Corrigendum: Necromys lasiurus: Lessons From a 38-Year Study in an Amazonian Savanna
 Clarissa Alves da Rosa1
Clarissa Alves da Rosa1 Pedro Henrique Salomão Ganança2*
Pedro Henrique Salomão Ganança2* Albertina Pimentel Lima1William Ernest Magnusson1
Albertina Pimentel Lima1William Ernest Magnusson1- 1Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
- 2Programa de Pós Graduação em Ecologia, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
Amazonian savannas are isolated patches of open habitats within an array of extensive tropical forest. The mammal fauna of the savannas in the Alter do Chão region (Santarém Municipality), is dominated by Necromys lasiurus, whose populations have been studied by researchers of the National Institute of Amazonian Research since 1983. Here, we summarize the studies and advances made so far to better understand aspects related to population dynamics and ecology of savanna rodents and the strategies they use to persist in an environment with frequent fires subject to global climatic influences. In the Amazonian savannas the species acts as a seed disperser and population fluctuations are related to invertebrate availability, but not with fire or vegetation structure. Global climate appears to affect N. lasiurus population dynamics at local scale (i.e., plot scale) but not at the regional scale of the Alter do Chão savannas. The long-term studies in Alter do Chão generated many advances about Necromys lasiurus population dynamics and ecology, including aspects relating to feeding, home range, animal-plant interactions, the effects of fire and climate change.
Introduction
Necromys lasiurus (Rodentia: Cricetidae) is a small (approximately 60 g, with males being slightly larger than females, Cangussu et al., 2002), terrestrial and diurnal rodent widely distributed in open habitats with seasonally dry and arid climates in South America (Paglia et al., 2012). It has been intensively studied in many open formations in South America, especially in the Brazilian Cerrado. It is a generalist species that eats roots, seeds, leaves, and invertebrates (Talamoni et al., 2008). The species is a major food source for predators, such as snakes and owls (Bueno and Motta-Júnior, 2015). In the Cerrado, Necromys lasiurus is terrestrial, with a mean displacement of 42 m in a 24-h period (Vieira et al., 2005), and males move greater distances than females between successive captures (Pires et al., 2010). The mean home range of males is larger than that of females (Pires et al., 2010). The species constructs nests on the ground, and sometimes uses armadillo burrows as nest sites (Vieira et al., 2005).
In Brazil, N. lasiurus is common in open habitats of the Cerrado, Pantanal and Caatinga (Souza and Alho, 1980; Becker et al., 2007), but also occurs in open patches within Amazonian and Atlantic forests (Gheler-Costa et al., 2012; Magnusson et al., 2021). In the Cerrado, N. lasiurus is usually the most abundant rodent within its area of occurrence and predominates in most vegetation physiognomies (Becker et al., 2007; Pires et al., 2010). In Amazonia, the species is found in savanna vegetation (Ghizoni et al., 2005) and in the Atlantic Forest it is often found in agroecosystems (Gheler-Costa et al., 2012). Santos et al. (2018) and Malange et al. (2016) noted that crops and pastures have favored the expansion of the species in the Atlantic Forest, and human-modified environments are associated with the occurrence of the species in all Brazilian domains (Santos et al., 2017).
Necromys lasiurus is considered to be one of the main hantavirus reservoirs in Brazil (Oliveira et al., 2013; Santos et al., 2017), and hence surveillance of its populations is of important. This species shows abrupt density fluctuations, and changes from extremely common to rare between seasons or years (Souza and Alho, 1980). Although N. lasiurus is a common and well-studied species, especially in Cerrado domain, there are few studies of the population dynamics of N. lasiurus in Amazonian savannas (e.g., Francisco et al., 1995; Magnusson et al., 2021).
Amazonian savannas are found as islands of vegetation between forest patches of varying size and are considered one of the most threatened Amazonian habitats because many of the savanna patches are being transformed by large-scale agriculture, increases in infrastructure, and uncontrolled fires, all of which are interlinked (Diniz and Santos, 2005).
Most of the studies carried out in Amazonian savannas have focused on plants, with only occasional studies of reptiles, birds, mammals, and invertebrates (De Carvalho and Mustin, 2017), suggesting that Amazonian-savanna biodiversity is underestimated. In 2004, the Brazilian Ministry for the Environment recognized the Amazonian savanna complexes as conservation priorities for the country (Ministério do Meio Ambiente [MMA], 2004), resulting in an increase of more than 60% in the number of protected areas in Amazonian savannas in Brazilian territory. Today, 36.8% of Brazilian Amazonian savannas are under some degree of protection, almost half of which are in indigenous lands (De Carvalho and Mustin, 2017). Despite the Brazilian policies to protect Amazonian savannas during the last 20 years, savannas are often the first environments lost, since major international attention has been focused on forests (Ratter et al., 1997). One exception to the paucity of knowledge of areas covered by Amazonian savannas is the Amazon savanna of Alter do Chão, located in the Brazilian state of Pará. Studied since 1983, the cumulative knowledge provided by the studies at Alter do Chão have turned the region into a laboratory for scientific investigation of ecological processes of savannas and fragmented tropical areas. An iconic example is comprised by the long-term studies carried out on Necromys lasiurus populations in the Alter do Chão savannas.
In this paper, we present the advances in knowledge of the biology of N. lasiurus and how the species has been used to study ecological and climate-change process in the Alter do Chão region since 1983. Necromys lasiurusis relatively abundant in the savannas of Alter do Chão and the installation of standardized plots and subsequent funding from diverse sources, allowed a program to study the abiotic and biotic factors that influence the long-term dynamics of populations of the species in Amazonian savannas.
History of Sampling Design of Necromys lasiurus Populations
Our study was conducted near Alter do Chão, a village in the Santarém municipality, Pará. The Alter do Chão region has supported typical savanna tree species for about 1.49 Ma (Buzatti et al., 2018), with a local landscape relatively stable for at least the last 7,000 to 6,000 years (Sanaiotti et al., 2002). The area is a project within the International Long-Term Ecological Research (LTER) Network (Mirtl et al., 2018) that seeks to investigate the local and global drivers of biodiversity change, such as fire and global warming, in savanna and forested environments.
Sampling of Necromys lasiurus began in 1983 in a single plot of approximately 4 ha using baited (Brazil nuts and peanut butter mixed with oats) Sherman traps (10 cm × 10 cm × 30 cm; H. B. Sherman Traps, Inc., Tallahassee, Florida) that were checked once daily, during the morning. The plot originally had a 150 m × 200 m size with a minimum spacing between capture points of 10 m. Traps were set on 25% of the plot intersections during 3 or 4 days and then advanced one trap position so that the whole plot was covered. In 1986, the plot size was increased to 200 m × 200 m, and in 1988 the plot was extended to 300 m × 340 m. However, researchers realized that the 200 m × 200 m plot was sufficient to have a good sample and, in 1997, the plot decreased to 200 m × 200 m. After this, the traps were placed every 20 m for 2 days, then moved 10 m to cover the intermediate positions for another 2 days. On that plot, 100 Sherman traps were used simultaneously and this plot has been sampled with this protocol since 1997. Temporal variation in abundance of N. lasiurus on this plot was obtained from 1985 to 1989, from 1997 to 1998, and 2000 to 2020, giving data in 23 of the 38 years. Details of the methods are given in Francisco et al. (1995); Magnusson et al. (1995), and Magnusson et al. (2010).
In 1999, 40 sampling plots were installed in the savannas across Alter do Chão, allowing studies on a regional scale to evaluate short and long-term patterns of biodiversity change and its drivers (Magnusson et al., 2013). Plots were sampled along four parallel 250-m-long transects that are 50 m apart (Figure 1). This standardized protocol has been repeated till now and can be used to evaluate fluctuations of Necromys lasiurus populations. In the first studies (Francisco et al., 1995; Magnusson et al., 1995), animals were marked by toe clipping. Already in 1999, the first year of sampling, many individuals were recaptured in each session and it became obvious that 4 days was more time than necessary, and the mice were becoming trap happy. Therefore, the effort in the 40 standardized plots was reduced to 2 days, allowing, in the following years, the sampling of the plots in a shorter time. Since 1999, in each of the regional plots, 50 Sherman live traps are set at 20-m intervals on four 250-m parallel transects spaced 50 m apart that covers the plot (Figure 1). To distribute the 50 traps into the four transects, we standardized the installation of 13 traps in the first and third transects and 12 traps in the second and fourth transects. Traps remained open for 2 days in each capture session and were checked each morning. Detailed descriptions of the methods are provided in Layme et al. (2004); Ghizoni et al. (2005), and Magnusson et al. (2021).
FIGURE 1
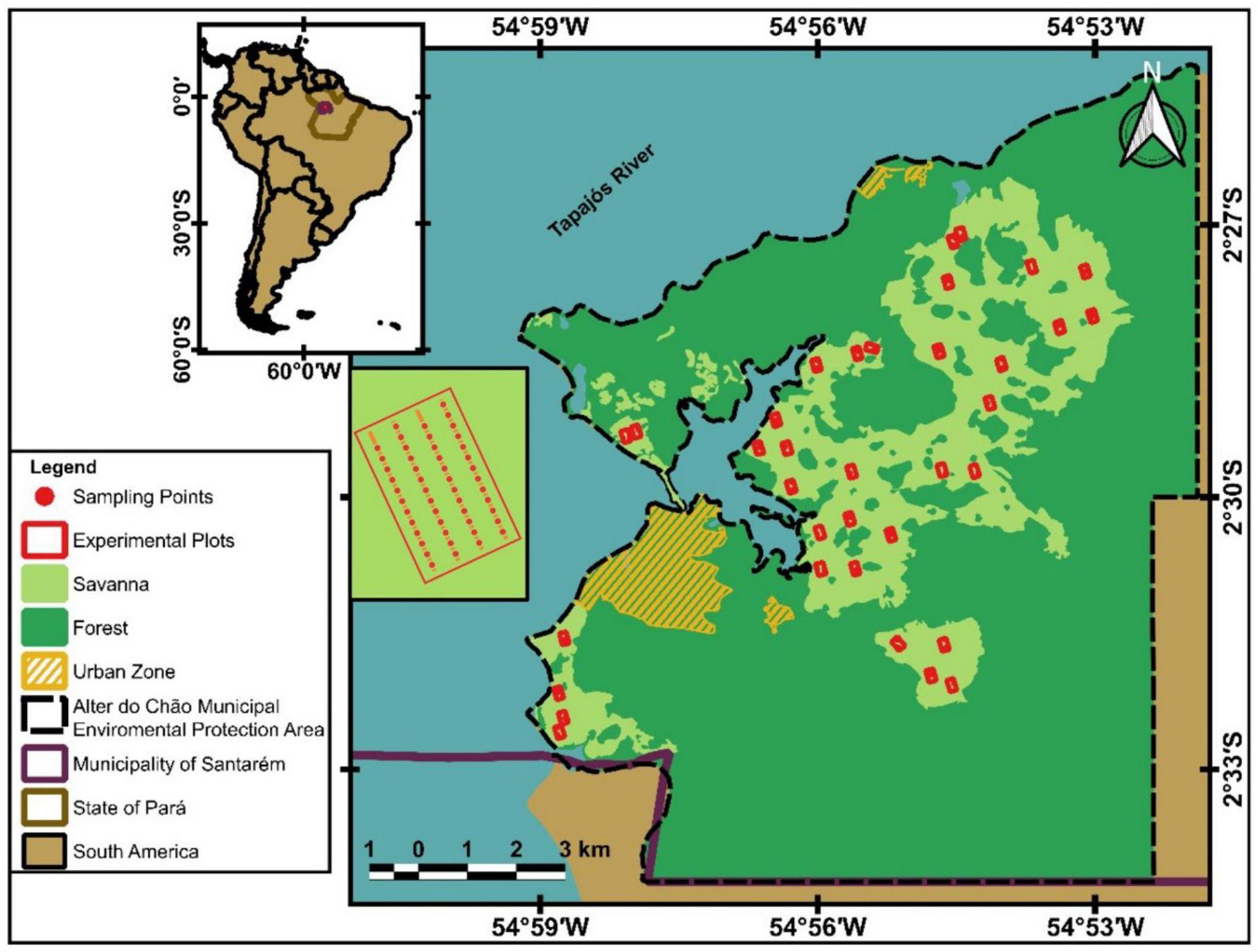
Figure 1. Sampling areas used to study Necromys lasiurus near Alter do Chão, Santarém, Pará.
In summary, two long-term studies with comparable protocols were carried out in the Alter do Chão savanna simultaneously. One study used a single 4-ha plot, with an effort of 4 days and 100 traps placed simultaneously (e.g., Francisco et al., 1995; Magnusson et al., 1995, 2010), and the other study used up to 40 plots (at present 35) with a sampling effort of 2 days and 50 traps placed simultaneously for each plot (e.g., Layme et al., 2004; Ghizoni et al., 2005; Magnusson et al., 2021). Between 2005 and 2020, five plots were deactivated because of illegal land clearing, resulting in 35 active plots today.
Natural History of Necromys lasiurus in the Alter Do Chão Savanna
During the two decades of sampling, N. lasiurus was the only rodent trapped regularly in the savanna in our study region. Individuals of Proechimys sp. and Didelphis marsupialis were occasionally caught in the traps, but were never recaptured and probably use the savanna only in transit, since both species are common in neighboring forest patches (Magnusson et al., 1995; Borges-Matos et al., 2016). In contrast, despite extensive trapping, only one individual of N. lasiurus has been captured in neighboring forest patches (Magnusson et al., 1995).
The first published paper about N. lasiurus near Alter do Chão by Magnusson and Sanaiotti (1987) found many Miconia albicans seeds in the feces of these mice. Miconia albicans is a common shrub in Amazonian savanna near Alter do Chão and, although its fruits are rarely eaten by birds or lizards, they are often consumed by N. lasiurus. The seeds can germinate after passing through the digestive tract of the mice, indicating that they may be important dispersal agents for this plant and, to the best of our knowledge, the study by Magnusson and Sanaiotti (1987) was the first to document seed dispersal through the digestive tract of a small rodent.
In addition to M. albicans seeds, Francisco et al. (1995) showed that the diet of N. lasiurus is composed of vegetable material and invertebrates (mostly centipedes, beetles, and termites). Fire at the end of the dry season appeared to affect the diet of the mice, causing increased consumption of invertebrates during the following wet season (Francisco et al., 1995). This consumption pattern is consistent with observations made for N. lasiurus in the Cerrado (Talamoni et al., 2008), and for Mastomys natalensis, a small rodent common in savannas of South Africa (Monadjem, 1998). In both cases, rodents consumed more invertebrates during the rainy season, and less during the dry season when there was an increase in vegetable items. These studies indicate that small rodents living in savanna environments are opportunistic, varying their diets depending on the abundance of food resources.
To determine the proportions of food sources that came from food chains based on grasses or bushes, Magnusson et al. (1999) measured the stable isotopes of carbon extracted from tissue samples. Although the mice spent most of their time among the grasses, where they would be protected from predators, such as raptors, the stable isotopes indicated that most of their food (∼60%) came from chains that originated in bushes and trees (Magnusson et al., 1999). A comparable pattern is found in African savanna rodent communities, where rodent activity and amount of food removed is greater under shrubs than under grasses, as an answer to the perception of low predation risk under shrubs (Loggings et al., 2019). Therefore, the balance between tree-bush clumps for nutrition and grasses for protection is likely to affect the densities of N. lasiurus and other rodents in savanna environments.
Near Alter do Chão, N. lasiurus has a mean home-range size that varies among years and between sexes. Mean home-range size for males varied from 4,138 to 21,880 m2 (one male, whose home range overlapped the perimeter of the study area, used 51,000 m2 within the plot) and for females varied from 1,317 to 5,480 m2 (Magnusson et al., 1995). The home-range sizes in Amazonian savanna reported by Magnusson et al. (1995) were higher than those reported in studies in the Cerrado vegetation in central Brazil (Souza and Alho, 1980; Alho and Pereira, 1985) and grassland among Atlantic-coastal-forest fragments in south-eastern Brazil (Pires et al., 2010).
In the Alter do Chão savanna, females have smaller, largely exclusive, home ranges, indicating that they are strongly territorial, and male home ranges are larger and overlapping (Magnusson et al., 1995). Because of a perpetual cycle of gestation and lactation, adult females probably need more food, so they defend their territories against potential intruders. In contrast, adult males probably have enough food resources, so they can spend more energy to find receptive females to reproduce (Magnusson et al., 1995). Males of N. lasiurus also had larger home ranges than females in the study by Pires et al. (2010) in grassland among Atlantic-coastal-forest fragments, but home-range sizes did not vary significantly between seasons for either sex.
In our study region, Necromys lasiurus has a short life cycle (a little over a year) and consequently the species has a fast rate of replacement of individuals in its populations (Francisco et al., 1995; Magnusson et al., 1995). Francisco et al. (1995) found a proportion of males and females of 1:1, with a quarter of the individuals being juveniles (weighing less than 30 g) and the remainder adults (weighing above 30 g). Reproduction and growth of N. lasiurus are strongly seasonal, with an increase in the rainy season (January to May), which may be related to the availability of fruits and invertebrates, which are the main food resources of N. lasiurus (Francisco et al., 1995; Layme et al., 2004). This pattern was also observed in a 6-year study in a savanna in central Brazil in which the captures of reproductive individuals of N. lasiurus were significantly higher in the rainy season (Rocha et al., 2017).
Necromys lasiurus as an Indicator of Ecological Process and Climate Change in the Savanna
The first studies showed that the species was relatively common in the Alter do Chão savanna and easily captured (Francisco et al., 1995; Magnusson et al., 1995; Layme et al., 2004), which appeared to make N. lasiurus an excellent study organism. The studies by Layme et al. (2004) and Ghizoni et al. (2005) carried out in the 40 sampling plots of the Alter do Chão savanna, investigated the densities and rates of increase of N. lasiurus in relation to food availability (invertebrates), fire, and vegetation structure. Densities of N. lasiurus were not influenced by vegetation structure or fire, but were strongly related to invertebrate availability (Layme et al., 2004). Rates of increase of N. lasiurus among years indicated that the mice were generally decreasing throughout the study area, but the degree and sometimes the direction of density changes varied among plots (Ghizoni et al., 2005). Reductions in density were smaller in plots where the availability of invertebrates increased and was also related to vegetation structure (proportion of shrubs, tall-grass species, shortgrass species, and open soil along the central line of each plot). The study of Layme et al. (2004), showed that, at large scale, there was a slight tendency to capture more individuals in areas with higher cover of tall-grass, and the densities of N. Lasiurus were not associated with short-grass or shrubs. At scales of tens of meters, more individuals were associated with areas that had a combination of short-grass and shrubs. However, Ghizoni et al. (2005) showed that the hypothesis of Layme et al. (2004) that densities of N. lasiurus are associated with tall grasses was not supported. Density and population growth rates were more associated with tree, shrub, and small grass, an effect that had not been detected when only density, and not density change, had been studied (Ghizoni et al., 2005). Fire appeared to have little direct effect on the densities or rates of increase of N. Lasiurus (Layme et al., 2004; Ghizoni et al., 2005). Also, fire did not influence reproduction or growth of N. lasiurus in a single 4 ha plot (Francisco et al., 1995), which is consistent with studies conducted in Cerrado areas of Central Brazil (Vieira and Marinho-Filho, 1998).
Previous studies in the Cerrado indicate that, although the species is captured frequently, populations fluctuate markedly and may disappear from trapping grids (e.g., Souza and Alho, 1980; Valle et al., 1982). However, during the first 6 years of sampling in the late 1980s and early 1990s in a single 4 ha plot, the number of mice captured in the Alter do Chão savanna was relatively constant and zero density was not registered in any trapping session (Francisco et al., 1995; Magnusson et al., 1995). Nevertheless, subsequent studies carried out in the early 2000s showed that the density of N. lasiurus was declining (Layme et al., 2004; Ghizoni et al., 2005) and the species disappeared from the plot previously sampled (Magnusson et al., 2010). Fluctuations in the population size of N. lasiurus over 22 years in a single plot were associated with local rainfall (Magnusson et al., 2010), which was largely controlled by the Southern Oscillation Index (SOI), which gives an indication of the development and intensity of El Niño or La Niña events in the Pacific Ocean by measuring the periodic interannual variation in global patterns of atmospheric and oceanic circulation that are correlated with variations in global climate patterns (Ropelewski and Halpert, 1987). Densities of N. lasiurus increased with the intensity of SOI and the extent of fire in the region. SOI affected rainfall and reduced the extent of fire. Nevertheless, the amount of rain had no measurable direct effect on mouse densities (Magnusson et al., 2010). The study by Magnusson et al. (2010) was carried out in a single plot of 4 ha, which is insufficient to capture regional fluctuations of the species, although studies on the population dynamics of rodents are often carried out in plots of similar or smaller size (e.g., Vieira, 2003; Rocha et al., 2011; Ribeiro et al., 2019). The studies by Layme et al. (2004); Ghizoni et al. (2005), and Magnusson et al. (2010) carried out in the Alter do Chão savanna suggested that the drivers of the population dynamics of the species can vary spatially and temporally.
To better understand the potential of climate change to impact the species, studies by Layme et al. (2004); Ghizoni et al. (2005), and Magnusson et al. (2010) were extended for another 9 years in eight permanent plots distributed over an area of approximately 16,000 ha (Magnusson et al., 2021). The aim of this study was to determine whether fluctuations in different areas were correlated, and whether these local fluctuations were related to climatic variables, such as local rainfall and SOI. The results showed that, considering all plots together, there was no effect of SOI or local rainfall. Overall, the density tended to decrease during the study period, with peaks in 2000, 2004, and 2013. However, a video of the density fluctuations throughout the period (Magnusson et al., 2021: supplement) indicated that there were three distinct clusters of plots in terms of geographic distribution and coordinated rates of change. Necromys lasiurus in individual plots do not form a population, and are influenced by immigration from neighboring plots. Grouping the data from each cluster, resulted in different conclusions. Density variations in one cluster were related to SOI, but those in the other clusters were not (Magnusson et al., 2021).
Since most studies of small mammals are carried out in a single plot of four hectares or less (e.g., Francisco et al., 1995; Magnusson et al., 2010; Rocha et al., 2011; Ribeiro et al., 2019), they might not reflect the regional population dynamics of small rodents. We are also wary of conclusions from studies undertaken when a species has very high densities. This facilitates field work and statistical analyses. Nevertheless, many species of rodents are at low density most of the time and what regulates densities of a species in the system may differ from the factors that affect its population dynamics when at high densities (Magnusson et al., 2021). Although our data clearly indicate that N. lasiurus at Alter do Chão is limited by invertebrate availability when at high densities, that might not explain most of the fluctuations in density. Global effects, such as SOI, and local rainfall may affect some areas, but much of the fluctuations in density seem to be unrelated to these phenomena. We suspect that the metapopulation dynamics of the mice are mainly affected by other factors, and a prime candidate for future studies is the effect of disease on local populations.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
In the last 3 years the data collection was financed by PELD (LTER)-POPA through the Conselho Nacional de Desenvolvimento Científco e Tecnológico (CNPq), grant number 441443/2016-8. Over the 20 years, INPA PPI-PRJ-1223 financed travel for AL. CNPq grants 446208/2015-9, 401120/2016-3, and 401327/2012-4 financed field assistants and material throughout the study. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil (CAPES)—PROEX n. 0742/2020, for PHSG (Ph.D. scholarship).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Laudelino Sardinha and Edivaldo Vasconcelos for helping with data collection and Isla Marialva for preparing the map.
References
Alho, C., and Pereira, L. (1985). Population ecology of a cerrado rodent community in central Brazil. Rev. Bras. Biol. 45, 597–607.
Becker, R. G., Paise, G., Baumgarten, L. C., and Vieira, E. M. (2007). Estrutura de comunidades de pequenos mamíferos e densidade de Necromys lasiurus (Rodentia, Sigmodontinae) em áreas abertas de cerrado no brasil central. Mastozool. Neotrop. 14, 157–168.
Borges-Matos, C., Aragón, S., Nazareth, M., Fortin, M., and Magnusson, W. E. (2016). Importance of the matrix in determining small-mammal assemblages in an Amazonian forest-savanna mosaic. Biol. Conserv. 204, 417–425. doi: 10.1016/j.biocon.2016.10.037
Bueno, A. A., and Motta-Júnior, J. C. (2015). Behavioural and morphological strategies by small savannah rodents to avoid predation. Mamm. Biol. 80, 401–408. doi: 10.1016/j.mambio.2015.05.005
Buzatti, R. S., de, O., Pfeilsticker, T. R., de Magalhães, R. F., Bueno, M. L., Lemos-Filho, J. P., et al. (2018). Genetic and historical colonization analyses of an endemic savanna tree, qualea grandiflora, reveal ancient connections between amazonian savannas and cerrado core. Front. Plant Sci. 9:981. doi: 10.3389/fpls.2018.00981
Cangussu, S. D., Vieira, F. G., and Rossoni, R. B. (2002). Sexual dimorphism and seasonal variation in submandibular gland histology of Bolomys lasiurus (Rodentia, Muridae). J. Morphol. 254, 320–327. doi: 10.1002/jmor.10041
De Carvalho, W. D., and Mustin, K. (2017). The highly threatened and little known Amazonian savannahs. Nat. Publ. Gr. 1:100. doi: 10.1038/s41559-017-0100
Diniz, A. M. A., and Santos, R. O. (2005). O vertiginoso crescimento populacional de Roraima e seus impactos socioambientais. Cad Geografia 15, 23–44.
Francisco, A., Magnusson, W. E., and Sanaiotti, T. M. (1995). Variation in growth and reproduction of Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna. J. Trop. Ecol. 11, 179–188. doi: 10.1017/S0266467400008622
Gheler-Costa, C., Vettorazzi, C. A., Pardini, R., and Verdade, L. M. (2012). The distribution and abundance of small mammals in agroecosystems of southeastern Brazil. Mammalia 76, 185–191. doi: 10.1515/mammalia-2011-0109
Ghizoni, I. R., Layme, V. M. G., Lima, A. P., and Magnusson, W. E. (2005). Spatially explicit population dynamics in a declining population of the tropical rodent, Bolomys lasiurus. J. Mammal. 86, 677–682. doi: 10.1644/1545-1542(2005)086[0677:sepdia]2.0.co;2
Layme, V. M. G., Lima, A. P., and Magnusson, W. E. (2004). Effects of fire, food availability and vegetation on the distribution of the rodent Bolomys lasiurus in an Amazonian savanna. J. Trop. Ecol. 20, 183–187. doi: 10.1017/S0266467403001263
Loggings, A. A., Shrader, A. M., Monadjem, A., and McCleery, R. A. (2019). Shrub cover homogenizes small mammals‘ activity and perceived predation risk. Sci. Rep. 9:16857. doi: 10.1038/s41598-019-53071-y
Magnusson, W. E., Braga-Neto, R., Baccaro, F. P. F., Penha, H. B. J., Verdade, D. R. L. M., Albernaz, A. L. A. L., et al. (2013). “Biodiversidade e monitoramento ambiental integrado,” in Biodiversity and Integrated Environmental Monitoring, eds M. L. Oliveira, F. Baccaro, R. Braga-Neto, and W. E. Magnusson (Santo André, SOA: Áttema Editorial).
Magnusson, W. E., De Araújo, M. C., Cintra, R., Lima, A. P., Martinelli, L. A., Sanaiotti, T. M., et al. (1999). Contributions of C3 and C4 plants to higher trophic levels in an Amazonian savanna. Oecologia 119, 91–96. doi: 10.1007/PL00008821
Magnusson, W. E., De Lima Francisco, A., and Sanaiotti, T. M. (1995). Home-range size and territoriality in Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna. J. Trop. Ecol. 11, 179–188. doi: 10.1017/S0266467400008622
Magnusson, W. E., Layme, V. M. G., and Lima, A. P. (2010). Complex effects of climate change: population fluctuations in a tropical rodent are associated with the southern oscillation index and regional fire extent, but not directly with local rainfall. Glob. Chang. Biol. 16, 2401–2406. doi: 10.1111/j.1365-2486.2009.02140.x
Magnusson, W. E., and Sanaiotti, T. M. (1987). Dispersal of miconia seeds by the rat bolomys lasiurus. J. Trop. Ecol. 3, 277–278. doi: 10.1017/S0266467400002169
Magnusson, W., Rosa, C., Layme, V. M. G., Ghizoni, I. R., and Lima, A. P. (2021). Local effects of global climate on a small rodent Necromys lasiurus. J. Mammal. 102, 188–194. doi: 10.1093/jmammal/gyaa140
Malange, J., Izar, P., and Japyassú, H. (2016). Personality and behavioural syndrome in Necromys lasiurus (Rodentia: Cricetidae): notes on dispersal and invasion processes. Acta Ethol. 19, 189–195. doi: 10.1007/s10211-016-0238-z
Mirtl, M. T., Borer, E., Djukic, I., Forsius, M., Haubold, H., Hugo, W., et al. (2018). Genesis, goals and achievements of long-term ecological research at the global scale: a critical review of ILTER and future directions. Sci. Total Environ. 626, 1439–1462. doi: 10.1016/j.scitotenv.2017.12.001
Ministério do Meio Ambiente [MMA] (2004). Portaria 126. Available online at: https://www.ibama.gov.br/component/legislacao/?view=legislacao&legislacao=111036 (accessed April 20, 2021).
Monadjem, A. (1998). Reproductive biology, age structure, and diet of Mastomys natalensis (Muridae: Rodentia) in a Swaziland grassland. Mamm. Biol. 63, 347–356.
Oliveira, S. V., Escobar, L. E., Peterson, A. T., and Gurgel-Gonçalves, R. (2013). Potential geographic distribution of hantavirus reservoirs in Brazil. PLoS One 8:e85137. doi: 10.1371/journal.pone.0085137
Paglia, A. P., Fonseca, G. A. B., Rylands, A. B., Herrmann, G., Aguiar, L. M. S., Chiarello, A. G., et al. (2012). Lista Anotada dos Mamíferos do Brasil. Arlington, VA: Conservation International.
Pires, A., dos, S., Fernandez, F. A., dos, S., Feliciano, B. R., and de Freitas, D. (2010). Use of space by Necromys lasiurus (Rodentia, Sigmodontinae) in a grassland among Atlantic Forest fragments. Mamm. Biol. 75, 270–276. doi: 10.1016/j.mambio.2009.04.002
Ratter, J. A., Ribeiro, J. F., and Bridgewater, S. (1997). The Brazilian cerrado vegetation and threats to its biodiversity. Ann. Bot. 80, 223–230. doi: 10.1006/anbo.1997.0469
Ribeiro, J. F., Guaraldo, A., Nardoto, G. B., Santoro, G., and Vieira, E. M. (2019). Habitat type and seasonality influence the isotopic trophic niche of small mammals in a neotropical savanna. Hystrix 30, 30–38. doi: 10.4404/hystrix-00150-2018
Rocha, C. R., Ribeiro, R., and Marinho-Filho, J. (2017). Influence of temporal variation and seasonality on population dynamics of three sympatric rodents. Mamm. Biol. 84, 20–29. doi: 10.1016/j.mambio.2017.01.001
Rocha, C. R., Ribeiro, R., Takahashi, F. S. C., and Marinho-Filho, J. (2011). Microhabitat use by rodent species in a central Brazilian cerrado. Mamm. Biol. 76, 651–653. doi: 10.1016/j.mambio.2011.06.006
Ropelewski, C. F., and Halpert, M. S. (1987). Global and Regional Scale Precipitation Patterns Associated with the El Niño/Southern Oscillation. Mon. Weather Rev. 115, 1606–1626. doi: 10.1175/1520-049319871152.0.co;2
Sanaiotti, A. T. M., Martinelli, L. A., Victoria, R. L., Trumbore, S. E., and Camargo, P. B. (2002). Past vegetation changes in amazon savannas determined using carbon isotopes of soil organic. Biotropica 34, 2–16. doi: 10.1646/0006-3606(2002)034[0002:pvcias]2.0.co;2
Santos, F. O., Teixeira, B. R., Cordeiro, J. L. P., Sousa, R. H. A., Lucio, C. S., Gonçalves, P. R., et al. (2018). Expansion of the range of Necromys lasiurus (Lund, 1841) into open areas of the Atlantic forest biome in Rio de Janeiro state, Brazil, and the role of the species as a host of the hantavirus. Acta Trop. 188, 195–205. doi: 10.1016/j.actatropica.2018.08.026
Santos, J. P., Oliveira, S. V., García-Zapata, M. T., and Steinke, V. A. (2017). Does land cover influence the spatial distribution of reservoir rodent Necromys lasiurus? SOJ Microbiol. Infect. Dis. 5, 1–5. doi: 10.15226/sojmid/5/4/00179
Souza, M., and Alho, C. (1980). Distribuiçao espacial do roedor silvestre Zygodontomys lasiurus em habitat natural do cerrado. Bras. Florest. 44, 31–74.
Talamoni, S. A., Couto, D., Cordeiro Júnior, D. A., and Diniz, F. M. (2008). Diet of some species of Neotropical small mammals. Mamm. Biol. 73, 337–341. doi: 10.1016/j.mambio.2007.09.008
Valle, C. M., de, C., Alves, M. C., Santos, I. B., and Varejão, J. B. M. (1982). Observacao sobre dinamica de populacao de Zygodontomys lasiurus (Lund, 1841), Colomys expulsus (Lund, 1841) e Oryzomys subflavus (Wagner,1842) em vegetaçãoo de cerrado no Vale do Rio das Velhas (Prudente de Morais, Minas Gerais, Brasil- 1981) Rodent. Lundian 2, 70–83.
Vieira, E. M., and Marinho-Filho, J. (1998). Pre- and post-fire habitat utilization by rodents of cerrado from central Brazil1. Biotropica 30, 491–496. doi: 10.1111/j.1744-7429.1998.tb00086.x
Vieira, M. V. (2003). Studies on neotropical fauna and environment seasonal niche dynamics in coexisting rodents of the Brazilian cerrado. Stud. Neotrop. Fauna Environ. 38, 7–15. doi: 10.1076/snfe.38.1.7.14034
Keywords: long-term studies, population dynamics, Rodent ecology, savanna mice, Tropical savanna
Citation: da Rosa CA, Ganança PHS, Lima AP and Magnusson WE (2021) Necromys lasiurus: Lessons From a 38-Year Study in an Amazonian Savanna. Front. Ecol. Evol. 9:716384. doi: 10.3389/fevo.2021.716384
Received: 28 May 2021; Accepted: 23 August 2021;
Published: 17 September 2021.
Edited by:
Ara Monadjem, University of Eswatini, EswatiniReviewed by:
Ricardo Bovendorp, Universidade Estadual de Santa Cruz, BrazilCaryne Braga, State University of the North Fluminense Darcy Ribeiro, Brazil
Copyright © 2021 da Rosa, Ganança, Lima and Magnusson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro Henrique Salomão Ganança, psalogan@hotmail.com