 Mounir Louhaichi
Mounir Louhaichi- 1Rangeland Ecology and Forages (REF), International Center for Agricultural Research in the Dry Areas (ICARDA), Tunis, Tunisia
- 2Department of Animal and Range Science, Oregon State University, Corvallis, OR, United States
- 3Office of Livestock and Pasture, Tunis, Tunisia
The aim of this study was to assess the impact of short grazing periods with high-stocking density on vegetation during weather conditions favorable to plant growth. Continuous grazing is widely practiced in Tunisian arid rangelands and across most drylands and deserts of the Middle East and North Africa. In the early 1990s, the Tunisian government combined a variety of incentives and restrictions on rangeland grazing practices and initiated a national strategy for rangeland improvement. The strategy emphasizes grazing exclusion for three consecutive years, a practice known locally as gdel. At the end of this period, grazing is allowed with no restrictions or with some guidelines. However, these regulations created discontent among pastoral communities, mainly when local rainfall conditions result in a considerable quantity of green biomass. High stocking-density grazing for a short period would help satisfy pastoralist concerns and achieve the government’s strategic goal. This study was implemented in three arid rangeland types of southern Tunisia that have been under restricted grazing for 2 years. Each area in the study was grazed for short periods in late May (about 7 days) with a flock of 150 head of sheep. Measurements of vegetation cover, forage productivity, density, and species richness were taken before and after grazing. Our results suggest that perennial vegetation is more stable under grazing than annual vegetation. The presence of annual species would enhance rangeland vegetation cover and diversity and at the same time offers an opportunity for livestock to select a high-quality diet rich in protein. Opportunistic grazing, applied to heterogeneous rangelands in late spring and the beginning of the annual temperature increase, encouraged animals to select annual plants due to their higher palatability, higher digestibility, and water content rather than heavily grazing perennial species. Perennial biomass materials that die and do not fall through rapid biological decay tended to decline with increased period of placement, resulting in further oxidation to CO2 which affects photosynthetic performance negatively and may eventually cause plant death. Trampling from high-density stocking enhances the litterfall rates and removes the oxidized plant material. These findings may contribute to strategies for addressing the extreme climatic variations that threaten rangeland and livestock sustainability.
Introduction
Rangelands cover nearly 45% of the world’s land area (Squires et al., 2018). As a common resource, they are mainly used for livestock grazing supporting 50% of the world’s livestock and 78% of global grazing area (Asner et al., 2004; MEA, 2005; Creamer and Horback, 2021). Despite the vital role of rangelands in sustaining the livelihoods of rural people, they are subject to human and climate drivers that can result in reduced production (Jama and Zeila, 2005; Ouled Belgacem and Louhaichi, 2013). Rangelands are especially vulnerable as semi-arid ecosystems experience extreme stress associated with low rainfall (Gamoun, 2016), prolonged periods of drought (Köchy et al., 2008; Gamoun et al., 2011), poor soil fertility, mismanagement, and other human-induced activities (Hanafi and Jauffret, 2008; Vetter, 2009; Zhou et al., 2013). All these factors negatively impact rangeland plant communities (Ouled Belgacem et al., 2013; Belgacem et al., 2019) and lead to changes in soil-plant-water dynamics (Fay et al., 2008; Heisler-White et al., 2009), sparse vegetation, and increases in the spread of invasive species (McNeely, 2004; Wang et al., 2018).
Rangelands offer habitats for wildlife and provide multiple goods and services of economic value (Yu et al., 2010). However, as rangelands become degraded, their potential to provide these services diminishes (Favretto et al., 2016). Overgrazing remains the primary anthropogenic factor impacting arid and semi-arid vegetation (Smet and Ward, 2005; Gamoun, 2013; Louhaichi et al., 2019). Associated with overstocking and drought, overgrazing further contributes to degradation resulting in reduction in biodiversity and rangeland productivity (Harris, 2010; Squires, 2010; Teague et al., 2011; Gamoun et al., 2016).
The impact of grazing on rangelands depends mainly on its intensity and timing (Holechek et al., 1998; Henkin et al., 2007). For example, light to moderate grazing may not affect productivity or species richness while heavy grazing often reduces vegetation by preferential removal of the more palatable species preferred by grazing animals (Louhaichi et al., 2009; Schönbach et al., 2009; Gamoun, 2014). Under heavy grazing, shrubs develop very dense small branches that protect their leaves and allow continued plant growth (van Duivenbooden, 1993).
In the late 1960s, the concept of high-stocking-density grazing for short periods was first introduced in North America by Goodloe (1969) and further developed by Savory in the 1980s (Savory, 1978). It quickly became a widely popular but controversial tool for grazing management. In this grazing scheme, a rangeland is divided into several paddocks grazed one at a time, allowing plants enough time to recover (Goodloe, 1969; Savory, 1978; Joseph et al., 2002). Another widespread grazing strategy is herd mobility. Livestock mobility is based on a deep knowledge of rangeland vegetation and is referred to as “opportunistic grazing” (Behnke et al., 1993; Scoones, 1995). This type of grazing management is often independent of stocking density and is guided more by inter-annual rainfall variation. Through their pastoral networks, herders know where the best rangelands are and when to use them (Bassett and Koné, 2006). Eventually, opportunistic grazing formalizes optimal relationships between livestock and the environment (McCabe, 2004) and has proven to be effective in terms of enhancing livestock performance and vegetation productivity (Suttie et al., 2005).
Rangelands in some areas of Tunisia continue to degrade with no restrictions on stocking rate (Belgacem et al., 2019). Their future sustainability will depend on identifying and promoting sustainable rangeland management practices that are environmentally sustainable and socially acceptable to local pastoral communities. Toward this end, the Government of Tunisia launched a national strategy for rangeland improvement in 1990. Since then, thousands of hectares covering natural rangelands, including private and collective lands, have been managed by simply excluding livestock grazing for 3 years. The program has played a paramount role in alleviating pressure on already fragile degraded arid rangelands, especially during dry years. Nevertheless, during years with sufficient rainfall, vegetation response to the wet season is phenomenal, even after only a few months of protection (Sullivan and Rohde, 2002).
Unfortunately, banning livestock grazing for three consecutive years has created conflict between administrations in charge of implementing the strategy and pastoral communities, especially when local rainfall conditions produce a considerable quantity of green biomass suitable for opportunistic livestock grazing.
To improve the current policy and procedures and to allow more flexibility in grazing practices when certain conditions are met, this study was undertaken to assess the effect of short-duration, high-stocking-rate opportunistic grazing on the arid rangelands of southern Tunisia. The findings of this study may prove useful to policymakers who make recommendations to national and international development agencies to enhance livestock production while conserving the natural resource base.
Methodology
Site Description
Tataouine Governorate is located in the extreme south of Tunisia and 38% of the land area is rangeland (1.5 million ha), which is roughly 27% of the total rangeland area in Tunisia. Rangelands in Tataouine are grazed by more than 1.3 million head of sheep and goats (18% of the total livestock population in Tunisia) with sheep the most dominant species (84.4% sheep, 15.6% goats) and 12,000 head of camels (25% of the total camel population in Tunisia). The study sites were 30 km east of Tataouine. Each site has a dominant plant type. Dominant plant types provide important information about the plant associations growing in one area due to factors that affect their growth and characteristics. This dominant plant species is often used to name the plant community and to indicate the rangelands’ site potentials. In this study, the target plant communities were classified as Anthyllis henoniana, Haloxylon schmittianum, and Retama raetam. Annual precipitation averaged over 80 mm with recorded extremes of 253 mm during 2017–2018 and 120 mm during 2018–2019 (Figure 1). Mean daily temperatures in the coldest (January) and warmest (August) months are 11 and 33°C, respectively. The major soils are Regosols and Sierozems. For over half a century, these rangelands have been prone to intense human use contributing to the continued deterioration of natural vegetation such that natural recovery is no longer possible (Jauffret and Visser, 2003).
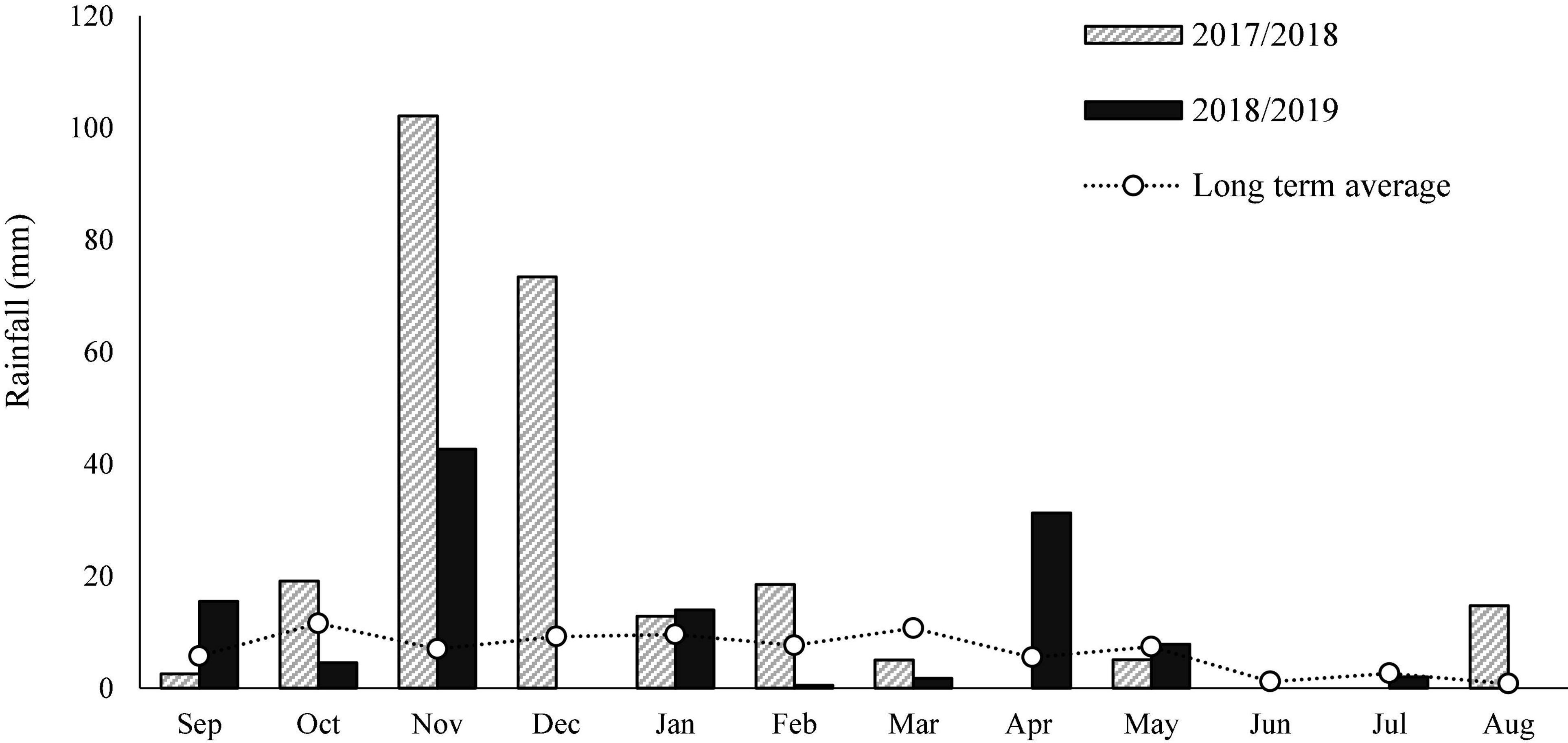
Figure 1. Amount and distribution of rainfall during 2017–2018 and 2018–2019 and the long-term average (20 years) monthly rainfall data in Tataouine, Tunisia.
Methods
The three sampled areas were excluded from livestock grazing for two consecutive years by implementing a resting technique known locally as gdel until the first vegetation sampling in May 2019. In desert environments, plants react quickly when heat and moisture trigger blooming. After a rainfall, annual species flower within a few weeks and seed pods burst and expel their seeds before the branches dry out.
Grazing on the study sites began toward the end of spring, just after seed dispersal and before plants began to dry out and become less nutritious for grazing animals. After the first sampling, livestock were allowed to graze for 1 week in late May. Each six-hectare paddock was grazed using a fixed density of 150 head of sheep. Measurements were taken before and after grazing using the line intersect method as described by Daget and Poissonet (1971).
To provide a more detailed understanding of the effects of overgrazing, for each plant community, there was a control site that was subjected to continuous grazing. However, since all these control sites were homogeneous in structure and dominated by different annuals with some perennial species, and due to heavy continuous grazing, the vegetation in these sites becomes sparsely distributed over time, resulting in a pattern of similar vegetation patches alternating with areas of bare soil, which made it difficult to identify reference vegetation types. Three permanent transects (50 m length) were laid randomly within each plant community and control sites. A pin was inserted into the ground at 50-cm intervals along the transect tape. At each of the 100 points on the line, the plant species were recorded. The percentage cover of each plant was obtained by dividing the number of points at which a particular species was encountered by the total number of samples taken for all species and multiplying by 100. Because habitat heterogeneity had a positive influence on species density, annual density was estimated by establishing nine quadrants (each 1 m2) per plant community and nine were placed in the continuously grazed sites. In sparsely vegetated communities, larger quadrats were required with the possibility of reduction of replication for perennial density measurement. Perennial plant density was determined by counting the number of tufts of each species within three quadrants of 50 m2 per plant community and three in each continuously grazed site.
In each plant community and continuously grazed site, total botanical composition and species richness were identified and measured by counting the number of species in the area. Plant species cover and the palatability or acceptability index determine rangeland productivity according to the following formula (INRA., 1978):
where P is total rangeland productivity in forage units or FU/ha/year, SCi is the specific cover of species I (%), PFi is the palatability factor of species i, and TPC is the total plant cover (%).
An ANOVA test was performed to determine the effects of the two-grazing systems in each plant community (A. henoniana, H. schmittianum, and R. raetam) with respect to vegetation cover, plant density, rangeland forage productivity, and species richness. To avoid the incidence of pseudo-replication, the transects and the samples taken inside the quadrats on each plant community and continuous grazing sites were considered as replicates for comparison and the sampling design was spatially autocorrelated as the transects were well spaced and sample locations were spatially separated covering an area of ∼60,000 m2 for each plant community and continues grazing site (Hurlbert, 1984; Waite, 2000).
Results
Vegetation Cover
Grazing exclusion led to a significant increase in both perennial and annual species cover in all plant communities (p > 0.05). Grazing exclusion results in up 2.7, 3.6, and 4.7-fold increase (p > 0.05) of perennial species cover in H. schmittianum, A. henoniana, and R. raetam plant communities, respectively, compared to their paired open grazing sites (controls), however, high-density grazing for a short period had no significant effect on the cover of perennial species for all plant communities and the perennials cover remained unchanged compared to before grazing. In the R. raetam community, the perennial grass Cynodon dactylon was consistently the most abundant species and its contribution to the total cover of perennial species reached 28%.
Compared with open grazing sites (controls), all plant communities exhibited significantly greater annual species cover after 2 years grazing exclusion (P < 0.05). Generally, restricted grazing increased the annual spices cover in A. henoniana, R. raetam, and H. schmittianum plant communities (41, 73, and 74%, respectively) compared with their paired open grazed sites. The cover of annual species after a short period of high-density grazing was significantly lower compared to before grazing for all plant communities (p < 0.05). When introducing sheep to graze at a high stocking rate for a short time, they are attracted to annual species. Therefore, the cover of annual species became extremely low in all plant communities, for example, the cover of annual species in the H. schmittianum community decreased from 25% before grazing to 1% after grazing which was much less than under continuous grazing H. schmittianum control site (Figure 2).
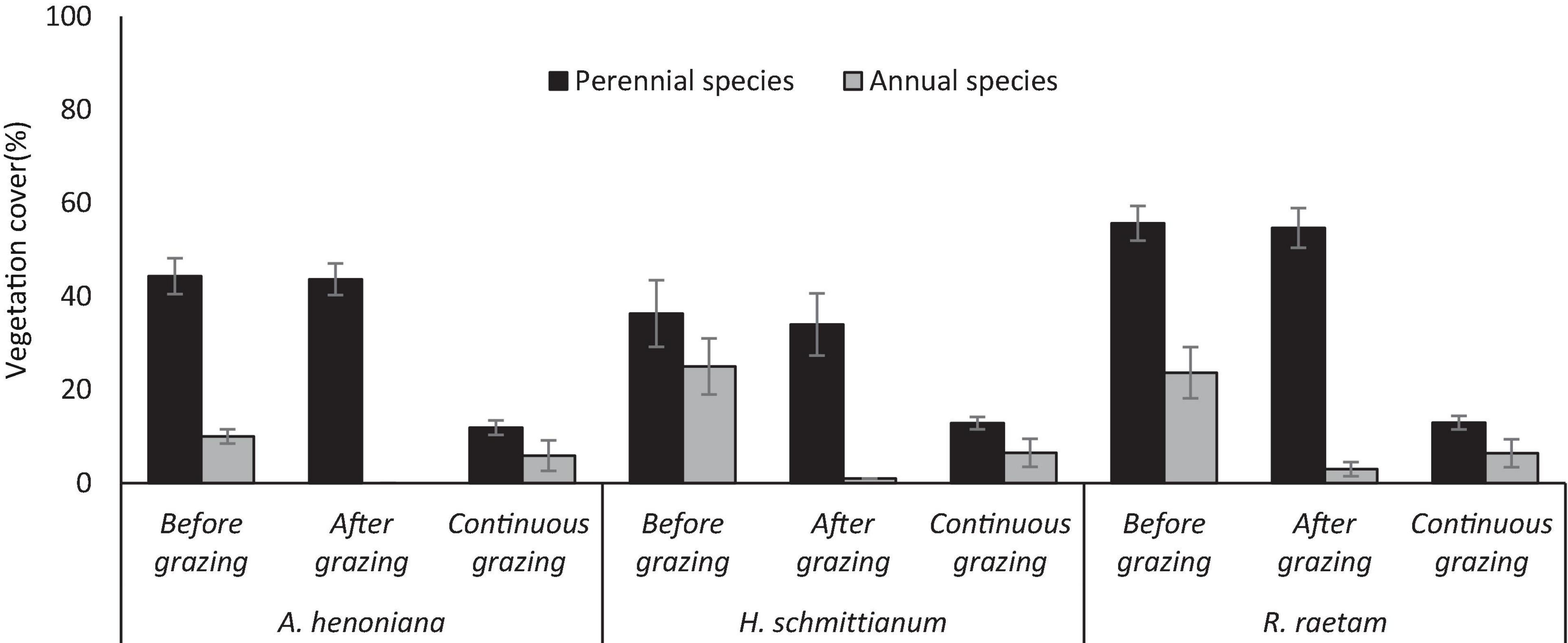
Figure 2. Mean percentage of perennial and annual species vegetation cover in 2 years protected plant communities (A. henoniana, H. schmittianum, and R. raetam) sites before and after subjecting to 1 week of intense grazing compared to continuous grazing sites in Tataouine, Tunisia.
Plant Density
After 2-year grazing exclusion, the values of the density of perennial species in H. schmittianum, A. henoniana, and R. raetam plant communities were 4, 6, and 9 plant m–2 respectively, and were significantly greater than those in their paired continuous grazing sites (controls) 1, 1, and 2 plant m–2 (P < 0.05). Moreover, the density of perennial species was not affected by high-density grazing for a short period and did not differ before and after grazing in all plant communities.
Annual species density increased significantly after 2-year grazing exclusion in all plant community sites. H. schmittianum, and R. raetam plant communities’ annual species density was substantially higher 28 plant m–2 in a 2-year grazing exclusion sites than in their paired continuous grazing sites (controls) 7 plant m–2. Similarly, grazing exclusion increased the annual density in A. henoniana plant community from 7 plant m–2 in continuous grazing site to 9 plant m–2 in grazing-excluded site. However, high-density grazing for a short period significantly affected the annual species density compared to before grazing in all plant communities. For example, the annual density in the H. schmittianum community decreased from 29 plant m–2 before grazing to 3 plant m–2 species after high-density grazing for a short period, while the annual density in continuous grazing site was 7 plant m–2 (Figure 3).
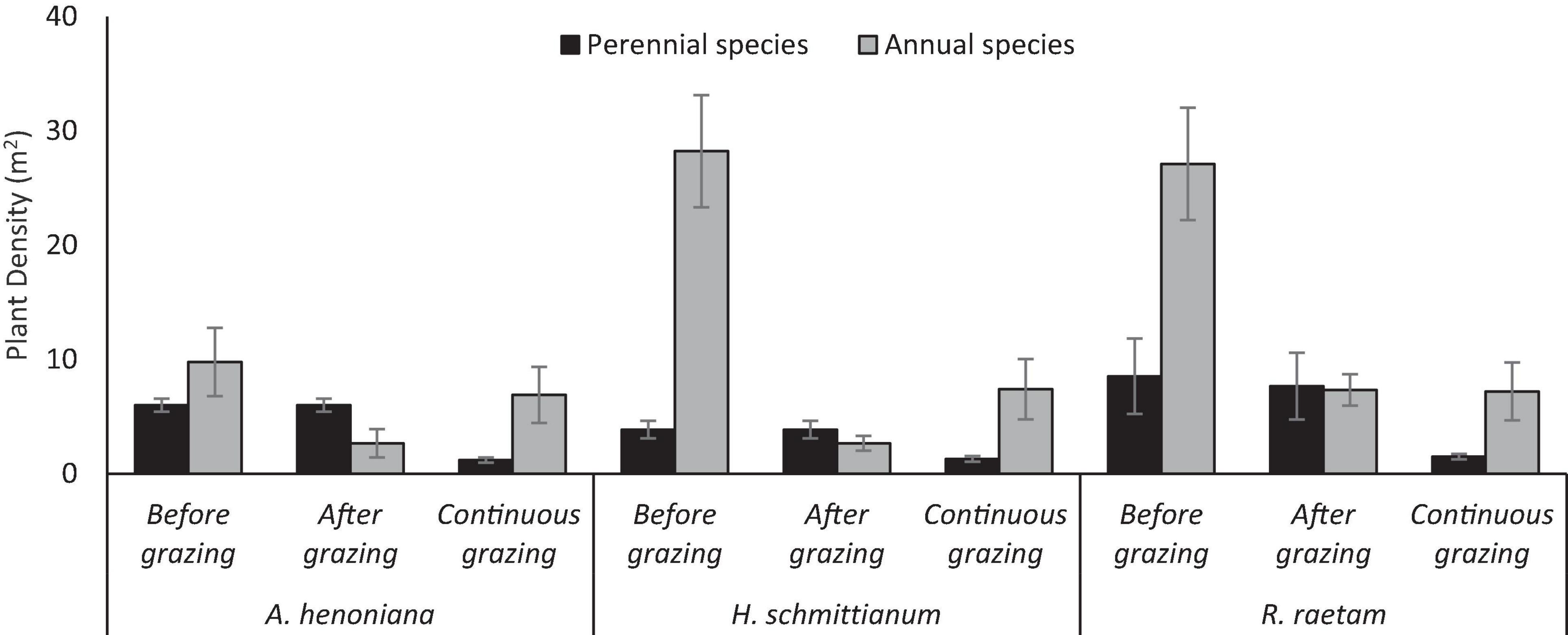
Figure 3. Perennial and annual species density in 2 years protected plant communities (A. henoniana, H. schmittianum, and R. raetam) sites before and after subjecting to 1 week of intense grazing compared to continuous grazing sites in Tataouine, Tunisia.
Rangeland Forage Productivity
Forage productivity varied significantly among plant community types and grazing systems (p < 0.05). Yet, there was no significant interaction between grazing systems and plant community types (p = 0.363). The forage productivity was notably higher after 2-year grazing exclusion of all plant communities than those continuous grazing sites. Forage productivity was the highest at R. raetam plant community, followed by A. henoniana plant community. Accordingly, R. raetam forage productivity of grazing-excluded site (341 FU/ha/year) was significantly higher compared to that of continuous grazing site (31 FU/ha/year). In A. henoniana plant community, mean forage productivity of grazing-excluded site was approximately 87% higher than that of continuous grazing site (control). Nevertheless, although forage productivity of all plant communities has seen a slight decline, the high-density grazing for a short period had no significant effect on rangeland forage productivity. In spite of the significantly higher stocking rate combined with the short-duration grazing system, R. raetam community recorded higher forage productivity at 274 FU/ha/year compared to other plant communities before and after grazing (Figures 4, 5).

Figure 4. Rangeland forage productivity (FU/ha/year) in 2 years protected plant communities (A. henoniana, H. schmittianum, and R. raetam) sites before and after subjecting to 1 week of intense grazing compared to continuous grazing sites in Tataouine, Tunisia.

Figure 5. Perennial grass (Cynodon dactylon) conditions before (A) and after (B) 1-week intense grazing in Tataouine, Tunisia.
Species Richness
Species richness of perennial species did differ greatly among plant community types. The highest proportion of species richness of perennial species was recorded in A. henoniana, followed by R. raetam (Figure 6). Grazing exclusion increased species richness in all plant community sites and had a positive effect on number of perennial species. The differences in perennial species richness between grazing-excluded sites and continuous grazing sites (controls) recorded 12, 8, and 3 species in A. henoniana, R. raetam, and H. schmittianum plant communities, respectively, (Supplementary Appendices 1, 2). In contrast, high-density grazing for a short period did not affect species richness of perennial species for all plant communities either positively or negatively.
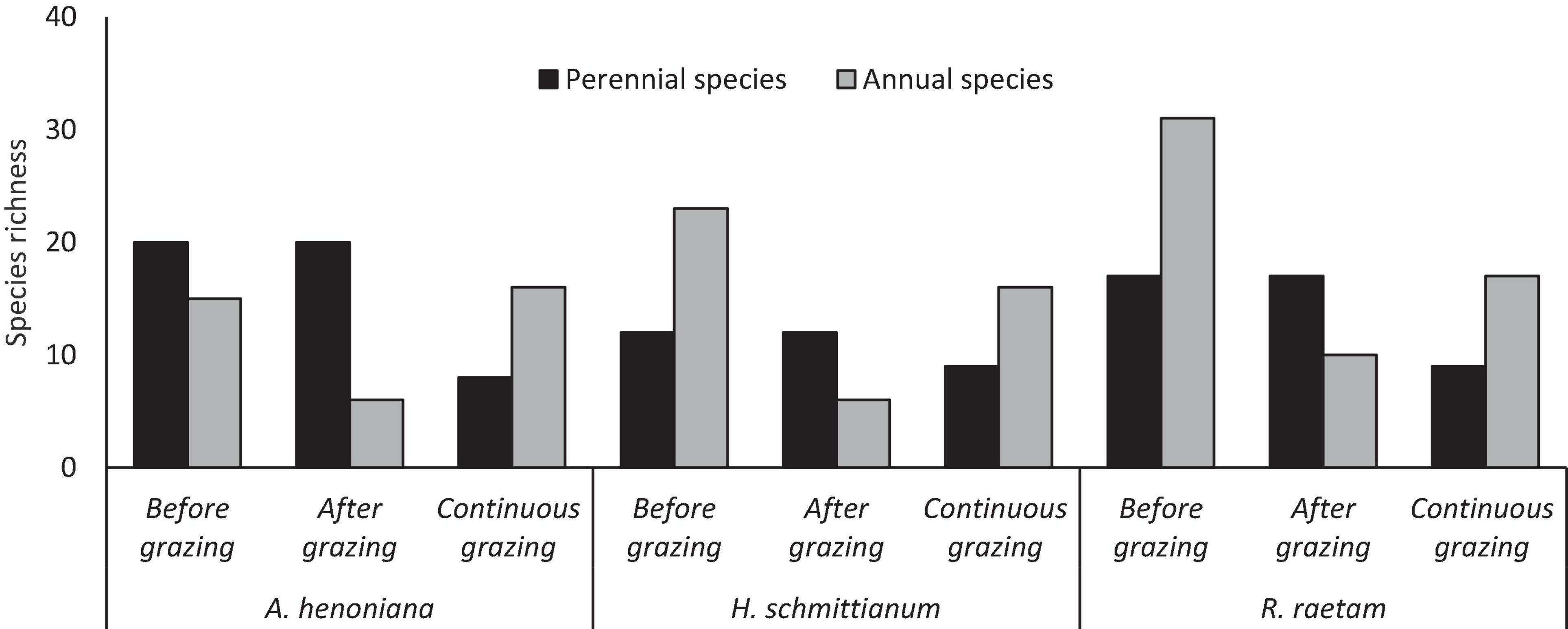
Figure 6. Species richness of perennial and annual species in 2 years protected plant communities (A. henoniana, H. schmittianum, and R. raetam) sites before and after subjecting to 1 week of intense grazing compared to continuous grazing sites in Tataouine, Tunisia.
The species richness of annual plants was affected significantly among plant community types and grazing systems (Figure 6). The annual vegetation was highly diverse in the R. raetam community, with 31 species identified before livestock grazing followed by H. schmittianum and A. henoniana (22 and 15 species respectively; Supplementary Appendix 3). The grazing exclusion increased the annual species numbers in R. raetam and H. schmittianum by 14 and 7 annual species when compared with their paired continuous grazing sites (controls). However, A. henoniana annual species number remained the same in both grazing-excluded site and continuous grazing site (control) without any hint of an exclosure effect (Figure 6). Compared to continuous grazing, annual plants were more sensitive and vulnerable to high-density grazing for a short period. This practice decreased the proportion of annual species. The annual vegetation diverse decreased in the R. raetam community to 10 species and to six annual species in both A. henoniana and H. schmittianum plant communities after livestock grazing at high stock-density for a short period (Supplementary Appendix 3).
Discussion
Rangeland management strategies can be used as a tool to improve sustainability (Louhaichi et al., 2019, 2021a). A growing number of studies have shown the positive contributions of grazing and how it can be a cost-effective practice for enhancing productivity, species diversity, and ecosystem health (Hodgson et al., 2005; Dorrough et al., 2007; Kohyani et al., 2008). This study highlighted the effects of a short period of high-density grazing on vegetation cover, rangeland productivity, plant density, and species richness. As found in previous studies, these results showed that continuous grazing was linked with poor rangeland conditions, scattered vegetation, low forage productivity, and a low level of diversity.
Grazing livestock at a high density for a short period provides different results in late spring. Under intensive short-duration grazing, there were no effects on perennial shrub species but a high reduction in annual species. The abundance of annuals increased rapidly during spring in the rested (protected) areas. During late spiring period, grazing animals focus mainly on annuals and rarely select woody species, particularly if they are less palatable. Allowing animals to browse the leaves of perennial species does not retard plant growth because they retain the capacity to provide enough energy for further growth and strengthening their root system reserves (Le Floc’h, 2001).
In general, green forage in late spring is high quality, particularly for annual plants. As summer progresses and temperatures increases, herbaceous plants begin to dry out and lose nutritional value. As plant growth progresses, high forage-quality indicators such as protein, energy, vitamins, and minerals decline. As forage plants mature, indicators of low quality or feed value such as fiber and lignin increase (George et al., 2001). The digestibility of annual plants declines with progression through flowering to seed-set and senescence (Kemp and Michalk, 1993), although Louhaichi et al. (2021b) found that the indigenous Tunisian range species contain high nutritional value due to moderate to high protein, low fiber, and high digestibility content and can be used as feed for livestock to enhance local livestock production and contribute to economic development.
For rangelands rested for 2 years, late spring grazing at high-density stocking rates for a short period had no effect on cover, density, and richness of perennial species. Delayed grazing after maturation of forage species and seed dispersal in late spring provides an opportunity for flash-grazing annual species. Skipping woody and perennial species provides an opportunity for selective grazing of annual species. It is well known that sheep are more selective of annual and herbaceous species in their diet than goats for example, which are more likely to browse perennial species (Yiakoulaki and Papanastasis, 2005).
The grazing regimes in this experiment were not detrimental to rangeland productivity because they reduced selectivity, which encouraged the growth of palatable species and enhanced rangeland diversity. Martin (1975) also reported that heavy grazing for a short period results in a more even use of forage, both from place to place and among different species. The grazing and trampling resulting from high-density grazing for a short period did not favor perennial over annual species. This is important because long-term sustained rangeland productivity is strongly dependent on conserving key perennial vegetation (Curry and Hacker, 1990). Grazing and trampling can improve rangeland conditions and plants may stay upright for a long time by removing accumulated dead forage due to senescence or disturbances such as drought and heavy grazing (Heukes and Cowling, 2000). A few months of rest immediately after each period of late spring-early summer grazing can be expected to reduce the effect of intensity of grazing and regrazing on favored plants in the spring.
The critical timing for high-intensity, short-duration grazing may vary from year to year and is influenced by multiple factors, including precipitation, temperature, and vegetation type. Therefore, it is important to base grazing plans on the state of the vegetation cover rather than simply on the length of resting times. This result supports findings highlighting the need for flexible grazing management depending on weather and climate variability (Louhaichi et al., 2021a) and offers an opportunity to develop grazing strategies that exploit these results.
Conclusion
The current national rangeland improvement programs simply ban livestock grazing for a minimum duration of 3 years at a time when sources of supplementary feed to fill feed-gap periods are expensive and where animal demand exceeds rangelands supply. During rainy years, the vegetation response is impressive. Such positive responses do not occur every year as the most common scenario under arid conditions is recurrent drought. Therefore, it would be a lost opportunity for pastoral communities if they are not allowed to use this abundant and rich biomass. From this perspective, development agencies have been keen to revise and update projects and programs for a more flexible approach linking administration with end-user needs based on scientific evidence. The suggested grazing scheme in this study produced satisfactory results compared to protected sites (livestock exclusion) or continuous grazing (control). These findings have implications for the health of the arid rangelands of Tunisia and the livelihoods of pastoral communities. Properly managed, arid rangelands can provide sustainable forage resources that reduce feeding costs and enhance livestock productivity. The findings of this pilot study may offer evidence for designing approaches that could be scaled out to other regions, such as the Middle East and North Africa.
Data Availability Statement
The data analyzed in this study are subject to the following licenses/restrictions: Data are restricted for 5 years. Requests to access these datasets should be directed to ML, m.louhaichi@cgiar.org.
Author Contributions
ML and MG conceptualized the study, did a general literature review, led the formal analysis, and contributed to writing manuscript drafts and revisions. MG and FG collected the data. ML acquired research funding. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Consultative Group on International Agricultural Research (CGIAR) Research Program on Livestock Agri-Food Systems – Livestock and Environment Flagship, agreement number 200173, and the article processing charge (APC) was also funded by the same source of funding.
Author Disclaimer
The views expressed in this article are those of the authors and not necessarily reflect those of ICARDA or the CGIAR.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This work was undertaken by the International Center for Agricultural Research in the Dry Areas (ICARDA) and in collaboration with the Office of Livestock and Pastures (OEP) in Tunisia.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.757764/full#supplementary-material
References
Asner, G. P., Elmore, A. J., Olander, L. P., Martin, R. E., and Harris, A. T. (2004). Grazing systems, ecosystem responses and global change. Annu. Rev. Environ. Resour 29, 261–299.
Bassett, T. J., and Koné, M. (2006). Grazing Lands and Opportunistic Models: The Political Ecology of Herd Mobility in Northern CÔTE d’Ivoire. ACTES du Colloque International Les Frontières de la Question Foncière – At the Frontier of Land Issues. Montpellier: International Union for Conservation of Nature.
Behnke, R. H., Scoones, I., and Kerven, C. (1993). Range Ecology at Disequilibrium: New Models of Natural Variability and Pastoral Adaptation in African Savannas. London: Overseas Development Institute.
Belgacem, A. O., Ben Salem, F., Gamoun, M., Chibani, R., and Louhaichi, M. (2019). Revival of traditional best practices for rangeland restoration under climate change in the dry areas: a case study from Southern Tunisia. Int. J. Clim. Chang. Strateg. Manag. 11, 643–659. doi: 10.1108/IJCCSM-02-2018-0019
Creamer, M., and Horback, K. (2021). Researching human-cattle interaction on rangelands: challenges and potential solutions. Animals 11:725. doi: 10.3390/ani11030725
Curry, P. J., and Hacker, R. B. (1990). Can pastoral grazing management satisfy endorsed conservation objectives in arid Western Australia? J. Environ. Manage. 30, 295–320. doi: 10.1016/0301-4797(90)90025-R
Daget, P., and Poissonet, J. (1971). An ecological analysis method of prairies criteria’s of application. Ann. Agron. 22, 5–41. doi: 10.1371/journal.pone.0170137
Dorrough, J. W., Ash, J. E., Bruce, S., and Mclntyre, S. (2007). From plant neighbourhood to landscape scales: how grazing modifies native and exotic plant species richness in grassland. Plant Ecol. 191, 186–198. doi: 10.1007/s11258-006-9236-y
Favretto, N., Stringer, L., Dougill, A., Dallimer, M., Perkins, J., Reed, M., et al. (2016). Multi-criteria decision analysis to identify dryland ecosystem service trade-offs under different rangeland land uses. Ecosyst. Serv. 17, 142–151. doi: 10.1016/j.ecoser.2015.12.005
Fay, P. A., Kaufman, D. M., Nippert, J. B., Carlisle, J. D., and Harper, C. W. (2008). Changes in grassland ecosystem function due to extreme precipitation events: implications for responses to climate change. Glob. Change Biol. 14, 1600–1608. doi: 10.1002/ece3.1662
Gamoun, M. (2013). Management and resilience of Saharan rangelands: south Tunisia. Fourrages 216, 321–328.
Gamoun, M. (2014). Grazing intensity effects on the vegetation in desert rangelands of Southern Tunisia. J. Arid. Land 6, 324–333. doi: 10.1007/s40333-013-0202-y
Gamoun, M. (2016). Rain use efficiency, primary production and precipitation relationships in desert rangelands of Tunisia. Land Degrad. Dev. 27, 738–747. doi: 10.1002/ldr.2418
Gamoun, M., Essifi, B., Dickens, C., and Hanchi, B. (2016). Interactive effects of grazing and drought on desert rangelands of Tunisia. Biologija 62, 105–115. doi: 10.6001/biologija.v62i2.3336
Gamoun, M., Tarhouni, M., Ouled Belgacem, A., Neffati, M., and Hanchi, B. (2011). Response of different arid rangelands to protection and drought. Arid. Land Res. Manag. 25, 372–378. doi: 10.1080/15324982.2011.611578
George, M., Nader, G., McDougald, N., Connor, M., and Frost, B. (2001). Annual Rangeland Forage Quality. Publication 8022. Davis, CA: University of California Division of Agriculture and Natural Resources.
Goodloe, S. (1969). Short duration grazing in rhodesia. Rangel. Ecol. Manag. 22, 369–373. doi: 10.2307/3895844
Hanafi, A., and Jauffret, S. (2008). Are long-term vegetation dynamics useful in monitoring and assessing desertification processes in the arid steppe, southern Tunisia. J. Arid Environ. 72, 557–572. doi: 10.1016/j.jaridenv.2007.07.003
Harris, R. B. (2010). Rangeland degradation on the Qinghai-Tibetan plateau: a review of the evidence of its magnitude and causes. J. Arid Environ. 74, 1–12. doi: 10.1016/j.jaridenv.2009.06.014
Heisler-White, J. L., Blair, J. M., Kelly, E. F., Harmoney, K., and Knapp, A. K. (2009). Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Glob. Change Biol. 15, 2894–2904. doi: 10.1111/j.1365-2486.2009.01961.x
Henkin, Z., Landau, S. Y., Ungar, E. D., Perevolotsky, A., Yehuda, Y., and Sternberg, M. (2007). Effect of timing and intensity of grazing on the herbage quality of a Mediterranean rangeland. J. Anim. Feed Sci. 16, 318–322. doi: 10.22358/jafs/74517/2007
Heukes, P. C., and Cowling, R. M. (2000). Impacts of non-selective grazing on cover, composition, and productivity of Nama-karoo grassy shrubland. Afr. J. Range Forage Sci. 17, 27–35. doi: 10.2989/10220110009485736
Hodgson, J. G., Montserrat-Martì, G., Tallowin, J., Thompson, K., Díaz, S., Cabido, M., et al. (2005). How much will it cost to save grassland diversity? Biol. Conserv. 122, 263–273. doi: 10.1016/j.biocon.2004.07.016
Holechek, J. L., Pieper, R. D., and Herbel, C. H. (1998). Range Management Principles and Practices, 3rd Edn. Hoboken, NJ: Prentice Hall.
Hurlbert, S. H. (1984). Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 54, 187–211. doi: 10.2307/1942661
Jama, B., and Zeila, A. (2005). Agroforestry in the drylands of Eastern Africa: A Call to Action. ICRAF Working Paper – No. 1. Nairobi: World Agroforestry Centre.
Jauffret, S., and Visser, M. (2003). Assigning life history traits to plant species to better qualify arid land degradation in presaharian tunisia. J. Arid Environ. 55, 1–28. doi: 10.1016/S0140-1963(02)00258-6
Joseph, J., Molinar, F., Galt, D., Valdez, R., and Holechek, J. (2002). Short duration grazing research in Africa an extensive review of the charter grazing trials and other short duration grazing studies on African rangelands. Rangelands 24, 9–12. doi: 10.2458/azu_rangelands_v24i4_joseph
Kemp, D. R., and Michalk, D. L. (1993). Pasture Management: Technology for the 21st Century. Melbourne: The Commonwealth Scientific and Industrial Research Organisation.
Köchy, M., Mathaj, M., Jeltsch, F., and Malkinson, D. (2008). Resilience of stocking capacity to changing climate in arid to Mediterranean landscapes. Reg. Environ. Change 8, 73–87. doi: 10.1007/s10113-008-0048-6
Kohyani, B. T., Bossuyt, B., Bonte, D., and Hoffmann, M. (2008). Grazing as a management tool in dune grasslands: evidence of soil and scale dependence of the effect of large herbivores on plant diversity. Biol. Conserv. 141, 1687–1694. doi: 10.1016/j.biocon.2008.04.010
Le Floc’h, E. (2001). Biodiversité et gestion pastorale en zone aride et semi-aride méditerranéenne du nord de l’Afrique. Bocconea 13, 223–237.
Louhaichi, M., Gamoun, M., Ben Salem, F., and Ouled Belgacem, A. (2021a). Rangeland biodiversity and climate variability: supporting the need for flexible grazing management. Sustainability 13:7124. doi: 10.3390/su13137124
Louhaichi, M., Gamoun, M., Sawsan, H., and Abdallah, M. A. B. (2021b). Characterizing biomass yield and nutritional value of selected indigenous range species from arid Tunisia. Plants 10:2031. doi: 10.3390/plants10102031
Louhaichi, M., Ouled Belgacem, A., Petersen, S. L., and Hassan, S. (2019). Effects of climate change and grazing practices on shrub communities of West Asian rangelands. J. Clim. Chang. Strateg. Manag. 11, 660–671. doi: 10.1108/IJCCSM-02-2018-0017
Louhaichi, M., Salkini, A. K., and Petersen, S. L. (2009). Effect of small ruminant grazing on the plant community characteristics of semi-arid Mediterranean ecosystems. Int. J. Agric. Biol. 11, 681–689.
Martin, S. C. (1975). Ecology and Management of Southwestern Semi Desert Grass-Shrub Ranges: the Status of our Knowledge. RM-156, Forest Service, Rocky Mountain Forest and Range Experiment Station Fort Collins. Fort Collins, CO: Department of Agriculture.
McCabe, J. T. (2004). Cattle Bring us to our Enemies: Turakana Ecology, Politics, and Raiding in a Disequilibrium System. Ann Arbor, MI: University of Michigan Press.
McNeely, J. A. (2004). Strangers in our midst: the problem of invasive alien species. Environment 46, 16–31. doi: 10.1080/00139157.2004.10545159
Ouled Belgacem, A., and Louhaichi, M. (2013). The vulnerability of native rangeland plant species to global climate change in the West Asia and North African regions. Clim. Change. 119, 451–463. doi: 10.1007/s10584-013-0701-z
Ouled Belgacem, A., Tarhouni, M., and Louhaichi, M. (2013). Effect of protection on plant community dynamics in the Mediterranean arid zone of Southern Tunisia: a case study from Bouhedma national park. Land Degrad Dev. 24, 57–62. doi: 10.1002/ldr.1103
Savory, A. (1978). “A holistic approach to ranch management using short duration grazing,” in Proceedings of the First International Rangeland Congress, ed. D. N. Hyder (Denver, CO: Society for Range Management), 555–557.
Schönbach, P., Wan, H., Schiborra, A., Gierus, M., Bai, Y., Müller, K., et al. (2009). Short-term management and stocking rate effects of grazing sheep on herbage quality and productivity of Inner Mongolia steppe. Crop Pasture Sci. 60, 963–974. doi: 10.1071/CP09048
Scoones, I. (1995). “Range management science and policy: politics, polemics, and pasture in southern Africa,” in The Lie of the Land: Challenging Received Wisdom on the African Environment, eds M. Leach and R. Mearns (Oxford: James Currey Ltd), 34–53.
Smet, M., and Ward, D. A. (2005). Comparison of the effects of different rangeland management systems on plant species composition, diversity and vegetation structure in a semi-arid savanna. Afr. J. Range Forage Sci. 22, 59–71. doi: 10.2989/10220110509485862
Squires, V. R. (2010). “People in rangelands: their role and influence on rangeland utilization and sustainable management,” in Range and Animal Sciences and Resources Management, ed. V. R. Squires (Paris: Encyclopedia of Life Support Systems), 408. doi: 10.1073/pnas.2002314117
Squires, V. R., Dengler, J., Hua, L., and Feng, H. (2018). Grasslands of the World: Diversity, Management and Conservation. Boca Raton, FL: CRC Press.
Sullivan, S., and Rohde, R. (2002). On non-equilibrium in arid and semi-arid grazing systems. J. Biogeogr. 29, 1595–1618. doi: 10.1046/j.1365-2699.2002.00799.x
Suttie, J. M., Reynolds, S. G., and Batello, C. (2005). Grasslands of the World. Food and Agriculture Organization of the United Nations. Available online at: http://www.fao.org/3/y8344e/y8344e.pdf (accessed August 4, 2021).
Teague, W. R., Dowhower, S. L., Baker, S. A., Haile, N., DeLaune, P. B., and Conover, D. M. (2011). Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 141, 310–322. doi: 10.1016/j.agee.2011.03.009
van Duivenbooden, N. (1993). Grazing as a tool for rangeland management in semiarid regions: a case study in the north-western coastal zone of Egypt. Agric. Ecosyst. Environ. 43, 309–324. doi: 10.1016/0167-8809(93)90094-6
Vetter, S. (2009). Drought, change and resilience in South Africa’s arid and semi-arid rangelands. S. Afr. J. Sci. 105, 29–33.
Waite, S. (2000). Statistical Ecology in Practice: A Guide to Analysing Environmental and Ecological Field Data. Harlow: Prentice Hall.
Wang, L., Gan, Y., Wiesmeier, M., Zhao, G., Zhang, R., Han, G., et al. (2018). Grazing exclusion- an effective approach for naturally restoring degraded grasslands in Northern China. Land Degrad. Dev. 29, 4439–4456. doi: 10.1002/ldr.3191
Yiakoulaki, M. D., and Papanastasis, V. P. (2005). “Diet selection of sheep and goats grazing on cereal stubble in northern Greece,” in Sustainable grazing, nutritional utilization and quality of sheep and goat products, eds E. Molina Alcaide, H. Ben Salem, K. Biala, and P. Morand-Fehr (Zaragoza: CIHEAM-IAMZ), 245–250.
Yu, L., Zhou, L., Liu, W., and Zhou, H. K. (2010). Using remote sensing and GIS technologies to estimate grass yield and livestock carrying capacity of alpine grasslands in Golog Prefecture, China. Pedosphere 20, 342–351. doi: 10.1016/S1002-0160(10)60023-9
Keywords: grazing exclusion, spring grazing, stocking rate, rapid grazing, gdel
Citation: Louhaichi M, Gamoun M and Gouhis F (2021) Benefits of Short-Duration, High-Stocking Rate Opportunistic Grazing on Arid Rangelands During Favorable Conditions. Front. Ecol. Evol. 9:757764. doi: 10.3389/fevo.2021.757764
Received: 12 August 2021; Accepted: 21 October 2021;
Published: 02 December 2021.
Edited by:
Stephen M. Mureithi, University of Nairobi, KenyaReviewed by:
Jennifer Lesley Silcock, The University of Queensland, AustraliaIlan Stavi, Dead Sea and Arava Science Center, Israel
Copyright © 2021 Louhaichi, Gamoun and Gouhis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mounir Louhaichi, M.louhaichi@cgiar.org