
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 09 December 2022
Sec. Biogeography and Macroecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.1068200
This article is part of the Research Topic Ecosystem Nutrients Stoichiometry From Local to Global Scales View all 8 articles
 Shuhan Yu1,2†Xiaolong Liu1,2†Xueling Chen1,2Meiling Sun1,2Yongqiang Cao1,2
Shuhan Yu1,2†Xiaolong Liu1,2†Xueling Chen1,2Meiling Sun1,2Yongqiang Cao1,2 Jinjiao Hu3Lixue Yang1,2*
Jinjiao Hu3Lixue Yang1,2* Jian Hu1,2*
Jian Hu1,2*Introduction: The alpine meadows are distributed widely and play a vital role in ecosystem service functions on the Qinghai-Tibet Plateau (QTP). Under the combined effect of climate change and overgrazing, shrubs display an apparent expansion trend, leading to the shrinking of alpine meadows, and directly affecting the structure and function of grassland ecosystems. However, the effects of shrub encroachment on the plant community and soil nutrients of alpine grassland ecosystems still need to be clarified.
Method: We aimed to determine differences in vegetation characteristics and nutrient distribution along the soil profile between shrub patches and their adjacent grassland at three sites, which were three typical types of shrub-encroached grassland, including Spiraea alpina Pall. (SA), Lonicera tubuliflora Rehd. (LT), and Salix cupularis Rehd. (ST).
Results: The results showed that shrub invasion changed the plant community structure of alpine grassland ecosystems, and shrub type was the critical factor driving this alteration. The expansion of the three shrubs reduced grassland species diversity. Shrub encroachment in SA positively impacted vegetation biomass but significantly decreased the soil organic content (SOC) and total nutrients. Shrub invasion in the ST had the most substantial impact on vegetation and soil, resulting in significantly lower nutrient content in shrubs than in grassland patches. The effect of LT was a significant reduction in vegetation biomass but no significant changes in biodiversity or soil nutrients. Grassland patches were more strongly correlated than shrub patches for SA and LT, while the opposite was true for ST. Vegetation characteristics were correlated least with soil nutrients for SA, while ST was most correlated, and LT was between them. Soil nutrients show more positive correlations with vegetation, enzyme activity, and microbial biomass in deeper soils (20–100 cm) than in shallow soils (0–20 cm). The deeper the soil layer is, the more significant the positive correlations in the shrub patches. Our findings indicated that shrubs play critical roles in the dynamics of vegetation patterns and soil environments for managing and sustainable utilization of shrubby alpine grasslands.
The grassland ecosystem is one of the significant global ecosystem types, accounting for approximately 20% of the global land area and covering 40% of China’s land area, with essential ecological functions such as climate regulation, soil and water conservation, pasture supply, and biodiversity maintenance (Naito and Cairns, 2011). However, grasslands are being invaded by shrubs due to global climate change and anthropogenic activities. These shrubs are gradually taking over the dominant position, a phenomenon known as shrub encroachment (Saintilan and Rogers, 2015). In semiarid regions, shrub encroachment has become the main form of vegetation change that has affected the stability of grassland ecosystems. This process generates unavoidable threats to the sustainable development of grassland production and ecological functions (Lorent et al., 2008).
Shrub encroachment is closely related to changes in the structure and function of grassland ecosystems. Shrub encroachment can change vegetation cover types and cause spatial heterogeneity of soil nutrients (Xiong and Han, 2005). Shrub expansion has been shown to destroy the original grassland vegetation, decrease species diversity, and reduce the primary productivity of grassland while increasing soil infiltration (Howard et al., 2012; Hu et al., 2022), changing soil nutrient patterns, and reducing soil nutrient pools (da Silva et al., 2016). However, it has also been shown that shrub invasion of grasslands increases grassland biomass, plant diversity, and richness (Erfanzadeh et al., 2021), promotes nitrogen mineralization (Maestre et al., 2009), and improves soil fertility (Ratajczak et al., 2012). Shrub species and the degree of shrub encroachment may explain these inconsistencies (Eldridge and Ding, 2020). Studies in semiarid grasslands have concluded that shrub species significantly influence the vegetation characteristics and soil properties of the shrub understory due to shrub height and leaf characteristics (Wang X. et al., 2019). The effects of shrub encroachment on plant communities and soil nutrients exhibit context-dependent variability due to differences in plant functional groups, dominant shrub types, and resource habitat conditions, which require further insight into ecosystem structural and functional changes in shrub encroachment, especially in alpine areas.
The natural grasslands on the QTP are widely distributed and large in area, with significant production and ecological functions, supporting the development of the plateau livestock industry (Loreau, 2000). Under the effect of climate change and overgrazing, 39% of the region’s alpine meadows have a noticeable tendency to expand, directly affecting alpine grassland resources and rural development at the cost of reducing alpine meadows (Klein et al., 2007; Geissler et al., 2019; Wang C. et al., 2019). The causes, processes, and ecological impacts of shrub encroachment in different climatic zones around the globe are highly variable, and shrub encroachment in the Tibetan Plateau alpine zone is unique compared to that in arid and semiarid regions (Collins et al., 2020; Criado et al., 2020). The effects of shrub encroachment can depend on ambient temperature and precipitation, which vary rapidly in alpine environments (Rammig et al., 2010). Because of these complexities, we lack a clear understanding of how alpine meadows will affect woody plant–soil interactions. ST, SA, LT, and other shrubs are widely distributed on the Zoige Plateau and are typical of the alpine shrub vegetation in the region, and they are the most influential of the shrubland types (Yang et al., 2017) and soil sampling depths (Chen et al., 2022). The effect of shrub invasion on soil is again determined by soil depth and grassland type, with deep soil organic carbon content showing higher levels in scrub than in grassland (Zhu et al., 2021). Most published studies have focused on the effects of climate (Gao et al., 2016) and grazing (Komac et al., 2013) on shrub encroachment, while the role of vegetation type and soil profile has rarely been considered (Yang et al., 2022). However, the formation mechanism and development trend of shrubby grassland at the whole soil profile in alpine grasslands remain unclear. Therefore, three types of typical shrubby grasslands in the Zoige Plateau, including SA, ST, and LT, are the subjects of this study. By comparing their shrub patches and neighboring grassland patches and combining vegetation characteristics parameters and deep soil physicochemical and functional indicators, the above- and belowground interactions of shrubby alpine grasslands were investigated. There is a theoretical basis for evaluating the nutrient cycling and removal control management of shrubby grasslands in alpine regions.
The study site was on an alpine meadow in Sichuan Zoige Alpine Wetland Ecosystem National Observation and Research Station of Hongyuan County, Sichuan Province of the eastern Qinghai-Tibet Plateau, China (32°49′48″N, 102°34′36″E, 3485 m; Figure 1). The study area is classified as a semihumid climate with an average annual precipitation of 749.1 mm, of which 80% falls during the growing season (i.e., from May to September), an average annual temperature of 1.4°C, average annual sunshine hours of 2158.7 h and average annual snowfall days of 76 days. The soil types are mainly alpine peat soil, subalpine, and alpine meadow soil. The main ecological types include wetlands, shrubs, and alpine meadows. SA, ST, and LT dominate the shrubby grassland ecosystem, with heights of 50–120 cm, 90–180 cm, and 120–280 cm, respectively, and shrub cover of 83, 85, and 90%, respectively (Table 1).
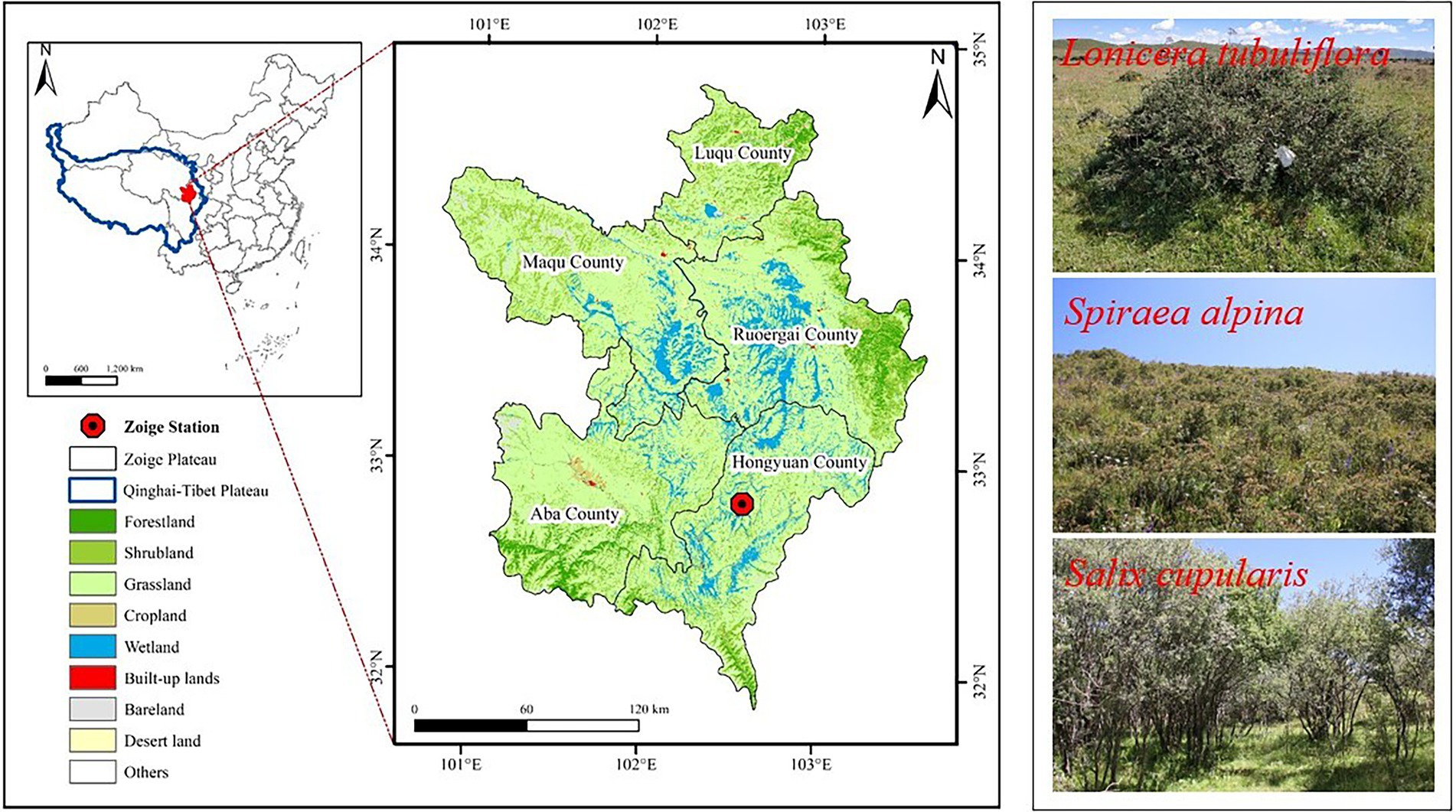
Figure 1. Distribution of shrubs and grasslands of the Zoige Plateau.
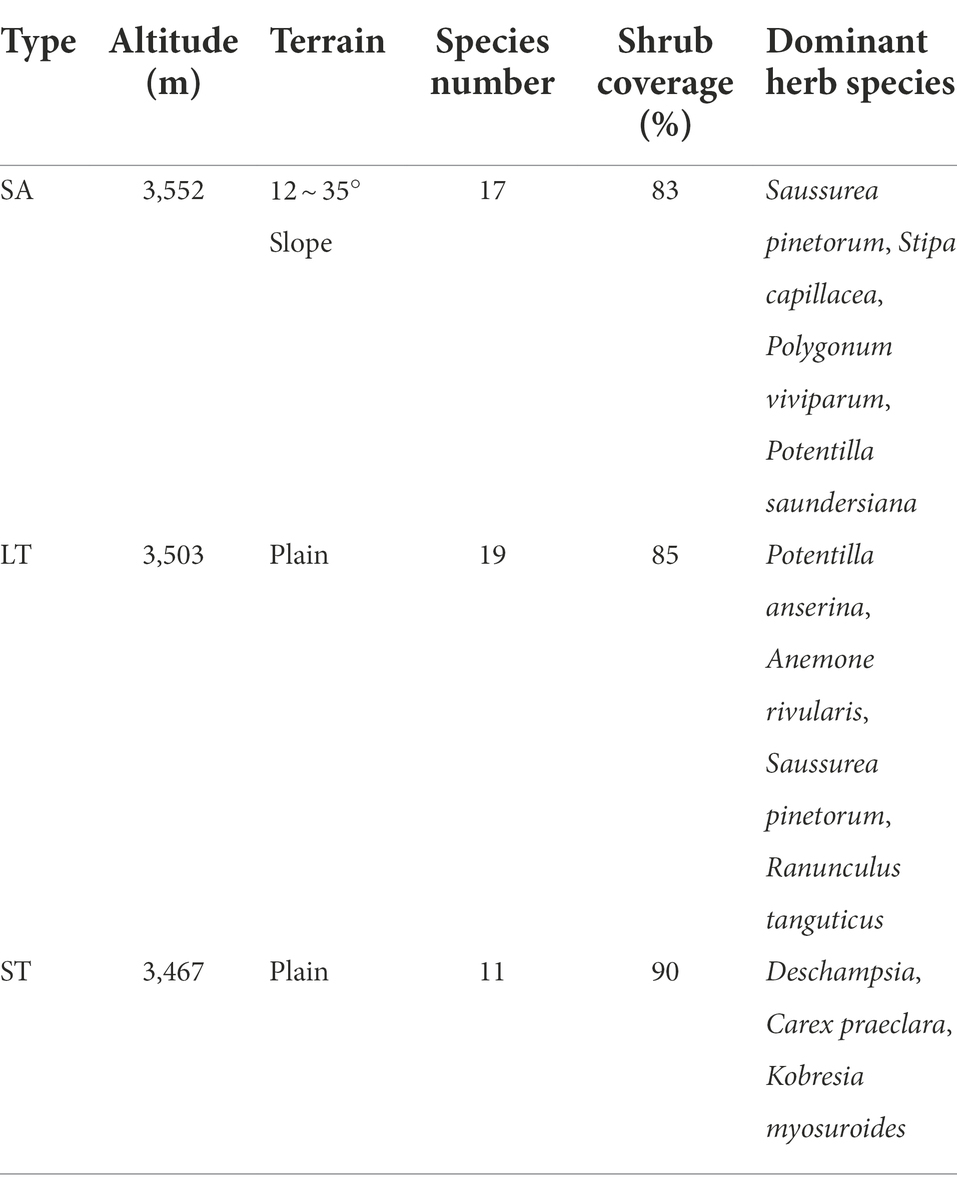
Table 1. General information of the study area.
In August 2019, quadrats of the herbaceous community under the shrub plots and grass plots adjacent to shrubland were randomly selected to survey the plant community with a size of 50 × 50 cm2 during the plant growing season in the winter grazing area (Table 1). We collected soil samples from the 0–100 cm soil profile with three replicates. The latitude, longitude, elevation, community species composition, total community cover, species cover, and the number of species were recorded for each quadrat. The aboveground herbaceous plants were harvested by mowing, brought back to the laboratory, heated at 105°C for 30 min, and then dried at 65°C to constant weight, and the dry weight of each plant was recorded to calculate the aboveground biomass. The Margalef index (R), Shannon-Wiener index (H), Pielou index (E), and Simpson index (D) were calculated as follows:
Where Pi is the relative importance value of species i, , ni is the number of individuals of species i, N is the total number of individuals observed, S is the total number of species in the sample site where species i is located and the diversity index is calculated using the importance value.
After removing aboveground plants, three soil samples were randomly collected from each quadrat at the 0–10 cm, 10–20 cm, 20–40 cm, 40–60 cm, and 60–100 cm soil layers, and we mixed a sample with each soil layer to determine the soil nutrients. SOC was determined via the dichromate oxidation method. Soil pH was measured with the electrode method using a soil: water ratio of 1: 2.5. Total nitrogen (TN) was determined using Kjeldahl analysis. Total phosphorus (TP) and total potassium (TK) were fused by NaOH and measured by molybdenum antimony anti-colorimetric and flame photometry, respectively. Available nitrogen (AN) was measured using the alkaline hydrolysis diffusion method. Available phosphorus (AP) was measured using the molybdenum antimony anti-colorimetric method after samples had been extracted with Na2HCO3, and available potassium (AK) was measured in NH4OAc extracts by flame photometry. Moreover, invertase, β-glucosidase, urease, and acid phosphatase were measured by colorimetric. We used volumetric and iodometric titration methods to analyze catalase and polyphenol oxidase, respectively.
Data analysis was carried out using SPSS 20.0, and all graphs with the mean ± SD were produced using Origin 2021. One-way ANOVA and the least significant difference (LSD) test were used to analyze the vegetation characteristics, soil chemical properties, soil enzyme activity, and soil microbial biomass of different types of shrub-encroached grassland. Furthermore, the Pearson correlation was used to examine the correlation among vegetation characteristics, soil chemical properties, soil enzyme activity, and soil microbial biomass. Redundancy analysis (RDA) was conducted using soil enzymes and soil microorganisms as response variables and soil nutrients such as soil pH, SOC, total nutrients, and available nutrients as explanatory variables by Canoco 5. The significance level of this study was 0.01 and 0.05.
The Margalef, Shannon-Wiener, and Simpson indices of the three types of grassland patches were the highest in LT (p < 0.05; Table 2). Shrub patches of SA showed the highest total coverage and biomass. The total vegetation cover and biomass of the shrub patches in SA increased significantly relative to the grassland patches by 14.12 and 68.97%, respectively (p < 0.05). In addition, all other plant community indices of shrub patches decreased relatively, but the differences were insignificant. The Margalef, Shannon-Wiener, and Simpson indices for the grassland patches of LT were the highest at 3.77, 2.75, and 0.92, respectively. Grassland patches of ST had the highest biomass. The indices of the ST shrub patch were significantly lower than those of the grassland patch with a 30.04% decrease in total cover, 61.11% decrease in biomass, 41.58% decrease in the Margalef index, 24.80% decrease in the Shannon-Wiener index, 7.06% decrease in the Simpson index, and 9.09% decrease in Pielou index (p < 0.05). Indices for shrub patches in ST all had the lowest recorded values. The total cover was 68.33%, a decrease of 30.04% relative to the highest value. The Margalef, Shannon-Wiener, and Simpson indices were 2.15, 1.91, and 0.80, a decrease of 42.97, 30.55, and 13.04% relative to the highest value (p < 0.05). Therefore, ST shrub expansion significantly reduced the vegetation cover, biomass, and biodiversity compared with grassland patches (p < 0.05). In contrast, SA shrub invasion reduced the cover and biomass in grassland patches.
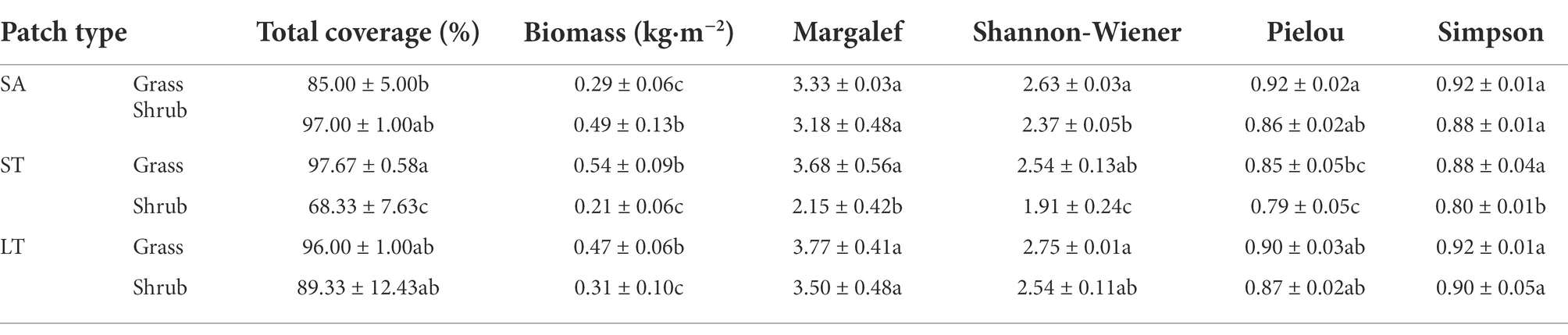
Table 2. The plant community characteristics of three species shrub encroachment grasslands.
The soil nutrients of different shrubby grassland types decreased significantly with the deepening of the soil layer (p < 0.05). The SOC of the LT shrub patches was significantly higher than that of the grassland patches at most soil depths (p < 0.05). The SOC of the ST shrub patches was lower than that of the grassland patches in shallow soil (0–40 cm) and deeper soil (60–100 cm) but higher in the 40–60 cm soil layer. SOC was 5.1% higher in the grass patches than in the SA shrub patches. As the soil depth increased, the pH of the SA shrub patches increased. Soil layers for the SA patches showed that shrub patches were lower than the grassland patches. Soil pH in the shrub patches of ST increased significantly by 18.6%, and SOC decreased significantly by 42.37% compared to the grassland patches (p < 0.05). In particular, pH increased with the depth of the soil layer. In addition, the SOC of grass patches was significantly higher than that of shrub patches at the 0–40 cm soil depth and lower than that of shrub patches at the 40–60 cm soil depth. The SOC of the shrub patches in LT was significantly higher by 83.91% relative to the grassland patches, and there was no significant difference in soil pH (p < 0.05; Figure 2). Soil total nutrients increased in the SA shrub patches by 46.86 and 13.99% for TP and TK but decreased by 1.96% for TN compared to the grassland patches (p < 0.05). The total nutrients of ST shrub patches decreased relative to grass patches by 9.22% for TN and 0.48% for TP but increased by 7.09% for TK. The TN of ST was higher in grassland patches than in shrub patches in the 0–20 cm soil layer but lower in the 20–60 cm soil layer (p < 0.05). ST shrub expansion reduced TP in the 0–20 cm soil depth but increased it in the 20–100 cm soil layer. In addition, TK was consistently higher in the shrub patches than in the grassland patches. LT shrub patches showed a significant increase of 9.49% in TK, 64.45% in TN, and 19.47% in TP compared to grassland patches. LT shrubs increased soil total nutrients in most soil layers (p < 0.05; Figure 3). AN decreased by 25.13% and AP by 12.83%, but AK increased by 2.02% in the SA shrub patches (p < 0.05). The available nutrients in the shrub patches of ST increased compared to the grassland patches, with 39.55% AN, 0.44% AP, and 11.56% AK (p < 0.05). In all soil layers, AN and AP were consistently higher in SA grassland patches than in shrub patches, but AK was lower than in shrub patches (p < 0.05). The available nutrient content was higher in shrub patches in LT in all soil layers. AN and AK were consistently higher in shrub patches than in grassland patches in ST, while AP was lower than in grassland patches in the 0–40 cm soil layer (p < 0.05; Figure 4).
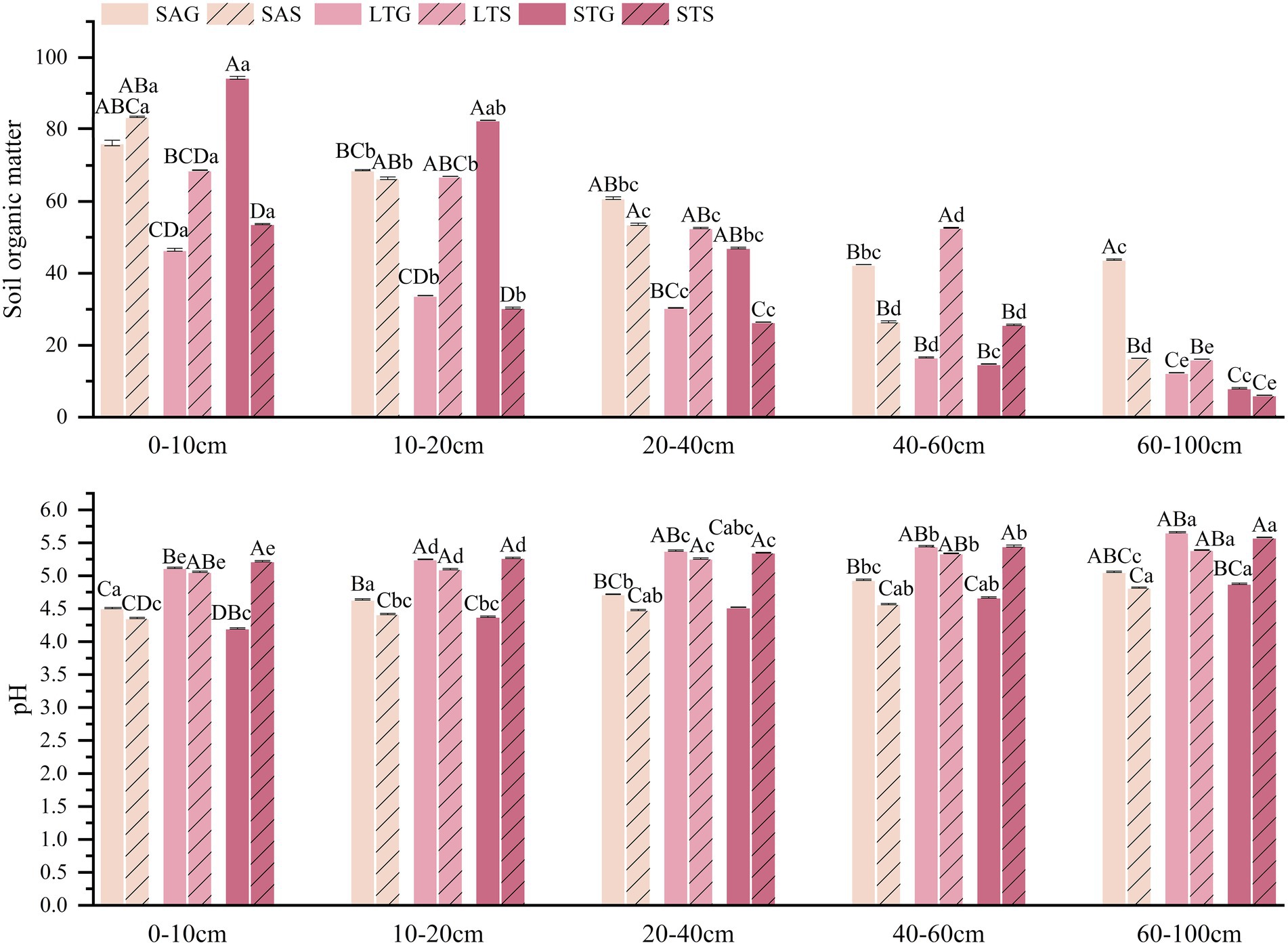
Figure 2. Characteristics of soil pH and SOC in the 0–100 cm soil profile under three shrub-encroached grasslands. SAS, shrub of SA; SAG, grassland of SA; LTS, shrub of LT; LTG, grassland of LT; STS, shrub of ST; STG, grassland of ST. Different lowercase letters indicates significant differences between different soil depths within the same patch at the 0.05 level (p < 0.05), and different uppercase letters indicate significant differences between different patch types at the 0.05 level (p < 0.05) for the same soil depth. The following figures are the same.
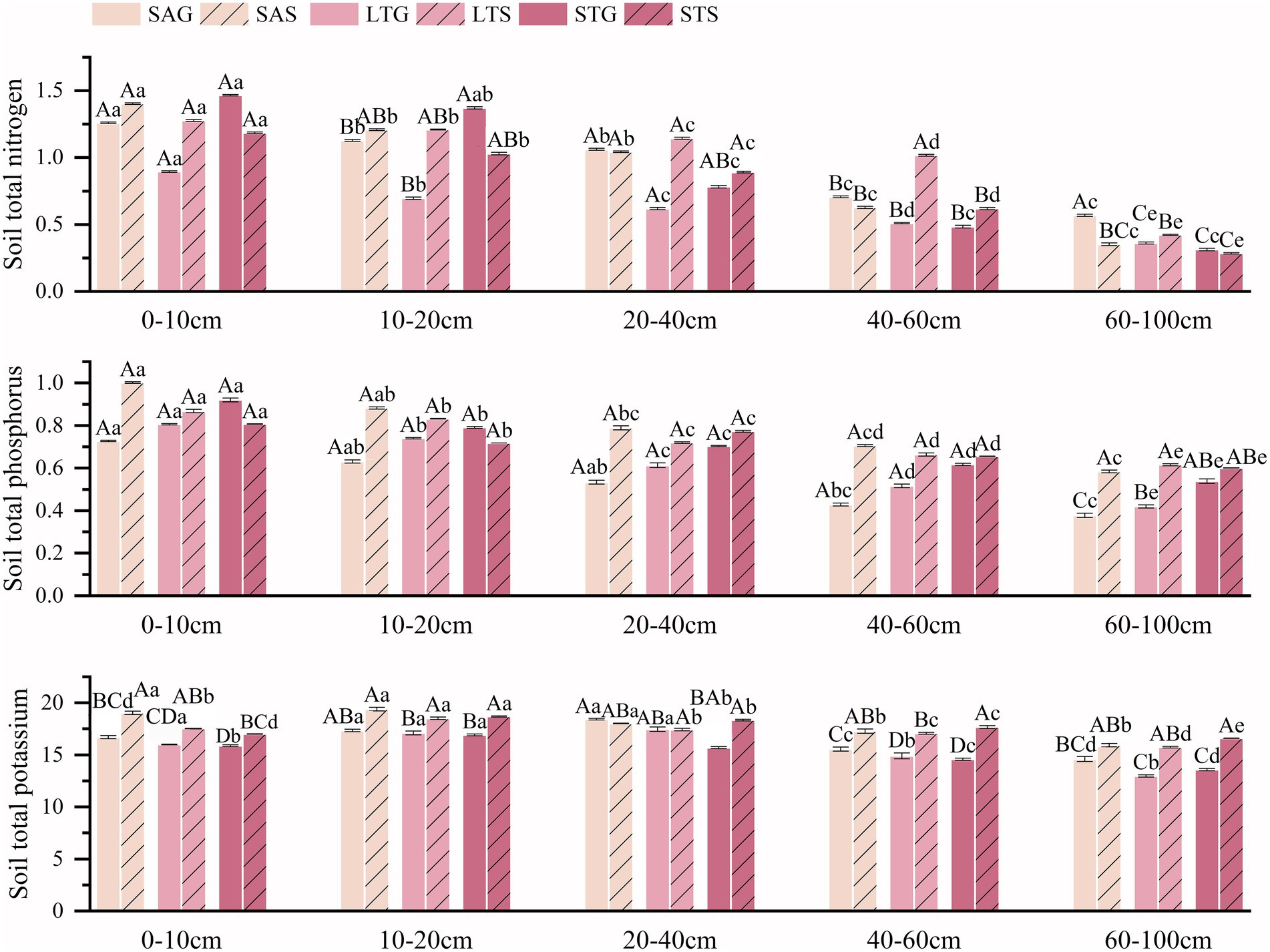
Figure 3. Characteristics of soil total nutrients in the 0–100 cm soil profile under three shrub-encroached grasslands.

Figure 4. Characteristics of soil available nutrients in the 0–100 cm soil profile under three shrub-encroached grasslands.
Shrub invasion by all three dominant shrub species resulted in lower microbial biomass in the deeper soil layer than in the surface soil layer. At the same time, there was a higher proportion of microbial biomass in the shallow soil layer under the three shrub patches than in the adjacent grassland patches (p < 0.05; Figure 5). SA shrub encroachment negatively affected soil microbial biomass carbon (MBC), microbial biomass phosphorus (MBP), and microbial biomass nitrogen (MBN), reducing MBC, MBP, and MBN in all soil layers by 23.56, 3.88, and 7.24%, respectively (p < 0.05). ST shrubs positively affected soil MBC and MBN by 13.69 and 13.68%, respectively, which were significantly higher in the shallow soil layer and lower along the soil depth, while the MBP of the soil reduced at 30.21% (p < 0.05). Differences in MBC were significant at 0–10 cm and 40–60 cm, while MBN was significantly weaker at 60–100 cm. LT shrub expansion positively affected MBC, MBN, and MBP, which increased by 37.82, 25.22, and 30.84%, respectively (p < 0.05). LT shrub patches were significantly higher than grassland patches in shallow and deep soil layers (p < 0.05). LT shrub encroachment increased MBC, which was substantially higher at 10–100 cm soil depths, MBN at 0–10 cm soil depth, and MBP at 10–60 cm (p < 0.05; Figure 5).
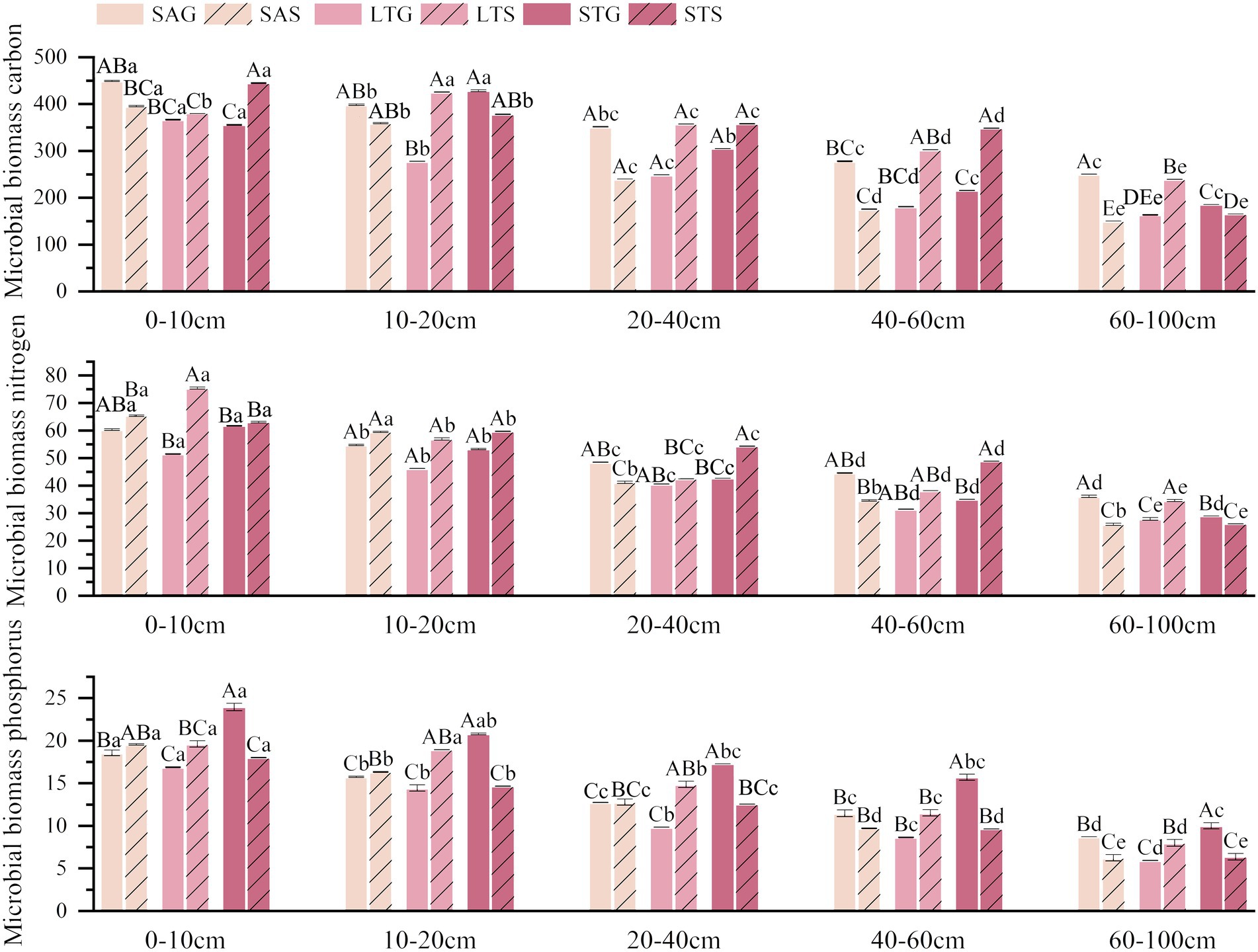
Figure 5. Characteristics of soil MBC, MBN and MBP in the 0–100 cm soil profile under three shrub-encroached grasslands.
The correlations were distinct in deeper soils than in shallow soils, with positive correlations between soil elements increasing significantly as the soil layer deepened (p < 0.01; Figure 6). There was no obvious correlation between vegetation characteristics and soil nutrients. The correlation between vegetation indices and soil nutrients decreased as the soil layer increased (p < 0.01). Grass patches in SA and LT were more strongly correlated than shrub patches, and the opposite was true for ST (p < 0.01). Vegetation characteristics in SA had the lowest correlation with soil nutrients, and ST had the strongest correlation (p < 0.01). The positive correlation between soil enzyme activity and microbial biomass, and soil nutrients increased as the soil layer increased (p < 0.01). In grassland and shrubland, pH was negatively correlated with soil enzymes and soil microbial biomass in all soil layers except for the shallow layers of the SAS, LTS, and STG patches. At shallow soil depths, pH, TK, and AK in SA and LT grassland patches were negatively correlated with soil enzyme activity and microbial biomass. In shallow soil layers, TK and AK were not significantly correlated with soil enzyme activity and microbial biomass in shrub patches of SA. Invertase, acid phosphatase, and MBC were positively correlated with pH, TK, and AK in shrub patches of LT (p < 0.01). In the grassland patches of ST, MBC and invertase were significantly positively correlated with pH, AK, and TK (p < 0.01). The shrub patches of ST, pH, TK, and AK were negatively correlated with enzyme activity and microbial biomass (p < 0.01). In deeper soils, all factors of soil nutrients, except pH, were positively correlated with enzyme activity and microbial biomass.
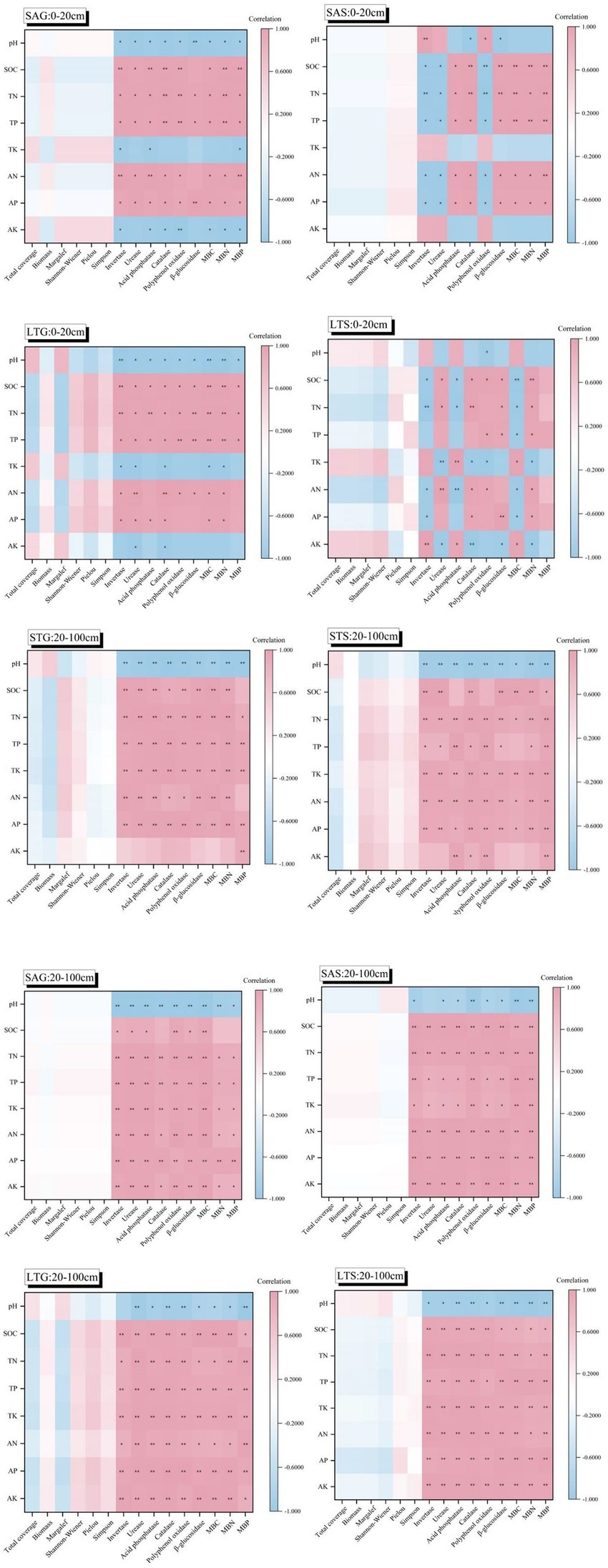
Figure 6. Relevance among vegetation characteristics, enzyme activity, and microbial biomass and soil nutrients under three shrub-encroached grasslands. ** indicates p < 0.01, * indicates p < 0.05.
The RDA of grass patches had 74.16, 87.53, 70.24, 61.96, and 67.95% first-axis interpretation and 25.54, 12.30, 29.55, 37.83, and 31.59% second-axis interpretation from shallow to deep soil layers. The RDA of shrub patches had 83.5, 65.72, 74.81, 88.36, and 69.10% first-axis interpretation and 15.31, 33.54, 24.29, 11.30, and 28.67% second-axis interpretation (Figure 7). According to the RDA ranking diagram, the positive correlations between soil enzyme activity, microbial biomass, and soil nutrients increased with increasing soil layer. The correlation was more substantial in the deep soil layer than in the shallow soil layer. pH and most soil microbial biomass were negatively correlated with enzyme activity, especially in grassland patches. Microbial biomass in grassland patches was mainly positively correlated, especially in the deeper soil layers. Microbial biomass was negatively correlated with SOC, TN, TK, AK, and most enzymes in the shallow soil layers. The deeper the soil layer is, the more pronounced the correlation. The negative correlations for microbial biomass in shrubs are significant in shallow soil layers and positive in deeper ones. They are positively correlated with a small proportion of soil nutrients and enzyme activity. The deeper the soil layer is, the more significant the positive correlations, but they are less significant than in the grassland patches.
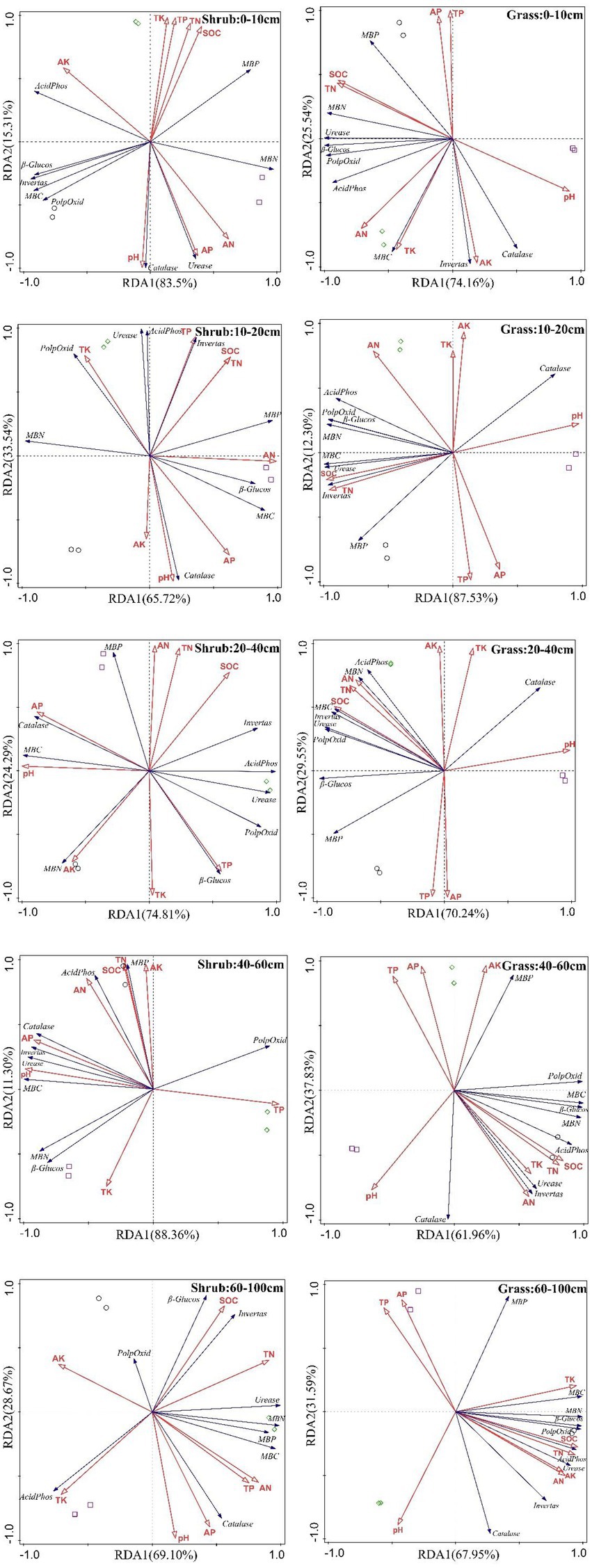
Figure 7. RDA analysis of soil enzyme activity, microbial biomass, and soil nutrients in all soil layers.
Shrubs have mixed effects on grassland ecosystems, with some studies showing that shrubs improve their canopy understory microenvironment and positively impact understory herbaceous growth (Archer et al., 2017). However, it has also been demonstrated that shrubs significantly reduce the net primary productivity, vegetation cover, and species richness of herbaceous plants under their canopy (Howard et al., 2012). In our study, SA shrub patches’ total vegetation cover and biomass were significantly higher than those of grassland patches, consistent with findings in Inner Mongolia grasslands in China (Li et al., 2013). The reasons for this may come from several sources. First, the poor palatability of shrubs increases the difficulty of livestock foraging and provides shelter for shrub understory herbs (Chen et al., 2022). Second, the shrub canopy can have a significant positive effect on understory herbaceous vegetation by improving the understory microenvironment, cover, biomass, and species diversity by improving the understory microenvironment (Howard et al., 2012). Furthermore, shrub patches have higher biomass and litter than their surrounding grassland patches, providing higher soil nutrient inputs (Aguirre et al., 2021). This study’s total cover, biomass, Margalef, Pielou, and Shannon-Wiener indices of LT and ST shrub patches were lower than those of their meadow patches. In particular, ST shrub invasion had a significant adverse effect on the understory herbaceous layer plant community, in contrast to the positive impact exhibited by SA. Studies in the northern grasslands of China have shown that shrub encroachment reduces the herbaceous abundance and aboveground biomass and that shrub type and climate are dominant factors (Zhou et al., 2019). Erfanzadeh also found that shrub type was an essential factor affecting shrub understory herbaceous communities’ productivity and species diversity in semiarid regions (Erfanzadeh et al., 2021). Shrub height and leaf characteristics significantly influence the vegetation characteristics and soil properties of the shrub understory (Wang X. et al., 2019). For example, shrub height, leaf area, and canopy structure affect rainfall redistribution (Yue et al., 2021). Studies on alpine shrub meadows on the Tibetan Plateau have shown that the two factors with the most significant effect of shrubs on the understory herbaceous layer are plant height and leaf area, mainly derived from the fact that the shrub canopy has a light competition effect on the understory herbs (Ackerly, 2004), while shade-tolerant plants have a higher positive response to shrubs (Liancourt et al., 2005). The three types of shrub height and canopy cover showed ST > LT > SA. ST and LT significantly negatively affected herbaceous layer biomass and species diversity due to stronger light competition relative to the herbaceous layer, with ST being the most significant. A study on golden dewberry shrubby meadows in the eastern Tibetan Plateau supports our results and showed that the Shannon-Wiener, Simpson, and Pielou indices of shrub patches were significantly lower than those of intershrub grassland patches, which indicated that grassland patches had higher species diversity and homogeneity relative to shrub patches (Maestre et al., 2009). Different magnitudes of impact due to the shrub characteristics and the contextual differences in climate, soil, and hydrology in various geographical areas cause different effects on the aboveground parts (Montane et al., 2010), but further examination is needed. In our findings, the differences between grasses and shrubs and between the three dominant shrubs may be due to various reasons, such as the characteristics of the shrubs, the physiological differences between the different shrubs, and soil nutrients.
Shrub cover often results in soil heterogeneity, especially in nutrient accumulation in shrubland. Many studies have shown that shrubs can form ‘fertile islands’ by accumulating C and N (Li et al., 2019). The effects of shrub encroachment on soil nutrients vary at the landscape level, ranging from positive to negative or neutral (Eldridge et al., 2011; Du et al., 2016). In contrast, some shrubs positively affect their understory soil conditions at the patch level (Ding et al., 2019). Our study showed that the soil nutrients of all three types of shrub patches decreased with the deepening of the soil layer because extensive root systems of shrubs can transfer soil nutrients from the deep soil layers to the shallow layers (Chen et al., 2022). Shrubs lead to greater litter biomass and N input to the soil, which will substantially affect soil microbial biomass, with microbes increasing the decomposition of fresh organic matter and decreasing the decomposition of SOM, leading to an accumulation of SOM (Liu et al., 2021). However, there is variability in soil nutrients and distribution among different shrub types. Compared to SA grassland patches, SA shrub patches had increased SOC in the shallow soil layer and significantly decreased SOC in the deeper layer. In comparison, LT shrub patches had increased SOC in all soil layers and significantly increased SOC in the lower soil layer. It has been shown that shrubs reduce SOC mineralization compared to grasses on grassland due to the nutrient limitation of microorganisms under shrubs (Jia et al., 2022). Studies also found that shrubs increased the shallow SOC, which is consistent with the results of our research (Rakhsh et al., 2017). The reason may be due to the lower lignin and cellulose content in shrub leaves compared to herbaceous plants resulting in higher decomposition rates of litter in shrub patches than grassland patches, increasing the accumulation of soil SOC (Lyon, 2012). In the present study, ST shrubs significantly reduced the SOC of the upper soil layer. Therefore, dominant shrub species in shrub-encroached grassland played a critical role in soil nutrients between shrub and grass patches. Vegetation types differ in nutrient requirements and utilization ability (Yang et al., 2010). The height, canopy surface, and distance from the canopy to the ground of the three studied shrubs have different effects on the soil and plant communities (Erfanzadeh et al., 2021). Many studies have found that shrubs increase soil N, P, and K (Turpin-Jelfs et al., 2019), and SA and LT shrubs also significantly increase soil TN, TP, and TK in the upper soil layer. ST shrubs significantly increased soil TK, consistent with some findings (Blaser et al., 2014; Zeng et al., 2017).
The ST shrubs significantly positively affected the shallow and deep soil AN and AK. Many studies have shown that the decrease in soil available nutrients is due to a reduction in soil microbial biomass or enzymatic activity, resulting in a slower litter decomposition rate (Wang Y. Q. et al., 2019). MBC, MBN, and MBP were higher in SA grassland patches in our study. Soil microbes are crucial in driving and regulating nutrient cycling in terrestrial ecosystems (Wang Z. Q. et al., 2019, 2022), such as soil carbon retention, organic matter decomposition, and nutrient cycling (Wang et al., 2021). Soil enzyme activity and MBC, MBN, and MBP decreased with increasing soil depth (Fu et al., 2018), and vegetation type can influence soil enzyme activity and microbial biomass (Fanin et al., 2013). In our study, soil MBC, MBN, and MBP were higher in shrub patches than grassland patches at the same soil depth and distributed mainly in the soil surface layer. This increase may be due to the large accumulation of carbon and nitrogen in the soil surface layer (Deng et al., 2018). This makes the surface layer of the soil rich in SOC, which can provide favourable conditions for the survival of microorganisms and soil enzyme activities. With increasing soil depth, SOC decreases rapidly, decreasing microorganisms and enzymes (Wang J. J. et al., 2022). In addition, the soil is rich in SOC due to the high litter biomass and well-developed root system of shrubs (Montane et al., 2010). The results support this finding that the amount and abundance of microorganisms in the soil surface layer were significantly higher in shrub patches than in herbaceous patche (Xie et al., 2021). It is commonly believed that enzyme activity and microbial biomass are positively correlated (Liang et al., 2010), and enzyme activity and microbial biomass are positively correlated with soil SOC and nutrient content (Chen et al., 2016). The present study also showed a similar relationship. Although the MBP of shrub patches in ST was significantly lower than that of ST grassland patches, it was not significantly different from the other two types of shrub patches, and the TP and AP were higher in ST shrub patches than in ST grassland patches. Therefore, it can be explained that soil MBP has a faster turnover rate than soil MBC and MBN and is more sensitive to environmental changes (Zhang et al., 2022), leading to an unconventional increase in MBP in ST grassland patches. The deep soil MBC and MBN of SA shrubs in this study showed a similar decreasing trend with the deep soil SOC, TN, and AN. Moreover, the soil oxidation process is closely connected with soil SOC synthesis and transformation, so catalase is closely related to soil SOC, nitrogen, and phosphorus cycling (Yu et al., 2019). In this study, the SOC AN and AP of SA shrub patches were significantly lower than those of grassland patches.
Our study showed that shrub encroachment had different impacts on the herbaceous community and soil nutrients under shrub and grass patches, while the dominant shrub species played a critical role in the interaction between the above- and belowground consequences of woody plant range expansion. Overall, the negative effect of shrub encroachment on the herbaceous layer was most substantial in ST, resulting in a significant reduction in both vegetation biomass and biodiversity, as well as the apparent spatial heterogeneity in soil nutrients, which is the most influential of the three shrubland types. The shrub invasion of SA increased the total vegetation cover and biomass but was accompanied by a significant decrease in AN and a weakening of richness. The impact of LT was between the three grassland types, with a significant reduction in vegetation biomass but no significant changes in biodiversity or soil nutrients and no apparent degradation of ecosystem function. Moreover, soil nutrients show more positive correlations with vegetation, enzyme activity, and microbial biomass in deeper soils than in shallow soils, and deep soils are more significantly correlated than shallow soils. In addition, the most significant heterogeneity in ecosystem function, structure, and soil nutrients in grasslands caused by shrub invasion in the ST should need long-term observational studies to reveal future changes further.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
SHY, XLL, JH, and LXY conceived and designed the experiments. SHY and XLL performed the experiments with the help of XLC, MLS, and YQC. SHY and XLL performed the data analysis. SHY wrote the manuscript. XLC, MLS, JJH, and JH revised the manuscript with advice from all authors. All authors contributed to the manuscript and approved the submitted version.
This research was funded by the National Natural Science Foundation of China (42007057), the Second Tibetan Plateau Scientific Expedition and Research (2019QZKK0307), the National Science and Technology Fundamental Resources Investigation program of China (2021FY100704), the Sichuan Science and Technology Program (2022JDR0307), the College students’ innovation project of Southwest Minzu University (ZD2022856), and the Fundamental Research Funds for the Central Universities of Southwest Minzu University (ZYN2022010).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ackerly, D. (2004). Functional strategies of chaparral shrubs in relation to seasonal water deficit and disturbance. Ecol. Monogr. 74, 25–44. doi: 10.1890/03-4022
Aguirre, D., Benhumea, A. E., and McLaren, J. R. (2021). Shrub encroachment affects tundra ecosystem properties through their living canopy rather than increased litter inputs. Soil Biol. Biochem. 153:108121. doi: 10.1016/j.soilbio.2020.108121
Archer, S. R., Andersen, E. M., Predick, K. I., Schwinning, S., Steidl, R. J., and Woods, S. R. (2017). Woody plant encroachment: causes and consequences. Rangeland Systems (David D, Springer Open), 25–84.
Blaser, W. J., Shanungu, G. K., Edwards, P. J., and Olde Venterink, H. (2014). Woody encroachment reduces nutrient limitation and promotes soil carbon sequestration. Ecol. Evol. 4, 1423–1438. doi: 10.1002/ece3.1024
Chen, J., Cui, H. Y., Jia, B., Gang, S., Li, Y. W., Li, F. C., et al. (2022). Soil sampling depth matters in assessing the impact of shrubification on soil organic carbon storage in grazed alpine meadows. Geoderma 426:116119. doi: 10.1016/j.geoderma.2022.116119
Chen, J., Luo, Y., Li, J., Zhou, X., Cao, J., Wang, R., et al. (2016). Costimulation of soil glycosidase activity and soil respiration by nitrogen addition. Glob. Chang. Biol. 23, 1328–1337. doi: 10.1111/gcb.13402
Collins, C. G., Spasojevic, M. J., Alados, C. L., Aronson, E. L., Benavides, J. C., Cannone, N., et al. (2020). Belowground impacts of alpine woody encroachment are determined by plant traits, local climate, and soil conditions. Glob. Chang. Biol. 26, 7112–7127. doi: 10.1111/gcb.15340
Criado, M. G., Myers-Smith, I. H., Bjorkman, A. D., Lehmann, C. E. R., and Stevens, N. (2020). Woody plant encroachment intensifies under climate change across tundra and savanna biomes. Glob. Ecol. Biogeogr. 29, 925–943. doi: 10.1111/geb.13072
da Silva, F. H. B., Arieira, J., Parolin, P., da Cunha, C. N., and Junk, W. J. (2016). Shrub encroachment influences herbaceous communities in flooded grasslands of a neotropical savanna wetland. Appl. Veg. Sci. 19, 391–400. doi: 10.1111/avsc.12230
Deng, L., Wang, K., Zhu, G., Liu, Y., Chen, L., and Shangguan, Z. (2018). Changes of soil carbon in five land use stages following 10 years of vegetation succession on the loess plateau, China. Catena 171, 185–192. doi: 10.1016/j.catena.2018.07.014
Ding, L., Wang, P., Zhang, W., Zhang, Y., Li, S., Wei, X., et al. (2019). Shrub encroachment shapes soil nutrient concentration, stoichiometry and carbon storage in an abandoned subalpine grassland. Sustain. Sci. 11:1732. doi: 10.3390/su11061732
Du, B., Ji, H., Peng, C., Liu, X., and Liu, C. (2016). Altitudinal patterns of leaf stoichiometry and nutrient resorption in Quercus variabilis in the Baotianman Mountains, China. Plant Soil 413, 193–202. doi: 10.1007/s11104-016-3093-9
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722. doi: 10.1111/j.1461-0248.2011.01630.x
Eldridge, D. J., and Ding, J. Y. (2020). Remove or retain: ecosystem effects of woody encroachment and removal are linked to plant structural and functional traits. New Phytol. 229, 2637–2646. doi: 10.1111/nph.17045
Erfanzadeh, R., Yazdani, M., and Arani, A. M. (2021). Effect of different shrub species on their sub-canopy soil and vegetation properties in semiarid regions. Land Degrad. Dev. 32, 3236–3247. doi: 10.1002/ldr.3977
Fanin, N., Fromin, N., Buatois, B., and Hättenschwiler, S. (2013). An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system. Ecol. Lett. 16, 764–772. doi: 10.1111/ele.12108
Fu, Z. Y., Sun, J. K., Li, C. R., Chi, Y., Xia, J. B., Qu, F. Z., et al. (2018). Responses of soil microbial biomass to different habitat factors in the chenier of the Yellow River Delta. Acta Ecol. Sin. 38, 6594–6602. doi: 10.5846/stxb201803220571
Gao, Q., Guo, Y., Xu, H., Ganjurjav, H., Li, Y., Wan, Y., et al. (2016). Climate change and its impacts on vegetation distribution and net primary productivity of the alpine ecosystem in the Qinghai-Tibetan Plateau. Sci. Total Environ. 554-555, 34–41. doi: 10.1016/j.scitotenv.2016.02.131
Geissler, K., Fiedler, S., Ni, J., Herzschuh, U., and Jeltsch, F. (2019). Combined effects of grazing and climate warming drive shrub dominance on the Tibetan Plateau. Rangel. J. 41:425. doi: 10.1071/rj19027
Howard, K. S. C., Eldridge, D. J., and Soliveres, S. (2012). Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland. Basic Appl. Ecol. 13, 159–168. doi: 10.1016/J.BAAE.2012.02.008
Hu, J., Cao, Q. H., Liu, X. L., Chen, X. L., Sun, M. L., Zhou, Q. P., et al. (2022). Research progress on the effect of the transition between shrub and grass vegetation on grassland ecosystem and its water-carbon processes. Acta Ecol. Sin. 42, 4324–4333. doi: 10.5846/stxb202106011441
Jia, B., Jia, L., Mou, X. M., Chen, J., Li, F. C., Ma, Q. J., et al. (2022). Shrubification decreases soil organic carbon mineralization and its temperature sensitivity in alpine meadow soils. Soil Biol. Biochem. 168:108651. doi: 10.1016/j.soilbio.2022.108651
Klein, J. A., Harte, J., and Zhao, X. Q. (2007). Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecol. Appl. 17, 541–557. doi: 10.1890/05-0685
Komac, B., Kefi, S., Nuche, P., Escós, J., and Alados, C. L. (2013). Modeling shrub encroachment in subalpine grasslands under different environmental and management scenarios. J. Environ. Manag. 121, 160–169. doi: 10.1016/j.jenvman.2013.01.038
Li, H., Shen, H. J., Zhou, L. H., Zhu, Y. K., Chen, L. Y., Hu, H. F., et al. (2019). Shrub encroachment increases soil carbon and nitrogen stocks in temperate grasslands in China. Land Degrad. Dev. 30, 756–767. doi: 10.1002/ldr.3259
Li, X. Y., Zhang, S. Y., Peng, H. Y., Hu, X., and Ma, Y. J. (2013). Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: results from Inner Mongolia steppe ecosystem of North China. Agric. For. Meteorol. 171-172, 20–30. doi: 10.1016/j.agrformet.2012.11.001
Liancourt, P., Callaway, R. M., and Michalet, R. (2005). Stress tolerance and competitive-response ability determine the outcome of biotic interactions. J. Ecol. 86, 1611–1618. doi: 10.1890/04-1398
Liang, B., Yang, X., He, X., and Zhou, J. (2010). Effects of 17-year fertilization on soil MBC and N and soluble organic C and N in loessial soil during maize growth. Biol. Fertil. Soils 47, 121–128. doi: 10.1007/s00374-010-0511-7
Liu, W. L., Pei, X. J., Peng, S. M., Wang, G. X., Smoak, J. M., and Duan, B. L. (2021). Litter inputs drive increases in topsoil organic carbon after scrub encroachment in an alpine grassland. Pedobiologia 85-86:150731. doi: 10.1016/j.pedobi.2021.150731
Loreau, M. (2000). Biodiversity and ecosystem functioning: recent theoretical advances. Oikos 91, 3–17. doi: 10.1034/j.1600-0706.2000.910101.x
Lorent, H., Evangelou, C., Stellmes, M., Hill, J., Papanastasis, V., Tsiourlis, G., et al. (2008). Land degradation and economic conditions of agricultural households in a marginal region of northern Greece. Glob. Planet. Change 64, 198–209. doi: 10.1016/j.gloplacha.2008.05.005
Lyon, A. J. (2012). Assessment of shrub encroachment on belowground carbon dynamics in tallgrass prairie using an isotopic approach [Master of Science].[Kansas]: Emporia State University.
Maestre, F. T., Bowker, M. A., Puche, M. D., Belén Hinojosa, M., Martínez, I., García-Palacios, P., et al. (2009). Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands. Ecol. Lett. 12, 930–941. doi: 10.1111/j.1461-0248.2009.01352.x
Montane, F., Romanya, J., Rovira, P., and Casals, P. (2010). Aboveground litter quality changes may drive soil organic carbon increase after shrub encroachment into mountain grasslands. Plant Soil 337, 151–165. doi: 10.1007/s11104-010-0512-1
Naito, A. T., and Cairns, D. M. (2011). Patterns and processes of global shrub expansion. Prog. Phys. Geogr. 35, 423–442. doi: 10.1177/0309133311403538
Rakhsh, F., Golchin, A., Al Agha, A. B., and Alamdari, P. (2017). Effects of exchangeable cations, mineralogy and clay content on the mineralization of plant residue carbon. Geoderma 307, 150–158. doi: 10.1016/j.geoderma.2017.07.010
Rammig, A., Jonas, T., Zimmermann, N. E., and Rixen, C. (2010). Changes in alpine plant growth under future climate conditions. Biogeosciences 7, 2013–2024. doi: 10.5194/bg-7-2013-2010
Ratajczak, Z., Nippert, J. B., and Collins, S. L. (2012). Woody encroachment decreases diversity across north American grasslands and savannas. J. Ecol. 93, 697–703. doi: 10.1890/11-1199.1
Saintilan, N., and Rogers, N. (2015). Woody plant encroachment of grasslands: a comparison of terrestrial and wetland settings. New Phytol. 205, 1062–1070. doi: 10.1111/NPH.13147
Turpin-Jelfs, T., Michaelides, K., Biederman, J. A., and Anesio, A. M. (2019). Soil nitrogen response to shrub encroachment in a degrading semi-arid grassland. Biogeosciences 16, 369–381. doi: 10.5194/bg-16-369-2019
Wang, C., Gao, Q., and Yu, M. (2019). Quantifying trends of land change in Qinghai-Tibet Plateau during 2001–2015. Remote Sens. 11:2435. doi: 10.3390/rs11202435
Wang, X., Michalet, R., Liu, Z., Guo, A., Zhou, X., Du, G., et al. (2019). Stature of dependent forbs is more related to the direct and indirect aboveground and belowground effects of a subalpine shrub than are foliage traits. J. Veg. Sci. 30, 403–412. doi: 10.1111/jvs.12739
Wang, J. J., Shu, K. L., Wang, S. Y., Zhang, C., Feng, Y. C., Gao, M., et al. (2022). Soil enzyme activities affect SOC and TN in aggregate fractions in sodic-alkali soils, Northeast of China. Agronomy 12:10. doi: 10.3390/agronomy12102549
Wang, Z. Q., Wang, M. C., Yu, K. L., Hu, H. F., Yang, Y. H., Ciais, P., et al. (2021). Global synthesis for the scaling of soil microbial nitrogen to phosphorus in terrestrial ecosystems. Environ. Res. Lett. 16:04403. doi: 10.1088/1748-9326/abed78
Wang, Y. Q., Yin, Y. L., and Li, S. X. (2019). Physicochemical properties and enzymatic activities of alpine meadow at different degradation degrees. Ecol. Environ. Sci. 28, 1108–1116. doi: 10.16258/j.cnki.1674-5906.2019.06.00
Wang, Z. Q., Yu, K. L., Lv, S. Q., Niklas, K. J., Mipam, T. D., Crowther, T. W., et al. (2019). The scaling of fine root nitrogen versus phosphorus in terrestrial plants: a global synthesis. Funct. Ecol. 33, 2081–2094. doi: 10.1111/1365-2435.13434
Wang, Z. Q., Zhao, M. Y., Yan, Z. B., Yang, Y. H., Niklas, K. J., Huang, H., et al. (2022). Global patterns and predictors of soil microbial biomass carbon, nitrogen, and phosphorus in terrestrial ecosystems. Catena 211:106037. doi: 10.1016/j.catena.2022.106037
Xie, L. N., Soliveres, S., Allan, E., Zhang, G. G., Man, L., Mei, X. F., et al. (2021). Woody species have stronger facilitative effects on soil biota than on plants along an aridity gradient. J. Veg. Sci. 32:e13034. doi: 10.1111/jvs.13034
Xiong, X. G., and Han, X. G. (2005). Spatial heterogeneity in soil carbon and nitrogen resources, caused by Caragana microphylla, in the thicketization of semiarid grassland, Inner Mongolia. Acta Ecol. Sin. 25, 1678–1683. doi: 10.00-0933(2005)07-1678-06
Yang, Y. H., Chen, Y. N., Li, W. H., and Chen, Y. P. (2010). Distribution of soil organic carbon under different vegetation zones in the Ili River Valley, Xinjiang. J. Geogr. Sci. 20, 729–740. doi: 10.1007/s11442-010-0807-4
Yang, X., Guo, Y. P., Mohhamot, A., Liu, H. Y., Ma, W. H., Yu, S. L., et al. (2017). Distribution of biomass in relation to environments in shrublands of temperate China. J. Plant Ecol. 41, 24–28. doi: 10.17521/cjpe.2016.0199
Yang, J. P., Su, P. X., Zhou, Z. J., Shi, R., and Qin, Y. Y. (2022). Shrub expansion is mainly affected by climate-dominated functional traits in alpine meadow. Front. Environ. Sci. 10:917597. doi: 10.3389/fenvs.2022.917597
Yu, D. L., Lei, Z. Y., Zhang, Y. S., Yu, D. W., Zhou, Y. P., and Jiang, J. W. (2019). Soil enzyme activity and its affecting factors under Pinus sylvestris var. mongolica plantation in sandy land. Arid Zone Res. 36, 621–629. doi: 10.13866/j.azr.2019.03.12
Yue, K., De Frenne, P., Fornara, D. A., Van Meerbeek, K., Li, W., Peng, X., et al. (2021). Global patterns and drivers of rainfall partitioning by trees and shrubs. Glob. Change Biol. 27, 3350–3357. doi: 10.1111/gcb.15644
Zeng, Q., Liu, Y., Fang, Y., Ma, R., Lal, R., An, S., et al. (2017). Impact of vegetation restoration on plants and soil C: N: P stoichiometry on the Yunwu Mountain Reserve of China. Ecol. Eng. 109, 92–100. doi: 10.1016/j.ecoleng.2017.10.003
Zhang, Y., Qiang, W., Luo, R. Y., Wang, M., and Pang, X. Y. (2022). Effects of nitrogen and phosphorus addition on soil microbial growth, turnover, and carbon use efficiency: a review. Chin. J. Appl. Environ. Biol. 28, 526–553. doi: 10.19675/j.cnki.1006-687x.2020.11019
Zhou, L. H., Shen, H. H., Chen, L. Y., Li, H., Zhang, P. J., Zhao, X., et al. (2019). Ecological consequences of shrub encroachment in the grasslands of northern China. Landscape J. Ecol. 34, 119–130. doi: 10.1007/s10980-018-0749-2
Keywords: shrub encroachment, plant community, soil nutrient, soil enzyme activity, soil microbial biomass
Citation: Yu S, Liu X, Chen X, Sun M, Cao Y, Hu J, Yang L and Hu J (2022) Effects of shrub encroachment on grassland community and soil nutrients among three typical shrubby grasslands in the alpine subhumid region of the Qinghai-Tibet Plateau, China. Front. Ecol. Evol. 10:1068200. doi: 10.3389/fevo.2022.1068200
Edited by:
Heng Huang, University of California, Berkeley, United StatesReviewed by:
Xukun Su, Research Center for Eco-environmental Sciences (CAS), ChinaCopyright © 2022 Yu, Liu, Chen, Sun, Cao, Hu, Yang and Hu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lixue Yang, eWFuZ2xpeHVlQHN3dW4uZWR1LmNu; Jian Hu, amlhbmh1QHN3dW4uZWR1LmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.