 10.1016/j.scitotenv.2021.150715
Montané, F., Rovira, P., and Casals, P. (2007). Shrub encroachment into mesic mountain grasslands in the Iberian peninsula: effects of plant quality and temperature on soil C and N stocks. Glob. Biogeochem. Cycles 21:GB4016.
Morán-Ordóñez, A., Bugter, R., Suárez-Seoane, S., de Luis, E., and Calvo, L. (2013). Temporal changes in socio-ecological systems and their impact on ecosystem services at different governance scales: a case study of heathlands. Ecosystems 16, 765–782. doi: 10.1007/s10021-013-9649-0
Morán-Ordóñez, A., Suárez-Seoane, S., Calvo, L., and de Luis, E. (2011). Using predictive models as a spatially explicit support tool for managing cultural landscapes. Appl. Geogr. 31, 839–848. doi: 10.1016/j.apgeog.2010.09.002
Mori, A. S., Isbell, F., Fujii, S., Makoto, K., Matsuoka, S., and Osono, T. (2016). Low multifunctional redundancy of soil fungal diversity at multiple scales. Ecol. Lett. 19, 249–259. doi: 10.1111/ele.12560
Nakagawa, S., and Schielzeth, H. (2013). A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 4, 133–142. doi: 10.1093/sysbio/syy060
Nelson, D. W., and Sommers, L. E. (1996). “Total carbon, organic carbon, and organic matter,” in Methods of Soil Analysis Part 3-chemical Methods, ed. D. L. Sparks (Madison, WI: American Society of Agronomy), 961–1010. doi: 10.2136/sssabookser5.3.c34
Ninyerola, M., Pons, X., and Roure, J. M. (2005). Atlas Climático Digital de la Península Ibérica. Metodología y Aplicaciones en Bioclimatología y Geobotánica. Bellaterra: Universidad Autónoma de Barcelona
Odriozola, I., García-Baquero, G., Fortin, M.-J., Laskurain, N. A., and Aldezabal, A. (2017). Grazing exclusion unleashes competitive plant responses in Iberian Atlantic mountain grasslands. Appl. Veg. Sci. 20, 50–61. doi: 10.1111/avsc.12277
Odriozola, I., García-Baquero, G., Laskurain, N. A., and Aldezabal, A. (2014). Livestock grazing modifies the effect of environmental factors on soil temperature and water content in a temperate grassland. Geoderma 236, 347–354. doi: 10.1016/j.geoderma.2014.08.002
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2020). vegan: Community Ecology Package. R package version 2.5-7. Available online at: https://CRAN.R-project.org/package=vegan (accessed January 10, 2022).
Öllerer, K., Varga, A., Kirby, K., Demeter, L., Biró, M., Bölöni, J., et al. (2019). Beyond the obvious impact of domestic livestock grazing on temperate forest vegetation – A global review. Biol. Conserv. 237, 209–219. doi: 10.1016/j.biocon.2019.07.007
Olsen, S. R., Cole, C. V., Frank, S. W., and Dean, L. A. (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. USDA Circular No. 939. Washington, DC: US Government Printing Office, 19.
Oñatibia, G. R., Aguiar, M. R., and Semmartin, M. (2015). Are there any trade-offs between forage provision and the ecosystem service of C and N storage in arid rangelands? Ecol. Eng. 77, 26–32. doi: 10.1016/j.ecoleng.2015.01.009
Pardo, I., Doak, D. F., García-González, R., Gómez, D., and García, M. B. (2015). Long-term response of plant communities to herbivore exclusion at high elevation grasslands. Biodivers. Conserv. 24, 3033–3047. doi: 10.1007/s10531-015-0996-3
Pe’er, G., Dicks, L., Visconti, P., Arlettaz, R., Báldi, A., Benton, T., et al. (2014). EU agricultural reform fails on biodiversity. Science 344, 1090–1092. doi: 10.1126/science.1253425
Perino, A., Pereira, H. M., Navarro, L. M., Fernández, N., Bullock, J. M., Ceausu, S., et al. (2019). Rewilding complex ecosystems. Science 364:eaav5570.
Pielou, E. C. (1966). Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 10, 370–383. doi: 10.1016/0022-5193(66)90133-0
Plue, J., Aavik, T., and Cousins, S. A. O. (2019). Grazing networks promote plant functional connectivity among isolated grassland communities. Divers. Distrib. 25, 102–115. doi: 10.1111/ddi.12842
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reich, P. B., Tilman, D., Isbell, F., Mueller, K., Hobbie, S. E., Flynn, D. F. B., et al. (2012). Impacts of biodiversity loss escalate through time as redundancy fades. Science 336, 589–592. doi: 10.1126/science.1217909
Ren, H., Eviner, V. T., Gui, W., Wilson, G. W. T., Cobb, A. B., Yang, G., et al. (2018). Livestock grazing regulates ecosystem multifunctionality in semi-arid grassland. Funct. Ecol. 32, 2790–2800. doi: 10.1111/1365-2435.13215
Rivas-Martínez, S., Bandullo, J. M., Allué, J. L., Montero de Burgos, J. L., and González, J. L. (1987). Memoria del Mapa Series de Vegetación de España. Madrid: ICONA.
Rivest, D., Rolo, V., López-Díaz, L., and Moreno, G. (2011). Shrub encroachment in Mediterranean silvopastoral systems: Retama sphaerocarpa and Cistus ladanifer induce contrasting effects on pasture and Quercus ilex production. Agric. Ecosyst. Environ. 141, 447–454. doi: 10.1016/j.agee.2011.04.018
Román-Trufero, A., Martínez, A., Ferreira, L. M. M., García-Prieto, V., Rosa-García, R., Osoro, K., et al. (2019). Foraging behaviour and performance of steers from two local breeds (Asturian Valley and Asturian Mountain) grazing in Cantabrian (N Spain) summer pastures. Span. J. Agric. Res. 17:e0601. doi: 10.5424/sjar/2019171-13541
Sardans, J., and Peñuelas, J. (2015). Potassium stoichiometry and global change. Glob. Ecol. Biogeogr. 24, 261–275. doi: 10.1111/geb.12259
Schultz, N. L., Morgan, J. W., and Lunt, I. D. (2011). Effects of grazing exclusion on plant species richness and phytomass accumulation vary across a regional productivity gradient. J. Veg. Sci. 22, 130–142. doi: 10.1111/j.1654-1103.2010.01235.x
Sebastià, M. T., de Bello, F., Puig, L., and Taull, M. (2008). Grazing as a factor structuring grasslands in the Pyrenees. Appl. Veg. Sci. 11, 215–222. doi: 10.3170/2008-7-18358
Semmartin, M., Aguiar, M. R., Distel, R. A., Moretto, A. S., and Ghersa, C. M. (2004). Litter quality and nutrient cycling affected by grazing-induced species replacements along a precipitation gradient. Oikos 107, 148–160. doi: 10.1111/j.0030-1299.2004.13153.x
Semmartin, M., Garibaldi, L. A., and Chaneton, E. J. (2008). Grazing history effects on above- and below-ground litter decomposition and nutrient cycling in two co-occurring grasses. Plant Soil 303, 177–189. doi: 10.1007/s11104-007-9497-9
Sepp, S.-K., Davison, J., Moora, M., Neuenkamp, L., Oja, J., Roslin, T., et al. (2021). Woody encroachment in grassland elicits complex changes in the functional structure of above- and belowground biota. Ecosphere 12:e03512.
Shannon, C. E., and Weaver, W. (1949). The Mathematical Theory of Communication. Urbana, IL: The University of Illinois Press.
Shi, X. M., Li, X. G., Li, C. T., Zhao, Y., Shang, Z. H., and Ma, Q. (2013). Grazing exclusion decreases soil organic C storage at an alpine grassland of the Qinghai–Tibetan Plateau. Ecol. Eng. 57, 183–187. doi: 10.1016/j.ecoleng.2013.04.032
Shrestha, G., and Stahl, P. D. (2008). Carbon accumulation and storage in semi-arid sagebrush steppe: Effects of long-term grazing exclusion. Agric. Ecosyst. Environ. 125, 173–181. doi: 10.1016/j.agee.2007.12.007
Speed, J. D. M., Martinsen, V., Mysterud, A., Mulder, J., Holand, Ø., and Austrheim, G. (2014). Long-term increase in aboveground carbon stocks following exclusion of grazers and forest establishment in an alpine ecosystem. Ecosystems 17, 1138–1150. doi: 10.1007/s10021-014-9784-2
Taboada, A., Calvo-Fernández, J., Marcos, E., and Calvo, L. (2018). Plant and vegetation functional responses to cumulative high nitrogen deposition in rear-edge heathlands. Sci. Total Environ. 638, 980–990. doi: 10.1016/j.scitotenv.2018.05.092
Tanentzap, A. J., and Coomes, D. A. (2012). Carbon storage in terrestrial ecosystems: do browsing and grazing herbivores matter? Biol. Rev. 87, 72–94. doi: 10.1111/j.1469-185X.2011.00185.x
Targetti, S., Staglianò, N., Messeri, A., and Argenti, G. (2010). A state-and-transition approach to alpine grasslands under abandonment. iForest 3, 44–51. doi: 10.3832/ifor0525-003
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schwager, M., et al. (2004). Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J. Biogeogr. 31, 79–92. doi: 10.1046/j.0305-0270.2003.00994.x
Thakur, M. P., Bakker, E. S., Veen, G. F., and Harvey, J. A. (2020). Climate extremes, rewilding, and the role of microhabitats. One Earth 2, 506–509. doi: 10.1016/j.oneear.2020.05.010
Valencia, E., Maestre, F. T., Le Bagousse-Pinguet, Y., Quero, J. L., Tamme, R., Börger, L., et al. (2015). Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands. New Phytol. 206, 660–671. doi: 10.1111/nph.13268
Velado-Alonso, E., and Gómez-Sal, A. (2016). “The current status of transhumance systems in the province of León (Spain), towards a multi-dimensional evaluation,” in Proceeding of the Meeting of the FAO-CIHEAM Mountain Pastures Subnetwork, Zaragoza.
Warncke, D., and Brown, J. R. (1998). “Potassium and other basic cations,” in Recommended Chemical Soil Test Procedures for the North Central Region. North Central Regional Research Publication Number 221, ed. J. L. Brown (Columbia, MI: University of Missouri), 31–33.
Yang, Y., Wu, L., Lin, Q., Yuan, M., Xu, D., Yu, H., et al. (2013). Responses of the functional structure of soil microbial community to livestock grazing in the Tibetan alpine grassland. Glob. Chang. Biol. 19, 637–648. doi: 10.1111/gcb.12065
Zarovali, M. P., Yiakoulaki, M. D., and Papanastasis, V. P. (2007). Effects of shrub encroachment on herbage production and nutritive value in semi-arid Mediterranean grasslands. Grass Forage Sci. 62, 355–363. doi: 10.1111/j.1365-2494.2007.00590.x
Zavaleta, E. S., Pasari, J. R., Hulvey, K. B., and Tilman, G. D. (2010). Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proc. Natl. Acad. Sci. U.S.A. 107, 1443–1446.
Zhang, R., Wang, Z., Han, G., Schellenberg, M. P., Wu, Q., and Gu, C. (2018). Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the Desert Steppe, Northern China. Agric. Ecosyst. Environ. 265, 73–83. doi: 10.1016/j.agee.2018.05.014
Keywords: abandonment, Cantabrian Mountains, ecosystem multifunctionality, grazing exclusion, livestock, mountain grassland, sheep
Citation: Fernández-Guisuraga JM, Fernández-García V, Tárrega R, Marcos E, Valbuena L, Pinto R, Monte P, Beltrán D, Huerta S and Calvo L (2022) Transhumant Sheep Grazing Enhances Ecosystem Multifunctionality in Productive Mountain Grasslands: A Case Study in the Cantabrian Mountains. Front. Ecol. Evol. 10:861611. doi: 10.3389/fevo.2022.861611
Received: 24 January 2022; Accepted: 21 March 2022;
Published: 12 April 2022.
Edited by:
Andres Cibils, New Mexico State University, United StatesReviewed by:
Lynn Huntsinger, University of California, Berkeley, United StatesCopyright © 2022 Fernández-Guisuraga, Fernández-García, Tárrega, Marcos, Valbuena, Pinto, Monte, Beltrán, Huerta and Calvo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Manuel Fernández-Guisuraga, am9mZWdAdW5pbGVvbi5lcw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.