 Mark J. MacDougall1*
Mark J. MacDougall1* Robert Seeger1
Robert Seeger1 Bryan Gee2,3
Bryan Gee2,3 Jasper Ponstein1,4Maren Jansen1Diane Scott2
Jasper Ponstein1,4Maren Jansen1Diane Scott2 Joseph J. Bevitt5
Joseph J. Bevitt5 Robert R. Reisz2
Robert R. Reisz2 Jörg Fröbisch1,4
Jörg Fröbisch1,4- 1Museum für Naturkunde Leibniz-Institute für Evolutions- und Biodiversitätsforschung, Berlin, Germany
- 2Department of Biology, University of Toronto Mississauga, Mississauga, CA, Canada
- 3Department of Biology, University of Washington, Seattle, WA, United States
- 4Department of Biology, Humboldt-Universität zu Berlin, Berlin, Germany
- 5Australian Centre for Neutron Scattering, Australian Nuclear Science and Technology Organisation, Lucas Heights, Sydney, NSW, Australia
The early Permian Richards Spur locality of Oklahoma has produced abundant material of numerous terrestrial fossil tetrapods, including various “microsaurs,” several of which are considered to belong to the clade Recumbirostra. We present a new partial skull of the recumbirostran “microsaur” Nannaroter mckinziei; through computed tomography (CT) analysis of both this new specimen and the holotype, we provide an updated description of the taxon. This new description provides novel information regarding several regions that could not be examined previously due to either being absent in the holotype or difficult to access. This includes missing and obscured aspects of the skull roof, braincase, lower jaw, and the palatal region. Furthermore, the new information obtained from this description was used to update phylogenetic character codings of Nannaroter, and a revised phylogenetic analysis was conducted. The results of this updated analysis are congruent with those of other recent phylogenetic analyses of recumbirostran “microsaurs.” This new information adds to the ever-growing body of early tetrapod CT data, which has been, and will continue to be, important in revealing details regarding early tetrapod anatomy, interrelationships, paleoecology, and evolution.
Introduction
Historically, “Microsauria” (Dawson, 1863) was considered to represent a diverse clade of small Palaeozoic terrestrial lepospondyl anamniotes found in North America and Europe, however, over the past few decades it has been proposed that the clade is not actually monophyletic but instead represents a polyphyletic assemblage (Anderson, 2001, 2007). Among “microsaurs” is the clade Recumbirostra, a group largely characterized by a recumbent rostrum (Anderson, 2007). Recently, it has been suggested that recumbirostrans possibly represent members of Reptilia (Pardo et al., 2017), instead of being closely related to the other lepospondyl “microsaurs,” providing further support for the hypothesis that “Microsauria” is not a monophyletic clade.
Within Recumbirostra, Ostodolepidae is a clade of “microsaurs” known primarily from south-central North America and currently comprises four genera. Ostodolepis, which forms the basis for the family-level name, was described from the Arroyo Formation of Texas by Williston (1913) on the basis of an articulated series of seven vertebrae, associated ribs, and scales (referred to as “scutes”) that he ascribed to a new genus of reptile. Because of the relative similarity of “microsaur” vertebrae, such material is not diagnostic, even at the genus level (Carroll and Gaskill, 1978); in a later work, Williston (1916) suggested that the taxon was synonymous with Pantylus, although this has not been corroborated or adopted by subsequent studies. The holotype of Ostodolepis features centra that are entirely fused to the neural arches, as in gymnarthrids and in contrast to other ostodolepids (Carroll and Gaskill, 1978). Case (1929) described a nearly complete specimen that he assigned to Ostodolepis and that he asserted was likely from the same locality as the holotype, though the material was obtained second-hand and little evidence was provided to support this claim beyond brief notes of a similar matrix lithology. Carroll and Gaskill (1978) separated this specimen from Ostodolepis and instead placed it within a new genus, Pelodosotis, on the basis of the presence of a clear neurocentral suture, more prominent intercentra, and different neural spine morphology. These authors noted that these notable differences could be reflective of the larger size (and presumed maturity) of Ostodolepis. Daly (1973) described a third taxon, Micraroter, from the South Grandfield locality in Oklahoma on the basis of a partial skull with an articulated jaw and an assortment of highly fragmentary cranial, mandibular, vertebral, and pectoral elements. Another specimen (BPI 3839) from around the same area as the holotype of Ostodolepis was also assigned to Micraroter by Carroll and Gaskill (1978) but demonstrates a few significant differences from the latter (e.g., fewer presacral vertebrae, stapedial foramen) as well as a number of more minor ones (e.g., prefrontal excluded from the narial opening) that cannot be excluded as the result of ontogeny. Schultze and Foreman (1981) maintained the taxonomic distinction of BPI 3839 from Micraroter, although they did not formalize the former as a new taxon; their position has been maintained in recent publications concerning ostodolepids (e.g., Anderson et al., 2009; Henrici et al., 2011).
Two ostodolepid recumbirostrans were described in more recent years on the basis of well-preserved isolated skulls: Nannaroter mckinziei from the fossiliferous karst deposits near Richards Spur, Oklahoma (Anderson et al., 2009), and Tambaroter carrolli, the first ostodolepid outside of North America, from the Bromacker locality of Thuringia, Germany (Henrici et al., 2011). In the context of the Richards Spur locality, Nannaroter is a small part of a diverse “microsaurian” assemblage that also includes the gymnarthrids Cardiocephalus and Euryodus, and the hapsidopareiid Llistrofus (Carroll and Gaskill, 1978; Bolt and Rieppel, 2009; Gee et al., 2020). Relative to all taxa at the site, material of Nannaroter is extremely rare, which has been proposed as possible evidence for fossoriality in conjunction with various morphological features (e.g., recumbent snout, reinforced basicranium).
Although the holotype of Nannaroter is nearly complete and well exposed in all profiles, it is missing several of the posteroventral cranial elements around the temporal emargination and the posterior mandibular region. Here, using CT data of both the holotype and a new specimen of Nannaroter mckinziei, we provide an updated description and phylogenetic analysis of the taxon. Newly described areas of the skull include previously inaccessible or absent parts of the braincase, posterior skull roof, mandible, and palate. These regions are both informative for comparisons with other ostodolepids and other “microsaurs” with temporal emarginations (e.g., hapsidopareiids) and for improving our understanding of the ecology of the taxon based on features such as a prominent coronoid process.
Institutional abbreviations: BPI—Evolutionary Studies Institute, formerly Bernard Price Institute for Palaeontological Research, University of the Witwatersrand, Johannnesburg, South Africa; OMNH—Sam Noble Oklahoma Museum of Natural History, Norman, Oklahoma, United States; ROMVP—Royal Ontario Museum, Toronto, Ontario, Canada.
Materials and Methods
Fossil Material
Both the holotype and a new referred specimen were examined as part of this study, these specimens were obtained from the karst fissures that make up the cave system found at the early Permian (∼289–286 Ma) Richards Spur locality in Oklahoma, United States. Details of the geology and taphonomy of the locality were described by MacDougall et al. (2017).
The new referred specimen consists of a partial skull with an articulated right mandible. The right side of the skull is mostly intact posterior to the narial opening and includes a complete mandible. The left side is broken off anterior to the frontals (except for a tiny portion of the nasal), and all of the lateral and ventral elements are lost. Medially, the skull is incomplete beyond the anterior portion of the parietals, but the right lateral side is mostly intact, though slightly dislodged. Portions of the basicranium are also preserved in a slightly dorsally shifted position, but the palatal view is partially obscured by broken fragments that likely represent a combination of skull roof and palatal elements of the specimen. Most of the elements posterior to the orbit, including those of the mandible, are a low-contrast beige coloration, while those anterior to the orbit are of the more typical dark black coloration that results from hydrocarbon enrichment at the Richards Spur locality.
Computed Tomography Data
The holotype of Nannaroter mckinziei, OMNH 73107, was scanned at the Museum für Naturkunde Berlin, using x-ray computed tomography (phoenix| x-ray nanotom s) at 75 kV, 170 μA for 1,440 projections and an exposition timing of 1000 ms/projection, with a magnification ratio of 5.77 x, and an effective voxel size of 0.00865 mm. Cone beam reconstruction was performed using datos| x- reconstruction software (GE Sensing & Inspection Technologies GmbH phoenix| x-ray). The multiscan of two parts was visualized, merged and segmented in VGStudio Max 3.0.
High-resolution tomographic analysis of specimen ROMVP 86541 was achieved using the Imaging and Medical Beamline at the Australian Nuclear Science and Technology Organisation’s (ANSTO) Australian Synchrotron, Melbourne, Victoria, Australia. For this measurement, a monochromatic beam energy of 70kV utilized, along with the “Ruby” detector, consisting of a PCO.edge sCMOS camera (16-bit, 2,560 × 2,160 pixels) coupled with a Nikon Makro Planar 100 mm lens, along with a 20 μm thick Gd2O3/CsI (Tl)/CdWO4 scintillator screen to yield a cubic voxel of 6.0 μm. A total of 1,800 equally spaced radiographs of 0.1 s exposure each were acquired as the sample was rotated 180° about its vertical axis. 100 dark (closed shutter) and beam profile (open shutter) images were obtained for calibration before and after shadow-radiograph acquisition. Normalization, spot removal and 3D reconstruction of the raw data was achieved using a combination of ImageJ v.1.51h and Octopus Reconstruction v8.8 (Inside Matters NV), to yield virtual slices perpendicular to the rotation axis. Unfortunately, as much of this specimen consisted of bone of white coloration it resolved poorly in the neutron scans, making it very difficult to identify sutures and internal features in the scans of this specimen, this has also been observed for other specimens from the locality (Gee et al., 2020).
Phylogenetic Analysis
The matrix of Gee et al. (2020), the latest published derivation of the original matrix of Pardo et al. (2017), was used for the phylogenetic analysis; the only modifications made to it were that some character codings of Nannaroter mckinziei were updated. The analysis was performed in PAUP 4.0a167 (Swofford, 2003) with maximum parsimony set as the optimality criterion, all branch lengths of less than zero were set to collapse, and a heuristic search with 10,000 random additional replicates and tree bisection and reconnection (TBR) branch swapping was used to search for trees. Updated characters for Nannaroter mckinziei are as follows: quadratojugal present (15) ?→0, widely separated internal nares (76) ?→0, and parasphenoid medial to stapes (82) ?→0.
Systematic Paleontology
LEPOSPONDYLI Zittel, 1888
TUDITANOMORPHA Carroll and Gaskill, 1978
RECUMBIROSTRA Anderson, 2007
OSTODOLEPIDAE Romer, 1945
NANNAROTER Anderson et al., 2009
NANNAROTER MCKINZIEI Anderson et al., 2009
Diagnosis (Anderson et al., 2009)—Small ostodolepid lepospondyl with high subtemporal recess so that squamosal and postorbital do not articulate, four premaxillary and 12 maxillary teeth, medial laminae of prefrontal, lacrimal, jugal, and postorbital forming solid orbital walls (and floor with palatine, pterygoid, ectopterygoid, and vomer), a ventral flange of frontal integrated with large orbitosphenoid, and massive epipterygoid laterally supporting posterior braincase.
Holotype—OMNH 73107, nearly complete skull with articulated mandibles.
Referred specimen—ROMVP 86541, partial skull with articulated right mandible.
Description
New information obtained from CT data of holotype
The following description will not be comprehensive, but rather will focus on information from areas of the holotype that could not be easily examined (Figure 1) in the original description of Nannaroter mckinziei (Anderson et al., 2009).
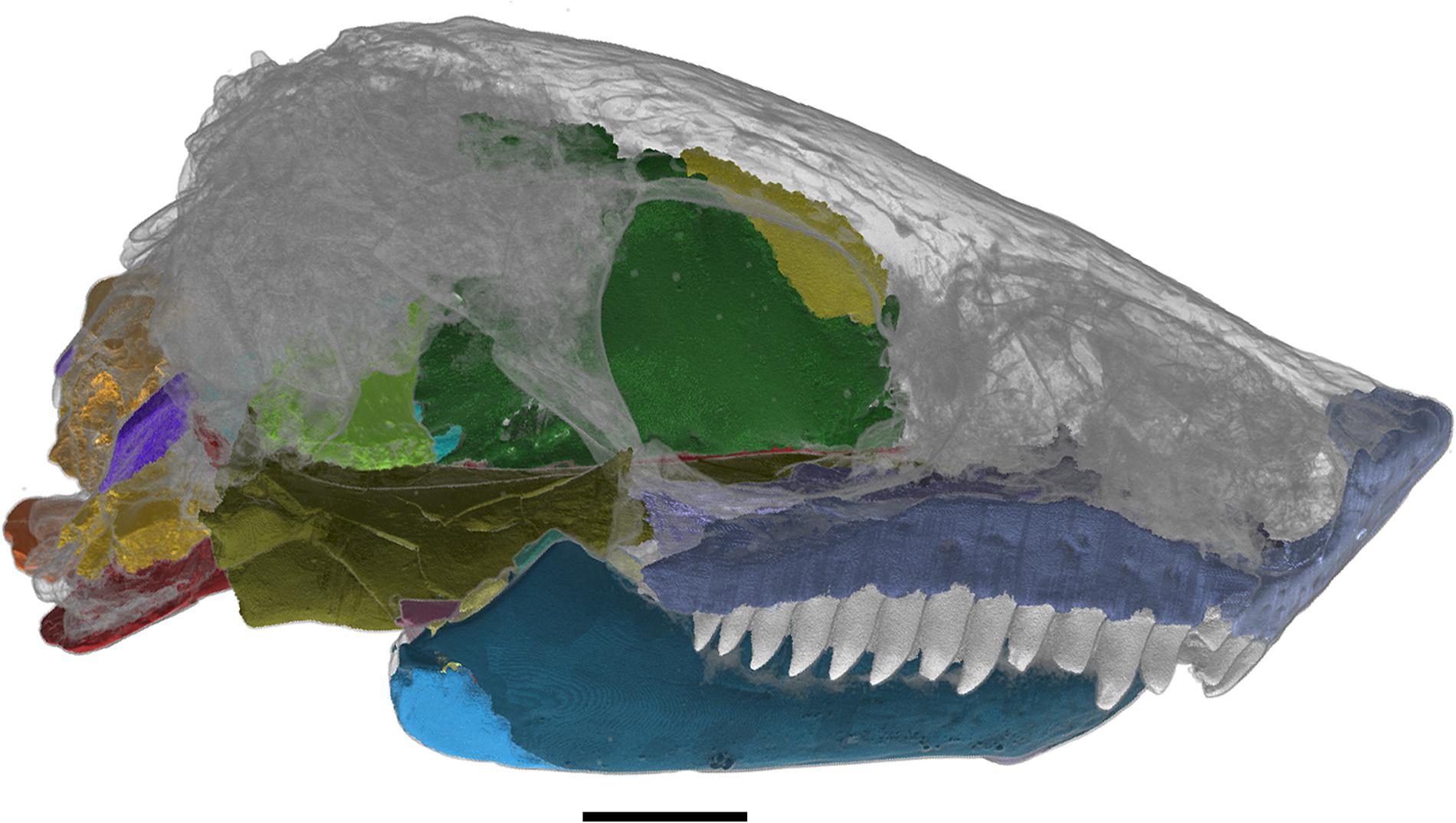
Figure 1. Skull of Nannaroter mckinziei, OMNH 73107, with the elements that were reconstructed using CT data shown. Scale bar equals 2 mm.
Palate
The vomer was only briefly described by Anderson et al. (2009), as much of it was obscured by the occluded jaws. The vomer is the anteriormost element of the palate (Figure 2); it is a flat rhomboidal bone with a long anteromedially placed dorsal flange. Dorsal view of the vomer reveals that this slender dorsal flange contacts with the ventral surface of the premaxilla, with the premaxilla interdigitating with the anterior section of the vomer with the former being wedged into it. The posteromedial edge of the vomer contacts the anterior portion of the palatine along a diagonal suture, however, the vomer and palatine do not meet at the posterior margin of the choana as erroneously described by Anderson et al. (2009), but rather further anteriorly. The posterolateral edge of the vomer contacts the anteromedial portion of the palatine, which combined with the aforementioned palatine contact, results in the triangular posterior portion of the vomer being wedged between the anterior ends of the palatine and pterygoid. As mentioned by Anderson et al. (2009) the vomer does appear to bear a single tooth row, though there are few teeth preserved in it. The vomer bears a large tooth on the posterior-most end of the bone and a smaller tooth that is more anteriorly placed on the medial edge of the element, near the anterior end of the pterygoid. In addition, the left vomer also exhibits a single large tooth anterior to the smaller one, whether this is the result of tooth replacement or is a consistent feature of the vomerine dentition cannot be currently determined due to the incomplete tooth rows on both vomers.
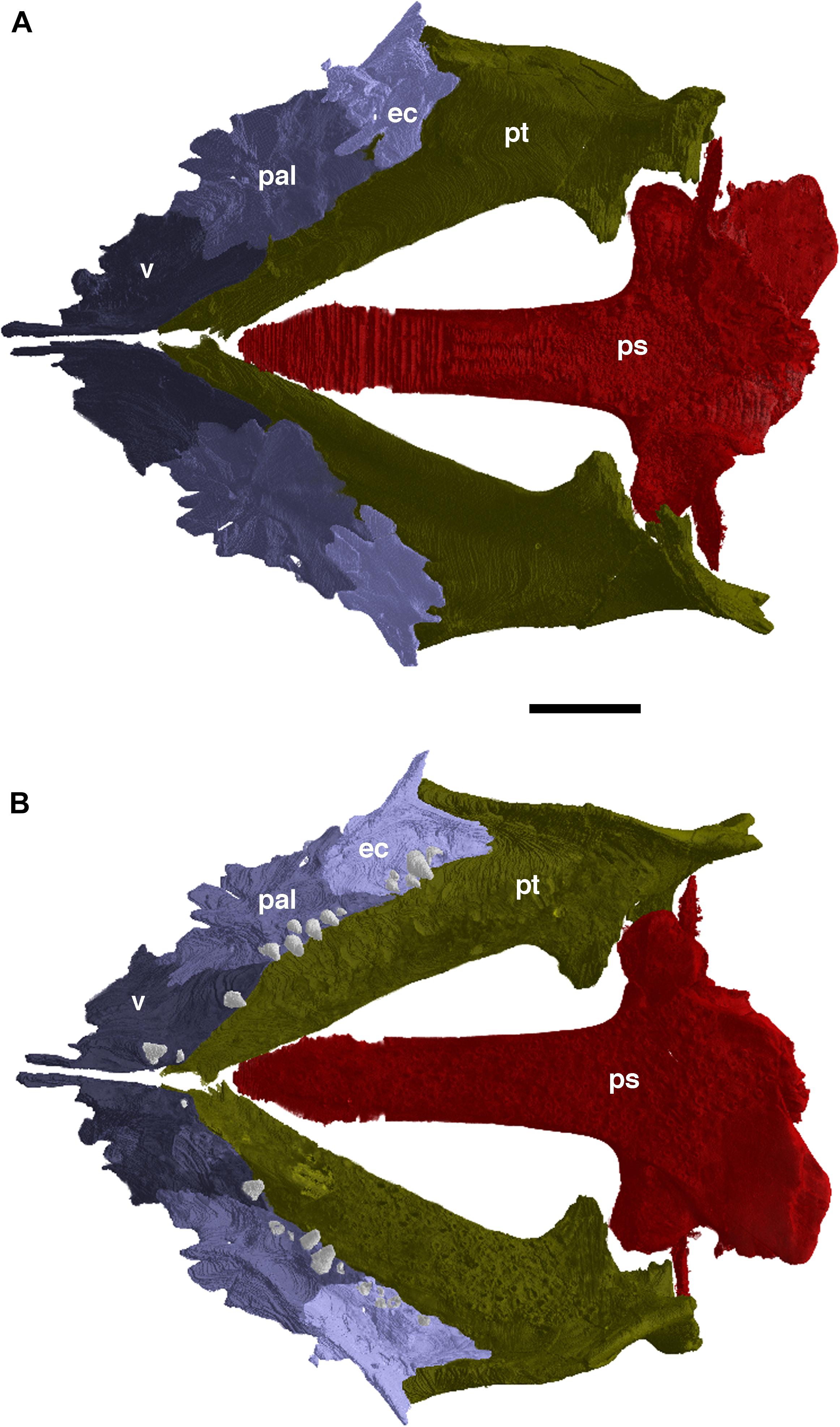
Figure 2. The palate of Nannaroter mckinziei, OMNH 73107, reconstructed from CT data. (A) Dorsal view, and (B) ventral view. ec, ectopterygoid; pal, palatine; ps, parasphenoid; pt, pterygoid; v, vomer. Scale bar equals 2 mm.
The palatine is a short, roughly rectangular element, smooth in ventral view (Figure 2). Its pointed anterior end interdigitates with the vomer and contributes to the posterior margin of the choana. Posteriorly, the palatine contacts the ectopterygoid along a curved suture in ventral view; in dorsal view the contact of the two elements is strongly interdigitated. The medial edge of the palatine contacts the vomer anteriorly and the pterygoid posteriorly, whereas the lateral edge of the element entirely contacts the maxilla. In ventral view, the articulation of both with the maxilla is along a relatively straight line, whereas the contact appears jagged in dorsal view, as well as curving slightly dorsally forming a larger surface of attachment with the maxilla. The palatine also possesses a medial tooth row of at least six teeth.
The ectopterygoid is similar in shape and size to the palatine, which the ectopterygoid contacts with anteriorly (Figure 2). As with the palatine, the lateral edge of the ectopterygoid contacts the maxilla, whereas the medial edge meets with the pterygoid. Posteriorly, the ectopterygoid splits into two processes, a medial one that entirely contacts the pterygoid, and a lateral one found wedged between the maxilla and pterygoid. The ectopterygoid possesses five teeth, which appears to be a continuation of the medially restricted teeth found on the palatine. This loose row of teeth is oriented parallel to the marginal dentition, and there is little variation between the sizes of the teeth, but they are noticeably smaller than the marginal dentition.
The pterygoid is much larger than the previously described palatal elements (Figure 2); it exhibits a steeply descending ventral flange on the lateral margin and reaches from the quadrate ramus anteriorly nearly touching the posteromedial margin of the maxilla with a small mediolateral tip of the ectopterygoid in between (Figure 1A). Both pterygoids reach far anterior and constrict toward the cultriform process of the parasphenoid, however they do not contact each other at their anterior ends due to slight taphonomic deformation. The pterygoid denticles and bone appear to have the same density in the CT data, which makes it difficult to identify and segment the small denticles mentioned by Anderson et al. (2009).
Mandible
As briefly mentioned by Anderson et al. (2009), the occluded mandible of the holotype of Nannaroter consists of a dentary, splenial, postsplenial, angular, surangular, and prearticular, with the posteriormost portion of the mandible missing. In addition to these elements, micro-CT revealed the presence of two distinct, tooth-bearing coronoid bones (Figure 3).

Figure 3. The partial right mandible of Nannaroter mckinziei, OMNH 73107, reconstructed from CT data. (A), Lateral view, (B) medial view, (C), dorsal view, and (D) ventral view. a, angular; c1, anterior coronoid; c2, posterior coronoid; d, dentary; pra, prearticular; psp, postsplenial; sa, surangular; sp., splenial. Scale bar equals 2 mm.
The dentary is the largest element of the mandibular ramus; in lateral view, it extends from the mandibular symphysis to just past the coronoid eminence. The element curves medially onto the ventral surface of the mandibular ramus where it contacts the splenial and postsplenial. In medial view, a small anterior portion of the dentary is visible, which ventrally contacts the splenial; posteriorly, it is obscured by the overlying coronoid elements. The marginal dentition of the dentary consists of 14 tooth positions, all of which are occupied in both dentaries, spanning from the mandibular symphysis to the point where the coronoid eminence begins. The edentulous posterior end of the element contributes to the coronoid eminence.
The coronoid of Nannaroter is divided into two distinct ossifications (Figure 3), with the second found immediately posterior to the first. Coronoid number appears to be quite variable among “microsaurs.” The presence of two coronoids in Nannaroter is similar to Rhynchonkos stovalli (Szostakiwskyj et al., 2015), but differs from taxa such as Euryodus dalyae, which possesses three coronoid elements on each mandibular ramus (Gee et al., 2020), and Aletrimyti gaskillae, which exhibits only a single coronoid (Szostakiwskyj et al., 2015). The anterior coronoid is a slender element and positioned dorsal to the posterior portion of the splenial, starting where the splenial broadens and extending posterior to its contact with the second coronoid. An anteroposterior row of at least 11 tooth positions can be observed on the anterior coronoid.
The second coronoid is positioned immediately posterior to the first coronoid, with the first coronoid slightly overlying its anteriormost end (Figure 3). It is roughly triangular in shape, being narrow anteriorly and broadening toward its posterior end. With its posterior end the second coronoid forms the anteromedial portion of the coronoid eminence. It has an indentation at the posterior end, normally obscured by the prearticular, in which the anterior tip of the fragmentary surangular is interlocked. The second coronoid bears only a single tooth, placed anteriorly on the element, it is in line with, but some distance from the tooth row of the anterior coronoid. The teeth of both coronoids are all similar in size and dorsomedially inclined.
The splenial is another large element of the mandibular ramus; its full extent is visible in medial view (Figure 3). Anteriorly, it contributes to the ventral portion of the mandibular symphysis, from there it extends posteriorly and slightly broadens until its contact with the prearticular and postsplenial. The splenial overlies the anterior half of the postsplenial and more than one third of the prearticular. It is also clear from the CT reconstruction of the mandibular ramus that in lateral view, a very small anteroventral portion of the splenial near the mandibular symphysis is exposed.
The postsplenial is similar in length to the splenial but is a much narrower element. In medial and ventral views, the full extent of the element cannot be observed normally due to being covered by the posterior portion of the splenial. Dorsally, it contacts the prearticular.
The prearticular is a broad element contributing to a substantial amount of the medial surface of the mandibular ramus, being widest underneath the tip of the coronoid process, covering medially most of the angular.
A small part of the angular can be seen in lateral view (Figure 3), however, the actual posterior extent of the element cannot be established exactly due to the posterior end of the mandibular ramus being absent. The anterior part of the angular is positioned ventral to the posterior end of the coronoid eminence, it tapers anteriorly to a narrow point. A large portion of the preserved angular is not visible externally, and is found wedged between the dentary and prearticular, this internal portion of the angular extends anteriorly to roughly the middle of the dentary.
In medial view, posterior to the prearticular and dorsal to the exposed part of the angular sits a small fragment of the surangular; it contributes to the posteromedial part of the coronoid eminence (Figure 3).
Marginal Dentition
In both examined specimens, the upper and lower marginal dentition are preserved as single and complete tooth row with no missing teeth. The premaxilla has four tooth positions, the maxilla has 12, and the dentary possesses 14. All teeth are homodont, monocuspid, and conical in shape; the posterior three teeth of the maxilla and dentary are slightly smaller than the other teeth, as is the case in many early reptiles. The cross-sections of the teeth are sub-rectangular at the base, become rounded moving toward the tip, and labiolingually compressed at the tip, which forms mesial and distal keels (carinae). There are no ridges on the keels and the tip is slightly recurved.
CT data reveals that the bases of the teeth exhibit gently folded plicidentine (Figure 4) similar to the recumbirostran “microsaur” Euryodus (Peyer, 1968) and the captorhinid reptile Captorhinus aguti (de Ricqlès and Bolt, 1983). The bases of the teeth of Nannaroter also exhibit numerous radial canals, which would have allowed nerve fibers and nutrient vessels to enter the pulp cavity. Overall, this type of plicidentine is very similar to what is observed in some early reptiles (de Ricqlès and Bolt, 1983; MacDougall et al., 2014) and appears to be quite common across the anamniote-amniote transition. The tooth implantation and attachment in Nannaroter is subthecodont, with the marginal teeth being held in small, shallow sockets along the jaws. This primitive type of attachment is common in various Paleozoic tetrapods. No direct evidence of tooth replacement could be found in the examined specimens, as there are no clear resorption pits or newly erupted teeth anywhere along the jaws. The complete lack of replacement teeth and empty tooth sockets may be indicative of a replacement pattern similar to that of some bolosaurid reptiles, in which tooth replacement is highly synchronized (Snyder et al., 2020). This is a slightly different pattern of tooth replacement than the simultaneous replacement that has been suggested for some “microsaurs” (Bolt and DeMar, 1983).
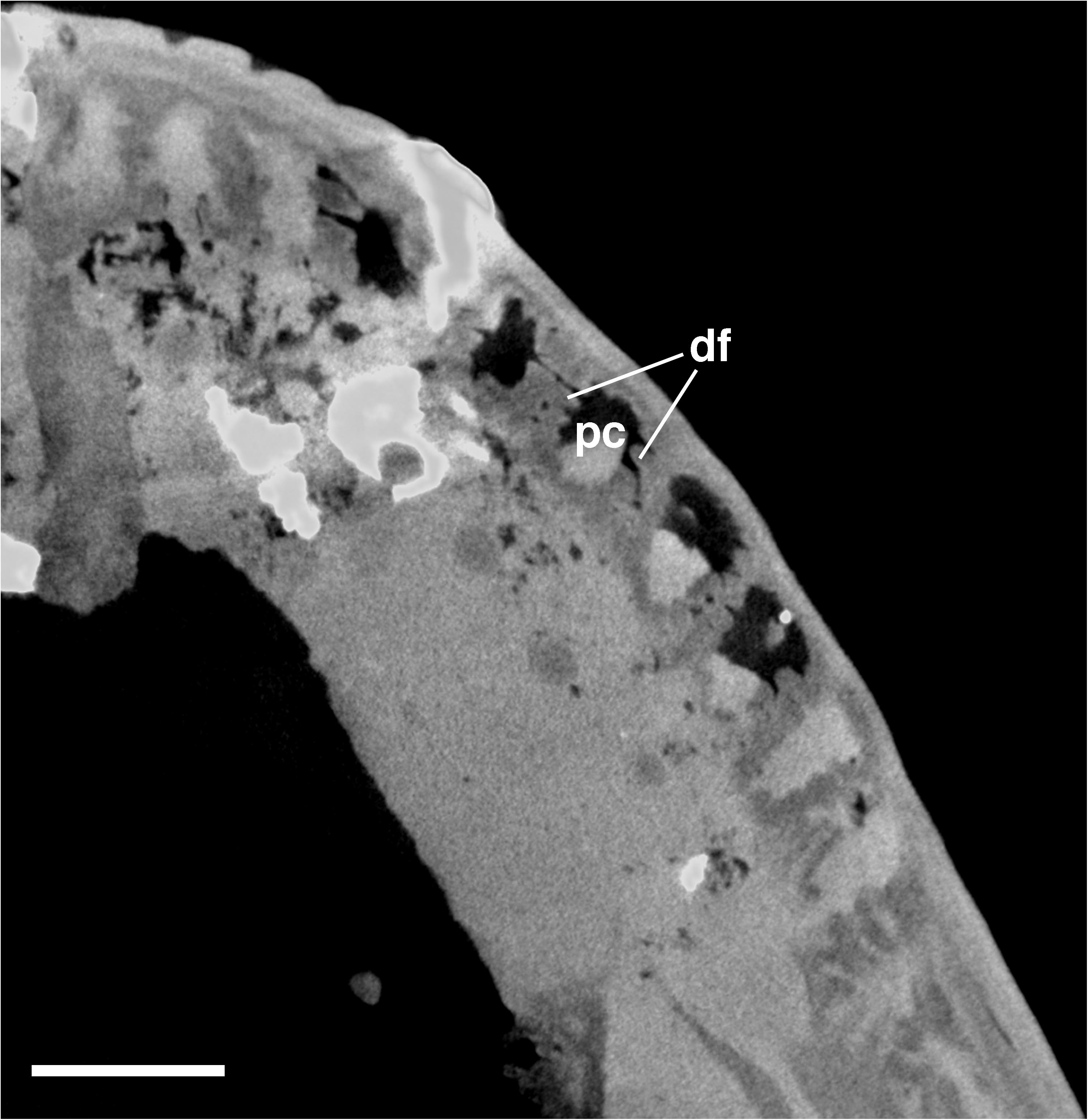
Figure 4. Cross-sectional view of the bases of the upper jaw marginal teeth in Nannaroter mckinziei, OMNH 73107, obtained from CT scans. df, dentine fold; pc, pulp cavity. Scale bar equals 1 mm.
Braincase
The external portions of the braincase that were preserved in the holotype of Nannaroter were described by Anderson et al. (2009), and some aspects of its internal anatomy was briefly described by Anderson et al. (2009); Huttenlocker et al. (2013), and Szostakiwskyj et al. (2015). Here, we add some further details regarding the braincase of Nannaroter. Much of the posterior braincase is crushed, damaged, and distorted. In addition, some elements appear to be absent. However, moving anterior from the occiput reveals much better preservation of the more interior braincase elements (Figure 5).

Figure 5. Braincase elements preserved in Nannaroter mckinziei, OMNH 73107, reconstructed from CT data. (A) Left lateral view, (B) dorsal view, and (C) posterior view. bo, basioccipital; eo, exoccipital; ep, epipterygoid; f, frontal; op, opisthotic; pls, pleurosphenoid; ps, parasphenoid; so, supraoccipital; s, stapes. Scale bar equals 1 mm.
The orbitosphenoid is a large U-shaped element that extends anteriorly from its contact with the pleurosphenoids to the anterior end of the underlying parasphenoid (Figure 5). The orbitosphenoid is one of the largest elements of the braincase and makes up a substantial portion of the lateral walls of the sphenoid region. Ventrally, the orbitosphenoid is underplated by the parasphenoid, with the two elements contacting each other along a wavy suture for much of the length of the orbitosphenoid. There also appears to be small gap in the ventral portion of the element, which may be due to the poor resolution of the scans in this region. It is also worth noting that the presence of a singular orbitosphenoid could potentially be the result of poor resolution and that it may actually be paired orbitosphenoids that are bridged by median ossifications, as is observed in other recumbirostran taxa (Szostakiwskyj et al., 2015), however, no sutures that would confirm this were identifiable. The posterolateral edges of the orbitosphenoid contact the pleurosphenoids along strongly interdigitated sutures. Furthermore, there is also a ventral flange on the frontals that contacts the anterodorsal portions of the orbitosphenoid. This descending flange of the frontal is small in comparison to the sphenoid elements, it is slightly curved and extends along the anterodorsal surface of the orbitosphenoid forming a tight articulation. As in other recumbirostrans (Szostakiwskyj et al., 2015), this contact between the frontal and orbitosphenoid serves to help support the sphenoid region.
The paired pleurosphenoids are thin elements about half the height of the orbitosphenoid, they also have a much shorter anteroposterior length. They make up the rest of the lateral walls of the sphenoid region and contact the dorsolateral edges of the parasphenoid ventrally. The paired nature of the pleurosphenoids is notable as in other recumbirostrans they instead form a single unpaired ossification (Szostakiwskyj et al., 2015).
Both of the epipterygoids are preserved in the holotype of Nannaroter; they are broad, robust, and well-ossified elements (Figure 5). Overall, they are very similar in structure to the epipterygoids of Carrolla (Maddin et al., 2011) and Euryodus (Gee et al., 2020). Ventrally, they possess an enormous facet for their contact with the basipterygoid processes, as well as a large dorsal process that extends toward the skull roof, but this extension appears to be too short to actually contact with it.
As noted by Anderson et al. (2009) the left stapes is present and is in its proper position (Figure 5), although it is damaged. It is a robust element with a broad concave footplate and no apparent dorsal process. There is no indication of a clear stapedial foramen, though there is a long gap between the footplate and the remaining lateral portion of the element, this is likely the result of the element being damaged.
A single median supraoccipital is present in the holotype, though it is not in its natural position, being both disarticulated and damaged. It is a broad flat element that would have acted as the roof of the braincase, it does not appear to have an anterior process that would have underlain the posteriormost elements of the skull roof, unlike what is observed in Huskerpeton (Huttenlocker et al., 2013) and Euryodus (Gee et al., 2020).
Both opisthotics are present, but the left opisthotic is considerably damaged (Figure 5). The more complete right opisthotic is quite short anteroposteriorly, but is very wide mediolaterally, being roughly three times wider than long. Ventrally, it contacts the posterior end of the basioccipital and would have contributed to part of the lateral wall of the foramen magnum. Dorsolaterally, they would have contacted with their associated exoccipital, however, as with several elements of the braincase in this specimen, the exoccipitals are disarticulated and not in their natural locations.
New specimen
This new material of Nannaroter mckinziei (ROMVP 86541) is a partial skull that largely consists of the right side of the skull roof and right mandible (Figures 6–8). Notably, it includes some of the more posteriorly positioned cranial and mandibular elements that were either damaged or not preserved in the holotype specimen. Comparative measurements between the holotype and new specimen can be found in Table 1.
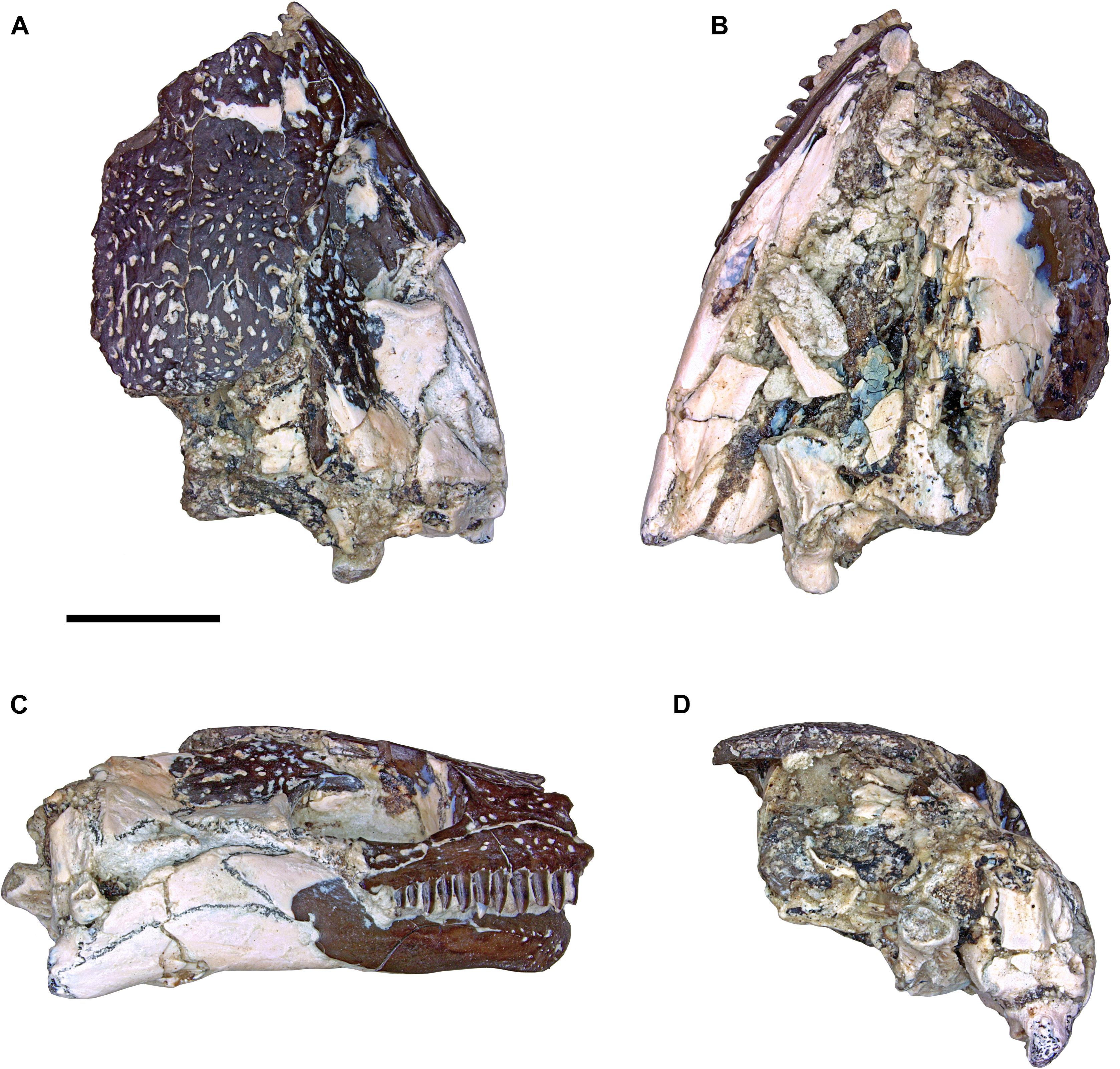
Figure 6. Photographs of the partial skull of Nannaroter mckinziei, ROMVP 86541. (A) Dorsal view, (B) ventral view, (C) right lateral view, and (D) occipital view. Scale bar equals 5 mm.

Table 1. Comparative skull measurements of areas that are preserved on both the holotype and new referred specimen of Nannaroter mckinziei.
Skull Roof
No premaxillae are preserved in the new specimen.
The nasals of the specimen are nearly entirely lost except at their posterior margin where they suture to the frontals. A small portion of the lateral suture with the prefrontal is preserved on the right side of the skull (Figures 6, 7).
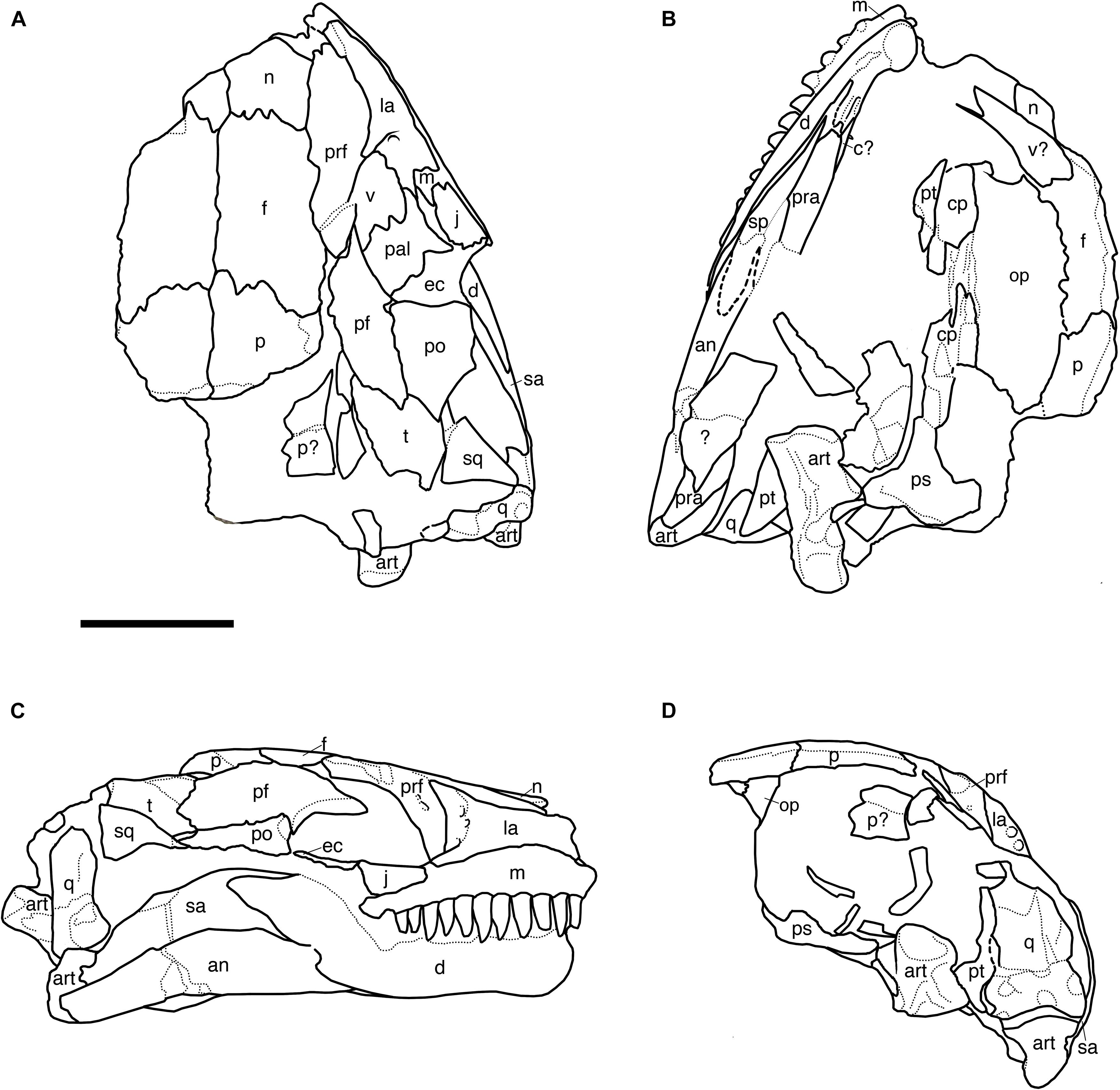
Figure 7. Line drawings of the partial skull of Nannaroter mckinziei, ROMVP 86541. (A) Dorsal view, (B) ventral view, (C) right lateral view, and (D) occipital view. an, angular; art, articular; c, coronoid; cp, cultriform process; d, dentary; ec, ectopterygoid; f, frontal; j, jugal; la, lacrimal; m, maxilla; n, nasal; op, opisthotic; p, parietal; pf, postfrontal; po, postorbital; pra, prearticular; prf, prefrontal; ps, parasphenoid; q, quadrate; qj, quadratojugal; sa, surangular; sq, squamosal; t, tabular. Scale bar equals 5 mm.
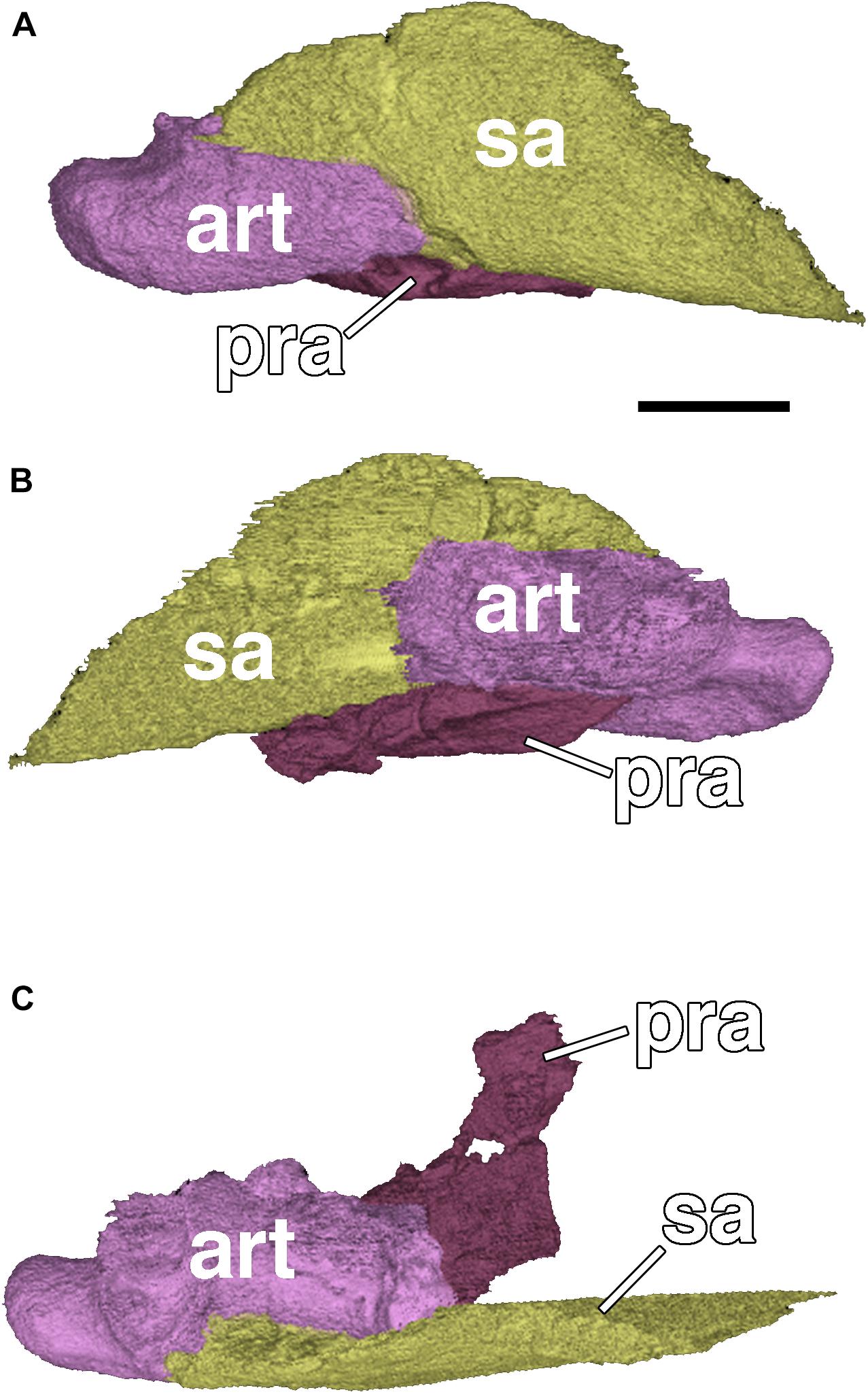
Figure 8. Posterior end of the right mandibular ramus of Nannaroter mckinziei, ROMVP 86541. (A) Lateral view, (B) medial view, and (C) dorsal view. art, articular; pra, prearticular; sa, surangular. Scale bar equals 2 mm.
The right prefrontal is incomplete, being preserved posteriorly where it sutures to the frontal posteromedially and at its mid-length where it sutures to the nasal medially and to the lacrimal laterally (Figures 6, 7). Based on the length of the preserved region, it appears that only the anteriormost portion that contributes to the posterior margin of the naris is absent. As with other ostodolepids, except for Pelodosotis (Carroll and Gaskill, 1978), the element is relatively elongate and rectangular, whereas in the latter, it is more trapezoidal as a result of the oblique suture with the frontal. The element also extends ventrally to contribute to the inner anterior orbital surface in a fashion that separates the lacrimal from the postfrontal.
The frontals are elongate, rectangular elements sutured to the nasals anteriorly, to the prefrontals anterolaterally, to the postfrontals posterolaterally, and to the parietals posteriorly (Figures 6, 7). As in most ostodolepids, except for Pelodosotis, the sutures with the nasals and the parietals are oriented nearly straight mediolaterally (Carroll and Gaskill, 1978); this is likely a result of more pronounced anterior tapering of the latter. As with most other “microsaurs” (the few exceptions include the co-occurring Llistrofus), the frontals are excluded from the orbital margin by a direct contact of the pre- and postfrontals.
The maxilla is an elongate dentulous element sutured to the jugal posterodorsally, to the lacrimal dorsally, and presumably to the premaxilla anteriorly, although this region of the snout is lost. The maxilla appears essentially complete (Figures 6, 7); it is shortest posteriorly, where it underlies the jugal, and gradually expands dorsally to about the mid-length of the lacrimal before shortening slightly. The teeth are conical, and slightly compressed and recurved at the tip of the crown. They appear to increase very slightly in size to the sixth position and then decrease posteriorly. Eleven teeth are present, and the posteriormost tooth socket is vacant, this makes for a total of 12 tooth positions; this count is the same as in the holotype (Anderson et al., 2009). This is the lowest tooth count among ostodolepids; Pelodosotis has 18, and Micraroter, BPI 3839, and Tambaroter are estimated to have 16 (Daly, 1973; Carroll and Gaskill, 1978; Henrici et al., 2011). Relative to other “microsaurs,” the tooth count is on the lower end, being comparable to many gymnarthrids that usually have 10–12 teeth.
The lacrimal is a robust, triangular element that contributes to the anterior orbital margin and is sutured to the prefrontal dorsomedially, to the maxilla ventrally, and to the jugal posteriorly (Figures 6, 7). The element is broadest posteriorly at the orbital margin and then narrows to a rectangular shape that maintains a consistent height. The morphology is similar to that of other ostodolepids except for BPI 3839 in which the element is rectangular throughout. The lacrimal of Pelodosotis continues to taper anteriorly to the narial opening. It is difficult to compare to that of Tambaroter because of the dorsoventral compression of the latter; by all accounts, the former appears to be generically similar. Also of note is that in Micraroter, BPI 3839, and Tambaroter, the lacrimal and the jugal are separately by a short region of the maxilla that contributes to the ventral orbital margin (Daly, 1973; Carroll and Gaskill, 1978; Henrici et al., 2011). This is considered a derived feature, but with regard to Micraroter, the restoration is somewhat suspect based on the figures of Daly (1973) and the poor preservational condition of the specimens. As with the holotype, several prominent foramina are present on the inner anterior orbital surface on the lacrimal in the referred specimen. The largest two are paired dorsoventrally near the mid-height of the anterior orbital margin and were interpreted as the openings of the nasolacrimal duct, and a third, slightly smaller and more ventromedially positioned foramen is interpreted as the passage site of the palatal branch of the facial nerve (Anderson et al., 2009). Similar foramina are reported in only a few “microsaurian” taxa, including the hapsidopareiids, Asaphestera, Pantylus, Cardiocephalus, and Micraroter (Daly, 1973; Carroll and Gaskill, 1978; Bolt and Rieppel, 2009; Gee et al., 2019). It is figured but not described for Pelodosotis (Carroll and Gaskill, 1978: Figure 48). In most of these taxa, the nasolacrimal duct is only a single opening, rather than the paired openings seen in Nannaroter, Micraroter, and Llistrofus, and a more medially positioned foramen for the medial nerve is not reported. Due to the small size of most “microsaurs” and the foramina, they may have been overlooked in other taxa that are more poorly preserved.
The postfrontal is a triangular element that has been slightly dislocated so as to be dipping into the right orbit (Figures 6, 7). In the natural condition, the element would be sutured to the frontal anteromedially, to the parietal posteromedially, to the tabular posteriorly, and to the postorbital ventrally. The postfrontal does not contact the lacrimal anteriorly, and the element contributes significantly to the dorsal orbital margin.
The postorbital is a triangular element that sutures to the postfrontal dorsomedially, to the tabular posterodorsally, possibly to the squamosal posteriorly, and to the jugal anteroventrally (Figures 6, 7). The contact with the jugal is difficult to discern because of the beige coloration of the element and the overlapping mandible. The element would also be predicted to suture to the squamosal posteroventrally based on other ostodolepids, but this contact is also not readily identifiable (further discussed below). The postorbital contributes to the dorsal margin of the temporal emargination and to the posterior orbital margin. The element is comparable in morphology to that of Pelodosotis (Carroll and Gaskill, 1978) and differs from that of Tambaroter and BPI 3839, in which the element is more elongate and rectangular (Carroll and Gaskill, 1978; Henrici et al., 2011).
The parietals are incomplete posteriorly in this specimen. The anterior suture with the frontals and the lateral suture with the postfrontals are preserved (Figures 6, 7). Due to the degree of damage it is impossible to confidently state whether a pineal opening was present (as in other ostodolepids) or absent (as in the holotype).
The tabular is a large, square element that forms the posterolateral corner of the skull roof. It is sutured to the postfrontal anteriorly, to the postorbital anteroventrally, and to the squamosal ventrally (Figures 6, 7). In the holotype, the tabular also sutures to the parietal dorsomedially, but because the posterior portion of the parietals is lost and the right temporal region is somewhat dislodged this contact is not present in this new specimen.
A small rectangular fragment of the squamosal was present in the holotype and was interpreted as not contacting the postorbital, a unique feature among ostodolepids that would have increased the relative dorsal extent of the temporal emargination (Anderson et al., 2009). In this specimen, a more complete squamosal is present and permits a greater characterization. The element is square and of a slightly smaller size than the tabular. Its anterior contact is difficult to discern because of the low-contrast beige coloration, but it approaches the narrow posterior process of the postorbital very closely and is separated only by the outward displacement of the squamosal (Figures 6, 7). Thus, it becomes apparent that the temporal emargination does not extend dorsally as previously suggested, but rather that it is of a more typical size and position to other ostodolepids.
The full extent of the temporal emargination, although fully enclosed posteriorly in this specimen in contrast to the holotype, is difficult to define ventrally due to slight dorsolateral displacement of the posterior end of the mandible that obscures this region. Underlying fragments in the palatal region prevent further exploration from the underside. The reconstruction of Tambaroter (Henrici et al., 2011: Figure 4) indicates that the emargination was relatively reduced in this taxon as a result of the posterior expansion of the postorbital and its horizontal, rather than posterodorsally angled, ventral margin, but it is important to note the extreme compression of the holotype. Similar considerations are noted for Micraroter, which is known from poorly preserved skull material in which the temporal emargination is not well defined (e.g., Daly, 1973: Figure 22; Carroll and Gaskill, 1978: Figure 52).
The quadratojugal is a slender, rectangular element that is normally sutured to the squamosal dorsally and to the quadrate ventrally. Although the element remains attached to the squamosal in this specimen, the dislodgment of the latter has altered the nature of the articulation such that the quadratojugal contacts the squamosal only posteriorly (Figures 6, 7). It frames the posterior margin of the temporal emargination, which is seen in other ostodolepids, hapsidopareiids, and lysorophians. The emargination is significantly smaller than that of hapsidopareiids in which it extends nearly to the dorsal skull roof and occupies the majority of the temporal region (Daly, 1973; Bolt and Rieppel, 2009), though the posterior margin of Pelodosotis and in this specimen are similarly framed by a bar formed primarily by the squamosal and the quadratojugal (Carroll and Gaskill, 1978). Although the squamosal is partially dislodged, it does not appear to overlap the quadratojugal posteroventrally, as is the condition in Micraroter, BPI 3839, and Tambaroter (Daly, 1973; Henrici et al., 2011).
The quadrate is a small but robust square element that is sutured to the quadratojugal dorsally and to the articular ventrally (Figures 6, 7). Because of its small size, it has proven difficult to characterize in many “microsaurs,” whether due to poor preservation of sutures, overlapping elements, or taphonomic loss. The element was not preserved in the holotype of Nannaroter but appears essentially indistinguishable from that of other ostodolepids in which it is known (e.g., Pelodosotis, Tambaroter) (Carroll and Gaskill, 1978; Henrici et al., 2011). The exception is Micraroter, which is reconstructed as having a rather elongate quadrate that extends far anterodorsally along the posteroventral margin of the quadratojugal (e.g., Carroll and Gaskill, 1978: Figure 103I), but evidence for this in the actual specimen figures is not unequivocal (e.g., Carroll and Gaskill, 1978: Figure 54D).
Mandible
The right mandible is complete and in articulation with the skull, although it is deflected such that the anterior end lies medial to the maxillary tooth row (Figures 6, 7). The anterior portion of the mandible is entirely comprised of the dentary through the anterior portion of the coronoid process; the element is mostly black except for the posterodorsal most portion that sutures to the surangular. Unlike the holotype there is no evidence for a thin lateral exposure of the ventrally positioned splenial and postsplenial, but this could be the result of taphonomic deformation. The posterior margin of the dentary is angled posterodorsally where it contacts the angular at the ventral portion of the mandible and the surangular at the dorsal portion. The angular is a rectangular element that is sutured to the dentary anteriorly, to the surangular dorsally, and to the articular posteriorly; it is partially damaged in the holotype. It is of a relatively uniform height throughout, comparable to Micraroter, Tambaroter, and BPI 3839 and in contrast to Pelodosotis (Carroll and Gaskill, 1978), and tapers only slightly at the posterior end. The surangular is a crescentic element that was mostly absent in the holotype, sutured to the dentary anteriorly, to the angular ventrally, and to the articular posteriorly (Figures 6, 7). It is broadly expanded dorsally to form the posterior region of the coronoid process. The medial profile of the mandible is obscured by other elements and matrix, and the ventral profile is relatively uninformative beyond identification of the slender splenial and postsplenial, which lie medial to the dentary and the angular, as in all ostodolepids. As with all other ostodolepids, a prominent retroarticular process is preserved on the articular, posterior to the articulation of the quadrate with the mandible (Figures 6, 7).
Results
The phylogenetic analysis produced a majority rule consensus tree (Figure 9) that is identical to that of the one obtained in Gee et al. (2020) when the species level operational taxonomic unit of Euryodus dalyae is used (the polytomy of the three genera Euryodus, Cardiocephalus, and Pariotichus is resolved). As in Gee et al. (2020), Nannaroter is recovered as the sister taxon to the clade containing Micraroter and Pelodosotis.
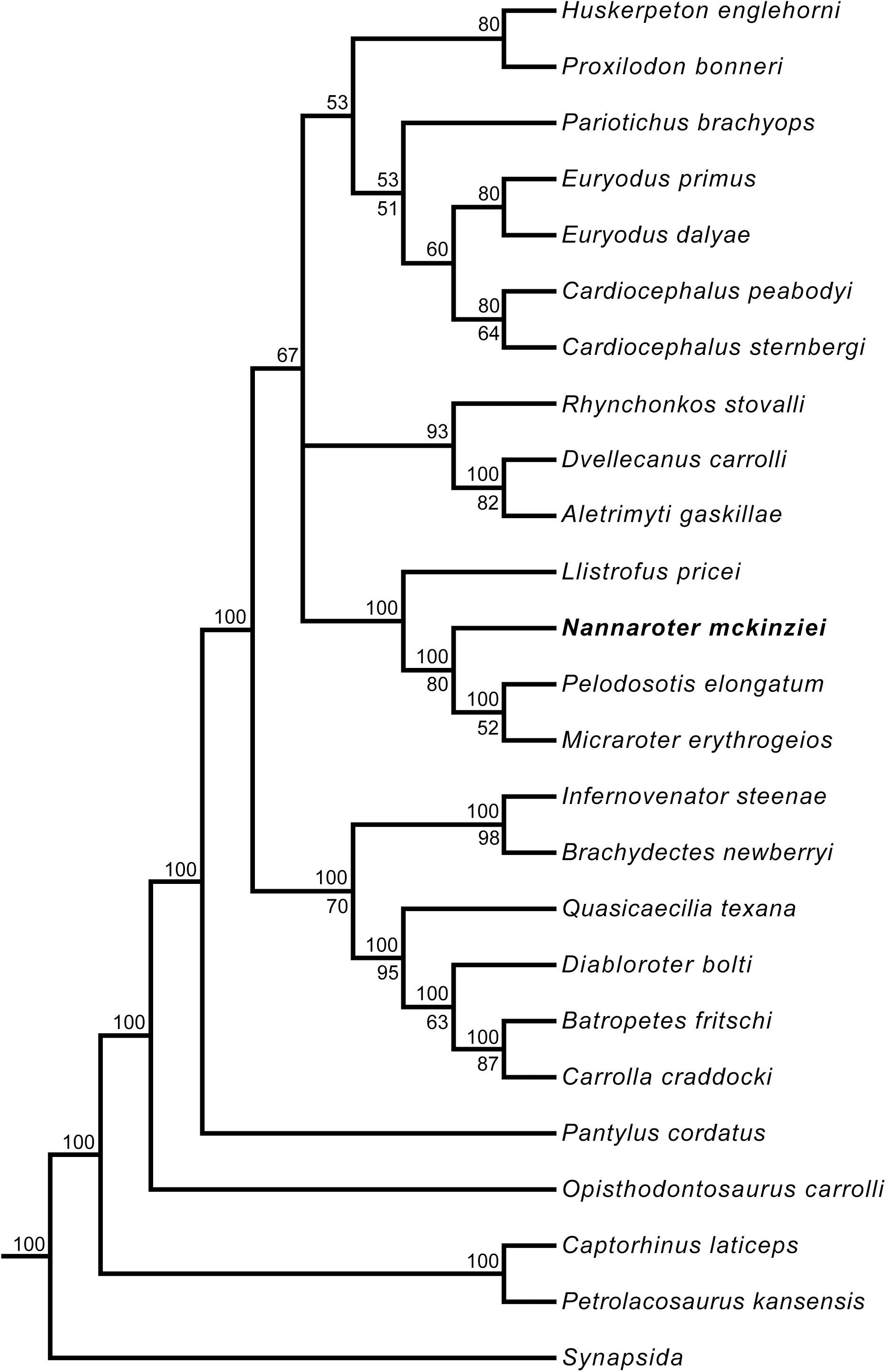
Figure 9. Majority rule consensus tree of the 270 optimal trees obtained from the phylogenetic analysis. Tree length = 1,744, consistency index = 0.2683, rescaled consistency index = 0.1764, retention index = 0.6574. Numbers above nodes represent the frequency of nodes (%). Bootstrap values are found below nodes, if no value is indicated it was less than 50%. Taxa have been trimmed to concentrate on Recumbirostra, the clade of interest.
Discussion
Ecology of Nannaroter
There is little doubt that Nannaroter was at least partially fossorial based on a suite of features (e.g., cranial ossification, reinforcement of orbital rims, heavily interdigitated sutures, recumbent snout) that have been identified in other clades of “microsaurs” and in extant fossorial taxa such as amphisbaenians and caecilians (Sherratt et al., 2014; Müller et al., 2016). The second specimen of Nannaroter and new CT data of the holotype specimen reinforces the original description and provides some new information regarding the mandible (full characterization of the coronoid process, confident presence of a robust retroarticular process) that further supports an inferred fossorial lifestyle. Anderson et al. (2009) suggested that the paucity of Nannaroter at the Richards Spur locality could be accordingly reflective of a low preservation potential associated with fossoriality or that it could be a naturally rare form among the assemblage. Based on cranial material, this taxon remains among the rarest forms at Richards Spur in spite of additional recovery and preparation of material, being of comparable rarity to other co-occurring small taxa, such as the amphibamids Pasawioops (Fröbisch and Reisz, 2008) and Tersomius dolesensis (Anderson and Bolt, 2013) and the phlegethontid Sillerpeton (Lund, 1978). There is no evidence suggesting paucity due to taphonomic sorting of small forms, as the amphibamid Doleserpeton and the eureptile Captorhinus are the most common taxa at the site, and the gymnarthrid Cardiocephalus is also reasonably well documented. Nannaroter is also rarer than the large dissorophoids, Cacops and Acheloma. However, it is worth considering that variable habitat occupation, rather than variable body size, acted as a taphonomic factor for the karstic setting at Richards Spur, as Gee et al. (2019) suggested for the similarly rare Llistrofus. Ostodolepids, and most recumbirostrans in general, are rare wherever they are recorded. Because the postcranium of Nannaroter is unknown, and many features of commonly preserved elements (e.g., vertebrae, ribs) are highly conserved among microsaurs, it is possible that postcranial material of the taxon may exist but is presently unidentifiable in the absence of an association with more informative skeletal elements.
If the rarity of Nannaroter is not attributable to taphonomic factors, the suggestion that it is a rare component of the fauna that was present at Richards Spur (Anderson et al., 2009) may hold more credence. The early Permian Richards Spur locality is dominated by numerous predatory taxa, and there are several small animals at the locality with a similar dentition to Nannaroter that were likely preying on small arthropods (Modesto et al., 2009; MM pers. obs.). These small predators include various eureptiles, parareptiles, and anamniote tetrapods, with Nannaroter being among them; thus it is quite apparent that this was a niche that was heavily exploited by tetrapods at the locality. The number of taxa occupying this niche may be part of the reason for the rarity of Nannaroter with their being a high degree of competition for prey items by small tetrapods.
Conclusion
This study adds to our knowledge of the enigmatic recumbirostran “microsaur” Nannaroter mckinziei, formerly known from only a single partial skull. The discovery of a second partial skull, combined with CT scanning of both this and the holotype specimen, has given us the opportunity to better understand the anatomy of this taxon, most notably the palate, lower jaw, and portions of the braincase. This further adds to our knowledge of the early Permian Richards Spur locality and the recumbirostran fauna that was present there.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
MM and JF conceived the study. MM, RS, and BG wrote the manuscript. MM, RS, and JP analyzed the data. MM, RS, and DS made the figures. DS prepared fossil material. MJ and JB scanned fossil material. RR and JF provided access to specimens and data. All authors edited the manuscript.
Funding
This work was supported by a grant from the Deutsche Forschungsgemeinschaft (FR 2457/9-1) to JF, a Humboldt postdoctoral fellowship to MM, and an Elsa-Neumann scholarship to JP. Access to the Australian Synchrotron’s Imaging and Medical Beamline was achieved under grant M13126.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Richard Cifelli, Bill May, and the OMNH for their assistance and support regarding our work on the Richards Spur locality. We also thank Anton Maksimenko for his exceptional technical assistance with the synchrotron imaging.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.739316/full#supplementary-material
References
Anderson, J. S. (2001). The phylogenetic trunk: maximal inclusion of taxa with missing data in an analysis of the lepospondyli (Vertebrata. Tetrapoda). Syst. Biol. 50, 170–193. doi: 10.1080/10635150119889
Anderson, J. S. (2007). “Incorporating ontogeny into the matrix: a phylogenetic evaluation of developmental evidence for the origins of modern amphibians,” in Major Transitions in Vertebrate Evolution, eds J. S. Anderson and H.-D. Sues (Bloomington, IN: Indiana University Press), 182–227.
Anderson, J. S., and Bolt, J. R. (2013). New information on amphibamids (Tetrapoda, Temnospondyli) from Richards Spur (Fort Sill), Oklahoma. J. Vertebr. Paleontol. 33, 553–567. doi: 10.1080/02724634.2013.726676
Anderson, J. S., Scott, D., and Reisz, R. R. (2009). Nannaroter mckinziei, a new ostodolepid ‘microsaur’ (Tetrapoda, Lepospondyli, Recumbirostra) from the early permian of richards spur (Ft. Sill), Oklahoma. J. Vertebr. Paleontol. 29, 379–388. doi: 10.1671/039.029.0222
Bolt, J. R., and DeMar, R. E. (1983). Simultaneous tooth replacement in Euryodus and Cardiocephalus (Amphibia; Microsauria). J. Paleontol. 57, 911–923.
Bolt, J. R., and Rieppel, O. (2009). The holotype skull of Llistrofus pricei carroll and gaskill, 1978 (Microsauria: Hapsidopareiontidae). J. Paleontol. 83, 471–483. doi: 10.1666/08-076.1
Carroll, R. L., and Gaskill, P. (1978). The Order Microsauria. Philadelphia, PA: American Philosophical Society, 232.
Case, E. C. (1929). Description of a nearly complete skeleton of ostodolepis brevispinatus williston, Contr. Mus. Paleontol. Univ. Mich. 3, 81–107.
Daly, E. (1973). A Lower permian vertebrate fauna from Southern Oklahoma. J. Paleontol. 47, 562–589.
Dawson, S. J. W. (1863). Air-Breathers Of The Coal Period: A Descriptive Account Of The Remains Of Land Animals Found In The Coal Formation Of Nova Scotia, With Remarks On Their Bearing On Theories Of The Formation Of Coal And Of The Origin Of Species, Dawson Brothers Burbank, CA: Creative Media Partners, LLC, 114. doi: 10.5962/bhl.title.167425
de Ricqlès, A., and Bolt, J. R. (1983). Jaw growth and tooth replacement in Captorhinus aguti (Reptilia: Captorhinomorpha): a morphological and histological analysis. J. Vertebr. Paleontol. 3, 7–24. doi: 10.1080/02724634.1983.10011952
Fröbisch, N. B., and Reisz, R. R. (2008). A new lower permian amphibamid (Dissorophoidea, Temnospondyli) from the fissure fill deposits near Richards Spur, Oklahoma. J. Vertebr. Paleontol. 28, 1015–1030. doi: 10.1671/0272-4634-28.4.1015
Gee, B. M., Bevitt, J. J., and Reisz, R. R. (2019). Dissorophid diversity at the early Permian cave system near Richards Spur, Oklahoma, USA. Palaeontol. Electronica 22, 1–32. doi: 10.26879/976
Gee, B. M., Bevitt, J. J., and Reisz, R. R. (2020). Computed tomographic analysis of the cranium of the early Permian recumbirostran “microsaur” Euryodus dalyae reveals new details of the braincase and mandible. Pap. Palaeontol. 7, 721–749. doi: 10.1002/spp2.1304
Henrici, A. C., Martens, T., Berman, D. S., and Sumida, S. S. (2011). An ostodolepid ‘microsaur’ (Lepospondyli) from the lower permian tambach formation of central germany. J. Vertebr. Paleontol. 31, 997–1004. doi: 10.1080/02724634.2011.596601
Huttenlocker, A., Pardo, J. D., Small, B. J., and Anderson, J. S. (2013). Cranial morphology of recumbirostrans (Lepospondyli) from the permian of kansas and nebraska, and early morphological evolution inferred by micro-computed tomography. J. Vertebr. Paleontol. 33, 540–552. doi: 10.1080/02724634.2013.728998
Lund, R. (1978). Anatomy and relationships of the family Phlegethontiidae (Amphibia. Aistopoda). Ann. Carnegie Mus. 47, 53–79.
MacDougall, M. J., LeBlanc, A. R. H., and Reisz, R. R. (2014). Plicidentine in the Early Permian parareptile Colobomycter pholeter, and its phylogenetic and functional significance among coeval members of the clade. PLoS One 9:e96559. doi: 10.1371/journal.pone.0096559
MacDougall, M. J., Tabor, N. J., Woodhead, J., Daoust, A. R., and Reisz, R. R. (2017). The unique preservational environment of the Early Permian (Cisuralian) fossiliferous cave deposits of the Richards Spur locality, Oklahoma. Palaeogeogr. Palaeoclimatol. Palaeoecol. 475, 1–11. doi: 10.1016/j.palaeo.2017.02.019
Maddin, H. C., Olori, J. C., and Anderson, J. S. (2011). A redescription of Carrolla craddocki (Lepospondyli: Brachystelechidae) based on high-resolution CT, and the impacts of miniaturization and fossoriality on morphology. J. Morphol. 272, 722–743. doi: 10.1002/jmor.10946
Modesto, S. P., Scott, D. M., and Reisz, R. R. (2009). Arthropod remains in the oral cavities of fossil reptiles support inference of early insectivory. Biol. Lett. 5, 838–840. doi: 10.1098/rsbl.2009.0326
Müller, J., Hipsley, C. A., and Maisano, J. A. (2016). Skull osteology of the Eocene amphisbaenian Spathorhynchus fossorium (Reptilia. Squamata) suggests convergent evolution and reversals of fossorial adaptations in worm lizards. J. Anat. 229, 615–630. doi: 10.1111/joa.12513
Pardo, J. D., Szostakiwskyj, M., Ahlberg, P. E., and Anderson, J. S. (2017). Hidden morphological diversity among early tetrapods. Nature 546, 642–645. doi: 10.1038/nature22966
Romer, A. S. (1945). Vertebrate Paleontology, 2nd Edn. Chicago, IL: University of Chicago Press, 687.
Schultze, H.-P., and Foreman, B. (1981). A new gymnarthrid microsaur from the Lower Permian of Kansas with a review of the tuditanomorph microsaurs (Amphibia). Occas. Pap. Mus. Nat. His. Univ. Kansas. 91, 1–25.
Sherratt, E., Gower, D. J., Klingenberg, C. P., and Wilkinson, M. (2014). Evolution of cranial shape in caecilians (Amphibia: Gymnophiona). Evol. Biol. 41, 528–545. doi: 10.1007/s11692-014-9287-2
Snyder, A. J., LeBlanc, A. R. H., Jun, C., Bevitt, J. J., and Reisz, R. R. (2020). Thecodont tooth attachment and replacement in bolosaurid parareptiles. PeerJ. 8:e9168. doi: 10.7717/peerj.9168
Swofford, D. L. (2003). PAUP∗. Phylogentic Analysis Using Parsimony (∗and Other Methods). Sunderland, MA: Sinauer Associates.
Szostakiwskyj, M., Pardo, J. D., and Anderson, J. S. (2015). Micro-CT Study of Rhynchonkos stovalli (Lepospondyli. Recumbirostra), with Description of Two New Genera. PLoS One 10:e0127307. doi: 10.1371/journal.pone.0127307
Williston, S. W. (1913). Ostodolepis brevispinatus, a new reptile from the Permian of Texas. J. Geol. 21, 363–368. doi: 10.1086/622067
Williston, S. W. (1916). Synopsis of the American Permo-Carboniferous Tetrapoda. Contributions From Walker Museum, Vol. 1, Chicago, IL: The University of Chicago Press. 193–236.
Keywords: Recumbirostra, Microsauria, Reptilia, Sauropsida, Lepospondyli, Palaeozoic, computed tomogaphy
Citation: MacDougall MJ, Seeger R, Gee B, Ponstein J, Jansen M, Scott D, Bevitt JJ, Reisz RR and Fröbisch J (2021) Revised Description of the Early Permian Recumbirostran “Microsaur” Nannaroter mckinziei Based on New Fossil Material and Computed Tomographic Data. Front. Ecol. Evol. 9:739316. doi: 10.3389/fevo.2021.739316
Received: 10 July 2021; Accepted: 26 August 2021;
Published: 22 September 2021.
Edited by:
Martin Daniel Ezcurra, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, ArgentinaReviewed by:
Alexander Wilhelm Armin Kellner, Federal University of Rio de Janeiro, BrazilGuang-Hui Xu, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (CAS), China
Copyright © 2021 MacDougall, Seeger, Gee, Ponstein, Jansen, Scott, Bevitt, Reisz and Fröbisch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mark J. MacDougall, bWFyay5tYWNkb3VnYWxsQG1mbi5iZXJsaW4=