 Mateus C. Silva
Mateus C. Silva Peter Moonlight2†
Peter Moonlight2† Rafael S. Oliveira
Rafael S. Oliveira Lucy Rowland
Lucy Rowland- 1Department of Geography, College of Life and Environmental Sciences, University of Exeter, Exeter, United Kingdom
- 2Tropical Diversity Section, Royal Botanic Garden Edinburgh, Edinburgh, United Kingdom
- 3Department of Plant Biology, Institute of Biology, University of Campinas, Campinas, Brazil
Seed markets are vital to scaling up ecosystem restoration in the Brazilian Cerrado, home of the world’s most species-rich grasslands and savannas. We compiled lists of species traded by four major Cerrado seed supply systems to investigate the representativeness of the species currently available for seed-based restoration. We also identified whether dominant ground-layer species are being sourced for seed production. Seeds from 263 Cerrado species can be purchased for restoration, of which 68% are trees, particularly legumes (24%). 63% of the traded species were found in only one seed supply system. The five most dominant graminoids of the Cerrado ground layer were available for sale, but two additional species uncommon in old-growth areas represented 44% of the sales of a key seed trader in Central Brazil. The expansion of Cerrado seed supply systems should be supported to further increase the number of species on the market. Sourcing seeds from a diversity of herbaceous species is central to facilitating the restoration of species-rich grasslands and savannas in the Cerrado. Recovering the diversity and functioning of old-growth open ecosystems through seeds will depend on increasing the supply and demand for species typical of Cerrado’s ground layer.
Introduction
Ecological restoration is central to tackling biodiversity loss and securing ecosystem services across the globe (Suding, 2011). Seeds provide a prominent avenue for recovering vegetation composition and functioning after degradation (Pedrini and Dixon, 2020). Seeds of native species can be sustainably collected from wild populations (Pedrini et al., 2020) and used to establish target species in restoration projects, easing dispersal limitations that may constrain natural regeneration (Shaw et al., 2020). Importantly, seed sowing can be more cost-effective than other restoration methods such as planting seedlings (Palma and Laurance, 2015; Raupp et al., 2020). Upscaling restoration efforts may depend on the number and identity of species available on the seed market, as well as the seed provenance, quality, and quantity (León-Lobos et al., 2020; Nef et al., 2021). A market that sources seeds from a diverse set of species spread across numerous locations can allow practitioners to choose a range of locally adapted species and genotypes for restoration projects (Erickson and Halford, 2020). Furthermore, seed markets should ideally cover a comprehensive variety of life forms and evolutionary lineages to foster the restoration of functionally and phylogenetically diverse ecosystems (Fremout et al., 2022). Finally, when the goal is to restore the ecosystem to resemble surrounding old-growth vegetation, it is essential to source seeds of species typical of old-growth areas.
Ecological restoration is of particular importance in Brazil. Brazil is home to almost 34,000 flowering plant species (Reflora, 2020). Yet, land-use changes have led to a loss of about one-third of Brazil’s native vegetation (Mapbiomas, 2021), putting this diversity under threat. Up to 19 million hectares of private land are required by law to be restored in Brazil, with deadlines varying by State (Guidotti et al., 2017). Additionally, Brazil’s government has committed to restoring 12 million hectares by 2030 (MMA, 2017). Seed supply systems for native species have emerged as a key strategy to achieve these national restoration targets (Urzedo et al., 2022). These systems consist mainly of cooperatives, including indigenous populations, which harvest, store, process, and sell the seeds. Seed markets provide income to local communities, supporting their livelihoods and the sustainable use of their lands (Schmidt et al., 2019). Brazil’s seed market will need to increase its seed sourcing capacity 6- to 30-times to achieve the 12 million hectare goal (Urzedo et al., 2020). Therefore, it is fundamentally important to design strategies to expand Brazilian seed supply systems before the end of this decade (2030).
The Brazilian Cerrado is a global biodiversity hotspot dominated by open ecosystems (i.e., grasslands and savannas; Myers et al., 2000). About 17–31% of Brazil’s restoration projects are expected to take place in the Cerrado. However, relative to the Atlantic Forest, another Brazilian biodiversity hotspot, restoration in the Cerrado is still in its infancy in terms of restored area and methodologies (see Pinto et al., 2014; Crouzeilles et al., 2019). Targeted and strategic expansion of restoration infrastructure is essential to meet Cerrado restoration ambitions (Strassburg et al., 2017), which range from two to 6 million hectares (Guidotti et al., 2017; MMA, 2017). Three factors will be key to this expansion. First, more than 12,000 plant species are found in the Cerrado (Zappi et al., 2015), and their distribution is often regionalized (Bridgewater et al., 2004; Françoso et al., 2020), so seed sourcing needs to take place throughout the Cerrado to represent this remarkable plant diversity. Second, savannas and grasslands cover about 70% of the Cerrado region (Mapbiomas, 2021), with herbaceous plants being the dominant life form. Supplying a diversity of herbaceous species will, consequently, be essential to restoring species-rich open ecosystems (Buisson et al., 2021). Third, it is unclear whether the dominant species characteristic of old-growth grasslands and savannas are available on the seed market, especially from the ground layer (i.e., vegetation strata composed of graminoids, forbs, and shrubs).
We aimed to assess how well the current seed market represents the diversity of the Brazilian Cerrado flora. We used this information to evaluate what the limits of the seed market may be and how this growing market can be expanded strategically. We focused on the four seed supply systems representing the main seed traders for restoration in the Cerrado (Caminhos da Semente, 2020): Rede de Sementes do Xingu (RSX), Rede de Sementes do Cerrado (RSC), VerdeNovo (VN), and Restauradores da RDS Nascentes Geraizeiras (RDS). We gathered data on the approximate location of seed collection and species on sale per system. This data was contrasted with estimates of species richness at the Cerrado scale. Additionally, we gathered ground layer vegetation survey data from the literature to identify its dominant species. We compared the species dominance rank with the 2017–2019 sales record of the RSC, the oldest and largest seed supplier of Cerrado species exclusively. We addressed the following questions.
1. How similar is the flora traded by different seed supply systems?
2. How are life forms and plant families represented in the systems?
3. Are dominant ground-layer species from old-growth open ecosystems available on the seed market?
Materials and methods
Seed harvesting sites
We mapped the centroid of the municipalities where the seed supply systems are active (Supplementary Table 1). The municipalities were obtained from the RSX website (6° Boletim, p. 2)1 and by directly contacting personnel from the RSC, VN, and RDS supply systems. We calculated the centroid of each municipality polygon using the function “st_centroid” from the package “sf” (Pebesma, 2018) on R version 4.1.2 (R Core Team, 2021). We displayed the municipality centroids alongside the Cerrado floristic regionalization map proposed by Françoso et al. (2020). The map depicts areas sharing a similar set of woody species, termed here as “biogeographical districts.”
Species richness
We accessed the species list of each seed supply system by directly contacting them (VN and RDS) or through their websites (RSX and RSC; Supplementary Table 2). The lists consisted of the species on sale for the first half of 2021. We used the R package “flora” (Carvalho, 2020) to standardize species’ accepted names and check their endemism and threatening status according to the Brazilian Flora 2020 checklist (version 393.291). We filtered the species that occur in the Cerrado region (263 of 305 species) as the RSX system is in the transitional zone between the Cerrado and Amazon. Additionally, we checked whether the species turnover between the systems might be a result of their distance. For that, we used the function “st_distance” from the R package “sf” to calculate the minimum geographical distance between each pair of systems (Karney, 2013). We also investigated whether the species richness of a given system might be a function of its age. The year of creation of each seed supply system was confirmed on their websites (RSX: https://www.sementesdoxingu.org.br/, RSC: https://www.rsc.org.br/, VN: https://consultoriaverdenovo.weebly.com/) and by contacting their staff (RDS).
Life forms and families
Species were grouped both by life form (tree, palm, liana, shrub, subshrub, forb, and graminoid) and botanical family, according to the Brazilian Flora 2020 checklist, which follows the APG IV (Flora do Brasil, 2020). Brazilian Flora 2020 checklist life form classification is based on the notes from herbarium collections. We considered graminoid all the herbaceous species belonging to the Poaceae (grasses), Cyperaceae (sedges), and Juncaceae (rushes) families. All the non-graminoid herbaceous species were assigned to the forb life form. We used the Brazilian Flora 2020 online platform2 to access the number of species per life form and family over the whole Cerrado region. We standardized the number of species per life form and family by the total number of species available on the seed supply systems and recorded in the Cerrado. The standardized (std.) species richness varied from 0 to 1. We then determined a metric of representativeness by calculating the difference between the std. species richness over the Cerrado region and the std. species richness within the seed supply systems for each life form and botanical family. We repeated the analysis within each seed supply system to verify whether the representation biases toward life forms or families were widespread among the systems.
Ground-layer dominant species
We searched for “Cerrado” and “Herbáceo” (“herbaceous” in Portuguese), in the Brazilian National Thesis and Dissertations repository (217 publications). We also searched for “Cerrado” and “Herbaceous” on the Web of Science (166 publications). We selected all the publications that included ground layer life forms (i.e., graminoid, forb, and shrub) and displayed a table with either plant cover or density at the species level (39 publications). We only considered data from old-growth ecosystems. We analyzed different study sites separately when the publication made that distinction. In the case of time series, we selected the data from the most recent time interval. When two or more publications used the same data set, we retained the latest publication to include the most updated and revised data. Since our focus was on open ecosystems, we removed data from the ground layer of closed-canopy ecosystems (e.g., gallery forests, woody encroached savannas; four study sites). We ended up with 66 study sites from 25 publications (Supplementary Table 3). We standardized the species name and removed all tree and liana species based on the Flora do Brasil (2020). We calculated the abundance index of each species in each study site by dividing their cover or density value by the total value across all species for the whole site. The abundance index varies from 0 to 1, with an index of 1 representing a monodominant species and 0 if the species was absent from the study site. We, thus, averaged the abundance indices at the species level. We calculated the relative frequency by dividing the number of study sites where a given species was found by the total number of sites. Finally, we calculated the importance value index (IVI) by averaging the abundance index and relative frequency (Munhoz and Araújo, 2011). IVIs closer to 1 mean species that occur in several sites and are abundant wherever they occur and hence are dominant. IVI was chosen to flag dominant species as it balances local abundance and regional commonness. We emphasize that IVI may not capture the importance of some species, such as ecosystem engineering or others with strong legacy effects. However, IVI is still useful to pinpoint species most characteristic of reference ecosystems, which is the goal of this study.
Rede de Sementes do Cerrado sales record
We obtained the total mass of seeds traded by the RSC in the period from 2017 to 2019 by directly contacting their personnel. Only ground-layer species were kept following the procedure mentioned in “Ground-layer dominant species.”
Results
Geographic distribution of seed sourcing
The seed supply systems are located approximately from 9°S to 16°S (Figure 1). RSX was the biggest seed supply system in terms of the number of municipalities (18), followed by RSC (6), RDS (5), and VN (4). The current extension of the four seed supply systems provided a good representation of the Cerrado central-west biogeographical district (6 RSC sites, 5 RSX sites) and the north of the southwest district (4 RDS sites, 1 VN site).
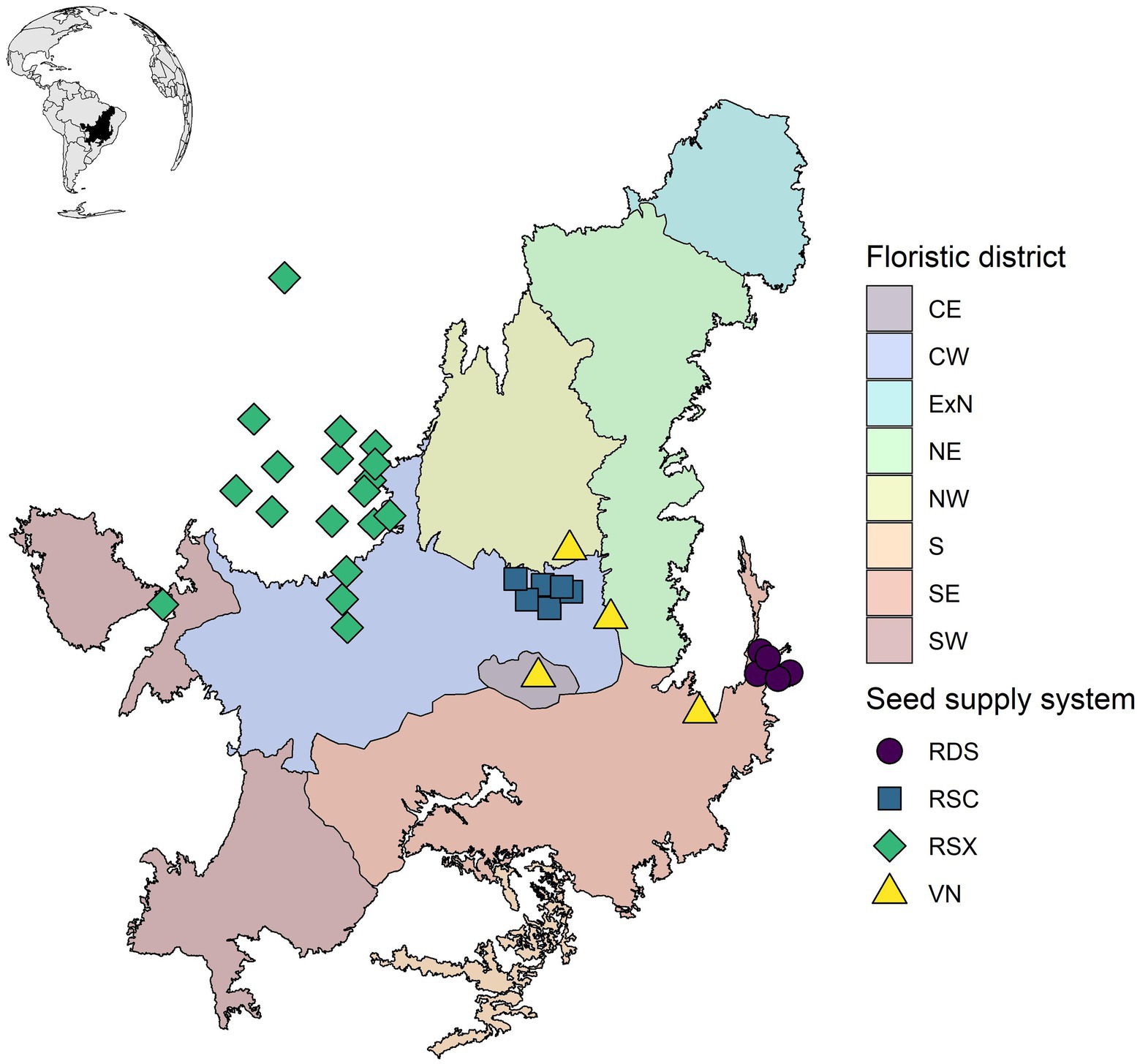
Figure 1. Geographical coverage of the main Cerrado seed supply systems for restoration. The points correspond to the centroid of the towns and cities with seed collection activities over the Cerrado region. The studied seed supply systems are the Restauradores da RDS Nascentes Geraizeiras (RDS), Rede de Sementes do Xingu (RSX; which also collect seeds outside of the Cerrado region), Rede de Sementes do Cerrado (RSC), and VerdeNovo (VN). Biogeographical districts depict areas with similar woody floras according to Françoso et al. (2020) and were represented by contrasting colors. The districts are central (CE), central-west (CW), external north (ExN), northeast (NE), northwest (NW), south (S), southeast (SE), and southwest districts (SW).
Floristic relationships
A total of 263 species were made available for restoration by the seed supply systems (Figure 2A). 12% of the traded species were endemic to the Cerrado (33 species). Regarding the conservation status of the traded species, one species was classified as endangered, five as vulnerable, four as near threatened, 40 as least concern, one as data deficient, and 212 were not evaluated. Only 13 out of 263 species (~4%) were sourced by all four systems. 167 species were found only in one system (i.e., unique species), representing around 63% of all traded species. The percentage of unique species per system ranged from 57 to 10% in the RSX and RSC systems, respectively. The VN system offered the greatest number of Cerrado species (165), followed by RSX (141), RSC (66), and RDS (48) systems. The number of shared species achieved its maximum (55) between the geographically closest systems (RSC and VN) and minimum (19) between the systems farthest apart (RSX and RDS; Figure 2B). RSC was the oldest system (2005), followed by RSX (2007), VN (2016), and RDS (2017). The VN and RDS traded the highest and lowest Cerrado species richness, respectively, even though they were created in a similar period, 2016 and 2017, respectively (Figure 2C).
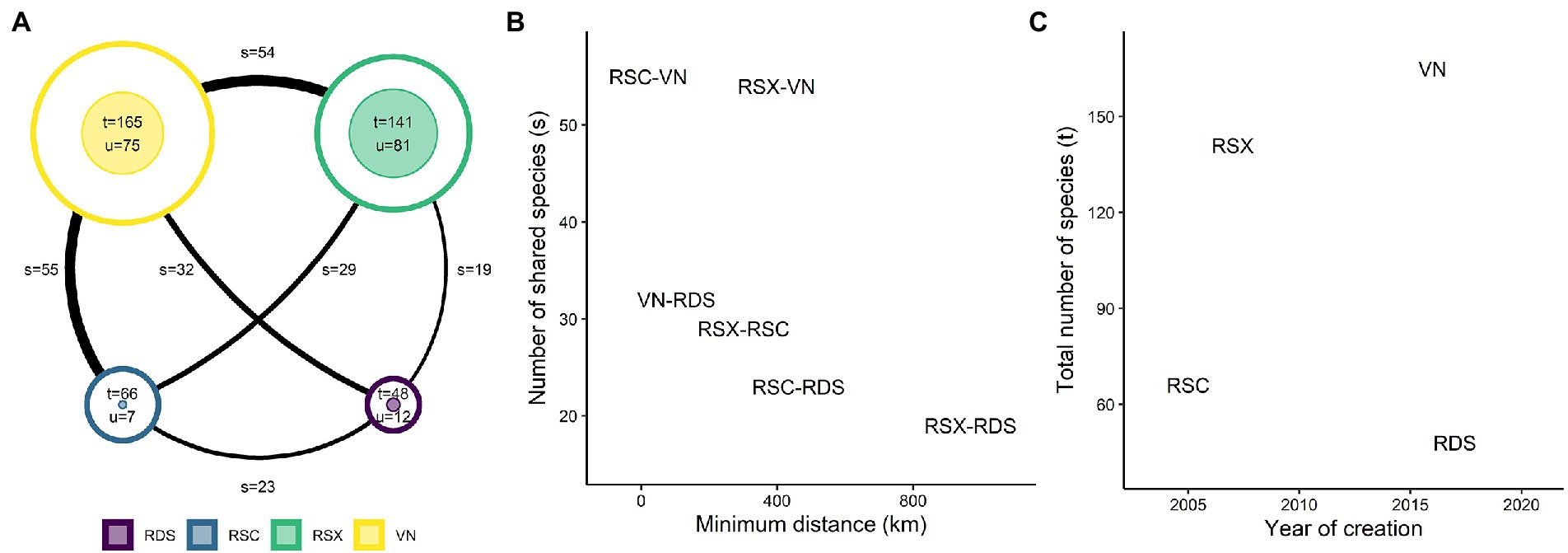
Figure 2. Floristic similarity between the Cerrado supply systems. (A) Diagram showing the number of total (t), unique (u), and shared (s) species between the systems. (B) Relationship between the number of shared species between a pair of seed supply systems and their minimum geographical distance. (C) Relationship between the total number of species of each seed supply system and the year of its creation. Unique species represent the ones traded by a single system. The size of the outer circle in (A) is proportional to t, the size of the inner circle to u, and the line thickness to s. The analysis includes only species that occur in the Cerrado. Seed supply systems were the Restauradores da RDS Nascentes Geraizeiras (RDS), Rede de Sementes do Xingu (RSX), Rede de Sementes do Cerrado (RSC), and VerdeNovo (VN).
Representativeness of life forms and families
Trees were the best-represented life form in the seed supply systems, while forbs were the worst according to the Brazilian Flora 2020 checklist (Figure 3A). 68% of all traded species were trees, though only 14% of the Cerrado flora belonged to this life form (1,761 species). In contrast, 32% of Cerrado flowering plants were forbs (3,948 species), but they represented just 3% of all traded species. Fabaceae (legumes) was the best represented of the 60 traded families (Figure 3B). Legumes accounted for 9% of the total Cerrado flora and 24% of the traded seed flora. No orchid species were traded, yet the Orchidaceae represented 5% of the Cerrado flowering plants. These patterns remained qualitatively similar when each seed supply system was analyzed separately (Figures 3C,D). All four systems had a good representation of trees and legumes and a lack of representation of forbs and orchids.
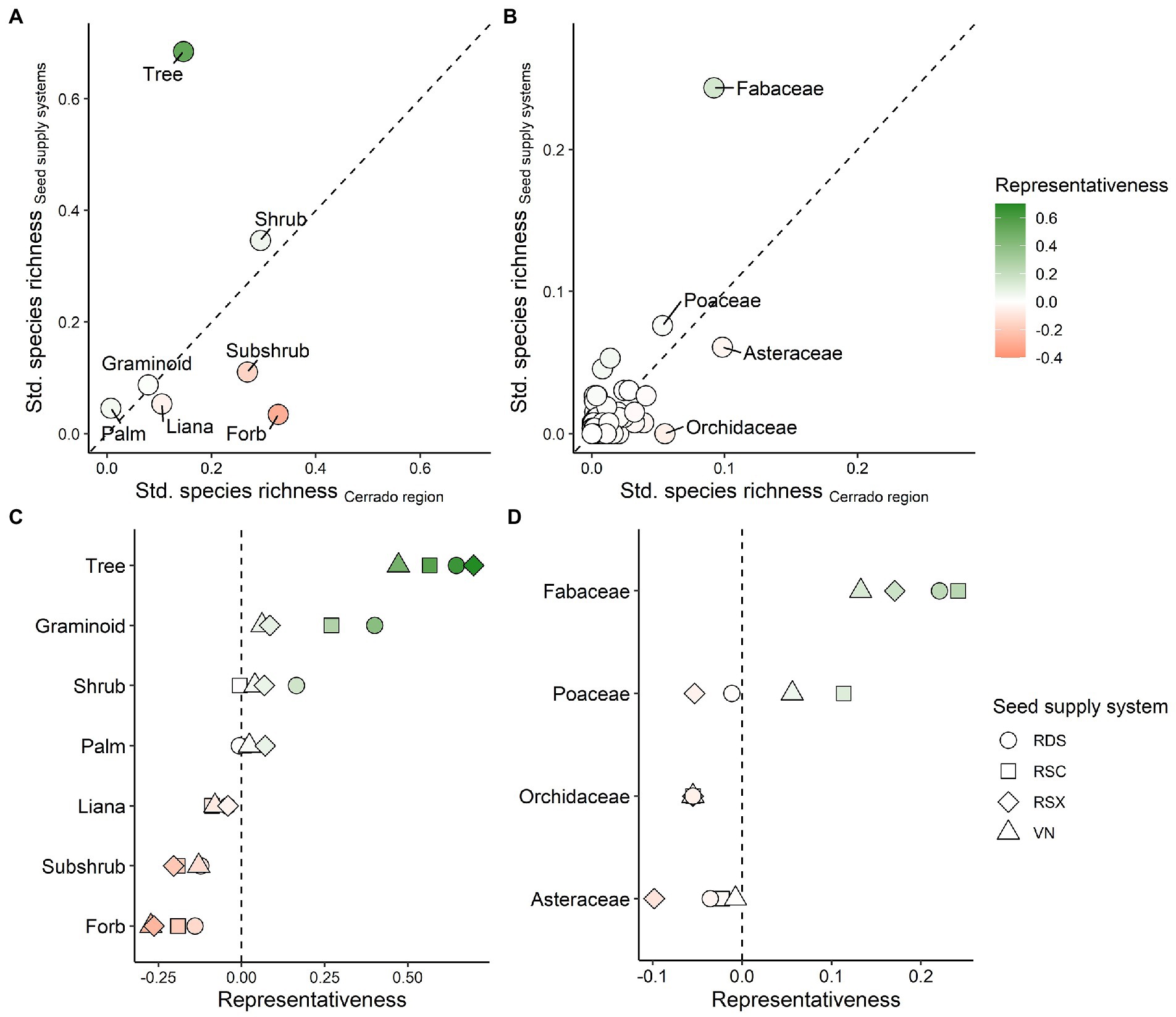
Figure 3. Representativeness of the flora available for seed-based restoration compared to the Cerrado totals. Relationship between the number of species traded by the systems versus the richness across the Cerrado for each life form in (A) and botanic family in (B). Difference between the observed and expected species richness (i.e., representativeness) for each life form in (C) and family in (D) per system. Species richness in (A,B) was standardized by the total number of species found in the Cerrado (x-axis) and available in the seed supply systems (y-axis). Representativeness in all panels consists of the difference between the standard species richness in the seed supply systems and the standard species richness in the Cerrado. The dashed line in (A,B) represent the 1:1 proportion. Representativeness > 0 indicates groups well represented by the seed supply systems given its richness in the Cerrado. Representativeness < 0 indicates groups lacking representation in the seed supply systems given its richness in the Cerrado. The top four species-rich families in the Cerrado are labeled in (B) and highlighted in (D). Seed supply systems were the Restauradores da RDS Nascentes Geraizeiras (RDS), Rede de Sementes do Xingu (RSX), Rede de Sementes do Cerrado (RSC), and VerdeNovo (VN).
Dominant ground-layer species on the market
The seed supply systems traded 7 out of the 15 ground-layer species with the greatest Importance Value Index (IVI) among 66 Cerrado savanna sites (Figures 4A,B; Supplementary Table 4). The top five species in terms of IVI—Trachypogon spicatus (IVI of 0.29), Echinolaena inflexa (0.28), Lagenocarpus rigidus (0.26), Rhynchospora globosa (0.25), and Axonopus brasiliensis (0.23)—had commercialized seeds. Paspalum lineare and Tristachya leiostachya were not traded and occupied the sixth and seventh positions in the IVI rank, respectively. RSC sold 11.63 tons of seeds from ground-layer species between 2017 and 2019, 55% of the total seed they sell. Two species, the shrub Lepidaploa aurea and the grass Andropogon fastigiatus, accounted for 44% of the RSC sales in terms of weight. L. aurea had an IVI of 0.008 (796th position in the IVI rank) and occurred in c. 1.4% of the studied sites. A. fastigiatus was absent from all the 66 sites used to calculate the species IVI. After L. aurea and A. fastigiatus, Schizachyrium sanguineum, Aristida riparia, and Aristida setifolia were the top-selling species, representing 9, 8.3, and 7% of the seeds sales and occupying the 78th, 492nd, and 339th IVI rank positions, respectively (Figure 4C; Supplementary Table 5).
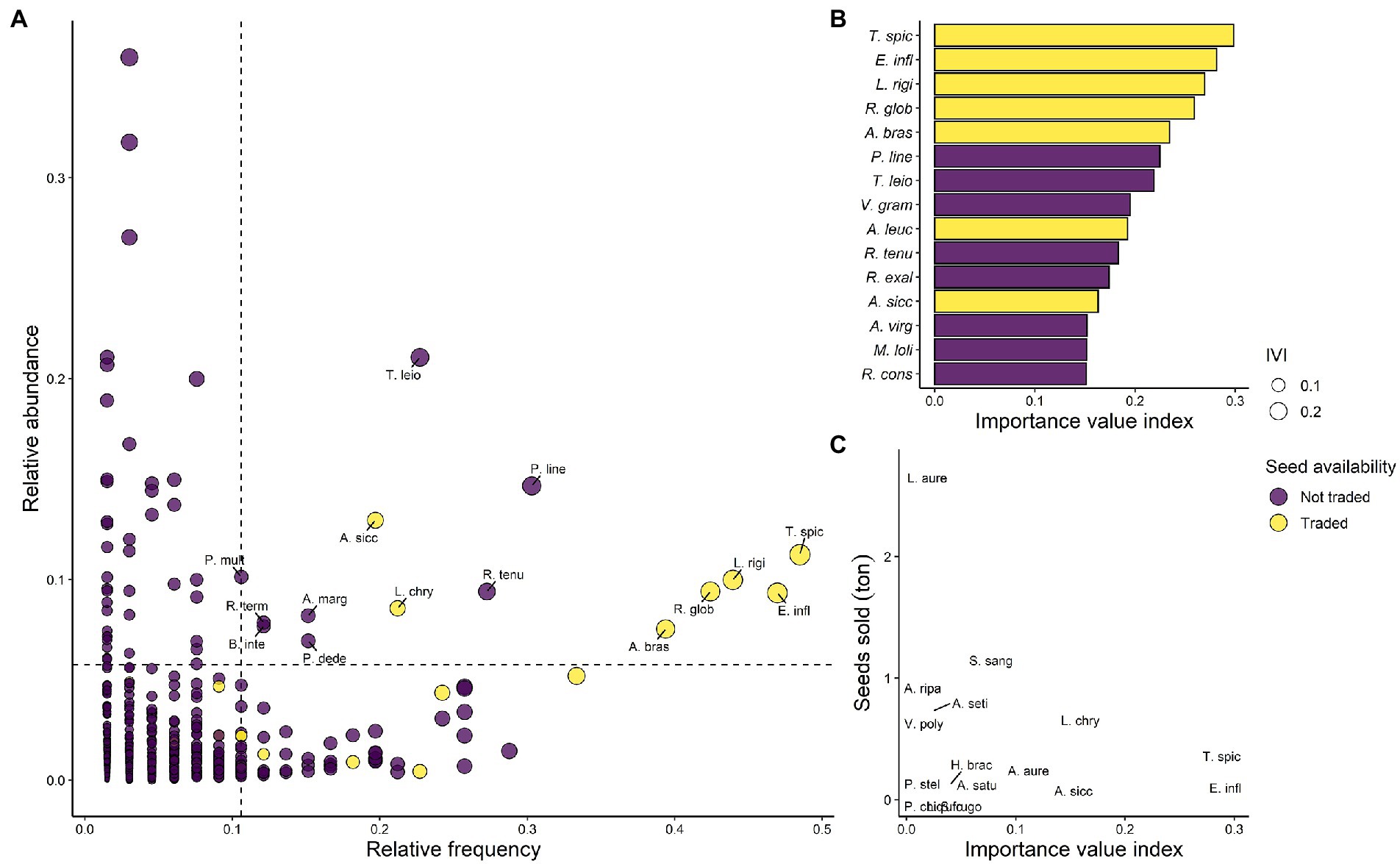
Figure 4. Availability of dominant ground-layer species among the seed supply systems. (A) The relationship between mean relative abundance and relative frequency of 1,108 ground-layer species over 66 Cerrado old-growth open ecosystem sites retrieved from the literature. (B) The top 15 species with the highest importance value index (IVI). (C) The relationship between the tons of seeds sold by the Rede de Sementes do Cerrado (RSC) from 2017 to 2019 per species and the species IVI. The species traded by the seed supply systems are colored in yellow and the non-traded species are in purple. The point size in (A) is proportional to the IVI. The dashed line in (A) mark the 95th percentile of the mean relative abundance (y-axis) and relative frequency (x-axis). Species in (A) located at the top-right quadrat represent locally abundant and geographically widespread species; top-left locally abundant and geographically restricted; bottom-left locally and geographically rare; and bottom-right locally rare and geographically widespread. T. spic is an abbreviation of Trachypogon spicatus, E. infl Echinolaena inflexa, L. rigi Lagenocarpus rigidus, R. glob Rhynchospora globosa, A. bras Axonopus brasiliensis, P. line Paspalum lineare, T. leio Tristachya leiostachya, V. gram Vellozia graminea, A. leuc Andropogon leucostachyus, R. tenu Rhynchospora tenuis, R. exal Rhynchospora exaltata, A. sicc Axonopus siccus, A. purp Axonopus purpusii, A. virg Andropogon virgatus, M. loli Mesosetum loliiforme, L. aure Lepidaploa aurea, S. sang Schizachyrium sanguineum, A. ripa Aristida riparia, V. poly Vernonanthura polyanthes, A. seti Aristida setifolia, L. chry Loudetiopsis chrysothrix, H. brac Hyparrhenia bracteata, A. aure Axonopus aureus, P. stel Paspalum stellatum, L. rufo Lepidaploa rufogrisea, A. satu Achyrocline satureioides, P. chiq Paepalanthus chiquitensis, S. rugo Senna rugosa, B. inte Byrsonima intermedia, P. dede Paspalum dedeccae, P. mult Paspalum multicaule, and R. term Rhynchospora terminalis.
Discussion
We assessed the current locations of the major Cerrado seed supply systems and their species portfolio. We found that these systems have expanded remarkably across the Cerrado’s central zone in less than 20 years, making up to 260 species available for ecological restoration. Trees, especially legumes, were well represented among seed traders, but relatively fewer forb species were available. Dominant ground-layer species were accessible on the seed market, but the seed sales of the RSC system, a key Cerrado seed supplier, were concentrated in two species rarely found in old-growth open ecosystems, the shrub Lepidaploa aurea and grass Andropogon fastigiatus.
Cerrado’s major seed suppliers have unique and complementary species portfolios
Each seed supply system traded a distinctive set of species, suggesting they are not interchangeable, as found in other seed supply systems across the world (Atkinson et al., 2021; Bosshard et al., 2021). Consequently, expanding the existing systems or creating new ones will probably increase the number of species available for seed-based restoration in the Cerrado. The seed supply systems were located in distinct biogeographical districts, which explains their floristic dissimilarity (Ratter et al., 1996; Bridgewater et al., 2004; Amaral et al., 2017; Françoso et al., 2020). The number of shared species decreased as the geographical distance increased, suggesting the broader the distribution of seed harvesting sites, the richer the flora available on seed markets for restoration. Additionally, there was no clear relationship between the year the seed supply systems were founded and their species richness. VN was created just ~7 years ago (2016) and already sells up to 160 species, suggesting the seed supply systems are flexible, innovative, and can expand their species portfolio over a few years (Schmidt et al., 2019). These findings reinforce that expanding the geographical coverage of the seed supply systems can be key to diversifying the seed market and this diversification can potentially take place until the end of this decade (2030).
Trees are overrepresented and forbs are underrepresented in the seed market
Trees are the best-represented life form in the seed supply systems evaluated here. The diversity of legume tree species on sale is one of the factors underpinning tree dominance in the seed markets for restoration. The overrepresentation of trees and legumes was found in all four seed supply systems. This finding suggests that currently the seed market is equipped to source plant material for the restoration of closed-canopy ecosystems (e.g., gallery forests) and the woody layer of savannas in the Cerrado. Yet, sourcing tree seeds is not the priority to restore the open ecosystems that cover up to 70% of the Cerrado region. Tree species often regenerate naturally in degraded lands within three decades in Cerrado savannas (Giles et al., 2021; Silva et al., 2021). Suggesting that assisted natural regeneration could be a prominent method to recover tree species. Forbs and graminoids, on the other hand, have a lower regeneration potential compared to trees (Cava et al., 2018; Overbeck et al., 2022), and they might be the life forms most reliant on active restoration, such as direct seeding.
Seeds from forbs were under-sourced given the richness of this life form in the Cerrado. Three causes may underly this pattern found in all the seed supply systems. First, open ecosystems are often undervalued compared to their closed-canopy counterparts (Parr et al., 2014; Silveira et al., 2022). By extension, forbs are likely to be overlooked in restoration science and practice, relative to trees. Second, harvesting seeds from forb species can be more laborious than trees. Forb seeds are often smaller than tree seeds, so collectors need to spend more time harvesting multiple populations of forbs to collect the same amount of seeds found in a single tree. Third, some species-rich forb lineages, such as Orchidaceae and Asteraceae, are composed mainly of micro-endemic narrow-ranged species (Neto and Forzza, 2013; Campos et al., 2019), which may be absent or rare in the harvesting sites. Yet, forbs should not be neglected when designing restoration interventions in open ecosystems. Forbs in families such as Eriocaulaceae, Xyridaceae, and Velloziaceae can amplify vegetation resilience to fires and drought (de Oliveira Joaquim et al., 2018; Pilon et al., 2021), support pollinator populations (Rabeling et al., 2019), and accelerate soil formation (Teodoro et al., 2019). Including forbs and not only graminoids in the seed mixes can, therefore, maximize the recovery of multiple ecosystem functions in Cerrado grasslands and savannas.
Dominant ground-layer species are available for sale but have low consumer demand
The species portfolio of the seed supply systems already includes plants that can dominate the ground layer of open ecosystems in the Cerrado. Previous studies have reported high ground cover by the grasses T. spicatus and E. inflexa in old-growth grasslands and savannas (de Souza et al., 2021; Nogueira et al., 2022; Teixeira et al., 2022). However, despite their ecological value, these species were not the most popular species for restoration, as evidenced by the RSC sales record. E. inflexa seeds cost c. USD 103 per kg, almost ten times more than the RSC top-selling species (L. aurea and A. fastigiatus), which may explain the low demand for E. inflexa seeds. E. inflexa spreads through rhizomes, which can facilitate its establishment through transplant techniques (Pilon et al., 2019). T. spicatus had a similar price to L. aurea and A. fastigiatus, so the price is unlikely to be the cause of its low demand. Instead, the low establishment rates of T. spicatus may underlie its low sale rank. Up to 80% of T. spicatus seeds can be empty (i.e., embryoless; Zanetti et al., 2020), clarifying its low germination and emergence rates in field conditions (Pellizzaro et al., 2017). Poor seeds and strong dormancy are common in Cerrado grasses (Le Stradic et al., 2015; Fontenele et al., 2020). It is, therefore, fundamental to improve seed quality control and enhancement techniques to establish key ground-layer species when direct seeding is chosen as the main restoration method (Buisson et al., 2021).
The biennial shrub L. aurea and the annual grass A. fastigiatus accounted for almost half of the seeds from ground-layer species sold by RSC from 2017 to 2019. The high demand for L. aurea and A. fastigiatus is probably related to their ability to cover the ground during the first 2 years after direct seeding (Pellizzaro et al., 2017; Coutinho et al., 2019; Sampaio et al., 2019), coupled with a relatively low price for their seed. These species can play an important role in immediately controlling soil erosion due to their fast ground cover. However, L. aurea and A. fastigiatus are rare in Cerrado open ecosystems, suggesting that these species might possess life history strategies that diverge from ones in old-growth areas. For instance, A. fastigiatus and L. aurea have an annual and biennial life cycle, respectively (Motta, 2017; Wolfsdorf et al., 2021), thus relying on seeds as the persistence strategy. Annual and biennial “seeder” species are often rare in Cerrado grasslands and savannas, where the perennial life cycle coupled with below-ground resprouting is the dominant strategy (Pilon et al., 2021). The abundant aboveground biomass produced by short-lived species, such as L. aurea and A. fastigiatus, can lead to high fuel loads, exposing the vegetation to intense fires early in the restoration process when “resprouter” species are not sufficiently abundant to confer ecosystem resilience to fire (Giles et al., 2022). Therefore, continuing to improve the availability and quality of seeds from species characteristic of old-growth sites, alongside continuing to develop techniques to successfully incorporate these species in the early stages of restoration, could increase the likelihood of restoring open ecosystems to a state similar to that of old-growth areas.
Concluding remarks and future perspectives
Our findings suggest that: (1) Brazilian Cerrado seed supply systems are irreplaceable and complement each other in the species they trade; (2) increasing the number of forb species on sale could lead to a more even representation of the flora available for open ecosystem restoration in the Cerrado; and (3) amplifying the demand for species typical of old-growth areas remains a challenge to effectively restore the ground layer of Cerrado grasslands and savannas. We acknowledge that, first, the present insights are based on a momentary picture of the four seed supply systems. However, a snapshot of the current state of the seed market for restoration is a vital step toward its development. Second, we stress that low-IVI species can also be targets in restoration projects, especially when they increase the desired ecosystem function or facilitate the establishment of other species. Third, our IVI rank does not diminish the need of surveying local reference ecosystems as species with low IVI at the scale of the entire Cerrado might be abundant in the region where restoration will take place. We advocate for: (1) more public and private support for creating new or expanding existing seed supply systems over the Cerrado region; (2) a better understanding of the motivations underlying species selection for seed-based restoration; (3) more awareness about the importance of sourcing and using a diverse set of herbaceous plants in open-ecosystem restoration; and (4) clear guidelines on which species should dominate in the seed mixes, potentially T. spicatus, E. inflexa, L. rigidus, R. globosa, or A. brasiliensis when the goal is restoring old-growth grasslands and savannas and/or boosting the multifunctionality of grassy ecosystems. We hope these suggestions provide a roadmap toward a strategic advancement of the seed markets for the restoration of Cerrado’s open ecosystems, especially during the UN Decade on Ecosystem Restoration (2021–2030).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
MS, RP, LR, and PM conceived the research. MS gathered and analyzed the data and drafted the manuscript. MS, PM, RO, RP, and LR edited and gave input to the final version of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
MS, RP, and LR thank WWF-UK and Exeter Alumni MS PhD scholarship (710015629). RO received a CNPq productivity scholarship and acknowledge CNPq grants 309709/2020, 303988/2018-5, and 312270/2017-8. RO, LR, and RP acknowledge the joint NERC-Fapesp grant 19/07773-1 and NE/S000011/1. LR acknowledges a NERC independent research fellowship NE/N014022/1. RP and PM acknowledge the NERC Newton FAPESP grant NE/N01247X/1.
Acknowledgments
We are grateful to ARATICUM Network for the discussions that inspired this study. We thank Maria Eduarda Camargo, Barbara Pachêco, and Fabricia Costa for kindly sharing the data. We thank Maria Eduarda Camargo, Camila Motta, and Daniel Vieira for the insightful conversations that helped interpret the data.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1045591/full#supplementary-material
Footnotes
References
Amaral, A. G., et al. (2017). Richness pattern and phytogeography of the Cerrado herb–shrub flora and implications for conservation. J. Veg. Sci. 28, 848–858. doi: 10.1111/jvs.12541
Atkinson, R. J., et al. (2021). Seeding resilient restoration: an indicator system for the analysis of tree seed systems. Diversity 13, 1–13. doi: 10.3390/d13080370
Bosshard, E., Jalonen, R., Kanchanarak, T., Yuskianti, V., Tolentino, E. Jr., Warrier, R. R., et al. (2021). Are tree seed Systems for Forest Landscape Restoration fit for purpose? An analysis of four Asian countries. Diversity 13:575. doi: 10.3390/d13110575
Bridgewater, S., Ratter, J. A., and Ribeiro, J. F. (2004). Biogeographic patterns, −diversity and dominance in the cerrado biome of Brazil. Biodivers. Conserv. 13, 2295–2317. doi: 10.1023/B:BIOC.0000047903.37608.4c
Buisson, E., Fidelis, A., Overbeck, G. E., Schmidt, I. B., Durigan, G., Young, T. P., et al. (2021). A research agenda for the restoration of tropical and subtropical grasslands and savannas. Restor. Ecol. 29:13292. doi: 10.1111/rec.13292
Caminhos da Semente (2020) Mapa de restauração ecológica com semeadura direta no Brasil. Available at: https://www.caminhosdasemente.org.br/mapa (Accessed July 28, 2021).
Campos, L., Freire Moro, M., Funk, V. A., and Roque, N. (2019). Biogeographical review of Asteraceae in the Espinhaço Mountain range, Brazil. Bot. Rev. 85, 293–336. doi: 10.1007/s12229-019-09216-9
Carvalho, G. (2020). ‘Flora: tools for interacting with the Brazilian Flora 2020’. Available at: https://cran.r-project.org/package=flora (Accessed September 09, 2021).
Cava, M. G. B., Pilon, N. A. L., Ribeiro, M. C., and Durigan, G. (2018). Abandoned pastures cannot spontaneously recover the attributes of old-growth savannas. J. Appl. Ecol. 55, 1164–1172. doi: 10.1111/1365-2664.13046
Coutinho, A. G., Alves, M., Sampaio, A. B., Schmidt, I. B., and Vieira, D. L. M. (2019). Effects of initial functional-group composition on assembly trajectory in savanna restoration. Appl. Veg. Sci. 22, 61–70. doi: 10.1111/avsc.12420
Crouzeilles, R., Santiami, E., Rosa, M., Pugliese, L., Brancalion, P. H. S., Rodrigues, R. R., et al. (2019). There is hope for achieving ambitious Atlantic Forest restoration commitments. Perspect. Ecol. Conserv. 17, 80–83. doi: 10.1016/j.pecon.2019.04.003
de Oliveira Joaquim, E., Moura Silva, T., Leone Figueiredo-Ribeiro, R. C., Gomes de Moraes, M., and Machado de Carvalho, M. A. (2018). Diversity of reserve carbohydrates in herbaceous species from Brazilian campo rupestre reveals similar functional traits to endure environmental stresses. Flora 238, 201–209. doi: 10.1016/j.flora.2017.01.001
de Souza, G. F., Almeida, R. F., Bijos, N. R., Fagg, C. W., and Munhoz, C. B. R. (2021). Herbaceous-shrub species composition, diversity and soil attributes in moist grassland, shrub grassland and savanna in Central Brazil. Rev. Bras. Bot. 44, 227–238. doi: 10.1007/s40415-020-00672-x
Erickson, V. J., and Halford, A. (2020). Seed planning, sourcing, and procurement. Restor. Ecol. 28, S216–S224. doi: 10.1111/rec.13199
Flora do Brasil (2020) List of species of the Brazilian Flora, Jardim Botânico do Rio de Janeiro. Available at: http://floradobrasil.jbrj.gov.br/ (Accessed July 27, 2021).
Fontenele, H. G. V., Cruz-Lima, L. F. S., Pacheco-Filho, J. L., and Miranda, H. S. (2020). Burning grasses, poor seeds: post-fire reproduction of early-flowering Neotropical savanna grasses produces low-quality seeds. Plant Ecol. 221, 1265–1274. doi: 10.1007/s11258-020-01080-7
Françoso, R. D., Dexter, K. G., Machado, R. B., Pennington, R. T., Pinto, J. R. R., Brandão, R. A., et al. (2020). Delimiting floristic biogeographic districts in the Cerrado and assessing their conservation status. Biodivers. Conserv. 29, 1477–1500. doi: 10.1007/s10531-019-01819-3
Fremout, T., Thomas, E., Taedoumg, H., Briers, S., Gutiérrez-Miranda, C. E., Alcázar-Caicedo, C., et al. (2022). Diversity for restoration (D4R): guiding the selection of tree species and seed sources for climate-resilient restoration of tropical forest landscapes. J. Appl. Ecol. 59, 664–679. doi: 10.1111/1365-2664.14079
Giles, A. L., Costa, P. B., Rowland, L., Abrahão, A., Lobo, L., Verona, L., et al. (2022). How effective is direct seeding to restore the functional composition of neotropical savannas? Restor. Ecol. 30, 1–13. doi: 10.1111/rec.13474
Giles, A. L., Flores, B. M., Rezende, A. A., Weiser, V. L., and Cavassan, O. (2021). Thirty years of clear-cutting maintain diversity and functional composition of woody-encroached Neotropical savannas. For. Ecol. Manag. 494:119356. doi: 10.1016/j.foreco.2021.119356
Guidotti, V., Freitas, F. L. M., Sparovek, G., Pinto, L. F. G., Hamamura, C., Carvalho, T., et al. (2017). “Números detalhados do novo código florestal e suas implicações para os PRAs,” in Sustentabilidade em debate (Piracicaba: Imaflora), 10.
Karney, C. F. F. (2013). Algorithms for geodesics. J. Geod. 87, 43–55. doi: 10.1007/s00190-012-0578-z
le Stradic, S., Silveira, F. A. O., Buisson, E., Cazelles, K., Carvalho, V., and Fernandes, G. W. (2015). Diversity of germination strategies and seed dormancy in herbaceous species of campo rupestre grasslands. Austral Ecol 40, 537–546. doi: 10.1111/aec.12221
León-Lobos, P., Bustamante-Sánchez, M. A., Nelson, C. R., Alarcón, D., Hasbún, R., Way, M., et al. (2020). Lack of adequate seed supply is a major bottleneck for effective ecosystem restoration in Chile: friendly amendment to Bannister et al. (2018). Restor. Ecol. 28, 277–281. doi: 10.1111/rec.13113
Mapbiomas (2021) Projeto MapBiomas Alerta–v.5.0–Sistema de Validação e Refinamento de Alertas de Desmatamento com Imagens de Alta Resolução. Available at: https://mapbiomas.org/ (Accessed June 25, 2021).
Motta, C. P. (2017). Dinâmica populacional de uma gramínea invasora e um arbusto nativo: implicações para a restauração ecológica no Cerrado. Brasília: Universidade de Brasília.
Munhoz, C., and Araújo, G. M. (2011). “Métodos de Amostragem do Estrato Herbáceo-subarbustivo,” in Fitossociologia no Brasil. eds. J. M. Felfili, P. V. Eisenlohr, M. M. R. F. Melo, L. A. Andrade, and J. A. A. Meira Neto (Viçosa: Editora UFV), 1st edn. 213–230.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Nef, D. P., Gotor, E., Wiederkehr Guerra, G., Zumwald, M., and Kettle, C. J. (2021). Initial Investment in Diversity is the efficient thing to do for resilient Forest landscape restoration. Frontiers in Forests and Global Change. 3, 1–10. doi: 10.3389/ffgc.2020.615682
Neto, L. M., and Forzza, R. C. (2013). Biogeography and conservation status assessment of Pseudolaelia (Orchidaceae). Bot. J. Linn. Soc. 171, 191–200. doi: 10.1111/j.1095-8339.2012.01304.x
Nogueira, E. V., Bijos, N. R., Trindade, V. L., Heusi, G. P., Togni, P. H. B., and Munhoz, C. B. R. (2022). Differences in soil properties influence floristic changes in the Veredas of the Brazilian Cerrado. Braz. J. Bot. 45, 763–774. doi: 10.1007/s40415-022-00795-3
Overbeck, G. E., Vélez-Martin, E., Menezes, L. S., Anand, M., Baeza, S., Carlucci, M. B., et al. (2022). Placing Brazil’s grasslands and savannas on the map of science and conservation. Perspect. Plant Ecol. Evol. Syst. 56:125687. doi: 10.1016/j.ppees.2022.125687
Palma, A. C., and Laurance, S. G. W. (2015). A review of the use of direct seeding and seedling plantings in restoration: what do we know and where should we go? Appl. Veg. Sci. 18, 561–568. doi: 10.1111/avsc.12173
Parr, C. L., Lehmann, C. E. R., Bond, W. J., Hoffmann, W. A., and Andersen, A. N. (2014). Tropical grassy biomes: misunderstood, neglected, and under threat. Trends Ecol. Evol. 29, 205–213. doi: 10.1016/j.tree.2014.02.004
Pebesma, E. (2018). Simple features for R: standardized support for spatial vector data. R J. 10, 439–446. doi: 10.32614/RJ-2018-009
Pedrini, S., and Dixon, K. W. (2020). International principles and standards for native seeds in ecological restoration. Restor. Ecol. 28, S286–S303. doi: 10.1111/rec.13155
Pedrini, S., Gibson-Roy, P., Trivedi, C., Gálvez-Ramírez, C., Hardwick, K., Shaw, N., et al. (2020). Collection and production of native seeds for ecological restoration. Restor. Ecol. 28, S228–S238. doi: 10.1111/rec.13190
Pellizzaro, K. F., Cordeiro, A. O. O., Alves, M., Motta, C. P., Rezende, G. M., Silva, R. R. P., et al. (2017). “Cerrado” restoration by direct seeding: field establishment and initial growth of 75 trees, shrubs and grass species. Rev. Bras. Bot. 40, 681–693. doi: 10.1007/s40415-017-0371-6
Pilon, N. A. L., Assis, G. B., Souza, F. M., and Durigan, G. (2019). Native remnants can be sources of plants and topsoil to restore dry and wet cerrado grasslands. Restor. Ecol. 27, 569–580. doi: 10.1111/rec.12902
Pilon, N. A. L., Cava, M. G. B., Hoffmann, W. A., Abreu, R. C. R., Fidelis, A., and Durigan, G. (2021). The diversity of post-fire regeneration strategies in the cerrado ground layer. The Journal of ecology 109, 154–166. doi: 10.1111/1365-2745.13456
Pinto, S., Melo, F., Tabarelli, M., Padovesi, A., Mesquita, C., de Mattos Scaramuzza, C., et al. (2014). Governing and delivering a biome-wide restoration initiative: the case of Atlantic Forest restoration pact in Brazil. Forests 5, 2212–2229. doi: 10.3390/f5092212
R Core Team (2021) ‘R: A language and environment for statistical computing’. Vienna, Austria. Available at: https://www.r-project.org/ (Accessed August 12, 2022).
Rabeling, S. C., Lim, J. L., Tidon, R., Neff, J. L., Simpson, B. B., and Pawar, S. (2019). Seasonal variation of a plant-pollinator network in the Brazilian Cerrado: implications for community structure and robustness. PLoS One 14, 1–22. doi: 10.1371/journal.pone.0224997
Ratter, J. A., Bridgewater, S., Atkinson, R., and Ribeiro, J. F. (1996). Analysis of the floristic composition of the Brazilian cerrado vegetation II: comparison of the woody vegetation of 98 areas. Edinb. J. Bot. 53, 153–180. doi: 10.1017/s0960428600002821
Raupp, P. P., Ferreira, M. C., Alves, M., Campos-Filho, E. M., Sartorelli, P. A. R., Consolaro, H. N., et al. (2020). Direct seeding reduces the costs of tree planting for forest and savanna restoration. Ecol. Eng. 148:105788. doi: 10.1016/j.ecoleng.2020.105788
Reflora (2020) Brazilian Flora 2020. Available at: http://floradobrasil.jbrj.gov.br/reflora/PrincipalUC/PrincipalUC.do (Accessed October 6, 2021).
Sampaio, A. B., Vieira, D. L. M., Holl, K. D., Pellizzaro, K. F., Alves, M., Coutinho, A. G., et al. (2019). Lessons on direct seeding to restore Neotropical savanna. Ecol. Eng. 138, 148–154. doi: 10.1016/j.ecoleng.2019.07.025
Schmidt, I. B., Urzedo, D. I., Piña-Rodrigues, F. C. M., Vieira, D. L. M., Rezende, G. M., Sampaio, A. B., et al. (2019). Community-based native seed production for restoration in Brazil – the role of science and policy. Plant Biol. 21, 389–397. doi: 10.1111/plb.12842
Shaw, N., Barak, R. S., Campbell, R. E., Kirmer, A., Pedrini, S., Dixon, K., et al. (2020). Seed use in the field: delivering seeds for restoration success. Restor. Ecol. 28, S276–S285. doi: 10.1111/rec.13210
Silva, T. R., Pena, J. C., Martello, F., Bettiol, G. M., Sano, E. E., and Vieira, D. L. M. (2021). Not only exotic grasslands: the scattered trees in cultivated pastures of the Brazilian Cerrado. Agric. Ecosyst. Environ. 314:107422. doi: 10.1016/j.agee.2021.107422
Silveira, F. A. O., Ordóñez-Parra, C. A., Moura, L. C., Schmidt, I. B., Andersen, A. N., Bond, W., et al. (2022). Biome awareness disparity is BAD for tropical ecosystem conservation and restoration. J. Appl. Ecol. 59, 1967–1975. doi: 10.1111/1365-2664.14060
Strassburg, B. B. N., Brooks, T., Feltran-Barbieri, R., Iribarrem, A., Crouzeilles, R., Loyola, R., et al. (2017). Moment of truth for the Cerrado hotspot. Nat. Ecol. Evol. 1:0099. doi: 10.1038/s41559-017-0099
Suding, K. N. (2011). Toward an era of restoration in ecology: successes, failures, and opportunities ahead. Annu. Rev. Ecol. Evol. Syst. 42, 465–487. doi: 10.1146/annurev-ecolsys-102710-145115
Teixeira, J., Souza, L., le Stradic, S., and Fidelis, A. (2022). Fire promotes functional plant diversity and modifies soil carbon dynamics in tropical savanna. Sci. Total Environ. 812:152317. doi: 10.1016/j.scitotenv.2021.152317
Teodoro, G. S., Lambers, H., Nascimento, D. L., de Britto Costa, P., Flores-Borges, D. N. A., Abrahão, A., et al. (2019). Specialized roots of Velloziaceae weather quartzite rock while mobilizing phosphorus using carboxylates. Funct. Ecol. 33, 762–773. doi: 10.1111/1365-2435.13324
Urzedo, D., Pedrini, S., Vieira, D. L. M., Sampaio, A. B., Souza, B. D. F., Campos-Filho, E. M., et al. (2022). Indigenous and local communities can boost seed supply in the UN decade on ecosystem restoration. Ambio 51, 557–568. doi: 10.1007/s13280-021-01593-z
Urzedo, D., Piña-Rodrigues, F., Feltran-Barbieri, R., Junqueira, R., and Fisher, R. (2020). Seed networks for upscaling Forest landscape restoration: is it possible to expand native plant sources in Brazil? Forests 11:259. doi: 10.3390/f11030259
Wolfsdorf, G., Abrahão, A., D'Angioli, A. M., de Sá Dechoum, M., Meirelles, S. T., F. L. Pecoral, L., et al. (2021). Inoculum origin and soil legacy can shape plant–soil feedback outcomes for tropical grassland restoration. Restor. Ecol. 29:2. doi: 10.1111/rec.13455
Zanetti, M., Dayrell, R. L. C., Wardil, M. V., Damasceno, A., Fernandes, T., Castilho, A., et al. (2020). Seed functional traits provide support for ecological restoration and ex situ conservation in the threatened Amazon ironstone outcrop Flora. Front. Plant Sci. 11:599496. doi: 10.3389/fpls.2020.599496
Keywords: ecosystem restoration, restoration planning, community-based seed production, old-growth savannas, tropical grassy biomes, open ecosystems
Citation: Silva MC, Moonlight P, Oliveira RS, Pennington RT and Rowland L (2022) Toward diverse seed sourcing to upscale ecological restoration in the Brazilian Cerrado. Front. Ecol. Evol. 10:1045591. doi: 10.3389/fevo.2022.1045591
Edited by:
Jasper A. Slingsby, University of Cape Town, South AfricaReviewed by:
Stefan Siebert, North-West University, South AfricaCésar Marín, Santo Tomás University, Chile
Copyright © 2022 Silva, Moonlight, Oliveira, Pennington and Rowland. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mateus C. Silva, bXMxMTkwQGV4ZXRlci5hYy51aw==
†ORCID: Mateus C. Silva, https://orcid.org/0000-0002-4281-3400
Peter Moonlight, https://orcid.org/0000-0003-4342-2089
Rafael S. Oliveira, https://orcid.org/0000-0002-6392-2526
R. Toby Pennington, https://orcid.org/0000-0002-8196-288X
Lucy Rowland, https://orcid.org/0000-0002-0774-3216