
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Ecol. Evol., 31 May 2022
Sec. Biogeography and Macroecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.850600
This article is part of the Research TopicAdaptation of Invasive Species to Islands and the Puerto Rican Honey BeeView all 14 articles
 Jose Marcelino1*†
Jose Marcelino1*† Charles Braese2
Charles Braese2 Krisztina Christmon3
Krisztina Christmon3 Jay D. Evans4Todd Gilligan5
Jay D. Evans4Todd Gilligan5 Tugrul Giray6Anthony Nearman3
Tugrul Giray6Anthony Nearman3 Elina L. Niño7Robyn Rose8Walter S. Sheppard9
Elina L. Niño7Robyn Rose8Walter S. Sheppard9 Dennis vanEngelsdorp3
Dennis vanEngelsdorp3 James D. Ellis1*†
James D. Ellis1*†Beekeeping is a cornerstone activity that has led to the human-mediated, global spread of western honey bees (Apis mellifera L.) outside their native range of Europe, western Asia, and Africa. The exportation/importation of honey bees (i.e., transfer of honey bees or germplasm between countries) is regulated at the national level in many countries. Honey bees were first imported into the United States in the early 1600’s. Today, honey bee movement (i.e., transport of honey bees among states and territories) is regulated within the United States at the state, territory, and federal levels. At the federal level, honey bees present in the country (in any state or territory) can be moved among states and territories without federal restriction, with the exception of movement to Hawaii. In contrast, regulations at the state and territory levels vary substantially, ranging from no additional regulations beyond those stipulated at the federal level, to strict regulations for the introduction of live colonies, packaged bees, or queens. This variability can lead to inconsistencies in the application of regulations regarding the movement of honey bees among states and territories. In November 2020, we convened a technical working group (TWG), composed of academic and USDA personnel, to review and summarize the (1) history of honey bee importation into/movement within the United States, (2) current regulations regarding honey bee movement and case studies on the application of those regulations, (3) benefits associated with moving honey bees within the United States, (4) risks associated with moving honey bees within the United States, and (5) risk mitigation strategies. This review will be helpful for developing standardized best practices for the safe movement of honey bees between the 48 contiguous states and other states/territories within the United States.
Honey bees, the most common pollinators across plant/pollinator networks worldwide (Hung et al., 2018), are crucial for human food production (Gallai et al., 2009). Western honey bees (Apis mellifera Linnaeus, 1758) are the only Apis species with a natural distribution that includes Europe, the Middle East, Africa, and parts of western Asia (Meixner et al., 2013). Beekeepers have transported honey bees around the world because they are easy to manage for production purposes (Crane, 1999). Apis mellifera now occurs on every continent except Antarctica (Hung et al., 2018) and is the dominant pollinator species used in agriculture (Williams, 1994; Aizen and Harder, 2009; Jordan et al., 2021). Globally, crop pollination and honey production services provided by managed honey bees are valued at $182–577 billion and $8 billion USD/year, respectively (Gallai et al., 2009; Lautenbach et al., 2012; Shahbandeh, 2021).
The human-mediated, global dispersal of western honey bees ultimately led to the development of rules and regulations created to mitigate potential negative outcomes associated with their dispersal. The exportation/importation of honey bees (i.e., transfer of honey bees between countries) is regulated at the national level in many countries. These measures range from outright bans on importation to the total absence of regulations. Regulatory authorities in many countries opt for structured permitting processes that, when met, allow the transportation of honey bees, or their germplasm, into a new area under strict protocols monitored by appropriate regulatory authorities. This occurs in the United States under the Honeybee Act, United States Code of Federal Regulation 7 CFR §281–§286 and §322, Subpart B—Importation of Adult Honeybees, Honeybee Germ Plasm, and Bees Other Than Honeybees From Approved Regions (Rules and Regulations, 2014). Regulations regarding the movement of other beneficial bees (i.e., bumble bees, Bombus spp.) are also included in this Federal Regulation. Currently, the importation of honey bees into areas where they do not exist is often prohibited worldwide, given concerns of their possible impact on native flora and fauna (Winter et al., 2006; Burgiel and Perrault, 2011).
Regulations concerning the transfer of live honey bees or germplasm within an area or political boundary in which they already occur (a process we term “movement” to distinguish from exportation/importation) vary within the United States. In the United States, the states and territories regulate the movement of live honey bees between themselves and other states/territories (Wehling and Flanders, 2005). At the federal level, honey bees already present in the United States (in any state or territory) are allowed to move among states and territories without federal restriction, with the exception of movement to Hawaii (see Code of Federal regulations 7 CFR Parts §319 and §322 at Rules and Regulations, 2014; Wehling and Flanders, 2005; Pernal, 2014). In contrast, regulations at the state and territory levels vary substantially, ranging from no additional regulations beyond the federal ones to strict regulations for the introduction of live colonies, packaged bees, or queens (Supplementary Table 1, Supplementary Figure 1 and interactive map1). This variability leads to the inconsistent application of regulations regarding the movement of honey bees among states and territories within the United States.
In November 2020, we convened a technical working group (TWG) formed by academic and USDA personnel, to review and summarize the current status of regulations regarding the human-mediated movement of honey bees within the United States. The TWG was composed of individuals (scientists/staff/graduate students) from the United States Department of Agriculture (USDA) Agriculture Research Service (ARS), the USDA Animal and Plant Health Inspection Service (APHIS), the USDA Farm Production and Conservation (FPAC) Business Center, and Land Grant Universities (see authors and acknowledgment sections). This manuscript represents the collective output of the TWG.
Herein, we review the (1) history of honey bee importation into and movement within the United States, (2) current regulations regarding honey bee movement and case studies on the application of those regulations, (3) benefits associated with moving honey bees within the United States, (4) risks associated with moving honey bees within the United States, and (5) risk mitigation strategies for bees moving within the United States. It is not our intention with this manuscript to suggest policies related to honey bee movement within the United States. We believe that is beyond the scope of this manuscript. Furthermore, we are not proposing a change in current policies regulating movement of honey bee colonies among states/territories under current management scenarios (i.e., for commercial pollination, honey production, or similar existing activities). Instead, we hope this review will be helpful for developing best practices for the safe movement of honey bees between the 48 contiguous states and other states/territories within the United States.
Honey bees are not native to the Americas and were imported into the United States as early as 1622 by English settlers (Supplementary Tables 2, 3; Sheppard, 1989a,b). The known imported stocks include A. m. mellifera Linnaeus, a subspecies with a natural distribution across northern Europe, and A. m. ligustica Spinola (Italian honey bee), introduced in the mid-1800’s and one of the most favored honey bee stocks (Sheppard, 1989a; Cobey et al., 2012). Other introductions occurred in the 19th century and included A. m. carnica Pollmann (Carniolan honey bee), A. m. caucasica Pollmann (Caucasian honey bee), A. m. lamarckii Cockerell (Egyptian honey bee), A. m. syriaca Skorikov (Syrian honey bee), A. m. cypria Pollmann (Cyprian honey bee), and A. m. intermissa Buttel-Reepen (a north African honey bee). See Supplementary Tables 2, 3 for more information.
Introductions of honey bee stocks into the United States over the past several decades are better documented and were done to benefit the beekeeping industry. Queen germplasm from the far-eastern Russian province of Primorsky was introduced into the United States after 7 months of quarantine and rigorous analyses of the biology, behavior, and pest resistance of queens and colonies from 1995 to 2000 (Danka et al., 1995; Rinderer et al., 1997, 1999, 2000, 2001a,b). The bees were reported as gentle, had high honey yields, and maintained low rates of infestation with the mite Varroa destructor (Anderson and Trueman, 2000; Rinderer et al., 2001a,b). Russian queen stock was made available to beekeepers in 2001 (Rinderer and Coy, 2020). Currently, a genetic stock certification protocol supports the Russian queen breeding industry (Bourgeois et al., 2020; Rinderer and Coy, 2020). However, this stock is not widely used by beekeepers (Haber et al., 2019).
Beekeepers readily moved various honey bee stocks throughout the United States after initial stock importation into the country (Supplementary Table 4). Furthermore, many colonies swarmed, producing feral colonies that established in many locations and environments. There were no efforts to slow the movement of introduced stock once it was present in the United States. The current United States feral and managed honey bee populations are a genetic mixture of the stocks initially imported into the United States and those subsequently introduced into the United States through the permitting process (Schiff and Sheppard, 1995).
The introduction of A. m. scutellata, a honey bee subspecies of African origin, into Brazil in 1956 greatly impacted the distribution of honey bees in the Americas (Kerr, 1967). Imported into Brazil to increase honey production, 26 colonies of A. m. scutellata escaped quarantine in 1957 and hybridized with other A. mellifera in the region. These hybrids (Africanized honey bees—AHBs) subsequently spread (e.g., natural dispersal without further human assistance) throughout South and Central America, the southwestern United States (Caron, 2001), and southern Florida (Hall, 1992). Hybridization is well documented across the Americas (Whitfield et al., 2006), and in the United States in particular (Calfee et al., 2020). Africanized honey bees exhibit heightened defensiveness and swarming/absconding behavior and are considered less desirable for commercial use by beekeepers in the United States. Furthermore, AHBs can outcompete and displace honey bees of European descent (EHBs) in areas where they co-occur (Breed et al., 2004; Schneider et al., 2004; Guzman-Novoa et al., 2020).
Africanized honey bee presence in an area does not always result in a negative outcome. This bee has documented resistance to V. destructor in Brazil (Carneiro et al., 2007) and in other areas (Strauss et al., 2015; Mondet et al., 2020). Africanized honey bees are generally the preferred honey bee among South and Central American beekeepers who value their honey production and pollination attributes (Roubik and Villanueva-Gutierrez, 2009). Africanized honey bees also occur in Puerto Rico (Puerto Rican Honey Bee or PRHB) where they are notably gentler than the Texas AHB population from which they are derived (Galindo-Cardona et al., 2013; Avalos et al., 2017, 2020; Acevedo-Gonzalez et al., 2019). This gentle demeanor has not been documented in other AHB populations in the Americas. Regulatory authorities in many states within the United States consider the AHB an “unwanted race or stock of honey bee,” and thereby prevent its movement into a given state, or its management within the state (Supplementary Table 1).
The possible accidental introduction of invasive honey bees and their pests/pathogens into the United States is a present and on-going threat (see section “Risks Associated With Moving Honey Bees Among United States and Territories”). APHIS personnel routinely inspect cargo and passengers entering the country for potential insect pest introductions through the Agricultural Quarantine Inspection (AQI) program. APHIS data on intercepts of individuals in the superfamily Apoidea (which includes bees and wasps) from air, maritime, and terrestrial arrivals between 2001 and 2021 show that honey bees (specifically, A. mellifera) compose the majority of interceptions and that most honey bee colonies and/or individuals arrive in the United States on plants and stored goods (Figure 1A). Furthermore, most of the intercepted honey bees arrive with materials originating from Central and South America. The volume of trade with nations in these regions may account for the number of honey bees intercepted from each country. Apart from Trypoxylon spp. (a crabronid wasp genus), most intercepted Apoidea arrived as live adults (Figure 1B). These data highlight the continued possibility of accidental or unlawful introductions of honey bees into the United States.
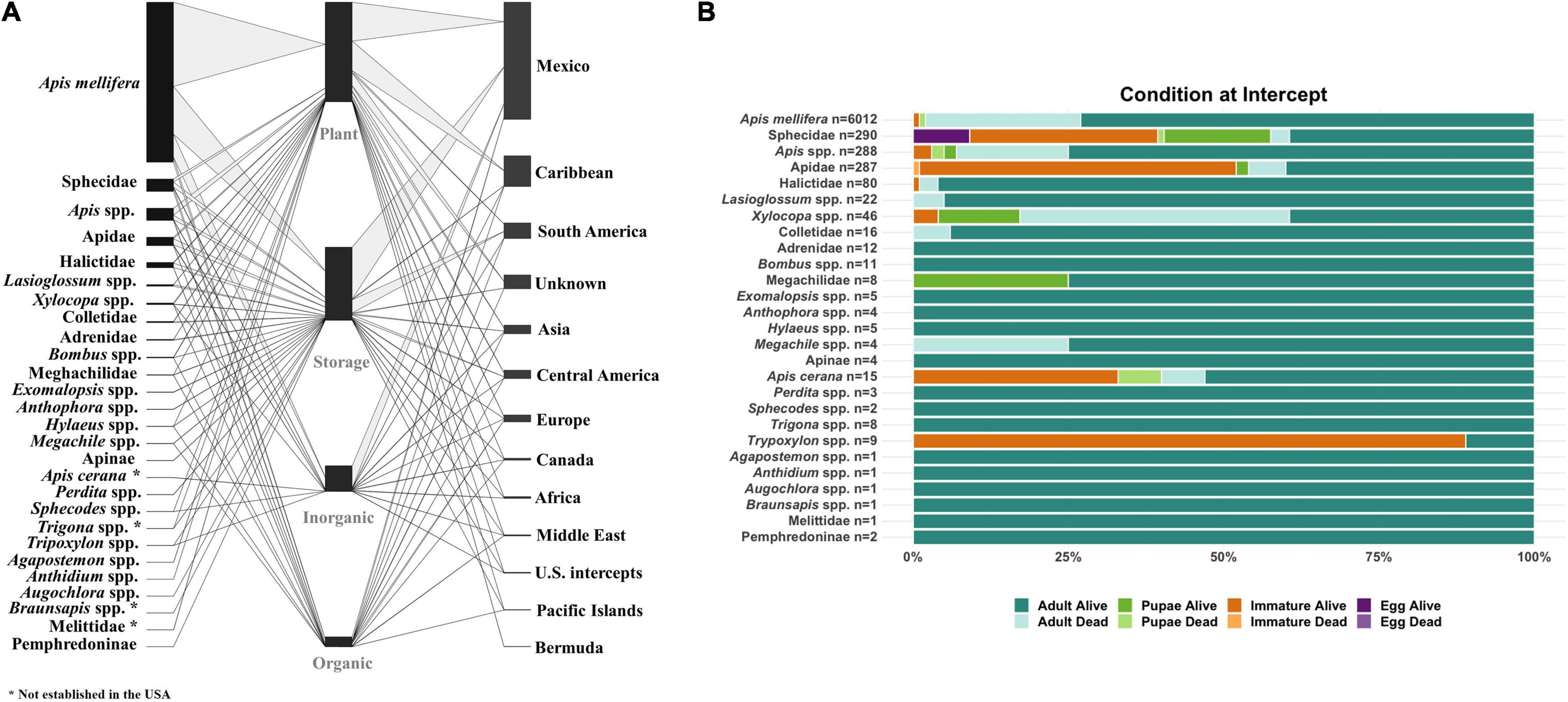
Figure 1. Apoidea intercepts at United States borders (2001–2021), adapted from APHIS’s Agricultural Quarantine Activity System (AQAS) data. (A) The bee or wasp species (left column) arrived on the material noted (middle column) from the region noted (right column). (B) The life stage and condition (dead or alive) of the individual that was found.
The United States Congress enacted the Honeybee Act (Federal Act) on 31 August 1922. Its purpose was to protect honey bees and the beekeeping industry from the tracheal mite, Acarapis woodi, the presumed cause of an unknown disease devastating honey bees in Europe at the time (Rennie, 1921). The Act has been promulgated multiple times (presently, 7 USCS §281-286 in effect on 3 January 2012). It has been broadened to regulate the importation of honey bee germplasm into the United States, to contain the spread of harmful diseases and parasites to honey bees within the country, and to eradicate/control undesirable species of Apis and subspecies of A. mellifera within the country and among imports (Rules and Regulations, 2014). Amendments in 1981 granted the Secretary of Agriculture the ability to approve bee-breeding stock and the release of bee germplasm. The Act now defines “honey bee” as all life stages and germplasm (with the exception of semen) of bees of the genus Apis (Rules and Regulations, 2014). As mentioned, there are no federal provisions to regulate honey bees currently present in the United States. This is the result of the perceived lack of risk associated with resident honey bee populations.
Regulations regarding interstate movement of honey bees differ at the state and territory levels within the United States (Supplementary Table 1). In general, honey bee movement is regulated by the issuance of permits from the receiving state based on certificates of bee health issued by the shipping state. Interstate movement ranges from no regulatory measures beyond those at the federal level to strict regulations for movement of live colonies, packaged bees, queens, or germplasm. This lack of uniformity has the potential to lead to the rapid spread of introduced pests, pathogens, parasites and/or deleterious honey bee subspecies across the United States, with repercussions for the beekeeping industry (Box 1).
Box 1. Case study on the spread of honey bee industry threats in the 1980s.
The lack of uniform policies and regulations among states and territories within the United States regarding honey bee movement (Supplementary Table 1 and Supplementary Figure 1) has failed to stop the spread of exotic pests and pathogens introduced to the mainland, with costly and irreversible impacts on the beekeeping industry in the United States. Here, we discuss the spread of honey bee pests and pathogens in the 1980s to illustrate this point, as significant new pests and pathogens established and spread in the United States during this decade.
The United States government passed the Honeybee Act in 1922 in response to the Isle of Wight bee disease in England (Nature, 1912). This act aimed to prevent the importation of honey bees from overseas. In addition, the spread and resulting impact of American foulbrood (bee disease caused by the bacterium Paenibacillus larvae) catalyzed the passage of state laws and the implementation of state inspection programs in the 1930s and 1940s (Surface, 1916; Phillips, 1920; Voorhies et al., 1933). In the United States, federal and state governments have distinct roles when protecting agriculture. The federal government, acting through APHIS (United States Department of Agriculture, Animal and Plant Health Inspection Service), attempts to prevent the introduction of honey bee threats into the country, helps develop technologies to detect and monitor potential or introduced threats, and works with state governments to contain newly identified introductions. However, state governments (via state departments of agriculture) have regulatory authority once a pest is considered established.
The 1980’s Annual Proceedings of the Apiary Inspectors of America reveals the complex and interacting factors that influenced the rate of spread within the United States of three honey bee pests during the 1980s (AIA, 1988d). These pests were honey bee tracheal mites (Acarapis woodi, HBTM, arrival date 1984), Varroa destructor (formerly V. jacobsoni, arrival date 1987) and Africanized honey bees (bees derived from A.m. scutellata, AHBs, arrival dates: feral colony intercept at FL ports in 1987 i.e., Panama City, Miami and Fort Lauderdale, followed by colonies found in Hidalgo, TX in 1990). See the timeline in this Box for more information.
For over 50 years since the passage of the Honey Bee Act (1922), the only biological threat to honey bee colonies in the United States was the fungal brood disease chalkbrood (Ascosphaera apis). It was first discovered in the United States in 1965, but was likely in the country prior to that time (Aronstein and Murray, 2010). At the same time, the predicted arrival of the AHB to southern states preoccupied lawmakers. The AHB quickly spread north throughout the Americas after its accidental release in Brazil in 1957 (Kerr, 1967). It is notorious for exhibiting heightened defensive responses, compared to those exhibited by A. mellifera of Eurasian descent managed by beekeepers in the United States. Identifying this undesirable stock was a difficult and time-consuming process. The inability to identify AHB stock efficiently and with confidence may have played some role in its dispersal. This was also true for bees infested by HBTMs. Proposals to certify stocks or bees as “HBTM free” largely failed because the mites were thought to be disseminated across the country already (AIA, 1987).
When state apiary laws were written, they were not designed to regulate the highly mobile apiary industry of the 1980s. Attempts to restrict the movement of bees, enforce quarantines, or await disease diagnosis were ineffective due to the economic and political demands of modern agriculture, i.e., the need to ship and/or truck bees, queens, and colonies across state lines to ensure the beekeeping industry’s viability and human food security through plant pollinated crops. Varroa destructor, when first detected on the continent (1987), was already in, or on its way to, several states, and its spread paralleled migratory operations coming out of Florida (ADCP, 1987a). A year after the mites were detected in Florida, they were found in most of the contiguous states (AIA, 1988d). Varroa destructor is now ubiquitous across the United States except for the territory of American Samoa and some of the islands of Hawaii, i.e., Maui, Kaua’i, Moloka’i, and Lâna’I (Rusert et al., 2021).
Timeline
Notable Honey Bee Threat Introductions, Response, and Spread in the United States Over the 1980’s
1979
• Varroa destructor scare, false positive in Maryland (AIA, 1981a).
• Varroa destructor response plan developed (AIA, 1981b).
1980
• United States Bee and Honey Act (1980) amended to include regulations to control the importation and interstate movement of diseases and other pests of honey bees (AIA, 1983).
• USDA initiates mite survey in Mexico, discovering the honey bee tracheal mite (Acarapis woodi, HBTM, Eischen et al., 1990).
1984
• HBTM first detected in Texas, and subsequently in nine states (AIA, 1985a).
• Florida rescinds all regulations on HBTM (AIA, 1985a). Regulators conclude HBTM cannot be eradicated.
• Consensus that it is best to contain HBTM until control measures are established, but now the pest is widespread, regulations controlling spread may cause more economic hardship than they would alleviate (AIA, 1985c).
• APHIS-PPQ recommends federal laws concerning HBTM be rescinded (AIA, 1985b).
• Nationwide HBTM survey discovering mites only in Florida (AIA, 1985c).
1985
• APHIS deregulates the HBTM (AIA, 1987). They are assumed widespread as migratory beekeepers and package producers continue to ship from Florida (AIA, 1987).
1986
• Each state implements its own protocols for HBTM detection and permitting for the shipment of packages, queens, and movement of colonies (AIA, 1986a).
• Florida queen and package industry “eliminated” by quarantine procedures by other states, halting movement of bees for pollination services and honey production in northern states (AIA, 1986b). (Continued)
1987
• Varroa destructor found in Wisconsin, from packaged bees from Florida (ADCP, 1987a).
• Varroa destructor in New York State, in migratory beekeeping from Florida (ADCP, 1987a).
• Concluded that restricting migratory movement is impossible to enforce (ADCP, 1987b).
• Canadian border closed to United States queen and package imports, devastating the queen and package industry in the United States (AIA, 1988b).
• Africanized honey bee intercepted at Florida ports (Hall, 1992).
1988
• USDA Action Plans-Varroa and Africanized Bees (AIA, 1988c).
1989
• Reaction to USDA action plan largely negative and meets resistance as too restrictive, or not restrictive enough, by different states (AIA, 1988e).
1990
• Feral Africanized honey bee colonies found in mainland United States, at Hidalgo, TX (Visscher et al., 1997).
• Varroa destructor arrives in Canada (AIA, 1990).
There are multiple potential benefits to moving honey bees among states and territories within the United States (Russo, 2016). First, migratory beekeeping (i.e., the movement of managed honey bee colonies by beekeepers) has direct value to agriculture because the pollination services provided by bees improves fruit set, crop yield, and quality. Second, moving bees provides direct economic benefit to beekeepers who transport their bees to areas where honey is produced and/or where they are paid to use their colonies to provide crop pollination services. Third, honey bees provide ecological value through the ecosystem service of pollination. Fourth, moving bees supports stock improvement initiatives that require an influx of new genetic material from which to select. Fifth, moving bees can compensate for colony losses at a given location. We provide context for each benefit below, demonstrating that moving bees is necessary within the United States to support a healthy economy, food supply, and beekeeping industry.
The United States agricultural sector is dependent on insect-mediated pollination for crop yield and/or quality (Jordan et al., 2021). Migratory beekeeping ensures that crop pollination demands are met (Rucker et al., 2012; Ferrier et al., 2018). In the last 50 years, the global demand for pollinator-dependent crops has increased by 300% (Aizen and Harder, 2009; Jordan et al., 2021). The estimated pollination services provided by honey bees to crop production in the United States averages $12–50 billion USD/year (Bauer and Wing, 2010; Calderone, 2012). To preempt pollination deficits, beekeepers move colonies to blooming crops that rely on insect-mediated pollination for fruit, vegetable, and nut production. Those colonies may originate from outside a state if an insufficient number of colonies exists within the state to provide the pollination services needed by the crops grown in that state.
Almond production in California requires a greater number of pollinating colonies than does any other single crop in the United States (Bond et al., 2021). Honey bee pollination of almonds contributed ∼$9.2 billion USD to the California’s gross state product (GSP) in the 2017/18 crop year (Matthews et al., 2020), and an estimated direct value of $6.09 billion USD in 2020 (Sumner et al., 2014; NASS, 2020). These are not marginal values; yet they account for a fraction of all marketable services and service fees that honey bees provide through migratory beekeeping. Almonds bloom in California in February, making this crop the starting point of an annual migratory route for many commercial beekeepers (Figure 2). Approximately 1.8 million managed honey bee colonies (∼60–75% of all United States commercial colonies) are moved to California each year to provide pollination services for almonds (Perez and Plattner, 2014; Goodrich et al., 2019).

Figure 2. Standard movement of honey bee colonies in migratory beekeeping operations in 1987 (A) and 2013 (B). Map lines represent the major movement patterns of honey bee colonies, rather than smaller movements that typically occur within state or contiguous states. Significant movement begins in February (east → west), triggered by California’s almond blooming season. Following this, movement occurs mid-March to May (west → northeast) to pollinate apple, pear and cherry orchards, among other crops, and to produce clover honey. A portion of these colonies will be moved to summer locations (northeast → northern plains) to pollinate other crops. During the summer months of June and July, colonies are moved (west coast + south-central → east coast + north) to pollinate apples, cherries, cranberries and vegetables. From mid-August to mid-October, major colony movement occurs (north + northeast →south) for overwintering in warmer latitudes (Rucker et al., 2012; Jabr, 2013; VPPC, 2016; Bond et al., 2021).
From almonds, beekeepers move these colonies across the country to provide pollination services for additional crops or to major nectar flows around the United States for honey production purposes. Additional colony movements occur during the fall season to allocate hives to warmer latitudes (e.g., Texas, Florida, etc.) for overwintering (Rucker et al., 2012; Jabr, 2013; VPPC, 2016; Bond et al., 2021). Only a subset of beekeepers are migratory. Many provide pollination services only within their region or state.
The movement of honey bees results in direct economic benefit to beekeepers. The revenue generated by providing honey bees for pollination services [rental fees ranging from $70–210 USD per colony, depending on the crop (USDA, 2020)] was estimated at $254.3 million USD in 2020 (NASS, 2021). This value was close to the $299.6 million USD generated from the production of honey in 2020 (NASS, 2021). Wax, pollen, queen, package, and nucleus (nuc) production represent additional value-added products originating from the hive and all generate income for beekeepers.
Non-marketable ecosystem services are provided via the movement of honey bees. These include the improvement of ecosystem function and stability (Losey and Vaughan, 2006). Honey bees are the most frequent visitor of non-crop plants worldwide, being present in about 89% of plant pollination networks (Hung et al., 2018). As a generalist species, honey bees visit a wide variety of flowering plants, while more specialized pollinators only visit a few species or families. Abundant pollination of native and wild vegetation increases the availability of plant resources which, in turn, benefit surrounding wildlife through the production of seeds, berries, nuts, and fruits (Pilati and Prestamburgo, 2016; Klein et al., 2017). In addition, honey bees are often used as bioindicators as they are exposed to trace contaminants such as pesticides and airborne heavy metals derived from anthropogenic industrial activities while foraging (Conti and Botrè, 2001; Van der Steen et al., 2011; Kennedy et al., 2013; Pilati and Prestamburgo, 2016; Goretti et al., 2020; Traynor et al., 2021). These contaminants can be traced in nectar/honey, wax, propolis and pollen samples, and can be used as indicators of environmental pollution for a given location (Conti and Botrè, 2001; Finger et al., 2014).
The movement of honey bees supports stock improvement programs that advance disease resistance and productivity in a managed stock (Spivak and Reuter, 2001; Traniello et al., 2002; Sadd et al., 2005; Blacquière and Panziera, 2018). Queen breeders and researchers monitor colonies for vigor and health. In response, they can select for traits such as productivity, gentleness and resistance to pests and pathogens (Ferrier et al., 2018). The movement of the selected stocks among states allows beekeepers in other states to incorporate the stocks into their beekeeping operations. As an example, scientists and queen breeders inseminated breeder queens in Hilo, Hawaii with drone germplasm from the USDA Varroa sensitive hygiene (VSH) stock (Harbo and Harris, 1999a,b; Ibrahim and Spivak, 2006). Movement of queens from this new stock, the Hilo bee, to the mainland United States is not restricted, but movement of honey bees to Hawaii is regulated.
Disease/pest resistant strains of honey bees can develop as a result of natural selection, making the movement of these bees of potential value to beekeepers. As an example, research suggests that the PRHB provides an opportunity to improve disease and pest resistance in the managed honey bee population in the United States (Rivera-Marchand et al., 2012; Avalos et al., 2017) by their inclusion in selective breeding programs.
Bee movement allows for the replacement of dead colonies. The individual, additive, and synergistic effects of colony stressors are responsible for annual gross colony loss rates averaging 45.5% (Steinhauer et al., 2020; BIP, 2021). Beekeepers mitigate these losses by splitting and/or purchasing colonies or queens of a preferred stock.
There are inherent risks associated with the movement of honey bees among states and territories within the United States. These include the dissemination of honey bee pathogens and pests, spread of resistance genes in a pest/pathogen population, trait dissemination or loss, impacts on other local flora/fauna, and others. It is important to recognize the risks associated with moving honey bees so that appropriate risk mitigation strategies can be developed.
The movement of a honey bee subspecies or stock risks introducing pests and pathogens that may impact the resident honey bee population (Ji et al., 2003; Moritz et al., 2005; Neumann, 2006; Cavigli et al., 2016; Owen, 2017). The reciprocal is likewise possible, with the introduced stock being naïve to a pest/pathogen endemic in an area, possibly being highly susceptible to it. The United States honey bee population harbors many of the major pests and pathogens known to impact honey bee colonies (Boncristiani et al., 2021), but their distribution and levels differ among states and territories within the United States. Puerto Rico and Hawaii can be used as examples of this difference. Deformed Wing Virus-A (DWV-A) and Acute Bee Paralysis Virus (ABPV) are the most prevalent honey bee viruses in Puerto Rico (APHIS, 2018), while Chronic Bee Paralysis Virus (CBPV), Israeli Acute Paralysis Virus (IAPV), and Slow Bee Paralysis Virus (SBPV) have not been detected there (APHIS, 2018). The movement of honey bees between the continental United States and Puerto Rico could result in the spread of these viruses from the continent to managed honey bees on the island (Grozinger and Flenniken, 2019). In Hawaii, V. destructor occurs in honey bee colonies on some islands but not on others (Ramadan et al., 2019; Beaurepaire et al., 2020; Boncristiani et al., 2021; Rusert et al., 2021). Moving bees among islands could spread V. destructor to islands where it does not occur.
Some honey bee stocks may harbor new pests/pathogens not broadly distributed in the United States. For example, an in-depth genetic analysis of United States mainland and Hawaiian honey bee samples collected during the 2015 APHIS National Honey Bee Pest and Diseases Survey found new viruses not linked to any reported clinical signs (Ray et al., 2020). The detection of these new viruses raises concerns that other undetected pests and pathogens exist, leading to their possible transmission when honey bee colonies are moved.
Honey bee movement may lead to the future dissemination of organisms likely to prove harmful to the beekeeping industry (see Box 1). These include Tropilaelaps spp., a genus of mites with members that are pests of honey bees in Asia (Anderson and Morgan, 2007), and Nosema neumanni, a new species of Nosema recently discovered in Africa (Chemurot et al., 2017). This list also includes A. mellifera species of concern, such as A. m. capensis (the Cape honey bee), known to be social parasite of managed honey bee colonies in South Africa (Neumann and Moritz, 2002).
Honey bee movement can result in the spread of resistance genes in pest/pathogen populations. As an example, some populations of V. destructor have documented resistance to amitraz, fluvalinate, and coumaphos in different areas within the United States (Kast et al., 2020; Rinkevich, 2020; Millán-Leiva et al., 2021). Correspondingly, moving honey bees that host miticide-resistant V. destructor could spread the resistance trait to miticide-susceptible V. destructor populations (Benito-Murcia et al., 2021).
The movement of honey bees could lead to the spread of negative traits from the introduced population to the resident managed population and vice versa. For example, introduced AHBs express heightened defensive behavior, absconding tendencies, etc. and these can be incorporated, via hybridization, into the managed honey bee population in areas where AHBs spread (Caron, 2001; Schneider et al., 2004).
Bee movement can impact heritable traits in other ways as well. A selected stock that exhibits beneficial traits can lose those traits when moved into a new area. This occurs because of open mating, i.e., newly produced queens leaving their hives to mate with multiple drones from different colonies among the local population (Koeniger et al., 2014). As an example, selected breeding stock could lose some of its desirable traits, e.g., gentleness and resistance to V. destructor, when moved into an area with established honey bee colonies. This may occur even when the original intent of the proposed move was to introduce selected traits into resident managed populations.
Honey bees (resident or introduced) may lose or lack adaptations that improve their health, productivity, and survival in a specific area. Multiple research teams have demonstrated that honey bees in a selection program can fail to perform as expected when moved to a different environment with different climatic conditions (Costa et al., 2012; Hatjina et al., 2014; Kovaèiæ et al., 2020). For example, a pan-European evaluation on behavioral traits across 16 genotypes of five honey bee subspecies in various environmental conditions across Europe showed that genotype and location can impact traits such as defensiveness and swarming behavior (Uzunov et al., 2014). The widespread movement of a large percentage of the managed honey bee population in the United States may limit the development of locally adapted honey bee stocks.
Honey bee colonies, shipped packages, and bees may harbor other arthropods, pests, and pathogens that can impact local bees, other animals, and plant communities (Graystock et al., 2016). Furthermore, plant communities can change when honey bees are moved into an area. This occurs when honey bees pollinate a variety of flowering species, resulting in increased weed species (Goulson and Derwent, 2004) or reduced ranges of range-restricted plants (Norfolk et al., 2018). Moved honey bees may also impact local pollinator communities through competition and displacement (Torné-Noguera et al., 2016; Cane and Tepedino, 2017). The placement of honey bee colonies in high densities at a given location reduces the connectedness of plant-pollinator networks, and in some cases may lead to a decline in ecosystem resilience (Goulson, 2003; Guimarães, Jordano and Thompson, 2011; Geslin et al., 2017; Valido et al., 2019). Additionally, there is evidence for population-level changes with native bee and flora decline in areas where honey bees are the dominant pollinator. This results in the homogenization of honey bee pollinated fauna (Thomson, 2004, 2016; Paini and Roberts, 2005; McKinney and La Sorte, 2007; Balfour et al., 2015; Goulson et al., 2015; Guzman-Novoa et al., 2020).
It is noteworthy that most of these studies document the potential for large-scale impacts on resident native pollinator populations from introduced non-native bees such as honey bees, and not empirical evidence of direct effects on native bee fitness, abundance, and diversity in response to honey bee competition. Usually, negative impacts are predicated on the assumption that natural communities are at capacity and all plants are pollinated efficiently. However, this is not always the case as has been shown in neotropical communities (Ackerman, 2021). Investigators, through long-term studies on the impact of AHBs on native solitary bees in Central and South America, failed to identify impacts of AHB introductions on native orchid bees. In fact, they found that orchid bee populations increased in numbers after AHBs became established. This finding may be explained by increased foraging resources resulting from pollinator services provided by honey bees (Roubik, 2009; Roubik and Villanueva-Gutierrez, 2009). If honey bees do negatively impact non-Apis bee communities in the United States, it is likely to be concentrated around apiaries, and the roadways that give beekeepers access to them, and influenced by local environmental characteristics. The magnitude of honey bee impacts on native pollinators and plant communities depends on local parameters such as foraging bloom density and patchiness, distance from forage, hive density and host specificity of the native pollinators (Cane and Tepedino, 2017; Mallinger et al., 2017).
Every year, beekeepers move more than 1.8 million honey bee colonies around the United States to provide good forage sources for their bees, manage them in more favorable climates/environments, and gain profit by making honey and/or providing pollination services (Ferrier et al., 2018; Goodrich et al., 2019; Bond et al., 2021). There is always a risk of accidents when moving bees on large trucks that typically hold ca. 400 colonies. An overturned trailer may contain 1.2 million or more bees and they pose significant risk to people involved in the accident, those passing it, and the first responders addressing it. Sensationalized press coverage of such events, including AHBs or “killer” bees as often called by the press, can harm the reputation of the industry, and may make the public more cautious about bees and beekeepers.
Transporting bees can harm them directly. Repetitive relocation of bees during migratory beekeeping seasons can induce oxidative stress, lead to overheating, and decrease the lifespan of a colony (Ahn et al., 2012; Simone-Finstrom et al., 2016). Multiple relocations, and travel itself, also leads to higher exposure to pesticides and other agrochemicals, which can affect colony survival (Mullin et al., 2010; vanEngelsdorp and Meixner, 2010; Doublet et al., 2015). In addition, high colony densities on moving trucks can lead to pest and pathogen transmission and intensify disease outbreaks. A stress response is also triggered by the need for foraging bees to reassess their changing environment continuously (Simone-Finstrom et al., 2016). vanEngelsdorp et al. (2012) inspected colonies in three migratory beekeeping operations in the eastern United States, quantifying survivorship and prevalence of pests and pathogens. The team identified two risk factors that were predictive of colony mortality in migratory operations, idiopathic brood disease syndrome (IBDS), where brood of different ages appears molten on the bottom of their cells, and queen replacement or failure. In addition, Zhu et al. (2014) reported that transportation increases the abundance and prevalence of N. ceranae in honey bees. Moving bees to better forage may counter these effects and ultimately improve the growth and survival of colonies.
There are means to mitigate the risks associated with moving honey bees. A successful risk mitigation approach integrates multiple risk reduction strategies. Here, we review risk mitigation strategies associated with bee movement and discuss their feasibility of adoption, which may depend on individual state regulations already in place (Supplementary Table 1).
An important risk mitigation strategy could include screening the honey bee stock/life stage (i.e., adult bees, immature bees, and germplasm) proposed for movement between the contiguous 48 states and outlying states/territories for pests and pathogens they may harbor, or undesirable behavioral traits they may display. The APHIS National Honey Bee Pests and Diseases Survey has expanded to screen for additional agents of concern (Traynor et al., 2016; Fahey et al., 2018, 2019; Ray et al., 2020). These types of surveys can target the detection of specific pathogens or pests present in the country (e.g., V. destructor, Nosema spp., etc.), or of major concern if they were introduced (e.g., Tropilaelaps spp., N. neumanni, SBPV).
High-throughput sequencing and additional molecular technologies can detect viruses and other microbes infecting honey bees (Cox-Foster et al., 2007). Novel and inexpensive metagenomic screening tools exist and are used to detect viruses in bee communities (Galbraith et al., 2018). These are also used for detection of viruses in honey bees (Runckel et al., 2011; Granberg et al., 2013; Beaurepaire et al., 2020). Environmental DNA (eDNA) has been used to screen for pathogens and pests in honey (Ribani et al., 2020), while shotgun sequencing of honey can identify traces of organisms that bees encounter while foraging (Bovo et al., 2020).
There are also techniques for screening for unwanted species/subspecies of honey bees, though they vary in degree of accuracy. For example, the African honey bee, A. m. scutellata, and its hybrids can be identified using a reduced set of single nucleotide polymorphisms (SNPs), a real-time qPCR assay, or combinations of morphological features (Pinto et al., 2014; Harpur et al., 2015; Munoz et al., 2015; Eimanifar et al., 2018, 2020; Boardman et al., 2021; Momeni et al., 2021). Geo-morphometric analyses of honey bee wings coupled with SNP data (Calfee et al., 2020; Henriques et al., 2020), or geo-morphometrics alone (Nawrocka et al., 2018; Bustamante et al., 2020) have been used to identify A. m. scutellata populations as well. However, there are problems with relying on the output from some of these screening techniques. For example, morphometric methods can fail to assign hybrid populations accurately (Guzman-Novoa et al., 1994). Furthermore, methods that rely on mitochondrial DNA are limited as they do not account for paternal contributions (Meusel and Moritz, 1993).
Colonies can also be screened for visible signs of infection/infestation by pathogens/pests and for the presence of undesirable traits using standard protocols (e.g., Shimanuki and Knox, 2000; Dietemann et al., 2013; Spivak and Reuter, 2016). Some behavioral traits could be screened for using molecular technologies (Avalos et al., 2020) or in the field (Giray et al., 2000; Avalos et al., 2014). There are caveats to these trait screening tools when genes governing a phenotype are not yet known, making a molecular screen impossible, or when a phenotype does not manifest in colony-based assays, as in the case of heightened absconding behavior.
A comprehensive monitoring program that profiled the microbiome and behavioral traits of honey bee stocks proposed for movement would be beneficial on a regional/country wide level. These activities could be considered as an integral component of the decision-making process for the inter-regional movement of honey bee stock to a new area.
Phased movement includes the movement of a honey bee stock into an area after being quarantined. Phased imports serve as an example of how phased movement within the United States could be implemented. APHIS developed procedures for phased imports to mitigate the spread of pests or pathogens from foreign stock to the United States. Similar procedures could be implemented when proposing the movement of honey bees from United States island territories to the mainland and vice versa. Here, we provide two examples of phased imports, the first for honey bee queens and the second for germplasm.
USDA scientists implemented a phased import strategy for the Russian honey bee (or Primorsky bee). Briefly, they imported 100 Russian honey bee queens and quarantined them on Grand Terre Island, USDA-ARS Honey Bee Quarantine Station, Louisiana, United States in 1997. The scientists re-queened colonies of local stock with the Russian queens and monitored the colonies for pest and disease resistance, behavior, offspring vigor and resistance to V. destructor. The quarantine lasted seven months until the new stock was certified free of notifiable pests and pathogens (Rinderer et al., 1999). A Russian queen commercialization and stock certification protocol is now established in the United States via the Russian Honey Bee Breeders Association (Bourgeois et al., 2020; Rinderer and Coy, 2020).
Phased movements of germplasm can follow the strategy implemented for phased imports of germplasm, which include the importation of germplasm into a quarantined stock before its release. Sheppard (2012) documented the importation of honey bee semen from managed stocks of A. m. ligustica (from Bologna, Italy, 2008, 2009), A. m. carnica (from Kirchhain, Germany, 2008, 2009) and A. m. caucasica (from the Tblisi and other locations, Republic of Georgia, 2010) to quarantine sites located in the wheat growing Palouse region of eastern Washington (Bald Butte and Smoot Hill, Whitman County). The imported semen was used to inseminate unmated queens for stock improvement purposes in the United States (Sheppard, 2012). The wheat fields consisted of several hundred hectares with restricted access and were surrounded by hectares of crops that do not provide resources to honey bees (e.g., lentils, barley, etc.). This experimental plan minimized interactions between the research stock and colonies managed by beekeepers in the area. The semen was screened for viruses prior to its use for the insemination of clipped queens. The queens were maintained in nucleus colonies under quarantine until approval was received from APHIS for their release.
Another mitigation strategy involves regulating interstate/territory movement following a standard protocol, similar to the one used at the national level for stock importation (Regulation 7 C.F.R. §322 at Rules and Regulations, 2014). In this case, bee movement is permitted when the honey bee stock or germplasm is accompanied by a permit or certificate that states the bees are free of pests, pathogens, and/or harmful phenotypes. This would trigger federal and state reviews of the risks and benefits of moving the bees. Such protocols exist but are not applied consistently across states (Supplementary Table 1).
Admittedly, regulating movement at the state level through inspections, certifications, and permits can be arduous and time-consuming. However, a vetted and implemented permitting process, triggered by notifying APHIS personnel with a request to move bees, could remove some of the inconsistencies between state and territory regulations regarding the movement of honey bees (Mailander and Grant, 2019). The adoption of a standardized permitting process across all states and territories may translate into a more uniform and judicious regulation of bee stock movement.
The movement of germplasm, rather than actual bees, can mitigate some risks associated with moving honey bees. Safeguards can be implemented to ensure pest-free germplasm. The movement of drone semen, for example, does not carry the risk of moving Varroa spp., Tropilaelaps spp., or other arthropod pests. Semen can be screened for pathogens using molecular tools and discarded at low cost if pathogens are discovered (Sheppard, 2012). Furthermore, germplasm is easier to transport, does not carry the risk of bee escape, and can be introduced into resident populations of honey bees through instrumental insemination (Hopkins and Herr, 2010; Hopkins et al., 2012; Sheppard, 2012). Due to these advantages, it was the protocol chosen for the introduction of Russian honey bees, A. m. ligustica, A. m. carnica, and A. m. caucasica germplasm into the United States (Sheppard, 2012). This method was also used for the Hilo and Pol-line stocks kept in Hawaii in closed mating systems for queen releases to commercial beekeepers (Danka et al., 2016; Bilodeau et al., 2020).
A stock certification program can include multiple risk mitigation strategies. First, a stock can be defined per its attributes, such as possessing a selected trait (i.e., Varroa Sensitive Hygiene) or combination of traits that make is resistant to a pest or pathogen. The latter happened in the case of the Russian honey bee, with its origin and general resistance to V. destructor being integral to the stock’s defined attributes (Bourgeois et al., 2020). Second, a certification program can include routine screening for pests, pathogens, and undesirable traits. If the stock is derived from an “unwanted race or subspecies” of honey bee (language often used at the state regulatory level), a demonstration that the stock is free of the “unwanted” trait could be included as part of the stock’s definition.
It is important to demonstrate that the genetic stock will not re-acquire, or revert to, undesirable phenotypes. It is also necessary to develop a maintenance plan for the stock. This could include a strategy to maintain and monitor the stock’s purity, along with a demonstration that it is not prone to broad geographic dissemination and competition with resident populations. The classic example of a failure to contain invasive bees is the introduction of the AHB into Brazil in 1956 (Kerr, 1967). Multiple AHB queens escaped from research colonies before adequate safeguards were put in place, leading to the establishment of one of the most successful invasive insects in the Americas (Smith, 1991; Visscher et al., 1997; Caron, 2001; Moritz et al., 2005).
Another risk mitigation strategy involves the prohibition of movement of honey bee stocks to areas they are not present. This represents the most extreme mitigation strategy and is employed only if the risk is so great that a negative outcome is almost ensured. It is necessary to ensure that a prohibition of this type would not affect the normal movement of honey bees for typical beekeeping purposes. In most cases, this mitigation strategy is probably not practical.
There are additional variables to consider when developing appropriate mitigation strategies. First, the level of mitigation can vary depending on what is being moved (i.e., queens, packages, colonies, germplasm, etc.). Second, certification requirements for a given move request would depend on its specific characteristics (e.g., island setting vs. mainland setting, variable regulatory frameworks between states, etc.). Third, a testing framework and appropriate certification process would be required for genetic, pest, and pathogen testing that ascribes to specific standards, such as the Good Laboratory Practices (GLP). Adherence to accepted standards could be demonstrated by private and federal laboratories that may conduct such tests. Fourth, it is important to consider the regulatory costs for monitoring movement. Consistency on movement policies and risk mitigation strategies at federal and state levels is needed for a country-wide regulatory framework to function in an efficient manner. The serious consideration of the issues presented in this manuscript can serve as the basis upon which to build a framework that will lead to a fair, economic, equitable and beneficial process for moving honey bees between states and territories within the United States, especially between the 48 contiguous states and the outlying states/territories.
Honey bees are vitally important to agriculture in the United States, ensuring a significant portion of the nation’s food supply through the pollination services they provide. Beekeepers move colonies to provide pollination services, produce honey, recover colony losses, improve stocks via new genetic material, and recuperate costs. The movement of honey bees is a necessary and critical component of agriculture in the United States and it supports the beekeepers whose incomes rely on healthy bees. Although necessary, the movement of honey bees is not without risk. This risk has led state and territory officials to develop a variety of rules and regulations imposed to ensure honey bee health and sustainability of the beekeeping industry.
The issues associated with the regulatory process and regulations regarding the movement of honey bees among states and territories within the United States led to this review. We believe bee movement remains a necessary and critical component of agriculture and the beekeeping industry in the United States. The information we present herein can inform future efforts designed to create standardized best practices for moving honey bees among states and territories within the United States.
JM and JEl conceived and developed the review. JEv, ToG, TuG, EN, RR, WS, and DvE provided empirical knowledge and scientific expertise regarding the application of current state and federal regulations on bee movement, present status of the bee industry and beekeeping in states and territories within the United States, and risks and mitigation strategies associated with moving honey bees within the United States. KC, AN, and CB performed data analysis, figure preparation, and interactive map assembly. All authors contributed to the article and approved the submitted version.
This project was funded, in part, by the United States Department of Agriculture’s Animal and Plant Health Inspection Service (APHIS) in a cooperative agreement (AP20PPQS&T00C105; PI, JEl) to the University of Florida. Statements and conclusions in this manuscript should not be construed as official APHIS policy. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor RG is currently organizing a Research Topic with one of the authors TuG.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank the scientific input of the following collaborators at the USDA Animal and Plant Health Inspection Services (APHIS) during the execution of this project: Anne LeBrun (APHIS National Policy Manager for Pollinator Pest Programs), Josie Ryan (APHIS National Operations Manager for Pollinator Pest Programs), Colin Stewart (Assistant Director of Pests, Pathogens, and Biocontrol Permits Branch at APHIS Plant Protection and Quarantine), Wayne Wehling (APHIS-PPQ Plant Health Programs), and Allan H. Smith-Pardo (APHIS-PPQ, Science and Technology). We would also like to thank Gloria DeGrandi-Hoffman (Carl Hayden Bee Research Center, USDA-ARS) and Marla Spivak (Department of Entomology, University of Minnesota) for project input, as well as Andy Brower (APHIS National Identification Services Assistant Director) and Roger Burks (APHIS Marketing and Regulatory Programs) for access to data on Apoidea intercepts at United States borders. We also thank the Canadian Association of Professional Apiculturists (CAPA) for providing access to their historical archives.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.850600/full#supplementary-material
Acevedo-Gonzalez, J. P., Galindo-Cardona, A., Avalos, A., Whitfield, C. W., Rodriguez, D. M., Uribe-Rubio, J. L., et al. (2019). Colonization history and population differentiation of the Honey Bees (Apis mellifera L.) in Puerto Rico. Ecol. Evol. 9, 10895–10902. doi: 10.1002/ece3.5330
Ackerman, J. D. (2021). Island invasions by introduced honey bees: what can be expected for Puerto Rico and the Caribbean? Front. Ecol. Evol. 10:556744. doi: 10.3389/fevo.2020.556744
ADCP (1987a). Proceedings of Apiary Disease Control Program, New York State 1987. New York, NY: Apiary Disease Control Program, 5–7. Varroa mites in New York State.
ADCP (1987b). Proceedings of Apiary Disease Control Program, New York State 1987. New York, NY: Apiary Disease Control Program, 294–225. Apiary advisory committee recommendations: Tracheal mites.
Ahn, K., Xie, X., Riddle, J., Pettis, J., and Huang, Z. Y. (2012). Effects of long-distance transportation on honey bee physiology. Psyche 2012:193029. doi: 10.1007/s00359-002-0374-y
AIA (1981a). “Proceedings of the Annual Conference Apiary Inspectors of America 1981,” in Annual Conference Apiary Inspectors of America, ed. I. Barton Smith Jr. (Las Vegas, NV), 26–28. Varroa scare in Maryland.
AIA (1981b). “Proceedings of the Annual Conference Apiary Inspectors of America 1981, January 12-14,” in Recommendations to the beekeeping industry concerning the mite Varroa jacobsoni. Annual Conference Apiary Inspectors of America, Las Vegas, NV, 28–30.
AIA (1983). “Proceedings of the Annual Conference Apiary Inspectors of America 1983, January,” in Amendments to the United States Bee and Honey Act Annual Conference Apiary Apiary Inspectors of America Orlando, Florida, 46.
AIA (1985a). “Proceedings of the Annual Conference Apiary Inspectors of America 1985, January 21-25,” in Federal Survey and Control Relating to African Bee, Varroa and Acarapis Mites by Mr. Richard L. Cowden, Senior Staff Officer, National Program Planning Staff, USDA Annual Conference Apiary Inspectors of America San Antonio, Texas, 33–34.
AIA (1985b). “Proceedings of the Annual Conference Apiary Inspectors of America 1985, January 21-25,” in Current status of honey bee tracheal mite, USDA-APHIS. Annual Conference Apiary Inspectors of America, San Antonio, Texas, 35–37.
AIA (1985c). “Proceedings of the Annual Conference Apiary Inspectors of America 1985 January 21-25,” in Technical Advisory committee for Africanized Bees and Honey Bee Mites Survey results. Annual Conference Apiary Inspectors of America San Antonio, Texas, 38–42.
AIA (1986a). “Proceedings of the Annual Conference Apiary Inspectors of America 1986, January 27-30,” in Tracheal mite Certification of Honey bees Annual Conference Apiary Inspectors of America Baton Rouge, Louisiana, 48–49.
AIA (1986b). “Proceedings of the Annual Conference Apiary Inspectors of America 1986, January 27-30,” in Beekeeping in Florida by Laurence P. Cutts. Annual Conference Apiary Inspectors of America Baton Rouge, Louisiana, 66–70.
AIA (1987). “Proceedings of the Annual Conference Apiary Inspectors of America 1987, January 26-29,” in Politics of Tracheal Mites by Dr. H. Shiminuki, Laboratory Director USDA, Beltsville MD. Annual Conference Apiary Inspectors of America, Biloxi, MS, 19–21.
AIA (1988a). “Interstate migratory beekeeper and colony survey,” in Proceedings Annual Conference of Apiary Inspectors of America. 1988 October 10-14, Denver, CO, 49–58.
AIA (1988b). “Proceedings of the Annual Conference Apiary Inspectors of America 1988, January 18-21,” in CAPA report By Mr. Will Kissinger. Annual Conference Apiary Inspectors of America, Baltimore, MD, 47.
AIA (1988c). “Proceedings of the Annual Conference Apiary Inspectors of America 1988, January 18-21,” in USDA Action Plans-Varroa and Africanized bees Annual Conference Apiary Inspectors of America, Baltimore, MD, 24.
AIA (1988d). “Proceedings of Fall Annual Conference Apiary Inspectors of America 1988, October 10-14,” in AIA General session Proceedings Annual Conference of Apiary Inspectors of America, Baltimore, MD, 16.
AIA (1988e). “Proceedings of Fall Annual Conference Apiary Inspectors of America 1988, October 10-14,” in Summary of Proposed Varroa rules responses by AIA (May1989) Washington State Department of Agriculture Proceedings Annual Conference of Apiary Inspectors of America, Denver, CO, 69–80.
AIA (1990). “Proceedings of the Annual Conference Apiary Inspectors of America 1990, January 12,” in CAPA report By Doug McRory Annual Conference Apiary Inspectors of America, Schaumburg, IL, 26.
Aizen, M. A., and Harder, L. D. (2009). The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 19, 915–918. doi: 10.1016/j.cub.2009.03.071
Anderson, D. L., and Trueman, J. W. H. (2000). Varroa jacobsoni (Acari: Varroidae) is more than one species. Exp. App. Acarol. 24, 165–189. doi: 10.1023/a:1006456720416
Anderson, Dl, and Morgan, M. J. (2007). Genetic and morphological variation of bee-parasitic Tropilaelaps mites (Acari: Laelapidae): new and re-defined species. Exp. Appl. Acarol. 43, 1–24. doi: 10.1007/s10493-007-9103-0
APHIS (2018). State Report, Puerto Rico. Bee Informed Partnership. Available online at https://bip2.beeinformed.org/reports/state_reports/state_report/?year=2018&state=PR (accessed April 26, 2022).
Aronstein, K. A., and Murray, K. D. (2010). Chalkbrood disease in honey bees. J. Invertebr. Pathol. 103, S20–S29.
Avalos, A., Fang, M., Pan, H., Lluch, A. R., Lipka, A. E., Zhao, S. D., et al. (2020). Genomic regions influencing aggressive behavior in honey bees are defined by colony allele frequencies. Proc. Natl. Acad. Sci. U.S.A. 117, 17135–17141. doi: 10.1073/pnas.1922927117
Avalos, A., Rodriguez-Cruz, Y., and Giray, T. (2014). Individual responsiveness to shock and colony-level aggression in honey bees: evidence for a genetic component. Behav. Ecol. Sociobiol. 68, 761–771. doi: 10.1007/s00265-014-1689-8
Avalos, H., Pan, C., Li, J. P., Acevedo-Gonzalez, G., Rendon, C. J., Fields, P. J., et al. (2017). A soft selective sweep during rapid evolution of gentle behavior in an Africanized honey bee. Nat. Commun. 8:1550. doi: 10.1038/s41467-017-01800-0
Balfour, N. J., Gandy, S., and Ratnieks, F. L. W. (2015). Exploitative competition alters bee foraging and flower choice. Bahav. Ecol. Sociobiol. 69, 1731–1738. doi: 10.1007/s00442-019-04576-w
Bauer, D. M., and Wing, I. S. (2010). Economic consequences of pollinator declines: a synthesis. Agric. Resour. Econ. Rev. 39, 368–383. doi: 10.1017/s1068280500007371
Beaurepaire, A., Piot, N., Doublet, V., Antunez, K., Campbell, E., and Chantawannakul, P. (2020). Diversity and global distribution of viruses of the western honey bee. Apis mellifera. Insects 11:239. doi: 10.3390/insects11040239
Benito-Murcia, M., Bartolomé, C., Maside, X., Bernal, J., Bernal, J. L., del Nozal, M. J., et al. (2021). Residual tau-fluvalinate in honey bee colonies is coupled with evidence for selection for Varroa destructor resistance to pyrethroids. Insects 12:731. doi: 10.3390/insects12080731
Bilodeau, L., Avalos, A., and Danka, R. G. (2020). Genetic diversity of the complementary sex-determiner (csd) gene in two closed breeding stocks of Varroa-resistant honey bees. Apidologie 51, 1125–1132. doi: 10.1007/s13592-020-00790-1
BIP (2021). Total Winter All Colony. Bee Informed Partnership. Available online at: https://beeinformed.org/2021/06/21/united-states-honey-bee-colony-losses-2020-2021-preliminary-results/ (accessed April 26, 2022).
Blacquière, T., and Panziera, D. (2018). A plea for use of honey bees’ natural resilience in beekeeping. Bee World 95, 34–38. doi: 10.1080/0005772X.2018.1430999
Boardman, L., Srivastava, P., Jeyaprakash, A., Moore, M. R., Whilby, L., and Ellis, J. D. (2021). A qPCR assay for sensitive and rapid detection of African A-lineage honey bees (Apis mellifera). Apidologie 52, 767–781. doi: 10.1007/s13592-021-00863-9
Boncristiani, H., Ellis, J. D., Bustamante, T., Graham, J., Jack, C., and Kimmel, C. B. (2021). World honey bee health: the global distribution of western honey bee (Apis mellifera L.) pests and pathogens. Bee World 98, 2–6. doi: 10.1080/0005772x.2020.1800330
Bond, J. K., Hitaj, C., Smith, D., Hunt, K., Perez, A., and Ferreira, G. (2021). Honey Bees on the Move: From Pollination to Honey Production and Back. ERR-290. Washington, DC: U.S. Department of Agriculture, Economic Research Service.
Bourgeois, L., Sheppard, W. S., Sylvester, H. A., and Rinderer, T. E. (2020). Genetic stock identification of russian honey bees. J. Econ. Entomol. 103, 917–924. doi: 10.1603/ec09335
Bovo, S., Utzeri, V. J., Ribani, A., Cabbri, R., and Fontanesi, L. (2020). Shotgun sequencing of honey DNA can describe honey bee derived environmental signatures and the honey bee hologenome complexity. Sci. Rep. 10:9279. doi: 10.1038/s41598-020-66127-1
Breed, M. D., Guzmán-Novoa, E., and Hunt, G. J. (2004). Defensive behavior of honey bees: organization, genetics, and comparisons with other bees. Ann. Rev. Entomol 49, 271–298. doi: 10.1146/annurev.ento.49.061802.123155
Burgiel, S. W., and Perrault, A. M. (2011). “Black, white, and gray lists,” in Encyclopedia Of Biological Invasions, eds M. Rejmánek and D. Simberloff (Berkeley: University of California Press), 75–77. doi: 10.1525/9780520948433-021
Bustamante, T., Baiser, B., and Ellis, J. D. (2020). Comparing classical and geometric morphometric methods to discriminate between the South African honey bee subspecies Apis mellifera scutellata and Apis mellifera capensis (Hymenoptera: Apidae). Apidologie 51, 123–136. doi: 10.1007/s13592-019-00651-6
Calderone, N. (2012). Insect pollinated crops, insect pollinators and US agriculture: trend analysis of aggregate data for the period 1992–2009. PLoS One 7:e37235. doi: 10.1371/journal.pone.0037235
Calfee, E., Agra, M. N., Palacio, M. A., Ramirez, S. R., and Coop, G. (2020). Selection and hybridization shaped the rapid spread of African honey bee ancestry in the Americas. PLoS Genet. 16:e1009038. doi: 10.1371/journal.pgen.1009038
Cane, J. H., and Tepedino, V. J. (2017). Gauging the effect of honey bee pollen collection on native bee communities. Conserv. Lett. 10, 205–210. doi: 10.1111/conl.12263
Carneiro, F. E., Torres, R. R., Strapazzon, R., Ramírez, S. A., Guerra, J. C. V. Jr., Koling, D. F., et al. (2007). Changes in the reproductive ability of the Varroa destructor (Anderson & Trueman) in Africanized honey bees (Apis mellifera L.) (Hymenoptera: Apidae) colonies in southern Brazil. Neotropical. Entomol. 36, 949–952. doi: 10.1590/s1519-566x2007000600018
Cavigli, I., Daughenbaugh, K. F., Martin, M., Lerch, M., Banner, K., Garcia, E., et al. (2016). Pathogen prevalence and abundance in honey bee colonies involved in almond pollination. Apidologie 47, 251–266. doi: 10.1007/s13592-015-0395-5
Chemurot, M., De Smet, L., Brunain, M., De Rycke, R., and de Graaf, D. C. (2017). Nosema neumanni n. sp. (Microsporidia, Nosematidae), a new microsporidian parasite of honeybees, Apis mellifera in Uganda. Eur. J. Protistol. 61(Pt A), 13–19. doi: 10.1016/j.ejop.2017.07.002
Cobey, S. W., Sheppard, W. S., and Tarpy, D. R. (2012). “Status of breeding practices and genetic diversity in domestic us honey bees,” in Honey Bee Colony Health, eds D. Sammataro and J. A. Yoder (Boca Raton, FL: CRC Press. Taylor and Francis).
Conti, M. E., and Botrè, F. (2001). Honeybees and their products as potential bioindicators of heavy metals contamination. Environ. Monit. Assessment 69, 267–282. doi: 10.1023/a:1010719107006
Costa, C., Büchler, R., Berg, S., Bienkowska, M., Bouga, M., Bubalo, D., et al. (2012). A Europe-wide experiment for assessing the impact of genotype–environment interactions on the vitality and performance of honey bee colonies: experimental design and trait evaluation. J. Apic. Sci. 56, 147–158. doi: 10.2478/v10289-012-0015-9
Cox-Foster, D. L., Conlan, S., Holmes, E. C., Palacios, G., Evans, J. D., Moran, N. A., et al. (2007). A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318, 283–287. doi: 10.1126/science.1146498
Crane, E. (1999). The World History of Beekeeping and Honey Hunting. Abingdon: Taylor and Francis Group, 704.
Danka, R. G., Dodds, G. E., and Harris, J. W. (2016). Selection of VSH-derived “Pol-line” honey bees and evaluation of their Varroa-resistance characteristics. Apidologie 47, 483–490. doi: 10.1007/s13592-015-0413-7
Danka, R. G., Rinderer, T. E., Kuznetsoy, V. N., and Delatte, G. T. (1995). A USDA-ARS project evaluate resistance to Varroa jacobsoni by honey bees of far-eastern Russia. Am. Bee J. 135, 746–748.
Dietemann, V., Nazzi, F., Martin, S. J., Anderson, D. L., Locke, B., Delaplane, K. S., et al. (2013). Standard methods for varroa research. In V Dietemann; JD Ellis; P Neumann (Eds) The Coloss Beebook. Volume I: standard methods for Apis mellifera research. J. Apic. Res. 52, 1–54. doi: 10.3896/ibra.1.52.1.09
Doublet, V., Labarussias, M., de Miranda, J. R., Moritz, R. F. A., and Pacton, R. J. (2015). Bees under stress: sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 17, 969–983. doi: 10.1111/1462-2920.12426
Eimanifar, A., Brooks, S. A., Bustamante, T., and Ellis, J. D. (2018). Population genomics and morphometric assignment of western honey bees (Apis mellifera L.) in the Republic of South Africa. BMC Genomics 19:615. doi: 10.1186/s12864-018-4998-x
Eimanifar, A., Pieplow, J. T., Asem, A., and Ellis, J. D. (2020). Genetic diversity and population structure of two subspecies of western honey bees (Apis mellifera L.) in the Republic of South Africa as revealed by microsatellite genotyping. PeerJ 8:e8280. doi: 10.7717/peerj.8280
Eischen, F. A., Wilson, W. T., Pettis, J. S., Suarez, A., Cardoso-Tamez, D., Maki, D. L., et al. (1990). The Spread of Acarapis woodi (Acari: Tarsonemidae) in Northeastern Mexico. J. Kansas Entomol. Soc. 63, 375–384.
Fahey, R., Rennich, K., Nessa, A., Swan, N., Steinhauer, N., Eversole, H., et al. (2018). 2016 – 2017 National Honey Bee Disease Survey Report. National Honey Bee Disease Survey. Available online at: https://www.aphis.usda.gov/plant_health/plant_pest_info/honey_bees/downloads/2016-2017-National-Survey-Report.pdf (accessed December 28, 2020).
Fahey, R., Rennich, K., Nessa, A., Swan, N., Steinhauer, N., Eversole, H., et al. (2019). 2017 – 2018 National Honey Bee Disease Survey Report. National Honey Bee Disease Survey. 36pp. Available online at: https://ushoneybeehealthsurvey.info/wp-content/uploads/sites/3/2020/04/2017-2018-Summary-Report.pdf (accessed December 28, 2020).
Ferrier, P. M., Rucker, R. R., Thurman, W. N., and Burgett, M. (2018). Economic Effects and Responses to Changes in Honey Bee Health. USDA Economic Research Service. Economic Research Report Nr. 246. 54 p. Available online at: https://www.ers.usda.gov/webdocs/publications/88117/err-246.pdf?v=601 (accessed February 15, 2021).
Finger, D., Filho, I. K., Torres, Y. R., and Quinaia, S. P. (2014). Propolis as an indicator of environmental contamination by metals. Bull. Environ. Contam. Toxicol. 92, 259–264. doi: 10.1007/s00128-014-1199-4
Galbraith, D. A., Fuller, Z. L., Ray, A. M., Brockmann, A., Frazier, M., Gikungu, M. W., et al. (2018). Investigating the viral ecology of global bee communities with high-throughput metagenomics. Sci. Rep. 8:8879. doi: 10.1038/s41598-018-27164-z
Galindo-Cardona, A., Acevedo-Gonzalez, J. P., Rivera-Marchand, B., and Giray, T. (2013). Genetic structure of the gentle Africanized honey bee population (gAHB) in Puerto Rico. BioMed Central Genet. 14:65. doi: 10.1186/1471-2156-14-65
Gallai, N., Salles, J. M., Settele, J., and Vaissiere, B. E. (2009). Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 68, 810–821. doi: 10.1016/j.ecolecon.2008.06.014
Geslin, B., Gauzens, B., Baude, M., Dajoz, I., Fontaine, C., Henry, M., et al. (2017). Massively introduced managed species and their consequences for plant–pollinator interactions. Adv. Ecol. Res. 57, 147–199. doi: 10.1016/bs.aecr.2016.10.007
Giray, T., Guzman-Novoa, E., Aron, C. W., Zelinsky, B., Fahrbach, S. E., and Robinson, G. E. (2000). Genetic variation in worker temporal polyethism and colony defensiveness in the honey bee, Apis mellifera. Behav. Ecol. 11, 44–55. doi: 10.1093/beheco/11.1.44
Goodrich, B. K., Williams, J. C., and Goodhue, R. E. (2019). The great bee migration: supply analysis of honey bee colony shipments into California for almond pollination Services. Am. J. Agric. Econ. 101, 1353–1372. doi: 10.1093/ajae/aaz046
Goretti, E., Pallottini, M., Rossi, R., La Porta, G., Gardi, T., Cenci Goga, B. T., et al. (2020). Heavy metal bioaccumulation in honey bee matrix, an indicator to assess the contamination level in terrestrial environments. Environ. Pollut. 256:113388. doi: 10.1016/j.envpol.2019.113388
Goulson, D. (2003). Effects of introduced bees on native ecosystems. Annu. Rev. Ecol. Evol. Syst. 43, 1–26. doi: 10.1146/annurev.ecolsys.34.011802.132355
Goulson, D., and Derwent, L. C. (2004). Synergistic interactions between an exotic honey bee and an exotic weed: pollination of Lantana camara in Australia. Weed Res. 44, 195–202. doi: 10.1111/j.1365-3180.2004.00391.x
Goulson, D., Nicholls, E., Botías, C., and Rotheray, E. L. (2015). Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, 1–16. doi: 10.1126/science.1255957
Granberg, F., Vicente-Rubiano, M., Rubio-Guerri, C., Karlsson, O. E., Kukielka, D., Belák, S., et al. (2013). Metagenomic detection of viral pathogens in spanish honeybees: co-infection by aphid lethal paralysis, israel acute paralysis and lake sinai viruses. PLoS One 2013:e57459. doi: 10.1371/journal.pone.0057459
Graystock, P., Blane, E. J., McFrederick, Q. S., Goulson, D., and Hughes, W. O. H. (2016). Do managed bees drive parasite spread and emergence in wild bees? Int. J. Parasitol. 5, 64–75. doi: 10.1016/j.ijppaw.2015.10.001
Grozinger, C. M., and Flenniken, M. L. (2019). Bee viruses: ecology, pathogenicity, and impacts. Annu. Rev. Entomol. 64, 205–226. doi: 10.1146/annurev-ento-011118-111942
Guimarães, P. R. Jr., Jordano, P., and Thompson, J. N. (2011). Evolution and coevolution in mutualistic networks. Ecol. Lett. 14, 877–885. doi: 10.1111/j.1461-0248.2011.01649.x
Guzman-Novoa, E., Morfin, N., De la Mora, A., Macías-Macías, J. O., Tapia-González, J. M., Contreras-Escareño, F., et al. (2020). The process and outcome of the Africanization of honey bees in Mexico: lessons and future directions. Front. Ecol. Evolut. 8:608091. doi: 10.3389/fevo.2020.608091
Guzman-Novoa, E., Page, R. E. Jr., and Fondrk, M. K. (1994). Morphometric techniques do not detect intermediate and low levels of Africanization in honey bee (Hymenoptera: Apidae) colonies. Ann. Entomol. Soc. Am. 87, 507–515. doi: 10.1093/aesa/87.5.507
Haber, A. I. H., Steinhauer, N. A., and vanEngelsdorp, D. (2019). Use of chemical and nonchemical methods for the control of Varroa destructor (Acari: Varroidae) and associated winter colony losses in U.S. beekeeping operations. J. Econ. Entomol. 112, 1509–1525. doi: 10.1093/jee/toz088
Hall, G. H. (1992). Suspected African honeybee Colonies in Florida tested for identifying DNA markers. Fla. Entomol. Soc. 75, 257–266. doi: 10.2307/3495628
Harbo, J. R., and Harris, J. W. (1999a). Selecting honey bees for resistance to Varroa jacobsoni. Apidologie 30, 183–196. doi: 10.1051/apido:19990208
Harbo, J. R., and Harris, J. W. (1999b). Heritability in honey bees (Hymenoptera: Apidae) of characteristics associated with resistance to Varroa jacobsoni Mesostigmata: Varroidae). J. Econ. Entomol. 92, 261–265. doi: 10.1093/jee/92.2.261
Harpur, B. A., Chapman, N. C., Krimus, L., Maciukiewicz, P., Sandhu, V. K., Sood, K., et al. (2015). Assessing patterns of admixture and ancestry in Canadian honey bees. Insectes Soc 62, 479–489. doi: 10.1007/s00040-015-0427-1
Hatjina, F., Costa, C., Büchler, R., Uzunov, A., Drazic, M., Filipi, J., et al. (2014). Population dynamics of European honey bee genotypes under different environmental conditions. J. Apic. Res. 53, 233–247. doi: 10.3896/ibra.1.53.2.05
Henriques, D., Chávez-Galarza, J., Teixeira, J., Galaschi, S., Ferreira, H., Neves, C. J., et al. (2020). Wing geometric morphometrics of workers and drones and single nucleotide polymorphisms provide similar genetic structure in the Iberian honey bee (Apis mellifera iberiensis). Insects 11:89. doi: 10.3390/insects11020089
Hopkins, B. K., and Herr, C. (2010). Factors affecting the successful cryopreservation of honey bee (Apis mellifera) spermatozoa. Apidologie. 41, 548–556. doi: 10.1051/apido/20010006
Hopkins, B. K., Herr, C., and Sheppard, W. S. (2012). Sequential generations of honey bee (Apis mellifera) queens produced using cryopreserved semen. Reprod. Fertil. Dev. 24, 1079–1083. doi: 10.1071/RD11088
Hung, K.-L. J., Kingston, J. M., Albrecht, M., Holway, D. A., and Kohn, J. R. (2018). The worldwide importance of honey bees as pollinators in natural habitats. Proc. R. Soc. B 285:20172140. doi: 10.1098/rspb.2017.2140
Ibrahim, A., and Spivak, M. (2006). The relationship between hygienic behavior and suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of resistance to Varroa destructor. Apidologie 37, 31–40. doi: 10.1051/apido:2005052
Jabr, F. (2013). The Mind-Boggling Math of Migratory Beekeeping. Scientific American, August 20, 2013. Available online at: http://www.scientificamerican.com/article/migratory-beekeeping-mind-boggling-math/ (accessed February 15, 2021).
Ji, R., Xie, B., Yang, G., and Li, D. (2003). From introduced species to invasive species—a case study on the Italian bee Apis mellifera L. J. Chin. Ecol. 5, 70–73.
Jordan, A., Patch, H. M., Grozinger, C. M., and Khanna, V. (2021). Economic Dependence and Vulnerability of United States Agricultural Sector on Insect-Mediated Pollination Service. Environ. Sci. Technol. 55, 2243–2253. doi: 10.1021/acs.est.0c04786
Kast, C., Kilchenmann, V., and Droz, B. (2020). Distribution of coumaphos in beeswax after treatment of honeybee colonies with CheckMite® against the parasitical mite Varroa destructor. Apidologie 51, 112–122. doi: 10.1007/s13592-019-00724-6
Kennedy, C. M., Lonsdorf, E., Neel, M. C., Williams, N. M., Ricketts, T. H., Winfree, R., et al. (2013). A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 16, 584–599. doi: 10.1111/ele.12082
Kerr, W. E. (1967). The history of introduction of African bees to Brazil. South Afr. Bee J. 39, 3–5.
Klein, S., Cabirol, A., Devaud, J.-M., Barron, A. B., and Lihoreau, M. (2017). Why bees are so vulnerable to environmental stressors. Trends Ecol. Evol. 32, 268–278. doi: 10.1016/j.tree.2016.12.009
Koeniger, G., Koeniger, N., Ellis, J., and Connor, L. (2014). Mating Biology of Honey Bees (Apis mellifera). Kalamazoo, MI: Wicwas Press, 155.
Kovaèiæ, M., Puškadija, Z., Dražiæ, M. M., Uzunov, A., Meixner, M. D., and Büchler, R. (2020). Effects of selection and local adaptation on resilience and economic suitability in Apis mellifera carnica. Apidologie 51, 1062–1073. doi: 10.1007/s13592-020-00783-0
Lautenbach, S., Seppelt, R., Liebscher, J., and Dormann, C. F. (2012). Spatial and temporal trends of global pollination benefit. PLoS One 7:e35954. doi: 10.1371/journal.pone.0035954
Losey, J. E., and Vaughan, M. (2006). The economic value of ecological services provided by insects. BioScience 56, 311–323. doi: 10.1641/0006-3568(2006)56[311:tevoes]2.0.co;2
Mailander, D., and Grant, Z. (2019). When Honey Bees Hit the Road: The Role of Federal, State, and Local Laws in Regulating Honey Bee Transportation. A White Paper of the University of Oregon School of Law Environmental and Natural Resources Law Center Food Resiliency Projec. University of Oregon. 55 p. Available online at: https://law.uoregon.edu/sites/law1.uoregon.edu/files/when_honey_bees_hit_the_road.pdf (accessed April 26, 2022).
Mallinger, R. E., Gaines-Day, H. R., and Gratton, C. (2017). Do managed bees have negative effects on wild bees? A systematic review of the literature. PLoS One 12:e0189268. doi: 10.1371/journal.pone.0189268
Matthews, W. A., Baratashvili, S., and Sumner, D. A. (2020). University of California, Agricultural Issues Center. Contributions of the California Almond Industry to the California Economy. Report for the Almond Board of California. Available online at: https://aic.ucdavis.edu/wp-content/uploads/2020/08/Economic-Impact-Study-Manuscript-Final.pdf (accessed April 26, 2022).
McKinney, M. L., and La Sorte, F. A. (2007). Invasiveness and homogenization: synergism of wide dispersal and high local abundance. Glob. Ecol. Biogeogr. 16, 394–400. doi: 10.1111/j.1466-8238.2007.000296.x
Meixner, M. D., Pinto, M. A., Bouga, M., Kryger, P., Ivanova, E., and Fuchs, S. (2013). Standard methods for characterizing subspecies and ecotypes of Apis mellifera. J. Apic. Res. 52, 1–27. doi: 10.3896/ibra.1.52.4.05
Meusel, M. S., and Moritz, R. F. A. (1993). Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs. Curr. Genet. 24, 539–543. doi: 10.1007/BF00351719
Millán-Leiva, A., Marín, Ó, Christmon, K., vanEngelsdorp, D., and González-Cabrera, J. (2021). Mutations associated with pyrethroid resistance in Varroa mite, a parasite of honey bees, are widespread across the United States. Pest. Manag. Sci. 77, 3241–3249. doi: 10.1002/ps.6366
Momeni, J., Parejo, M., Nielsen, R. O., Langa, J., Montes, I., Papoutsis, L., et al. (2021). Authoritative subspecies diagnosis tool for European honey bees based on ancestry informative SNPs. BMC Genomics 22:101. doi: 10.1186/s12864-021-07379-7
Mondet, F., Beaurepaire, A., McAfee, A., Locke, B., Alaux, C., Blanchard, S., et al. (2020). Honey bee survival mechanisms against the parasite Varroa destructor: a systematic review of phenotypic and genomic research efforts. Int. J. Parasitol. 50, 433–447. doi: 10.1016/j.ijpara.2020.03.005
Moritz, R. F. A., Hartel, S., and Neumann, P. (2005). Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 12, 289–301. doi: 10.2980/i1195-6860-12-3-289.1
Mullin, C. A., Frazier, M., Frazier, J. L., Ashcraft, S., Simonds, R., vanEngelsdorp, D., et al. (2010). High levels of miticides and agrochemicals in North American apiaries: implications for honey bee health. PLoS One 5:e9754. doi: 10.1371/journal.pone.0009754
Munoz, I., Henriques, D., Johnston, J. S., Chavez-Galarza, J., Kryger, P., and Pinto, M. A. (2015). Reduced SNP panels for genetic identification and introgression analysis in the dark honey bee (Apis mellifera mellifera). PLoS One 10:e0124365. doi: 10.1371/journal.pone.0124365
NASS (2020). National Agricultural Statistics Service. United States Department of Agriculture. National Statistics for Almonds. 2020 California Almond Objective Measurement Report. Available online at: https://www.nass.usda.gov/Statistics_by_State/California/Publications/Specialty_and_Other_Releases/Almond/Objective-Measurement/202007almom.pdf (accessed February 16, 2021].
NASS (2021). National Agricultural Statistics Service. United States Department of Agriculture. United States Honey Production. Available online at: https://downloads.usda.library.cornell.edu/usda-esmis/files/hd76s004z/7h14bh90x/w9505v43v/hony0321.pdf (accessed April 26, 2022).
Nawrocka, A., Kandemir, I., Fuchs, S., and Tofilski, A. (2018). Computer software for identification of honey bee subspecies and evolutionary lineages. Apidologie 49, 172–184.
Neumann, P. (2006). Infestation levels of Apis mellifera scutellata swarms by socially parasitic Cape honeybee workers (Apis mellifera capensis). Apidologie 37, 462–470. doi: 10.1051/apido:2006012
Neumann, P., and Moritz, R. F. A. (2002). The cape honeybee phenomenon: the sympatric evolution of a social parasite in real time? Beahv. Ecol. Sociobiol. 52, 271–281. doi: 10.1007/s00265-002-0518-7
Norfolk, O., Gilbert, F., and Eichhorn, M. P. (2018). Alien honey bees increase pollination risks for range-restricted plants. Divers. Distrib. 24, 705–713. doi: 10.1111/ddi.12715
Owen, R. (2017). Role of human action in the spread of honey bee (Hymenoptera: Apidae) pathogens. J. Econ. Entomol. 110, 797–801. doi: 10.1093/jee/tox075
Paini, D. R., and Roberts, J. D. (2005). Commercial honey bees (Apis mellifera) reduce the fecundity of an Australian native bee (Hylaeus alcyoneus). Biol. Conserv. 123, 103–112. doi: 10.1016/j.biocon.2004.11.001
Perez, A., and Plattner, K. (2014). Fruit and Tree Outlook. USDA Economic Research Service. Fruit and Tree Nuts Outlook Special Article/FTS-357SA/September 26, 2014. Available online at: https://www.ers.usda.gov/webdocs/outlooks/37059/49130_fts-357.pdf?v=6225.6 (accessed September 26, 2014).
Pernal, S. F. (2014). “National regulations for beekeeping in north america (Canada and the United States of America),” in Bee Health and Veterinarians, ed. W. Ritter (Paris: OIE World Organization for Animal Health), 275–280.
Pilati, L., and Prestamburgo, M. (2016). Sequential relationship between profitability and sustainability: the case of migratory beekeeping. Sustainability 8:94. doi: 10.3390/su8010094
Pinto, M. A., Henriques, D., Chaves-Galarza, J., Kryger, P., Garnery, L., Van der Zee, M., et al. (2014). Genetic integrity of the Dark European honey bee (Apis mellifera mellifera) from protected populations: a genome-wide assessment using SNPs and mtDNA sequence data. J. Apic. Res. 53, 269–278. doi: 10.3896/ibra.1.53.2.08
Ramadan, M. M., Reimer, N. J., Oishi, D. E., Young, C. L., and Heu, R. A. (2019). Varroa mite Varroa destructor Anderson & Trueman (Acari: Varroidae). New York, NY: Springer.
Ray, A. M., Lopez, D. L., Iturralde Martinez, F. J., Galbraith, D. A., Rose, R., Van Engelsdorp, D., et al. (2020). Distribution of recently identified bee-infecting viruses in managed honey bee (Apis mellifera) populations in the USA. Apidologie 51, 736–745. doi: 10.1007/s13592-020-00757-2
Rennie, J. (1921). Isle of wight disease in hive bees – acarine disease: the organism associated with the disease Tarsonemus woodi, n. sp. Trans. R. Soc. Edinburgh 52, 768–779. doi: 10.1017/s0080456800016008
Ribani, A., Utzeri, V. J., Taurisano, V., and Fontanesi, L. (2020). Honey as a source of environmental DNA for the detection and monitoring of honey bee pathogens and parasites. Vet. Sci. 7:113. doi: 10.3390/vetsci7030113
Rinderer, T. E., de Guzman, L. I., Harris, J., Kuznetsov, V., Delatte, G. T., Stelzer, J. A., et al. (2000). The release of ARS Russian honey bees. Am. Bee J. 140, 305–307.
Rinderer, T. E., Delatte, G. T., De Guzman, L. I., Williams, J., Stelzer, J. A., and Kuznetsov, V. N. (1999). Evaluations of the Varroa-resistance of honey bees imported from Far-Eastern Russia. Am. Bee J. 139, 287–290.
Rinderer, T. E., Guzman, L. I., Delatte, G. T., Stelzer, J. A., Williams, J. L., Beaman, L. D., et al. (2001a). Multi-state field trials of ARS Russian honey bees 1. Response to Varroa destructor 1999, 2000. Am. Bee J. 141, 658–661.
Rinderer, T. E., Guzman, L. I., Delatte, G. T., Stelzer, J. A., Williams, J. L., Beaman, L. D., et al. (2001b). Multi-state field trials of ARS Russian honey bees 2. Honey production 1999, 2000. Am. Bee J. 141, 726–729.
Rinderer, T. E., Kuznetsoy, V., Danka, R. G., and Delatte, G. T. (1997). An importation of potentially Varroa-resistant honey bees from Far-Eastern Russia. Am. Bee J. 137, 787–789.
Rinkevich, F. D. (2020). Detection of amitraz resistance and reduced treatment efficacy in the Varroa Mite, Varroa destructor, within commercial beekeeping operations. PLoS One 15:e0227264. doi: 10.1371/journal.pone.0227264
Rivera-Marchand, B., Oskay, D., and Giray, T. (2012). Gentle Africanized bees on an oceanic island. Evol. Appl. 5, 746–756. doi: 10.1111/j.1752-4571.2012.00252.x
Roubik, D. (2009). Ecological impact on native bees by the invasive Africanized honey bee. Acta Biol. Colombiana 14, 114–124.
Roubik, D., and Villanueva-Gutierrez, R. (2009). Invasive Africanized honey bee impact on native solitary bees: a pollen resource and trap nest analysis: responses to invasive honey bees. Biol. J. Linnean Soc. 98, 152–160. doi: 10.1111/j.1095-8312.2009.01275.x
Rucker, R. R., Thurman, W. N., and Burgett, M. (2012). Honey bee pollination markets and the internalization of reciprocal benefits. Am. J. Agric. Econ. 94, 956–977. doi: 10.1093/ajae/aas031
Rules and Regulations (2014). United States department of agriculture. Animal and plant health inspection services. 7 CFR parts 319 and 322. Bees and related articles. Federal Register 69, 61735–61755.
Runckel, C., Flenniken, M. L., Engel, J. C., Ruby, J. G., Ganem, D., Andino, R., et al. (2011). Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS One 6:e20656. doi: 10.1371/journal.pone.0020656
Rusert, L. M., Pettis, J. S., and Tarpy, D. R. (2021). Introduction of Varroa destructor has not altered honey bee queen mating success in the Hawaiian archipelago. Sci. Rep. 11:1366. doi: 10.1038/s41598-020-80525-5
Russo, L. (2016). Positive and negative impacts of non-native bee species around the world. Insects 7:69. doi: 10.3390/insects7040069
Sadd, B. M., Kleinlogel, Y., Schmid-Hempel, R., and Schmid-Hempel, P. (2005). Trans-generational immune priming in a social insect. Biol. Lett. 1, 386–388. doi: 10.1098/rsbl.2005.0369
Schiff, N. M., and Sheppard, W. S. (1995). Genetic Analysis of commercial honey bees (Hymenoptera: Apidae) from the southeastern United States. J. Econ. Entomol. 88, 1216–1220. doi: 10.1093/jee/88.5.1216
Schneider, S. S., DeGrandi-Hoffman, G., and Smith, D. R. (2004). The African Honey bee. Factors contributing to a successful biological invasion. Ann. Rev. Entomol. 49, 351–376. doi: 10.1146/annurev.ento.49.061802.123359
Shahbandeh, M. (2021). Honey Market Worldwide And In The U.S. – Statistics & Facts. Statista. Available online at: https://www.statista.com/topics/5090/honey-market-worldwide/#dossierKeyfigures (accessed October 1, 2021).
Sheppard, W. S. (1989a). A history of the introduction of honey bee races into the United Sates: Part I. Am. Bee J. 129, 617–619.
Sheppard, W. S. (1989b). A history of the introduction of honey bee races into the United Sates: Part II. Am. Bee J. 129, 664–667.
Sheppard, W. S. (2012). Honey bee genetic diversity and breeding: towards the reintroduction of european germplasm. Bee Health 140, 24–27.
Shimanuki, H., and Knox, D. A. (2000). Diagnosis of Honey Bee Diseases. Agriculture Handbook No. AH690. Beltsville, MD: United States Department of Agriculture.
Simone-Finstrom, M., Li-Byarlay, H., Huang, M. H., Strand, M. K., Rueppell, O., and Tarpy, D. R. (2016). Migratory management and environmental conditions affect lifespan and oxidative stress in honey bees. Sci. Rep. 6:32023. doi: 10.1038/srep32023
Smith, D. R. (1991). African bees in the americas: insights from biogeography and genetics. Trends Ecol. Evol. 6, 17–21. doi: 10.1016/0169-5347(91)90142-K
Spivak, M., and Reuter, G. S. (2001). Resistance to American foulbrood disease by honey bee colonies Apis mellifera bred for hygienic behavior. Apidologie 32, 555–565. doi: 10.1051/apido:2001103
Spivak, M., and Reuter, G. S. (2016). Honey bee Diseases and Pests: A companion to Beekeeping in Northern Climates. St Paul, MN: University of Minesota Extension, 42.
Steinhauer, N., vanEngelsdorp, D., and Saegerman, C. (2020). Prioritizing changes in management practices associated with reduced winter honey bee colony losses for US beekeepers. Sci. Total Environ. 753:141629. doi: 10.1016/j.scitotenv.2020.141629
Strauss, U., Dietemann, V., Human, H., Crewe, R. M., and Pirk, C. W. W. (2015). Resistance rather than tolerance explains survival of savannah honeybees (Apis mellifera scutellata) to infestation by the parasitic mite Varroa destructor. Parasitology 143, 374–387. doi: 10.1017/S0031182015001754
Sumner, D. A., Matthews, W. A., Medellín-Azuara, J., and Bradley, A. (2014). The Economic Impacts of the California Almond Industry. The Almond Board of California. Davis, CA: University, California.
Thomson, D. (2004). Competitive interactions between the invasive European honey bee and native bumble bees. Ecology 85, 458–470. doi: 10.1890/02-0626
Thomson, D. M. (2016). Local bumble bee decline linked to recovery of honey bees, drought effects on floral resources. Ecol. Lett. 19, 1247–1255. doi: 10.1111/ele.12659
Torné-Noguera, A., Rodrigo, A., Osorio, S., and Bosch, J. (2016). Collateral effects of beekeeping: Impacts on pollen-nectar resources and wild bee communities. Basic Appl. Ecol. 17, 199–209. doi: 10.1016/j.baae.2015.11.004
Traniello, J. F. A., Rosengaus, R. B., and Savoie, K. (2002). The development of immunity in a social insect: evidence for the group facilitation of disease resistance. Proc. Natl. Acad. Sci. U.S.A. 99, 6838–6842. doi: 10.1073/pnas.102176599
Traynor, K. S., Rennich, K., Forsgren, E., Rose, R., Pettis, J., Kunkel, G., et al. (2016). Multiyear survey targeting disease incidence in US honey bees. Apidologie 47, 325–347. doi: 10.1007/s13592-016-0431-0
Traynor, K. S., Tosi, S., Rennich, K., Steinhauer, N., Forsgren, E., Rose, R., et al. (2021). Pesticides in honey bee colonies: establishing a baseline for real world exposure over seven years in the USA. Environ. Pollut. 279:116566. doi: 10.1016/j.envpol.2021.116566
Uzunov, A., Costa, C., Panasiuk, B., Meixner, M., Kryger, P., Hatjina, F., et al. (2014). Swarming, defensive and hygienic behaviour in honey bee colonies of different genetic origin in a Pan-European experiment. J. Apic. Res. 53, 248–260. doi: 10.3896/ibra.1.53.2.06
Valido, A., Rodríguez-Rodríguez, M. C., and Jordano, P. (2019). Honeybees disrupt the structure and functionality of plant-pollinator networks. Sci. Rep. 9:4711. doi: 10.1038/s41598-019-41271-5
Van der Steen, J., de Kraker, J., and Grotenhuis, T. (2011). Spatial and temporal variation of metal concentrations in adult honeybees (Apis mellifera L.). Environ. Monit. Assessment 184, 4119–4126. doi: 10.1007/s10661-011-2248-7
vanEngelsdorp, D., and Meixner, M. D. (2010). A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 103, S80–S95. doi: 10.1016/j.jip.2009.06.011
vanEngelsdorp, D., Tarpy, D. R., Lengerich, E. J., and Pettis, J. S. (2012). Idiopathic brood disease syndrome and queen events as precursors of colony mortality in migratory beekeeping operations in the eastern United States. Prev. Vet. Med. 108, 225–233. doi: 10.1016/j.prevetmed.2012.08.004
Visscher, P. K., Vetter, R. S., and Baptista, F. C. (1997). Africanized bees, 1990-1995: Initial rapid expansion has slowed in the U.S. Calif. Agric 51, 22–25. doi: 10.3733/ca.v051n01p22
Voorhies, E. C., Todd, Frank, E., and Galbraith, J. K. (1933). Economic aspects of the bee industry. Univer.Calif. Coll. Agric. Bull. 555, 1–117.
VPPC (2016). Vermont’s pollination Protection committee. Report to the Vermont Legislature as required by Act 83 of 2016 Session. 67 p. Available online at: https://agriculture.vermont.gov/sites/agriculture/files/documents/Pollinator%20Protection%20Report-FINAL.pdf (accessed February 16 2021).
Wehling, W. F., and Flanders, R. V. (2005). The Risks of International Trade in Bees: Laws And Regulations on The Import, Movement and Release Of Bees in The United States. Proceedings of The Bee Research and Virus in Europe (BRAVE), Sophia-Antipolis, France, 24-26 April 2005, page 301-309. Available online at: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.115.4234&rep=rep1&type=pdf#page=301 (accessed April 26, 2022).
Whitfield, C. W., Behura, S. K., Berlocher, S. H., Clark, A. G., Johnston, J. S., Sheppard, W. S., et al. (2006). Thrice out of Africa: ancient and recent expansions of the honey bee, Apis mellifera. Science 314, 642–645. doi: 10.1126/science.1132772
Williams, I. H. (1994). The dependences of crop production within the European Union on pollination by honey bees. Agric. Zool. Rev. 6, 229–257.
Winter, K., Adams, L., Thorp, R., Inouye, D., Day, L., Ascher, J., et al. (2006). Importation of Non-Native Bumble Bees into North America: Potential Consequences of Using Bombus terrestris and Other Non-Native Bumble Bees for Greenhouse Crop Pollination in Canada, Mexico, and the United States. A White Paper of the North American Pollinator Protection Campaign (NAPPC). Available online at: https://www.pollinator.org/pollinator.org/assets/generalFiles/BEEIMPORTATION_AUG2006.pdf (accessed April 26, 2022).
Keywords: honey bee stock, Apis mellifera, human mediated movement, regulations, benefits, risks, mitigation strategies
Citation: Marcelino J, Braese C, Christmon K, Evans JD, Gilligan T, Giray T, Nearman A, Niño EL, Rose R, Sheppard WS, vanEngelsdorp D and Ellis JD (2022) The Movement of Western Honey Bees (Apis mellifera L.) Among U.S. States and Territories: History, Benefits, Risks, and Mitigation Strategies. Front. Ecol. Evol. 10:850600. doi: 10.3389/fevo.2022.850600
Received: 07 January 2022; Accepted: 19 April 2022;
Published: 31 May 2022.
Edited by:
Rosanna Giordano, Science, Technology and Research Trust, Puerto RicoReviewed by:
Carlos A. Blanco, University of New Mexico, United StatesCopyright © 2022 Marcelino, Braese, Christmon, Evans, Gilligan, Giray, Nearman, Niño, Rose, Sheppard, vanEngelsdorp and Ellis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jose Marcelino, am1hcmNlbGlub0B1ZmwuZWR1; James D. Ellis, amRlbGxpc0B1ZmwuZWR1
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.