
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 18 January 2023
Sec. Paleoecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.1067432
This article is part of the Research TopicMethods in Paleoecology: 2021View all 8 articles
 Giliane G. Rasbold1*†
Giliane G. Rasbold1*† Ludimila Calheira2†
Ludimila Calheira2† Leandro Domingos-Luz1†
Leandro Domingos-Luz1† Luiz C. R. Pessenda3†Ulisses Pinheiro4†Michael M. McGlue1†
Luiz C. R. Pessenda3†Ulisses Pinheiro4†Michael M. McGlue1†Freshwater sponges (Porifera: Spongillida) are sessile invertebrates with skeletons composed of siliceous elements termed spicules. Sponge spicules (megascleres, microscleres, and gemmuloscleres) are characterized by widely varying sizes and shapes. These spicules are well-preserved in lacustrine, wetland, and riverine sediments and hold significant ecological and limnological information that can be applied as diagnostic tools in reconstructions of Quaternary environments. However, problems with taxonomy and the absence of systematic guidelines and standards of identification represent major challenges to utilizing freshwater sponges as a paleo-proxy. Here, we present a well-illustrated extraction protocol and morphological guide to the Neotropical freshwater sponge fauna. This guide is intended to introduce researchers and students to the study of freshwater sponges and their use as a diagnostic tool in paleoecology and paleolimnology.
Freshwater sponges are sessile animals whose skeletons are composed of microscopic siliceous spicules (needle-like forms) that are often well preserved in lacustrine and riverine sediments (Harrison, 1988). The freshwater sponge fauna is widespread globally, occurring in all zoogeographic zones with exception of Antarctica. These animals are known to colonize nearly all natural or artificial water bodies, from freshwater springs, wetlands, and lakes to thermal vents and saline-alkaline brine (Manconi and Pronzato, 2008). In the last five decades, the number of studies and publications in freshwater sponge ecology has grown exponentially, and many studies have illustrated the global biodiversity of these fascinating organisms (Penney and Racek, 1968; Manconi and Pronzato, 2008). Similarly, freshwater sponge spicules preserved in lake sediments have been used as ecological tools in paleoenvironmental studies (Harrison, 1988). Since then, significant progress has been made by applying freshwater sponge spicules to answer paleoenvironmental, paleoecological, evolutionary, and taxonomic questions (Pronzato et al., 2017; Łukowiak, 2020; Docio et al., 2021; Łukowiak et al., 2022).
The Order Spongillida corresponds to sponges living exclusively in freshwater and is believed to be monophyletic, with a possible marine Haplosclerida ancestor (Manconi and Pronzato, 2002; Pronzato et al., 2017). The fossil record of these sponges extends at least to the late Paleozoic, with confirmed specimens from the Upper Carboniferous (Schindler et al., 2008; Pisera et al., 2013, 2016; Pronzato et al., 2017). To date, a total of 268 species of freshwater sponges have been described globally (De Voogd et al., 2022). The Order Spongillida consists of seven families comprising 47 genera: Spongillidae Gray, 1867 (23 genera); Lubomirskiidae Rezvoi, 1936 (3 genera); Malawispongiidae Manconi & Pronzato, 2002 (5 genera); Metaniidae Volkmer-Ribeiro, 1986 (5 genera); Metschnikowiidae Czerniawsky, 1880 (1 genus); Paleospongillidae Volkmer-Ribeiro & Reitner, 1991 (3 genera); Potamolepidae Brien, 1967 (7 genera). In addition, four genera are incertae sedis (Arinosaster Volkmer-Ribeiro, Tavares-Frigo, Ribeiro & Bichuette, 2021; Balliviaspongia Boury-Esnault & Volkmer-Ribeiro, 1991; Makedia Manconi, Cubeddu & Pronzato, 1999; and Ohridospongilla Gilbert & Hadzische, 1984) (Manconi and Pronzato, 2002; De Voogd et al., 2022). The Neotropical zoogeographic region has the highest species richness with 77 species from 3 families (Spongillidae, Metaniidae, and Potamolepidae) (Pinheiro and Calheira, 2020; De Voogd et al., 2022), followed by the Palearctic (59 species) and Afrotropics (49 species) (Manconi and Pronzato, 2002).
The Neotropical region consists of Central America, the Caribbean, and parts of South America and is well known for its freshwater biodiversity (Antonelli and Sanmartín, 2011; Albert et al., 2020). Sponges play an important role as sessile filter feeders in many inland waters, aiding in circulation, trapping of particulates, and serving as a microhabitat for other organisms, including autotrophs that contribute to primary production (Manconi and Pronzato, 2016). Fossil remains of aquatic plants and animals recovered from continental sediments are particularly valuable for assessing the patterns, timing, and causes of late Quaternary environmental change in the Neotropics and may be insightful for the conservation of threatened ecosystems (Sifeddine et al., 2001; Parolin et al., 2007; Bush et al., 2016; Guerreiro et al., 2019; Rasbold et al., 2021). In this context, sponges are underutilized. It is common, for example, that researchers note the presence of sponge spicules in studies of other siliceous microfossils, such as diatoms, chrysophytes, and plant phytoliths, without attempting a description of spicule morphologies or identification (Trombold and Israde-Alcantara, 2005; Bush et al., 2016). To date, many if not most studies that have incorporated insights from sponge fossils have done so using a semi-quantitative approach, with species presence/absence quantitative and relative abundance of different species qualitative (e.g., McGlue et al., 2012; Rasbold et al., 2019; Rasbold et al., 2021). The methodology outlined here will allow more quantitative assessments of species assemblages in Neotropical sediment sequences, although we caution that new autecological studies are needed to help refine the environmental tolerances of the known taxa, and additional taxonomic and genetic studies are likewise needed to improve knowledge of the Porifera. Descriptions of living sponges from lowland rivers in Brazil, for example, have informed paleoecological analyses in the region, and much of what is known about ecological relationships is owed to these foundational studies (Volkmer-Ribeiro and Maciel, 1983; Volkmer-Ribeiro, 1992; Volkmer-Ribeiro and Costa, 1992). Sponge spicule preservation varies and may be affected by taphonomic processes, including fragmentation, abrasion, and dissolution (particularly in saline-alkaline brines). Preservation biases may influence interpretations, and new analysts are cautioned against overinterpreting broken, abraded spicules, or assemblages marked by selected preservation. Nonetheless, even patterns of spicule damage can be revealing of hydrodynamic energy, chemistry, and bioturbation, and in conjunction with additional proxies, reliable inferences on past environments can be achieved (Guerreiro et al., 2017).
Freshwater sponges are composed of three distinct morphological spicules, namely, megascleres, microscleres, and gemmuloscleres. These skeletal elements are often well preserved in lacustrine and riverine sediments. For identification to the genus level, megascleres and microscleres are usually sufficient, but gemmuloscleres are the most important structure for the determination of species (Potts, 1887). In sediments, spicules from different species can be deposited together after skeletal dissociation, which can be a challenge for the correct species identification, not to mention the potentially complicate effects of postmortem preservational biases (taphonomy). However, few guides for sponge morphology are available to assist researchers and students, which presents a roadblock to fully gaining insights into paleoecology and paleolimnology from these fossils. A few studies are available on the use of sponges as a proxy in paleoenvironmental studies (Docio et al., 2021), taxonomy, geographic distribution, and critical review of the fossil freshwater sponges (Pronzato et al., 2017; Łukowiak, 2020), and more recently the terminology of sponge spicules (Łukowiak et al., 2022). This stands in contrast to those available for other siliceous microfossils such as diatoms and phytoliths (e.g., Neumann et al., 2019; Spaulding et al., 2021).
Here, we present an illustrated guide for extracting, identifying, and quantifying Neotropical freshwater sponge spicules for non-specialists. Sponge spicules are often encountered with pollen grains (when the extraction method does not use hydrofluoric acid), phytoliths, or diatom frustules in paleolimnological studies but few analysts are trained to identify sponge spicules. The motivation for this guide is to expand access and agency for paleoecologists and students who encounter these fossils in their studies.
To extract sponge spicules from sediments, the wet oxidation methodology is often used (e.g., Battarbee, 1986; Harrison, 1988; Volkmer-Ribeiro and Turcq, 1996; Figure 1). Through this method, it is possible to extract and prepare other siliceous microfossils (e.g., plant phytoliths and diatoms) as well. The extraction steps are as follows:
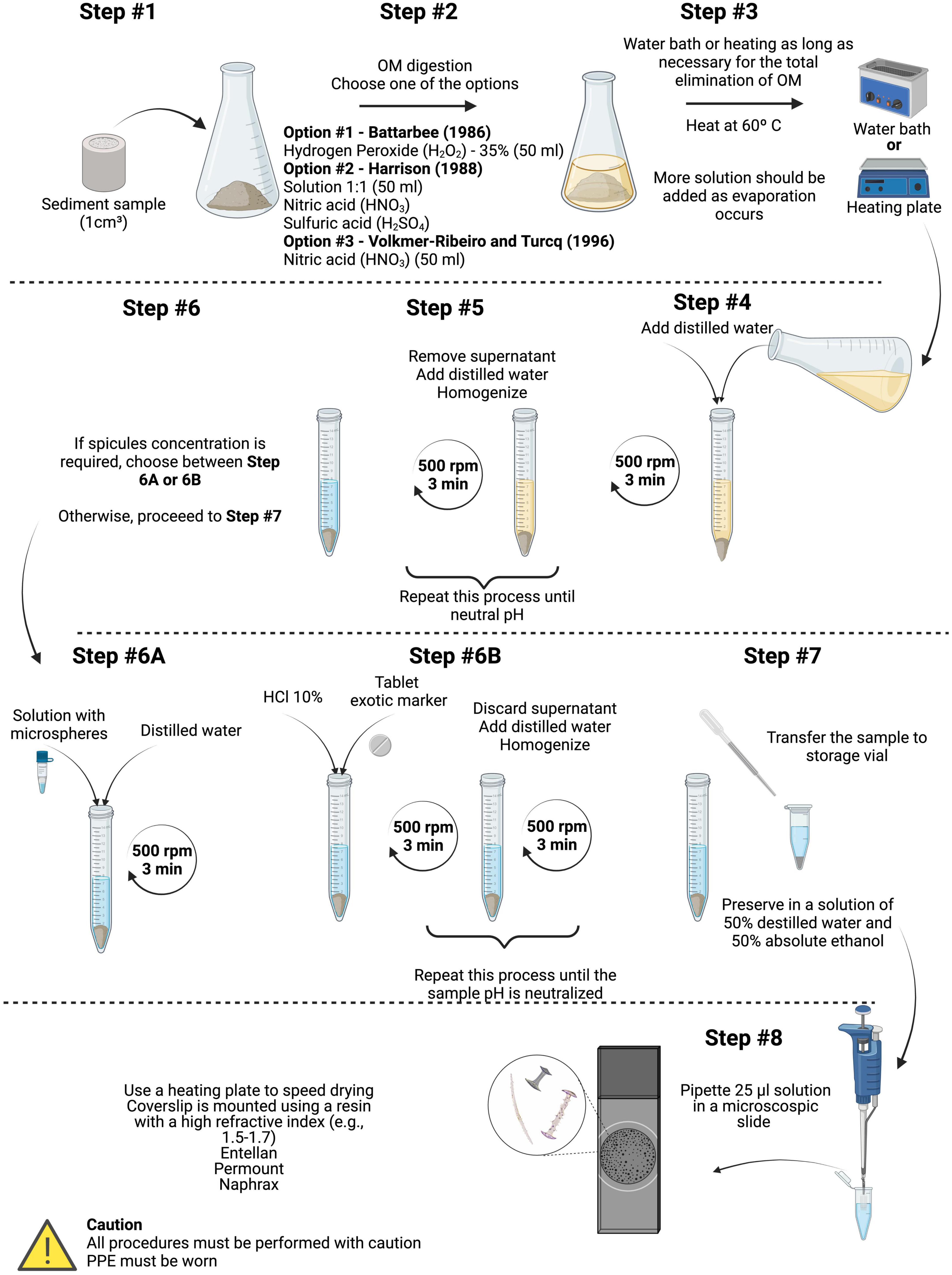
Figure 1. Flow chart of the wet oxidation method of extracting sponge spicules from sediments. Methodology adapted from Battarbee (1986), Harrison (1988), and Volkmer-Ribeiro and Turcq (1996). OM, organic matter; PPE, personal protective equipment.
Step #1: Measure and weigh ∼1 cm3 of wet sediment, in practice the initial amount of sediment used may vary based on the depositional environment and richness of spicules. Wash the samples using distilled water (DI) heated to 80–90°C in a 250-ml Erlenmeyer flask (or similar). Hot distilled water helps to break up clays and remove calcium ions that may interfere with some heavy liquids (Zhao and Pearsall, 1998).
Step #2: Under a fume hood and using the proper personal protective equipment (PPE), proceed to organic matter (OM) digestion using one of the following techniques: a) adding 50 ml of hydrogen peroxide (H2O2) (Battarbee, 1986); b) adding 50 ml of 1:1 solution of nitric acid (HNO3) and sulfuric acid (H2SO4) (Harrison, 1988); or c) adding 50 ml of nitric acid (HNO3) (Volkmer-Ribeiro and Turcq, 1996).
Step #3: Move the samples to a water bath or hot plate at 60°C to accelerate the digestion of the OM. Turn off the heat source after the sample ceases to react with the acid (fizzing/bubbles are no longer generated at the surface). Note that it may be necessary to add more acid solution to complete OM removal, again depending on the depositional environment.
Steps #4 and #5: After cooling, transfer the samples to 50-ml centrifuge tubes, fill them with DI, and centrifuge for 3 min under 500 rpm. Decant and repeat this step until neutral pH (∼3 to 5 times) is achieved. Alternatives, such as allowing the spicules to settle under the influence of gravity under longer periods, are also possible.
Step #6: If the study requires spicule concentration, choose one of the following sub-steps; otherwise, proceed to step #7. Step #6A: Add a solution of glass microspheres, complete with DI, and centrifuge the sample for 3 min under 500 rpm and decant. Or step #6B: Add a tablet of exotic marker and 10 ml of hydrochloric acid (10%), and centrifuge for 3 min under 500 rpm. Decant, refill with DI water, and centrifuge for 3 min under 500 rpm (repeat until a neutral pH is obtained).
Step #7: Decant and transfer the contents to storage vials with proper labeling in a 1:1 solution of DI water and 95% ethanol.
Step #8: Prepare microscope slides by pipetting 25-μl sample solution onto a cleaned glass slide and drying it on a hot plate. After cooling, cover with a coverslip using a resin with a high refractive index of 1.5–1.7 (e.g., Entellan, Permount, or Naphrax).
This guide has been designed based on the available taxonomic literature for the Neotropical region, including the World Porifera Database (De Voogd et al., 2022), personal collections of the authors, and microscope slides archived in the Porifera Laboratory (LABPOR) at Pernambuco Federal University (Recife, Brazil). This guide does not replace the need to consult the proper taxonomic keys, as well as the literature describing the species since morphological variations are possible and cannot be fully explored here. We describe here the spicules of 27 genera of freshwater sponges from the Neotropical region (Figures 3–14). Freshwater sponges (Neotropics fauna) were identified based on their morphology, exception given to the genus Rosulaspongilla (Sokolova et al., 2021), which was identified based on DNA (Supplementary Material 1). Some genera may have morphological differences or similarities when compared to species from other zoogeographic regions. In total, 15% of the species found in the Neotropical region also occur in other areas of the world, such as in the Oriental Region (OL), Nearctic Region (NA), Palaearctic Region (PA), Australian Region (AU), and Afrotropical Region (AT) (Supplementary Material 1).
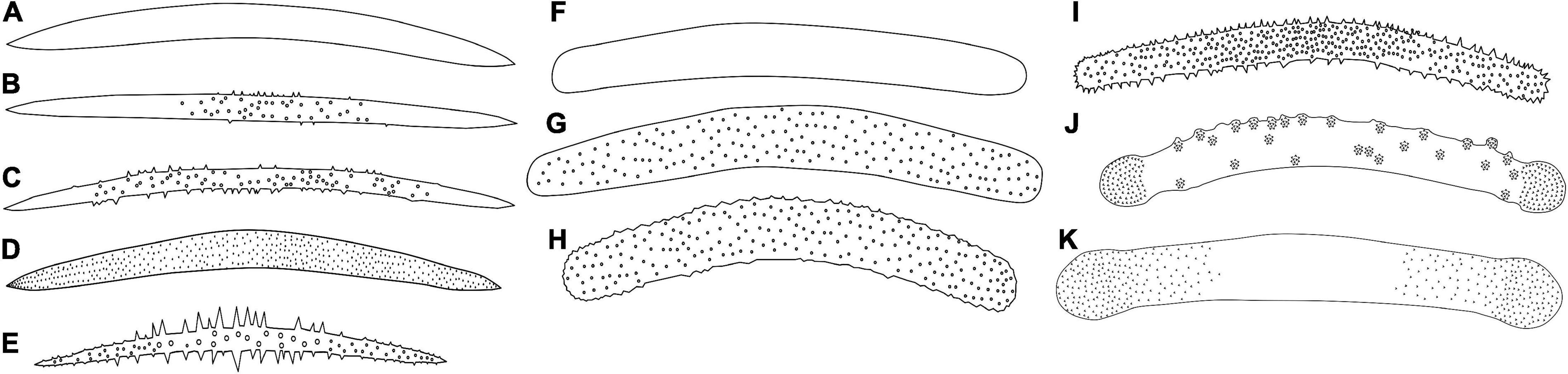
Figure 2. Megascleres descriptors. (A) Oxea. (B) Oxea with microspines in the center of the spicule. (C) Acanthoxeas. (D) Acanthoxeas are covered with spines. (E) Acanthoxeas, covered by spines, with conical spines in the center region. (F) Smooth strongyle. (G–I) Acantostrongyle. (J) Strongyles microgranulated, the spines grouped to form spots with inflated tips. (K) Strongyles microgranulated with inflated ends.

Figure 3. (A–E) Anheteromeyenia Schröder, 1927. Gemmuloscleres. (A) Acanthostrongyle; (B) acanthoxea; (C) pseudobirotules; (D,E) grading from long to very short, from stout to slim, acanthostrongyles and acanthoxeas. (F–P) Corvoheteromeyenia Ezcurra de Drago, 1979. Microscleres. (F–H) Pseudobirotule with long hooks; (I,J) pseudobirotule with short hooks; (K,L) acanthoxea with simple and/or compound spines; Gemmuloscleres. (M) Birotules with rotules microspines on their margins to convex; (N) birotules with rotules microspines on their margins to flat; (O,P) sanidaster. (Q–S) Corvospongilla Annandale, 1911. Microscleres. (Q) Pseudobirotule with four to seven radial hooks on each pseudorotule and smooth shaft; Gemmuloscleres. (R) Microspined strongyles. (S) Microspined oxea. Scale: 10 μm.
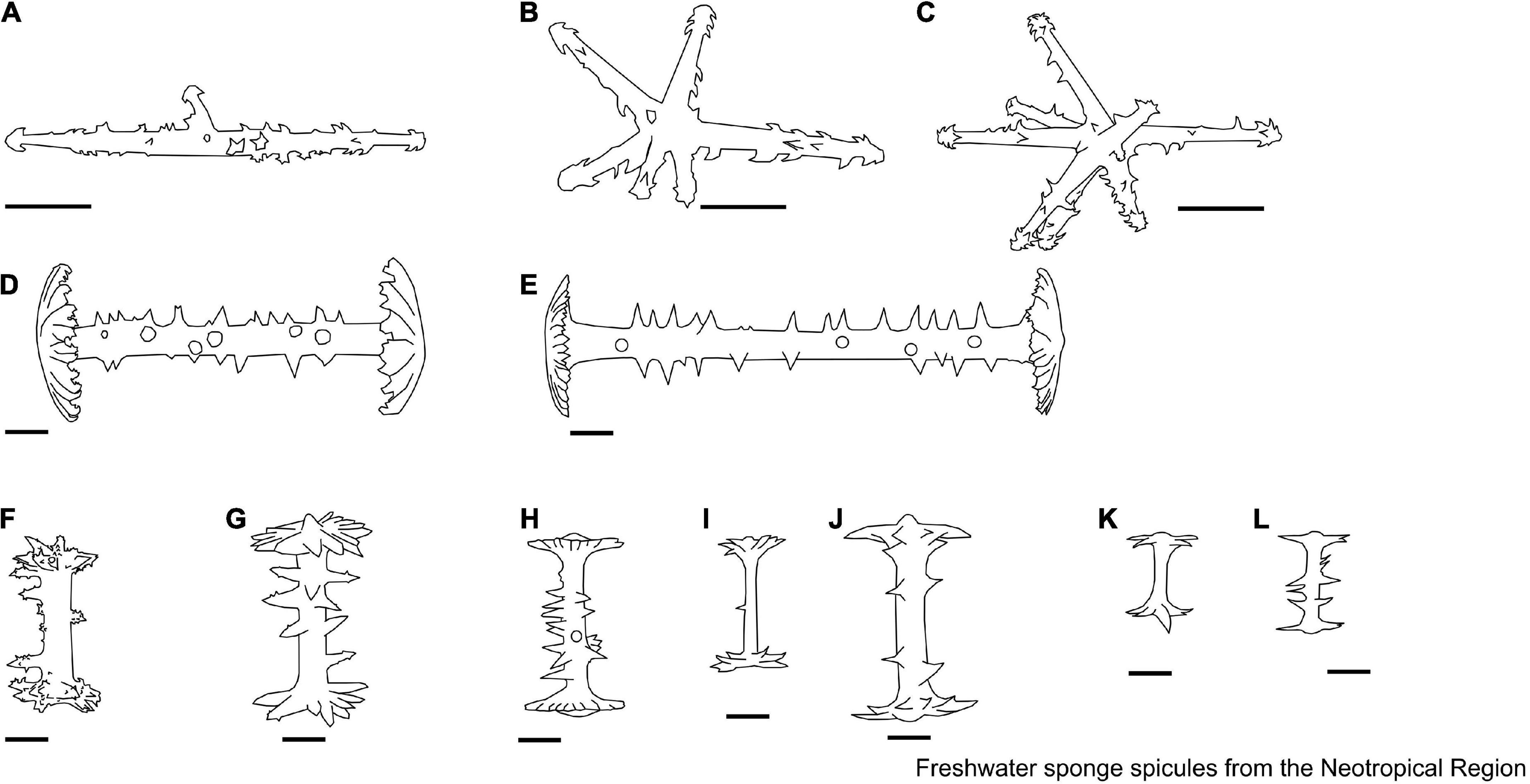
Figure 4. (A–E) Dosilia Gray, 1867. Microscleres. (A) Aster from simple acerates with one more long divergent spined branch; (B,C) spherical aster-radiated bodies. Gemmuloscleres. (D) Birotules, spined shaft, rotules convex; (E) birotules, spined shaft, rotules of size and shape equal, flat or slightly umbonate with margins thin and numerous spines. (F–L) Ephydatia Lamouroux, 1816. Gemmuloscleres. (F) Birotules spined shaft, with secondary spines; (G) birotules with irregular rotules, shaft with long and conical spines; (H) birotules with spined shaft, flat rotules; (I–L) birotules with a smooth shaft, flat rotules, not deep irregular incisions. Scale: 10 μm.
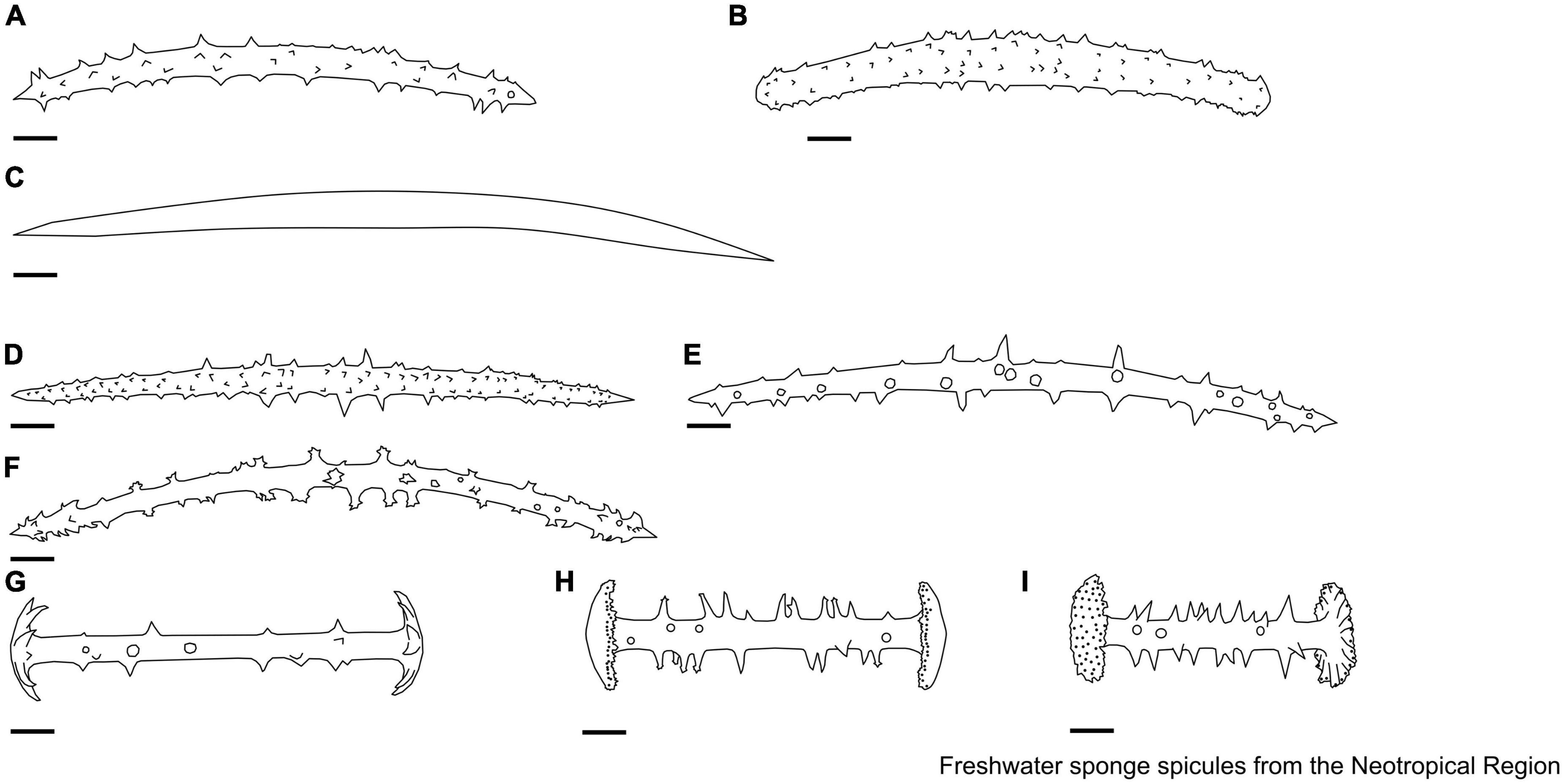
Figure 5. (A–C) Eunapius Gray, 1867. Gemmuloscleres. (A) Acanthoxea; (B) acanthostrongyles; (C) oxea. (D–I) Heteromeyenia Potts, 1881. Microscleres. (D–F) Acanthoxeas, spines can be straight or curved, simple or compound (have a bouquet-like structure); gemmuloscleres. (G) Pseudobirotules; (H) birotules, shafts with conical spines (simple or compound). Rotules are smooth with microspines on their margins; (I) birotules with the spined shaft. Rotules are entirely covered in microspines, with serrate margins. Scale: 10 μm.
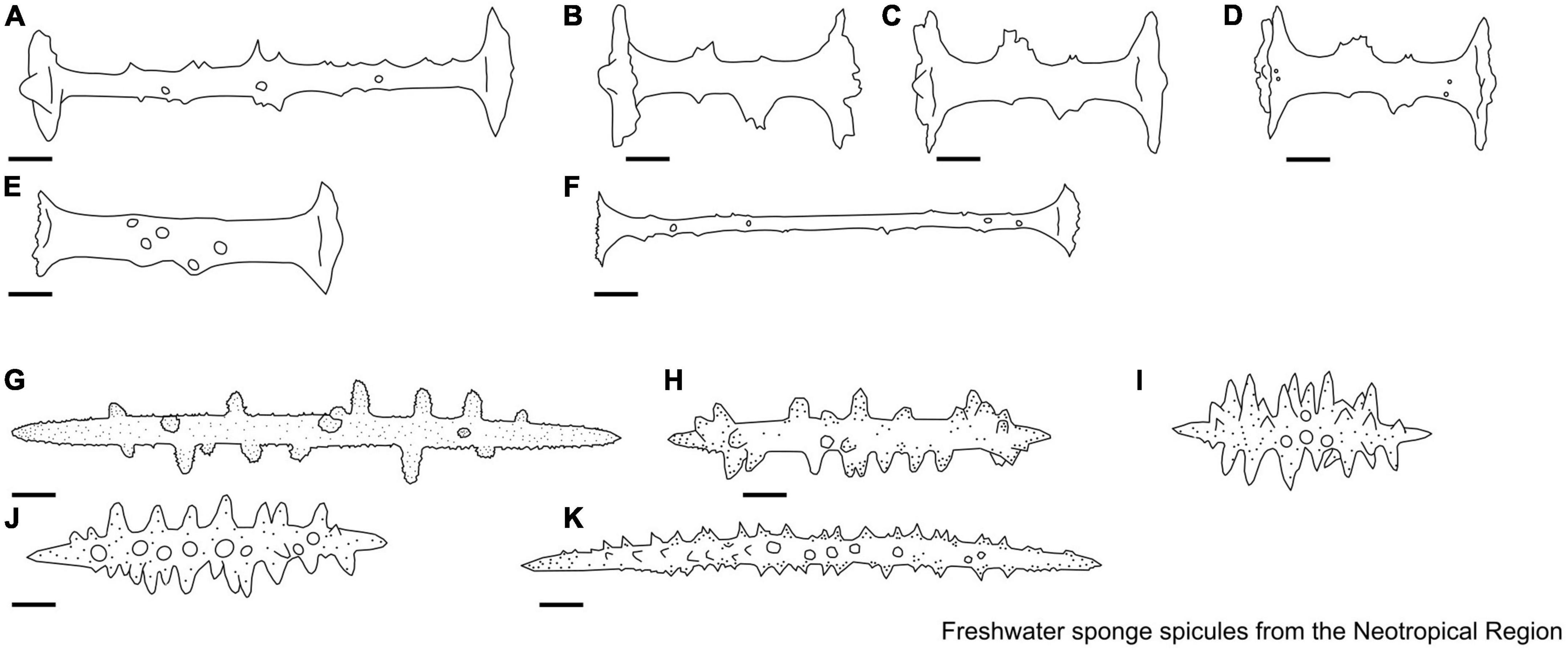
Figure 6. (A–F) Heterorotula Penney & Racek, 1968. Gemmuloscleres. (A) Birotule with long axis, with almost flat rotules; (B–D) birotules with short axis, rotules with irregular edges, some large and irregular spines; (E) birotules with short axis, with incomplete rotules; (F) birotules with long axis, with incomplete rotules. (G–K) Pottsiela Volkmer-Ribeiro, Machado, Fürstenau-Oliveira & Soares, 2010. Microscleres. (G–K) Acanthoxeas with microspined bearing along its length, conical spines also microspined. Scale: 10 μm.
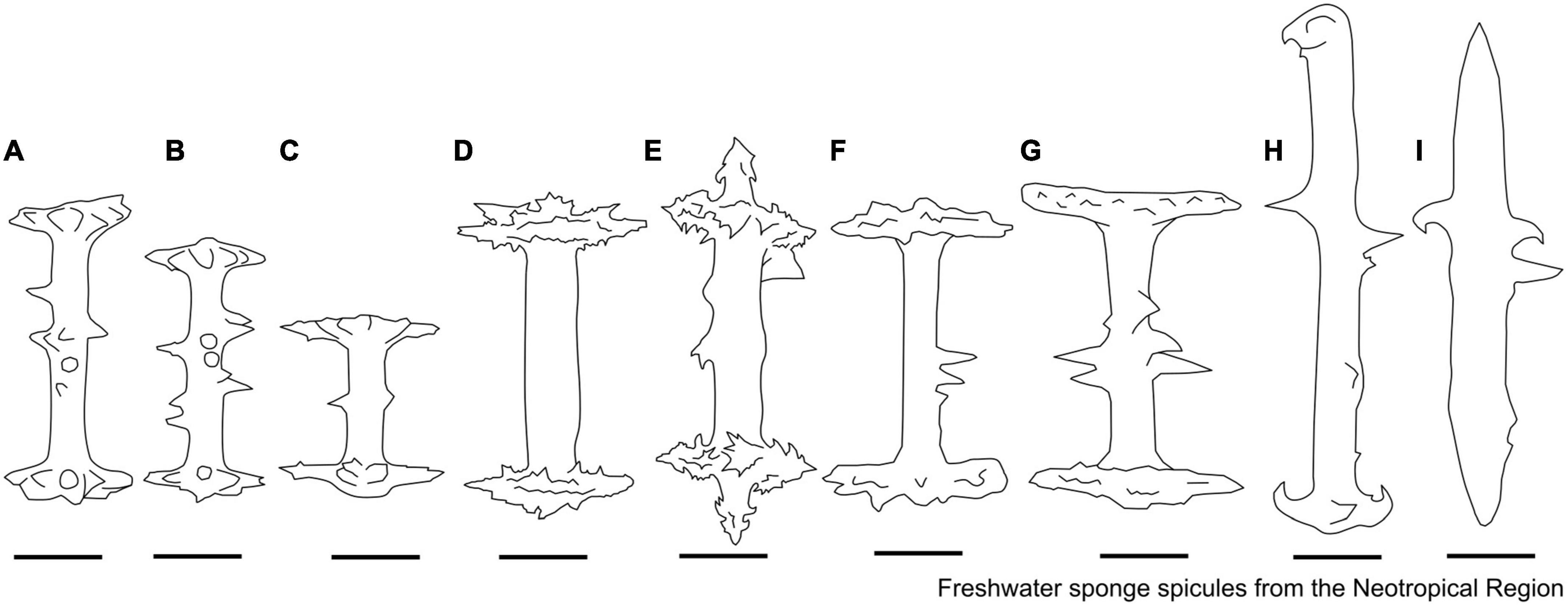
Figure 7. (A–I) Racekiela Bass & Volkmer-Ribeiro, 1998. Gemmuloscleres. (A–G) Birotules, short thin, or robust shafts with smooth or with few spines; (H,I) pseudobirotules with long and spiny shaft. Scale: 10 μm.
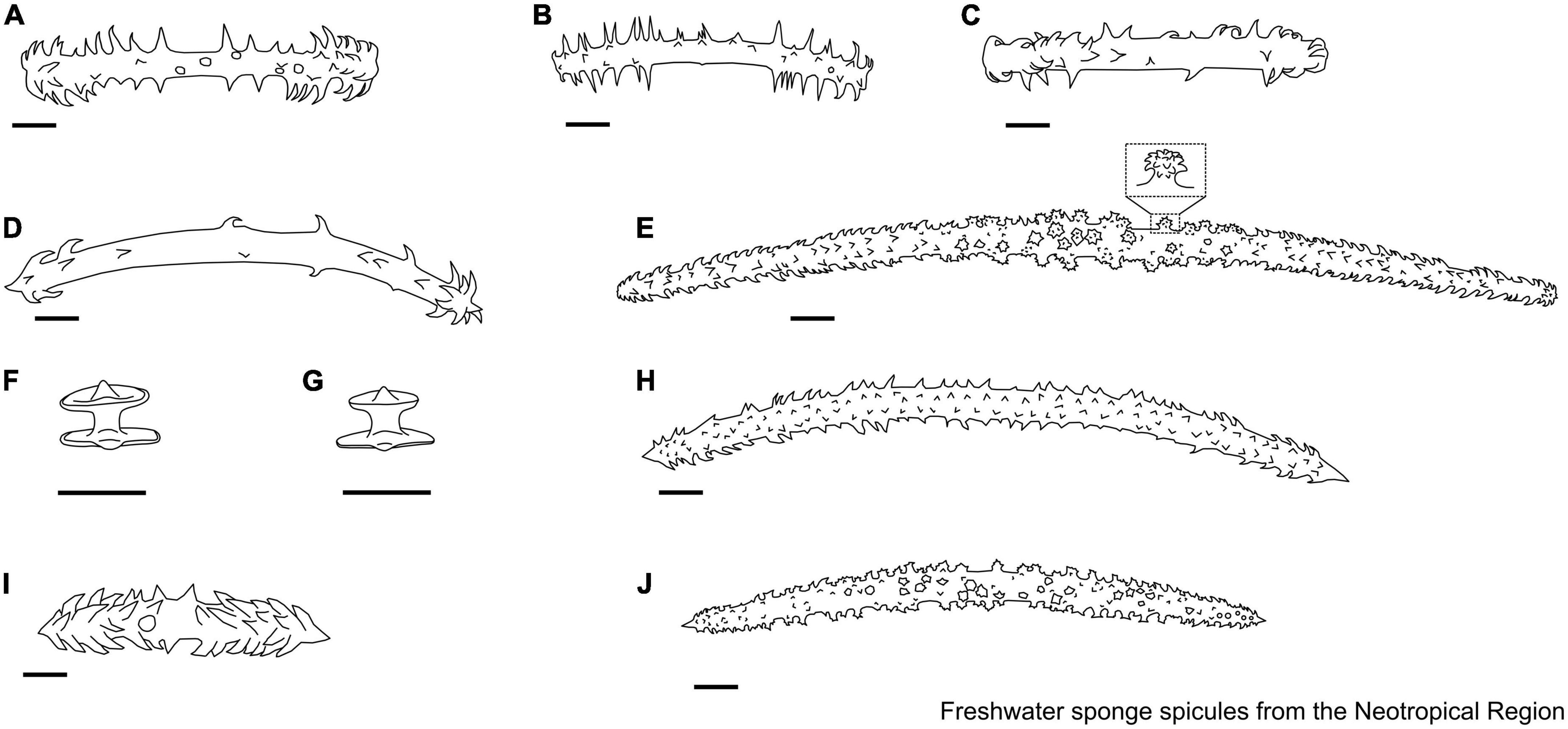
Figure 8. (A–C) Radiospongilla Penney & Racek, 1968. Gemmuloscleres. (A) Acanthostrongyles, the spines are concentrated at the tips, forming hooks; (B) acanthostrongyles, the spines are straight and sharp; (C) pseudobirotule with spiny shaft and bent long spines at the apices. (D,E) Rosulaspongilla Sokolova, Palatov, Masuda & Itskovich, 2021. Gemmuloscleres. (D) Acanthoxeas with large, curved spines more thickly accumulated at the tips and often form mace-shaped structures; Microscleres. (E) Fusiform acanthoxeas densely spined with complex spines in the middle and simple spines at the tips. (F–H) Saturnospongilla Volkmer-Ribeiro, 1976. Gemmuloscleres. (F,G) Birotules with the smooth shaft and entire margins; (H) Acanthoxeas with conical spines. (I,J) Spongilla (Lamarck, 1816). Gemmuloscleres. (I) Acanthoxeas is straight and curved, entirely covered with recurved spines; Microscleres. (J) Acanthoxeas with simple and composed spines. Scale: 10 μm.
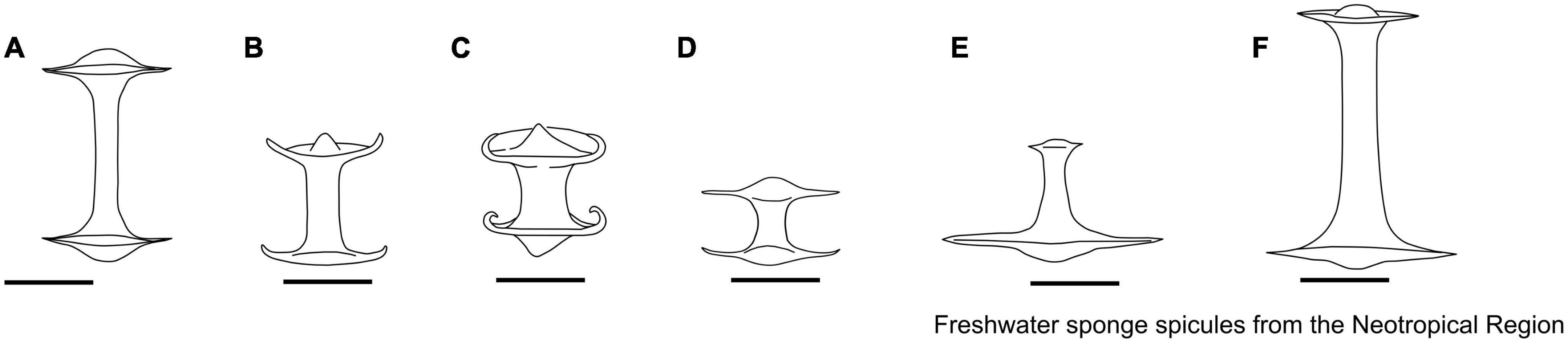
Figure 9. (A–F) Tubella Carter, 1881. Gemmuloscleres. (A–D) Birotules with a smooth shaft, equal rotules, and entire margins; (E) birotule with the smooth shaft, with only one rotule (the other rotule is vestigial); (F) birotules with a smooth shaft, unequal diameters, and entire margins. Scale: 10 μm.
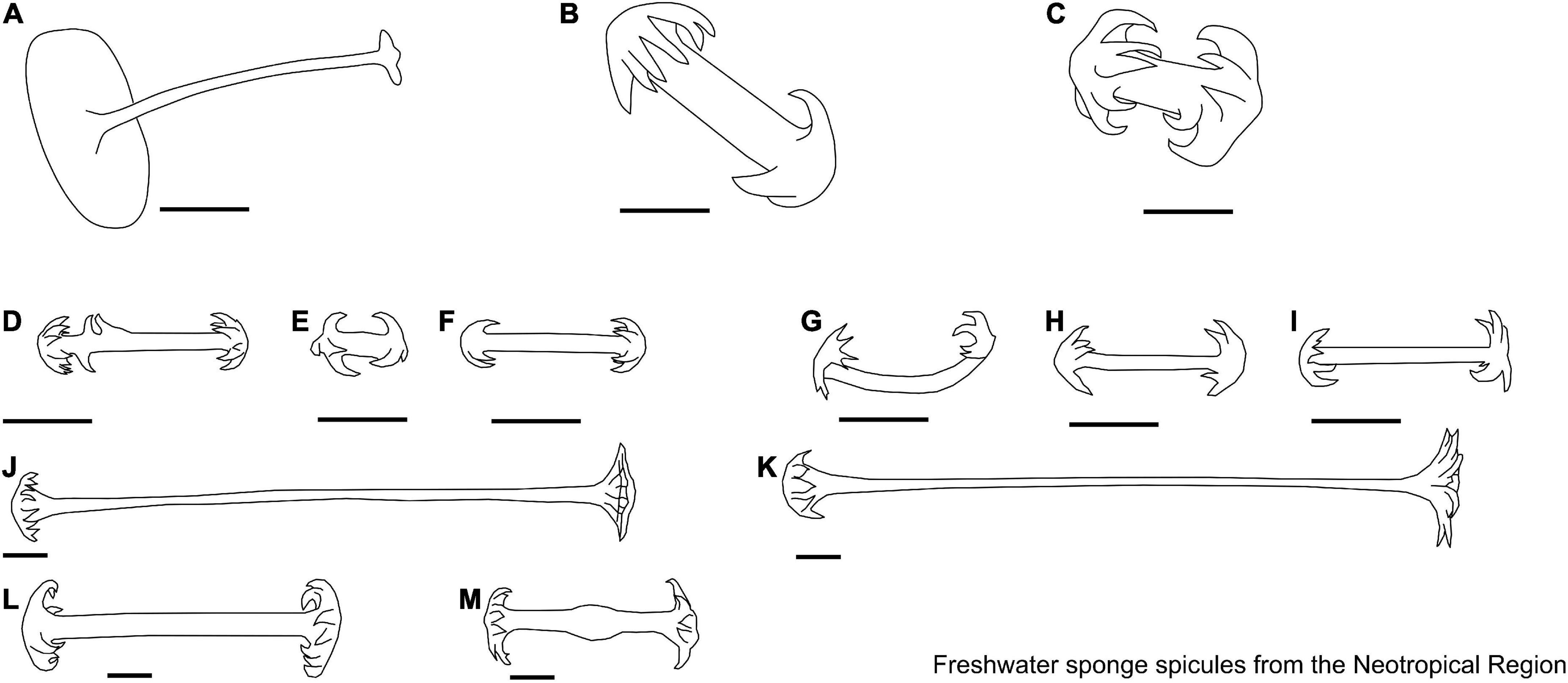
Figure 10. (A–C) Acalle Gray, 1867. Gemmuloscleres. (A) Tubelliform; (B,C) pseudobirotule. (D–M) Corvomeyenia Weltner, 1913. Microscleres. (D–F) Microbirotules with regularly cropped rotules; (G–I) pseudobirotules. Gemmuloscleres. (J,K) Pseudobirotules with long, smooth, delicate shafts, and strongly unbonneted rotules; (L,M) pseudobirotules have small thicknesses, rotules formed, and bearing at their edge six small or quite large, incurved hooks. Scale: 10 μm.
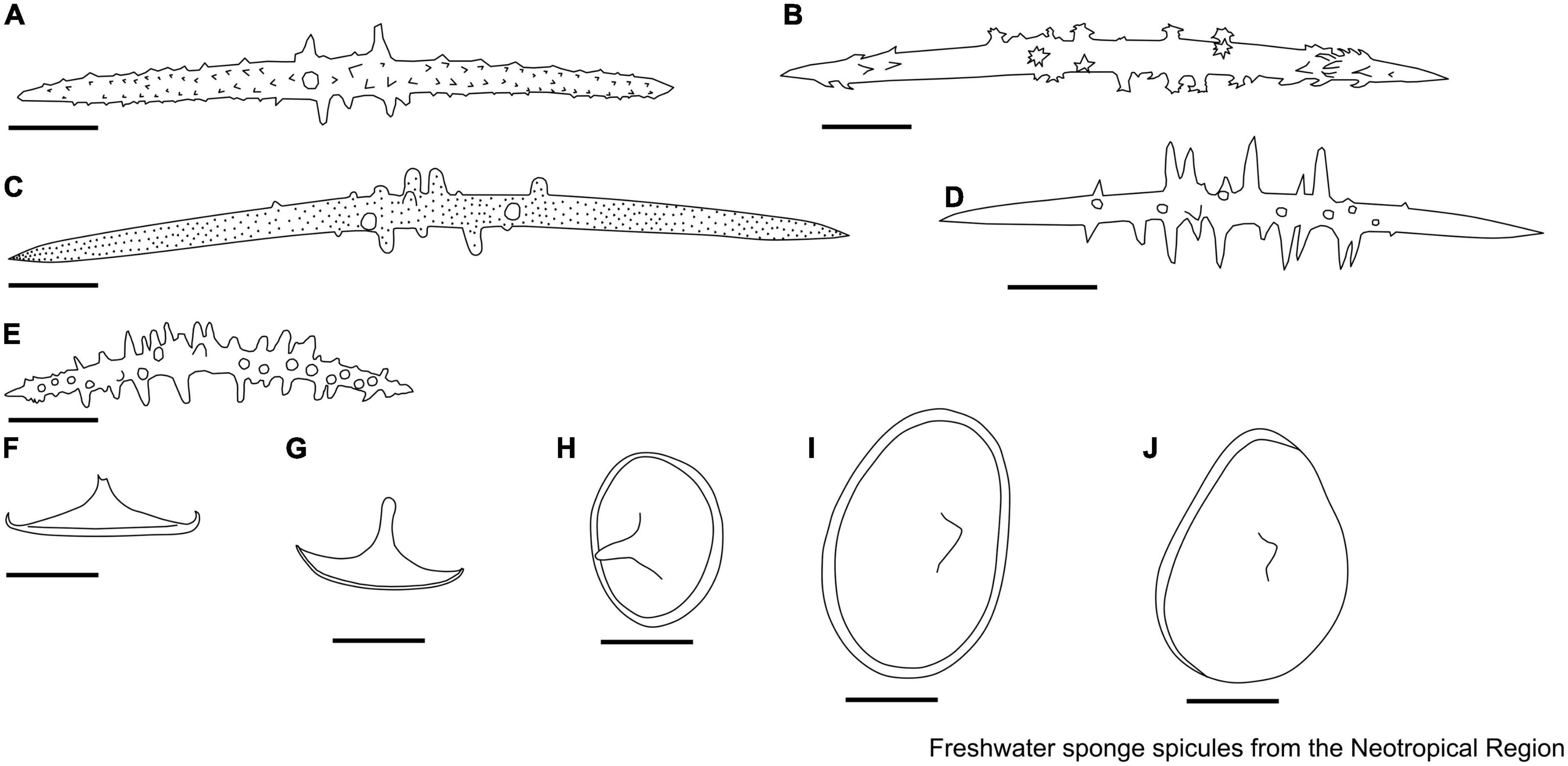
Figure 11. (A–J) Drulia Gray, 1867. Microscleres. (A) Acanthoxeas, covered with spines, with conical spines in the center of the spicule; (B) acanthoxeas with spines flat-ended and harpoon-shaped at extremities; (C) microspined oxea, a few larger spines with rounded extremities irregularly grouped near the central part of spicules; (D) acanthoxeas, with conical spines in the center of the spicule, spines with lanceolate tips; (E) acanthoxeas are covered almost to their extremities with large, straight, conical spines. Gemmuloscleres. (F–J) Parmuliform. Scale: 10 μm.
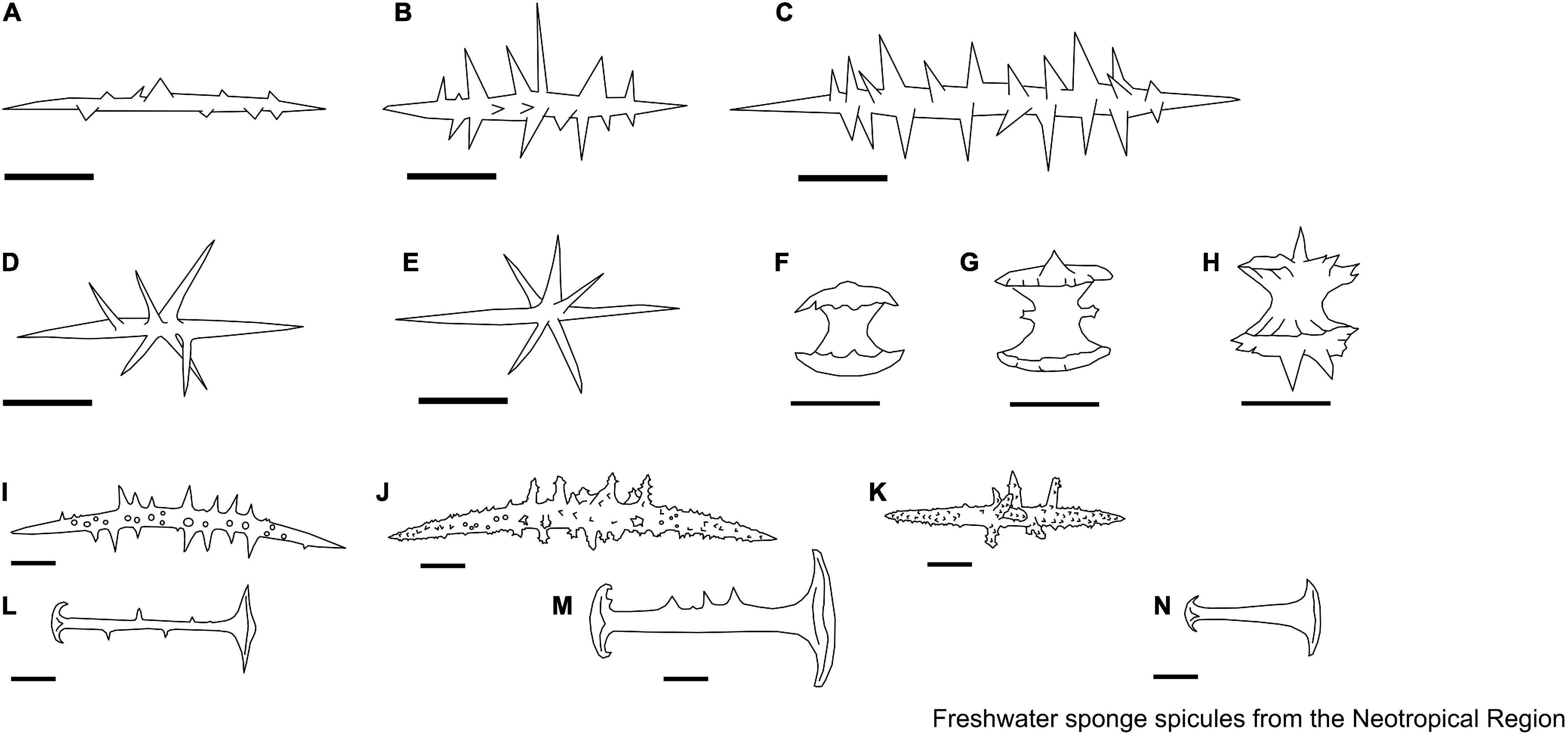
Figure 12. (A–H) Houssayella Bonetto & Ezcurra de Drago, 1966. Microscleres. (A–C) Acanthoxeas; (D,E) aster. Gemmuloscleres. (F–H) Birotules. (I–N) Metania (Gray, 1867). Microscleres. (I–K) Acanthoxeas. Gemmuloscleres. (L–N) Boletiform. Scale: 10 μm.
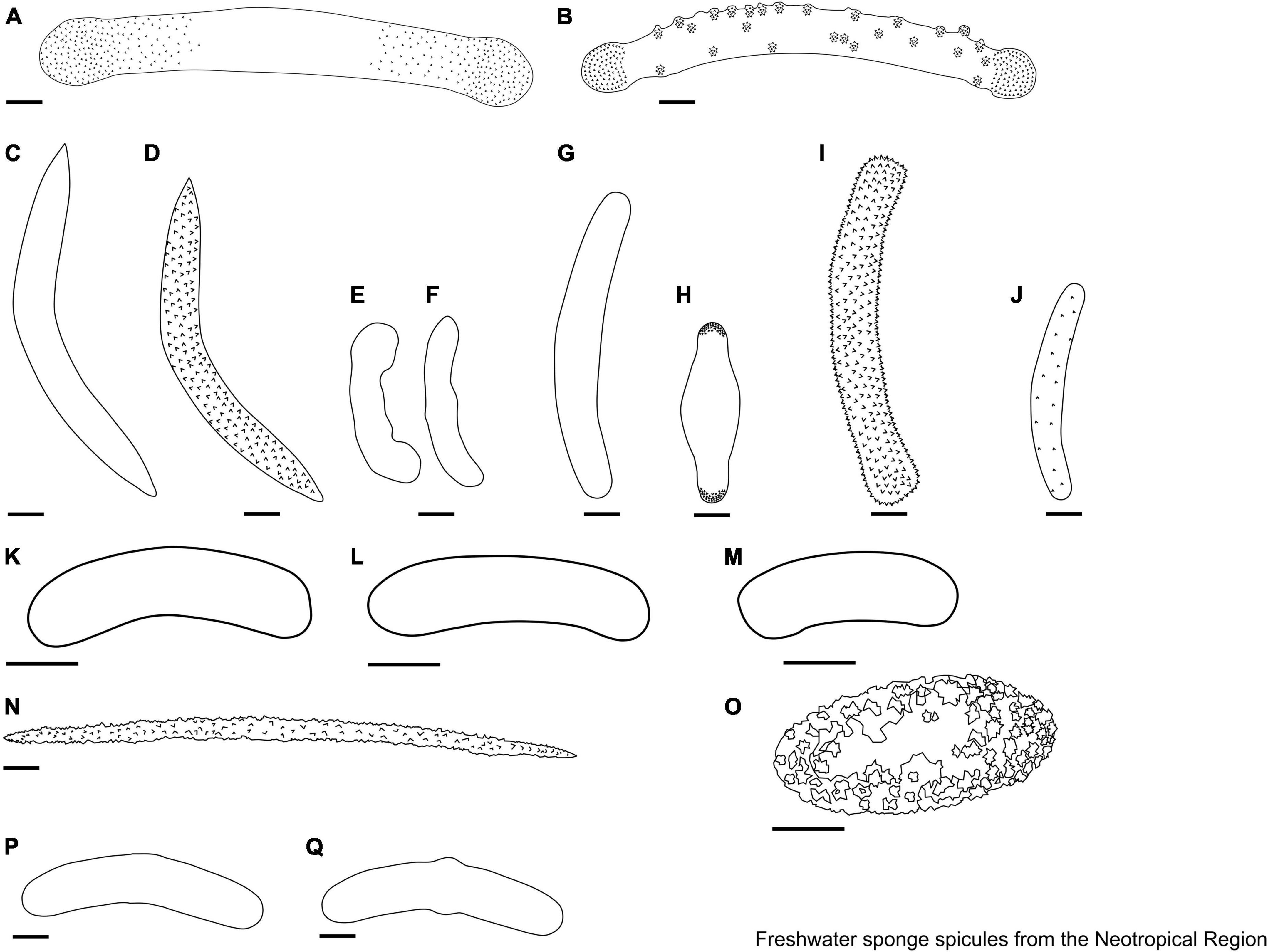
Figure 13. (A,B) Acanthotylotra Volkmer-Ribeiro, Tavares & Fürstenau-Oliveira, 2009. Megascleres. (A) Strongyles microgranulated with inflated tips; (B) Beta megascleres. (C–J) Oncosclera Volkmer-Ribeiro, 1970. Gemmuloscleres. (C) Smooth oxeas; (D) spined oxeas; (E,F) strongyles irregular; (G) strongyles; (H) strongyles with their median portion inflated, with spines concentrated on the extremities; (I) acanthostrongyles; (J) microspined strongyles. (K–M) Potamophloios Brien, 1970 [1969]. Gemmuloscleres. (K–M) Strongyles. (N,O) Sterrastrolepis Volkmer-Ribeiro & Rosa-Barbosa, 1978. Microscleres. (N) Acanthoxea; gemmuloscleres. (O) Sterrasters. (P,Q) Uruguaya Carter, 1881. Gemmuloscleres. (P,Q) Smooth strongyles can be slightly centrotylotus. Scale: 10 μm.
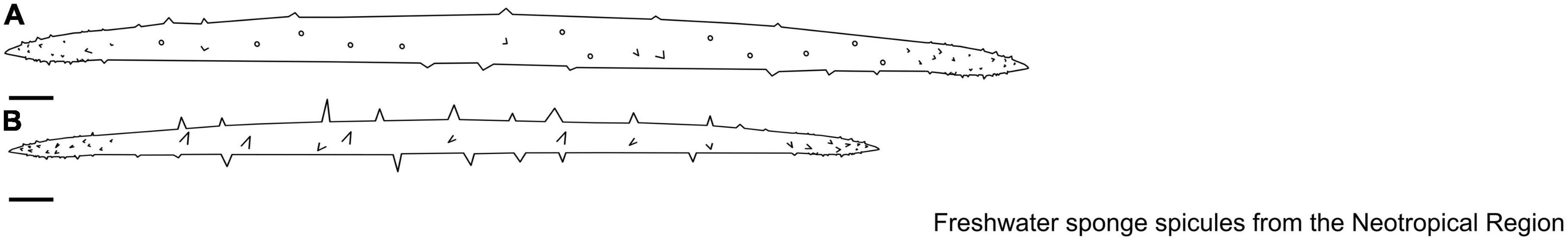
Figure 14. (A,B) Balliviaspongia Boury-Esnault & Volkmer-Ribeiro, 1991. Megascleres. (A,B) Acanthoxeas. Scale: 10 μm.
The skeleton of freshwater sponges consists of a fibrous network of spongin and siliceous spicules (also known as the spicular complement). The megascleres are relatively large and form the main structural component of sponges. In contrast, microscleres are relatively small and often lack a structural function when present (Manconi and Pronzato, 2002). Megascleres can be oxeas (monaxon diactinal spicule pointed at both ends) to strongyles (an isodiametric, diactinal megasclere with rounded end), smooth, spiny, or granular, occasionally with larger tuberculate ornamentations, with sharply pointed, rounded, or occasionally inflated points liked tylote (diactinal megasclere with swelling on each end) (Boury-Esnault and Rützler, 1997; Manconi and Pronzato, 2002; Figure 2). Microscleres can be present or absent, including smooth or spined oxeas or strongyles, aster-like (spicules in which the rays radiate from a central point), and pseudobirotules (birotules that have curved hooks at the ends) spicules. Freshwater sponges can also produce gemmules, a resistant body and asexual propagule produced by Spongillidae, Metaniidae, and Potamolepidae, to survive potentially stressful seasonal environmental conditions (e.g., desiccation and ice-cover). This body is armed with gemmuloscleres and is the most important structure for species identification (Manconi and Pronzato, 2002). Gemmuloscleres encompass various morphologies, ranging from oxeas to strongyles, birotules (a type of spicule with a straight shaft and umbrella-shaped ends), pseudobirotules (does not show a developed rotule but a group of curved hooks radiating from the apices of the shaft), sanidasters (straight spicule having spines at intervals along the entire length). Spines along the shaft are perpendicular to the axis and may or may not be spirally arranged (those at the ends diverge obliquely), parmuliform (spicule with a single rotule supporting a short acute conical stem) to boletiform-tubelliform (characterized by a proximal large, irregularly circular, flat rotule with the entire margin supporting a smooth shaft decreasing in thickness toward the distal apex), and shaped as umbonate (pseudo-rotule with large hooks bearing microspines at their apices) forms (Boury-Esnault and Rützler, 1997; Manconi and Pronzato, 2002).
It is important to differentiate the categories of spicules present in the sediment if they are megascleres, microscleres, and/or gemmuloscleres. It is preferable for the analyst to measure the size range of the spicule morphotype (e.g., length, width, and diameter) and include illustrations or scanning electron microscope (SEM) images to generate the most robust characterizations.
Important remarks regarding some sponge species that should be considered:
• Heterorotula fistula Volkmer-Ribeiro & Costa, 1995, has been found only in spongilites from peat-bog ponds in the southwestern part of Minas Gerais state, Brazil.
• Balliviaspongia wirrmanni Boury-Esnault & Volkmer-Ribeiro, 1991, is only found in Lake Titicaca (Bolivian Altiplano) and may be endemic.
• The genera Balliviaspongia and Acanthotylotra Volkmer-Ribeiro, Tavares & Fürstenau-Oliveira, 2009, have only megascleres spicules, with other structures absent or unknown.
• Similarities between the smooth strongyles gemmuloscleres of some Oncosclera species, Potamophloios, and Uruguaya can result in misidentification; consultation of the primary literature and details about the distributions of these sponges will aid in accurate identification.
The species Arinosaster patriciae Volkmer-Ribeiro, Tavares-Frigo, Ribeiro & Bichuette, 2021, presents megascleres long, robust, smooth, and abruptly pointed oxea. Gemmuloscleres have not been detected. Microscleres fall into three categories; the most abundant are the heavily spined oxea; spines can be straight or curved, and simple or compound (have a bouquet-like structure); rare microscleres include euasters, spherasters, and spheres in more than one size category, as well as rare smooth to spined spirasters. These microscleres were detected only in the preparations containing the dissociated spicules (Volkmer-Ribeiro et al., 2021). The highlighted spicules have a strong similarity to phytoliths, namely, SPHEROID ECHINATE, SPHEROID ORNATE, and SPHEROID PSILATE (Neumann et al., 2019). Phytoliths are structures of silica accumulated by plants and are also preserved in sediments. Their postmortem and their morphologies are directly related to specific botanical families (Piperno, 2006). The SPHEROID ECHINATE morphotype is most often associated with palms (Arecaceae) and Bromeliaceae (Albert et al., 2009; Strömberg et al., 2013); the SPHEROID ORNATE are used as indicators of woody vegetation, and SPHEROID PSILATE have been used as evidence of non-grass plants (Strömberg, 2004, 2005; Strömberg et al., 2018). Concerning Arinosaster patriciae, images of this genus are not included in this guide, as confusion remains regarding the identification of the spicules and their differentiation from phytoliths.
The Neotropical zoogeographic region has the highest species richness. Here, we provide a taxonomic description and information for the genera of freshwater sponges in the Spongillidae, Metaniidae, and Potamolepidae families, and the incertae sedis genus. For each genus, we present a list of species, drawings of gemmuloscleres and/or microscleres that attribute specific taxonomic characteristics, and the appropriate bibliography for reference.
Spongillidae Gray, 1867.
Anheteromeyenia Schöder, 1927
Anheteromeyenia cheguevarai Manconi & Pronzato, 2005
Anheteromeyenia diamantina Calheira & Pinheiro, 2018
Anheteromeyenia ornata (Bonetto & Ezcurra de Drago, 1970)
Anheteromeyenia vitrea Buso, Volkmer-Ribeiro, Pessenda & Machado, 2012
Spicules: Megascleres fall into one category (acanthoxeas or smooth oxeas) or two categories (alpha megascleres, acanthoxeas or smooth oxeas; and beta megascleres, acanthoxeas, and acanthostrongyles). Microscleres are absent. Gemmuloscleres are acanthoxeas, acanthostrongyles, pseudobirotules with a spiny shaft, and bent smooth hooks, acanthoxeas, and grading from long to very short, from stout to slim, acanthostrongyles and acanthoxeas (Figures 3A–E).
Species information: Bonetto and Ezcurra de Drago (1970), Manconi and Pronzato (2005), Buso et al. (2012), Calheira and Pinheiro (2018).
Corvoheteromeyenia Ezcurra de Drago, 1979
Corvoheteromeyenia australis (Bonetto & Ezcurra de Drago, 1966)
Corvoheteromeyenia heterosclera (Ezcurra de Drago, 1974)
Corvoheteromeyenia sanidosclera Pinheiro, Silva & Calheira, 2015
Spicules: Megascleres are oxeas, generally smooth, sometimes irregularly microspined. Microscleres are pseudobirotules with long hooks, pseudobirotule with short hooks, and/or acanthoxeas varying in shape and size, with a variable number of spines, simple (straight or curved) and/or compound (Figures 3F–L). Gemmuloscleres are birotules with rotules microspines on their margins, convex to flat or sanidasters (Figures 3M–S).
Species information: Bonetto and Ezcurra de Drago (1966), Ezcurra de Drago (1974b), Pinheiro et al. (2015b), Calheira and Pinheiro (2016).
Corvospongilla Annandale, 1911
Corvospongilla seckti Bonetto & Ezcurra de Drago, 1966
Spicules: Megascleres are predominantly microspined strongyles, and rarely microspined strongyles. Microscleres are pseudobirotules with four to seven radial hooks on each pseudorotule and a smooth shaft (Figure 3Q). Gemmuloscleres are microspined strongyles and acanthoxeas (Figures 3R, S).
Species information: Bonetto and Ezcurra de Drago (1966), De Rosa-Barbosa (1988), Manconi and Pronzato (2002), Pinheiro et al. (2013).
Dosilia Gray, 1867
Dosilia palmeri (Potts, 1885)
Dosilia pydanieli (Volkmer-Ribeiro, 1992)
Spicules: Megascleres are smooth oxeas or with microspines. Microscleres range from aster from simple acerates with one or more long divergent branch spines to true “euasters” with spiny rays (Figures 4A–C). Gemmuloscleres are birotules, with straight cylindrical shafts and large spines (Figures 4D, E). Rotules vary from flat to slightly umbonate with margins bearing numerous small blunt and recurved teeth or spines.
Species information: Potts (1885), Penney and Racek (1968), Volkmer-Ribeiro (1992), Manconi and Pronzato (2002), Cândido et al. (2010).
Ephydatia Lamouroux, 1816
Ephydatia caatingae Nicacio & Pinheiro, 2015
Ephydatia chileana Pisera & Sáez, 2003†
Ephydatia facunda Weltner, 1895
Ephydatia fluviatilis (Linnaeus, 1759)
Ephydatia robusta (Potts, 1888)
Spicules: Megascleres are oxeas that range from smooth to microspined. Microscleres are absent. Gemmuloscleres are birotules with smooth or spined shafts (simple and compound spines), with secondary spines (Figures 4F–L). Flat rotules with incised irregular margins may be present.
Species information: Penney and Racek (1968), Ezcurra de Drago (1975a), De Rosa-Barbosa (1979), Ricciardi and Reiswig (1993), Manconi and Pronzato (2002), Pisera and Sáez (2003), Pinheiro et al. (2004), Nicacio and Pinheiro (2015).
Eunapius Gray, 1867
Eunapius carteri (Bowerbank, 1863)
Eunapius fragilis (Leidy, 1851)
Eunapius igloviformis (Potts, 1884)
Spicules: Megascleres are smooth or spined oxeas. Microscleres absent. Gemmuloscleres are smooth oxeas or acanthoxeas to acanthostrongyles (Figures 5A–C).
Species information: Penney and Racek (1968), Ezcurra de Drago (1974a), Manconi and Pronzato (2002), Nicacio and Pinheiro (2015).
Heteromeyenia Potts, 1881
Heteromeyenia barlettai Pinheiro, Calheira & Hajdu, 2015
Heteromeyenia cristalina Batista, Volkmer-Ribeiro & Melão, 2007
Heteromeyenia horsti Ezcurra de Drago, 1988
Heteromeyenia insignis Weltner, 1895
Spicules: Megascleres are acanthoxeas. Microscleres are acanthoxeas, with spines that can be straight or curved, and simple or compound with a bouquet-like structure (Figures 5D–F). Gemmuloscleres fall into one or two categories: pseudobirotules (with teeth projecting from the center of the pseudorotule and can be simple or anastomosing) (Figure 5G), and birotules, and shafts with conical spines (simple or compound) (Figures 5H, I). Rotules are smooth or covered in microspines, circular, convex, and identical, with microspines on their margins, and the margins can be serrated. Heteromeyenia horsti is a possible synonym of H. insignis (Pinheiro et al., 2015a; Calheira et al., 2020).
Species information: Manconi and Pronzato (2002), Batista et al. (2007), Pinheiro et al. (2015a).
Heterorotula Penney & Racek, 1968
Heterorotula fistula Volkmer-Ribeiro & Motta, 1995
Spicules: Megascleres oxeas are microspined except at the ends. Microscleres are absent. Gemmuloscleres birotules fall into two categories (long and short), with rotules flat and microspined, one always larger than the other (Figures 6A–F). Birotules have long margins with serrations, and birotules exhibit short margins that range from serrated to toothed.
Species information: Volkmer-Ribeiro and Motta (1995), Manconi and Pronzato (2002).
Pottsiela Volkmer-Ribeiro, Machado, Fürstenau-Oliveira & Soares, 2010
Pottsiela pesae Volkmer-Ribeiro, Machado, Fürstenau-Oliveira & Soares, 2010
Pottsiela spoliata (Volkmer-Ribeiro & Maciel, 1983)
Spicules: Megascleres are smooth to spined oxeas. Microscleres are acanthoxeas, microspine-bearing along their length, with conical projections that are also microspined (Figures 6G–K). Gemmuloscleres are absent. Gemmules are missing pneumatic layers and gemmuloscleres, with the megascleres forming cages to contain the gemmules, or else they irregularly adhere to the gemmular wall.
Species information: Volkmer-Ribeiro and Maciel (1983), Manconi and Pronzato (2002), Volkmer-Ribeiro et al. (2010c).
Racekiela Bass & Volkmer-Ribeiro, 1998
Racekiela andina Hernandez & Barreat, 2017
Racekiela cavernicola Volkmer-Ribeiro, Bichuette & Machado, 2010
Racekiela sheilae (Volkmer-Ribeiro, Rosa-Barbosa & Tavares, 1988)
Spicules: Megascleres are acanthoxeas. Microscleres are absent. Gemmuloscleres are present in two types: birotules (short thin or robust shafts with smooth or with few spines) and pseudobirotules (long and spiny shafts) (Figure 7). Birotules can have rotules with small slightly umbonate to flat and deeply cut into several microspined long rays.
Species information: Volkmer-Ribeiro et al. (1988), Manconi and Pronzato (2002), Volkmer-Ribeiro et al. (2010a), Hernández and Barreat (2017), Gómez et al. (2019).
Radiospongilla Penney & Racek, 1968
Radiospongilla amazonensis Volkmer-Ribeiro & Maciel, 1983
Radiospongilla crateriformis (Potts, 1882)
Radiospongilla inesi Nicacio & Pinheiro, 2011
Spicules: Megascleres are spined oxeas, rarely spined strongyles. Microscleres are absent. Gemmuloscleres are acanthostrongyles with tips bearing apical spines, where spines are concentrated at the tips; they are curved, forming hooks directed toward the center of the spicule, or are straight and sharp, which can form small umbonate rotules (pseudorotules). Pseudobirotules with spiny shafts and bent long spines are at the apices (Figures 8A–C).
Species information: Penney and Racek (1968), Ezcurra de Drago (1975b), Volkmer-Ribeiro and Maciel (1983), Ricciardi and Reiswig (1993), Bass and Volkmer-Ribeiro (1998), Manconi and Pronzato (2002), Volkmer-Ribeiro and Machado (2009), Nicacio et al. (2011).
Rosulaspongilla Sokolova, Palatov, Masuda & Itskovich, 2021
Rosulaspongilla alba (Carter, 1849)
Spicules: Megascleres are predominantly smooth oxeas. Microscleres are fusiform acanthoxeas densely spined with complex spines in the middle and simple spines at the tips (Figure 8E). Gemmuloscleres are acanthoxeas with large, curved spines more thickly accumulated at the tips and often form mace-shaped structures (Figure 8D).
Species information: Penney and Racek (1968), Manconi and Pronzato (2005), Volkmer-Ribeiro and Machado (2009), Pinheiro et al. (2015c), Sokolova et al. (2021).
Remark: Sokolova et al. (2021) demonstrated from genetic analysis and confirmed with morphological data that the part of the genus Spongilla, “S. alba group” should be separated into another genus, Rosulaspongilla, which reveals a new group of brackish-water sponges. Thus, Spongilla alba from the Neotropical region now is accepted as Rosulaspongilla alba (junior synonym).
Saturnospongilla Volkmer-Ribeiro, 1976
Saturnospongilla carvalhoi Volkmer, 1976
Spicules: Megascleres are smooth oxeas. Microscleres are absent. Gemmuloscleres are acanthoxeas with conical spines (Figure 8H) and short birotule with smooth shafts (Figures 8F, G). Rotule with smooth margins of the same size (predominant) or different size (rare) perforated on both sides by the shaft end.
Species information: Volkmer-Ribeiro (1976), Manconi and Pronzato (2002).
Spongilla Lamarck, 1816
Spongilla cenota Penney & Racek, 1968
Spicules: Megascleres are stout, smooth oxeas, with abruptly pointed extremities. Microscleres are slender, slightly curved, spiny oxeas, all spines capped by a rosette of microspines (Figure 8J). Gemmuloscleres are short, stout, spiny oxeas, straight to slightly curved, covered by large spines curved toward the middle part of the spicule, and their extremities split into a few smaller spines (Figure 8I).
Species information: Penney and Racek (1968), Manconi and Pronzato (2002), Volkmer-Ribeiro and Machado (2009).
Tubella Carter, 1881
Tubella amazonica (Weltner, 1895)
Tubella delicata Bonetto & Ezcurra de Drago, 1967
Tubella gregaria (Bowerbank, 1863)
Tubella horrida Weltner, 1893
Tubella lanzamirandai Bonetto & Ezcurra de Drago, 1964
Tubella leidii (Bowerbank, 1863)
Tubella minuta (Potts, 1887)
Tubella paulula (Bowerbank, 1863)
Tubella pennsylvanica (Potts, 1882)
Tubella repens (Hinde, 1888)
Tubella variabilis Bonetto & Ezcurra de Drago, 1973
Spicules: Megascleres are spined or smooth strongyles and/or oxeas. Microscleres are absent. Gemmuloscleres are birotules with a short stout smooth shaft (Figure 9). The rotules can be with equal or unequal diameters, or only with only one rotule (the other rotule is vestigial) and are circular with entire margins.
Species information: Bonetto and Ezcurra de Drago (1964), Bonetto and Ezcurra de Drago (1965), Penney and Racek (1968), Bonetto and Ezcurra de Drago (1973b), Volkmer-Ribeiro (1973), Volkmer-Ribeiro and De Rosa-Barbosa (1985), Batista et al. (2007), Manconi and Pronzato (2002), Nicacio and Pinheiro (2015).
Metaniidae Volkmer-Ribeiro, 1986.
Acalle Gray, 1867
Acalle recurvata (Bowerbank, 1863)
Spicules: Megascleres are smooth or microspined strongyles to smooth oxeas. Microscleres are absent. Gemmuloscleres present as two types: tubelliform (Figure 10A) and pseudobirotules (Figures 10B, C). Tubelliforms have a proximal large irregularly circular flat rotule with an entire margin supporting a smooth shaft, decreasing in thickness toward the distal end and shaped as an umbonate knob-like rotule with few teeth. Pseudobirotules have cylindrical shafts and hooks of umbonate pseudo-rotules stout and notably recurved bearing microspines at their apices.
Species information: Volkmer-Ribeiro and De Rosa-Barbosa (1972), Manconi and Pronzato (2002).
Corvomeyenia Weltner, 1913
Corvomeyenia epilithosa Volkmer-Ribeiro, de Rosa-Barbosa & Machado, 2005
Corvomeyenia thumi (Traxler, 1895)
Spicules: Megascleres are oxeas, generally smooth. Microscleres fall into one or two categories: in the first case with microbirotules straight, with smooth or spiny axis, with regularly cropped rotules, and in the second case with strongly curved and reduced pseudobirotule-like rotules (Figures 10D–I). Gemmuloscleres with one or two categories of pseudobirotules (Figures 10J–M). Pseudobirotules with long, smooth, delicate shafts, and strongly umbonate rotules, are dissimilar in size and can be cut out in a variable number of hooks or teeth, and these are with irregular dispositions. Pseudobirotules are small thick, conspicuously umbonate, usually well-formed, and bearing at their edge six small or quite large, incurved hooks. However, this rotule may be reduced to a knob with a few irregularly formed hooks or spines.
Species information: Manconi and Pronzato (2002), Volkmer-Ribeiro et al. (2005).
Drulia Gray, 1867
Drulia brownii (Bowerbank, 1863)
Drulia conifera Bonetto & Ezcurra de Drago, 1973
Drulia cristata (Weltner, 1895)
Drulia cristinae Volkmer-Ribeiro, Drago, Machado & Sabaj, 2017
Drulia ctenosclera Volkmer-Ribeiro & Mothes de Moraes, 1981
Drulia uruguayensis Bonetto & Ezcurra de Drago, 1968
Spicules: Megascleres fall in two size classes: alpha megascleres range from smooth straight to curved oxea with abruptly pointed extremities to nanospined and conspicuously curved strongyles, and beta megascleres, when present, usually are curved oxea, two-thirds of the size of the alpha megascleres, and may bear sparse spines. Microscleres are minute, straight to curved, slender to thick spiny oxea with harpoon-shaped extremities and the central portion bearing few to several larger spines; or they are straight, uniformly nanospined oxea (Figures 11A–E). Gemmuloscleres are minute, flat, or umbonate parmuliform spicules with circular or ellipsoid outlines; both faces have a smooth or outer face with a central conical or rounded projection, or a shallow crest and thin or thick, incurved borders (Figures 11F–J).
Species information: Bowerbank (1863), Weltner (1895), Bonetto and Ezcurra de Drago (1968a), Bonetto and Ezcurra de Drago (1973a), Volkmer-Ribeiro and Mothes de Moraes (1981), Volkmer-Ribeiro and Tavares (1995), Manconi and Pronzato (2002), Volkmer-Ribeiro et al. (2017).
Houssayella Bonetto & Ezcurra de Drago, 1966
Houssayella iguazuensis Bonetto & Ezcurra de Drago, 1966
Spicules: Megascleres range from oxeas to strongyles that range from densely spined to smooth, particularly at the apices. Microscleres range from acanthostrongyles to oxeas and acanthoxeas, with long perpendicular or slanting spines to aster-like shaped spicules (Figures 12A–E). Gemmuloscleres are stout microspined birotulates (Figures 12F–H).
Species information: Bonetto and Ezcurra de Drago (1966), Manconi and Pronzato (2002).
Remark: Bonetto and Ezcurra de Drago (1966) erected Houssayella and suggested this genus is closely related to the Dosilia Gray, 1967, since both present aster-like microscleres. Volkmer-Ribeiro and Rützler (1997) erected Pachyrotula and suggested Houssayella and Heterorotula closely related to a new genus, due to the similar spicular set.
Metania Gray, 1867
Metania fittkaui Volkmer-Ribeiro, 1979
Metania kiliani Volkmer-Ribeiro & Costa, 1992
Metania reticulata (Bowerbank, 1863)
Metania spinata (Carter, 1881)
Metania subtilis Volkmer-Ribeiro, 1979
Spicules: Megascleres fall into two distinct classes: alpha megascleres range from smooth, stout oxea to strongyles, and beta megascleres, when present, range from spiny oxea to strongyles. Microscleres fall into two distinct classes: spiny minute oxea displaying large spines in the middle, with simple and compound spines, sometimes grouped in rosettes, and can show a microgranulation at the extremities (Figures 12I–K). The large spines have lanceolate endings. Gemmuloscleres are boletiform with shafts long to short, smooth to spined, with a variable number of spines, and a collar of spines under the lower rotule (Figures 12L–N). Lower rotules are large, stout, and polygonal, with curved, undulated margins. Upper rotules are knob-like, smooth, or with a few recurved, irregularly placed spines or hooks, or approaching a true rotule with marginal incurved spines.
Species information: Bowerbank (1863), Carter (1881), Volkmer-Ribeiro (1979), Volkmer-Ribeiro (1984), Volkmer-Ribeiro and Costa (1992), Manconi and Pronzato (2002), Castello-Branco et al. (2015).
Potamolepidae Brien, 1967.
Acanthotylotra Volkmer-Ribeiro, Tavares & Fürstenau-Oliveira, 2009
Acanthotylotra alvarengai Volkmer-Ribeiro, Tavares & Fürstenau-Oliveira, 2009
Acanthotylotra xingu Pinheiro, Martins & Calheira, 2020
Spicules: Megascleres fall into two categories: alpha megascleres are acanthotylostrongyles, with spines grouped in small spots or forming half rings on the convex section of the spicule, the tylote extremities entirely covered with minute spines, or they can be strongyles microgranulated with inflated tips (Figure 13A). Beta megascleres are acanthostrongyles, with microspined tubercules along the spicule length except at the extremities, which are invariably covered with minute spines (Figure 13B). Microscleres are unknown or absent. Gemmules are unknown.
Species information: Volkmer-Ribeiro et al. (2009), Pinheiro et al. (2020).
Oncosclera Volkmer-Ribeiro, 1970
Oncosclera atrata (Bonetto & Ezcurra de Drago, 1970)
Oncosclera intermedia (Bonetto & Ezcurra de Drago, 1973)
Oncosclera jewelli (Volkmer, 1963)
Oncosclera navicella (Carter, 1881)
Oncosclera petricola (Bonetto & Ezcurra de Drago, 1967)
Oncosclera ponsi (Bonetto & Ezcurra de Drago, 1968)
Oncosclera rosariae Tavares-Frigo, Volkmer-Ribeiro, Oliveira & Machado, 2015
Oncosclera schubarti (Bonetto & Ezcurra de Drago, 1967)
Oncosclera spinifera (Bonetto & Ezcurra de Drago, 1973
Oncosclera stolonifera (Bonetto & Ezcurra de Drago, 1973)
Oncosclera tonollii (Bonetto & Ezcurra de Drago, 1968)
Spicules: Megascleres can be microspined, smooth or spines oxeas, and/or strongyles microgranules, smooth or covered by spines (Figures 13C–J). Microscleres are absent. Gemmuloscleres are highly variable from smooth oxea curved in the shape of a boomerang to microspined oxeas with variable shapes (curved, with or without pointed tips, folded, completely microspined irregularly spherical). They could be smooth strongyles that may present with an inflated medial portion to spined strongyles, with spines concentrated mainly on the extremities. Irregular strongyles also can be present; spiny strongyles resembling peanut shells. Spines are more abundant at the tips. A bumped middle region is present in the gemmuloscleres so that quite cylindrical ones are rare. Spherical forms are rare.
Species information: Carter (1881), Volkmer-Ribeiro (1963), Bonetto and Ezcurra de Drago (1967), Bonetto and Ezcurra de Drago (1968b), Bonetto and Ezcurra de Drago (1970), Volkmer-Ribeiro (1970), Bonetto and Ezcurra de Drago (1973a), Tavares and Volkmer-Ribeiro (1997), Manconi and Pronzato (2002), Tavares-Frigo et al. (2015).
Potamophloios Brien, 1970 [1969].
Potamophloios guairensis Volkmer-Ribeiro, Parolin, Fürstenau-Oliveira & Menezes, 2010
Spicules: The neotropical species has megascleres that are large, stout, smooth, straight to curved or angular strongyles, which initiate as slim sharply pointed oxea. Microscleres are absent. Gemmuloscleres vary from straight to curved, smooth, small, stout strongyles, with smaller examples reaching a spheric shape (Figures 13K–M).
Species information: Manconi and Pronzato (2002), Volkmer-Ribeiro et al. (2010b).
Sterrastrolepis Volkmer-Ribeiro & Rosa-Barbosa, 1978
Sterrastrolepis brasiliensis Volkmer-Ribeiro & Rosa-Barbosa, 1978
Spicules: Megascleres are stout and slightly curved, from smooth to uniformly granulated strongyles with inflated tips. Rare oxeas with blunt tips are also present. Microscleres are slender, slightly curved acanthoxeas (reported as tornotes in the original description) entirely ornamented by tubercules or spines with microspines arranged in rosettes (Figure 13N). Gemmuloscleres are spherical to sub-spherical, most often ellipsoid sterrasters, with an irregular surface apart from a unilateral smooth area (Figure 13O).
Species information: Volkmer-Ribeiro and De Rosa-Barbosa (1978), Manconi and Pronzato (2002), Volkmer-Ribeiro and Parolin (2005).
Uruguaya Carter, 1881
Uruguaya corallioides (Bowerbank, 1863)
Spicules: Megascleres range from stout curved strongyles with granulated surfaces and a very thin axial canal, to stout smooth oxeas. Microscleres are absent. Gemmuloscleres are smooth curved strongyles, spherules of silica, and deformed strongyles are also present (Figures 13P, Q).
Species information: Bonetto and Ezcurra de Drago (1964), Bonetto and Ezcurra De Drago (1969), Manconi and Pronzato (2002), Nicacio and Pinheiro (2015).
Spongillidae incertae sedis (Manconi & Pronzato, 2002).
Balliviaspongia Boury-Esnault & Volkmer-Ribeiro, 1991
Balliviaspongia wirrmanni Boury-Esnault & Volkmer-Ribeiro, 1991
Spicules: Megascleres are oxeas ranging from slender to stout, from straight to slightly curved, from smooth to spined, with acerate tips. Microscleres and gemmules are absent (Figures 14A, B).
Species information: Boury-Esnault and Volkmer-Ribeiro (1991), Manconi and Pronzato (2002).
Sponge assemblages have considerable value for deciphering the influence of riverine connectivity with floodplain, wetland, and/or lake environments; reconstruction of flood pulse paleohydrology is enhanced by the inclusion of this fossil record, as has been demonstrated in the Pantanal wetlands (McGlue et al., 2012; Rasbold et al., 2019). Given the vast tracts of lowland floodplains in the Neotropics, considerable opportunity exists to expand paleoecological studies using sponge fossils. Sponge paleoecology is scarce in larger lakes, such as those in tectonically formed basins on the Altiplano or in Central America, despite reliable indications of the presence of these animals in nearshore environments (Binford, 1982; Martens and Harrison, 1993; Erpenbeck et al., 2020). The expansion of scientific drilling technology for sampling ancient sediments opens opportunities for learning about ecological history in deep time, and uncovering the role sponges may have played in the evolution of lake and river environments (e.g., Sawakuchi et al., 2015).
This study provides systematic guidelines for sediment sample preparation and identification of sponges in continental sediments from the Neotropics. Sponges constitute important components of benthic life in many inland Neotropical waters, and their siliceous skeletons are frequently well preserved as fossils in Quaternary and older sediments. Our recommendations stem from combined several decades of collaborative research that has resulted in the development of a set of practices for sponge paleoecology, which has led to insights into aquatic ecosystem response to a hydroclimate change in the Neotropics. Freshwater sponge taxonomy as a discipline is not widespread, and additional autecological research will help improve and extend the utility of sponge fossils for paleoecological purposes.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
GR, LC, LD-L, and MM contributed to the conception and design of the study and wrote sections of the manuscript. GR and LC organized the database and wrote the first draft of the manuscript. LP and UP contributed to the supervision. All authors contributed to manuscript revision, read, and approved the submitted version.
GR was supported by the São Paulo Research Foundation (FAPESP) (Grant No. 2020/07726-0). LC was supported by the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (BFP-FACEPE 0031-2.04/21).
We thank all researchers and taxonomists on freshwater sponges, especially Dr. Cecilia Volkmer-Ribeiro and Dr. Mauro Parolin for the dissemination of these studies in Brazil.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor was currently organizing a Research Topic with the author MM.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1067432/full#supplementary-material
Albert, J. S., Tagliacollo, V. A., and Dagosta, F. (2020). Diversification of neotropical freshwater fishes. Annu. Rev. Ecol. Evol. Syst. 51, 27–53. doi: 10.1146/annurev-ecolsys-011620-031032
Albert, R. M., Bamford, M. K., and Cabanes, D. (2009). Palaeoecological significance of palms at Olduvai Gorge, Tanzania, based on phytolith remains. Quaternary Int. 193, 41–48. doi: 10.1016/j.quaint.2007.06.008
Antonelli, A., and Sanmartín, I. (2011). Why are there so many plant species in the Neotropics? Taxon 60, 403–414. doi: 10.1002/tax.602010
Bass, D., and Volkmer-Ribeiro, C. (1998). Radiospongilla crateriforms (Porifera, Spongillidae) in the West Indies and taxonomic notes. Iheringia Sér. Zool 85, 123–128.
Batista, T. C., Volkmer-Ribeiro, C., and Melão, M. D. G. G. (2007). Espongofauna da área de proteção ambiental meandros do rio araguaia (GO, MT, TO), Brasil, com descrição de Heteromeyenia cristalina sp. nov.(Porifera, Demospongiae). Rev. Bras. Zool. 24, 608–630. doi: 10.1590/S0101-81752007000300013
Battarbee, R. W. (1986). “Diatom analysis,” in Handbook of holocene palaeoecology and palaeohydrology, ed. B. E. Berglund (Chichester: John Wiley & Sons), 527–570.
Binford, M. W. (1982). Ecological history of Lake Valencia, Venezuela: Interpretation of animal microfossils and some chemical, physical, and geological features. Ecol. Monogr. 52, 307–333. doi: 10.2307/2937333
Bonetto, A. A., and Ezcurra de Drago, I. (1967). Esponjas del noreste argentino. Acta Zool. Lilloana 23, 331–347.
Bonetto, A., and Ezcurra de Drago, I. (1964). Nuevas esponjas de agua dulce de la República Argentina. Physis 24, 329–336.
Bonetto, A., and Ezcurra de Drago, I. (1965). El género Trochospongilla vejdowski en el alto paraná argentino (Porifera, Spongillidae). Physis 25, 95–98.
Bonetto, A., and Ezcurra de Drago, I. (1966). Nuevos aportes al conocimiento de las esponjas argentinas. Physis 26, 129–140.
Bonetto, A., and Ezcurra de Drago, I. (1968a). El género Drulia gray en el río uruguay (Porifera, Spongillidae). Physis 28, 211–216.
Bonetto, A., and Ezcurra de Drago, I. (1968b). El genero Spongilla lamarck en el rio uruguay (Porifera, Spongillidae). Physis 27, 429–436.
Bonetto, A., and Ezcurra De Drago, I. (1969). Notas sistemáticas sobre el género Uruguaya carter (Porifera, Spongillidae). Physis 28, 351–357.
Bonetto, A., and Ezcurra de Drago, I. (1970). Esponjas de los afluentes del alto parana en la Provincia de Misiones. Acta Zool. Lilloana 27, 37–61.
Bonetto, A., and Ezcurra de Drago, I. (1973a). Aportes al conocimiento de las esponjas del Orinoco. Physis 28, 211–216.
Bonetto, A., and Ezcurra de Drago, I. (1973b). Las esponjas del género Trochospongilla vejdovsky en aguas argentinas. Physis 32, 13–18.
Boury-Esnault, N., and Rützler, K. (1997). Thesaurus of sponge morphology. Smithson. Contrib. Zool. 596, 1–55. doi: 10.5479/si.00810282.596
Boury-Esnault, N., and Volkmer-Ribeiro, C. (1991). “Las esponjas,” in Lake titicaca: A synthesis of limnological knowledge, eds C. Dejoux and A. Iltis (Dordrecht: Kluwer Academic), 297–301.
Bush, M. B., Correa-Metrio, A., McMichael, C. H., Sully, S., Shadik, C. R., Valencia, B. G., et al. (2016). A 6900-year history of landscape modification by humans in lowland Amazonia. Quaternary Sci. Rev. 141, 52–64. doi: 10.1016/j.quascirev.2016.03.022
Buso, A. A. Jr., Volkmer-Ribeiro, C., Pessenda, L. C. R., and Machado, V. (2012). Anheteromeyenia vitrea (Porifera: Demospongiae) new species of continental sponge in Brazil. Neotrop. Biol. Conserv. 7, 148–157. doi: 10.4013/nbc.2012.73.01
Calheira, L., and Pinheiro, U. (2016). Corvoheteromeyenia Ezcurra de Drago, 1979 (Spongillidae, Porifera): Genus review with proposal of a neotype of Corvoheteromeyenia heterosclera (Ezcurra de Drago, 1974). Zootaxa 4126, 351–374. doi: 10.11646/zootaxa.4126.3.3
Calheira, L., and Pinheiro, U. (2018). A new species of Anheteromeyenia (Porifera, Demospongiae) with an emended diagnosis of the genus. Zootaxa 4378, 129–136. doi: 10.11646/zootaxa.4378.1.9
Calheira, L., Santos, P. J., and Pinheiro, U. (2020). Hatchability of gemmules of two neotropical freshwater sponges (Porifera: Spongillida). Iheringia Sér. Zool. 110, 1–5. doi: 10.1590/1678-4766e2020001
Cândido, J. L., Volkmer-Ribeiro, C., and Fürstenau-Oliveira, K. (2010). Dosilia (Porifera, Demospongiae) redefined. Iheringia Sér. Zool. 100, 425–448. doi: 10.1590/S0073-47212010000400017
Carter, H. (1881). History and classification of the known species of Spongilla. J. Nat. Hist. 7, 77–107. doi: 10.1080/00222938109459483
Castello-Branco, C., Calor, A. R., and Menegola, C. (2015). Microscleres and gemmoscleres as phylogenetic signals in Spongillida: Phylogeny and biogeography of the genus Metania Gray, 1867 (Porifera, Metaniidae). Invertebrate Syst. 29, 369–385. doi: 10.1071/IS14060
De Rosa-Barbosa, R. (1979). Redescriçao do tipo de Ephydatia facunda Weltner, 1895 (Porifera-Spongillidae). Iheringia Sér. Zool. 54, 27–34.
De Rosa-Barbosa, R. (1988). Corvospongilla volkmeri sp. n. e registro de Corvospongilla seckti Bonetto & Ezcurra de Drago, 1966 no Brasil (Porifera, Spongillidae). Iheringia Sér. Zool. 67, 109–122.
De Voogd, N., Alvarez, B., Boury-Esnault, N., Carballo, J., Cárdenas, P., Díaz, M.-C., et al. (2022). World Porifera Database [Online]. Available online at: https://www.marinespecies.org/porifera (accessed July 1, 2022).
Docio, L., Parolin, M., and Pinheiro, U. (2021). A contribution to adequate use of freshwater sponges as a proxy in paleoenvironmental studies. Zootaxa 4915, 506–528. doi: 10.11646/zootaxa.4915.4.3
Erpenbeck, D., Galitz, A., Wörheide, G., Albrecht, C., Pronzato, R., and Manconi, R. (2020). Having the balls to colonize–The Ephydatia fluviatilis group and the origin of (ancient) lake “endemic” sponge lineages. J. Great Lakes Res. 46, 1140–1145. doi: 10.1016/j.jglr.2019.09.028
Ezcurra de Drago, I. (1974a). La presencia de Spongilla (Eunapius) fragilis Leidy en Chile (Porifera, Spongillidae). Physis 33, 249–252.
Ezcurra de Drago, I. (1974b). Las especies sudamericanas de Corvomeyenia Weltner (Porifera, Spongillidae). Physis 33, 233–240.
Ezcurra de Drago, I. (1975a). El genero Ephydatia Lamouroux (Porifera, Spongillidae). Sistemática y distribuccion. Physis 34, 157–174.
Ezcurra de Drago, I. (1975b). “Freshwater sponges of suriname,” in Studies on the fauna of suriname and other guyanas, eds D. Geijskes and P. Hummelinck (Dordrecht: Springer), 175–183. doi: 10.1007/978-94-017-7106-1_2
Gómez, P., Carballo, J. L., Cruz-Barraza, J. A., and Camacho-Cancino, M. (2019). On the genus Racekiela in Mexico: Molecular and morphological description of Racekiela cresciscrystae n. sp. J. Nat. Hist. 53, 1351–1368. doi: 10.1080/00222933.2019.1637475
Guerreiro, R. L., Bergier, I., McGlue, M. M., Warren, L. V., Abreu, U. G. P. D., Abrahão, J., et al. (2019). The soda lakes of Nhecolândia: A conservation opportunity for the Pantanal wetlands. Perspect. Ecol. Conserv. 17, 9–18. doi: 10.1016/j.pecon.2018.11.002
Guerreiro, R. L., McGlue, M. M., Stone, J. R., Bergier, I., Parolin, M., da Silva Caminha, S. A. F., et al. (2018). Paleoecology explains Holocene chemical changes in lakes of the Nhecolândia (Pantanal-Brazil). Hydrobiologia 815, 1–19. doi: 10.1007/s10750-017-3429-3
Harrison, F. W. (1988). Utilization of freshwater sponges in paleolimnological studies. Palaeogeogr. Palaeoclimatol. Palaeoecol. 62, 387–397. doi: 10.1016/0031-0182(88)90063-6
Hernández, V. M., and Barreat, J. G. (2017). Racekiela andina sp. nov. (Spongillida: Spongillidae): First report of a freshwater sponge from the Venezuelan Andes. Zootaxa 4341, 275–278. doi: 10.11646/zootaxa.4341.2.8
Łukowiak, M. (2020). Utilizing sponge spicules in taxonomic, ecological and environmental reconstructions: A review. PeerJ 8:e10601. doi: 10.7717/peerj.10601
Łukowiak, M., Van Soest, R., Klautau, M., Pérez, T., Pisera, A., and Tabachnick, K. (2022). The terminology of sponge spicules. J. Morphol. 283, 1517–1545. doi: 10.1002/jmor.21520
Manconi, R., and Pronzato, R. (2002). “Suborder spongillina subord. nov.: Freshwater sponges,” in Systema porifera: A guide to the classification of sponges, eds J. N. A. Hooper and R. W. M. van Soest (Berlin: Springer), 921–1019. doi: 10.1007/978-1-4615-0747-5_97
Manconi, R., and Pronzato, R. (2005). Freshwater sponges of the West Indies: Discovery of Spongillidae (Haplosclerida, Spongillina) from Cuba with biogeographic notes and a checklist for the Caribbean area. J. Nat. Hist. 39, 3235–3253. doi: 10.1080/00222930500307327
Manconi, R., and Pronzato, R. (2008). Global diversity of sponges (Porifera: Spongillina) in freshwater. Hydrobiologia 595, 27–33. doi: 10.1007/s10750-007-9000-x
Manconi, R., and Pronzato, R. (2016). “Phylum porifera,” in Keys to nearctic fauna: Thorp and Covich’s freshwater invertebrates, 4ed Edn, Vol. II, eds J. Thorp and D. C. Rogers (San Diego, CA: Elsevier), 39–83. doi: 10.1016/B978-0-12-385028-7.00003-2
Martens, K., and Harrison, K. (1993). An interesting association between a freshwater sponge (Porifera) and an ostracod (Crustacea) in Lake Titicaca (Bolivia). Int. Ver. Theor. Angew. Limnol. Verh. 25, 923–924. doi: 10.1080/03680770.1992.11900282
McGlue, M. M., Silva, A., Zani, H., Corradini, F. A., Parolin, M., Abel, E. J., et al. (2012). Lacustrine records of holocene flood pulse dynamics in the upper paraguay river watershed (Pantanal wetlands, Brazil). J Quaternary Res. 78, 285–294. doi: 10.1016/j.yqres.2012.05.015
Neumann, K., Strömberg, C. A. E., Ball, T., Albert, R. M., Vrydaghs, L., and Cummings, L. S. (2019). International code for phytolith nomenclature (ICPN) 2.0. Ann. Bot. 124, 189–199. doi: 10.1093/aob/mcz064
Nicacio, G., and Pinheiro, U. (2015). Biodiversity of freshwater sponges (Porifera: Spongillina) from northeast Brazil: New species and notes on systematics. Zootaxa 3981, 220–240. doi: 10.11646/zootaxa.3981.2.4
Nicacio, G., Severi, W., and Pinheiro, U. (2011). New species of Radiospongilla (Porifera: Spongillidae) from Brazilian inland waters. Zootaxa 3132, 56–63. doi: 10.11646/zootaxa.3132.1.2
Parolin, M., Volkmer-Ribeiro, C., and Stevaux, J. C. (2007). Sponge spicules in peaty sediments as paleoenvironmental indicators of the Holocene in the upper Paraná River, Brazil. Rev. Bras. Paleontol. 10, 17–26. doi: 10.4072/rbp.2007.1.02
Penney, J. T., and Racek, A. A. (1968). Comprehensive revision of a worldwide collection of freshwater sponges (Porifera, Spongillidae). Bull. U.S. Natl Museum 272, 1–184. doi: 10.5479/si.03629236.272.1
Pinheiro, U., and Calheira, L. (2020). “Phylum porifera,” in Thorp and Covich’s freshwater invertebrates, eds C. Damborenea, J. H. Thorp, and D. C. Rogers (New York, NY: Academic Press), 79–92. doi: 10.1016/B978-0-12-804225-0.00003-4
Pinheiro, U., Calheira, L., and Hajdu, E. (2015a). A new species of freshwater sponge, Heteromeyenia barlettai sp. nov. from an aquarium in São Paulo, Brazil (Spongillida: Spongillidae). Zootaxa 4034, 351–363. doi: 10.11646/zootaxa.4034.2.7
Pinheiro, U., Calheira, L., Martins, C., Janson, L., Taylor, R., and Samaai, T. (2020). Two new species of freshwater sponges from Neotropical and Afrotropical Regions. Zootaxa 4728, 363–371. doi: 10.11646/zootaxa.4728.3.5
Pinheiro, U., Docio, L., Nicacio, G., and Hajdu, E. (2013). A reassessment of neotropical species of Corvospongilla (Porifera: Spongillidae). J. Nat. Hist. 47, 2373–2384. doi: 10.1080/00222933.2013.798700
Pinheiro, U., Hajdu, E., and Correa, M. (2004). First description of gemmules of Ephydatia facunda Weltner, 1895 (Porifera, Haplosclerida, Spongillidae) by scanning electron microscopy, with underwater observations of a large population from north-eastern Brazil. J. Nat. Hist. 38, 1071–1080. doi: 10.1080/0022293031000064404
Pinheiro, U., Nicacio, G., and Muricy, G. (2015c). An example of the importance of labels and fieldbooks in scientific collections: A freshwater sponge misunderstood for a marine new genus and species. Zootaxa 3974, 447–450. doi: 10.11646/zootaxa.3974.3.12
Pinheiro, U., Silva, C., and Calheira, L. (2015b). Sanidaster in freshwater sponges: An unexpected spicule for the birotuled Genus Corvoheteromeyenia Ezcurra de Drago, 1979. Zoosystema 37, 449–456. doi: 10.5252/z2015n3a2
Piperno, D. R. (2006). Phytoliths: a comprehensive guide for archaeologists and paleoecologists. London: Rowman Altamira.
Pisera, A., and Sáez, A. (2003). Paleoenvironmental significance of a new species of freshwater sponge from the Late Miocene Quillagua Formation (N Chile). J. South Am. Earth Sci. 15, 847–852. doi: 10.1016/S0895-9811(03)00012-9
Pisera, A., Siver, P., and Wolfe, A. (2013). A first account of freshwater potamolepid sponges (Demospongiae, Spongillina, Potamolepidae) from the middle Eocene: Biogeographic and paleoclimatic implications. J. Paleontol. 87, 373–378. doi: 10.1666/12-079.1
Pisera, A., Siver, P., and Wolfe, A. (2016). The sponge genus Ephydatia from the high-latitude middle Eocene: Environmental and evolutionary significance. Palaeontol. Z. 90, 673–680. doi: 10.1007/s12542-016-0328-2
Potts, E. (1885). Fresh-water sponges from Mexico. Proc. U.S Natl Museum 8, 587–589. doi: 10.5479/si.00963801.8-542.587
Potts, E. (1887). Contributions towards a synopsis of the American forms of fresh water sponges with descriptions of those named by other authors and from all parts of the world. Proc. Acad. Nat. Sci. Philad. 39, 158–279.
Pronzato, R., Pisera, A., and Manconi, R. (2017). Fossil freshwater sponges: Taxonomy, geographic distribution, and critical review. Acta Palaeontol. Polonica 62, 467–495. doi: 10.4202/app.00354.2017
Rasbold, G. G., McGlue, M. M., Stevaux, J. C., Parolin, M., Silva, A., and Bergier, I. (2019). Sponge spicule and phytolith evidence for late quaternary environmental changes in the tropical Pantanal wetlands of western Brazil. Palaeogeogr. Palaeoclimatol. Palaeoecol. 518, 119–133. doi: 10.1016/j.palaeo.2019.01.015
Rasbold, G. G., McGlue, M. M., Stevaux, J. C., Parolin, M., Silva, A., and Bergier, I. (2021). Enhanced middle Holocene organic carbon burial in tropical floodplain lakes of the Pantanal (South America). J. Paleolimnol. 65, 181–199. doi: 10.1007/s10933-020-00159-5
Ricciardi, A., and Reiswig, H. M. (1993). Freshwater sponges (Porifera, Spongillidae) of eastern Canada: Taxonomy, distribution, and ecology. Can. J. Zool. 71, 665–682. doi: 10.1139/z93-091
Sawakuchi, A. O., Hartmann, G. A., Sawakuchi, H. O., Pupim, F. D. N., Bertassoli, D. J., Parra, M., et al. (2015). The Volta Grande do Xingu: Reconstruction of past environments and forecasting of future scenarios of a unique Amazonian fluvial landscape. Sci. Dril. 20, 21–32. doi: 10.5194/sd-20-21-2015
Schindler, T., Wuttke, M., and Poschmann, M. (2008). Oldest record of freshwater sponges (Porifera: Spongillina) spiculite finds in the Permo-Carboniferous of Europe. Paläontl. Z. 82, 373–384. doi: 10.1007/BF03184428
Sifeddine, A., Martin, L., Turcq, B., Volkmer-Ribeiro, C., Soubiès, F., Cordeiro, R. C., et al. (2001). Variations of the Amazonian rainforest environment: A sedimentological record covering 30,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 168, 221–235. doi: 10.1016/S0031-0182(00)00256-X
Sokolova, A. M., Palatov, D. M., Masuda, Y., and Itskovich, V. B. (2021). Investigation of the spongillid Spongilla alba Carter, 1849 reveals a new group of brackish-water sponges. Syst. Biodivers. 19, 976–992. doi: 10.1080/14772000.2021.1958948
Spaulding, S. A., Potapova, M. G., Bishop, I. W., Lee, S. S., Gasperak, T. S., Jovanoska, E., et al. (2021). Diatoms.org: Supporting taxonomists, connecting communities. Diatom Res. 36, 291–304. doi: 10.1080/0269249X.2021.2006790
Strömberg, C. A. (2004). Using phytolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the great plains of North America during the late Eocene to early Miocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 207, 239–275. doi: 10.1016/j.palaeo.2003.09.028
Strömberg, C. A. (2005). Decoupled taxonomic radiation and ecological expansion of open-habitat grasses in the Cenozoic of North America. Proc. Natl Acad. Sci. U.S.A. 102, 11980–11984. doi: 10.1073/pnas.0505700102
Strömberg, C. A. E., Dunn, R. E., Crifò, C., and Harris, E. B. (2018). “Phytoliths in paleoecology: Analytical considerations, current use, and future directions,” in Methods in paleoecology: Reconstructing cenozoic terrestrial environments and ecological communities, eds D. A. Croft, D. F. Su, and S. W. Simpson (Cham: Springer International Publishing), 235–287. doi: 10.1007/978-3-319-94265-0_12
Strömberg, C. A., Dunn, R. E., Madden, R. H., Kohn, M. J., and Carlini, A. A. (2013). Decoupling the spread of grasslands from the evolution of grazer-type herbivores in South America. Nat. Commun. 4:1478. doi: 10.1038/ncomms2508
Tavares, M. C., and Volkmer-Ribeiro, C. (1997). Redescrição das esponjas de água doce Oncosclera navicella (Carter, 1881) e Spongilla spoliata Volkmer-Ribeiro & Maciel, 1983 (Spongillidae). Biociências 5, 97–111.
Tavares-Frigo, M., Volkmer-Ribeiro, C., Oliveira, A. E. Z., and Machado, V. (2015). Freshwater sponges from the Pampa Biome, Brazil, with description of a new species of Oncosclera. Neotrop. Biol. Conserv. 10, 110–122. doi: 10.4013/nbc.2015.103.01
Trombold, C. D., and Israde-Alcantara, I. (2005). Paleoenvironment and plant cultivation on terraces at La Quemada, Zacatecas, Mexico: The pollen, phytolith and diatom evidence. J. Archaeol. Sci. 32, 341–353. doi: 10.1016/j.jas.2004.10.005
Volkmer-Ribeiro, C. (1963). Spongilla jewelli n. sp. from fresh-water sponge at Brazil. Anais Acad. Bras. Ciência 35, 271–273.
Volkmer-Ribeiro, C. (1970). Oncosclera-a new genus of freshwater sponges (Porifera-Spongillidae) with redescription of two species. Amazoniana 2, 435–442.
Volkmer-Ribeiro, C. (1973). Redescription and ecomorphic variations of the freshwater sponge Trochospongilla minuta (Potts, 1887). Proc. Acad. Nat. Sci. Philad. 125, 137–144.
Volkmer-Ribeiro, C. (1976). A new monotypic genus of neotropical freshwater sponges (Porifera-Spongillidae) and evidence of a speciation via hybridism. Hydrobiologia 50, 271–281. doi: 10.1007/BF00021002
Volkmer-Ribeiro, C. (1979). Evolutionary study of genus Metania Gray, 1867 (Porifera-Spongillidae) 1. The new species. Amazoniana 6, 639–649.
Volkmer-Ribeiro, C. (1984). Evolutionary study of the genus Metania Gray, 1867 (Porifera: Spongillidae): II. Redescription of two neotropical species. Amazoniana 8, 541–553.
Volkmer-Ribeiro, C. (1992). The freshwater sponges in some peat-bog ponds in Brazil. Amazoniana 12, 317–335.
Volkmer-Ribeiro, C., and Costa, P. R. C. (1992). On Metania spinata (Carter, 1881) and Metania kiliani n. sp.: Porifera, Metaniidae Volkmer-Ribeiro, 1986. Amazoniana 12, 7–16.
Volkmer-Ribeiro, C., and De Rosa-Barbosa, R. (1972). On Acalle recurvata (Bowerbank, 1863) and an associated fauna of other freshwater sponges. Rev. Bras. Biol. 32, 303–317.
Volkmer-Ribeiro, C., and De Rosa-Barbosa, R. (1978). A new genus and species of Neotropical freshwater sponges. Iheringia Ser. Zool. 52, 103–107.
Volkmer-Ribeiro, C., and De Rosa-Barbosa, R. (1985). Redescription of the freshwater sponges Trochospongilla repens (Hinde, 1888) and Trochospongilla amazonica (Weltner, 1895) with an account of the South American species of Trochospongilla (Porifera, Spongillidae). Iheringia Sér. Zool. 65, 77–93.
Volkmer-Ribeiro, C., and Machado, V. D. S. (2009). Freshwater sponges (Porifera, Demospongiae) in a benthic filter feeding community at the Guanacaste Dry Forest, Costa Rica. Iheringia Sér. Zool. 99, 335–344. doi: 10.1590/S0073-47212009000400001
Volkmer-Ribeiro, C., and Maciel, S. B. (1983). New freshwater sponges from Amazonian waters. Amazoniana 8, 255–264.
Volkmer-Ribeiro, C., and Mothes de Moraes, B. (1981). Drulia ctenosclera, a new species of neotropical spongillid. Iheringia Sér. Zool. 60, 117–121.
Volkmer-Ribeiro, C., and Motta, J. F. M. (1995). Esponjas formadoras de espongilitos em lagoas no Triângulo Mineiro e adjacências, com indicação de preservação de habitat. Biociências 3, 145–169.
Volkmer-Ribeiro, C., and Parolin, M. (2005). Segundo registro de Sterrastrolepis brasiliensis Volkmer-Ribeiro & de Rosa-Barbosa (Demospongiae, Potamolepidae) com descrição do habitat e de assembléia, Bacia do Rio Paraná, Brasil. Rev. Bras. Zool. 22, 1003–1013. doi: 10.1590/S0101-81752005000400028
Volkmer-Ribeiro, C., and Tavares, M. (1995). Redescrição de Drulia uruguayensis Bonetto & Ezcurra de drago, 1968 com redefinição do gênero Drulia Gray, 1867 (Porifera: Metaniidae). Biociências 3, 183–205.
Volkmer-Ribeiro, C., and Turcq, B. (1996). SEM analysis of silicious spicules of a freshwater sponge indicate paleoenvironmental changes. Acta Microscopica 5, 186–187.
Volkmer-Ribeiro, C., Bichuette, M. E., and Machado, V. D. S. (2010a). Racekiela cavernicola (Porifera: Demospongiae) new species and the first record of cave freshwater sponge from Brazil. Neotrop. Biol. Conserv. 5, 53–58. doi: 10.4013/nbc.2010.51.08
Volkmer-Ribeiro, C., de Rosa-Barbosa, R., and Machado, V. D. S. (2005). Corvomeyenia epilithosa sp. nov.(Porifera, Metaniidae) no Parque Nacional da Serra Geral, Rio Grande do Sul, Brasil. Rev. Brasl. Zool. 22, 844–852. doi: 10.1590/S0101-81752005000400007
Volkmer-Ribeiro, C., de Rosa-Barbosa, R., and Tavares, M. C. (1988). Anheteromeyenia sheilae sp. n. e outras esponjas dulciaquícolas da região costeira do Rio Grande do Sul. (Porifera, Spongillidae). Iheringia Sér. Zool. 68, 83–98.
Volkmer-Ribeiro, C., Ezcurra de Drago, I., Machado, V., and Sabaj, M. H. (2017). Drulia cristinae, new species of sponge from the rio Xingu, Amazonas Basin, Brazil (Porifera: Demospongiae: Poecilosclerida: Metaniidae Volkmer-Ribeiro, 1986). Proc. Acad. Nat. Sci. Philad. 166, 1–17. doi: 10.1635/053.166.0101
Volkmer-Ribeiro, C., Machado, V. D. S., Oliveira, K. F., and Soares, F. V. (2010c). A new genus of freshwater sponges with a new species from Amazonian waters (Porifera, Demospongiae). Rev. Ciências Ambientais 4, 47–64.
Volkmer-Ribeiro, C., Parolin, M., Fürstenau-Oliveira, K., and Menezes, E. R. (2010b). Colonization of hydroelectric reservoirs in Brazil by freshwater sponges, with special attention on Itaipu. Interciencia 35, 340–347.
Volkmer-Ribeiro, C., Tavares, M. D. C. M., and Oliveira, K. F. (2009). Acanthotylotra alvarengai (Porifera, Demospongiae) new genus and species of sponge from Tocantins river, Pará State, Brazil. Iheringia Sér. Zool. 99, 345–348. doi: 10.1590/S0073-47212009000400002
Volkmer-Ribeiro, C., Tavares-Frigo, M., Ribeiro, A. C., and Bichuette, M. E. (2021). Arinosaster patriciae (Porifera, Demospongiae): New genus and species and the second record of a cave freshwater sponge from Brazil. Neotrop. Biol. Conserv. 16:45. doi: 10.3897/neotropical.16.e50156
Volkmer-Ribeiro, C., and Rützler, K. (1997). Pachyrotula, a new genus of freshwater sponges from New Caledonia (Porifera: Spongillidae). Proc. Biol. Soc. Washington 110, 489–501.
Weltner, D. (1895). Spongillidenstudien III.” Katalog und Verbreitung der bekannten Suswasserschwamme. Arch. Naturgesh. 61, 114–144.
Keywords: Porifera, paleoecology, proxies, lacustrine sediments, wetlands
Citation: Rasbold GG, Calheira L, Domingos-Luz L, Pessenda LCR, Pinheiro U and McGlue MM (2023) A morphological guide of neotropical freshwater sponge spicules for paleolimnological studies. Front. Ecol. Evol. 10:1067432. doi: 10.3389/fevo.2022.1067432
Received: 11 October 2022; Accepted: 15 December 2022;
Published: 18 January 2023.
Edited by:
Irene Tunno, Lawrence Livermore National Laboratory (DOE), United StatesReviewed by:
Andrzej Pisera, Institute of Paleobiology, Polish Academy of Sciences, PolandCopyright © 2023 Rasbold, Calheira, Domingos-Luz, Pessenda, Pinheiro and McGlue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giliane G. Rasbold,  Z3Jhc2JvbGRAZ21haWwuY29t
Z3Jhc2JvbGRAZ21haWwuY29t
†ORCID: Giliane G. Rasbold, orcid.org/0000-0001-7375-6261; Ludimila Calheira, orcid.org/0000-0003-4585-2373; Leandro Domingos-Luz, orcid.org/0000-0001-8477-1010; Luiz C. R. Pessenda, orcid.org/0000-0001-9119-8195; Ulisses Pinheiro, orcid.org/0000-0003-3658-1372; Michael M. McGlue, orcid.org/0000-0002-0725-7250
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.