 Yahkat Barshep
Yahkat Barshep Adewale G. Awoyemi
Adewale G. Awoyemi Jacinta Abalaka
Jacinta Abalaka Ulf Ottosson
Ulf Ottosson- 1A.P. Leventis Ornithological Research Institute, Jos, Nigeria
- 2Forest Centre, International Institute of Tropical Agriculture, Ibadan, Nigeria
The Bergmann’s rule predicts that in endotherms, body sizes will differ with respect to thermal gradients. Larger bodied individuals will inhabit colder environments while in warmer environments, individuals will be smaller-bodied. This hypothesis has been proved and disproved many times due to inconsistencies in body size differences along latitudinal gradients. We tested this hypothesis in 30 Afrotropical resident bird species inhabiting two vegetation types at different latitudes (southern guinea forests and northern savanna) and at different altitudes in Nigeria, West Africa. Using principal component analyses of body mass and wing length, the first principal component, the component of size, indicated that individuals in montane areas were larger than lowland populations in southern guinea forests. However, in the northern guinea savanna, there was no significant difference in body sizes between lowland and montane populations. General linear models show that body size increases as temperature decreases. In species found in both southern guinea forests and northern savanna (i.e., African Thrush Turdus pelios and Snowy-crowned Robin Chat Cossypha niveicapilla), variations in body sizes were significantly dependent on sites. Our study indicates that other macro-scale factors such as vegetation and rainfall patterns might modulate conformity to Bergmann’s rule in Afrotropical environments.
Introduction
Bergmann’s rule, an eco-evolutionary generalization predicting that endothermic animals will be larger in cold climates and smaller in warm climates, gives an explanation for the evolution of body size variation among similar groups of organisms (Bergmann, 1847; Salewski and Watt, 2017), which has been deployed to understand various aspects of ecology, such as predator-prey relationships (e.g., Mcnab, 1971). Although this hypothesis has been tested across different taxa, such as mammals (Clauss et al., 2013), birds (Ashton, 2002), and insects (Scriven et al., 2016), there is still an ongoing debate on its suitability for intra- and inter-specific studies (see Shelomi, 2012; Clauss et al., 2013). Simultaneous investigation of intra- and inter-specific variations in body size will enhance our understanding of the generalization of the Bergmann’s rule. This is particularly important if such studies are conducted on little known species and regions such as Afrotropical regions.
The contrasting results obtained from the investigation of Bergmann’s hypothesis suggest influence of other confounding factors, such as microclimates along latitudinal and altitudinal gradients (e.g., Shelomi, 2012; Bhusal et al., 2019). With the global climate change affecting all life forms on earth (Crozier et al., 2008; Şekercioĝlu et al., 2012), understanding species adaptation is important from an applied perspective. Species would need to adjust their physiology and behavior in order to cope with changing climatic conditions (Tieleman and Williams, 2000). Such studies are seldom reported from tropical areas, particularly Africa, which holds huge biodiversity, but projected to be adversely affected by global warming in the nearest future (Midgley and Bond, 2015; Sintayehu, 2018).
Due to their relative ease of trapping, birds are an important model for physiological studies. Here, we tested the Bergmann’s rule within and across 30 avian species along vegetation and altitudinal differences in Nigeria, West Africa. While several studies have supported the Bergmann’s rule, demonstrating that average body size within a species tends to increase toward cooler and higher latitudes (Ashton, 2002), we chose to test this rule along an altitudinal gradient because the north-south temperature gradient at the same elevation in Nigeria is small (an average difference of 7°C) and a previous study has already demonstrated a lack of relationship between latitude and body size in Nigeria (Nwaogu et al., 2018). We hypothesized a larger avian body size at high altitudes compared to lower altitudes and that the magnitude of size difference between lowland and montane populations will differ with respect to vegetation type (savanna vs. rainforest) and species.
Materials and Methods
Study Sites
The data analyzed in this study were collected as part of the regular Bird Ringing activities of the A.P. Leventis Ornithological Research Institute (APLORI) located in northern Nigeria (Mwansat et al., 2011; Cresswell, 2018), on the Jos Plateau (9.5196°N, 8.5897°E; 1280 m asl) and at Yankari Game Reserve (9.7567°N, 10.5094°E; 330 m asl). In southern Nigeria, birds were trapped on the Obudu Plateau (6.3858°N, 9.3745°E; 1400 m asl) and at the lowland forests at International Institute of Tropical Agriculture (IITA; 7.2985°N, 3.5333°E) and Emerald Forest Reserve (EFR: 7.1780°N, 4.0806°E) both at 230 m asl (Figure 1). Because of close proximity (51 km apart), similar habitats and elevation, data from IITA and EFR were merged and analyzed as a single location (IITA). Data from Jos Plateau was from the period 200 to 2018, Yankari data was from 2011 to 2017, Obudu data was from the period 2005 to 2017, and the data from both IITA and Emerald was from the period 2017 to 2020.
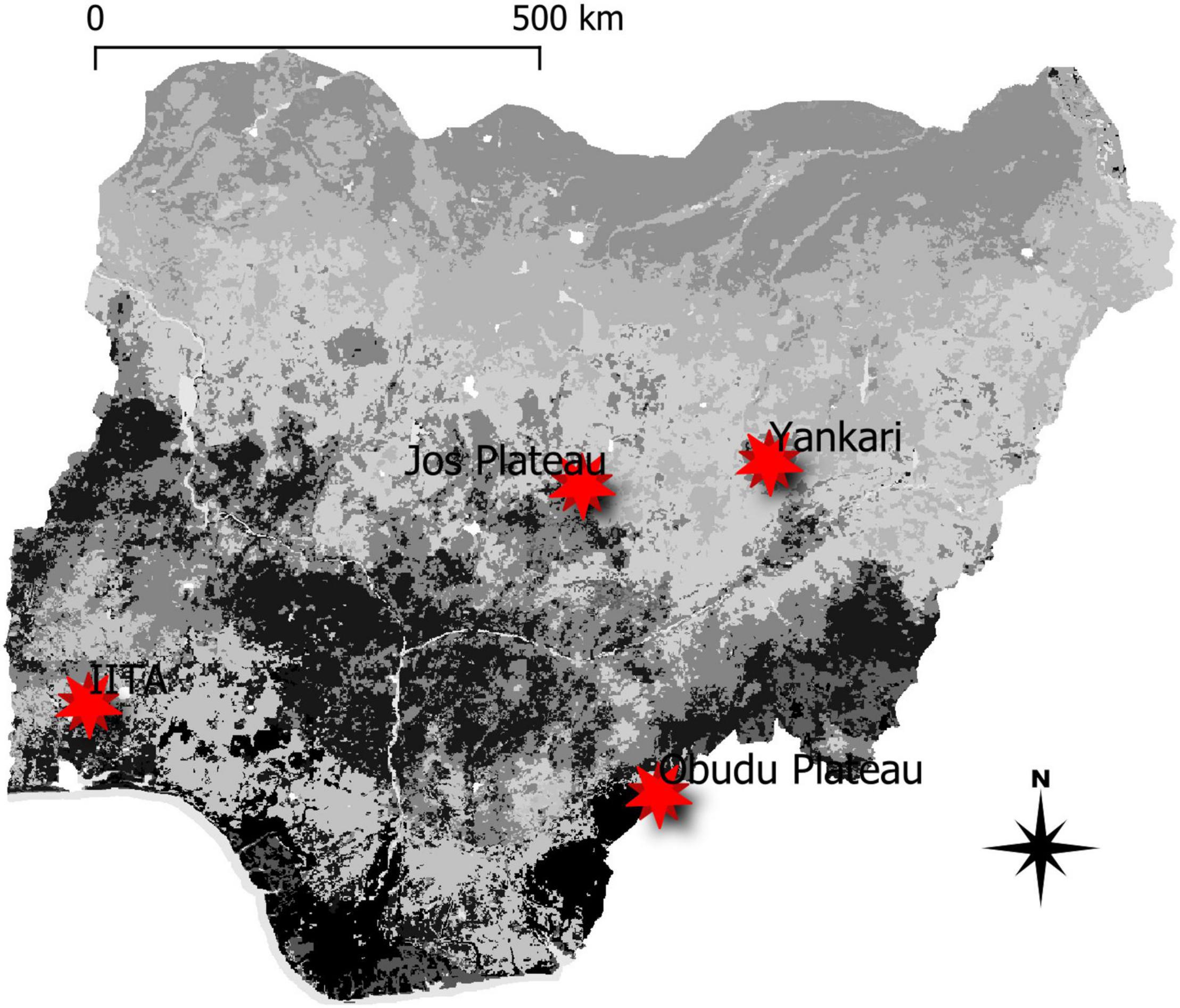
Figure 1. Map of Nigeria with study sites indicated as stars.
The northern Nigeria sites are within the northern Guinea Savannah vegetation zone characterized by a mixture of trees and grass and receives annual rainfall of between 600 and 1,000 mm per mostly concentrated within 5–7 months (Ezealor, 2001; Omotoriogun et al., 2011; Braimoh et al., 2018). The southern sites are within the Guinea Forest vegetation zone characterized by dense evergreen forest of tall trees with thick undergrowth and receives annual rainfall of between 1,500 and 2,000 mm per annum with about 6–8 months of rainfall (Ezealor, 2001; Adeyanju et al., 2014; Awoyemi et al., 2020).
Data Collection and Analyses
Morphometric data of non-migratory bird species at low and high elevations at the northern and southern sites were used to test the Bergmann’s rule. At all sites, birds were trapped with mist nets of various lengths at these different sites, tagged with numbered metal rings, aged, sexed, weighed (g), and wing length (mm) measured. Only adult individuals of non-migratory or nomadic species were used for this study. Monthly precipitation and temperature data were obtained from the Bioclime website,1 measured at 1 km2 resolution, and extracted using the open-source program QGIS. These weather variables were used as covariates in further analyses.
For species with at least 10 records at both lowland and montane areas, we used the R package (R Core Team, 2016) to perform a principal component analysis (PCA) (Cozzolino et al., 2019) based on a correlation matrix using body mass and wing length. Both wing length and body mass are an indication of body size and have been consistently recorded at all ringing data but rather than analyze them separately, by using PCA, we can combine these two response variables into a single component to produce an index of body size. This procedure produces principal component scores for each individual; negative and low vales of scores indicate individuals with shorter wings and lighter body mass (i.e., smaller body sized individuals) while positive and higher values indicate larger body sized individuals. Because principal component scores had a Gaussian distribution, we used a general linear model to investigate if body size was affected by altitude (m), average monthly temperature (°C), total monthly rainfall (mm), using latitude as weighted variable. We used latitude as a weighting factor rather than an explanatory variable because a previous study testing the Bergmann’s Rule along latitudinal gradient indicated that latitude had a slight but non-statistically significant effect on body size of birds in Nigeria (Nwaogu et al., 2018). By doing this, we also avoided over-parameterization of models while still adjusting for possible, if only slight, effect of latitude. To also avoided over-parameterization of models, the northern and southern sites were analyzed separately. Because temperature and rainfall were correlated, we analyzed their effect on body size in two separate models in order to avoid effects multicollinearity.
Results
We analyzed data of six species in the southern Nigerian sites and 24 species in the northern Nigerian sites (Supplementary Table 1). The results of the principal component showed high and positive loadings on wing length and body mass (0.71 and 0.71, respectively) while PC2 had a high and negative loading on wing length (–0.60). Based on this result of the principal component analyses, Principal component 1 (PC1) represents the component of body size, accounted for 97.1% variability in the data while PC2 represents the component of wing length which made up 2.9% variability in the data.
A plot of PC1 against PC2 and the general linear model with site as predictor indicated that for the southern zone, montane populations were on average significantly heavier than lowland populations, but with lowland species having a comparable longer wing length (Figure 2 and Table 1). Of the six species analyzed, only the Olive Sunbird Cyanomitra olivacea had a higher body size in the lowland population (Supplementary Table 1). However, since PCA 2 is the component of wing length and since lowland species have more positive scores along PCA 2 axis (Figure 2), it means that even though they are smaller than their montane counterparts, their wing length are comparatively longer. The GLM indicated that temperature was a significant predictor of body size, with body size and temperature having a negative relationship, The three-way interaction of location, species, and temperature was significant, indicating that differences in size between species was due to temperature variations between location (Table 1, Model a). Rainfall was not a significant predictor of body size and the three-way interaction between location, species, and rainfall was not significant either (Table 1, Model b), therefore, rainfall cannot explain the differences in body size between lowland and montane sites in the southern zone.
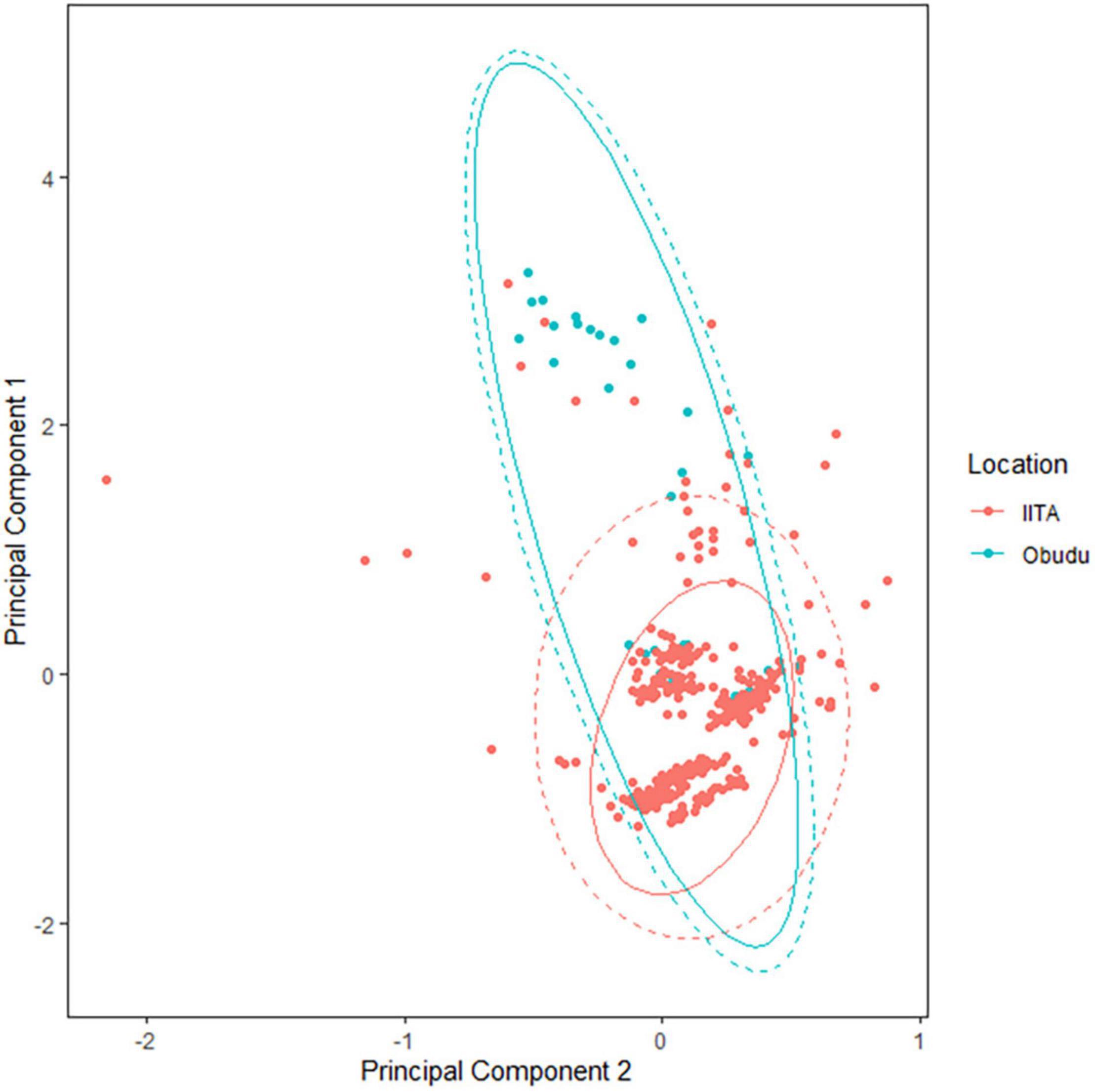
Figure 2. Plots of principal component 1 vs. principal component 2 of lowland and montane species of southern guinea forest of Nigeria.
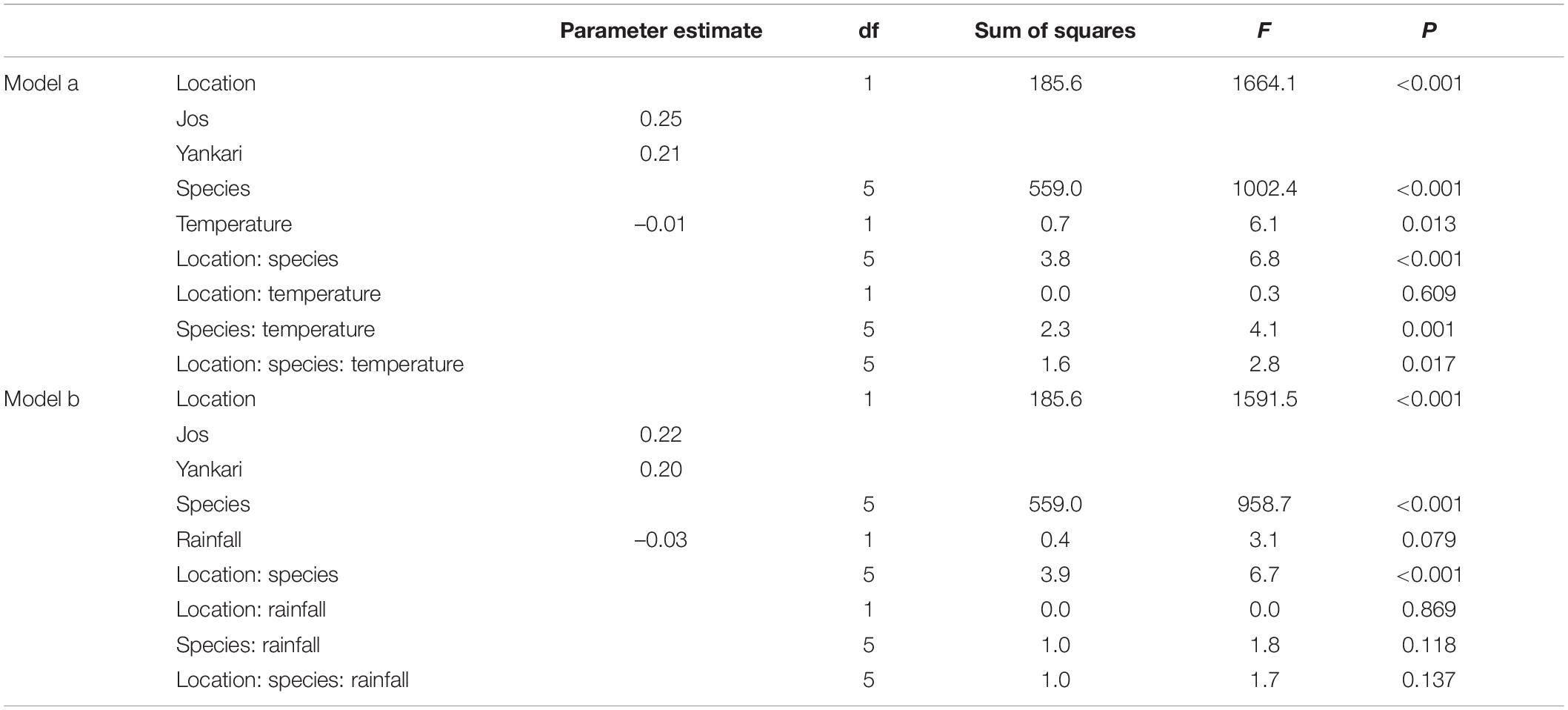
Table 1. General linear model investigating the relationship between body size (PC1) with sites (lowland vs. montane), species, temperature (model a), and rainfall (model b) in southern Nigeria.
In northern sites, a plot of PC1 against PC2 showed a high degree of overlap in body sizes of lowland and montane populations (Figure 3), though montane populations were statistically heavier (Table 2). Of the 24 species considered in the study, populations of 17 species were heavier in montane areas while populations of seven species were heavier in lowland areas (Supplementary Table 1). A general linear model showed that both temperature and rainfall were significant predictors of body size (Table 2, Models a and b) with temperature showing a negative effect on body size. The three-way interaction of location, species, and temperature was significant indicating that temperature was responsible for the difference in body size of species at the different elevations. Although there was a significant effect of rainfall on the body sizes of species but the three-way interaction of location, species, and rainfall was not significant (Table 2, Model b). This is supported by weather data which shows that temperatures were lower at higher altitudes compared to lower (Figure 4A) latitudes whereas rainfall pattern did not follow and altitudinal trend (Figure 4B).
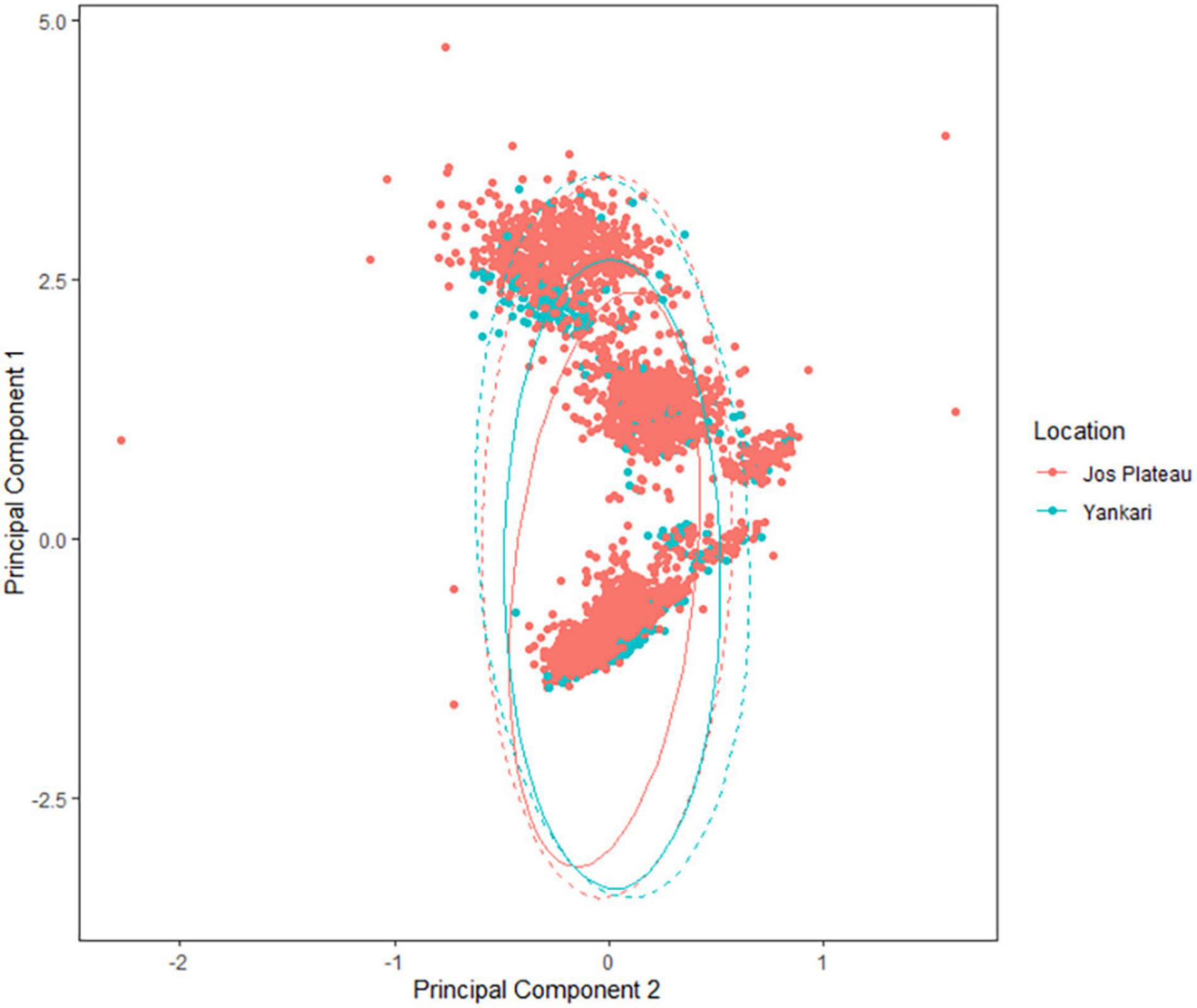
Figure 3. Plots of principal component 1 vs. principal component 2 of lowland and montane species of northern savanna region of Nigeria.
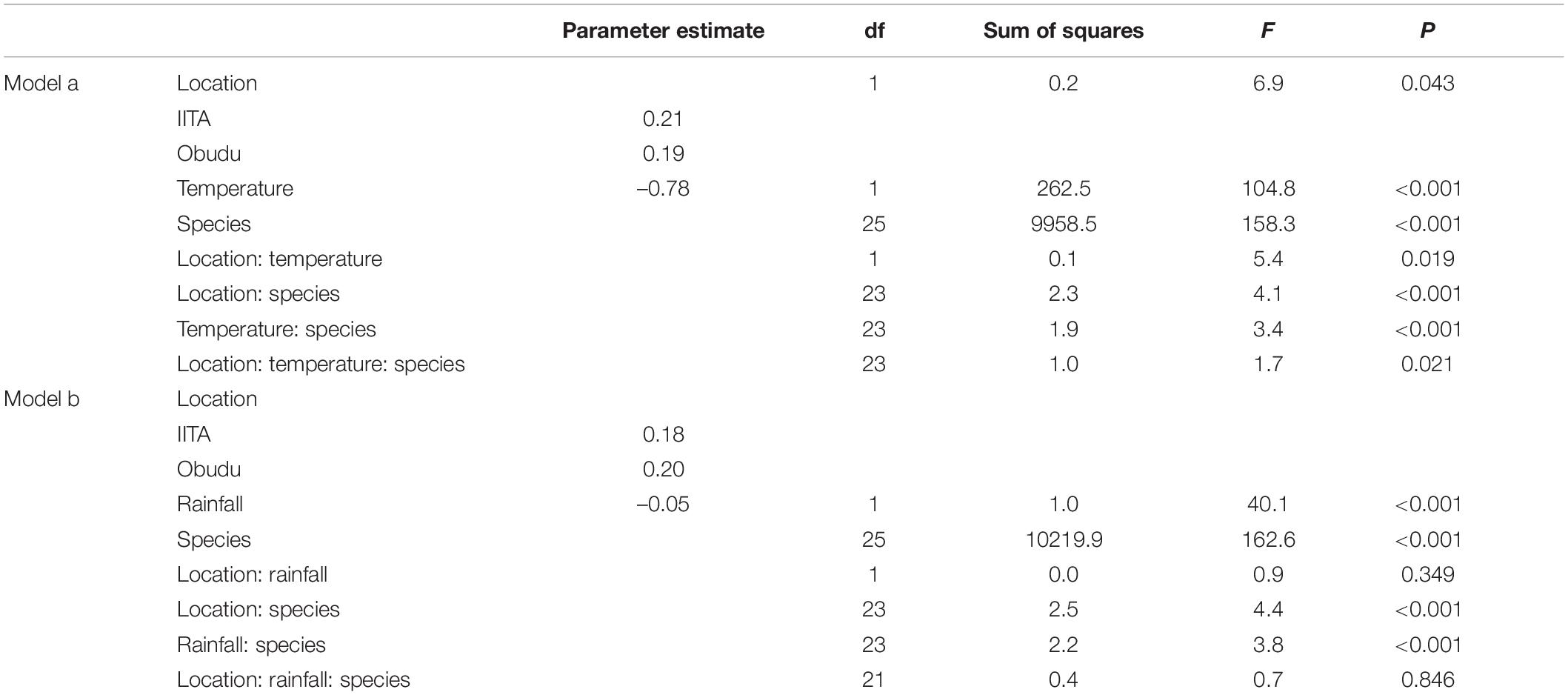
Table 2. General Linear Model investigating the relationship between body size (PC1) with sites (lowland vs. montane), species, temperature (model a), and rainfall (model b) in northern Nigeria.
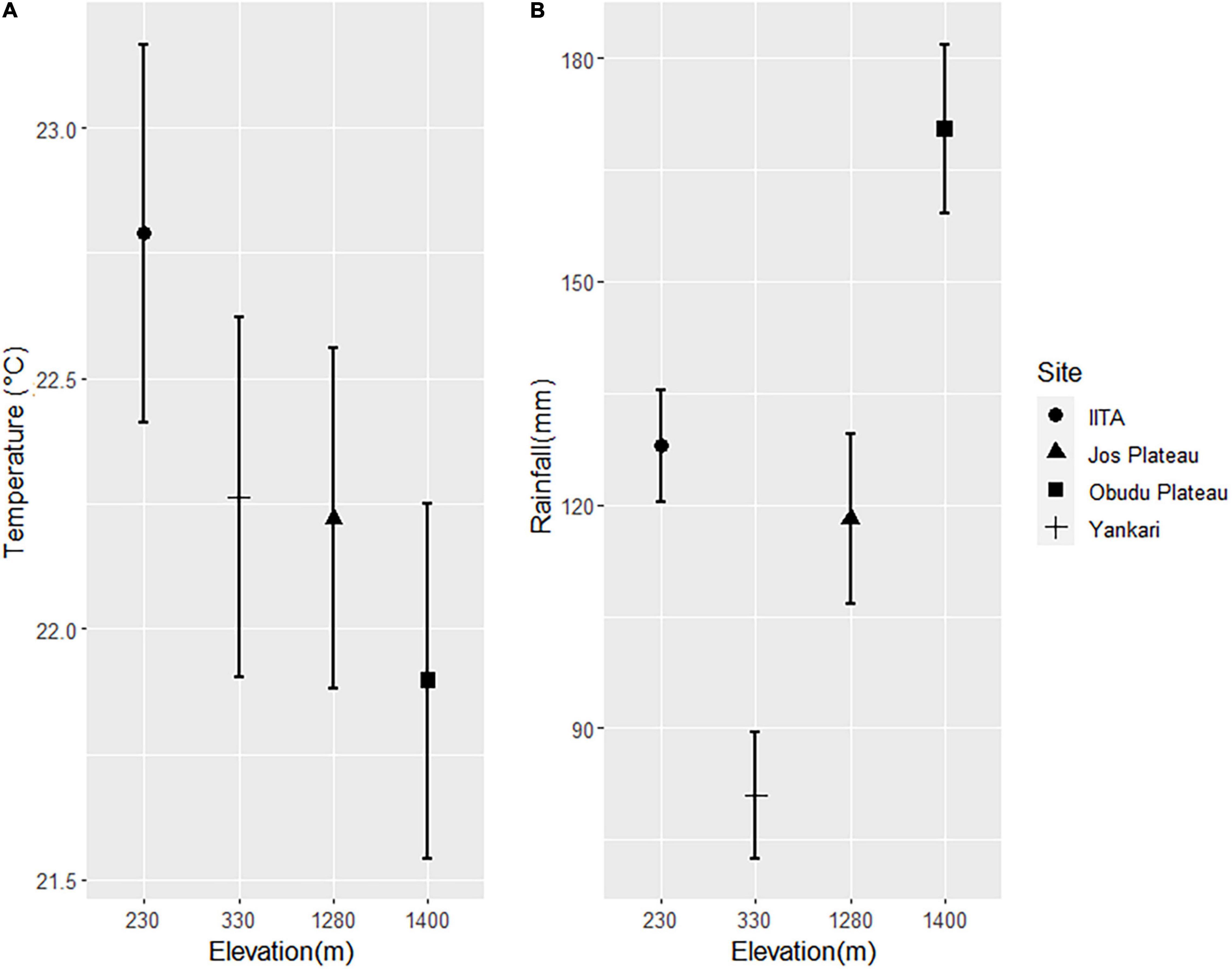
Figure 4. Mean temperature (A) and rainfall (B) ranges across elevation gradients.
Discussion
The findings of this study enhance our understanding of the validity of the Bergmann’s rule, particularly from the understudied afrotropical region. In addition to determining the applicability of this rule among new afrotropical bird species (c.f. Nwaogu et al., 2018), our findings shed more light on the importance of microclimates while testing the Bergmann’s rule along altitudinal gradients. To our understanding, this is the first study to simultaneously consider the interplay of these factors in the afrotropical context.
Although several studies have proven the effects of latitude on body size (see Olson et al., 2009; Shelomi, 2012; Clauss et al., 2013), we argue that these effects are dependent on habitat types. In the present study, montane populations were significantly heavier than lowland populations in the Guinea forest region. However, in the savanna region, there was extensive overlap in body sizes between montane and lowland populations. This pattern suggests the influence of other microclimates and macroclimate factors.
For the species that were found in both northern and southern zones, (African Thrush Turdus pelios and Snowy-crowned Robin Chat Cossypha niveicapilla), we used a t-test to also do a between-elevations comparison of body sizes across ecozones, i.e., body sizes of montane populations were compared between the north and southern zones and the same for lowland populations. Indices of body sizes indicated that populations in the southern zone were larger than the northern zone; body size on the Obudu Plateau were significantly larger than birds on the Jos Plateau (difference = 1.29, CI = 0.06–1.91, p < 0.001, Figure 4) and birds from IITA were also significantly larger than those of Yankari (difference = 0.36, CI = 0.12–0.59, p < 0.001; Figure 5).
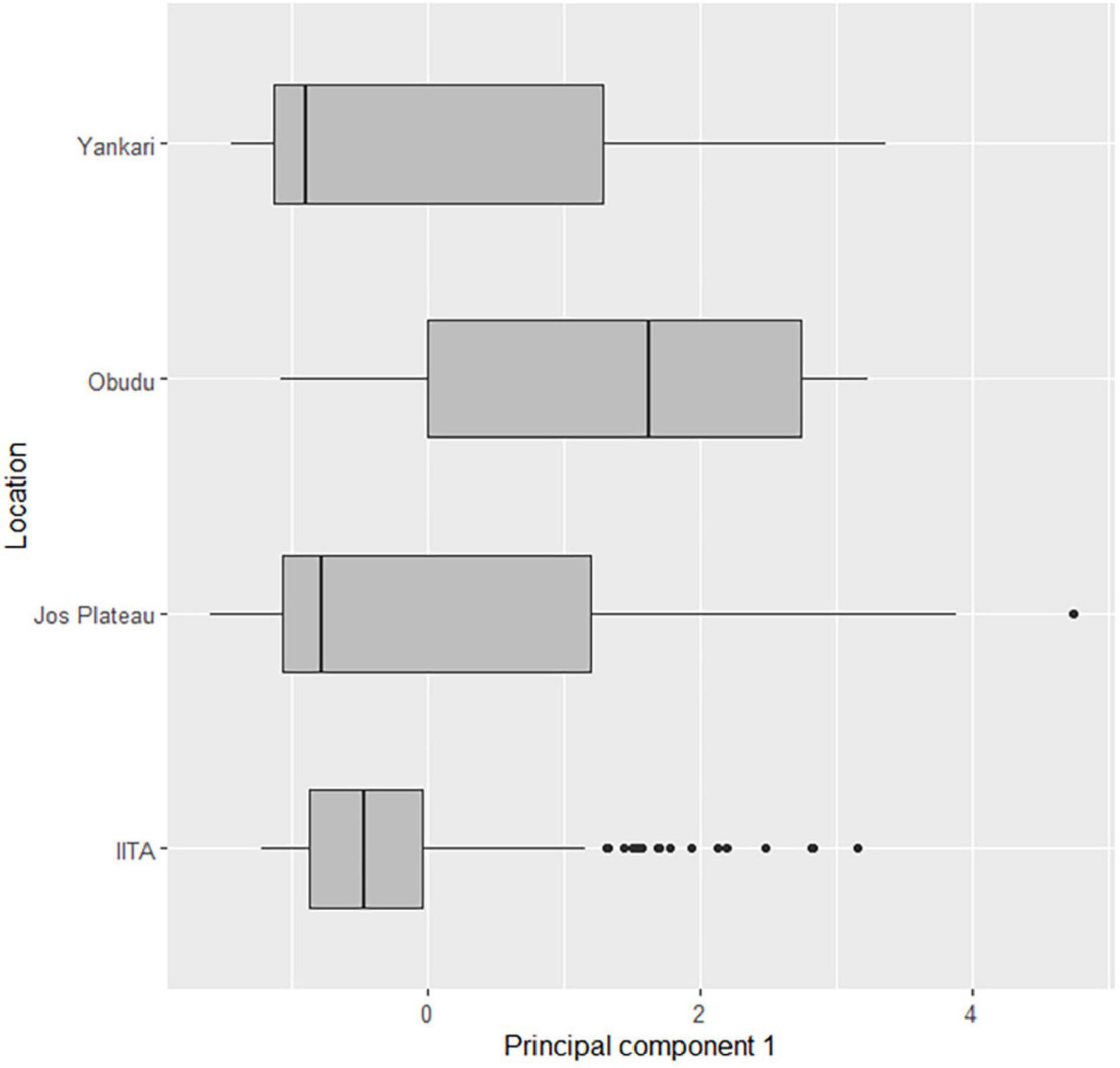
Figure 5. Body size comparisons between northern and southern montane and lowland areas in Nigeria.
For instance, we found that temperature significantly influenced body size in both rainforest and savanna regions, whereas, rainfall only had a significant impact on savanna species populations in the north. This is hardly surprising. Unlike temperature, the duration and volume of rainfall in Nigeria is significantly higher in the southern rainforests in comparison with the northern savanna (Anyadike, 1993; Obot et al., 2010). Shorter rainfall duration and intensity in the north would result in shorter periods of food availability, thus resulting in a lower body size index. In highly variable environments when food availability is limited or unpredictable, a larger body size is a disadvantage because larger bodied individuals require high energy intake in order to meet their energy requirement (Yom-Tov and Geffen, 2006). This premise is supported by our results which show that in the same species, the northern populations were smaller than the southern populations for both lowland and montane regions. In addition, it is likely that the temperature difference between the montane and the lowland habitat in the savanna region was not wide enough to impact on body size difference in the studied populations (Figure 4). Although there was a wide altitudinal gradient between the Jos Plateau and the lowland Yankari, the temperature difference between these sites was less than 1°C.
Our study has not only shown the applicability of the Bergmann’s rule across afrotropical bird species, but also of its potential in understanding the impacts of climate change, which is important from a conservation standpoint. Although we attempted to fill knowledge gaps, our findings have exposed many others, which should provoke further studies. Since Africa supports a host of ecoregions and microclimates along latitudinal and altitudinal gradients (Olson et al., 2001), we would benefit more if further studies are conducted in understudied regions and species, particularly those that are sexually dimorphic (Blanckenhorn et al., 2006; Scriven et al., 2016). This study has also highlighted the need to study the potential impacts of climate change on avian species due to increasing temperature and erratic rains particularly its effect on the physiological state of organisms (Brown, 1996). Body condition, for instance, is known to influence survival and breeding in many species (Kitaysky et al., 1999; Souchay et al., 2013).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by SAFRING, South Africa and the A.P. Leventis Ornithological Research Institute, University of Jos.
Author Contributions
YB and UO: conceptualization. YB: methodology and analyses. YB, AA, JA, and UO: validation and writing—original draft. UO: project supervision. All authors have read and agreed to the published version of the manuscript.
Funding
The data collected for this study are part of the Research activities of the A.P. Leventis Ornithological Research Institute, funded by the Leventis Foundation.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank visiting researchers, colleagues and staff at A.P. Leventis Ornithological Research Institute, and at IITA, who were part of the trapping and ringing of birds at the four different localities. This contribution no. 210 from A.P. Leventis Ornithological Research Institute. We would also like to thank bird ring staff and visiting researchers at the A.P. Leventis Ornithological Research Institute for help in the field capturing and sampling the individual birds.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.741755/full#supplementary-material
Footnotes
References
Adeyanju, T. A., Ottosson, U., Adeyanju, T. E., Omotoriogun, T. C., Hall, P., Manu, S. A., et al. (2014). Birds of the international institute of tropical agriculture campus, a stronghold of avian diversity in the changing Cityibadan area (country-regionplaceNigeria) over the last 50 years. Malimbus 36, 82–101.
Anyadike, R. N. C. (1993). Seasonal and annual rainfall variations over country-regionplaceNigeria. Int. J. Climatol. 13, 567–580.
Ashton, K. G. (2002). Patterns of within-species body size variation of birds : strong evidence for Bergmann’s rule. Glob. Ecol. Biogeogr. 11, 505–523.
Awoyemi, A. G., Olubodun, O., Awoyemi, O. A., Manu, S., Abalaka, J., Abayomi, A., et al. (2020). Emerald forest reserve: a potential important bird and biodiversity area in south-west Nigeria. ABC Bull. 27, 196–203.
Bergmann, C. (1847). Ueber die verhältnisse der wärmeökonomie der thiere zu ihrer grösse. Göttinger Stud. Part 1, 595–708.
Bhusal, D. R., Patel, P., Ghimire, K. C., Bista, M., and Kumar, B. (2019). Size-based intraspecific variations along altitudinal gradient and follow of Bergmann’s rule in Parthenium beetle, Zygogramma bicolorata Pallister. J. Asia. placePac. Entomol. 22, 1173–1179. doi: 10.1016/j.aspen.2019.10.008
Blanckenhorn, W. U., Stillwell, R. C., Young, K. A., Fox, C. W., and Ashton, K. G. (2006). When rensch meets bergmann: does sexual size dimorphism change systematically with latitude? Evolution (N. Y) 60:2004. doi: 10.1554/06-110.1
Braimoh, B., Iwajomo, S., CityplaceWilson, M., Chaskda, A., Ajang, A., and Cresswell, W. (2018). Managing human disturbance: factors influencing flight-initiation distance of birds in a West African nature reserve. Ostrich 89, 59–69. doi: 10.2989/00306525.2017.1388300
Brown, M. E. (1996). “Assessing body condition in birds,” in Current Ornithology. Current Ornithology, eds V. Nolan and E. D. Ketterson (CityplaceBoston, StateMA: Springer).
Clauss, M., Dittmann, M. T., Müller, D. W. H., Meloro, C., and Codron, D. (2013). Bergmann’s rule in mammals: a cross-species interspecific pattern. Oikos 122, 1465–1472. doi: 10.1111/j.1600-0706.2013.00463.x
Cozzolino, D., Power, A., and Chapman, J. (2019). Interpreting and reporting principal component analysis in food science analysis and beyond. Food Anal. Methods 12, 2469–2473. doi: 10.1007/s12161-019-01605-1605
Cresswell, W. (2018). The continuing lack of ornithological research capacity in almost all of placeWest Africa. Ostrich 89, 123–129. doi: 10.2989/00306525.2017.1388301
Crozier, L. G., Hendry, A. P., Lawson, P. W., Quinn, T. P., Mantua, N. J., Battin, J., et al. (2008). Potential responses to climate change in organisms with complex life histories: evolution and plasticity in Pacific salmon. Evol. Appl. 1, 252–270. doi: 10.1111/j.1752-4571.2008.00033.x
Ezealor, E. A. (2001). “Nigeria,” in Important Bird Areas in Africa and Associated Islands- Nigeria: Priority sites for Conservation, eds L. D. C. Fishpool and M. I. Evans (Newbury: Newbury and Cambridge Pisces).
Kitaysky, A. S., Wingfield, J. C., and Piatt, J. F. (1999). Dynamics of food availability, body condition and physiological stress response in breeding Black-legged Kittiwakes. Funct. Ecol. 13, 577–584. doi: 10.1046/j.1365-2435.1999.00352.x
Midgley, G. F., and Bond, W. J. (2015). Future of African terrestrial biodiversity and ecosystems under anthropogenic climate change. Nat. Clim. Chang. 5, 823–829. doi: 10.1038/nclimate2753
Mwansat, G. S., Lohdip, Y. N., and Dami, F. D. (2011). Activities of the A. P. Leventis, the West African foremost ornithological research center. Sci. World J. 6, 9–12. doi: 10.4314/swj.v6i1.70308
Nwaogu, C. J., Tieleman, B. I., Bitrus, K., and Cresswell, W. (2018). Temperature and aridity determine body size conformity to Bergmann’s rule independent of latitudinal differences in a tropical environment. J. Ornithol. 159, 1053–1062. doi: 10.1007/s10336-018-1574-1578
Obot, N. I., Chendo, M. A. C., Udo, S. O., and Ewona, placeI. O. (2010). Evaluation of rainfall trends in country-regionplace Nigeria for 30 years (1978-2007). Int. J. Phys. Sci. 5, 2217–2222.
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial ecoregions of the world: a new map of life on Earth. Bioscience 51, 933–938.
Olson, V. A., Davies, R. G., Orme, C. D. L., Thomas, G. H., Meiri, S., Blackburn, T. M., et al. (2009). Global biogeography and ecology of body size in birds. Ecol. Lett. 12, 249–259. doi: 10.1111/j.1461-0248.2009.01281.x
Omotoriogun, T. C., Onoja, J. D., Tende, T., Manu, S., and Ottosson, U. (2011). Density and diversity of birds in the wetlands of Yankari Game Reserve Bauchi country-regionplaceNigeria. J. Wetl. Ecol. 5, 48–58. doi: 10.3126/jowe.v5i0.4778
R Core Team (2016). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. CityplaceVienna: R Core Team.
Salewski, V., and Watt, C. (2017). Bergmann’s rule: a biophysiological rule examined in birds. Oikos 126, 161–172. doi: 10.1111/oik.03698
Scriven, J. J., Whitehorn, P. R., Goulson, D., and Tinsley, M. C. (2016). Bergmann’s body size rule operates in facultatively endothermic insects: evidence from a complex of cryptic bumblebee species. PLoS One 11:e0163307. doi: 10.1371/journal.pone.0163307
Şekercioĝlu, ÇH., Primack, R. B., and Wormworth, J. (2012). The effects of climate change on tropical birds. Biol. Conserv. 148, 1–18. doi: 10.1016/j.biocon.2011.10.019
Shelomi, M. (2012). Where are we now? Bergmann’s rule Sensu Lato in insects. Am. Nat. 180, 511–519. doi: 10.1086/667595
Sintayehu, D. W. (2018). Impact of climate change on biodiversity and associated key ecosystem services in placeAfrica: a systematic review. Ecosyst. Heal. Sustain. 4, 225–239. doi: 10.1080/20964129.2018.1530054
Souchay, G., Gauthier, G., and Pradel, R. (2013). Temporal variation of juvenile survival in a long-lived species: the role of parasites and body condition. Oecologia 173, 151–160. doi: 10.1007/s00442-013-2613-z
Tieleman, B. I., and Williams, J. B. (2000). The adjustment of avian metabolic rates and water fluxes to desert environments. Physiol. Biochem. Zool. 73, 461–479. doi: 10.1086/317740
Keywords: Bergmann’s hypothesis, Afrotropics, thermal gradients, endotherms, altitude
Citation: Barshep Y, Awoyemi AG, Abalaka J and Ottosson U (2022) Bergmann’s Rule Holds in Birds Inhabiting Southern Guinea Forests but Not in the Northern Savanna of Nigeria. Front. Ecol. Evol. 10:741755. doi: 10.3389/fevo.2022.741755
Received: 15 July 2021; Accepted: 11 May 2022;
Published: 09 June 2022.
Edited by:
Jon Fjeldså, University of Copenhagen, DenmarkReviewed by:
Adriaan Engelbrecht, University of the Western Cape, South AfricaSaeed Mohammadi, University of Zabol, Iran
Copyright © 2022 Barshep, Awoyemi, Abalaka and Ottosson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yahkat Barshep, YmFyc2hlcC55YWhrYXRAZ21haWwuY29t