 Leonardo Ordóñez-Delgado1,2
Leonardo Ordóñez-Delgado1,2 Carlos Iñiguez-Armijos1
Carlos Iñiguez-Armijos1 Mario Díaz3
Mario Díaz3 Adrián Escudero4
Adrián Escudero4 Elyce Gosselin5
Elyce Gosselin5 Lisette P. Waits5
Lisette P. Waits5 Carlos Iván Espinosa1*
Carlos Iván Espinosa1*
- 1Laboratorio de Ecología Tropical y Servicios Ecosistémicos (EcoSs-Lab), Departamento de Ciencias Biológicas y Agropecuarias, Universidad Técnica Particular de Loja, Loja, Ecuador
- 2Programa de Doctorado en Conservación de Recursos Naturales, Escuela Internacional de Doctorado, Universidad Rey Juan Carlos, Madrid, Spain
- 3Departamento de Biogeografía y Cambio Global (BGC–MNCN), Museo Nacional de Ciencias Naturales, CSIC, Madrid, Spain
- 4Department of Science, Rey Juan Carlos University, Madrid, Spain
- 5Department of Fish and Wildlife Sciences, University of Idaho, Moscow, ID, United States
Urbanization constitutes one of the most aggressive drivers of habitat and biodiversity loss worldwide. However, studies focused on determining the response of local biodiversity to urbanization are still scarce, especially in tropical ecosystems. Urban ecosystems are characterized by low biological productivity which in turn leads to a reduction in biodiversity. However, the responses to urbanization should be species dependent. For instance, changes in the availability of resources can favor certain species with specific characteristics. We assessed the effects of the urbanization process on a bird community in a city located in the Tropical Andes of southern Ecuador, a region widely recognized for its diversity and endemism of birds. We selected three independent localities in each of the four levels of the urbanization gradient in the study area (forest, forest-pasture, pasture, and urban). In each locality, we sampled the bird community by visual and auditory surveys along 1 km transects between 2016 and 2017. We recorded a total of 1,257 individuals belonging to 74 bird species. We evaluated if the responses of richness and abundance of birds are dependent on trophic guild and foraging strata. We found a significant decrease in bird species richness and abundance from forest to urban sites. However, the response of birds was dependent on the trophic guild and foraging strata. Granivorous birds showed a positive response associated with the urbanization gradient while insectivorous birds showed a negative response. Insectivorous birds were more abundant in forest sites and decreased in abundance across the urbanization gradient. We found that the proportion of birds using different foraging strata drastically changed along urban gradient. Forest sites exhibited a bird community using a variety of habitats, but the bird community became simpler toward the most urbanized sites. Our findings showed different effects of urbanization on bird communities. The ugly: urbanization leads to a dramatic reduction in the diversity of birds, which is consistent in cities with different characteristics and ecological contexts. On the other hand, the responses of bird guilds to urbanization are species dependent. Some guilds are positively impacted by urbanization and show increases in species richness and abundance while other guilds are negatively impacted.
Introduction
The expansion of urban areas is one of the greatest and most widespread threats to biodiversity worldwide (Lee and Rotenberry, 2015; Banville et al., 2017). The shift from original natural habitats to urban habitats triggers profound changes in many environmental factors (McDonnell and Hahs, 2008; Seress et al., 2014). The alteration of vegetation cover; an increase of chemical, light, and noise pollution; and higher environmental temperatures have been reported as the main effects of urbanization (Yow, 2007; Fox, 2013; Díaz et al., 2022). Those changes have strong cascading effects on species composition of plant and animal communities at both regional and global scales (Grimm et al., 2008; Arroyo-Rodríguez et al., 2013; Aronson et al., 2014) and usually involve a dramatic reduction of biodiversity (McDonnell and Hahs, 2008; Seress et al., 2014; Beninde et al., 2015; Sherry, 2021).
The effects of urbanization have been evaluated using different taxonomic groups, such as plants (Kowarik, 2008), insects (McIntyre, 2000; Blair, 2001; Gaona et al., 2021), aquatic macroinvertebrates (Zúñiga-Sarango et al., 2020), mammals (McIntyre, 2000), amphibians and reptiles (Hutto and Barrett, 2021), and birds (Pennington and Blair, 2011; McClure et al., 2015; Maure et al., 2018; Souza et al., 2019; Morelli et al., 2021; Valente-Neto et al., 2021). Birds tend to be especially affected by urbanization due to their high degree of specialization (Beaumont et al., 2011; Hermes et al., 2018). As a general rule, the effects of urbanization on bird communities are negative (Pennington and Blair, 2011; McClure et al., 2015; Ibáñez-Álamo et al., 2017; Maure et al., 2018; Morelli et al., 2021), evidencing decreases in species richness and abundance (Chiari et al., 2010; MacGregor-Fors et al., 2010), as well as decreases in phylogenetic and functional diversity (Carvajal-Castro et al., 2019; Morelli et al., 2021; Valente-Neto et al., 2021). Nevertheless, there is evidence for some positive effects of urbanization on specific bird species (Chiari et al., 2010; MacGregor-Fors et al., 2010; Møller and Díaz, 2018a). Therefore, the responses of birds to urbanization are difficult to predict (Rodewald and Yahner, 2001) because such responses are species-specific (Forsman et al., 2010; Montaño-Centellas and Garitano-Zavala, 2015).
Negative effects of urbanization on birds are especially significant for frugivores and nectarivores (de Oliveira Ramos and dos Anjos, 2014; Souza et al., 2019) because flower and fruit abundance drops in urban areas (Schütz and Schulze, 2015; Concepción et al., 2017). In contrast, generalist species such omnivores, granivores, or invasive species (Sol et al., 2017), and those foraging on the ground (MacGregor-Fors and Escobar-Ibáñez, 2017), usually take advantage of these urban habitats (Chiari et al., 2010; MacGregor-Fors et al., 2010). Increased abundance of generalist birds within cities (Møller and Díaz, 2018a) is likely the result of a dramatic reduction of predators, nest parasites, and competitors (de Oliveira Ramos and dos Anjos, 2014; Møller et al., 2016; Møller and Díaz, 2018b), an increase in refuges and/or nesting sites (Leveau, 2018), a higher supply of food associated with humans (McKinney, 2002; De León et al., 2019; Díaz et al., 2022), and a longer breeding season due to a warmer climate, especially in higher latitudes (Møller et al., 2015).
On the other hand, the trophic-guild organization of birds is directly related to the vegetation structure, which in turn is determined by the degree of anthropic modification (Laurance and Bierregaard, 1997). Several studies showed a positive association between structural complexity of vegetation and bird diversity (Fajardo et al., 2009; Carranza-Quiceno et al., 2018). Forested areas have greater plant diversity, occupying different strata and providing a complex canopy structure (de Toledo et al., 2012; Leveau et al., 2015). Conversely, vegetation within cities becomes simpler and sparser with few, scattered trees—mainly exotic species—and with the middle and lower vegetation strata disappearing (Chace and Walsh, 2006). Therefore, modified habitats allow the presence of an important number of bird species of different feeding guilds which take advantage of the available resources depending on the phenology and seasonality of the vegetation (Ramírez-Albores, 2004; Aycart and Díaz, 2021).
In spite of the effort to understand the effects of urbanization on birds, the information for the Tropical regions such as the Andes is still scarce (see Escobar-Ibáñez and MacGregor-Fors, 2017). This is disconcerting because the tropical Andes are the richest hotspots of bird biodiversity globally (Mittermeier et al., 2011; Bax and Francesconi, 2019). However, the major concern is that the cities in these regions are experiencing rapid expansion, disturbing their adjacent natural habitats (Cincotta et al., 2000; Luck, 2007). The Tropical Andes host more than 25% of the world’s bird species in a territory that corresponds to approximately 3% of the planet’s surface (BirdLife International and Conservation International, 2005). Despite its importance for birds, this region is one of the most threatened, mainly due to a rapid land transformation (Jetz et al., 2007), urbanization being one of the most critical pressures on biodiversity (Ramirez-Villegas et al., 2014; Zúñiga-Sarango et al., 2020). Consequently, more than 10% of the bird species in the region are seriously threatened (IUCN, 2021).
Conscious of the dramatic loss of bird diversity and of the extreme habitat degradation due to the expansion of cities in the Tropical Andes, we have conducted a study of a bird community along an urbanization gradient. The study was carried out in one middle size Andean city of Ecuador in sites ranging from the best-preserved areas (peri-urban forests) to the most disturbed areas dominated by urban development. We specifically evaluated (1) to what extent the bird diversity is negatively affected by urbanization, (2) do these negative effects differ among trophic guilds and foraging strata of birds? We hypothesized that the diversity and abundance of birds would exhibit a drastic decrease along the urbanization gradient as a consequence of the simplification of the landscape. Also, we predicted that the effects would likely differ among trophic guilds and foraging strata. For instance, granivores or birds that forage on the ground would be positively impacted rather than negatively affected by urbanization due to food subsidies from human sources, whereas insectivores would be negatively affected within cities.
Materials and Methods
Study Area and Sampling Sites
The study was carried out in the city of Loja and its surroundings in a watershed located in the southern Ecuadorian Andes (Figure 1). This city is located between two important areas of bird endemism (BirdLife International, 2021a,b), the Southern Central Andes (EBA046) and the Central Andean Paramo (EBA043). These areas are recognized globally for their high diversity of birds and the presence of several highly threatened species with narrow ranges, such as the bearded guan (Penelope barbata), the rainbow starfrontlet (Coeligena iris), the neblina metaltail (Metallura odomae), or the mouse-colored thistletail (Asthenes griseomurina) (BirdLife International, 2021a,b).
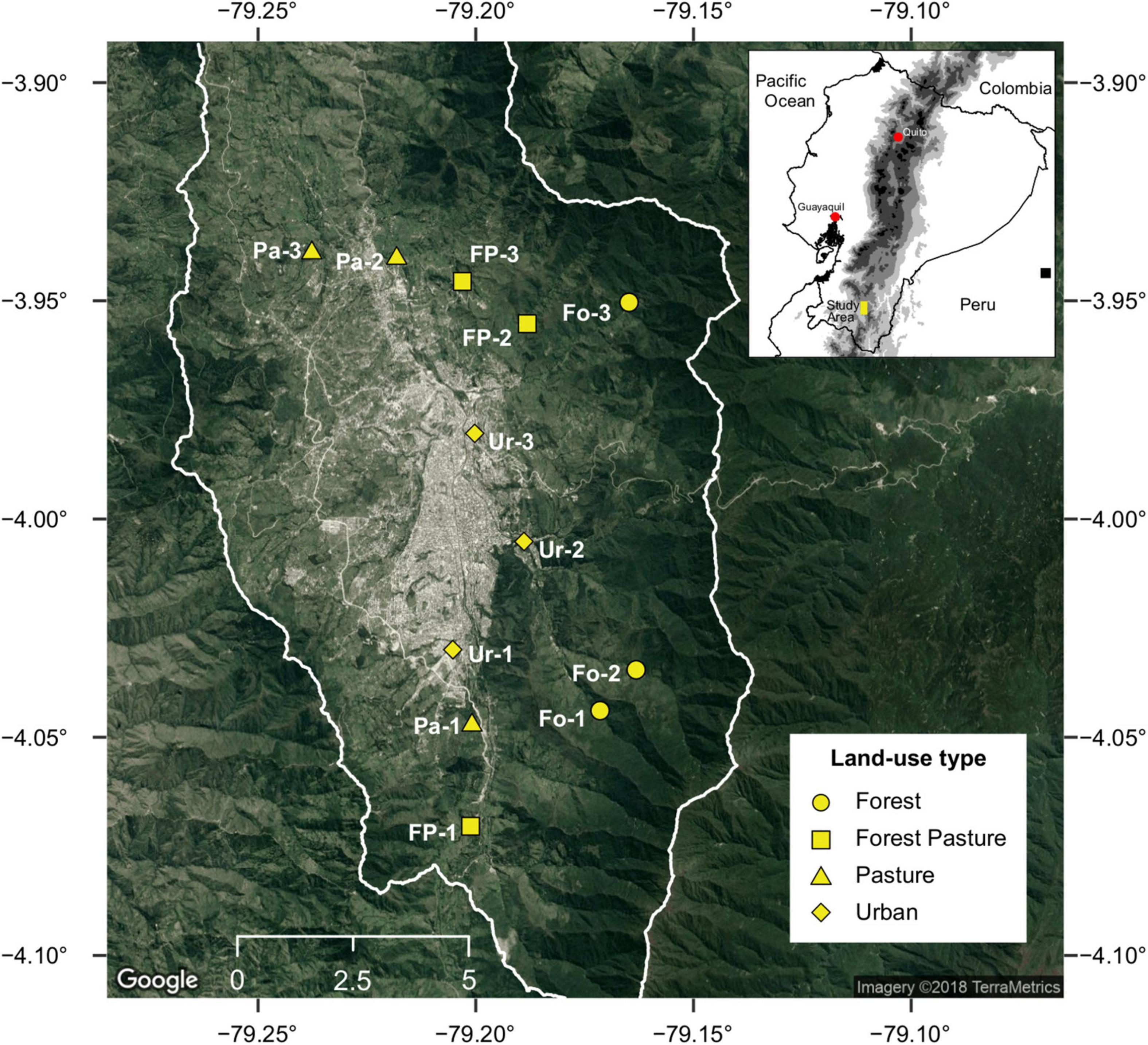
Figure 1. Location of the study area, the city of Loja, and the sampling sites (yellow symbols) within different land-use types in southern Ecuadorian Andes. Fo, forest; FP, forest-pasture; Pa, pasture; Ur, urban. Modified from Chuquimarca et al. (2019).
Altitude in the watershed ranges from 2,000 to 3,250 m. Climate is influenced by humid air masses from the Amazon and dry air masses from the Tumbes Dry Region. The annual precipitation varies between 800 and 2,500 mm and with a range of annual temperature between 7 and 15°C (Iñiguez-Armijos et al., 2014). The landscape of the study area has been transformed from native vegetation to other land uses throughout history but with a sharp intensification in recent decades (Ochoa-Cueva et al., 2015; Tapia-Armijos et al., 2015). The city of Loja, with ∼215,000 inhabitants, occupies approximately 36% (99 km2) of a watershed of 276 km2 (Gaona et al., 2021). The population growth of the city in general has been very low with a significant deceleration between 1974 and 2001. The number of inhabitants in 1950 was ∼50,000 and by 2001 it reached ∼175,000 inhabitants. Loja is a small-medium sized city relative to other Ecuadorian cities, with a contribution of around 6% of the national population (Information taken from the INEC 2022 web portal). It is located in the valley and has spread out toward the outskirts, replacing native vegetation first into pasture and then to urban areas (Figure 1), displaying a gradient of transformation to urban areas (Zúñiga-Sarango et al., 2020). The actual landscape in the area exhibits a city surrounded by pastureland dedicated to cattle ranching with the native vegetation restricted to the headwaters (Iñiguez-Armijos et al., 2016; Ordóñez-Delgado et al., 2016).
Experimental Design
Since the urbanization process in the study area has been defined by the transformation of native forest vegetation into cleared forest, pasture, and then to urban areas, we have defined four levels of disturbance based on the landscape pattern (see Iñiguez-Armijos et al., 2022). We selected 12 watersheds, hereafter sampling sites, arranged in watersheds that are characterized by different levels of entry within the city and that include different levels of urbanization. Forest sites (Fo) correspond to the least disturbed areas and are dominated by montane forests (≥75% of the watershed). These watersheds have isolated houses and are the farthest sites from the city. Forest-pasture sites (FP) are areas dominated by a mixture of disturbed montane forest (∼60% of the watershed) and pasture (∼40% of the watershed) and are occasionally used by livestock. In these watersheds, there are small neighborhoods that have not been completely integrated with the city. Pasture sites (Pa) correspond to areas that are dominated by pastures (>65%) and are entirely dedicated to cattle ranching. These watersheds are located on the edge of the city and include some urbanized areas connected to the city; and although urbanization is not yet dominant upstream these sites are directly influenced by the urban areas. Finally, urban sites (Ur) correspond to watersheds with around 30% of their area composed of impervious surfaces and the presence of few green areas which are dominated by exotic and ornamental vegetation. These watersheds are part of the city, despite that they are not entirely covered by urban areas. Our experimental set up is similar to others that studies have previously used to assess the effects of this urbanization gradient on terrestrial (Gaona et al., 2021) and aquatic (Zúñiga-Sarango et al., 2020) insects, as well as on riparian vegetation (Chuquimarca et al., 2019) and stream ecosystems (Iñiguez-Armijos et al., 2022).
Bird Sampling
Birds were surveyed along 1 km transects set at each sampling site. Since changes in the structural conditions of the study sites may influence the detectability of the birds, we have used two different sampling methods (auditory and visual surveys), which allowed us to reduce potential detectability bias. We carried out the sampling in two periods between 2016 (May–July) and 2017 (June–August). Surveys started at 05:45 a.m. and finished at 10:00 a.m., and birds were recorded while walking at a speed of 100 m/6 min. All the surveys were led by the same person (LO-D), an experienced researcher with several years of experience studying the birds of southern Ecuador and contributing to on-line databases for bird vocalizations. Birds were identified using field guides (Ridgely and Greenfield, 2006), collections of bird songs (Moore et al., 2013), and the reference collection for the city of Loja.1 Trophic guilds (frugivores, granivorous, insectivorous, nectarivores, and omnivorous) and foraging strata (aerial, canopy, midstory, understory, and terrestrial) of each species detected were assigned to each recorded species according to Stotz et al. (1996) and Billerman et al. (2021). Although birds may have varied diets or use other strata than those used here for analysis, we used the primary guild and stratum for each species. For example, Tyrannus melancholicus (Tyrannidae), a species that feeds mainly on insects in the canopy, is defined here as a canopy-foraging insectivore, although it may sporadically feed on ripe fruits found in lower strata. Aerial strata refer mainly to insectivorous birds that feed in the air, canopy refers to birds that forage in the upper canopy of trees, midstory refers to birds that forage within the canopy and trunk of trees, understory refers to birds that forage in the shrubbier part of the forest, and terrestrial refers to birds that forage on the ground.
Data Analysis
Sampling effort was assessed by calculating the coverage for each level of urbanization using the rarefy function (Oksanen et al., 2020). The species richness was extrapolated using the Chao2 estimator and was used to calculate the sampling coverage as observed species richness divided by estimated species richness. We built generalized linear models (GLMs) to evaluate the effect of each land-use type on the species richness, abundance and evenness. We included in the model the sampling year as covariate. We pooled the visual and auditory recordings at the transect level. We assumed each auditory record registered at least 100 m apart as an independent individual. In the case of birds with a loud vocalization, we took care to only record them once. We calculated the species richness and abundance for each sampling site and year. The species evenness was calculated as the division of the Shannon diversity index by the logarithm of species richness.
Generalized linear models were fitted assuming a Poisson error distribution for count data and gamma error distribution for continuous data. Additionally, we calculated the proportion of individuals and species by foraging strata and trophic guild. In this case, we adjusted the GLMs using a binomial error distribution for proportions. Parameters for the GLMs were estimated using Laplace approximations (Bolker et al., 2008). We then performed a post hoc test using the function glht of the “multcomp” R package (Hothorn et al., 2008, 2017) to examine significant differences between land-use types. Data analyses were performed in R (R Core Team, 2020).
Results
We recorded a total of 1,257 individuals of 74 bird species belonging to 30 families (Supplementary Material). We found changes in the total species richness along the urbanization gradient, with a total of 54 species in forest sites, 47 in forest-pasture sites, 36 in pasture sites, and 29 in urban sites. According to rarefaction, the differences between urbanization levels were significant (95% confidence intervals did not overlap) (Figure 2). The Chao2 estimate of species richness varies from 55 to 30 species along the urbanization gradient, showing a sampling coverage greater than 90%, being highest in forest (0.98) and lowest in forest-pasture (0.90), indicating an adequate sampling effort for all urbanization levels. Most bird species recorded use the understory (44%) followed by the terrestrial stratum (36%), with no difference in the proportion of species using the aerial, canopy, and midstory strata (c.a. 7% each).
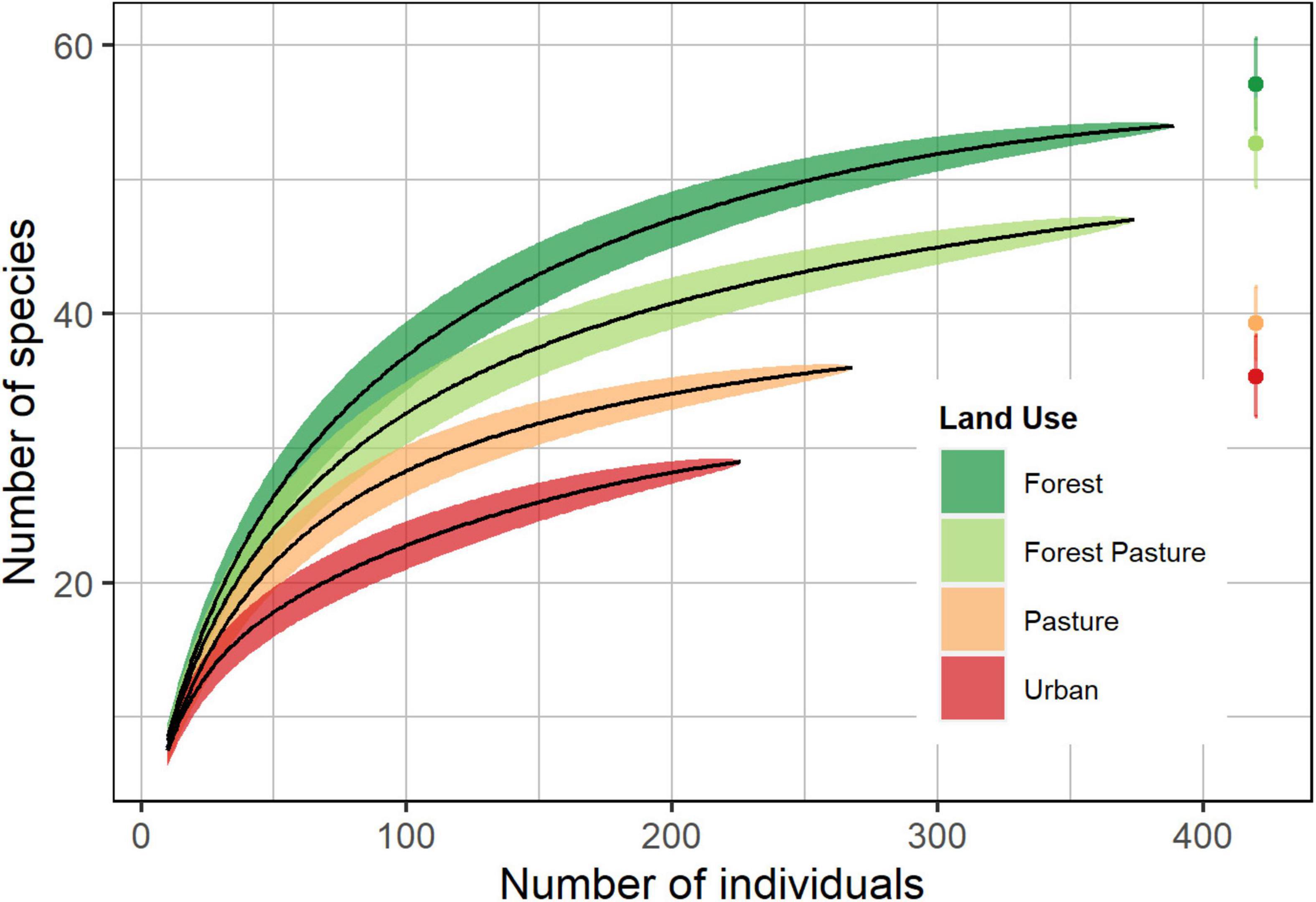
Figure 2. Rarefaction curves (solid lines) for species richness calculated by using the Chao2 estimator with 95% confidence intervals (shaded areas) for the four land-use types along an urbanization gradient in the southern Ecuadorian Andes.
Our models showed a strong negative effect of urbanization on bird species richness (Figure 3A). Species richness was significantly higher in forest sites, followed by the forest-pasture sites, and with lower numbers of species at pastures and urban sites, respectively. Bird abundance showed a similar pattern (Figure 3B) with the number of individuals decreasing from forest to urban sites. Significantly higher abundance was detected in forest and forest-pasture sites compared to pasture and urban sites. Species evenness only showed significant differences between the forest and the urban sites, with the bird community being more even toward the most disturbed sites (Figure 3C).
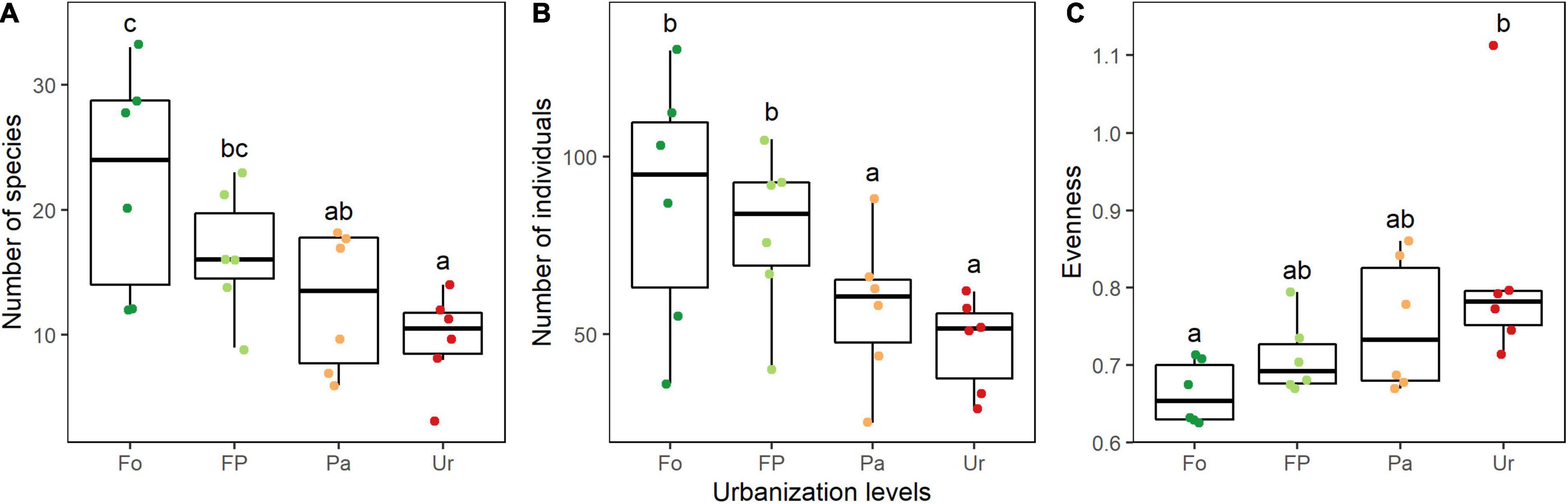
Figure 3. Land-use effects on richness (A), abundance (B), and evenness (C) of the bird communities along an urbanization gradient in the southern Ecuadorian Andes. Data is depicted through quartiles and different lowercase letters above the whiskers indicate significant differences between land uses (GLM p < 0.05). Fo, forest sites; FP, forest-pasture sites; Pa, pasture sites; Ur, urban sites. All sampling times (two) and sites (three) for each land use are used for plotting.
The proportion of the richness of the trophic guilds was significantly affected by urbanization only in granivorous birds (Figure 4A). We found a large amount of variation in sampling sites within the same level of urbanization. Proportion of abundance was affected in three of the four trophic guilds (Figures 4A–H). Granivorous birds increased in abundance in urban sites, while insectivorous birds decreased in abundance from forest to urban sites (Figures 4A,B,E,F). The omnivores showed a reduction in abundance in the first levels of urbanization, but the abundance of this guild was higher in the urban areas (Figures 4E,F,H).
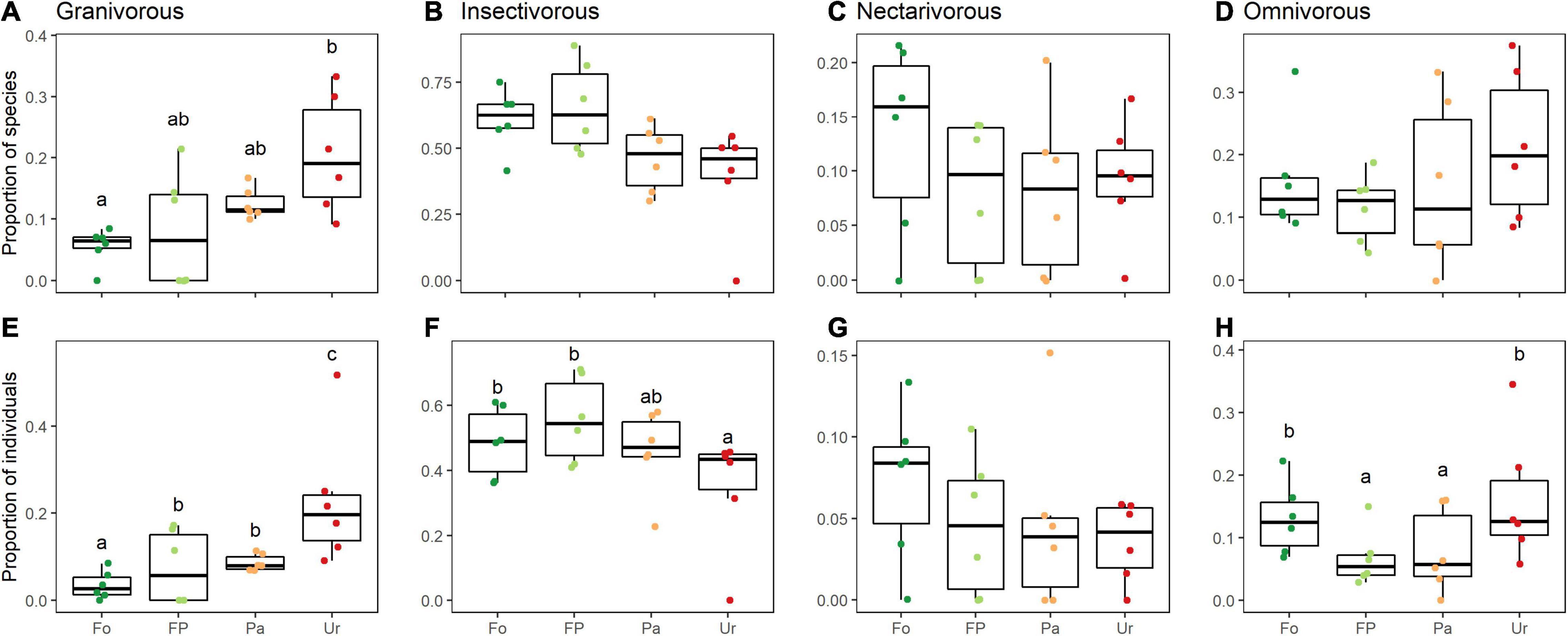
Figure 4. Land-use effects on richness (A–D) and abundance (E–H) of bird trophic guilds along an urbanization gradient in the southern Ecuadorian Andes. Carnivore (A), frugivore (B), granivore (C), insectivore (D), nectarivore (E), and omnivore (F) birds. Data is depicted through quartiles and different lowercase letters above (or below) the whiskers indicate significant differences between land uses (GLM p < 0.05). Fo, forest sites; FP, forest-pasture sites; Pa, pasture sites; Ur, urban sites.
Our models showed significant effects of urbanization in proportion of richness in two foraging guilds and proportion of abundance in all foraging guilds (Figures 5A–J). The proportion of richness in the understory and terrestrial foraging guilds changed significantly along the urbanization gradient. Understory species decreased and terrestrial species increased (Figures 5D,E). Abundance of all foraging guilds was significantly affected by urbanization, although the direction of the response depended on the guild. The midstory and terrestrial guilds increased in abundance along the gradient, while aerial, canopy, and understory guilds decreased in abundance (Figures 5F–J).
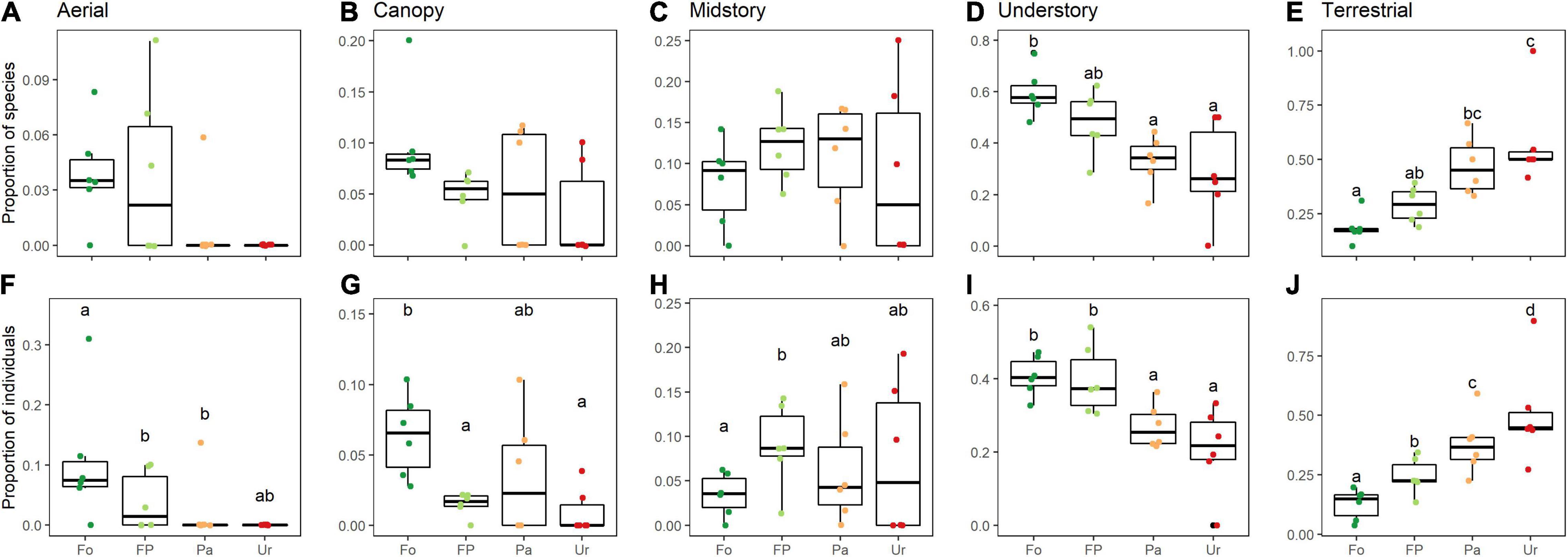
Figure 5. Land-use effects on richness (A–E) and abundance (F–J) of bird foraging strata along an urbanization gradient in the southern Ecuadorian Andes. Aerial (A,F), canopy (B,G), midstory (C,H), understory (D,I), and terrestrial (E,J) habitats. Panels (A–E) show models from species richness and (F–J) show models from abundance. Data is depicted through quartiles and different lowercase letters above (or below) the whiskers indicate significant differences between land uses (GLM p < 0.05). Fo, forest sites; FP, forest-pasture sites; Pa, pasture sites; Ur, urban sites.
We included the year of sampling in all models to test interannual changes and quantify the possible effect of temporal collinearity between samples in the same study site. In general, the sampling year did not affect the richness and abundance of birds and guilds.
Discussion
Urban areas across the planet contain substantially lower species densities as compared to non-urban surroundings (Chace and Walsh, 2006; Aronson et al., 2014; Lee and Rotenberry, 2015). Species loss can exceed 75% relative to species pools in the original habitats that cities have replaced (Newbold et al., 2015). Our work also demonstrates a drastic loss of bird diversity along the urbanization gradient in a megadiverse tropical region. However, these effects are not the same for all species and can change depending on the guild they belong to. The urbanization was “good” for some guilds, such as granivorous and terrestrial species; the areas most urbanized show higher richness and/or abundance for these guilds. The urbanization was “bad” for the others guilds, with a reduction in the species richness and/or abundance. Finally, the urbanization was “ugly” for whole bird community, the richness and abundance decreasing drastically with the increase of urbanization.
According to our expectations, tropical areas should show a greater loss of diversity as a consequence of the levels of specialization that birds present in tropical ecosystems. Although comparison with other ecosystems and studies with different methods and sampling efforts have limitations, we found some indications that the magnitude of loss is greater in our ecosystem. In the studies by Mikami and Mikami (2014) in Fukuoka, Japan and by Blair (1996) in Palo Alto, California, they found a decrease in 10 of the 22 and 21 species found in forested areas, respectively. In the case of cities of similar size to Loja such as Örebro in Sweden (155,989 inhabitants), the bird community showed similar losses reaching values as high as 66% (Sandström et al., 2006). The reported changes are similar to those found in our study where 25 of the 54 species found in the forest sites are lost. Although the loss ratios of these three studies are similar, the disturbance gradient that is used in each is very different. While the studies in Fukuoka, Örebro, and Palo Alto cover fully urbanized areas, in our study the areas with the highest urbanization are not fully urbanized, thus we would expect this loss to be even stronger. On the other hand, the history of intervention and the size of the cities is very different, while Loja is a relatively new small city, Palo Alto and Fukuoka are older and larger than Loja. Other studies have found high losses of bird diversity in temperate cities. In Chile (Silva et al., 2016), where three cities were studied (Temuco, Osorno, and Valdivia) with populations between 100,000 and 300,000 inhabitants, results showed bird species loss of 40, 17, and 15%, respectively. In Argentina, bird species losses of at least 40% of the total species richness were recorded for the cities of Mar del Plata (500,000 inhabitants), Balcarce (35,000 inhabitants), and Miramar (24,000 inhabitants), when comparing natural and the urban sites (Leveau et al., 2017). Although the data presented here show some dissimilarities in the percentages of species loss between less undisturbed areas and urban areas, all the studies reviewed here are consistent in their findings. Urban environments always tend to harbor a smaller number of species than forested areas or areas with greater vegetation cover surrounding the cities.
While bird community response has been consistent across different regions, these effects are expected to be species dependent, with certain species being favored by new habitats and resources, and others being harmed by habitat-level alterations (Chace and Walsh, 2006; Walker and Shochat, 2010; Valente-Neto et al., 2021). Our results showed trophic guild-dependent responses. Granivorous bird diversity and abundance increased toward the most urbanized sites. This response is consistent with what was found in other urbanized areas (Chace and Walsh, 2006; Fox and Hockey, 2007; Blair and Johnson, 2008). The group of granivorous terrestrial-feeding birds within the city of Loja is composed mainly of generalist species (e.g., Rock Dove Columba livia, Eared Dove Zenaida auriculata, Rufous-collared Sparrow Zonotrichia capensis), which have adapted quite well to coexistence with humans. These conditions have favored an increase in the population densities of these species in the last decade, similar to what has been reported for other generalist species, like Passer domesticus and Sturnus vulgaris in cities in the United Kingdom (Evans et al., 2009), C. livia, and P. domesticus in Chile (Silva et al., 2016) or in Mexico (Escobar-Ibáñez et al., 2020).
In the case of omnivorous birds, we found an “U” shape response with a decrease in abundance in the first levels of urbanization, but another increase of abundance in more urbanized areas. These results differ from some studies that reported that abundance and species richness were lower in the least urbanized areas (Walker and Shochat, 2010; Huang et al., 2015). The shift in this pattern could be a consequence of the appearance of exotic generalist bird species, associated with the consumption of food provided directly or indirectly by humans. The omnivorous birds found in the forest sites are native species (e.g., Aulacorhynchus prasinus, Cyclarhis gujanensis), while the bird community in the urban sites is dominated by exotic species associated with humans and the resources that they provide (e.g., C. livia). This change in the trophic organization, again, is influenced by the difference in the presence of food resources and perching or nesting sites. Previous studies have already reported that urbanization favors terrestrial-feeding omnivorous and granivorous birds (Beissinger and Osborne, 1982; Lim and Sodhi, 2004). Omnivorous birds in forested areas use different strata to forage, while omnivorous birds in the city efficiently use the resources available in the urban areas, feeding in trees in the parks or on the ground.
As for insectivorous birds, our hypothesis was corroborated. In the Tropical Andes, the richness and abundance of this trophic guild decreased toward sites with higher degrees of urbanization, as reported by Amaya-Espinel et al. (2019) in Chile or by Máthé and Batáry (2015) in Romania. Many resident insectivorous birds are dependent on insect populations, both for their own food (Tallamy, 2004) and for feeding their nestlings (Dickinson, 1999). Hence the scarcity of vegetation, and particularly the scarcity of native species in urban green spaces, directly causes a decrease in insect diversity, which in turn negatively influences the diversity of birds, particularly insectivorous birds (Burghardt et al., 2009; Sherry, 2021). In our study area, Gaona et al. (2021) found a severe decrease in the diversity, abundance and species richness of moth species, which could reflect a reduction of insects because of urbanization in the city of Loja. Despite the decreases in insectivorous birds, some studies have showed an increased diversity of insectivorous birds in the most-urbanized areas (Hayes et al., 2020), for example, in Guyana (Escobar-Ibáñez and MacGregor-Fors, 2016) and various locations in Mexico (Escobar-Ibáñez et al., 2020). Leveau (2013) attributed the presence and distribution of remnants of native vegetation within or near urban areas as the main factor causing this phenomenon. Cities with the presence of native vegetation in their parks have the possibility of providing a greater amount of food resources to bird species. Many of the changes that were observed in trophic guilds can be partially explained by changes in the supply of food resources. However, further research is needed to quantify the changes in the supply of resources along the urbanization gradient and how these affect the trophic guilds.
Our results showed very clear responses to urbanization of birds using the lower strata (understory and terrestrial). As urbanization increases, the species richness and abundance of these groups changes abruptly, but in opposite directions. While urbanization favors terrestrial bird species, understory bird species show a reduction in diversity and abundance. We believe that the lack of trees within the city has a direct effect on birds that occupy the upper strata. Although we did not find significant differences in the richness of birds using the canopy and aerial strata between urbanization levels, the trend of reduction in their abundance at the most urbanized sites suggests that canopy birds and those that feed during flight are negatively affected by the urbanization process. This could be also explained by the lack of trees and food resources (e.g., insects) that have been drastically reduced in these urban sites (Gaona et al., 2021). Although the changes in resource availability in response to the structural changes of vegetation and other resources along the urbanization gradient is a plausible explanation, other authors have proposed that the restriction of nesting sites in urban areas could explain the changes in the diversity and abundance of the birds. It is necessary to carry out further studies to assess the extent to which nesting habits could be a taxonomic diversity filtering factor.
Our work shows the “ugly” side of the urbanization: a severe reduction in the diversity of birds along the urbanization gradient, which is consistent in cities with different demographic characteristics and environmental features. On the other hand, the effect of urbanization showed that the responses of the birds are species-dependent. The “bad” side of the urbanization is the reduction of richness and abundance of most guilds. Finally, for granivorous and terrestrial trophic guilds, the urbanization shows its “good” side as there is an increase of species richness and abundance. Although the answers found here are consistent with other studies, it has been observed that in certain cities the favored or affected groups may vary. It is necessary to develop a more in-depth study that allows us to understand what the factors within the urbanization process are that are causing this variation in the responses of the bird community.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval was not required for the animal study because the study did not contemplate the direct manipulation of the animals, only their visual and/or auditory recording.
Author Contributions
CI-A and LW provided the funding acquisition. CI-A, CE, and LO-D contributed to the conception and design of the study and wrote the first draft of the manuscript. LO-D and EG performed the fieldwork. CE and LO-D organized the database. CE and CI-A performed the statistical analysis. MD, AE, LW, and EG revised and edited the new versions of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This work was funded by the Universidad Técnica Particular de Loja through project: Convive con los ríos (PY2066) and a National Science Foundation grant #1460079.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
To the Departamento de Ciencias Biológicas y Agropecuarias of the Universidad Técnica Particular de Loja for the institutional support and backing in the development of this work. LO-D thanks Leonardo and Alejandra Ordóñez-Jaramillo for their invaluable support in the execution of the field work. This manuscript is a contribution by MD to the project URBILAND (PID2019-107423GA-I00) by the MCIN/AEI/10.13039/501100011033.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.844944/full#supplementary-material
Footnotes
References
Amaya-Espinel, J. D., Hostetler, M., Henríquez, C., and Bonacic, C. (2019). The influence of building density on neotropical bird communities found in small urban parks. Landsc. Urban Plan. 190:103578. doi: 10.1016/j.landurbplan.2019.05.009
Aronson, M. F. J., La Sorte, F. A., Nilon, C. H., Katti, M., Goddard, M. A., Lepczyk, C. A., et al. (2014). A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 281:20133330. doi: 10.1098/rspb.2013.3330
Arroyo-Rodríguez, V., Rös, M., Escobar, F., Melo, F. P. L., Santos, B. A., Tabarelli, M., et al. (2013). Plant β-diversity in fragmented rain forests: testing floristic homogenization and differentiation hypotheses. J. Ecol. 101, 1449–1458. doi: 10.1111/1365-2745.12153
Aycart, P., and Díaz, M. (2021). Low-intensity landscaping of research facilities increased taxonomic, functional and phylogenetic bird diversity in a lowland rainforest. Ardeola 68, 355–371. doi: 10.13157/arla.68.2.2021.ra3
Banville, M. J., Bateman, H. L., Earl, S. R., and Warren, P. S. (2017). Decadal declines in bird abundance and diversity in urban riparian zones. Landsc. Urban Plan. 159, 48–61. doi: 10.1016/j.landurbplan.2016.09.026
Bax, V., and Francesconi, W. (2019). Conservation gaps and priorities in the tropical andes biodiversity hotspot: implications for the expansion of protected areas. J. Environ. Manage. 232, 387–396. doi: 10.1016/j.jenvman.2018.11.086
Beaumont, L. J., Pitman, A., Perkins, S., Zimmermann, N. E., Yoccoz, N. G., and Thuiller, W. (2011). Impacts of climate change on the world’s most exceptional ecoregions. Proc. Natl. Acad. Sci. U S A. 108, 2306–2311. doi: 10.1073/pnas.1007217108
Beissinger, S. R., and Osborne, D. R. (1982). Effects of urbanization on avian community organization. Condor 84:75. doi: 10.2307/1367825
Beninde, J., Veith, M., and Hochkirch, A. (2015). Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 18, 581–592. doi: 10.1111/ele.12427
Billerman, S. M., Keeney, B. K., Rodewald, P. G., and Schulenberg, T. S. (2021). Birds of the World. Ithaca, NY: Cornell Laboratory of Ornithology.
BirdLife International and Conservation International (2005). Áreas Importantes para la Conservación de las Aves en los Andes Tropicales: Sitios Prioritarios Para la Conservación de la Biodiversidad (Serie de Conservación de BirdLife No 14). Ecuador: Quito.
BirdLife International (2021a). Endemic Bird Areas factsheet: Central Andean páramo. Cambridge: BirdLife International.
BirdLife International (2021b). Endemic Bird Areas factsheet: Southern Central Andes. Cambridge: BirdLife International.
Blair, R. B. (1996). Land-use and avian species diversity along an urban gradient. Ecol. Appl. 6, 506–519. doi: 10.2307/2269387
Blair, R. B. (2001). “Birds and butterflies along urban gradients in two ecoregions of the united states: is urbanization creating a homogeneous fauna?,” in Biotic Homogenization, eds J. L. Lockwood and M. L. McKinney (Boston, MA: Springer), 33–56. doi: 10.1007/978-1-4615-1261-5_3
Blair, R. B., and Johnson, E. M. (2008). Suburban habitats and their role for birds in the urban–rural habitat network: points of local invasion and extinction? Landsc. Ecol. 23, 1157–1169. doi: 10.1007/s10980-008-9267-y
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, H. H., et al. (2008). Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135. doi: 10.1016/j.tree.2008.10.008
Burghardt, K. T., Tallamy, D. W., and Gregory Shriver, W. (2009). Impact of native plants on bird and butterfly biodiversity in suburban landscapes. Conserv. Biol. 23, 219–224. doi: 10.1111/j.1523-1739.2008.01076.x
Carranza-Quiceno, J. A., Henao-Isaza, J. R., and Castaño, J. H. (2018). Avifauna de un paisaje rural heterogéneo en Risaralda, cordillera Central de Colombia. Biota Colomb. 19, 92–104. doi: 10.21068/c2018.v19n02a08
Carvajal-Castro, J. D., Ospina-L, A. M., Toro-López, Y., Pulido-G, A., Cabrera-Casas, L. X., Guerrero-Peláez, S., et al. (2019). Birds vs bricks: patterns of species diversity in response to urbanization in a Neotropical Andean city. PLoS One 14:e0218775. doi: 10.1371/journal.pone.0218775
Chace, J. F., and Walsh, J. J. (2006). Urban effects on native avifauna: a review. Landsc. Urban Plan. 74, 46–69. doi: 10.1016/j.landurbplan.2004.08.007
Chiari, C., Dinetti, M., Licciardello, C., Licitra, G., and Pautasso, M. (2010). Urbanization and the more-individuals hypothesis. J. Anim. Ecol. 79, 366–371. doi: 10.1111/j.1365-2656.2009.01631.x
Chuquimarca, L., Gaona, F. P., Iñiguez-Armijos, C., and Benítez, Á (2019). Lichen responses to disturbance: clues for biomonitoring land-use effects on riparian andean ecosystems. Diversity 11:73. doi: 10.3390/d11050073
Cincotta, R. P., Wisnewski, J., and Engelman, R. (2000). Human population in the biodiversity hotspots. Nature 404, 990–992. doi: 10.1038/35010105
Concepción, E. D., Götzenberger, L., Nobis, M. P., de Bello, F., Obrist, M. K., and Moretti, M. (2017). Contrasting trait assembly patterns in plant and bird communities along environmental and human-induced land-use gradients. Ecography (Cop.) 40, 753–763. doi: 10.1111/ecog.02121
De León, L. F., Sharpe, D. M. T., Gotanda, K. M., Raeymaekers, J. A. M., Chaves, J. A., Hendry, A. P., et al. (2019). Urbanization erodes niche segregation in Darwin’s finches. Evol. Appl. 12, 1329–1343. doi: 10.1111/eva.12721
de Oliveira Ramos, C. C., and dos Anjos, L. (2014). The width and biotic integrity of riparian forests affect richness, abundance, and composition of bird communities. Nat. Conserv. 12, 59–64. doi: 10.4322/natcon.2014.011
de Toledo, M. C. B., Donatelli, R. J., and Batista, G. T. (2012). Relation between green spaces and bird community structure in an urban area in Southeast Brazil. Urban Ecosyst. 15, 111–131. doi: 10.1007/s11252-011-0195-2
Díaz, M., Ramos, A., and Concepción, E. (2022). Changing urban bird diversity: how to manage adaptively our closest relation with wildlife. Ecosistemas Preprint 1–21. doi: 10.13140/RG.2.2.24140.74888
Dickinson, M. B. (1999). Field Guide to the Birds of North America, 3rd Edn. Washington, D.C: National Geographic Society.
Escobar-Ibáñez, J. F., and MacGregor-Fors, I. (2016). Peeking into the past to plan the future: assessing bird species richness in a neotropical city. Urban Ecosyst. 19, 657–667. doi: 10.1007/s11252-015-0517-x
Escobar-Ibáñez, J. F., and MacGregor-Fors, I. (2017). “What’s new? an updated review of avian ecology in urban Latin America,” in Avian Ecology in Latin American Cityscapes, (Cham: Springer International Publishing), 11–31. doi: 10.1007/978-3-319-63475-3_2
Escobar-Ibáñez, J. F., Rueda-Hernández, R., and MacGregor-Fors, I. (2020). The greener the better! avian communities across a neotropical gradient of urbanization density. Front. Ecol. Evol. 8:500791. doi: 10.3389/fevo.2020.500791
Evans, K., Newson, S., and Gaston, K. (2009). Habitat influences on urban avian assemblages. Ibis (Lond. 1859) 151, 19–39. doi: 10.1111/j.1474-919X.2008.00898.x
Fajardo, D., Johnston González, R., Neira, L., Chará, J., and Murgueitio, E. (2009). Influencia de sistemas silvopastoriles en la diversidad de aves en la cuenca del río La Vieja, Colombia. Recur. Nat. y Ambient. 58, 9–16.
Forsman, J. T., Reunanen, P., Jokimäki, J., and Mönkkönen, M. (2010). The effects of small-scale disturbance on forest birds: a meta-analysis. Can. J. For. Res. 40, 1833–1842. doi: 10.1139/X10-126
Fox, R. (2013). The decline of moths in Great Britain: a review of possible causes. Insect Conserv. Divers. 6, 5–19. doi: 10.1111/j.1752-4598.2012.00186.x
Fox, S.-J., and Hockey, P. (2007). Impacts of a South African coastal golf estate on shrubland bird communities. South Afr. J. Sci. 103, 27–34.
Gaona, F. P., Iñiguez-Armijos, C., Brehm, G., Fiedler, K., and Espinosa, C. I. (2021). Drastic loss of insects (Lepidoptera: Geometridae) in urban landscapes in a tropical biodiversity hotspot. J. Insect Conserv. 25, 395–405. doi: 10.1007/s10841-021-00308-9
Grimm, N. B., Faeth, S. H., Golubiewski, N. E., Redman, C. L., Wu, J., Bai, X., et al. (2008). Global change and the ecology of cities. Science 319, 756–760. doi: 10.1126/science.1150195
Hayes, W. M., Fisher, J. C., Pierre, M. A., Bicknell, J. E., and Davies, Z. G. (2020). Bird communities across varying landcover types in a neotropical city. Biotropica 52, 151–164. doi: 10.1111/btp.12729
Hermes, C., Keller, K., Nicholas, R. E., Segelbacher, G., and Schaefer, H. M. (2018). Projected impacts of climate change on habitat availability for an endangered parakeet. PLoS One 13:e0191773. doi: 10.1371/journal.pone.0191773
Hothorn, T., Bretz, F., and Westfall, P. (2008). Simultaneous inference in general parametric models. Biometrical J. 50, 346–363. doi: 10.1002/bimj.200810425
Hothorn, T., Bretz, F., Westfall, P., Heiberger, R. M., Schützenmeister, A., and Scheibe, S. (2017). Multcomp: Simultaneous Inference in General Parametric Models. R package version 1. 4–8.
Huang, Y., Zhao, Y., Li, S., and von Gadow, K. (2015). The effects of habitat area, vegetation structure and insect richness on breeding bird populations in Beijing urban parks. Urban For. Urban Green 14, 1027–1039. doi: 10.1016/j.ufug.2015.09.010
Hutto, D., and Barrett, K. (2021). Do urban open spaces provide refugia for frogs in urban environments? PLoS One 16:e0244932. doi: 10.1371/journal.pone.0244932
Ibáñez-Álamo, J. D., Rubio, E., Benedetti, Y., and Morelli, F. (2017). Global loss of avian evolutionary uniqueness in urban areas. Glob. Chang. Biol. 23, 2990–2998. doi: 10.1111/gcb.13567
Iñiguez-Armijos, C., Leiva, A., Frede, H., Hampel, H., and Breuer, L. (2014). Deforestation and benthic indicators: how much vegetation cover is needed to sustain healthy andean streams? PLoS One 9:e105869. doi: 10.1371/journal.pone.0105869
Iñiguez-Armijos, C., Rausche, S., Cueva, A., Sánchez-Rodríguez, A., Espinosa, C., and Breuer, L. (2016). Shifts in leaf litter breakdown along a forest–pasture–urban gradient in Andean streams. Ecol. Evol. 6, 4849–4865. doi: 10.1002/ece3.2257
Iñiguez-Armijos, C., Tapia-Armijos, M. F., Wilhelm, F., and Breuer, L. (2022). Urbanisation process generates more independently-acting stressors and ecosystem functioning impairment in tropical Andean streams. J. Environ. Manage. 304:114211. doi: 10.1016/j.jenvman.2021.114211
Jetz, W., Wilcove, D. S., and Dobson, A. P. (2007). Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol. 5:e157. doi: 10.1371/journal.pbio.0050157
Kowarik, I. (2008). “On the role of alien species in urban flora and vegetation,” in Urban Ecology, eds J. M. Marzluff, et al. (Boston, MA: Springer), 321–338. doi: 10.1007/978-0-387-73412-5_20
Laurance, W. F., and Bierregaard, R. O. Jr. (1997). “Tropical forest remnants,” in Ecology, Management, and Conservation of Fragmented Communities, eds W. F. Laurance and R. O. Bierregaard Jr. (Chicago, IL: University of Chicago Press).
Lee, M. B., and Rotenberry, J. T. (2015). Effects of land use on riparian birds in a semiarid region. J. Arid Environ. 119, 61–69. doi: 10.1016/j.jaridenv.2015.04.001
Leveau, L. (2013). Relaciones aves–habitat en el sector suburbano de mar del plata, Argentina. Ornitol. Neotrop. 24, 201–212.
Leveau, L. (2018). Urbanization, environmental stabilization and temporal persistence of bird species: a view from Latin America. PeerJ 6:e6056. doi: 10.7717/peerj.6056
Leveau, L., Isla, F. I., and Bellocq, M. I. (2015). Urbanization and the temporal homogenization of bird communities: a case study in central Argentina. Urban Ecosyst. 18, 1461–1476. doi: 10.1007/s11252-015-0469-1
Leveau, L., Leveau, C., Villegas, M., Cursach, J., and Suazo, C. (2017). Bird communities along urbanization gradients: a comparative analysis among three neotropical cities. Ornitol. Neotrop. 28, 77–87.
Lim, H. C., and Sodhi, N. S. (2004). Responses of avian guilds to urbanisation in a tropical city. Landsc. Urban Plan. 66, 199–215. doi: 10.1016/S0169-2046(03)00111-117
Luck, G. W. (2007). A review of the relationships between human population density and biodiversity. Biol. Rev. 82, 607–645. doi: 10.1111/j.1469-185X.2007.00028.x
MacGregor-Fors, I., and Escobar-Ibáñez, J. F. (2017). “Birds from urban latin america, where economic inequality and urbanization meet biodiversity,” in Avian Ecology in Latin American Cityscapes, (Cham: Springer International Publishing), doi: 10.1007/978-3-319-63475-3_1
MacGregor-Fors, I., Morales-Pérez, L., and Schondube, J. E. (2010). Migrating to the city: responses of neotropical migrant bird communities to urbanization. Condor 112, 711–717. doi: 10.1525/cond.2010.100062
Máthé, O., and Batáry, P. (2015). Insectivorous and open-cup nester bird species suffer the most from urbanization. Bird Study 62, 78–86. doi: 10.1080/00063657.2014.1000262
Maure, L. A., Rodrigues, R. C., Alcântara, ÂV., Adorno, B. F. C. B., Santos, D. L., Abreu, E. L., et al. (2018). Functional redundancy in bird community decreases with riparian forest width reduction. Ecol. Evol. 8, 10395–10408. doi: 10.1002/ece3.4448
McClure, C. J. W., Korte, A. C., Heath, J. A., and Barber, J. R. (2015). Pavement and riparian forest shape the bird community along an urban river corridor. Glob. Ecol. Conserv. 4, 291–310. doi: 10.1016/j.gecco.2015.07.004
McDonnell, M. J., and Hahs, A. K. (2008). The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes: current status and future directions. Landsc. Ecol. 23, 1143–1155.
McIntyre, N. E. (2000). Ecology of urban arthropods: a review and a call to action. Ann. Entomol. Soc. Am. 93, 825–835.
McKinney, M. L. (2002). Urbanization, biodiversity, and conservation: the impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 52, 883–890.
Mikami, O. K., and Mikami, K. (2014). Structure of the Japanese avian community from city centers to natural habitats exhibits a globally observed pattern. Landsc. Ecol. Eng. 10, 355–360. doi: 10.1007/s11355-012-0201-8
Mittermeier, R. A., Turner, W. R., Larsen, F. W., Brooks, T. M., and Gascon, C. (2011). “Global biodiversity conservation: the critical role of hotspots,” in Biodiversity Hotspots, (Berlin: Springer), 3–22. doi: 10.1007/978-3-642-20992-5_1
Møller, A. P., and Díaz, M. (2018a). Avian preference for close proximity to human habitation and its ecological consequences. Curr. Zool. 64, 623–630. doi: 10.1093/cz/zox073
Møller, P., and Díaz, M. (2018b). Niche segregation, competition, and urbanization. Curr. Zool. 64, 145–152. doi: 10.1093/cz/zox025
Møller, A. P., Díaz, M., and Liang, W. (2016). Brood parasitism and proximity to human habitation. Behav. Ecol. 27, 1314–1319. doi: 10.1093/beheco/arw049
Møller, A., Díaz, M., Grim, T., Dvorská, A., Flensted-Jensen, E., Ibáñez-Álamo, J., et al. (2015). Effects of urbanization on bird phenology: a continental study of paired urban and rural populations. Clim. Res. 66, 185–199. doi: 10.3354/cr01344
Montaño-Centellas, F. A., and Garitano-Zavala, Á (2015). Andean bird responses to human disturbances along an elevational gradient. Acta Oecol. 65–66, 51–60. doi: 10.1016/j.actao.2015.05.003
Moore, J. V., Krabbe, N., and Jahn, O. (2013). Bird Sounds of Ecuador, a Comprehensive Collection. San José, CA: John V. Moore Nat. Rec.
Morelli, F., Reif, J., Díaz, M., Tryjanowski, P., Ibáñez-Álamo, J. D., Suhonen, J., et al. (2021). Top ten birds indicators of high environmental quality in European cities. Ecol. Indic. 133:108397. doi: 10.1016/j.ecolind.2021.108397
Newbold, T., Hudson, L. N., Hill, S. L. L., Contu, S., Lysenko, I., Senior, R. A., et al. (2015). Global effects of land use on local terrestrial biodiversity. Nature 520, 45–50. doi: 10.1038/nature14324
Ochoa-Cueva, P., Fries, A., Montesinos, P., Rodríguez-Díaz, J. A., and Boll, J. (2015). Spatial estimation of soil erosion risk by land-cover change in the Andes of Southern Ecuador. L. Degrad. Dev. 26, 565–573. doi: 10.1002/ldr.2219
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2020). Vegan: Community Ecology Package. Available online at: https://cran.r-project.org/package=vegan (accessed November 13, 2021).
Ordóñez-Delgado, L., Reyes-Bueno, F., Orihuela-Torres, A., and Armijos-Ojeda, D. (2016). Registros inusuales de aves en la hoya de Loja, Andes sur del Ecuador. ACI Av. Ciencias Ing. 8, 26–36. doi: 10.18272/aci.v8i14.276
Pennington, D. N., and Blair, R. B. (2011). Habitat selection of breeding riparian birds in an urban environment: untangling the relative importance of biophysical elements and spatial scale. Divers. Distrib. 17, 506–518. doi: 10.1111/j.1472-4642.2011.00750.x
Ramírez-Albores, J. E. (2004). Efecto de la Estructura del Paisaje Sobre la Diversidad Alfa, Beta y Gama de Comunidades de Aves en San Fernando. tesis de Maestría, Chiapas: El Colegio de la Frontera Sur. San Cristóbal de las Casas.
Ramirez-Villegas, J., Cuesta, F., Devenish, C., Peralvo, M., Jarvis, A., and Arnillas, C. A. (2014). Using species distributions models for designing conservation strategies of Tropical Andean biodiversity under climate change. J. Nat. Conserv. 22, 391–404. doi: 10.1016/j.jnc.2014.03.007
Ridgely, R., and Greenfield, P. (2006). Aves del Ecuador: Guía de Campo. Quito: Fundación de Conservación Jocotoco.
Rodewald, A. D., and Yahner, R. H. (2001). Influence of landscape composition on avian community structure and associated mechanisms. Ecology 82:3493. doi: 10.2307/2680167
Sandström, U. G., Angelstam, P., and Mikusiński, G. (2006). Ecological diversity of birds in relation to the structure of urban green space. Landsc. Urban Plan. 77, 39–53. doi: 10.1016/j.landurbplan.2005.01.004
Schütz, C., and Schulze, C. H. (2015). Functional diversity of urban bird communities: effects of landscape composition, green space area and vegetation cover. Ecol. Evol. 5, 5230–5239. doi: 10.1002/ece3.1778
Seress, G., Lipovits, Á, Bókony, V., and Czúni, L. (2014). Quantifying the urban gradient: a practical method for broad measurements. Landsc. Urban Plan 131, 42–50. doi: 10.1016/j.landurbplan.2014.07.010
Sherry, T. W. (2021). Sensitivity of tropical insectivorous birds to the anthropocene: a review of multiple mechanisms and conservation implications. Front. Ecol. Evol. 9:662873. doi: 10.3389/fevo.2021.662873
Silva, C. P., Sepúlveda, R. D., and Barbosa, O. (2016). Nonrandom filtering effect on birds: species and guilds response to urbanization. Ecol. Evol. 6, 3711–3720. doi: 10.1002/ece3.2144
Sol, D., Bartomeus, I., González-Lagos, C., and Pavoine, S. (2017). Urbanisation and the loss of phylogenetic diversity in birds. Ecol. Lett. 20, 721–729. doi: 10.1111/ele.12769
Souza, F. L., Valente-Neto, F., Severo-Neto, F., Bueno, B., Ochoa-Quintero, J. M., Laps, R. R., et al. (2019). Impervious surface and heterogeneity are opposite drivers to maintain bird richness in a Cerrado city. Landsc. Urban Plan. 192:103643. doi: 10.1016/j.landurbplan.2019.103643
Stotz, D. F., Parker, T. A. III, Fitzpatrick, J. W., and Moskovits, D. K. (1996). Neotropical Birds: Ecology and Conservation. Chicago, IL: University of Chicago Press.
Tallamy, D. W. (2004). Do alien plants reduce insect biomass? Conserv. Biol. 18, 1689–1692. doi: 10.1111/j.1523-1739.2004.00512.x
Tapia-Armijos, M. F., Homeier, J., Espinosa, C. I, Leuschner, C., and de la Cruz, M. (2015). Correction: deforestation and forest fragmentation in South Ecuador since the 1970s - losing a hotspot of biodiversity. PLoS One 10:e0142359. doi: 10.1371/journal.pone.0142359
Valente-Neto, F., Roque, F., de, O., Pauliquevis, C. F., Oliveira, A. K. M., and de, et al. (2021). Loss of cultural and functional diversity associated with birds across the Urbanization gradient in a tropical city. Front. Ecol. Evol. 9:615797. doi: 10.3389/fevo.2021.615797
Walker, J. S., and Shochat, E. (2010). Scalar effects of vegetation on bird communities in an urbanizing desert ecosystem. Urban Ecosyst. 13, 155–167. doi: 10.1007/s11252-009-0112-0
Yow, D. M. (2007). Urban heat islands: observations, impacts, and adaptation. Geogr. Compass 1, 1227–1251. doi: 10.1111/j.1749-8198.2007.00063.x
Keywords: birds, diversity, trophic guilds, foraging strata, land use, Andes, Ecuador
Citation: Ordóñez-Delgado L, Iñiguez-Armijos C, Díaz M, Escudero A, Gosselin E, Waits LP and Espinosa CI (2022) The Good, the Bad, and the Ugly of Urbanization: Response of a Bird Community in the Neotropical Andes. Front. Ecol. Evol. 10:844944. doi: 10.3389/fevo.2022.844944
Received: 29 December 2021; Accepted: 25 February 2022;
Published: 22 March 2022.
Edited by:
Lucas Leveau, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), ArgentinaReviewed by:
Maria Cecília Toledo, Universidade de Taubaté, BrazilThomas W. Sherry, Tulane University, United States
Scott Wilson, Wildlife Research and Landscape Science, Environment and Climate Change Canada, Canada
Copyright © 2022 Ordóñez-Delgado, Iñiguez-Armijos, Díaz, Escudero, Gosselin, Waits and Espinosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carlos Iván Espinosa, Y2llc3Bpbm9zYUB1dHBsLmVkdS5lYw==