
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 18 April 2022
Sec. Paleoecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.838839
This article is part of the Research Topic Formation and Taphonomy of Quaternary Fossil Accumulations: Advances and New Perspectives View all 5 articles
 Anna Clara Arboitte de Assumpção1
Anna Clara Arboitte de Assumpção1 Felipe Caron2†
Felipe Caron2† Fernando Erthal1,3†
Fernando Erthal1,3† Eduardo Guimarães Barboza1,4†
Eduardo Guimarães Barboza1,4† Raphael Mathias Pinotti5†
Raphael Mathias Pinotti5† Matias do Nascimento Ritter1,2*
Matias do Nascimento Ritter1,2*
The study of sambaquis (ancient shell mounds) in conservation paleobiology is a complicated subject, especially when comparing body sizes of current and past mollusk valves to observe possible changes. There is a lack of information regarding how ancient fisher-gardeners collected these shellfish. Another obstacle is finding the hypothesis tests and data that can be used to compare current and past body sizes. To this end, we use the t-test (tt), Mann-Whitney (mw), and Bootstrap (bt) analyses to determine data scenarios for yellow clam (Amarilladesma mactroides) shells from two sambaquis units, and from two transects along the beaches of Rio Grande do Sul in southern Brazil. The study finds that the average body size of an Amarilladesma mactroides adult specimen is greater in sambaquis than the current valves across all tests (tt, p = 0.0005; mw, p = 0.006; bt, p = 0.04), even after reducing the sample number. However, when the comparison is only between valves that are larger than average, this does not hold true for two of the statistics (tt, p = 0.24; mw, p = 0.04; bt, p = 0.25). By refining the comparison for recent valves from different levels in the swash zone (upper, middle, and lower levels) with the past valves, we find that body sizes only differ at the middle level of the swash zone (tt, p = 0.004; mw, p = 0.004; bt, p = 0.005), where smaller adults, recruits, and juveniles live. This result could indicate that fisher-gardeners collected large organisms at distinct levels depending on the season (i.e., adults are collected in the upper level during summer and in the lower level during winter).
Humans have increasingly been affecting the environment since their origin, causing direct and indirect damage to living beings around them (Kowalewski et al., 2000; Millenium Ecosystem Assessment [MEA], 2005; Ewers and Didham, 2006; Diaz and Rosenberg, 2008; Halpern et al., 2008; Barnosky et al., 2011; Conservation Paleobiology Workshop, 2012; Dirzo et al., 2014; Boivin et al., 2016). Conservation studies have established baselines for sustainable conditions prior to human impact, or where there is less environmental interference (Monsarrat et al., 2019). Therefore, it is possible to understand how the environments used to be, which species existed, and how they responded to variations; in this way, species management can be more effective and clearer (Willis and Birks, 2006; Dietl and Flessa, 2009, 2011; Kowalewski, 2017; Kusnerik et al., 2018; Dietl, 2019; Albano et al., 2020). The name given to this science is conservation paleobiology. It uses paleontology techniques and theories to unravel conservation issues, restoration of biodiversity, and ecosystem services. It aims to develop a long-term scale (Dietl and Flessa, 2009, 2011); and studies using archeological constructions are increasingly common, as they are the geo-archives of past species data (e.g., Mannino and Thomas, 2002; Cannarozzi and Kowalewski, 2019; Fossile et al., 2019; Mendes et al., 2019).
One such construction is the ancient shell mounds, locally named “sambaquis” (from “Tupi-Guarani,” tamba = shell, ki = huddle), which are distributed plentifully all along the Brazilian coast. These sambaquis were built by indigenous groups of fisher-gardeners between circa 10,000 and 1,000 years ago, and are composed predominantly of mollusk shells (Lima, 1999; Figuti, 2005; Schmitz, 2006; Wagner, 2009; Ferrasso et al., 2016; Silva et al., 2017; Scheel-Ybert and Boyadjian, 2020). Presently, the sambaquis are interpreted as funerary monuments and ceremonial spaces, which signifies that the mollusks were collected principally as building material and their role as food was secondary—the shells are great building material and food supplements at once (DeBlasis et al., 1998, 2007; Scheel-Ybert and Boyadjian, 2020). The use of these Brazilian shell mounds in conservation paleobiology, as a baseline for when human activities were less expressive, has already showed that it is possible to establish a taxonomic census of species that occurred in past environments (Fossile et al., 2019; Mendes et al., 2019), but no effort has been made yet to evaluate the body-size difference between the valves of present-day mollusks and of sambaquis for conservation purposes. These information from the past can help us conserve the environment today; in other words, the past is also the key to the present and the future (Dietl and Flessa, 2017). The fisher-gardeners may have acted as researchers of the past, although they lacked discernible scientific methods.
However, working with research concerning the comparison of the valves’ body size (an intrinsic characteristic of mollusks) from the sambaquis and of the current populations proved to be a convoluted task. Although the body size of the valves contains relevant information about what the environment was like in their lifetime (Fiori and Cazzaniga, 1999; Mannino and Thomas, 2002; Campbell, 2008; Álvarez-Fernández et al., 2011), the information acquired from the sambaquis may be biased, because they were made by humans. There is no real evidence from sambaquis in south Brazil about how the gathering was done by the fisher-gardeners, but factors such as cultural preferences, food taboos, discarded materials, and technique can affect the data that was collected (Silva et al., 2017; Mendes et al., 2019). For the result to be as reliable as possible, other important questions, such as the type of data and tests that are appropriate for this comparison, must be addressed.
For these reasons, this research aims to assess whether it is possible to identify differences in body size between the Amarilladesma mactroides (Reeve, 1854) valves found in two sambaquis, and in two transects in the foreshore areas of the state of Rio Grande do Sul in the southernmost part of Brazil. Furthermore, we verify the existence of significant differences between valve sizes of the sambaquis and the current mollusk population at distinct swash zone levels (from one of the current transects only). These analyses were made using parametric (t-test, tt), non-parametric (Mann-Whitney test, mw) analyses, and Bootstrap (bt) techniques for the following scenarios: normal comparison (equal sample numbers) and decreasing the sample number (n = 100, n = 50); comparison with valves bigger than average, and comparison of the valves of sambaquis with those from different parts of the swash zone, which is the beach area that is covered and exposed, alternatively, by the uprush and backwash of the water and is thus limited by it (Masselink and Puleo, 2006; Hughes and Baldock, 2020). Therefore, we aim to contribute to conservation paleobiology by assessing the problems enumerated above and trying to obtain clues regarding how groups of ancient fisher-gardeners exploited these shells (and the swash zones they were collected in).
The Coastal Plain of Rio Grande do Sul (CPRS) is 620 km long (from Chuí Creek at Santa Vitória do Palmar to Mampituba River at Torres), with a slightly wavy shape and SW-NE orientation (Tomazelli and Villwock, 1992). According to Fernandez et al. (2019), this coastal plain is characterized by sandy barriers and lagoons formed during the Quaternary through the juxtaposition of the sedimentary deposits of four barrier-lagoon depositional systems (Villwock et al., 1986). Dillenburg et al. (2020) presented data that indicated a smaller intermediary scale system (the Pleistocene barrier), formed in response to a sea-level highstand between 47.7 and 36.2 ka BP, before the construction of the sambaquis. Each barrier-lagoon system corresponds to a high-frequency depositional sequence (Rosa et al., 2011, 2017). The beach is sandy and dissipative, controlled by a microtidal regime with semidiurnal tides and a mean range of only 0.5 m (Dillenburg et al., 2009). The prevailing wind is from NE (Tomazelli and Villwock, 1992). The sand is fine to very fine-grained; however, between the Rio Grande and Chuí, bioclastic gravel is an important constituent of the sediment (Villwock, 1972; Calliari and Klein, 1993). The beaches of southern Brazil are among the richest and most productive sandy environments; this high productivity allows high diversity and increased survival of organisms, including mollusks such as Amarilladesma mactroides, Donax hanleyanus Philippi, 1847, Eucallista purpurata (Lamarck, 1818), Tivela spp. Link, 1807, and Mactra isabelleana d’ Orbigny (1846) (Calliari et al., 1997; Pinotti et al., 2014).
No endangered or protected species are included here, neither are shell mounds with restricted access; thus, no special permits were required. This study integrates two distinct types of data: (a) samples of specimens from current populations, and (b) archeological samples from the shell-rich layers of two sambaquis. As for the species’ distribution, the recent as well as the archeological samples are conspecific and from sympatric regions.
The transects from where the recent Amarilladesma mactroides valves were collected are located in the northern and southern part of the CPRS foreshore in southernmost Brazil, between Tramandaí (29°58′46.11″S, 50° 7′12.46″W) and Palmares do Sul (30°23′19.93″S, 50°17′2.98″W), and between the Altair shipwreck (32°17′S, 52°15′W) and Sarita Lighthouse (32°37′S, 52°25′W) (Figure 1A). This last transect has information on organisms at different levels of the swash zone (upper, middle and lower levels). Instead of carrying out samplings in Arroio do Sal (where the sambaquis are located), we chose these transects because of time constraints, prevent death of organisms, and monetary reservations; additionally, the material was already available in the case of the Altair shipwreck-Sarita Lighthouse transect (Pinotti, 2014).
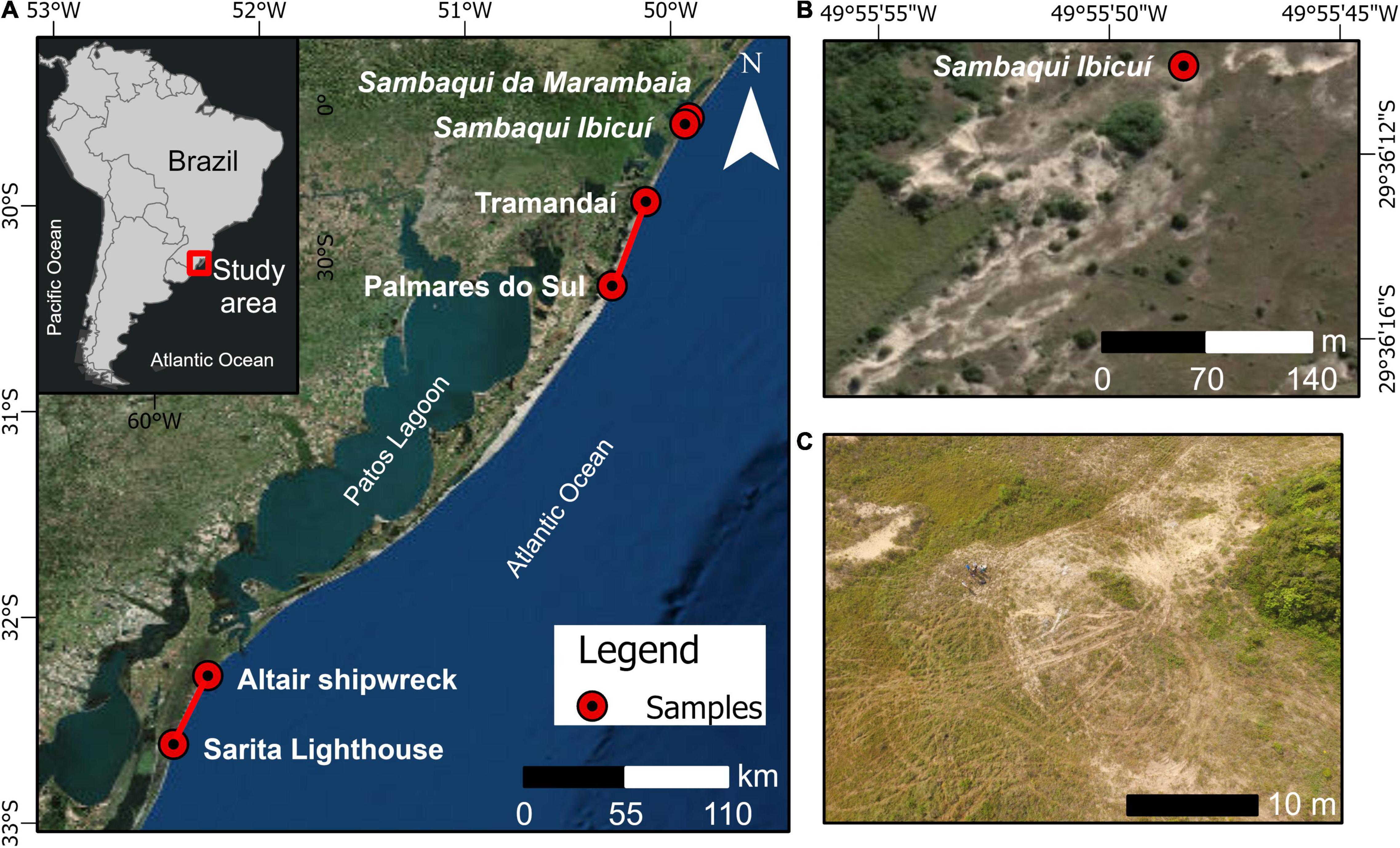
Figure 1. Map of the study area showing (A) sample sites at the Coastal Plain of Rio Grande do Sul–Brazil, (B) the location of Sambaqui Ibicuí, and (C) a drone aerial photograph from Sambaqui Ibicuí depicting off-road vehicles tracks. Basemap–ArcView.
The sampling in the Tramandaí-Palmares do Sul transect entailed collecting empty valves (those of dead organisms) (Figure 2A) in the sand strip in an extension of 10 m at 11 points, 5 km equidistant from each other. After the collection, they were stored in plastic bags labeled with collection point. Arriving at the Laboratory of Sedimentology – Centro de Estudos Costeiros, Limnológicos e Marinhos (CECLIMAR/UFRGS), the samples were then washed, placed in trays to dry naturally, and then stored in separately identified pots. All shells were then archived at the malacological collection of the Museu de Ciências Naturais (MUCIN), UFRGS–CECLIMAR.

Figure 2. Right Amarilladesma mactroides valves from (A) Tramandaí-Palmares do Sul transect and (B) Sambaqui Ibicuí. Scale: 1 cm.
In the Altair shipwreck-Sarita Lighthouse transect, living organisms were collected from different areas of the beach between 2010 and 2011 by Pinotti (2014), with PVC tube of 20 cm diameter (0.03 m2), and sieved in the field with a 0.5 mm mesh. The collected organisms are stored in alcohol at Laboratório de Ecologia de Invertebrados Bentônicos of the Oceanographic Institute of the Universidade Federal do Rio Grande (FURG).
The sambaquis studied for the sampling of ancient valves (Figure 2B) are the Sambaqui Ibicuí (29°36′10.09″S, 49°55′48.39″W; detailed in Figure 1B) and Sambaqui da Marambaia (29°34′23.42″S, 49°54′34.11″W), both located in the Arroio do Sal (Figure 1A). In the last shell mound, an age of 3,050 ± 40 BP was obtained through 14C-graphite dating (Rogge and Schmitz, 2010). It is believed that the Sambaqui Ibicuí was of a similar age, as both the mounds were found on the same dune line and were located close by. These shell mounds were chosen, after a bibliographical review of the sambaquis in the northern part of the CPRS and from field exploration, because they are accessible without restricted access and contain preserved shells. It is noteworthy that while sambaquis are protected and provide relevant information for archeology and paleontology, they are still depredated by people who collect materials illegally, for real estate construction, and off-road vehicle traffic (Figure 1C; Lima, 1999; Wagner, 2009; Afonso, 2017).
The samplings of yellow clam shells in Sambaqui Ibicuí and Sambaqui da Marambaia were acquired by removing blocks on the exposed north face [where a profile had already been carried out on the first shell mound by Wagner (2009)], resulting in a total volume of 298.46 cm3, after which the preserved shells were taken. The samples were then brought to the Laboratory of Sedimentology (UFRGS–CECLIMAR), where they were washed with a 1 mm mesh sieve to remove sediments and plant material. The material that remained in the sieve was dried naturally in trays at the laboratory. Thereafter, the shells were stored in pots labeled with the date of collection, name of the sambaqui, and the collection box. A volume of raw material (37.31 cm3) was removed from the Sambaqui Ibicuí and washed over the sieve; the vertebrate fragments found were separated for possible future studies, and no ceramic artifacts were found. The material is kept at UFRGS–CECLIMAR together with the current valves that are to be listed. It is worth noting that using blocks, rather than archeological stratigraphic columns, would diminish the ability to detect spatial and temporal fine variations. However, it gives a general idea of time-averaged pooled sample shells from fisher-gardeners: samples collected over different seasons and over several years (Parker et al., 2019/2020).
After collecting and cleaning the material (when necessary), the length and height of Amarilladesma mactroides left valves were measured (in mm) using a digital caliper, with an accuracy of 0.1 mm. Bivalve body size was estimated based on maximum length, maximum height, and sample number using the geometric mean (Kosnik et al., 2006). Body size, rather than length or height, is a more reliable method of estimating mollusk size (Kosnik et al., 2006) and has been used in several works globally and throughout geological time, as it is an important comparative ecological variable between different taxonomic levels (e.g., Heim et al., 2017; Payne and Heim, 2020). All the abovementioned data are available in the electronic Supplementary Material.
Comparisons of body size between Amarilladesma mactroides valves from the Tramandaí-Palmares do Sul and Altair shipwreck-Sarita Lighthouse transects, and the valves from sambaquis were performed in the free statistical environment R (R Core Team, 2021). Only the adult specimens (larger than 4.3 cm–Masello and Defeo, 1986) were used for analysis, since juvenile specimens in sambaquis are rare or have not been preserved, possibly due to fragmentation and chemical dissolution (e.g., Claassen, 1998; Ritter et al., 2013; Fiori et al., 2019). Prior to pairwise tests, all datasets were checked for normal distribution through a Shapiro–Wilk test and the sample number (n) was adjusted for each type of comparison, such that the data (body size from sambaquis or recent valves) that had n greater than its comparable were resampled without replacement. The pairwise tests we used for all comparisons are the parametric t-test (tt), the non-parametric Mann-Whitney (mw), and the Bootstrap (bt) (Efron, 1979).
The two-sample Bootstrap was conducted to test the hypothesis that the two samples are not significantly different. In this test, original observations from sambaquis and modern shells were combined, and Bootstrap iterations were drawn (10,000, with replacement) to replicate pairs of the samples. The probability of the difference between the sample means (p-value) was then computed for the two Bootstrap samples. The R scripts implementing these methods are based on Kowalewski and Novack-Gottshall (2010).
The following scenarios were tested for body size comparisons: (a) all sambaquis and recent adult left valves; (b) recent and ancient valves with n = 100; (c) recent and ancient shell mound valves with n = 50; (d) recent and ancient valves with body size greater than average; (e) valves from upper, middle and lower levels of the swash zone of the Altair shipwreck-Sarita Lighthouse transect and valves from sambaquis; (f) valves from the upper level of the swash zone and ancient valves; (g) middle level swash zone and old valves; and (h) valves from the lower level of the swash zone and sambaqui valves.
The sample size for comparing old (from sambaquis) and recent (from the Tramandaí-Palmares do Sul, and Altair shipwreck-Sarita Lighthouse transects) valves was 233, and 177 for valves larger than average (mean sambaquis: 45.95; mean of recent valves: 44.30). The sample size was 74 for comparing body size between the valves of the shell mound, with valves from all levels of the swash zone, from the Altair shipwreck-Sarita Lighthouse transect. When we refine for the different zones, the values are as follows: n = 31 in the comparison shell mound and upper-swash zone; n = 30 in sambaqui and middle level; and n = 13 in the sambaqui and lower swash zone level.
The Shapiro-Wilk test showed that the data from the normal comparison between the body size of shell mound valves (p < 0.001) with the current ones (from the two transects) (p < 0.001) do not have normality, even when performing the Box-Cox conversion; therefore, we did not transform the data that were not normal. When we reduced the sample size to 100 and 50, the body size data for shell mound valves were normal (p = 0.28, and p = 0.71, respectively), but this was not the case for recent valves (p = 0.01 and p = 0.004). Comparison data with sambaquis and recent valves above the mean were also abnormal (p = 0.0004 and p = 0.01). In comparing sambaquis valves with those of all levels in the swash zone (from Altair shipwreck-Sarita Lighthouse transect), the data showed a normal distribution (p = 0.66 and p = 0.11). For the comparisons with the different levels of the swash zone, the data from the shell mounds were all normal (upper: p = 0.37; middle: p = 0.30; lower: p = 0.92) and the data on the current valves had normality only at the upper and lower levels (upper: p = 0.77; middle: p = 0.04; lower: p = 0.09).
There is a significant difference (p < 0.05) between the means (the body size is greater in the sambaquis) of the Amarilladesma mactroides valves present in the shell mounds and those in the current coastal plain in all statistical tests, even when the sample number is reduced (Table 1 and Figure 3). However, when comparing the size of valves with a body size greater than average, we obtained a significant difference only in the Mann-Whitney test (tt, p = 0.24; mw, p = 0.04; bt, p = 0.25) (Table 1 and Figure 3). The means do not differ significantly when the body sizes of sambaqui are compared with those of the Altair shipwreck-Sarita Lighthouse transect (tt, p = 0.21; mw, p = 0.28; bt, p = 0.21) (Table 1 and Figure 4A). Thus, when refining the comparisons for each level of the swash zone (Figure 4B), there is no significant difference in the means when comparing sambaquis valves with the upper level (tt, p = 0.84; mw, p = 0.63; bt, p = 1), or with the lower level (tt, p = 0.16; mw, p = 0.44; bt, p = 0.15); there is a significant difference only a when comparing sambaqui valves with the middle level (tt, p = 0.004; mw, p = 0.004; bt, p = 0.005) (Table 1 and Figure 4A).
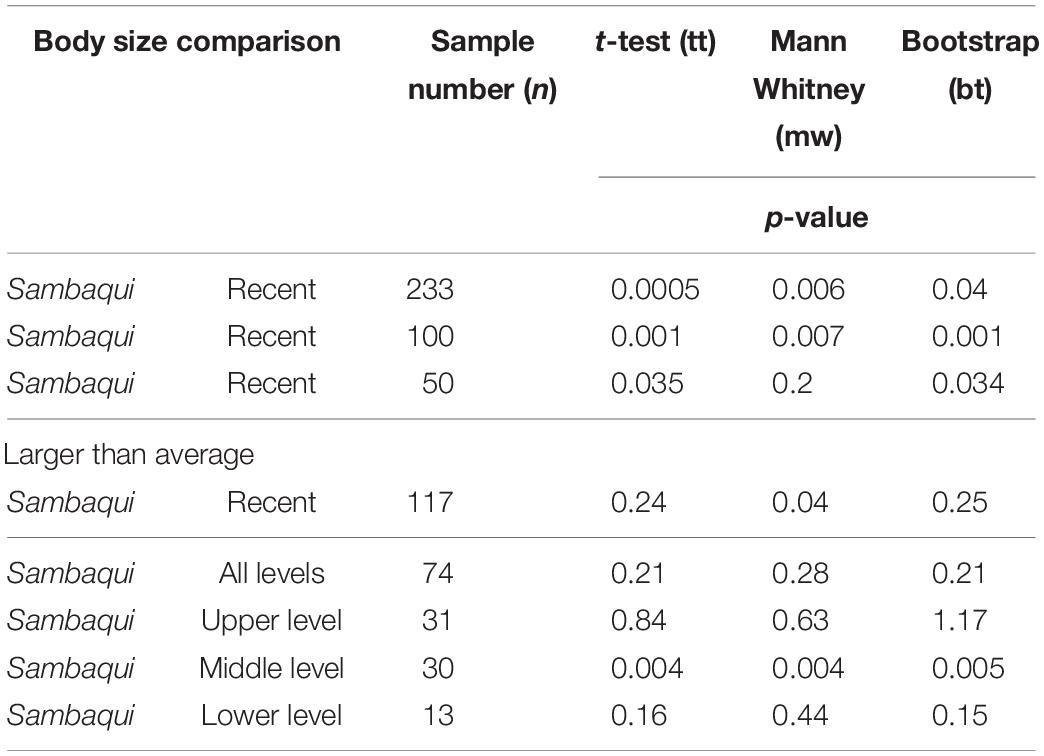
Table 1. Statistical pairwise values between the body size of ancient and recent valves.
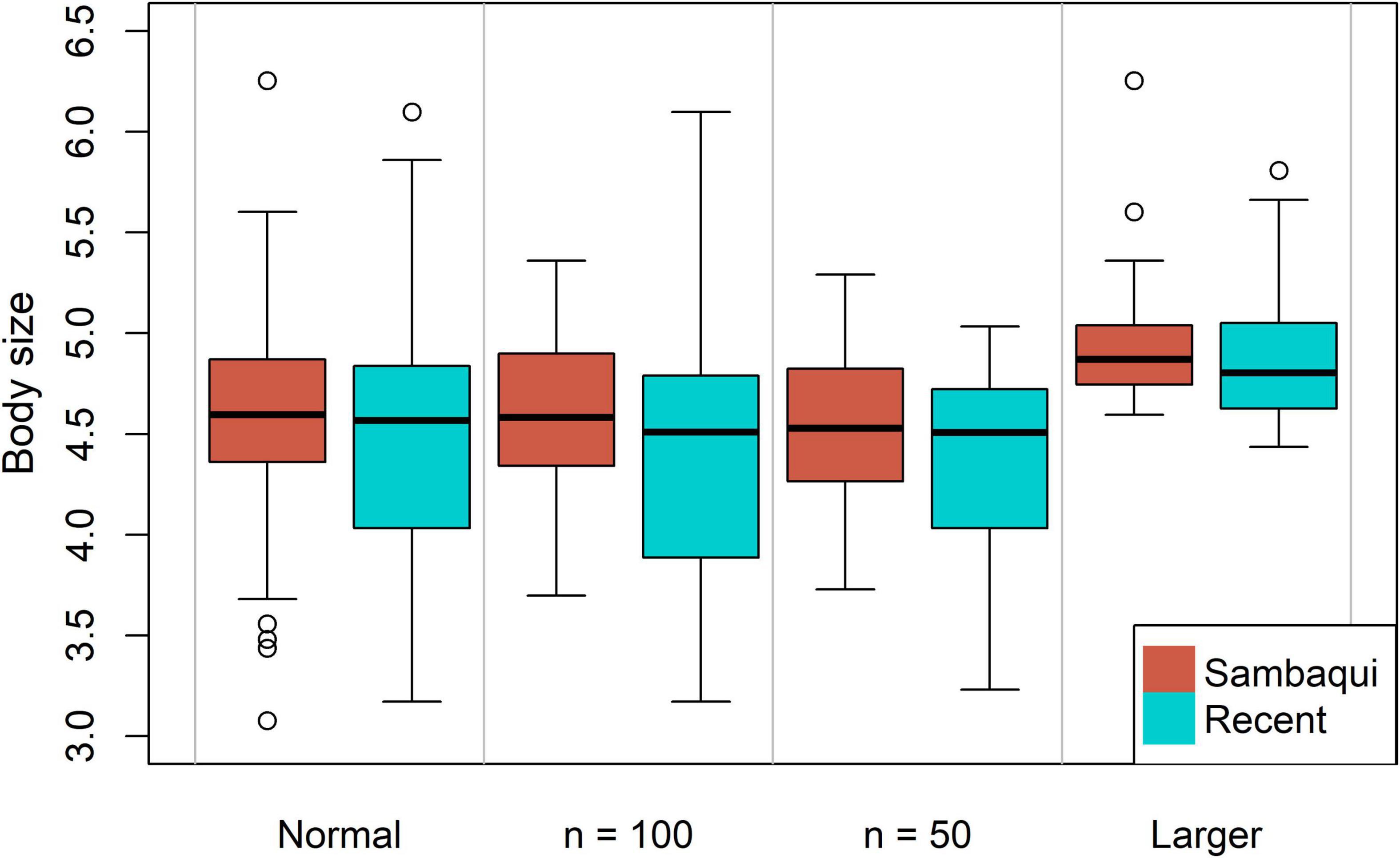
Figure 3. Comparison between body sizes from all sambaquis and recent adult left valves (n = 233, recent data were resampled), recent and ancient valves with n = 100 (both data were resampled), recent and sambaquis valves with n = 50 (both data were resampled), and recent and ancient valves larger than average (n = 117, both data were resampled).
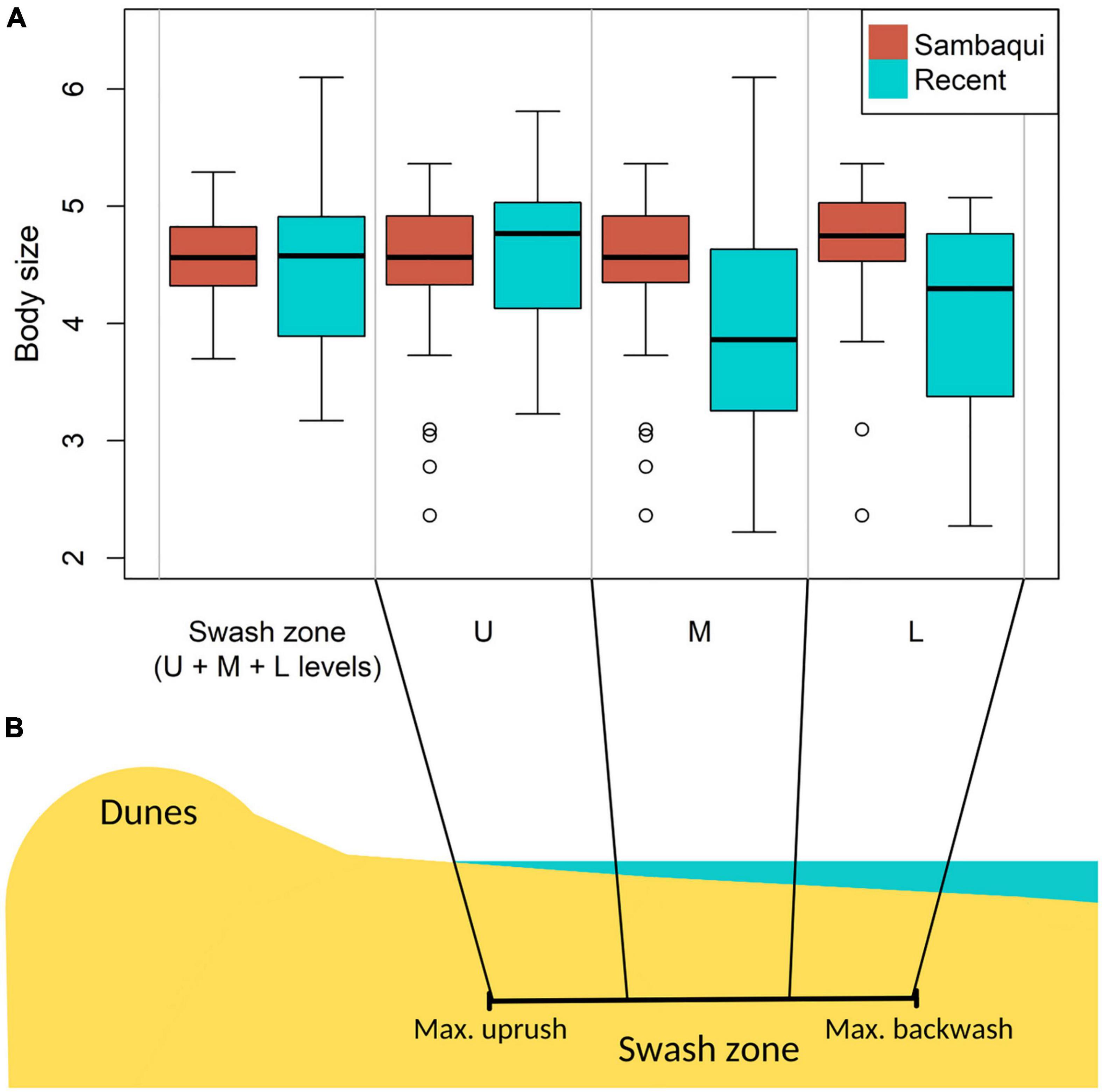
Figure 4. Box plot of comparison between valves body sizes from all levels of the swash zone (upper: U, middle: M, and lower: L) of the transect Altair shipwreck–Sarita Lighthouse and valves from sambaquis (n = 74, sambaqui data were resampled), valves from the upper level of the swash zone and ancient valves (n = 31, sambaqui data were resampled), middle-level swash zone and ancient valves (n = 30, sambaqui data were resampled), and valves from the lower level of the swash zone and sambaqui valves (n = 13, sambaqui data were resampled) (A); and location of the upper, middle and lower levels within swash zone (B).
Body size is an intrinsic characteristic of mollusk valves related to the physiology, lifestyle, and ecology of organisms, and is controlled by abiotic and biotic factors (Campbell, 2008). Specifically, from the body size of fossil valves or those present in archeological sites, relevant information can be obtained regarding paleoenvironmental conditions (Balbo et al., 2011), population size estimates of ancient fisher-gardener groups (Klein and Steele, 2013), the establishment of baselines, and contributions to species management (Kusnerik et al., 2018). However, when comparing this parameter at different times, it is necessary to take some precautions, such as knowing which data and methods to use, the location and its history, considering processes that can lead to biased data, and similar environmental conditions; however, such information is not always available.
An important problem for comparing body size between recent and ancient Amarilladesma mactroides valves is being unsure of how and in which areas of the beach the fisher-gardeners collected or discarded unwanted shell sizes. Ancient populations from different areas of the world collected the shells with their bare hands or simple tools in areas close to the camp, where there were banks of abundant organisms (reviewed in Waselkov, 1987). However, there are no reports or studies on how this was done by the people who collected from sambaquis at the North Coast of CPRS. Thus, the approach taken here uses body size and conservation paleobiology to provide information on how fisher-gardeners exploited these mollusks (Campbell, 2008; Cannarozzi and Kowalewski, 2019).
Interestingly, the statistical tests—and even the t-test that requires data normality, unlike the Mann-Whitney and Bootstrap—had significantly similar results to at least one of the non-parametric tests, especially the Bootstrap approach. Studies suggest that sometimes, due to moderate deviations from normality and large and equal sample size in comparatives, the t-test works without normality (Bartlett, 1935; Kowalewski and Novack-Gottshall, 2010; Poncet et al., 2016); however, there are also studies that show otherwise (Kim and Park, 2019). The t-test makes this dubious, and there are comparisons to non-normal data with small sample sizes; therefore, this test is not indicated here. Additionally, the Mann–Whitney test, a non-parametric rank-based test, is not the best to use, as it consists of a loss of statistical power, increasing the chance of false results (Kowalewski and Novack-Gottshall, 2010); further, there are data with normality. Alternatively, the Bootstrap is a resampling method and there is little or no loss of statistical power (Kowalewski and Novack-Gottshall, 2010). The resulting sample from the bootstrap estimates, even with non-normal data, generally form a Gaussian distribution and are more robust and valid for various conditions (Efron and Tibshirani, 1985). Hence, it is indicated to a greater extent in this paper.
Considering all this, and despite having found a difference between the means of the body size of Amarilladesma mactroides valves from the sambaquis and current population (even with the decrease in the sample size), this result may not be credible and indeed, may be biased, which makes it impossible to discern whether this change in body size is due to human or environmental changes. This is due to the fact that when a more refined comparison was made between the different swash zones (upper, middle, and lower zones) of the Altair shipwreck-Sarita Lighthouse transect, we observed no difference in means between the lower and the upper zones, where the larger organisms stay during winter and summer, respectively, and/or where larger organisms are found at low tide (Fiori and Morsán, 2004; Bergonci and Thomé, 2008).
Thus, we perceive that the groups of fisher-gardeners chose larger shells, possibly for the construction of the sambaquis, when we consider that the organisms of Amarilladesma mactroides were distributed in this manner in the past. Nevertheless, it remains unclear whether or not they fed only on larger shells and discarded the smaller ones. This, however, seems unlikely due to the energy expended in collecting and searching (Waselkov, 1987; Campbell, 2008), since Amarilladesma mactroides is infaunal. Alternatively, they may have collected all the shell sizes and fed on them, using only the biggest ones to make the shell mounds to preserve energy. Finally, they may have collected the larger shells seasonally, at only one level of swash (e.g., Waselkov, 1987; Andersen, 2000; Bailey and Craighead, 2003; Colonese et al., 2011; Cannarozzi and Kowalewski, 2019).
Therefore, when working with the body size of shellfish valves in sambaquis, one must be careful to discern whether the results are biased toward a better similarity with current data, as attempted here. However, it would be valuable to carry out samplings in the different swash zones on the Arroio do Sal, the location of the sambaquis, as it presents the most accurate conditions for comparing the organisms of the present and the past. Additionally, to conduct more expeditions to ascertain whether there are Amarilladesma mactroides valves preserved in the depositional barriers (retro barrier, lagoonal systems) on the coastal plain (e.g., Fiori et al., 2019; Frontini et al., 2021), we would have to use older samples while comparing the body size to prevent artificial biases. As a precaution, it would be important to investigate the body size and horizontal distribution of Amarilladesma mactroides before and after the mass mortality that occurred between 1993 and 2004 (Fiori and Cazzaniga, 1999; Dadon, 2005; Vázquez et al., 2016) because it may change those parameters; however, mass mortalities could also exist continually through time.
The Brazilian shell mounds have been destroyed since the invasion by the Europeans in the 16th century, as they were used in lime making, as mortar in constructions, fertilizer, and in road paving (Lima, 1999). This exploration for industrial purposes ended in the second half of the 20th century, when sambaquis began to be legally protected (Lima, 1999). However, they are still depredated in the present day because, in most cases, there is neither the implementation of in loco protective measures, nor any contact with the local population and visitors (e.g., Costa, 2019; Guimarães et al., 2021), which is why the “know to protect” phrase is important. Hence, the results herein can help future conservation paleobiology studies using sambaqui to access more archeological and paleontological information. This may improve the importance of these mounds and bring them to the current and accessible media, where the population are aware of their responsibility to protect them.
In conclusion, we use conservation paleobiology procedures to assess the problems brought about when comparing the body size of recent and past mollusk valves that are present in shell mounds, in an attempt to show that the choice of tests and data collection sites are essential in this regard. Additionally, we are able to propose that the groups of ancient fisher-gardeners exploited the larger shells for the construction of shell mounds in the upper and lower part of the swash zone; hence, the body size data from sambaquis are biased and is not reliable to infer morphometrics of past populations of mollusks in these shell mounds from the southern Brazilian coast. This research contributes to the existing information on paleontological and archeological areas of the CPRS, and to future worldwide studies that may use shell mounds as geo-archives. Thus, the sambaquis will have increased attention and protection through the responsible agencies, as currently they are somewhat degraded and depredated for not having in loco protection.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
AA and MR designed the study and analyzed the data. FC, FE, and EB helped with field exploration and data collection. RP helped by sharing live datasets. All authors were involved in data interpretation, drafting of the manuscript, and read and approved the final manuscript.
This study was primarily supported by the National Council of Technological and Scientific Development (CNPq/MCTI, Brazil) (422766/2018-6, to the senior authorship, MNR). AA was also grateful for a scholarship supported by the Coordination for the Improvement of Higher Education Personnel (CAPES; process number: 88887.503702/2020-00). The funds had no role in study design, data collection, analyses, the decision to publish, or preparation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.838839/full#supplementary-material
Data Sheet 1 | Data raw from yellow clam (Amarilaldesma mactroides) shells sampled at Sambaqui Ibicuí, Sambaqui da Marambaia and at the transects Tramandaí-Palmares do Sul and Altair shipwreck-Sarita Lighthouse.
Afonso, M. (2017). Arqueologia dos sambaquis no litoral de São Paulo: análise da distribuição dos sítios e cronologia. Especiaria Cad. de Ciências Hum. 17, 203–227.
Albano, P. G., Azzarone, M., Amati, B., Bogi, C., Sabelli, B., and Rilov, G. (2020). Low diversity or poorly explored? Mesophotic molluscs highlight undersampling in the Eastern Mediterranean. Biodivers. Conserv. 29, 4059–4072. doi: 10.1007/s10531-020-02063-w
Álvarez-Fernández, E., Chauvin, A., Cubas, M., Arias, P., and Ontañón, R. (2011). Mollusc shell sizes in archaeological contexts in Northern Spain (13 200 to 2600 cal BC): new data from la garma A and los gitanos (Cantabria). Archaeometry 53, 963–985. doi: 10.1111/j.1475-4754.2011.00589.x
Andersen, S. H. (2000). ‘Køkkenmøddinger’ (shell middens) in Denmark: a survey. Proc. Prehis. Soc. 66, 361–384. doi: 10.1017/S0079497X00001857
Bailey, G. N., and Craighead, A. S. (2003). Late Pleistocene and Holocene coastal palaeoeconomies: a reconsideration of the molluscan evidence from northern Spain. Geoarchaeology 18, 175–204. doi: 10.1002/gea.10057
Balbo, A., Madella, M., Godino, I. B., and Álvarez, M. (2011). Shell midden research: an interdisciplinary agenda for the quaternary and social sciences. Quat. Int. 239, 147–152. doi: 10.1016/j.quaint.2011.03.032
Barnosky, A. D., Matzke, N., Tomiya, S., Wogan, G. O., Swartz, B., Quental, T. B., et al. (2011). Has the Earth’s sixth mass extinction already arrived? Nature 471, 51–57. doi: 10.1038/nature09678
Bartlett, M. S. (1935). The effect of non-normality on the t distribution. Math. Proc. Camb. Phil. Soc. 31, 223–231. doi: 10.1017/S0305004100013311
Bergonci, P. E. A., and Thomé, J. W. (2008). Vertical distribution, segregation by size and recruitment of the yellow clam Mesodesma mactroides Deshayes, 1854 (Mollusca, Bivalvia, Mesodesmatidae) in exposed sandy beaches of the Rio Grande do Sul state, Brazil. Braz. J. Biol. 68, 297–305. doi: 10.1590/S1519-69842008000200010
Boivin, N. L., Zeder, M. A., Fuller, D. Q., Crowther, A., Larson, G., Erlandson, J. M., et al. (2016). Ecological consequences of human niche construction: examining long-term anthropogenic shaping of global species distributions. Proc. Natl. Acad. Sci. U.S.A. 113, 6388–6396. doi: 10.1073/pnas.1525200113
Calliari, L. J., Garcia, C. A. E., Niencheski, L. F., Fillmann, G., Seeliger, U., Gianuca, N. M., et al. (1997). “Coastal and marine environments and their biota,” in Subtropical Convergence Environments, eds U. Seeliger, C. Odebrecht, and J. P. Castello (Heidelberg: Springer), 91–160. doi: 10.1007/978-3-642-60467-6_6
Calliari, L. J., and Klein, A. H. (1993). Características morfodinâmicas e sedimentológicas das praias oceânicas entre Rio Grande e Chuí, RS. Pesqui. em Geociênc. 20, 45–56. doi: 10.22456/1807-9806.21281
Campbell, G. (2008). Beyond means to meaning: using distributions of shell shapes to reconstruct past collecting strategies. Environ. Archaeol. 13, 111–121. doi: 10.1179/174963108X343236
Cannarozzi, N. R., and Kowalewski, M. (2019). Seasonal oyster harvesting recorded in a late Archaic period shell ring. PLoS One 14:e0224666. doi: 10.1371/journal.pone.0224666
Colonese, A. C., Camarós, E., Verdún, E., Estevez, J., Giralt, S., and Rejas, M. (2011). Integrated archaeozoological research of shell middens: new insights into hunter-gatherer-fisher coastal exploitation in Tierra del Fuego. J. Isl. Coast. Archaeol. 6, 235–254. doi: 10.1080/15564894.2011.586088
Conservation Paleobiology Workshop (2012). Conservation Paleobiology: Opportunities for the Earth Sciences. Report to the Division of Earth Sciences, National Science Foundation. New York, NY: Paleontological Research Institution.
Costa, B. R. (2019). “Percebendo o sambaqui: simetria aplicada à gestão do patrimônio arqueológico em Joinville/SC,” in Seminário Preservação de Patrimônio Arqueológico, orgs, eds G. N. Campos and M. Granato (Rio de Janeiro: Museu de Astronomia e Ciências Afins), 403–415.
Dadon, J. R. (2005). Changes in the intertidal community structure after a mass mortality event in sandy beaches of Argentina. Contrib. Zool. 74, 27–39. doi: 10.1163/18759866-0740102002
d’ Orbigny, A. D. (1846). “Mollusques,” in Voyage Dans l’Amérique Méridionale (Le Brésil, La République Orientale de L’Uruguay, La République Argentine, La Patagonie, La République du Chili, La République de Bolivia, La République du Pérou), exécuté pendant les années, 81, ed. C. P. Bertrand (Paris: Chez Ve. Levrault), 1835–1847.
DeBlasis, P., Fish, S. K., Gaspar, M. D., and Fish, P. R. (1998). Some references for the discussion of complexity among the sambaqui moundbuilders from the southern shores of Brazil. Revista de Arqueol. Am. 15, 75–105.
DeBlasis, P., Kneip, A., Scheel-Ybert, R., Giannini, P. C., and Gaspar, M. D. (2007). Sambaquis e paisagem: dinâmica natural e arqueologia regional no litoral do Sul do Brasil. Arqueol. Suram. 3, 29–61.
Diaz, R. J., and Rosenberg, R. (2008). Spreading dead zones and consequences for marine ecosystems. Science 321, 926–929. doi: 10.1126/science.1156401
Dietl, G. P. (2019). Conservation palaeobiology and the shape of things to come. Philos. Trans. R. Soc. Lond. 374:20190294. doi: 10.1098/rstb.2019.0294
Dietl, G. P., and Flessa, K. W. (2009). Conservation Paleobiology: Using the Past to Manage for the Future. Lubbock, TX: Paleontological Society.
Dietl, G. P., and Flessa, K. W. (2011). Conservation Paleobiology: putting the dead to work. Trends Ecol. Evol. 26, 30–37. doi: 10.1016/j.tree.2010.09.010
Dietl, G. P., and Flessa, K. W. (2017). Conservation Paleobiology: Science and Practice. Chicago, IL: University of Chicago Press.
Dillenburg, S. R., Barboza, E. G., Rosa, M. L. C. C., Caron, F., Cancelli, R., Santos Fischer, C. B., et al. (2020). Sedimentary records of Marine Isotopic Stage 3 (MIS 3) in southern Brazil. Geo Mar. Lett. 40, 1099–1108. doi: 10.1007/s00367-019-00574-2
Dillenburg, S. R., Barboza, E. G., Tomazelli, L. J., Hesp, P. A., Clerot, L. C. P., and Ayup-Zouain, R. N. (2009). The holocene coastal barriers of Rio Grande do Sul. geology and geomorphology of holocene coastal barriers of Brazil. Lect. Notes Earth Sci. 107, 53–91. doi: 10.1007/978-3-540-44771-9_3
Dirzo, R., Young, H. S., Galetti, M., Ceballos, G., Isaac, N. J., and Collen, B. (2014). Defaunation in the Anthropocene. Science 345, 401–406. doi: 10.1126/science.1251817
Efron, B. (1979). Bootstrap methods: another look at the jackknife. Ann. Statist. 7, 1–26. doi: 10.1214/aos/1176344552
Efron, B., and Tibshirani, R. (1985). The bootstrap method for assessing statistical accuracy. Behaviormetrika 12, 1–35. doi: 10.2333/bhmk.12.17_1
Ewers, R. M., and Didham, R. K. (2006). Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. Camb. Philos. Soc. 81, 117–142. doi: 10.1017/S1464793105006949
Fernandez, G. B., Rocha, T. B., Barboza, E. G., Dillenburg, S. R., Rosa, M. L. C. C., Angulo, R. J., et al. (2019). “Natural landscapes along brazilian coastline,” in The Physical Geography of Brazil - Environment, Vegetation and Landscape, eds A. A. R. Salgado, L. J. C. Santos, and J. C. Paisani (Salgado: American Academy of Religion), 199–218. doi: 10.1007/978-3-030-04333-9_10
Ferrasso, S., Fiorentin, G. L., and Schmitz, P. I. (2016). Identificação de remanescentes conquiliológicos de um assentamento holocênico na planície costeira do Rio Grande do Sul: contribuições sob o enfoque zooarqueológico. Pesqui. Antropol. São Leopoldo. 72, 225–266.
Figuti, L. (2005). A recipe for a sambaqui: considerations on Brazilian shell mound composition and building. Early Hum. Impact Megamollusks. 1, 67–80.
Fiori, S., Frontini, R., and Bayón, C. (2019). Morphometric analysis of the yellow clam (Amarilladesma mactroides) to interpret fragmentary archaeological specimens. J. Isl. Coast. Archaeol. 14, 291–300. doi: 10.1080/15564894.2018.1521482
Fiori, S. M., and Cazzaniga, N. J. (1999). Mass mortality of the yellow clam, Mesodesma mactroides (Bivalvia: Mactracea) in Monte Hermoso beach, Argentina. Biol. Conser. 89, 305–309. doi: 10.1016/S0006-3207(98)00151-7
Fiori, S. M., and Morsán, E. M. (2004). Age and individual growth of Mesodesma mactroides (Bivalvia) in the southernmost range of its distribution. ICES J. Mar. Sci. 61, 1253–1259. doi: 10.1016/j.icesjms.2004.07.025
Fossile, T., Ferreira, J., da Rocha Bandeira, D., Dias-da-Silva, S., and Colonese, A. C. (2019). Integrating zooarchaeology in the conservation of coastal-marine ecosystems in Brazil. Quat. Int. 545, 38–44. doi: 10.1016/j.quaint.2019.04.022
Frontini, R., Vecchi, R., Bayón, C., and Fiori, S. (2021). Arqueología de la laguna sauce grande (partido de monte hermoso, provincia de buenos aires). Relaciones. 46, 121–140.
Guimarães, G. M., Zamparetti, B. C., Farias, D. S. E., and Anjos, F. A. (2021). “Preservação e valorização dos sambaquis de Laguna-SC na perspectiva do turismo arqueológico,” in Patrimônio Cultural, Direito e Meio Ambiente: Arqueologia e Turismo Sustentável, eds J. B. Campos, M. H. S. G. Rodrigues, N. I. Ladwig, P. P. A. Funari, and L. Oosterbeek (Criciúma, SC: UNESC). doi: 10.18616/pcultura11
Halpern, B. S., Walbridge, S., Selkoe, K. A., Kappel, C. V., Micheli, F., D’Agrosa, C., et al. (2008). A global map of human impact on marine ecosystems. Science 319, 948–952. doi: 10.1126/science.1149345
Heim, N. A., Payne, J. L., Finnegan, S., Knope, M. L., Kowalewski, M., Lyons, S. K., et al. (2017). Hierarchical complexity and the size limits of life. Proc. R. Soc. B. Biol. Sci. 284:20171039. doi: 10.1098/rspb.2017.1039
Hughes, M. G., and Baldock, T. E. (2020). “The swash zone,” in Sandy Beach Morphodynamics, eds D. W. T. Jackson and A. D. Short (Amsterdam: Elsevier), 155–186. doi: 10.1016/B978-0-08-102927-5.00008-4
Kim, T. K., and Park, J. H. (2019). More about the basic assumptions of t-test: normality and sample size. Korean J. Anesthesiol. 72:331. doi: 10.4097/kja.d.18.00292
Klein, R. G., and Steele, T. E. (2013). Archaeological shellfish size and later human evolution in Africa. Proc. Natl. Acad. Sci. U.S.A. 110, 10910–10915. doi: 10.1073/pnas.1304750110
Kosnik, M. A., Jablonski, D., Lockwood, R., and Novack-Gottshall, P. M. (2006). Quantifying molluscan body size in evolutionary and ecological analyses: maximizing the return on data-collection efforts. Palaios 21, 588–597. doi: 10.2110/palo.2006.p06-012r
Kowalewski, M. (2017). “The youngest fossil record and conservation biology: Holocene shells as eco-environmental recorders,” in Conservation Paleobiology: Science and Practice, eds G. P. Dietl and K. W. Flessa (Chicago, IL: The University of Chicago Press), 7–29. doi: 10.7208/chicago/9780226506869.003.0001
Kowalewski, M., and Novack-Gottshall, P. (2010). Resampling methods in paleontology. Paleontol. Soc. Pap. 16, 19–54. doi: 10.1017/s1089332600001807
Kowalewski, M., Serrano, G. E. A., Flessa, K. W., and Goodfriend, G. A. (2000). Dead delta’s former productivity: two trillion shells at the mouth of the Colorado River. Geology 28, 1059–1062. doi: 10.1130/0091-7613200028<1059:DDFPTT>2.0.CO;2
Kusnerik, K. M., Lockwood, R., and Grant, A. N. (2018). “Using the fossil record to establish a baseline and recommendations for oyster mitigation in the Mid-Atlantic US,” in Marine Conservation Paleobiology, eds C. L. Tyler and C. L. Schneider (Cham: Springer), 75–103. doi: 10.1007/978-3-319-73795-9_5
Lamarck, [J.-B. M]. (1818). Histoire Naturelle des Animaux Sans vErtèbres. Tome 5. Paris: Deterville/Verdière.
Lima, T. A. (1999). Em busca dos frutos do mar os pescadores-coletores do litoral centro-sul do Brasil. Revista USP 44, 270–327. doi: 10.11606/issn.2316-9036.v0i44p270-327
Link, D. H. F. (1807). Beschreibung der naturalien-sammlung der universität zu rostock. Adlers Erben. 1 Abt, 1–50. doi: 10.1159/000404737
Mannino, M. A., and Thomas, K. D. (2002). Depletion of a resource? The impact of prehistoric human foraging on intertidal mollusc communities and its significance for human settlement, mobility and dispersal. World Archaeol. 33, 452–474. doi: 10.1080/00438240120107477
Masello, A., and Defeo, O. (1986). Determinación de la longitud de primera madurez sexual en Mesodesma mactroides (Deshayes 1854). Com. Soc. Malacológica Urug. 6, 387–395.
Masselink, G., and Puleo, J. A. (2006). Swash-zone morphodynamics. Contin. Shelf Res. 26, 661–680. doi: 10.1016/j.csr.2006.01.015
Mendes, A. B., Silva, E. P., and Duarte, M. R. (2019). Can sambaquis (shell mounds) be used as records of the Holocene marine fish biodiversity? Biodivers. Conserv. 29, 39–56. doi: 10.1007/s10531-019-01868-8
Millenium Ecosystem Assessment [MEA] (2005). Ecosystems and Human Well-Being: Synthesis. Washington, DC: Island Press.
Monsarrat, S., Jarvie, S., and Svenning, J. C. (2019). Anthropocene refugia: integrating history and predictive modelling to assess the space available for biodiversity in a human-dominated world. Philos. Trans. R. Soc. 374, 20190219. doi: 10.1098/rstb.2019.0219
Parker, W. G., Yanes, Y., Mesa Hernández, E., Hernández Marrero, J. C., Pais, J., and Surge, D. (2019/2020). Scale of time-averaging in archaeological shell middens from the Canary Islands. Holocene 30, 258–271. doi: 10.1177/0959683619883020
Payne, J. L., and Heim, N. A. (2020). Body size, sampling completeness, and extinction risk in the marine fossil record. Paleobiology. 46, 23–40. doi: 10.1017/pab.2019.43
Philippi, R. A. (1847). Donacidae: Bivalvia. Bol. Zool. Univ. São Paulo. 3, 121–142. doi: 10.11606/issn.2526-3358.bolzoo.1978.121656
Pinotti, R. M. (2014). A Estrutura do Macrozoobentos em Uma Praia Arenosa no Extremo Sul do Brasil: a Influência da Morfodinâmica de Feições Geomorfológicas e de Eventos Meteorológicos. [Dissertation/Master’s Thesis]. Rio Grande TX: Universidade Federal do Rio Grande.
Pinotti, R. M., Minasi, D. M., Colling, L. A., and Bemvenuti, C. E. (2014). A review on macrobenthic trophic relationships along subtropical sandy shores in southernmost Brazil. Biota Neotrop. 14, 1–12. doi: 10.1590/1676-06032014006914
Poncet, A., Courvoisier, D. S., Combescure, C., and Perneger, T. V. (2016). Normality and sample size do not matter for the selection of an appropriate statistical test for two-group comparisons. Methodology 12, 61–71. doi: 10.1027/1614-2241/a000110
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reeve, L. A. (1854). “Monograph of the genus Mesodesma,” in Conchologia Iconica, or, Illustrations of the Shells of Molluscous Animals (London: Reeve & Co). Available online at: https://www.biodiversitylibrary.org/bibliography/8129
Ritter, M. N., Erthal, F., and Coimbra, J. C. (2013). Taphonomic signatures in molluscan fossil assemblages from the Holocene lagoon system in the northern part of the coastal plain, Rio Grande do Sul State, Brazil. Quat. Int. 305, 5–14. doi: 10.1016/j.quaint.2013.03.013
Rogge, J. H., and Schmitz, P. I. (2010). Projeto Arroio do Sal: a ocupação indígena pré-histórica no litoral norte do RS. Pesqui. Antropol. 68, 167–225.
Rosa, M. L. C. C., Barboza, E. G., Abreu, V. S., Tomazelli, L. J., and Dillenburg, S. R. (2017). High-frequency sequences in the Quaternary of Pelotas Basin (coastal plain): a record of degradational stacking as a function of longer-term base-level fall. Braz. J. Geol. 47, 183–207. doi: 10.1590/2317-4889201720160138
Rosa, M. L. C. C., Barboza, E. G., Dillenburg, S. R., Tomazelli, L. J., and Ayup-Zouain, R. N. (2011). The Rio Grande do Sul (southern Brazil) shoreline behavior during the Quaternary: a cyclostratigraphic analysis. J. Coast. Res. SI 64, 686–690.
Scheel-Ybert, R., and Boyadjian, C. (2020). Gardens on the coast: considerations on food production by Brazilian shell mound builders. J. Anthropol. Archaeol. 60:101211. doi: 10.1016/j.jaa.2020.101211
Schmitz, P. I. (2006). Arqueologia do Rio Grande do Sul, Brasil. São Leopoldo: Instituto Anchietano de Pesquisas – UNISINOS.
Silva, E. P., Pádua, S. C., Souza, R. C. C. L., and Duarte, M. R. (2017). “Shell mounds of the Southeast coast of Brazil: recovering information on past malacological biodiversity,” in Zooarchaeology in the Neotropics, eds M. Mondini, A. Muñoz, and P. Fernández (Cham: Springer). doi: 10.1007/978-3-319-57328-1_4
Tomazelli, L., and Villwock, J. (1992). Considerações sobre o ambiente praial e a deriva litorânea de sedimentos ao longo do litoral norte do Rio Grande do Sul. Brasil. Pesqui. Geoci. 19, 3–12. doi: 10.22456/1807-9806.21318
Vázquez, N., Fiori, S. M., Arzul, I., Carcedo, C., and Cremonte, F. (2016). Mass mortalities affecting populations of the yellow clam Amarilladesma mactroides along its geographic range. J. Shellfish Res. V. 35, 739–745. doi: 10.2983/035.035.0403
Villwock, J. A. (1972). Contribuição a Geologia do Holoceno da Província Costeira do Rio Grande do Sul-Brasil. [Dissertation/Master’s Thesis]. Porto Alegre: Universidade Federal do Rio Grande do Sul.
Villwock, J. A., Tomazelli, L. J., Loss, E. L., Dehnhardt, E. A., Horn Filho, N. O., Bachi, F. A., et al. (1986). “Geology of the Rio Grande do Sul Coastal province,” in Quaternary of South America and Antarctic Peninsula, 4, ed. J. Rabassa (Rotterdam: A. A. Balkema), 79–97. doi: 10.1201/9781003079316-5
Wagner, G. P. (2009). Sambaquis da Barreira da Itapeva: uma Perspectiva Geoarqueológica. [Dissertation/Master’s Thesis]. Porto Alegre: Pontifícia Universidade Católica do Rio Grande do Sul.
Waselkov, G. A. (1987). Shellfish gathering and shell midden archaeology. Adv. Archaeol. Method Theory 10, 339–362. doi: 10.1016/B978-0-12-003110-8.50006-2
Keywords: Sambaqui, body size, mollusks, statistical analysis, swash zone
Citation: Assumpção ACA, Caron F, Erthal F, Barboza EG, Pinotti RM and Ritter MN (2022) The Tricky Task of Fisher-Gardener Research in Conservation Paleobiology. Front. Ecol. Evol. 10:838839. doi: 10.3389/fevo.2022.838839
Received: 18 December 2021; Accepted: 15 March 2022;
Published: 18 April 2022.
Edited by:
Hermínio Araújo-Júnior, Rio de Janeiro State University, BrazilReviewed by:
Thomas Mark Cronin, United States Geological Survey (USGS), United StatesCopyright © 2022 Assumpção, Caron, Erthal, Barboza, Pinotti and Ritter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matias do Nascimento Ritter, bWF0aWFzLnJpdHRlckB1ZnJncy5icg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.