
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 04 April 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.844251
 Viviana Di Pietro1*†
Viviana Di Pietro1*† Helena Mendes Ferreira1†
Helena Mendes Ferreira1† Annette Van Oystaeyen2†Felix Wäckers2†Tom Wenseleers1†
Annette Van Oystaeyen2†Felix Wäckers2†Tom Wenseleers1† Ricardo Caliari Oliveira1,3†
Ricardo Caliari Oliveira1,3†The buff-tailed bumblebee Bombus terrestris presents two distinct colony phenotypes in which some colonies already start producing males very early in the season, while others switch to producing sexuals much later in the season, and specialize mainly in the production of virgin queens. Despite having been extensively investigated in the past, the mechanisms underlying these two distinct phenotypes remain poorly understood. Here we use a combination of behavioral and genetic data to demonstrate that early switchers are in fact diploid male producing colonies, while late switchers produced normal, haploid males. Hence, at a proximate level, early switching to male production was caused by queens mating with males that by chance carried the same sex allele. These patterns were not the result of strong inbreeding within the study population, as the average inbreeding coefficient was very low and in fact slightly negative (−0.11). Moreover, an effect of inbreeding was further excluded because hybrids produced by crossing two distinct outbred populations also produced diploid males in early switching colonies. Finally, we found that diploid males only emerged after a first cohort of workers was produced, while we would have expected diploid males to be produced simultaneously with workers in a 50:50 ratio. We demonstrate that adult diploid males had a distinct cuticular hydrocarbon profile and propose that queens likely cull diploid males during early colony development to reduce the cost of diploid male production.
Eusocial Hymenoptera are characterized by the presence of a remarkable reproductive division of labor between queens, who specialize in reproduction, and mostly sterile workers, who focus on tasks such as brood care, foraging, and nest defense (Wilson, 1971). In some species, however, workers can develop their ovaries and also lay unfertilized male eggs under certain conditions (Wilson, 1971). Nevertheless, the haplodiploid sex-determination system found in hymenopterans leads to workers being more related to their sisters (r = 0.75) than to their brothers (r = 0.25) or even to their own offspring (r = 0.50) in colonies headed by one single-mated queen. Due to this relatedness asymmetry, workers in such species would be expected to invest three times more in the production of new queens than in males, while the queen would be expected to invest equally in the production of both sexes, as she is equally related to sons and daughters (r = 0.50). Empirical data show that in several species this conflict over sex allocation is resolved in favor of the workers, since they care for the brood and can therefore manipulate the sex ratio to their own advantage (Hamilton, 1964; Trivers and Hare, 1976; Bourke and Franks, 1995; Wenseleers et al., 2021).
A striking exception to these predicted female-biased sex ratios is observed in the buff-tailed bumblebee Bombus terrestris (Hymenoptera: Apidae), where it is observed a highly male-biased population sex investment ratios (Bourke, 1997; Beekman and Van Stratum, 1998; Duchateau et al., 2004). The case of B. terrestris is broadly studied for investigating queen-worker conflict (Bloch and Hefetz, 1999; Bourke and Ratnieks, 2001; Alaux et al., 2004b; Amsalem et al., 2009) as this species constitute a perfect model study due to its colony features. First, it is a monogynous species with a singly mated queen (Schmid-Hempel and Schmid-Hempel, 2000) thus the conflict is expected to occur. Second, it is an annual species, thus a higher probability of an actual conflict between queen and workers, as both parties do not have interest in maintaining the colony in the later years (Bulmer, 1981). Third, the workers start to lay their own eggs in a specific moment of the colony lifecycle, called the “competition point”. This phase normally follows the “switch point” which occurs when the queen starts to lay haploid eggs for producing males (Duchateau and Velthuis, 1988; Lopez-Vaamonde et al., 2009), although some diploid eggs may also be laid after the switch point (Van der Blom, 1986; Van Doorn and Heringa, 1986; Duchateau and Velthuis, 1988). Over the years, several hypotheses have been put forward to try to explain this observed male-biased sex ratio. For example, the protandry hypothesis states that males produced early in the season have higher opportunities to mate than late-emerging males, and that colonies producing their males early would therefore gain greater fitness benefits (Bulmer, 1983; Bourke, 1997). However, the protandry hypothesis alone cannot explain the occurrence of a split sex ratio observed in this species, whereby some colonies specialize in the production of males and others in females (Meunier et al., 2008; Kümmerli and Keller, 2009). Indeed, B. terrestris presents two distinct social phenotypes correlated to differential reproductive strategies (Beekman and Van Stratum, 1998; Duchateau et al., 2004). In some colonies the queen “switches” to produce haploid male eggs early in the season (early switching colonies) when the worker/larvae ratio is small and specializes in male production, while other “late switching” colonies produce mostly queens toward the end of the season and start to produce males after queen larvae are detected (Duchateau and Velthuis, 1988; Alaux et al., 2004a,2005). As the two colony phenotypes were found with a frequency of 0.5 in nature (Duchateau and Velthuis, 1988), it was hypothesized that in half of the colonies, the mother queens could prevent the production of new queens by starting to lay haploid eggs earlier. In this scenario, in male-specialized colonies, workers would have no choice but to comply, given that they would have no diploid female eggs available to rear into queens and the replacement with workers’ eggs would not be profitable by the time the workers would be able to detect them (Bulmer, 1981; Pamilo, 1982; Bourke and Ratnieks, 2001). By contrast, in the remaining half of the colonies, the evolutionary interest of both the workers and the mother queen would coincide in benefiting from specializing in queen production (Bourke and Ratnieks, 2001). Although this mechanism seems plausible and a similar system was observed in some ant species (Helms, 1999), it still does not explain why workers in the late switching colonies begin to lay haploid male eggs at the onset of queen production (Bourke and Ratnieks, 2001; Alaux et al., 2004a,2005), resulting in a male-biased sex ratio at the population level (Goulson, 2010).
A further development of Bulmer’s model (Bulmer, 1983) proposes that the timing of the switch point, and consequently the split sex ratios, could be the result of an optimal sex allocation of resources since queens are 1.69–2.11 times more costly to produce than males (Beekman and Van Stratum, 1998; Duchateau et al., 2004). Therefore, colonies with more limited resources available could benefit from switching earlier and specializing in the production of the sex that is least costly to produce–males (Beekman and Van Stratum, 1998). However, in situations with overall low resource availability, such a mechanism would then cause the majority of colonies to produce mainly males, leading to a low reproductive success (Goulson, 2010), and so it is also hard to see how this theory would explain the male-biased sex ratios seen in bumblebees. In support of this, it was shown how environmental factors, including resources availability, have limited or no effect on reproductive decisions in B. terrestris (Schmid-Hempel and Schmid-Hempel, 1998; Baer and Schmid-Hempel, 2003).
Environmental stressors, such as climate change, do not only affect the availability of resources but can also lead to genetic variability constraints (Templeton et al., 1990; Gienapp et al., 2008; Hoffmann and Willi, 2008; Perez and Aron, 2020). A particularly important constraint in social insects is linked to the production of diploid males, which in extreme cases may result in the so-called “diploid male extinction vortex” (Zayed and Packer, 2005). In most hymenopteran species, including ants, bees, wasps, and sawflies, sex is determined by a complementary sex-determination system (CSD), in which heterozygous eggs at the CSD locus develop as females while hemizygous (haploid) or homozygous eggs develop as males (Whiting, 1943). Diploid male production comes at a large cost in social Hymenoptera, as they are produced at the expense of the production of new workers, thereby compromising colony growth (Duchateau et al., 1994; Van Wilgenburg et al., 2006), and on top of that, diploid males are also typically sterile or unable to produce viable offspring (Duchateau and Mariën, 1995; Beekman et al., 1999; Ayabe et al., 2004; Gerloff and Schmid-Hempel, 2005; Whitehorn et al., 2009). Nevertheless, some species have developed mechanisms to reduce the fitness costs of diploid male production to the colony. Such mechanisms are usually linked with inbreeding avoidance (Van Wilgenburg et al., 2006), but in some corbiculate bee species, different strategies have evolved to prevent colony collapse due to diploid male production. In the honeybee Apis mellifera, the queen mates with several males which spreads the risk of making a matched mating i.e., mating with a male carrying the same CSD allele (Page and Metcalf, 1982). In addition, workers are able to detect and cull diploid male larvae early during their development, therefore preventing a higher cost to the colony (Woyke, 1962, 1963). Stingless bees rear their brood in mass provisioned sealed cells and hence workers do not have access to the developing brood (Engels and Imperatriz-Fonseca, 1990). Nevertheless, it has been shown that matched mated queens are swiftly killed and replaced by one of their daughters once diploid males are detected (Vollet-Neto et al., 2017). Differently to honeybees, most of stingless bees and bumblebees are monandrous. Polyandrous species in fact have the advantage to dilute the effect of diploid males production due to a matched allele (Page, 1980; Ratnieks, 1990), while a monandrous mating system, such as in B. terrestris (Estoup et al., 1995; Schmid-Hempel and Schmid-Hempel, 2000), decreases the genetic variation and therefore increases the vulnerability to inbreeding (Chapman and Bourke, 2001). Due to the costs associated with inbreeding in bumblebees, the evolution favored mechanism of inbreeding avoidance, such as nestmate recognition and scent-marking during patrolling for a female (Foster, 1992). However, although B. terrestris males display scent-marking and patrolling behavior to find a partner (Brown and Baer, 2005; Goulson, 2010), it was shown that males do not avoid mating with related individuals (Gerloff and Schmid-Hempel, 2005; Bogo et al., 2018) to the detriment of certain colony traits such as the colony foundation success and the colony size, (Gerloff and Schmid-Hempel, 2005; Gosterit, 2016), although in some occasions inbred colonies can have a normal development (Duchateau et al., 1994; Bortolotti et al., 2020). Honeybees, stingless bees and bumblebees have been shown to be resilient to genetic bottlenecks with examples of successful populations being established from only a few colonies (Winston, 1996; Schmid-Hempel et al., 2007; Alves et al., 2011). These mechanisms reducing the costs of diploid males are likely to play an important role in the maintenance of populations with relatively lower diversity caused by genetic bottlenecks. Here, we use a combination of behavioral, chemical, and genetic data analysis to test if diploid male production causes a switch to early male production in the buff-tailed bumblebee B. terrestris. We hypothesize that the competition point, when workers begin to compete with the queen over male production, in early switching colonies could actually be a chance to produce haploid males and reduce the fitness cost of diploid male production.
To test the role of a genetic component resulting in the distinct colony phenotypes we used a total of 20 colonies of B. terrestris, obtained from the main stock of Biobest Group NV (Westerlo, Belgium) in August 2020. The colonies were selected by the breeding facility to be 10 early switching and 10 late switching. Overall, only 6% of the colonies produced in Biobest are early switching. Identical nest-boxes (23 cm × 30 cm × 20 cm) were placed in a controlled-climate room at constant temperature and relative humidity of 28°C and 50%, respectively, and 16L: 8D light: dark photoperiod. Honeybee-collected pollen and a 50% sugar solution (Biogluc, Belgosuc, Belgium) were provided ad libitum (supplied by Biobest Group NV, Westerlo, Belgium). Colonies were observed once per day to detect the presence of males. We then counted the number of workers present in the nest when the first males were observed. Early switching colonies were frozen 3 weeks after the emergence of the first male to reduce the probability of collecting workers’ sons (considering that male development from egg to adult takes on average 26 days; Duchateau and Velthuis, 1988). We followed the same protocol for the late switching colonies, except that the colonies were frozen 4 weeks after the first male emergence in order to obtain the required sample size of ten males per colony. All colonies had reached the competition phase at the end of the experimental period. Ten males and one worker were randomly sampled and, together with the queen, stored at −80°C for microsatellite genotyping to check ploidy. The remaining workers present in the colonies were stored at −20°C for later dissection to assess the ovary development status.
To check for differences in ovary development between the two colony phenotypes, all workers of the early switching colonies were dissected (total of 789) together with a randomly selected sample of 50 workers per colony of the late switching ones (total of 500). Newly emerged or callow individuals are easily detected by their faded coloration (less than 12 h old) and were excluded, since they would not have had enough time to develop their ovaries (Gosterit et al., 2016). The degree of ovary development of workers from early and late switching colonies was obtained via dissection in a saline solution under a Leica stereomicroscope and photographed with a digital camera (Leica MSV266) connected to the computer. Ovaries were scored based on the stage of the terminal oocytes according to the scale and description provided by Duchateau and Velthuis (1989). Ovaries from 0 to 2 were scored as undeveloped while from 3 to 5 as developed. Regressed ovaries were included in the undeveloped category. We further measured the thorax width (inter-tegulae span) as a proxy of the body size using the software Leica Application Suite. The dissections were performed randomly by the same operator that was blind to the colony type.
The statistical analyses were performed using R v4.0.4 (R Core Team, 2021). To test if the sex ratio (proportion of virgin queens produced) was different between colony types at the end of the experimental period we ran a binomial generalized linear mixed model (GLMM) with the package “lme4” (Bates et al., 2015) with the number of virgin queens and males coded as dependent variable, colony type (i.e., early or late switching) included as a factor and colony ID included as a random intercept. To test whether the number of workers and males produced at the beginning and at the end of the experimental period deviated from a 1:1 ratio expected if the queen had made a matched mating, we ran a binomial generalized linear model, using the number of workers and males coded as the dependent variable, the type of colony (i.e., early or late switching) included as a factor and colony ID coded as a random intercept. We further used the package “emmeans” (Lenth, 2016) to test the significance of a deviation from 50% diploid male production expected if the queen had made a matched mating.
To test for differences in the proportion of workers with developed ovaries in early and late switching colonies we used a binomial GLMM (Bates et al., 2015) using developed or undeveloped ovaries as dependent variable colony type, thorax length and colony size as explanatory variables and colony ID as a random intercept.
To analyze the ploidy of the bumblebees from both colony phenotypes, we genotyped 10 males randomly selected from each colony together with a worker and the mother queen. Three legs per sample were placed in a 2 mL microcentrifuge tube for subsequent DNA extraction. The extraction was performed following the Chelex method (Walsh et al., 2018) and samples were genotyped at six microsatellite loci with the highest levels of heterozygosity described for B. terrestris (BTMS0127, BTMS0141, BTMS0153, BT01, BT09, and BT24; Reber Funk et al., 2006; Stolle et al., 2009). Multiplex polymerase Chain Reaction (PCR) reactions were conducted in a thermocycler Biometra TOne (analytik jena) in a 10 μl total volume of which 1 μl of DNA, using the QIAGEN Multiplex PCR Kit. Samples were amplified using touchdown PCR with the following regime: 15 min of initial denaturation at 95°C, followed by 30 s at 94°C, followed by 3 cycles each of 90 s at 54°C with the annealing temperature decreasing 1°C per cycle and 90 s extension at 72°C and then with 30 cycles each of 30 s at 94°C, 90 s at 51°C and 90 s at 72°C. The PCR reaction was finalized with a final extension of 10 min at 72°C. In order to confirm that the result obtained was not an outcome of inbreeding, we further selected seven early switching colonies for genotyping that were specifically crossed between different populations at Biobest and were frozen 3 weeks after the emergence of the first male, consistent with the previous colonies.
Alleles were analyzed and scored using GeneMapper v5 (APPLIED BIOSYSTEMS). The observed and expected heterozygosity (HO and HS; Nei, 1987), the fixation index (FST; Weir and Cockerham, 1984), and the inbreeding coefficient (FIS; Nei, 1987) values were calculated with the “hierfstat” package (Goudet, 2005) in R v4.0.4, including only the diploid individuals, i.e., queens, workers and diploid males from the two colony types (R Core Team, 2021).
A gas chromatography and mass spectrometry (GC-MS) assay was used to analyze the cuticular profiles of the bumblebees (males, mother queens, and workers) and test for differences among the sexes, castes and colony phenotypes. The same individuals sampled for microsatellite genotyping were used for cuticular hydrocarbon analysis (CHC) plus five randomly selected workers from each colony, which totaled to 320 individuals of which 200 were males, 100 were workers and 20 were mother queens. Of the total samples, the extraction of the compounds did not produce a readable chromatogram for ten workers and two queens, which were excluded from the analysis. Workers and males were extracted in 2 ml while queens were extracted in 6 ml of HPLC-grade hexane (SIGMA-ALDRICH) for 1 min. The samples were evaporated at room temperature under a fume hood overnight until dryness. After that, 1 ml of hexane was used to resuspend the samples which were then analyzed with a gas chromatograph (Thermo Fisher Scientific Trace 1,300 series) coupled with a mass spectrometer (Thermo Fisher Scientific ISQ series MS), using a Restek RXi-5sil MS 20 m with an internal diameter of 0.18 mm and a film thickness of 0.18 μm. The program had an initial temperature of 80°C held for 2 min, followed by four temperature ramps from 80°C to 220°C at 25°C min–1, from 220°C to 250°C at 10°C min–1, from 250°C to 300°C at 3°C min–1 and from 300°C to 350°C at 5°C min–1, after which the final temperature of 350°C was held for 4 min. Helium was used as a carrier gas at a flow rate of 0.9 ml min–1, and a splitless injection with an inlet temperature of 320°C. External linear alkane ladders ranging from octane (n-C8) up to tetracontane (n-C40) injected at three different concentrations (0.1, 0.01, and 0.05 μg/μL) were used to infer cubic spline interpolated retention indices. Peaks in the total ion chromatogram were aligned and integrated using a custom R script. Compounds were identified based on their mass spectra, retention indices and comparisons with library references (using the NIST2014 mass spectral and retention index libraries).
For the chemical analysis, the raw peak areas were transformed to the relative abundances and the 95% confidence intervals (Mean, LCI, UCI) were calculated with the t-distribution. To verify a possible cluster formation based on the CHC profile, an explanatory principal component analysis (PCA) was performed with the package “FactoMineR” (Lê et al., 2008) and visualized with “factoextra” package (Kassambara and Mundt, 2017). The overall differences between groups were calculated through a PERMANOVA with 9,999 permutations using the package “vegan” (Oksanen et al., 2013) and the package “pairwiseAdonis” (Martinez Arbizu, 2020) for the pairwise comparison. To check for univariate differences in the abundance of the cuticular compounds in early and late switching colonies the raw peak areas were first transformed by applying Aitchison transformation (Aitchison, 1982) and then single linear mixed models were run for each compound coded as response variable, colony phenotype as the explanatory variable and colony ID as a random factor. Significance levels were post-hoc corrected using a Bonferroni correction with the package “multcomp” (Hothorn et al., 2008). In addition, we performed a linear discriminant analysis (package “MASS,” Venables and Ripley, 2002) with the subset of compounds that were expressed significantly differently between the two male groups to check if these compounds could be reliably used as recognition cues by the nestmates. Finally, the log2 fold difference in individual relative CHCs abundances in males, workers and queens compared to the average of the three groups was plotted in a heatmap using the “pheatmap” package (Kolde, 2015), to visually display the observed differences between the two colony phenotypes.
The colonies used for this experiment showed a clear bimodal pattern in terms of male emergence. First male emergence started on average 46 days after the starting of the colony and 12 days after first worker emergence in the early switching colonies, while first males only appeared on average 84 days since the colony foundation and 54 days after first worker emergence in late switching colonies. This supports the existence of distinct colony phenotypes in B. terrestris (Figure 1A). At the end of the experiment, all the colonies had reached the competition point, observed through one of the following events: (1) worker oviposition, (2) oophagy, and (3) egg-cell destruction (Van Doorn and Heringa, 1986; Duchateau and Velthuis, 1988; Bloch and Hefetz, 1999). There was no significant difference in the proportion of workers with developed ovaries in early and late switching colonies (GLMM, Z ratio = 0.323, p = 0.747). In fact, we detected a virtually identical percentage of workers with developed ovaries in each colony type: 44% (33 ± 18.3) and 43% (21 ± 4.1) in early and late switching colonies, respectively (Table 1).
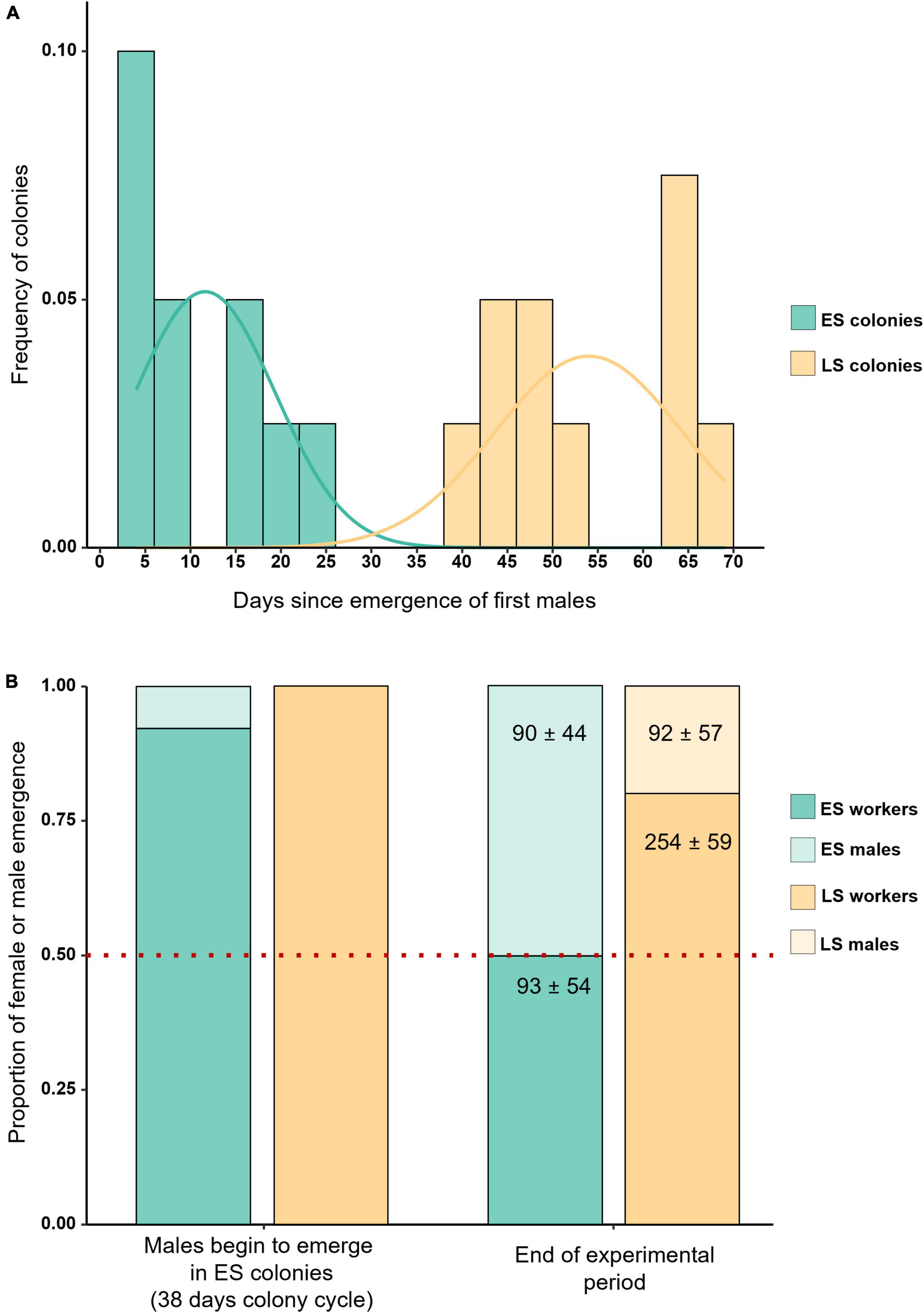
Figure 1. The two colony types present distinct life-history traits. Density plot showing the bimodal distribution of the two colony types according to the date of emergence of the males. Males begin to emerge much earlier after the first worker cohort in early switching (ES) as opposed to late switching colonies (LS) as shown in the density histogram fitted with normal distribution per phenotype (A). Effect plot of the proportion of female or male emergence (y-axis) in two different moments of the observation: when the first males started to emerge in early switching colonies, and at the end of the experimental period when the colonies were frozen (x-axis). The proportion of males being produced by each phenotype varies throughout the cycle with early switching colonies shifting from producing almost exclusively workers earlier on to 50% males and workers at the end of the experiment. Late switching colonies specialize in the production of females (workers and virgin queens) and begin producing males only at the end of the cycle (B).
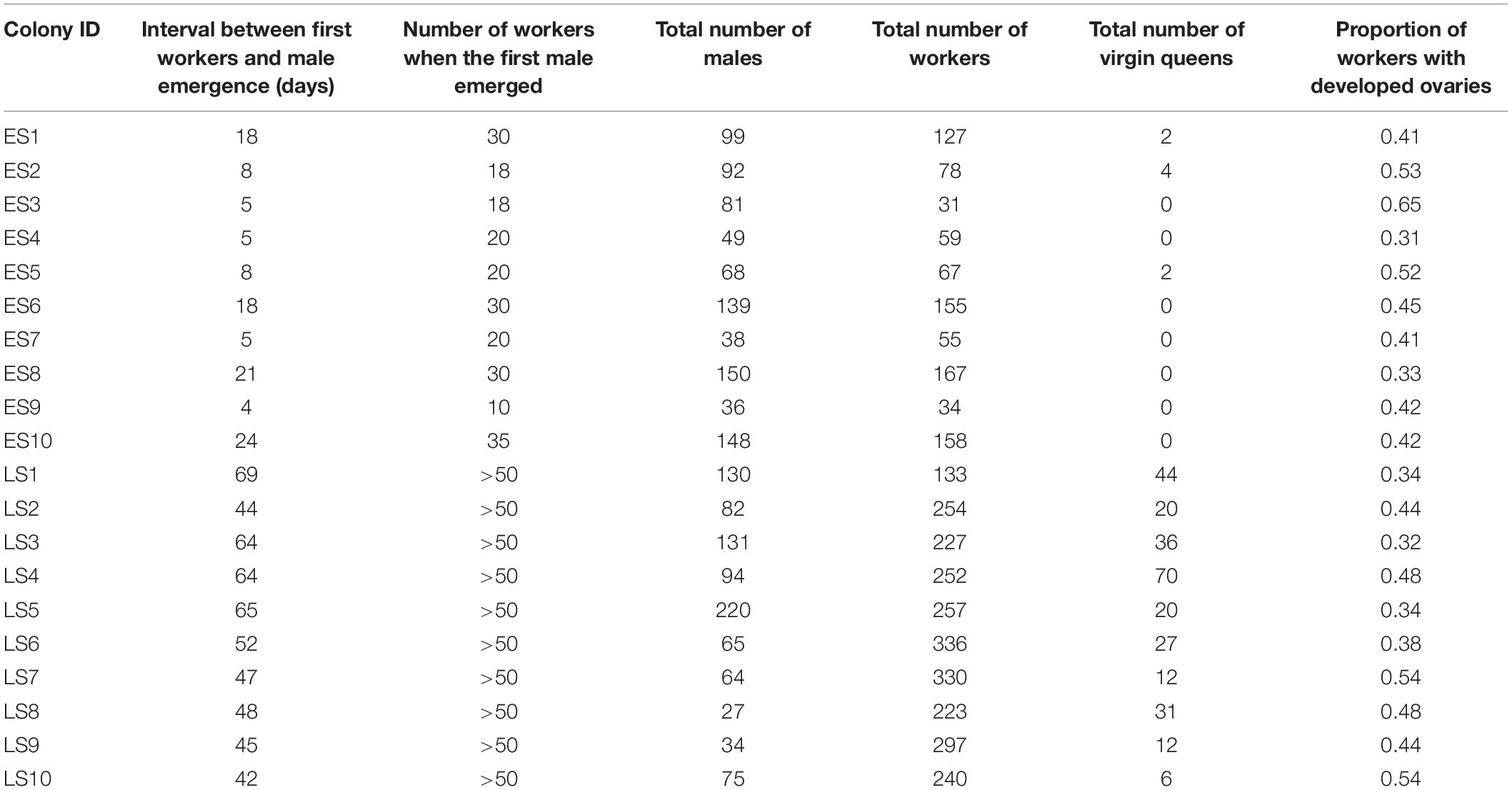
Table 1. Detailed data on the twenty early switching (ES) and late switching (LS) colonies including the number of days between worker and male emergence, the total number of workers present in the colony when males were first observed and at the end of the experiment, the total number of virgin queens, as well as the proportion of workers with developed ovaries.
All males successfully genotyped in our study were queens’ sons (complete genotype table presented in Supplementary Table 1). From the total of 324 sampled individuals, 21 failed to produce detectable microsatellite PCR products and were excluded from the analysis. The observed overall heterozygosity (HO) was 0.85, slightly higher than the expected heterozygosity (HS = 0.77; Supplementary Table 2). The overall fixation index (FST) was 0.04 suggesting no genetic differentiation between the bumblebee populations. Furthermore, the low and slightly negative inbreeding coefficient (FIS = −0.11) was consistent with random mating (Keller and Waller, 2002). A remarkable outcome of our study is that all genotyped males in early switching colonies were in fact diploid while all males analyzed in late switching colonies were haploid. According to the single locus CSD sex determination system, a matched mating should result in 50% of the diploid brood developing into males. In the diploid male producing early switching colonies, however, males began emerging only after the first cohort of workers. When the first males started to appear, colonies were highly biased toward producing workers, and there was a highly significant deviation from the expected 1:1 male:female ratio (binomial GLMM, mean prop. of workers produced = 0.959, LCL = 0.921, UCL = 0.979, significance of deviation from 1:1 ratio: p < 0.001; Figure 1B). Toward the end of the experimental period, hence when the colonies where frozen, however, colonies were producing diploid males and workers at a 1:1 ratio expected if a single-mated queen makes a matched mating (binomial GLMM, mean prop. of workers produced = 0.498, LCL = 0.423, UCL = 0.573, significance of deviation from 1:1 ratio: p = 0.962; Figure 1B and Supplementary Table 3), with only one exception in which one colony was producing almost three times more males than workers (Table 1). Conversely, late switching colonies produced almost exclusively workers throughout the experimental period, with haploid males emerging together with virgin queens (ca 25% males and 7% of virgin queens) at the end of the bioassay. In order to confirm that our results were not caused by inbreeding, we further selected seven early switching colonies from an experimental hybrid population created by crossing two genetically distinct outbred populations. The proportion of early switching colonies in the hybrid population is around 1%, according to the data provided by the breeding company. Consistent with the early switching colony of the main production, the males started to emerge on average 47 days after the colony initiation. Likewise, these colonies were also producing the diploid brood in a 1:1 ratio at the end of the experiment (binomial GLMM, mean prop. of workers produced = 0.459, LCL = 0.368, UCL = 0.553, significance of deviation from 1:1 ratio: p = 0.3936; Supplementary Table 3).
When comparing the overall chemical profiles of mother queens, workers, and males in both colony phenotypes, only males presented significant differences between early switching and late switching colonies (PERMANOVA F = 12.855, R2 = 0.061, p = 0.015), consistent with their difference in ploidy resulting in distinct cuticular profiles. A PCA shows that the first two dimensions explain more than 70% of the cuticular chemical variance, with the majority of the observed variance (Dim1 45.9%) being strongly linked with sex, caste membership and colony phenotype (Figure 2). Diploid and haploid males from early switching and late switching colonies differ in several CHCs belonging to all classes of compounds present on their cuticle, i.e., linear and branched alkanes, alkenes, alkadienes, and esters (Figure 3). Furthermore, the discriminant analysis was able to successfully distinguish the diploid male chemical profile from the haploid male chemical profile, with an accuracy of 98%. Interestingly, despite the overall CHC profile of workers and queens not being significantly different between early and late switching colonies (PERMANOVA ES workers × LS workers, F = 3.23, R2 = 0.035, p = 0.465; ES queens x LS queens, F = 4.212, R2 = 0.208, p = 0.285), some specific compounds were expressed significantly differently between the two colony phenotypes. Workers in late switching colonies presented two upregulated esters (tetracosyl oleates) when compared to workers in early switching colonies. A similar pattern was observed in queens, with late switching colonies upregulating two other esters (tetracosyl stearates) that are chemically similar to those observed in workers, but lacking the double bonds (Figure 3).
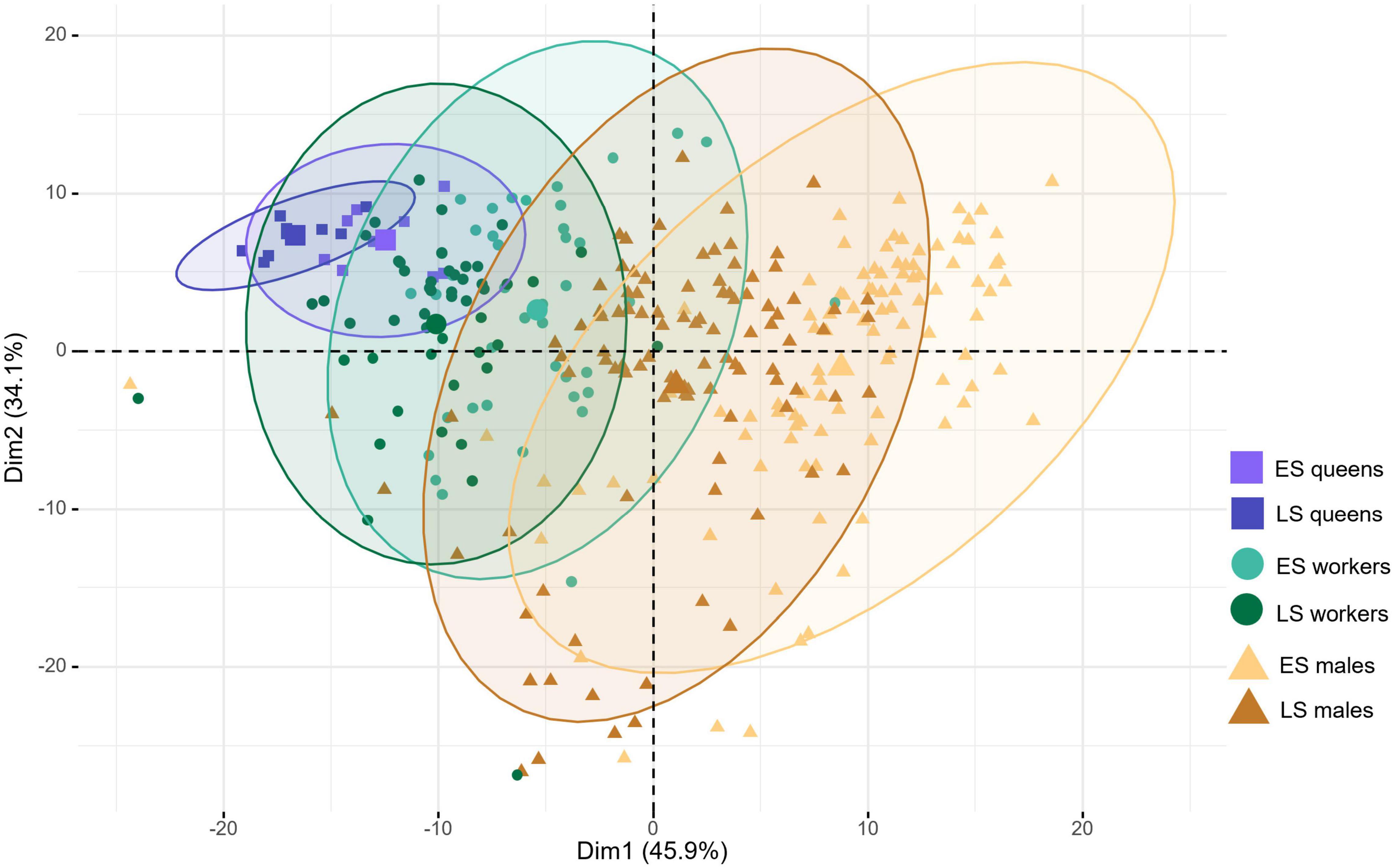
Figure 2. Principal component analysis of CHCs extracted from queens, males and workers of early (ES) and late switching (LS) colonies. Sex and caste membership are represented by the different symbols (queens = squares, workers = circles and males = triangles) with the centroid of the overall profile groups shown as the large symbol in the center of each 95% confidence interval ellipse. Different shades of each color show the two colony phenotypes for each caste or sex.

Figure 3. Heatmap with differences in the expression of individual CHCs in males, workers and queens between the two colony phenotypes. The results are presented in terms of the log2 fold change between the relative abundance of a given compound between late switchers and early switchers. Compounds with colors in the shades of green (between 0 and –3) are more abundant in early switching colonies while compounds with colors in the shade of brown (between 0 and 3) are overrepresented in late switching colonies. Stars indicate significance levels after Bonferroni correction for multiple comparisons: *p < 0.05; **p < 0.01; and ***p < 0.001. A complete table with chemical compounds, 95% CI intervals and significance levels is shown in Supplementary Table 4.
The buff-tailed bumblebee B. terrestris is a remarkable model species to study colony-level reproductive strategies. A particularly interesting trait of this species is the existence of two distinct colony phenotypes with highly divergent reproductive patterns. Some of the colonies are characterized by the production of males early in the season and others by the late emergence of both virgin queens and males (Duchateau and Velthuis, 1988). Several hypotheses have been proposed in the past to explain the evolutionary mechanisms underlying these different colony phenotypes. Yet, to date, this phenomenon remains poorly understood. Here, we investigated if a diploid male production could be behind this bimodal pattern.
A striking outcome of our study is that all genotyped males belonging to the early switching type colonies from both the main stock and the experimental hybrid colonies obtained by mating males and queens from different populations of the commercial rearing facility were in fact diploid, thereby showing that early production of males was in fact caused by queens having made a matched mating with a male that by chance carried the same sex allele. The fact that males of early switching hybrid colonies were diploid as well indicates that this was not a consequence of strong inbreeding within the main stock of the commercial bumblebee rearing but a mating between individuals carrying by chance the same allele at the CSD locus. Although previous studies showed that colonies producing diploid males can have a normal development and reach a large colony size (Duchateau et al., 1994; Bortolotti et al., 2020), diploid males exact a large fitness cost, as in social Hymenoptera their production comes at a cost of the production of new workers or queens, and on top of that diploid males are usually sterile, bringing no fitness benefits to the colony (Cook and Crozier, 1995), or, when fertile, imposing a hefty fitness cost to a potential female mate which will then produce triploid sterile progeny (Ayabe et al., 2004; Darvill et al., 2012; Harpur et al., 2013). It is therefore in the colony’s interest to adopt strategies to reduce or avoid diploid male production. In the honeybee A. mellifera for example, workers can detect a quantitative shift in the pattern of tricosane, pentacosane, heptacosane, nonacosane, and the terpenoid squalene present on the cuticle of diploid males larvae (Santomauro et al., 2004), promptly triggering their cannibalization (Woyke, 1963). In two stingless bee species, Melipona quadrifasciata and Scaptotrigona depilis, the brood is reared in mass provisioned sealed cells and, consequently, diploid males cannot be recognized during their larval stage. Nevertheless, workers are able to detect the presence of adult diploid males in the colony, presumably by making use of chemical cues (Borges et al., 2012; Vollet-Neto et al., 2017), eliminating the matched mated queen and replacing her with one of her daughters (Ratnieks, 1990). According to the CSD system, when a matched mating occurs, 50% of the diploid eggs laid by the queen should result in homozygous males (Cook and Crozier, 1995; Heimpel and De Boer, 2008; Whidden and Owen, 2011). An interesting outcome of our study was that we only observed the expected 1:1 worker to diploid male ratio at the end of the experimental period, and that the first batch of brood still almost entirely consisted of workers, even though a priori one would expect workers and diploid males to be produced in a 1:1 ratio throughout the entire colony cycle (Whiting, 1943). This suggests that mother queens could be able to selectively remove diploid male brood in an early developmental stage during the early phases of the colony cycle. Mechanistically, this could be achieved by the detection of specific chemical cues emitted by either diploid male eggs or first instar larvae. Indeed, it has been shown that in B. terrestris, eggs have caste and colony-specific chemical signals which are largely composed of alkanes, alkenes and alkadienes (Ayasse et al., 1999). The precise mechanism preventing diploid male brood from developing into adults at the beginning of the colony cycle remains nevertheless an interesting topic for future studies.
Some morphological traits can be used as diagnostics to differentiate diploid from haploid males of B. terrestris such as body size (Duchateau and Mariën, 1995; Gerloff et al., 2003) and the wing venation pattern (Gerard et al., 2015; Bortolotti et al., 2021; Bogo et al., 2022). Here, we show that their cuticular hydrocarbon profiles can also be used to reliably distinguish adult diploid and haploid B. terrestris males with an accuracy of 98%. The CHC reveals that diploid (early switching colonies) and haploid (late switching colonies) males present no qualitative differences on their CHC profile, but shows significant quantitative differences, with some compounds being more expressed in diploid males while others being overrepresented in haploid individuals. Therefore, these specific differences in the chemical profile of diploid males could potentially be used by workers as cues to start a premature competition over the male parentage in the colony with the queen. At the end of our experiment, nearly 50% of the workers had developed ovaries in both colony types. Previous studies have shown that, despite the high levels of ovary activation in workers, only around 4.5% of the total males produced are worker sons (Alaux et al., 2004b; Lopez-Vaamonde et al., 2004a), which implies that the queen wins the competition over the male parentage (Bourke and Ratnieks, 2001). This could be an indication that workers have a limited potential to mitigate the fitness loss due to diploid male production. Nevertheless, worker matricide toward the end of the season is not uncommon in B. terrestris and allows workers to reproduce without interference by the mother queen (Van Honk et al., 1981; Van Doorn and Heringa, 1986; Bourke, 1994; Wenseleers et al., 2021), which could constitute an advantage for early switching colonies to produce workers’ haploid males instead of the queen’s diploid ones. Nevertheless, the difference in the frequency of matricide between early and late switching colonies is currently unknown (Goulson, 2010).
In conclusion, in this study we show that in our study population, early switching colonies of B. terrestris, might be resulting from queens mated with males that by chance carry the same sex allele. In our study the frequency of early switching colonies was relatively low, being observed in only 6% of the colonies, which is in accordance with a previous study (Lopez-Vaamonde et al., 2003). However, there is an overwhelming bias toward laboratory studies with respect to the proportions of the two colony types, which might not fully reflect the situation in natural conditions. While the frequency of early and late switching colonies was rarely assessed in the field, some studies show that the proportion of early switching colonies was much higher than what is observed in laboratory conditions (50% in Duchateau and Velthuis, 1988; 14% in Yeninar and Kaftanoglu, 1996). Unfortunately, the ploidy of the first male cohort emerging in early switching colonies was not determined in any of these occasions. In addition, the diploid brood was assumed to be females in experiments determining the ploidy of eggs (Lopez-Vaamonde et al., 2003, 2004b). Our results alone cannot preclude the existence of different populations with early switching colonies producing haploid males early in the season. Therefore, testing the generality of our findings in other B. terrestris populations is an interesting topic for future investigations.
The presence of diploid males is linked with the number of sex alleles in the population, which was estimated to be 24 for B. terrestris (Duchateau et al., 1994). However, this number could be an overestimate of the alleles actually available in wild that can vary from population to population. Furthermore, habitat fragmentation is another factor that increases population susceptibility to the loss of genetic diversity (Frankham et al., 2002; Keller and Waller, 2002). Many bumblebees population have suffered a sharp decline in recent decades, in which it is already possible to observe diploid males in wild populations at detectable frequencies (B. muscorum, Darvill et al., 2006; B. sylvarum, Ellis et al., 2006; B. florilegus, Takahashi et al., 2008; B. occidentalis, B. perplexus, Whidden and Owen, 2011). Therefore, the detection of diploid males in wild is an essential parameter in hymenopteran conservation genetics since high frequencies of homozygotes may signal genetic depletion resulting from matched mating, small habitat size, habitat fragmentation, loss of genetic diversity, or a combination of these factors (Packer and Owen, 2001).
Finally, our results suggest the existence of a mechanism by which a matched mated queen can prevent the emergence of diploid males during the first batch of brood but is no longer able to do so later on during the colony cycle. In addition, our data suggest that the early onset of the competition point (Bloch, 1999), in which workers engage in direct competition over the male parentage with the queen, might be a consequence of the premature appearance of males in the colony. Together, our findings demonstrate that, similarly to the other two groups of highly eusocial corbiculate bees (i.e., honeybees and stingless bees), the buff-tailed bumblebee B. terrestris might display a behavioral adaptation to prevent population decline due to diploid male production.
The datasets and R scripts generated during the current study are available in the Mendeley Data repository, doi: https://doi.org/10.17632/FDM8924TKP.3 (Di Pietro et al., 2021).
RCO and TW conceived the original idea. VDP, AVO, FW, TW, and RCO designed the study. AVO and FW selected and provided the colonies. VDP performed the experiments and performed the dissections. VDP and HMF performed the chemical analysis. VDP, RCO, HMF, and TW performed data analysis and interpretation. VDP, HMF, and RCO prepared the first draft. All authors contributed to revision, proofreading, and the approved version before submission.
This study was supported by the Research Foundation Flanders (FWO) with the research grant G064120N, FWO postdoctoral grant postdoctoral grant 12R9619N and research grant 1502119N to RCO.
AVO and FW were employed by company Biobest Group NV.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Bart Hellemans for his assistance with the microsatellite genotyping and Cátia José Neves for her helpful insights in the microsatellite analysis.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.844251/full#supplementary-material
Aitchison, J. (1982). The statistical analysis of compositional data. J. R. Stat. Soc. Ser. B Methodol. 44, 139–160. doi: 10.1111/j.2517-6161.1982.tb01195.x
Alaux, C., Savarit, F., Jaisson, P., and Hefetz, A. (2004b). Does the queen win it all? Queen-worker conflict over male production in the bumblebee, Bombus terrestris. Naturwissenschaften 91, 400–403. doi: 10.1007/s00114-004-0547-3
Alaux, C., Jaisson, P., and Hefetz, A. (2004a). Queen influence on worker reproduction in bumblebees (Bombus terrestris) colonies. Insectes Soc. 51, 287–293. doi: 10.1007/s00040-004-0741-5
Alaux, C., Jaisson, P., and Hefetz, A. (2005). Reproductive decision-making in semelparous colonies of the bumblebee Bombus terrestris. Behav. Ecol. Sociobiol. 59, 270–277. doi: 10.1007/s00265-005-0035-6
Alves, D. A., Imperatriz-Fonseca, V. L., Francoy, T. M., Santos-Filho, P. S., Billen, J., and Wenseleers, T. (2011). Successful maintenance of a stingless bee population despite a severe genetic bottleneck. Conserv. Genet. 12, 647–658. doi: 10.1007/s10592-010-0171-z
Amsalem, E., Twele, R., Francke, W., and Hefetz, A. (2009). Reproductive competition in the bumble-bee Bombus terrestris: do workers advertise sterility? Proc. R. Soc. B Biol. Sci. 276, 1295–1304. doi: 10.1098/rspb.2008.1688
Ayabe, T., Hoshiba, H., and Ono, M. (2004). Cytological evidence for triploid males and females in the bumblebee, Bombus terrestris. Chromosome Res. 12, 215–223. doi: 10.1023/B:CHRO.0000021880.83639.4b
Ayasse, M., Birnbaum, J., Tengö, J., van Doorn, A., Taghizadeh, T., and Francke, W. (1999). Caste-and colony-specific chemical signals on eggs of the bumble bee, Bombus terrestris L. (Hymenoptera: Apidae). Chemoecology 9, 119–126. doi: 10.1007/s000490050042
Baer, B., and Schmid-Hempel, P. (2003). Effects of selective episodes in the field on life history traits in the bumblebee Bombus terrestris. Oikos 101, 563–568. doi: 10.1034/j.1600-0706.2003.12395.x
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Beekman, M., and Van Stratum, P. (1998). Bumblebee sex ratios: why do bumblebees produce so many males? Proc. R. Soc. B Biol. Sci. 265, 1535–1543. doi: 10.1098/rspb.1998.0469
Beekman, M., Van Stratum, P., and Veerman, A. (1999). Selection for non-diapause in the bumblebee Bombus terrestris, with notes on the effect of inbreeding. Entomol. Exp. Appl. 93, 69–75. doi: 10.1046/j.1570-7458.1999.00563.x
Bloch, G. (1999). Regulation of queen–worker conflict in bumble bee (Bombus terrestris) colonies. Proc. R. Soc. Lond. Ser. B Biol. Sci. 266, 2465–2469. doi: 10.1098/rspb.1999.0947
Bloch, G., and Hefetz, A. (1999). Regulation of reproduction by dominant workers in bumblebee (Bombus terrestris) queenright colonies. Behav. Ecol. Sociobiol. 45, 125–135. doi: 10.1007/s002650050546
Bogo, G., Cejas, D., De la Rúa, P., and Bortolotti, L. (2022). How to get rid of diploid bumblebee males–variability in wing size and shape does not allow within-colony ploidy discrimination. Entomol. Exp. Appl. 170, 182–192. doi: 10.1111/eea.13128
Bogo, G., De Manincor, N., Fisogni, A., Galloni, M., Zavatta, L., and Bortolotti, L. (2018). No evidence for an inbreeding avoidance system in the bumble bee Bombus terrestris. Apidologie 49, 473–483. doi: 10.1007/s13592-018-0575-1
Borges, A. A., Ferreira-Caliman, M. J., Nascimento, F. S., Campos, L. A. O., and Tavares, M. G. (2012). Characterization of cuticular hydrocarbons of diploid and haploid males, workers and queens of the stingless bee Melipona quadrifasciata. Insectes Soc. 59, 479–486. doi: 10.1007/s00040-012-0242-x
Bortolotti, L., Fiorillo, F., Dall’Olio, R., Cejas, D., De la Rúa, P., and Bogo, G. (2021). Ploidy determination in Bombus terrestris males: cost-efficiency comparison among different techniques. J. Apic. Res. 61, 180–189. doi: 10.1080/00218839.2021.1959753
Bortolotti, L., Fiorillo, F., Zavatta, L., De Rogatis, A., and Bogo, G. (2020). Influence of inbreeding in the early stages of artificially reared colonies of Bombus terrestris. J. Appl. Entomol. 144, 637–646. doi: 10.1111/jen.12765
Bourke, A. F. G. (1994). Worker matricide in social bees and wasps. J. Theor. Biol. 167, 283–292. doi: 10.1006/jtbi.1994.1070
Bourke, A. F. G. (1997). Sex ratios in bumble bees. Philos. Trans. R. Soc. B Biol. Sci. 352, 1921–1933. doi: 10.1098/rstb.1997.0179
Bourke, A. F. G., and Franks, N. R. (1995). Social Evolution in Ants. Princeton, NJ: Princeton University Press. doi: 10.1515/9780691206899
Bourke, A. F. G., and Ratnieks, F. L. W. (2001). Kin-selected conflict in the bumble-bee Bombus terrestris (Hymenoptera: Apidae). Proc. R. Soc. B Biol. Sci. 268, 347–355. doi: 10.1098/rspb.2000.1381
Brown, M. J., and Baer, B. (2005). The evolutionary significance of long copulation duration in bumble bees. Apidologie 36, 157–167. doi: 10.1051/apido:2005008
Bulmer, M. G. (1981). Worker-queen conflict in annual social Hymenoptera. J. Theor. Biol. 93, 239–251. doi: 10.1016/0022-5193(81)90068-0
Bulmer, M. G. (1983). The significance of protandry in social Hymenoptera. Am. Nat. 121, 540–551. doi: 10.1086/284081
Chapman, R. E., and Bourke, A. F. G. (2001). The influence of sociality on the conservation biology of social insects. Ecol. Lett. 4, 650–662. doi: 10.1046/j.1461-0248.2001.00253.x
Cook, J. M., and Crozier, R. H. (1995). Sex determination and population biology in the Hymenoptera. Trends Ecol. Evol. 10, 281–286. doi: 10.1016/0169-5347(95)90011-X
Darvill, B., Ellis, J. S., Lye, G. C., and Goulson, D. (2006). Population structure and inbreeding in a rare and declining bumblebee, Bombus muscorum (Hymenoptera: Apidae). Mol. Ecol. 15, 601–611. doi: 10.1111/j.1365-294X.2006.02797.x
Darvill, B., Lepais, O., Woodall, L. C., and Goulson, D. (2012). Triploid bumblebees indicate a direct cost of inbreeding in fragmented populations. Mol. Ecol. 21, 3988–3995. doi: 10.1111/j.1365-294X.2012.05679.x
Di Pietro, V., Ferreira, H. M. Van Oystaeyen, A. Wäckers, F., Wenseleers, T., and Oliveira, R. C. (2021). Distinct Colony Types Caused by Diploid Male Production in the Buff-Tailed Bumblebee Bombus terrestris. V3 [Dataset]. doi: 10.17632/FDM8924TKP.3
Duchateau, M. J., Hoshiba, H., and Velthuis, H. H. W. (1994). Diploid males in the bumble bee Bombus terrestris. Sex determination, sex alleles and viability. Entomol. Exp. Appl. 71, 263–269. doi: 10.1111/j.1570-7458.1994.tb01793.x
Duchateau, M. J., and Mariën, J. (1995). Sexual biology of haploid and diploid males in the bumble bee Bombus terrestris. Insectes Soc. 42, 255–266. doi: 10.1007/BF01240420
Duchateau, M. J., and Velthuis, H. H. W. (1988). Development and reproductive strategies in Bombus terrestris colonies. Behaviour 107, 186–207. doi: 10.1163/156853988X00340
Duchateau, M. J., and Velthuis, H. H. W. (1989). Ovarian development and egg laying in workers of Bombus terrestris. Entomol. Exp. Appl. 51, 199–213. doi: 10.1111/j.1570-7458.1989.tb01231.x
Duchateau, M. J., Velthuis, H. H. W., and Boomsma, J. J. (2004). Sex ratio variation in the bumblebee Bombus terrestris. Behav. Ecol. 15, 71–82. doi: 10.1093/beheco/arg087
Ellis, J. S., Knight, M. E., Darvill, B., and Goulson, D. (2006). Extremely low effective population sizes, genetic structuring and reduced genetic diversity in a threatened bumblebee species, Bombus sylvarum (Hymenoptera: Apidae). Mol. Ecol. 15, 4375–4386. doi: 10.1111/j.1365-294X.2006.03121.x
Engels, W., and Imperatriz-Fonseca, V. L. (1990). “Caste development, reproductive strategies, and control of fertility in honey bees and stingless bees,” in Social Insects, ed. W. Engels (Berlin: Springer), 167–230. doi: 10.1007/978-3-642-74490-7_9
Estoup, A., Scholl, A., Pouvreau, A., and Solignac, M. (1995). Monoandry and polyandry in bumble bees (Hymenoptera; Bombinae) as evidenced by highly variable microsatellites. Mol. Ecol. 4, 89–94. doi: 10.1111/j.1365-294x.1995.tb00195.x
Foster, R. L. (1992). Nestmate recognition as an inbreeding avoidance mechanism in bumble bees (Hymenoptera: Apidae). J. Kansas Entomol. Soc. 65, 238–243.
Frankham, R., Ballou, S. E. J. D., Briscoe, D. A., and Ballou, J. D. (2002). Introduction to Conservation Genetics. Cambridge: Cambridge university press.
Gerard, M., Michez, D., Fournier, D., Maebe, K., Smagghe, G., Biesmeijer, J. C., et al. (2015). Discrimination of haploid and diploid males of Bombus terrestris (Hymenoptera; Apidae) based on wing shape. Apidologie 46, 644–653. doi: 10.1007/s13592-015-0352-3
Gerloff, C. U., Ottmer, B. K., and Schmid-Hempel, P. (2003). Effects of inbreeding on immune response and body size in a social insect, Bombus terrestris. Funct. Ecol. 17, 582–589. doi: 10.1046/j.1365-2435.2003.00769.x
Gerloff, C. U., and Schmid-Hempel, P. (2005). Inbreeding depression and family variation in a social insect, Bombus terrestris (Hymenoptera: Apidae). Oikos 111, 67–80. doi: 10.1111/j.0030-1299.2005.13980.x
Gienapp, P., Teplitsky, C., Alho, J. S., Mills, J. A., and Merilä, J. (2008). Climate change and evolution: disentangling environmental and genetic responses. Mol. Ecol. 17, 167–178. doi: 10.1111/j.1365-294X.2007.03413.x
Gosterit, A. (2016). Adverse effects of inbreeding on colony foundation success in bumblebees, Bombus terrestris (Hymenoptera: Apidae). Appl. Entomol. Zool. 51, 521–526. doi: 10.1007/s13355-016-0427-2
Gosterit, A., Koskan, O., and Gurel, F. (2016). The relationship of weight and ovarian development in Bombus terrestris workers under different social conditions. J. Apic. Sci. 60, 51–58. doi: 10.1515/jas-2016-0016
Goudet, J. (2005). Hierfstat, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 5, 184–186. doi: 10.1111/j.1471-8286.2004.00828.x
Goulson, D. (2010). Bumblebees: Behaviour, Ecology and Conservation. New York, NY: Oxford University Press. doi: 10.1086/655083
Hamilton, W. D. (1964). The genetical evolution of social behaviour. II. J. Theor. Biol. 7, 17–52. doi: 10.1016/0022-5193(64)90039-6
Harpur, B. A., Sobhani, M., and Zayed, A. (2013). A review of the consequences of complementary sex determination and diploid male production on mating failures in the Hymenoptera. Entomol. Exp. Appl. 146, 156–164. doi: 10.1111/j.1570-7458.2012.01306.x
Heimpel, G. E., and De Boer, J. G. (2008). Sex determination in the Hymenoptera. Annu. Rev. Entomol. 53, 209–230. doi: 10.1146/annurev.ento.53.103106.093441
Helms, K. R. (1999). Colony sex ratios, conflict between queens and workers, and apparent queen control in the ant Pheidole desertorum. Evolution 53, 1470–1478. doi: 10.1111/j.1558-5646.1999.tb05411.x
Hoffmann, A. A., and Willi, Y. (2008). Detecting genetic responses to environmental change. Nat. Rev. Genet. 9, 421–432. doi: 10.1038/nrg2339
Hothorn, T., Bretz, F., and Westfall, P. (2008). Simultaneous inference in general parametric models. Biom. J. 50, 346–363. doi: 10.1002/bimj.200810425
Kassambara, A., and Mundt, F. (2017). ”factoextra: Extract and Visualize the Results of Multivariate Data Analyses”, in: CRAN repository.
Keller, L. F., and Waller, D. M. (2002). Inbreeding effects in wild populations. Trends Ecol. Evol. 17, 230–241. doi: 10.1016/S0169-5347(02)02489-8
Kümmerli, R., and Keller, L. (2009). Patterns of split sex ratio in ants have multiple evolutionary causes based on different within-colony conflicts. Biol. Lett. 5, 713–716. doi: 10.1098/rsbl.2009.0295
Lê, S., Josse, J., and Husson, F. (2008). FactoMineR: an R Package for multivariate analysis. J. Stat. Softw. 25, 1–18. doi: 10.18637/jss.v025.i01
Lenth, R. V. (2016). Least-squares means: the R package lsmeans. J. Stat. Softw. 69, 1–33. doi: 10.18637/jss.v069.i01
Lopez-Vaamonde, C., Koning, J. W., Brown, R. M., Jordan, W. C., and Bourke, A. F. G. (2004a). Social parasitism by male-producing reproductive workers in a eusocial insect. Nature 430, 557–560. doi: 10.1038/nature02769
Lopez-Vaamonde, C., Koning, J. W., Jordan, W. C., and Bourke, A. F. G. (2004b). A test of information use by reproductive bumblebee workers. Anim. Behav. 68, 811–818. doi: 10.1016/j.anbehav.2004.01.007
Lopez-Vaamonde, C., Koning, J. W., Jordan, W. C., and Bourke, A. F. G. (2003). No evidence that reproductive bumblebee workers reduce the production of new queens. Anim. Behav. 66, 577–584. doi: 10.1006/anbe.2003.2205
Lopez-Vaamonde, C., Raine, N. E., Koning, J. W., Brown, R. M., Pereboom, J. J., Ings, T. C., et al. (2009). Lifetime reproductive success and longevity of queens in an annual social insect. J. Evol. Biol. 22, 983–996. doi: 10.1111/j.1420-9101.2009.01706.x
Martinez Arbizu, P. (2020). ”pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis”, in: CRAN Repository.
Meunier, J., West, S. A., and Chapuisat, M. (2008). Split sex ratios in the social Hymenoptera: a meta-analysis. Behav. Ecol. 19, 382–390. doi: 10.1093/beheco/arm143
Nei, M. (1987). Molecular Evolutionary Genetics. New York, NY: Columbia university press. doi: 10.7312/nei-92038
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’hara, R. B., et al. (2013). ”vegan: Community Ecology Package”, in: CRAN repository.
Packer, L., and Owen, R. (2001). Population genetic aspects of pollinator decline. Conserv. Ecol. 5:4. doi: 10.5751/ES-00267-050104
Page, R. E. Jr. (1980). The evolution of multiple mating behavior by honey bee queens (Apis mellifera L.). Genetics 96, 263–273. doi: 10.1093/genetics/96.1.263
Page, R. E. Jr., and Metcalf, R. A. (1982). Multiple mating, sperm utilization, and social evolution. Am. Nat. 119, 263–281. doi: 10.1086/283907
Pamilo, P. (1982). Genetic evolution of sex ratios in eusocial Hymenoptera: allele frequency simulations. Am. Nat. 119, 638–656. doi: 10.1086/283940
Perez, R., and Aron, S. (2020). Adaptations to thermal stress in social insects: recent advances and future directions. Biol. Rev. 95, 1535–1553. doi: 10.1111/brv.12628
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Ratnieks, F. L. W. (1990). The evolution of polyandry by queens in social Hymenoptera: the significance of the timing of removal of diploid males. Behav. Ecol. Sociobiol. 26, 343–348. doi: 10.1007/BF00171100
Reber Funk, C., Schmid-Hempel, R., and Schmid-Hempel, P. (2006). Microsatellite loci for Bombus spp. Mol. Ecol. Notes 6, 83–86. doi: 10.1111/j.1471-8286.2005.01147.x
Santomauro, G., Oldham, N. J., Boland, W., and Engels, W. (2004). Cannibalism of diploid drone larvae in the honey bee (Apis mellifera) is released by odd pattern of cuticular substances. J. Apic. Res. 43, 69–74. doi: 10.1080/00218839.2004.11101114
Schmid-Hempel, P., Schmid-Hempel, R., Brunner, P. C., Seeman, O. D., and Allen, G. R. (2007). Invasion success of the bumblebee, Bombus terrestris, despite a drastic genetic bottleneck. Heredity 99, 414–422. doi: 10.1038/sj.hdy.6801017
Schmid-Hempel, R., and Schmid-Hempel, P. (2000). Female mating frequencies in Bombus spp. from Central Europe. Insectes Soc. 47, 36–41. doi: 10.1007/s000400050006
Schmid-Hempel, R., and Schmid-Hempel, P. (1998). Colony performance and immunocompetence of a social insect, Bombus terrestris, in poor and variable environments. Funct. Ecol. 12, 22–30. doi: 10.1046/j.1365-2435.1998.00153.x
Stolle, E., Rohde, M., Vautrin, D., Solignac, M., Schmid-Hempel, P., Schmid-Hempel, R., et al. (2009). Novel microsatellite DNA loci for Bombus terrestris (Linnaeus, 1758). Mol. Ecol. Resour. 9, 1345–1352. doi: 10.1111/j.1755-0998.2009.02610.x
Takahashi, J., Ayabe, T., Mitsuhata, M., Shimizu, I., and Ono, M. (2008). Diploid male production in a rare and locally distributed bumblebee, Bombus florilegus (Hymenoptera, Apidae). Insectes Soc. 55, 43–50. doi: 10.1007/s00040-007-0976-z
Templeton, A. R., Shaw, K., Routman, E., and Davis, S. K. (1990). The genetic consequences of habitat fragmentation. Ann. Mo. Bot. Garden 1, 13–27. doi: 10.2307/2399621
Trivers, R. L., and Hare, H. (1976). Haploidploidy and the evolution of the social insect. Science 191, 249–263. doi: 10.1126/science.1108197
Van der Blom, J. (1986). Reproductive dominance within colonies of Bombus terrestris (L.). Behaviour 97, 37–49. doi: 10.1163/156853986x00306
Van Doorn, A., and Heringa, J. (1986). The ontogeny of a dominance hierarchy in colonies of the bumblebee Bombus terrestris (Hymenoptera, Apidae). Insectes Soc. 33, 3–25. doi: 10.1007/BF02224031
Van Honk, C. G. J., Röseler, P. F., Velthuis, H. H. W., and Hoogeveen, J. C. (1981). Factors influencing the egg laying of workers in a captive Bombus terrestris colony. Behav. Ecol. Sociobiol. 9, 9–14. doi: 10.2307/4599404
Van Wilgenburg, E., Driessen, G., and Beukeboom, L. W. (2006). Single locus complementary sex determination in Hymenoptera: an” unintelligent” design? Front. Zool. 3:1. doi: 10.1186/1742-9994-3-1
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics with S. Berlin: Springer. doi: 10.1007/b97626
Vollet-Neto, A., Oliveira, R. C., Schillewaert, S., Alves, D. A., Wenseleers, T., Nascimento, F. S., et al. (2017). Diploid male production results in queen death in the stingless bee Scaptotrigona depilis. J. Chem. Ecol. 43, 403–410. doi: 10.1007/s10886-017-0839-7
Walsh, P. S., Metzger, D. A., and Higuchi, R. (2018). Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 54, 134–139. doi: 10.2144/000114018
Weir, B. S., and Cockerham, C. C. (1984). Estimating F-statistics for the analysis of population structure. Evolution 38, 1358–1370. doi: 10.2307/2408641
Wenseleers, T., Princen, S., Caliari Oliveira, R., and Oi, C. A. (2021). “Conflicts of interest within colonies,” in Encyclopedia of Social Insects, ed. C. K. Starr (Cham: Springer), 1–15. doi: 10.1007/978-3-319-90306-4_28-1
Whidden, T. L., and Owen, R. E. (2011). Frequencies of diploid males in natural populations of three North American bumble bee (Bombus) species (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 104, 83–87. doi: 10.1603/AN10092
Whitehorn, P. R., Tinsley, M. C., Brown, M. J. F., Darvill, B., and Goulson, D. (2009). Impacts of inbreeding on bumblebee colony fitness under field conditions. BMC Evol. Biol. 9:152. doi: 10.1186/1471-2148-9-152
Whiting, P. W. (1943). Multiple alleles in complementary sex determination of Habrobracon. Genetics 28, 365–382. doi: 10.1093/genetics/28.5.365
Winston, M. L. (1996). Killer bees : the africanized honey bee in the Americas. Perspect. Biol. Med. 39:312. doi: 10.1353/pbm.1996.0049
Woyke, J. (1962). The hatchability of ‘lethal’eggs in a two sex-allele fraternity of honeybees. J. Apic. Res. 1, 6–13. doi: 10.1080/00218839.1962.11100040
Woyke, J. (1963). What happens to diploid drone larvae in a honeybee colony. J. Apic. Res. 2, 73–75. doi: 10.1080/00218839.1963.11100063
Yeninar, H., and Kaftanoglu, O. (1996). “Colony development of anatolian bumblebees (Bombus terrestris) under laboratory conditions,” in Paper Presented at the VII International Symposium on Pollination, Vol. 437, Lethbridge, AB, 277–282. doi: 10.17660/actahortic.1997.437.32
Keywords: Bombus terrestris, cuticular hydrocarbons, diploid males, complementary sex determination, sex ratio
Citation: Di Pietro V, Ferreira HM, Van Oystaeyen A, Wäckers F, Wenseleers T and Oliveira RC (2022) Distinct Colony Types Caused by Diploid Male Production in the Buff-Tailed Bumblebee Bombus terrestris. Front. Ecol. Evol. 10:844251. doi: 10.3389/fevo.2022.844251
Received: 27 December 2021; Accepted: 28 February 2022;
Published: 04 April 2022.
Edited by:
Isabel Marques, University of Lisbon, PortugalReviewed by:
Fehmi Gurel, Akdeniz University, TurkeyCopyright © 2022 Di Pietro, Ferreira, Van Oystaeyen, Wäckers, Wenseleers and Oliveira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Viviana Di Pietro, dml2aWFuYS5kaXBpZXRyb0BrdWxldXZlbi5iZQ==
†ORCID: Viviana Di Pietro, orcid.org/0000-0003-1116-9795; Helena Mendes Ferreira, orcid.org/0000-0003-2090-4528; Annette Van Oystaeyen, orcid.org/0000-0002-7756-8628; Felix Wäckers, orcid.org/0000-0002-9748-0615; Tom Wenseleers, orcid.org/0000-0002-1434-861X; Ricardo Caliari Oliveira, orcid.org/0000-0002-8996-1291
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.