
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Ecol. Evol., 17 March 2022
Sec. Urban Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.834436
This article is part of the Research TopicCognition and Adaptation to Urban EnvironmentsView all 6 articles
 Ernő Vincze1,2,3*
Ernő Vincze1,2,3* Bálint Kovács1,4
Bálint Kovács1,4
Cognitive abilities are often assumed to be advantageous in urban habitats, but relatively few studies tested this assumption. In a meta-analysis, we tested whether urban animals have better problem-solving abilities compared to their less urbanized conspecifics. After screening 210 papers we collected by keyword search and forward search, we found 12 studies that compared the ability to solve food-extraction or obstacle-removal problems between urban and non-urban populations of the same animal species. These studies were published between 2009 and 2021, and were performed mostly on birds, whereas a quarter of them used mammals as study species. We found a statistically non-significant trend that urban animals are more successful and faster problem-solvers compared to their less urbanized conspecifics. However, both solving success and solving latency effect sizes were highly heterogeneous, therefore hard to generalize. Though the sample was too low to test the factors explaining this high heterogeneity, we suggest that it may be explained by variation in task types, study species, definitions of urbanization, whether the study was performed on captive or free-living animals, geographical location, or publication bias in both directions. Altogether, more studies are needed to either confirm or disprove this trend.
Urbanization is one of the most profound environmental changes of our time, affecting numerous species (Grimm et al., 2008; Sol et al., 2013; Seress and Liker, 2015). Although the majority of species is declining in these anthropogenic environments (Grimm et al., 2008), many species successfully exploit these urban habitats (Sol et al., 2013). What makes an animal a successful urban exploiter is a hot topic in behavioral ecology. A number of factors have been suggested, from diet (Evans et al., 2011) to body size (Bateman and Fleming, 2012) to stress tolerance (Partecke et al., 2006).
Cognitive abilities have often been suggested to be advantageous in urban habitats (Lee and Thornton, 2021), for several reasons. Urban habitats are complex environments where animals face novel challenges on a regular basis, and cognitive abilities, such as innovation and learning, are advantageous in such novel habitats (Sol et al., 2002). Good cognitive abilities can also be beneficial during human-wildlife interactions, which are the most common in urban habitats with high human population density (Goumas et al., 2020). This leads to two different predictions. First, species with better cognitive abilities should be more successful urban exploiters compared to other species. Brain size is often a good proxy for cognitive abilities, and bird species with larger brains appear to be better at colonizing urban habitats (Carrete and Tella, 2011; Maklakov et al., 2011; Sayol et al., 2020). Second, within a species, individuals from urban populations should have more advanced cognitive abilities than those from less urbanized habitats. This idea is supported by the larger brain size of urbanized small mammals compared to their non-urban conspecifics (Snell-Rood and Wick, 2013). By contrast, urban environment can also have environmental factors that hinder cognitive performance, such as artificial light at night (Jha and Kumar, 2017, but see Yorzinski et al., 2017). Therefore, it is unclear how urbanization is expected to affect cognitive performance, and the results so far have been equivocal (Griffin et al., 2017).
In this study, we aim to explore the differences in cognitive abilities of urban and non-urban conspecifics. We focus on one specific cognitive skill: problem-solving, which has been suggested to be a good proxy of cognitive abilities in general (Morand-Ferron et al., 2016). We review the literature available on urbanization’s effects on cognitive abilities and test with meta-analytical methods whether animals from urban populations are better problem-solvers compared to their non-urban conspecifics.
We followed the preferred reporting items for systematic reviews and meta-analysis (PRISMA) protocol (Moher et al., 2009) to screen the literature on urbanization’s effects on animal cognition (Figure 1). First, on 24th of January, 2021, we ran a search in Web of Science with the following keyword combination: urban* + (cogni* OR solv*). Additionally, on 9th of March, we performed a forward search in Google Scholar by looking at all papers that cited one of the earliest studies focusing on the topic (Papp et al., 2015). After removing duplicates, we thoroughly screened the abstracts of these papers (BK and EV screened the abstracts from the keyword search and the forward search, respectively). During the screening we checked whether or not the papers met the following criteria: (i) the study system was a non-human animal species; (ii) the study compared urban and non-urban individuals of the same species; and (iii) the behavior tested was either food extraction from a container, or removal of an obstacle.
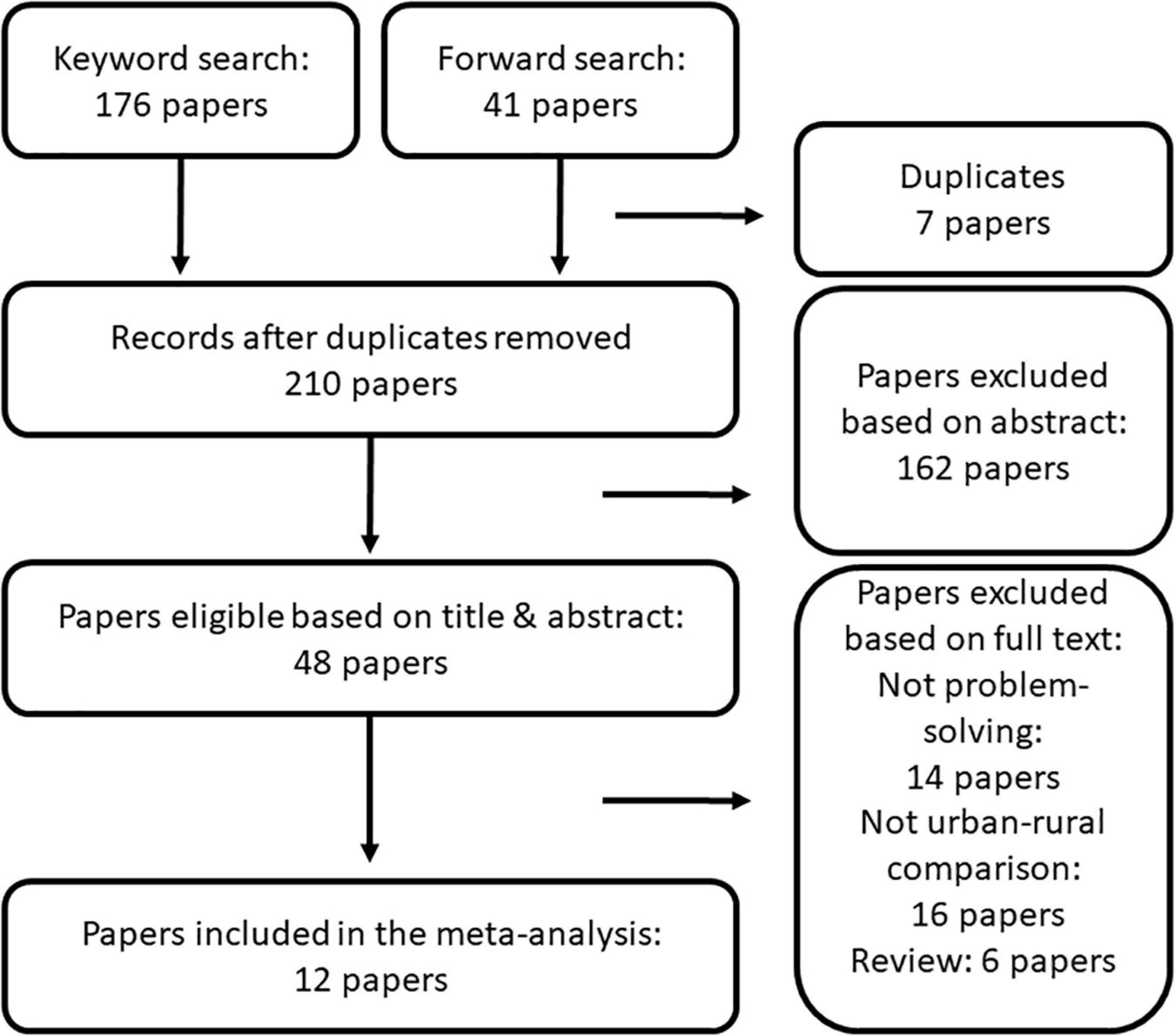
Figure 1. PRISMA diagram of our literature screening.
When defining our criteria, we decided to exclude papers that tested cognitive tasks other than problem-solving, because these tasks require qualitatively very different cognitive and motoric abilities and thus are not comparable to problem-solving skills, and there were too few of each type to be tested in a separate model. These tasks include discrimination between dangerous and non-dangerous humans (Vincze et al., 2015; Vincze et al., 2019); spatial memory (Morand-Ferron et al., 2019; Thompson and Morand-Ferron, 2019); or discriminating between safe and unsafe shelters (Kang et al., 2018; Batabyal and Thaker, 2019). However, we included one test where the birds had to remove an obstacle to enter their nest instead of accessing food, because it still required the bird to manipulate an obstacle to access a reward, thus presumably required similar cognitive and motoric skills to the food access tests (Preiszner et al., 2017). We also decided to exclude papers that, rather than comparing urban and non-urban populations of the same species, defined “urbanization” as the distance from the edge of a residential area (Grunst et al., 2020) or compared subspecies with different history in urban habitats (Vrbanec et al., 2021).
After excluding the papers that did not meet the criteria based on their titles and abstracts, we read the Methods and Results section of the remaining papers, and excluded further papers that did not meet the above criteria, ending with a final set of 12 papers. We contacted the authors for additional information (interpretation of published results as well as additional data).
We collected two different types of effect sizes. Urbanization’s effect on solving success was estimated by calculating logarithmic odds ratios (Nakagawa and Cuthill, 2007) from the exact numbers of urban and non-urban individuals that were successful and unsuccessful in solving the task. In one case, instead of testing the animals individually, the study compared the solving success of breeding pairs of birds (Preiszner et al., 2017). In this study, we calculated the odds ratios from the numbers of successful and unsuccessful urban and rural pairs rather than individuals. Standard errors of these logarithmic odds ratios were calculated as the square root of the sum of the reciprocals of the number of successful urban, unsuccessful urban, successful non-urban, and unsuccessful non-urban individuals (or pairs), following the formula of Nakagawa and Cuthill (2007). When animals were given multiple tasks in the same study, we extracted separate effect sizes for each task type, except for one paper where they only provided the cumulative number of successful and unsuccessful tests (Mazza and Guenther, 2021).
Urbanization’s effect on solving latency was extracted from the parameter estimates (Z-value or T-value) and standard errors of Cox models (11 effect sizes), linear models (one effect size), linear mixed-effects models (two effect sizes), or generalized linear mixed-effects models (six effect sizes). As the models differed in whether they used the more urbanized or the less urbanized population as the reference group, we inverted some of the parameter estimates (seven out of 20) so that positive effect sizes always indicated that the urban animals were more successful (had shorter latencies). We did not include estimates from models where urbanization was in an interaction with another predictor. Again, when there were multiple tasks in the study, we extracted separate effect sizes for each task type, except for one study where all task types were tested in the same model and only one latency estimate was provided (Mazza and Guenther, 2021).
Due to the high heterogeneity in task types and urbanization gradients in the study, as well as multiple effect sizes in the same study, we ran random-effect meta-analyses (Hedges and Vevea, 1998) where we tested whether the mean of effect sizes differs from zero. We could not run a sub-group analysis of problem-solving test types because of the low sample sizes in each test type (see Section “Results”). To check whether there was publication bias or any outliers, we created funnel plots for both models and visually interpreted them, but we had too few effect sizes to statistically test for publication bias. All tests were conducted in RStudio (Racine, 2012), using the rmeta (Lumley, 2018), dmetar (Harrer et al., 2021), meta (Schwarzer, 2007), and metafor (Viechtbauer, 2015) packages.
We found 176 papers with the keyword search and 41 papers with the forward search. After removing seven duplicates, we checked the titles and the abstracts of 210 papers, based on which we excluded 162 papers and checked the full text of 48 papers, out of which 12 papers met our full criteria. We extracted 17 solving success odds ratios from nine papers and 20 solving latency estimates from 11 papers (Table 1).
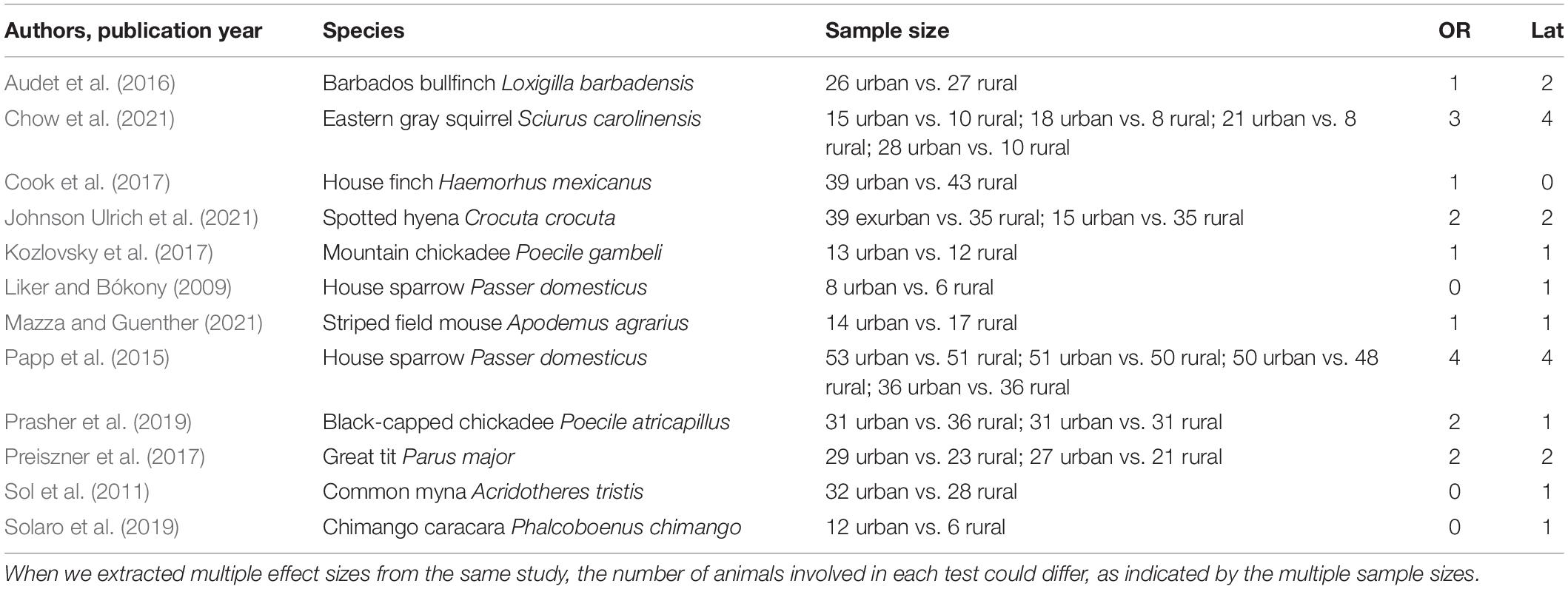
Table 1. List of papers included, with study species, sample size [flocks in Liker and Bókony (2009); pairs in Preiszner et al. (2017); individuals in every other study], and number of solving success odds ratios (OR) and solving latencies (Lat) collected from the study.
The oldest paper we found was published in 2009 (Liker and Bókony, 2009); the most recent ones were published in 2021 (Chow et al., 2021; Johnson Ulrich et al., 2021; Mazza and Guenther, 2021). Nine out of 12 studies (75%) were performed on birds (out of which, eight studies were on Passeriformes and only one on Falconiformes); the remaining three studies (25%) were on mammals (two on Rodentia and one on Carnivora). Nine studies were performed on captive individuals, whereas three were performed in the wild. Geographically, five of the studies were performed partly or completely in Europe (three in Hungary, one in Germany, one partly in the United Kingdom), five in North America (including one from Barbados in the Caribbean region), one in Africa, one in South America, and one in Australia (Supplementary Table S1).
All effect sizes were from food-extraction experiments, except for one (Preiszner et al., 2017) where the task was the removal of an obstacle from the entrance of a nest box. Task types included: removing a lid from a container (Liker and Bókony, 2009; Sol et al., 2011; Papp et al., 2015; Audet et al., 2016; Preiszner et al., 2017; Chow et al., 2021; Mazza and Guenther, 2021); opening one or more doors on a container (push door, pull door, sliding door, lift door; Papp et al., 2015; Cook et al., 2017; Solaro et al., 2019; Johnson Ulrich et al., 2021; Mazza and Guenther, 2021); pulling a plug or other obstacle from a tube (Papp et al., 2015; Kozlovsky et al., 2017; Mazza and Guenther, 2021); pulling out a drawer (Audet et al., 2016; Johnson Ulrich et al., 2021); ripping a paper covering a container (Papp et al., 2015; Prasher et al., 2019); pulling or pushing a lever (Prasher et al., 2019; Chow et al., 2021). In a few studies, the same task could be solved multiple ways, i.e., the food container having both a lid and a drawer (Audet et al., 2016) or multiple doors (Solaro et al., 2019; Johnson Ulrich et al., 2021).
In the random-effect meta-analysis where we used log-odds ratios as effect sizes, the standard mean difference was 0.39 (95% confidence interval = −0.18 to 0.97; Figure 2). This indicates that in our sample, urban animals tended to be better problem-solvers compared to their rural conspecifics, but this trend was statistically non-significant due to the confidence interval overlapping with 0. We found relatively high heterogeneity in our effect sizes (I2 = 67%, p ≤ 0.01, df = 16, Chi2 = 47.83, Tau2 = 0.994). This resulted in a broad prediction interval (−1.81 to 2.59; Figure 2).
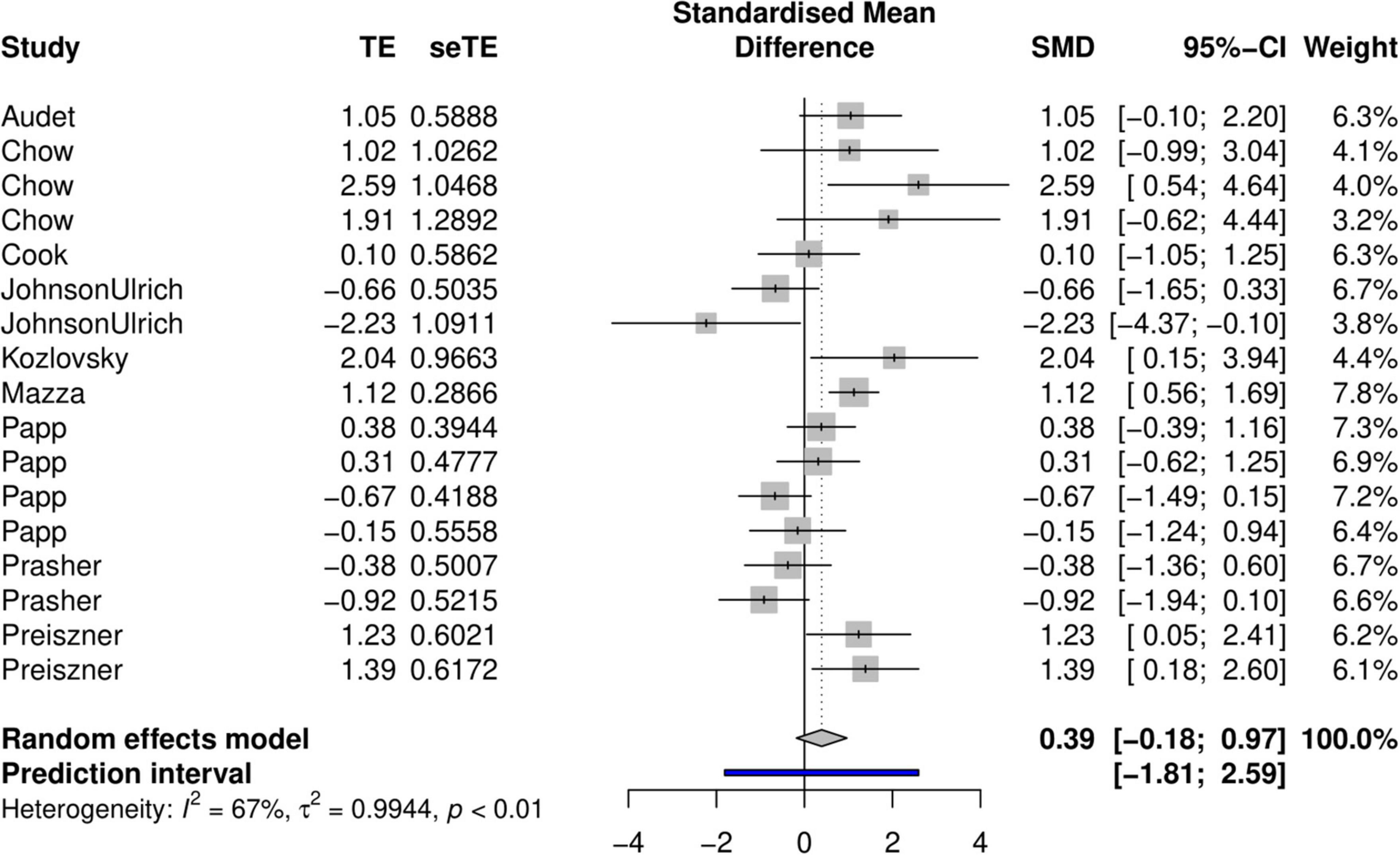
Figure 2. Forest plot showing effect sizes (logarithmic odds ratios indicating the difference in problem-solving success between urban and rural animals) with 95% confidence intervals. Positive effect size values indicate that animals from urban environments were more successful problem-solvers, whereas negative values indicate better problem-solving abilities in rural populations.
In the model with the latency parameters as effect sizes, the standard mean difference was 0.31 (95% confidence interval: −0.02 and 0.64, Figure 3.), which suggests that urban animals tended to solve the problems faster than their non-urban conspecifics, but this trend was also statistically non-significant. Heterogeneity was also large in this sample (I2 = 70%, p ≤ 0.01, df = 19, Chi2 = 64.30, Tau2 = 0.4716), and the prediction interval was also broad (−1.17 to 1.79; Figure 3).
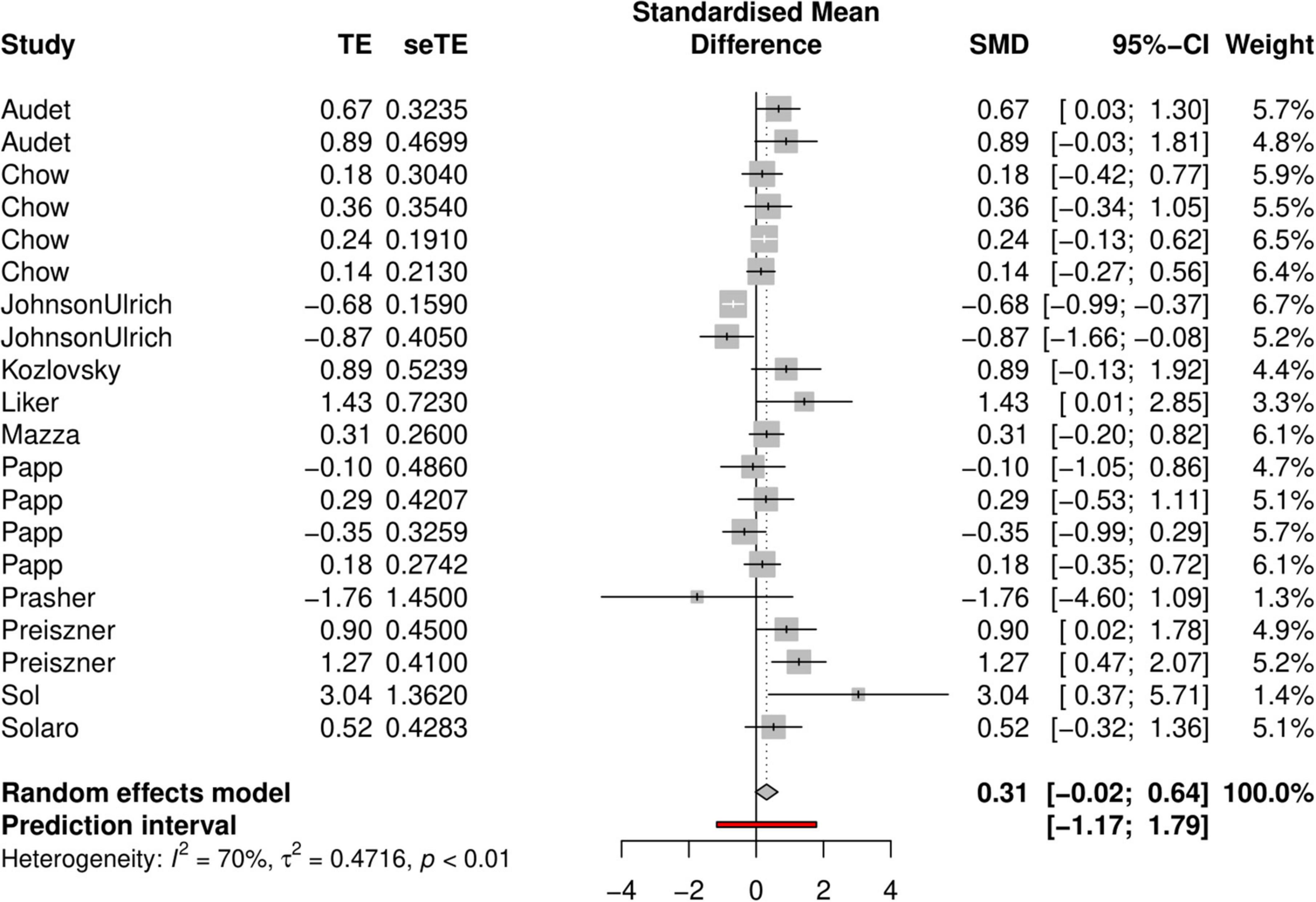
Figure 3. Forest plot showing effect sizes (Z values from Cox models or T values from linear models, indicating the differences in problem-solving latency between urban and rural animals) with 95% confidence intervals. Positive values indicate that animals from urban environments were faster problem-solvers whereas negative values indicate that animals from rural populations solved faster.
The funnel plots were not skewed in either direction, indicating that there was no clear publication bias in either the solving success odds ratios or the latency estimates (Figure 4). However, in both models we found more outliers than expected by random chance (35% of the solving success odds ratios and 20% of the latency estimates, opposed to 5%), which is in line with the high heterogeneity and the broad prediction intervals.
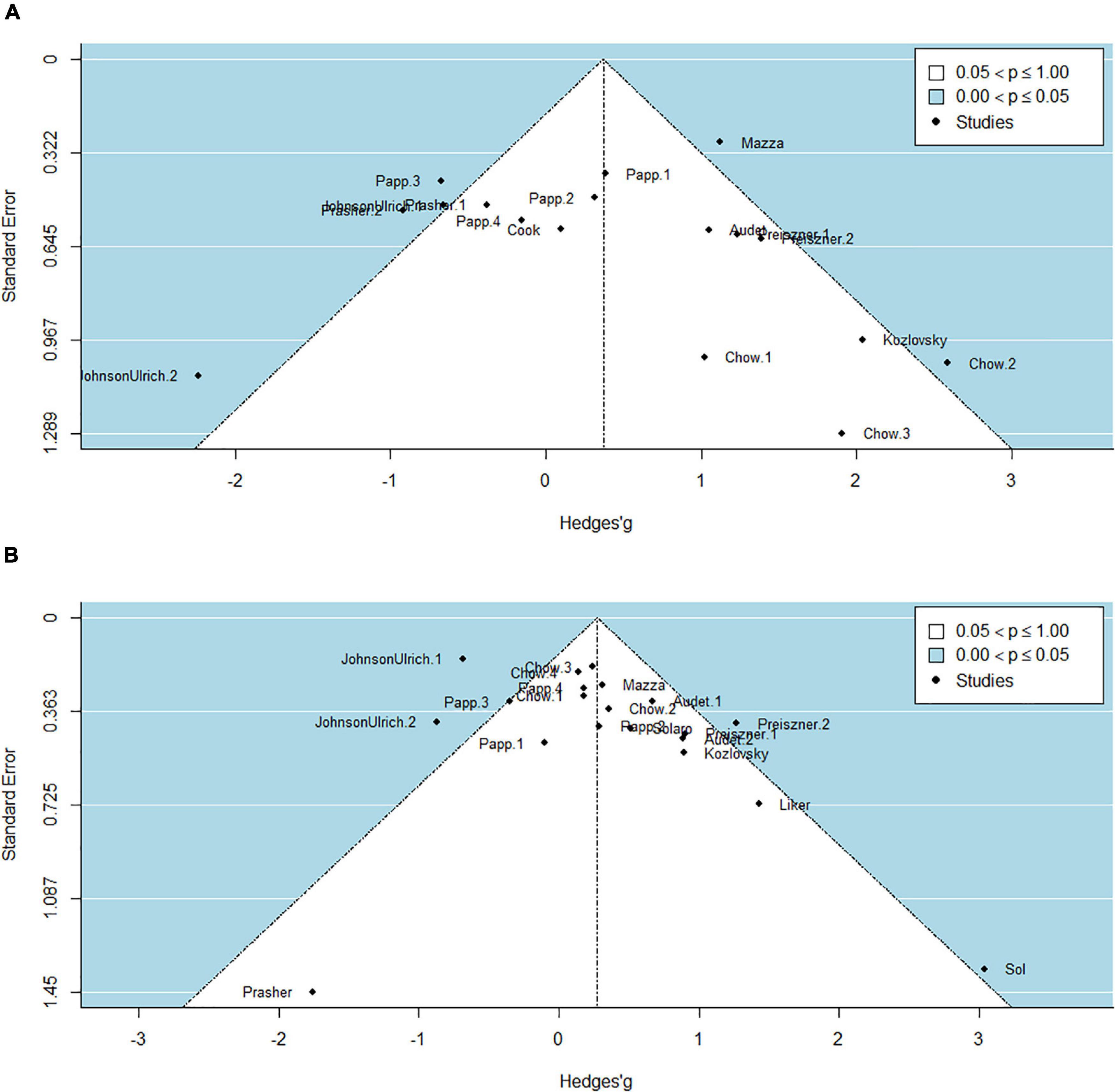
Figure 4. Funnel posts for testing potential publication bias and outliers for solving success odds ratios (A) and solving latency estimates (B).
In a meta-analysis, we found that urban animals tended to be more successful and faster in food-extraction or obstacle-removal tasks compared to their non-urban conspecifics, though these trends were statistically non-significant. This positive mean effect size is in line with our initial prediction, i.e., that urban animals would be better at problem-solving than their non-urban conspecifics. However, we also found high heterogeneity despite the low sample size, with broad prediction intervals from the random-effects models and a high number of outliers, indicating that the pattern is far from general. There are several possible explanations for this high heterogeneity, both biological and statistical. Unfortunately, due to the low sample size, it is not possible to disentangle these effects and their potential interactions by statistical methods.
First, it is possible that the heterogeneity is due to the variation between task types. To reduce the possibility of this option, we limited our sample to tasks where an animal had to manipulate some kind of obstacle (door, lever, plug, drawer, etc.) to access a resource (food, or in one case, nest), as we predicted these tasks would require similar cognitive and motoric skills. We excluded other cognitive tasks such as discriminating between dangerous and non-dangerous humans (Vincze et al., 2015; Vincze et al., 2019), spatial memory (Morand-Ferron et al., 2019; Thompson and Morand-Ferron, 2019), discriminating between safe and unsafe shelters (Batabyal and Thaker, 2019), and learning directions in a maze (Kang et al., 2018) from our analysis, as these require different cognitive and motoric skills, and would have been represented only by one or two effect sizes in our models. Nevertheless, we still ended up with at least six distinct task types: lid-removal, door-opening, drawer-pulling, plug-opening, paper-ripping, and lever-pulling. Unfortunately, we had too few effect sizes per task type to reliably test whether different task types are affected differently by urbanization. Some studies tested multiple problem-solving tasks on the same individuals; some of them found correlation between the solving success in the two tasks (Audet et al., 2016), but others indicate that solving success of an individual animal in one task type does not predict solving success in the other (Preiszner et al., 2017; Prasher et al., 2019).
Second, the heterogeneity may come from the variation between study species, as urbanization can affect different animal species in various ways. For a decade, the studies were exclusively on passerine birds; the only non-passerine bird study we found (Solaro et al., 2019) was as recent as 2019, and all mammal studies were published in 2021, on two rodent species (Chow et al., 2021; Mazza and Guenther, 2021) and one carnivore (Johnson Ulrich et al., 2021). However, even very closely related species responded in vastly different ways to urbanization: urban mountain chickadees (Poecile gambeli) were more successful in a plug-opening task than their forest-dwelling conspecifics (Kozlovsky et al., 2017), whereas urban black-capped chickadees (Poecile atricapillus) were no more successful in a lever-pulling task and less successful in a paper-ripping task than their rural conspecifics (Prasher et al., 2019). It is likely that if urbanization has different effects on the problem-solving ability of different species, it has less to do with the species’ phylogeny and more with its ecological characteristics such as diet, daily activity or social behavior, or the time or extent the species adapted to urban environments.
Third, whether a population was considered “urban” or “rural” depended very much on the study. Some studies compared residential areas to farmlands (e.g., Papp et al., 2015; Solaro et al., 2019), others compared urban parks to forests (e.g., Preiszner et al., 2017; Prasher et al., 2019), or the outskirts of a developing town adjacent to a national park to a savanna at the center of a national park (e.g., Johnson Ulrich et al., 2021). It is arguable whether the outskirts of a developing town can be considered as more urbanized than a farmland. The relationship between anthropogenic disturbance and behavior is not necessarily linear (e.g., Fernández-Juricic et al., 2003), which may explain some of the variation in the results.
Fourth, whether the experiment was performed on animals in their natural habitat or in captivity could have also affected the results. On the one hand, different animals could respond differently to captivity (e.g., have different tolerance toward capturing stress), which may affect their performance in the problem-solving tasks; on the other hand, performing the study on natural populations, where the animal voluntarily approaches the problem-solving device, can lead to sampling bias toward the more explorative or neophilic individuals. In our sample, only three studies were performed in the animals’ natural habitat (Preiszner et al., 2017; Chow et al., 2021; Johnson Ulrich et al., 2021), whereas the rest were performed on captive animals.
Fifth, some of the heterogeneity can potentially be explained by geographical variation. As the bulk of our effect sizes come from Europe and North America, a hemisphere bias (Martin et al., 2012) is certainly present in these studies. As urbanization is a global phenomenon, but different continents and countries have vastly different urbanization history, it would be important to include more studies from other parts of the world.
Finally, it is possible that the higher-than-expected heterogeneity is due to a special case of publication bias: though we did not find publication bias in either direction (i.e., the funnel plot was relatively symmetrical), we may have a two-way publication bias, i.e., studies in which urban animals were either significantly more successful or significantly less successful than rural animals were more likely to get published than non-significant results. Notably, in the studies where urbanization, by itself, had no significant effect on problem-solving abilities also reported a statistically significant interaction between urbanization and another explanatory variable, such as body mass (Papp et al., 2015), dominance rank (Prasher et al., 2019), or disturbance in captivity (Cook et al., 2017). Therefore, it is possible that studies where urbanization had neither significant effect on problem solving abilities by itself nor a significant interaction with another explanatory variable remained unpublished.
Despite the relevance of the topic, we only managed to find a handful of studies investigating whether problem solving abilities are affected by urbanization. We need more studies on this topic to test whether or not the positive trend we found is generalizable, and to investigate the underlying mechanisms of the heterogeneity. We recommend to extend the studies on additional taxa, and include more non-passerine birds (such as raptors or parrots), mammals and even reptiles. There are two studies indicating that that tested the effects of urbanization on some sort of cognitive abilities in lizards, specifically learning the difference between a “safe” and “unsafe” shelter. The two studies came to different conclusions, one finding that urban lizards were more successful in this task (Batabyal and Thaker, 2019), the other finding no significant difference between the populations (Kang et al., 2018). We also suggest more studies performed in Asia and the Southern hemisphere, to test how globally generalizable the patterns are. Finally, beside problem-solving tasks, we also recommend additional studies on other cognitive tasks such as spatial memory and individual recognition of humans, to test how general these patterns are across behavioral contexts.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
EV conceived the study idea, collected literature, collected effect sizes, and wrote the first draft of the manuscript. BK screened the abstracts and run the meta-analytical models. Both authors wrote the final version of the manuscript.
This study was supported by the PD-134985 grant from the National Research, Development and Innovation Office of Hungary and the MSCA Seal of Excellence 2021-01102 grant from Vinnova, the Swedish Innovation Agency, both awarded to EV. Article processing fee was funded by Lund University’s APC Fund.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Jean-Nicolas Audet for inviting us to this special issue of Frontiers in Ecology and Evolution, and for providing additional information on his earlier study. We also thank Veronika Bókony, Anja Guenther, Lily Johnson-Ulrich, Pizza Ka Yee Chow, Julie Morand-Ferron, Sanjay Prasher, and Claudina Solaro for providing additional information on their respective studies, and Nikolay Oksolkov and Lars Råberg for advice in interpreting our results.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.834436/full#supplementary-material
Audet, J.-N., Ducatez, S., and Lefebvre, L. (2016). The town bird and the country bird: problem solving and immunocompetence vary with urbanization. Behav. Ecol. 27, 637–644. doi: 10.1093/beheco/arv201
Batabyal, A., and Thaker, M. (2019). Lizards from suburban areas learn faster to stay safe. Biol. Lett. 15:20190009. doi: 10.1098/rsbl.2019.0009
Bateman, P. W., and Fleming, P. A. (2012). Big city life: carnivores in urban environments. J. Zool. 287, 1–23. doi: 10.1111/j.1469-7998.2011.00887.x
Carrete, M., and Tella, J. L. (2011). Inter-individual variability in fear of humans and relative brain size of the species are related to contemporary urban invasion in birds. PLoS One 6:e18859. doi: 10.1371/journal.pone.0018859
Chow, P. K. Y., Clayton, N. S., Steele, M. A., and Ashton, B. J. (2021). Cognitive performance of wild Eastern gray squirrels (Sciurus carolinensis) in rural and urban, native, and non-native environments. Front. Ecol. Evol. 9:615899. doi: 10.3389/fevo.2021.615899
Cook, M., Weaver, M., Hutton, P., and McGraw, K. (2017). The effects of urbanization and human disturbance on problem solving in juvenile house finches (Haemorhous mexicanus). Behav. Ecol. Sociobiol. 71:85.
Evans, K. L., Chamberlain, D. E., Hatchwell, B. J., Gregory, R. D., and Gaston, K. J. (2011). What makes an urban bird? Glob. Change Biol. 17, 32–44. doi: 10.1111/j.1365-2486.2010.02247.x
Fernández-Juricic, E., Sallent, A., Sanz, R., and Rodriguez-Prieto, I. (2003). Testing the risk-disturbance hypothesis in a fragmented landscape: nonlinear responses of House Sparrows to humans. Condor 105, 316–326. doi: 10.1093/condor/105.2.316
Goumas, M., Lee, V. E., Kelley, L. A., and Thornton, A. (2020). The role of animal cognition in human-wildlife interactions. Front. Psychol. 11:589978. doi: 10.3389/fpsyg.2020.589978
Griffin, A. S., Netto, K., and Peneaux, C. (2017). Neophilia, innovation and learning in an urbanized world: a critical evaluation of mixed findings. Curr. Opin. Behav. Sci. 16, 15–22. doi: 10.1016/j.cobeha.2017.01.004
Grimm, N. B., Faeth, S. H., Golubiewski, N. E., Charles, L., Wu, J., Bai, X., et al. (2008). Global change and the ecology of cities. Science 319, 756–760. doi: 10.1126/science.1150195
Grunst, A. S., Grunst, M. L., Pinxten, R., and Eens, M. (2020). Sources of individual variation in problem-solving performance in urban great tits (Parus major): exploring effects of metal pollution, urban disturbance and personality. Sci. Total Environ. 749:141436. doi: 10.1016/j.scitotenv.2020.141436
Harrer, M., Cuijpers, P., Furukawa, T. A., and Ebert, D. D. (2021). Doing Meta-Analysis with R: a Hands-on Guide. Abingdon: Taylor and Francis.
Hedges, L. V., and Vevea, J. L. (1998). Fixed- and random-effects models in meta-analysis. Psychol. Methods 3, 486–504.
Jha, N. A., and Kumar, V. (2017). Effect of no-night light environment on behaviour, learning performance and personality in zebra finches. Anim. Behav. 132, 29–47. doi: 10.1016/j.anbehav.2017.07.017
Johnson Ulrich, L., Yirga, G., Strong, R. L., and Holekamp, K. E. (2021). The effect of urbanization on innovation in spotted hyenas. Anim. Cogn. 24, 1027–1038. doi: 10.1007/s10071-021-01494-4
Kang, F., Goulet, C. T., and Chapple, D. G. (2018). The impact of urbanization on learning ability in an invasive lizard. Biol. J. Linn. Soc. 123, 55–62. doi: 10.1093/biolinnean/blx131
Kozlovsky, D. Y., Weissgerber, E. A., and Pravosudov, V. V. (2017). What makes specialized food-caching mountain chickadees successful city slickers? Proc. R. Soc. B Biol. Sci. 264:20162613. doi: 10.1098/rspb.2016.2613
Lee, V. E., and Thornton, A. (2021). Animal cognition in an urbanised world. Front. Ecol. Evol. 9:633947. doi: 10.3389/fevo.2021.633947
Liker, A., and Bókony, V. (2009). Larger groups are more successful in innovative problem solving in house sparrows. Proc. Natl. Acad. Sci. U.S.A. 106, 7893–7898. doi: 10.1073/pnas.0900042106
Maklakov, A. A., Immler, S., Gonzalez-Voyer, A., Rönn, J., and Kolm, N. (2011). Brains and the city: big-brained passerine birds succeed in urban environments. Biol. Lett. 7, 730–732. doi: 10.1098/rsbl.2011.0341
Martin, L. J., Blossey, B., and Ellis, E. (2012). Mapping where ecologists work: biases in the global distribution of terrestrial ecological observations. Front. Ecol. Environ. 10, 195–201. doi: 10.1890/110154
Mazza, V., and Guenther, A. (2021). City mice and country mice: innovative problem solving in rural and urban noncommensal rodents. Anim. Behav. 172, 197–210. doi: 10.1016/j.anbehav.2020.12.007
Moher, D., Liberati, A., Tetzlaff, J., and Altman, D. G., and PRISMA Group (2009). Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. Ann. Intern Med. 151, 264–269. doi: 10.1371/journal.pmed.1000097
Morand-Ferron, J., Cole, E. F., and Quinn, J. L. (2016). Studying the evolutionary ecology of cognition in the wild: a review of practical and conceptual challenges. Biol. Rev. 91, 367–389. doi: 10.1111/brv.12174
Morand-Ferron, J., Hermer, E., Jones, T. B., and Thompson, M. J. (2019). Environmental variability, the value of information, and learning in winter residents. Anim. Behav. 147, 137–145. doi: 10.1016/j.anbehav.2018.09.008
Nakagawa, S., and Cuthill, I. C. (2007). Effect size, confidence interval and statistical significance: a practical guide for biologists. Biol. Rev. 82, 591–605. doi: 10.1111/j.1469-185X.2007.00027.x
Papp, S., Vincze, E., Preiszner, B., Liker, A., and Bókony, V. (2015). A comparison of problem-solving success between urban and rural house sparrows. Behav. Ecol. Sociobiol. 69, 471–480. doi: 10.1007/s00265-014-1859-8
Partecke, J., Schwabl, I., and Gwinner, E. (2006). Stress and the city: urbanization and its effects on the stress physiology in European blackbirds. Ecology 87, 1945–1952. doi: 10.1890/0012-9658(2006)87[1945:satcua]2.0.co;2
Prasher, S., Evans, J. C., Thompson, M. J., and Morand-Ferron, J. (2019). Characterizing innovators: ecological and individual predictors of problem-solving performance. PLoS One 14:e0217464. doi: 10.1371/journal.pone.0217464
Preiszner, B., Papp, S., Pipoly, I., Seress, G., Vincze, E., Liker, A., et al. (2017). Problem-solving performance and reproductive success of great tits in urban and forest habitats. Anim. Cogn. 20, 53–63. doi: 10.1007/s10071-016-1008-z
Racine, J. S. (2012). Rstudio: A platform-independent IDE for R and sweave. J. Appl. Econ. 27, 167–172. doi: 10.1002/jae.1278
Sayol, F., Sol, D., and Pigot, A. L. (2020). Brain size and life history interact to predict urban tolerance in birds. Front. Ecol. Evol. 8:58. doi: 10.3389/fevo.2020.00058
Seress, G., and Liker, A. (2015). Habitat urbanization and its effects on birds. Acta Zool. Academ. Sci. Hung. 61, 373–408. doi: 10.17109/AZH.61.4.373.2015
Snell-Rood, E. C., and Wick, N. (2013). Anthropogenic environments exert variable selection on cranial capacity in mammals. Proc. R. Soc. B Biol. Sci. 280:20131384. doi: 10.1098/rspb.2013.1384
Sol, D., Griffin, A. S., Bartomeus, I., and Boyce, H. (2011). Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS One 6:e19535. doi: 10.1371/journal.pone.0019535
Sol, D., Lapiedra, O., and González-Lagos, C. (2013). Behavioural adjustments for a life in the city. Anim. Behav. 85, 1101–1112. doi: 10.1016/j.anbehav.2013.01.023
Sol, D., Timmermans, S., and Lefebvre, L. (2002). Behavioural flexibility and invasion success in birds. Anim. Behav. 63, 495–502. doi: 10.1006/anbe.2001.1953
Solaro, C., Sarasola, J. H., Aves, D., de Ciencias, F., Nacional, U., and La Pampa, D. (2019). Urban living predicts behavioural response in a neotropical raptor. Behav Processes 169:103995. doi: 10.1016/j.beproc.2019.103995
Thompson, M. J., and Morand-Ferron, J. (2019). Food caching in city birds: urbanization and exploration do not predict spatial memory in scatter hoarders. Anim. Cogn. 22, 743–756. doi: 10.1007/s10071-019-01271-4
Vincze, E., Papp, S., Preiszner, B., Seress, G., Liker, A., and Bókony, V. (2015). Does urbanization facilitate individual recognition of humans by house sparrows? Anim. Cogn. 18, 291–298. doi: 10.1007/s10071-014-0799-z
Vincze, E., Pipoly, I., Seress, G., Preiszner, B., Papp, S., Németh, B., et al. (2019). Great tits take greater risk toward humans and sparrowhawks in urban habitats than in forests. Ethology 125, 686–701. doi: 10.1111/eth.12922
Vrbanec, L., Matijevi, V., and Guenther, A. (2021). Enhanced problem-solving ability as an adaptation to urban environments in house mice. Proc. R. Soc. B Biol. Sci. 288:20202504. doi: 10.1098/rspb.2020.2504
Keywords: urbanized environment, animal cognition, problem-solving abilities, interspecific comparison, meta-analysis
Citation: Vincze E and Kovács B (2022) Urbanization’s Effects on Problem Solving Abilities: A Meta-Analysis. Front. Ecol. Evol. 10:834436. doi: 10.3389/fevo.2022.834436
Received: 13 December 2021; Accepted: 16 February 2022;
Published: 17 March 2022.
Edited by:
Jayna Lynn DeVore, The University of Sydney, AustraliaReviewed by:
Daniela Campobello, University of Palermo, ItalyCopyright © 2022 Vincze and Kovács. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ernő Vincze, ZXJuby52aW5jemVAYmlvbC5sdS5zZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.