 Ronglei Duan1,2,3
Ronglei Duan1,2,3 Haicheng Wei
Haicheng Wei Guangliang Hou
Guangliang Hou Jingyi Gao
Jingyi Gao Zhanjie Qin
Zhanjie Qin- 1Key Laboratory of Comprehensive and Highly Efficient Utilization of Salt Lake Resources, Qinghai Institute of Salt Lakes, Chinese Academy of Sciences, Xining, China
- 2Qinghai Provincial Key Laboratory of Geology and Environment of Salt Lakes, Xining, China
- 3Qinghai Provincial Key Laboratory of Physical Geography and Environmental Processes, School of Geographic Science, Qinghai Normal University, Xining, China
Long-term evolution of human-environment interaction in the Qinghai-Tibetan Plateau (QTP) has been discussed intensively in recent years. The identification of human-related pollen types may help explore the coupled process of climate change, ecological response and anthropogenic activities on the QTP. The aim of this study is to evaluate the impact of anthropogenic activities on surface pollen assemblages and identify pollen indicators associated with grazing and cultivation in typical agro-pastoral ecotone of the eastern QTP. Totally 84 surface samples were analyzed from five vegetational communities in the eastern QTP, which are identifiable based on surface pollen assemblages. Principal component analysis of 29 pollen types and two supplementary variables of human influences were used to assess the impact of anthropogenic activities on surface pollen assemblages in eastern QTP. Severe grazing led to increase of Rosaceae, Ranunculaceae, Fabaceae, Taraxacum-Type, Aster-type and Saussurea-Type in pollen assemblages. Highland agricultural cultivation can be identified by the content and morphological characteristic of Brassicaceae and Cerealia Poaceae pollen. The aerial dispersed distances of Brassicaceae and Cerealia Poaceae pollen are very short, and domesticated herbivores are important factors, which disperse the Cerealia Poaceae pollen into pastures in agro-pastoral ecotone of the QTP. Modern pollen indicators of cultivation and grazing activity will provide precise references for fossil pollen study of prehistoric human activities in the QTP.
Introduction
The Qinghai-Tibetan Plateau (QTP), with an average altitude over 4000 m a.s.l., is the highest physical geographical unit in the world and is known as the “third pole” of the earth (Sun et al., 2012; Yao et al., 2017), its alpine cold environment and fragile ecosystem are sensitive to global climate change and anthropogenic activities (Schwalb et al., 2010; Yao et al., 2012). Since the late Pleistocene, a large number of archeological sites of prehistoric human activities have been preserved in the QTP (Madsen et al., 2006; Chen et al., 2015; Zhang et al., 2018; Hou et al., 2020), and extensive anthropogenic activities, including fire use, hunting, farming and grazing, have significantly impacted on the fragile ecology of the QTP (Miehe et al., 2014; Miao et al., 2015; Huang et al., 2017; Wei et al., 2020). Thus, the coupled process of past climate change, ecological response and anthropogenic activities has recently becomes a focus for research on the QTP, due to its extremes of climate and biotic communities (e.g., Schlütz and Lehmkuhl, 2009; Dong et al., 2013; Miehe et al., 2014; Chen et al., 2015; Guedes et al., 2015; Meyer et al., 2017; Gao et al., 2020; Wei et al., 2020; Wang et al., 2020). Pollen is an effective proxy and plays an important role in the study of paleoclimate, vegetation evolution and anthropogenic activities (Ma et al., 2020). Long-term survival and adaptation of human beings on the QTP have significantly impacted on the regional vegetation, which left important clues in the pollen spectrum of sedimentary archives (Huang et al., 2017; Wei et al., 2020). Therefore, accurate application of human-related pollen indicator in paleo-archives is significant for revealing the coupling process and mechanism of climate change, ecological response and anthropogenic activities on the QTP, as well as tracing the adaptation and survival strategies of prehistoric human on the QTP.
In the last decade, palynologists have used human-related pollen types to explore the history of anthropogenic activities on the QTP. Miehe et al. (2009, 2014) suggested that pastoralism in the highlands of the QTP commenced from the 8th millennium BP and ancient pastoralists converted original vegetation into sedge dominated pastures, while Schlütz and Lehmkuhl (2009) identified signs of nomadic activity as early as 7,200 a BP in pollen assemblages at the eastern margin of the QTP. Huang et al. (2017) used the record of Stellera pollen in a sediment core from Genggahai Lake in the northeastern NE-QTP and revealed that the grazing-induced grassland degradation has been started at 4.7 ka and intensified during 3.6–3.0 ka. Similarly, Wei et al. (2020) analyzed the pollen record from Langgeri (LGR) aeolian section in Qinghai Lake basin, and elucidated the intensification of grazing activity since ca. 3.5 ka and extensive grazing activity had affected regional plant community structure, and Cerealia Poaceae pollen appeared since 2.2 ka, suggesting the development of agricultural cultivation in the Qinghai Lake basin.
Nevertheless, modern pollen process is important premise and foundation for accurate interpretation of the fossil pollen spectrum. Assessing the impact of human activities with different pattern and intensity on the surface pollen assemblages, and accurately identifying the human-related pollen signals, are the key points to study the coupling process of climate change, ecological response and anthropogenic activities on the QTP using fossil pollen. However, to date the research works of assessing the impact of human activities on surface pollen assemblages in different vegetation communities and human-related pollen indicators are still lacking on the QTP (Wei et al., 2018). With the development of the local economy over the past decades, the intensification of human activities on the QTP has caused a significant disturbance to the regional vegetation (Zhang et al., 2020). Extensive grazing activity led to changes in vegetation community structure, grassland degradation and reduction in biodiversity (Dong et al., 2004, 2007; Wang et al., 2018). Meanwhile, the native vegetation has been changed by extensive cultivation in the agro-pastoral ecotone in the eastern QTP. Previous studies suggested that the intensification of aeolian activity in the last millennium likely related to the ecological destruction of human activities (Chen et al., 2019, 2020).
Therefore, in this study we collected topsoil samples from different vegetation communities of the typical agro-pastoral ecotone in the eastern QTP. The objective of this study was: (1) to reveal the characteristics of pollen assemblage and distribution patterns in different vegetation communities of the typical agro-pastoral ecotone in the eastern QTP; and (2) to identify the typical pollen indicators of cultivation and grazing activity, which can provide important references for fossil pollen study of paleoecology and prehistoric human activities on the QTP.
Materials and Methods
Region Characteristics
The study area is located in the eastern QTP, with sample sites ranging between 34°27′10″–35°35′E, 100°37′53.26″–102°8′N and elevations of 2,760–3,967 m (Figure 1). The regional climate is relatively cold, with mean annual temperature ranging between −3.7 and 6.1°C. Mean annual precipitation is 230–560 mm/a, with strong summer monsoonal rainfall so that more than 60% of annual precipitation occurs from May to September.
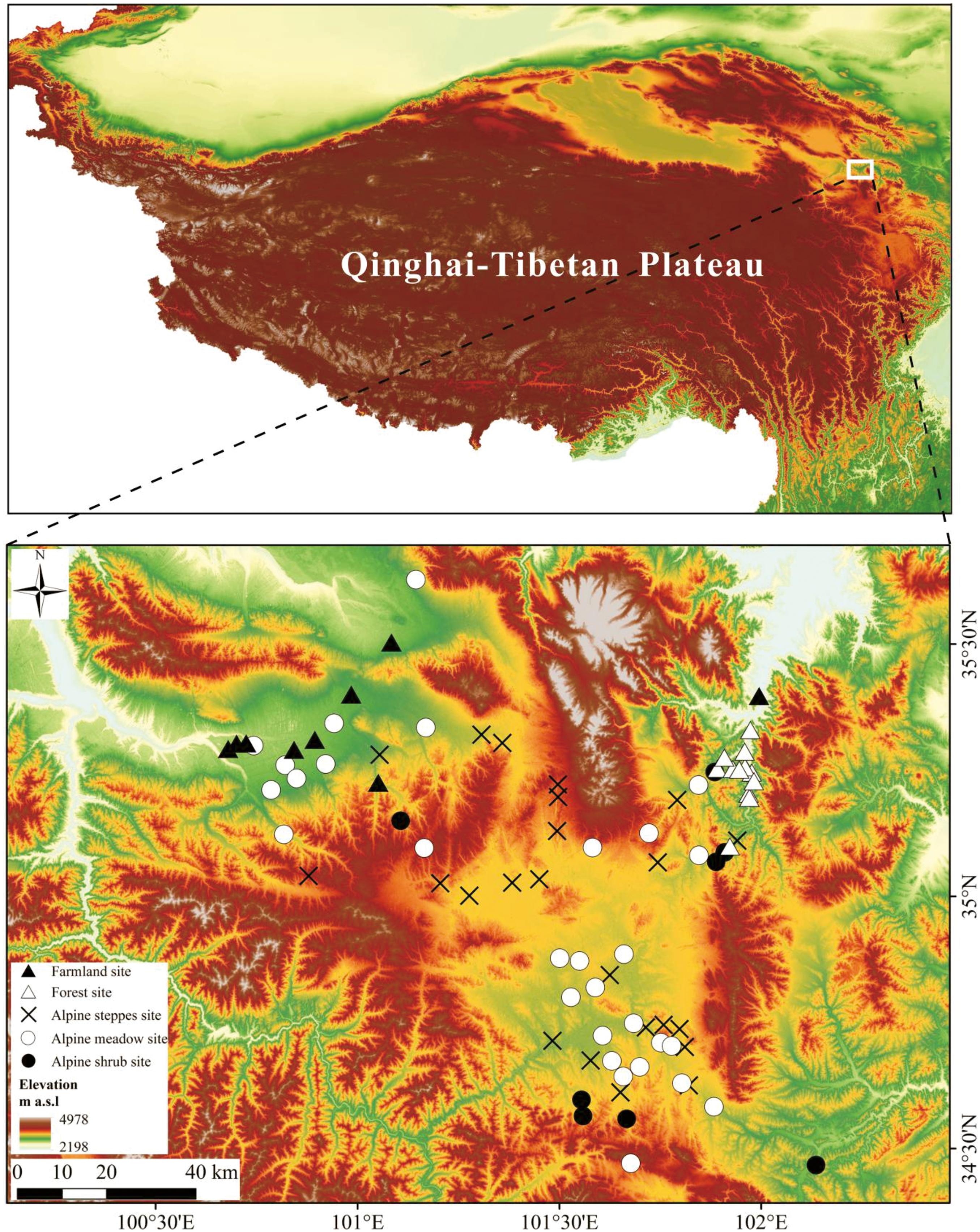
Figure 1. Location map of sampling sites in eastern Qinghai-Tibetan Plateau.
The main vegetation communities in the study area comprise alpine meadow, alpine steppe, alpine shrub, mountain forest, and cropland. Alpine meadows are composed of Kobresia spp. (K. humilis, K. capillifolia), Carex sp., Anemone cathayensis, Trollius chinensis, Gentiana sp. (Gentiana macrophylla, Gentiana siphonantha Maxim., Gentiana futtereri Diels), Potentilla chinensis, Poaceae (Stipa purpurea Griseb., Poa annua L., Elymus nutans Griseb), Trollius chinensis, Leontopodium leontopodioides, Polygonum capitatum Buch., Pedicularis longiflora, Spiraea alpine, Thalictrum sp., and Nardostachys chinensis. Alpine steppe primarily contains Stipa spp., Elymus nutans, Poa annua, Trollius chinensis, Artemisia spp. (Artemisia frigida, Artemisia scoparia, Artemisia desertorum), Sausssurea sp., Taraxacum sp., Oxytropis sp. (Oxytropis ochrocephala Bunge, Oxytropis imbricata), Ligularia virgaurea Maxim., and Stellera chamaejasme Linn. The dominant species in alpine shrub mainly include Rhododendron spp., Dasiphora fruticosa, and Hippophae rhamnoides; Mountain forests are found in the mountain valleys of the eastern study area and mainly composed of Sabina chinensis, Picea purpurea, Betula sp. (Betula platyphylla Suk), Pinus sp., Populus sp., and Salix sp.; While croplands are found in the northwest of the study area, with cultivation primarily of highland barley (Hordeum vulgare var. nudum) and rapeseed (Brassica campestris) (Zhou et al., 1986; Hou, 2001; Wang et al., 2018; Wei et al., 2021).
Fieldwork
Totally 84 topsoil samples were collected from the study area, including 13 samples from mountain forests, 8 samples from alpine shrub, 29 samples from alpine meadow, 23 samples from alpine steppe, and 11 samples from croplands. The sampling sites were selected using a preferential design based on degree of human disturbance and current use. Vegetation communities and major plant taxa were recorded in the field using quadrats of 1 × 1 m for alpine meadow, alpine steppe, and cropland sites, 5 × 5 m for alpine shrub, and 10 × 10 m for forest sites. At each topsoil sampling plot, at least five sub-samples of topsoil (0–1 cm depth, surface soil with mosses) were randomly collected and homogenized into one sample, the location and elevation of each sampling site was determined by GPS. The distances between each sampling site and the nearest cropland have determined on the Google Earth. Furthermore, quantitative assessment of browsing rate, dung coverage, trampling pressure and pika burrows have also carried out in the field. We used the method of Wei et al. (2018) classified the abovementioned variables and allocated the weights to each variable, and then calculated the Grazing Influence Index (GII) and Cultivating Influence Index (CII) for each quadrate. The GII was defined as follows: GII = B × WB + D × WD + T × WT + H × WH. Where B is the herbivore plant browsing rate at each sampling site, D is the dung coverage of each sampling site, T is trampling pressure and H is plateau pika burrow density. W is the weight assigned to each variable. The CII values for individual pollen sampling sites were derived from the distance between the sampling site and closest croplands (Table 1). Detailed sampling site, vegetation community and human disturbance information were provided in Supplementary Table 1.

Table 1. Variables included in the cultivation influence index (CII) and grazing influence index (GII) for sampling sites in the study.
Laboratory Work
A standard pollen extract procedure was used in this study (Fægri and Iversen, 1989; Moore et al., 1991). 2 g dry subsamples are weighted from each sample, calcareous minerals and humic components were dissolved with 10% HCl and 10% NaOH, respectively. Samples were then sieved through a 200 μm mesh screen and treated with 40% HF to digest fine silica. Finally, samples were sieved with 7 μm meshes in an ultrasonic bath to remove clay-sized particles, and mounted in glycerin jelly. Before chemical treatment, a known number of exotic Lycopodium spores (27,637 ± 563 grains/sample) were added to each sample to enable estimation of pollen concentrations. Pollen types were identified under an optical microscope at a magnification of 400×. Pollen identifications were based on published palynological literature (Wang et al., 1997, 2018; Tang et al., 2016) and reference collections from the study area. Approximately 400 terrestrial pollen grains were counted per sample. Pollen diagrams were constructed using Tilia and Tilia-Graph software (Grimm, 2011).
Numerical Analysis
Ordination techniques were used to examine the relationships among pollen assemblages, as well as that between pollen assemblages and potential environmental factors. A detrended correspondence analysis (DCA) was initially performed to estimate the underlying linearity of the surface pollen data. The gradient lengths of the first four axes were less than 1.9 SDs, suggesting that the underlying responses were linear between surface pollen sample and pollen types. Therefore, we use the principal component analysis (PCA) to analyze the differences between the samples from different communities, and the correlation between different pollen types. GII and CII values were selected as supplementary variables to determine the effects of anthropogenic activities on the surface pollen assemblage. In this study, pollen types in which the percentage of that taxon exceeded 2% in at least one sample were selected in the numerical analysis, and 29 pollen types of 84 pollen samples were selected for PCA analysis. PCA was performed using the Canoco 5.0 software package (Ter Braak and Smilauer, 2002).
Results
Pollen Assemblages From Different Vegetation Communities
A total of 45 pollen types were identified from 84 samples in this study, the main pollen types are shown in the Figure 2. According to the characteristics of vegetation communities, the topsoil pollen samples were divided into five zones, and the content ranges and average values of main pollen types are provided in Table 2. The pollen assemblage characteristics of each community are as follows:
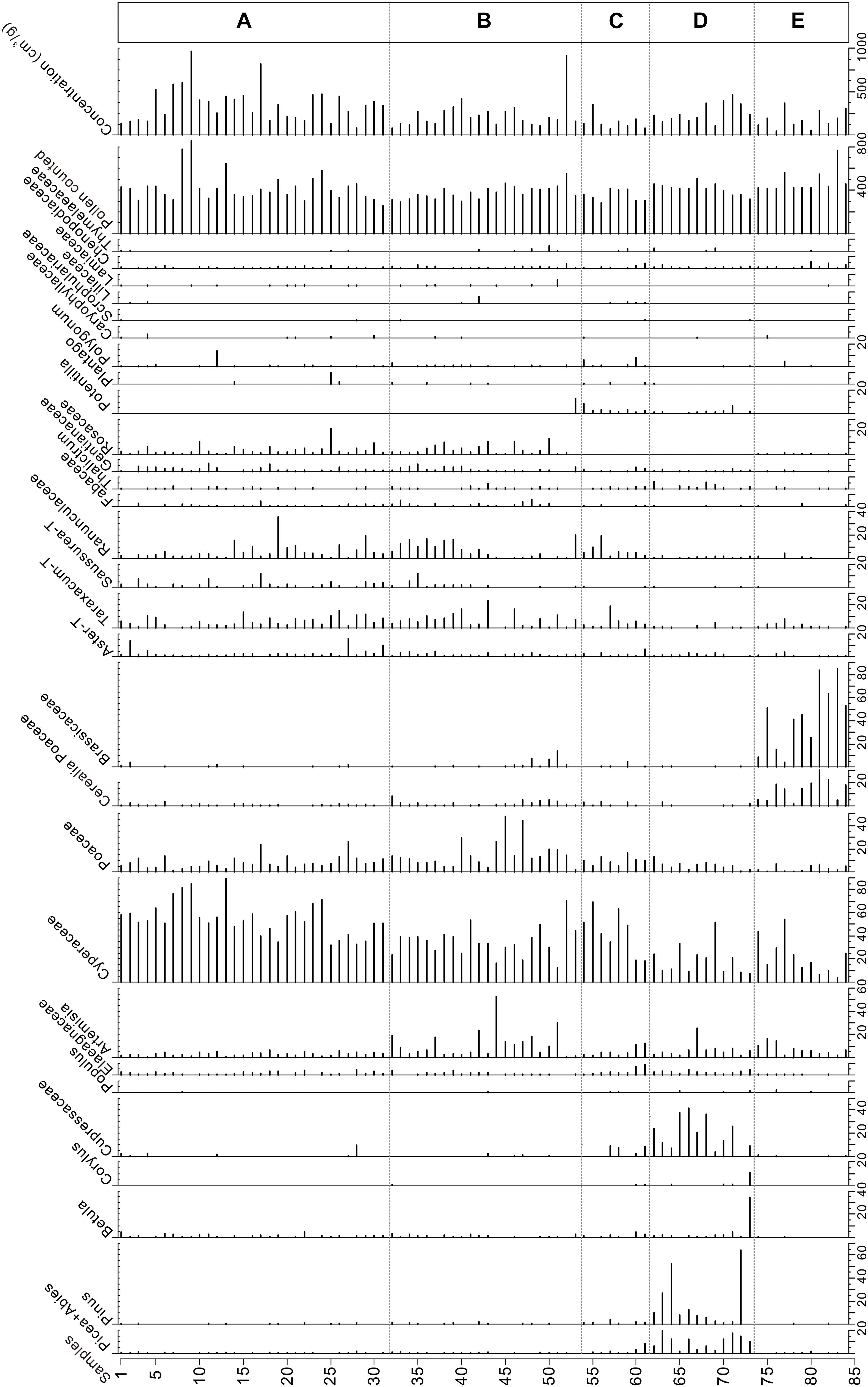
Figure 2. Modern pollen assemblages in surface samples obtained from five vegetation communities in eastern Qinghai-Tibetan Plateau.
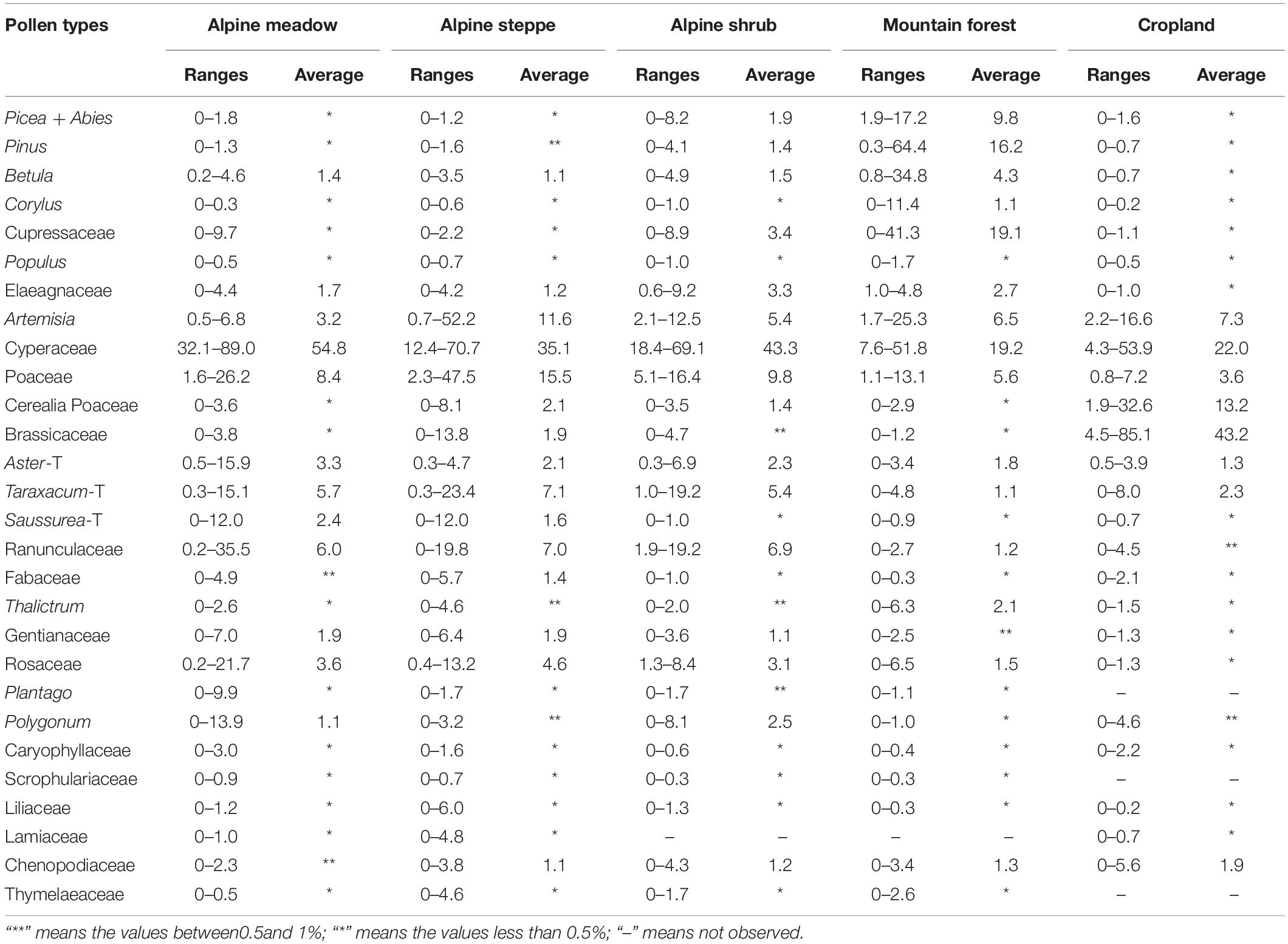
Table 2. Ranges and average percentages (%) of the main pollen types on different vegetational communities in typical agro-pastoral ecotone in the eastern Tibetan Plateau.
In alpine meadow, Cyperaceae is the predominant taxon, while other important pollen types are Poaceae, Ranunculaceae, Taraxacum-T, Artemisia, Aster-T, Saussurea-T, Gentianaceae, Rosaceae, Elaeagnaceae, and Polygonum. Cyperaceae, Poaceae, and Artemisia are dominant pollen types in the alpine steppe. While other pollen types are Taraxacum-T, Ranunculaceae, Rosaceae, Aster-T, Saussurea-T, Gentianaceae, Cerealia Poaceae, Brassicaceae, Elaeagnaceae, Fabaceae, and Chenopodiaceae. In alpine shrub, the Cyperaceae remains the most abundant pollen type, while other common pollen types, including Ranunculaceae and Poaceae, Artemisia, Taraxacum-T, Polygonum and Rosaceae. The content of arboreal pollen increased moderately in this zone and accounted for 6.6% of the total.
In mountain forest, topsoil samples are collected from different vegetation communities, which include 11 samples from Sabina chinensis and Juniperus komarovii dominated community, one sample from Pinus and Picea mixed needle forest, and Betula, Picea, Populus and Cupressaceae mixed forest, respectively. Among these vegetation communities, in Cupressaceae dominated forest sites the dominant pollen are Cupressaceae, Pinus and Picea, with average content of 20.8, 14.0, and 7.8%, respectively. The percentage of Pinus pollen is up to 64.2% in pine and spruce mixed site, and the content of Picea pollen is 14.6%. In Betula, Picea, Populus and Cupressaceae mixed site the content of Betula pollen is up to 33.9%. Other herb and shrub pollen types are mainly include Cyperaceae, Artemisia, Poaceae, Elaeagnaceae, and Ericacea.
In croplands, the pollen assemblages are dominated by Brassicaceae and Cerealia Poaceae. Other pollen types commonly found in croplands are Cyperaceae, Poaceae, Artemisia, Taraxacum-T, Chenopodiaceae, and Aster-T. Arboreal pollen types appeared sporadically with the percentages less than 1%.
Numerical Analysis
The PCA ordination for the 29 pollen types from 84 samples show that the eigenvalues of the first four axes are 0.311, 0.222, 0.099 and 0.07, respectively, which explained 70.2% of the total variation of pollen assemblages. The cumulative percentage of variation in the dataset explained by the first and second axes is 31.1 and 53.3%, respectively. Pseudo-canonical correlation coefficients of the supplementary variables of CII and GII with the first and second axes are 0.81 and 0.72, respectively.
Cropland and forest samples are clearly differentiated from other sampling sites in the PCA plot (Figure 3). The cropland samples are grouped in the fourth quadrant, while forest samples are scattered in the first quadrant of the PCA plot. Alpine shrubs, alpine meadow and alpine steppe samples are mixed and scattered in the central part of the PCA plot. However, the alpine steppe samples are closer to the positive side of first axis, while alpine meadow samples are concentrated on the negative side of the first axis, and the alpine shrub samples are closer to the positive side of the second axis.
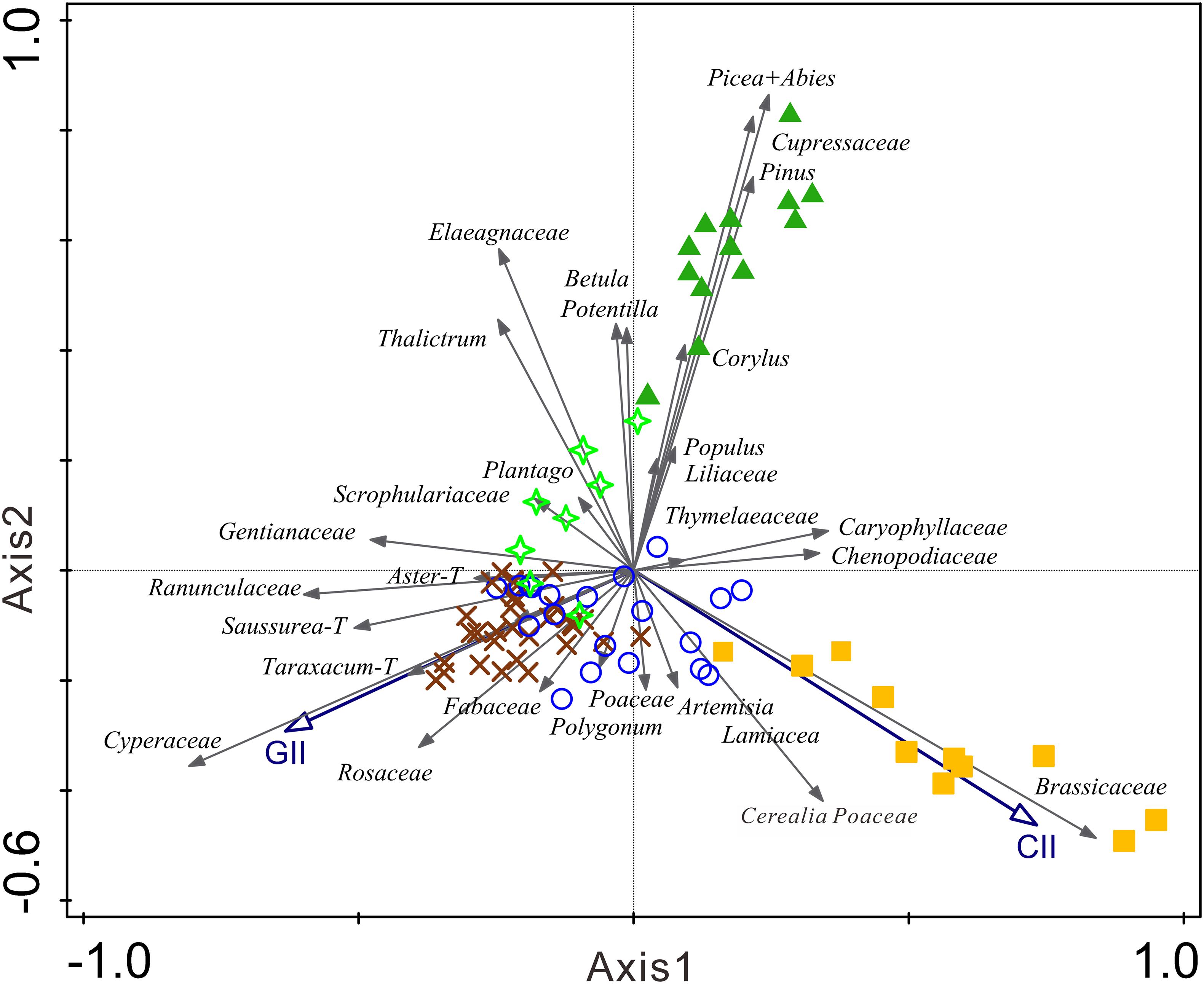
Figure 3. Principal component analysis (PCA) results for pollen assemblages of topsoil samples, CII and GII are included as supplementary variables. Green triangles are samples from mountain forest, green stars are samples from alpine shrub, yellow squares are samples from croplands, brown crosses are samples from alpine meadow, and blue circles are samples from alpine steppe.
Tree pollen types including Picea, Abies, Cupressaceae, Corylus, Betula, and Populus are significantly correlated to the forest samples. Brassicaceae and Cerealia Poaceae are significantly correlated to the cropland samples. Potentilla, Elaeagnaceae, Scrophulariaceae, Thalictrum and Plantago are spread in the second quadrant of the PCA plot, and correlated to the alpine shrub samples. Cyperaceae, Taraxacum-T, Aster-T, Saussurea-T, Ranunculaceae, Fabaceae, Polygonum and Gentianaceae are grouped on the negative side of the plot and are correlated with the alpine meadow samples, whereas Artemisia and Poaceae are correlated well with the alpine steppe samples. The PCA results revealed that CII was strongly correlated with Brassicaceae and Cerealia Poaceae pollen. Meanwhile, GII was significantly correlated with Rosaceae, Ranunculaceae, Fabaceae, Taraxacum-T, Aster-T, and Saussurea-T pollen.
Discussion
Representation of Vegetation Communities by Pollen
In the forest sites, the arboreal pollen averagely account for 57.6% of the total. Among these forest communities, the Cupressaceae pollen is the dominant taxon with average content of 20.8% in the cypress forest, while the percentage of Pinus pollen is up to 64.2% in pine and spruce mixed sites, and the percentage of Betula pollen is up to 33.9% in birch mixed forest sites. Previous study of surface pollen from Qilian Mountains in northeastern QTP indicates that the percentage of Cupressaceae pollen in cypress forest is 11.7% (Wei et al., 2011), and its ranges between 2 and 7% in conifer woodland of Lhasa Valley (Zhang et al., 2018). Thus, Cupressaceae pollen is under representative to vegetation, due to its low pollen productivity, thin pollen wall and easy to be corroded. Conversely, Pinus pollen is exhibiting over representative to vegetation, previous studies indicated that the content of Pinus pollen is higher than 40% in most of pine or pine mixed forest in north China (Li et al., 2005; Xu et al., 2007). Additionally, Betula pollen shows a moderate representative to vegetation. Surface pollen from Qilian Mountains (Wei et al., 2011) and northern China (Li et al., 2005, 2009; Geng et al., 2019) indicated that the content of Betula pollen is up to 60% in birch forest and higher than 40% birch mixed forest, which is consistent with this study.
Cyperaceae is the predominant taxon in the alpine meadow sites, with the content higher than 50% in most of sampling sites. In the alpine shrub sites, the content of Cyperaceae pollen is as high as 42%, while the content of Potentilla pollen is up to 8.4% in alpine shrub sites, and shows under representative to its vegetation. In this study, alpine shrub samples are mainly collected from Dasiphora fruticosa community, which are embedded in alpine meadow vegetation community. These vegetational distribution patterns resulted in a higher content of Cyperaceae pollen in the surface samples of Dasiphora fruticosa dominated alpine shrub. Numerous surface pollen studies from QTP have also indicated that Cyperaceae pollen is the predominant pollen taxon in the alpine meadow (Herzschuh, 2007; Lu et al., 2011; Wei et al., 2011, 2018; Zhang et al., 2017; Lu, 2018; Li et al., 2019). In alpine steppe sites, pollen assemblages are dominated by Cyperaceae (average 35%), Poaceae (average 15.5%), and Artemisia (average 10%). This is obviously different from surface pollen assemblages from the northeastern QTP, which reported that the content of Cyperaceae pollen in alpine meadow steppe and temperate steppe is lower than 30%, and Artemisia higher than 30% on the whole (Zhao and Herzschuh, 2009; Wei et al., 2011, 2018). The main reason for this difference is that the present study area is wetter than the northeastern QTP, which imply that the Artemisia/Cyperaceae ratio can not only distinguish alpine meadow from the temperate steppe vegetation, but also is an effective indicator for moisture estimation in the alpine meadow-steppe regions of the QTP. Higher values of Artemisia/Cyperaceae ratio correspond to lower effective moisture, and vice versa.
The dominant taxa represented by the pollen in cropland are Brassicaceae and Cerealia Poaceae, the average value of Brassicaceae pollen in rapeseed farmland is 67.3%, and it also shows a high percentage (average 34.3%) in samples collected from highland barley farmlands, mainly because most farmlands in the northeastern QTP have adopted a highland barley and rape rotation tillage system. On the contrary, Cerealia Poaceae (Hordeum-type) generally shows lower percentages (average 14.9%) in samples from highland barley farmlands. Similar results were obtained from northern slopes of Qilian Mountain (Huang et al., 2011, 2018) and Qinghai Lake basin (Wei et al., 2018) in the northeastern QTP. In conclusion, surface pollen assemblages generally represent the composition and dominant taxa of modern vegetation in the eastern QTP.
The Significance of Pollen Assemblage in Topsoil to Human Activities
The Influence of Grazing Activity on the Topsoil Pollen Assemblages
Grazing is an important factor affecting the vegetation community. Livestock have significant impact on the structure of vegetation community and the cycle of soil nutrients through selective feeding, trampling and fecal excretion (Marcelo et al., 2000; Li et al., 2002; Su et al., 2005; Diaz et al., 2007; Du et al., 2008; Wang et al., 2018; Zhang et al., 2018; Gao et al., 2020).
PCA result indicates that Rosaceae, Asteraceae, Fabaceae and Ranunculaceae pollen have a strong relationship with GII. The vegetation survey in the QTP showed that Kobresia spp. is the dominant species and Potentilla spp. is the subdominant species under light and moderate grazing conditions, but Potentilla spp. became dominant species with the increase of grazing intensity (Dong et al., 2004). Our field investigation has also found that the plant species of Potentilla anserina, Ligularia spp., Leontopodium, Oxytropis spp., and Ranunculus spp. are significantly increased in the severe grazed sites. Thus, the disturbances of grazing activity on the regional vegetation are well reflected in the topsoil pollen assemblages in the eastern QTP. In this study, Cyperaceae is the dominant component in topsoil pollen assemblages in some overgrazed sampling sites from alpine meadow and alpine steppe, while the other pollen types, including Rosaceae, Asteraceae, Fabaceae and Ranunculaceae, increased significantly in these sampling sites. Similar conclusions have also obtained from surface pollen study in Qinghai Lake basin, which suggesting the pollen of Rosaceae and Asteraceae are the most important grazing-indicator (Wei et al., 2018). Nonetheless, the response of vegetational degradation to overgrazing is also various in different communities and vegetation zones, which reflected in the surface pollen assemblages. For example, our results indicate that severe pastoral activity causes the increase of Asteraceae, Rosaceae, Ranunculaceae and Fabaceae in surface pollen assemblages of alpine meadow and alpine steppe in eastern QTP, while in temperate steppe of northeastern QTP and northern China, overgrazing led to the significant increase of Artemisia, Chenopodiaceae, Stellera and Plantago pollen (Liu et al., 2008; Huang et al., 2017; Wei et al., 2018). Additionally, some human-related pollen types, such as Artemisia and Chenopodiaceae, in sedimentary sequences have dual denotative meaning (Liu et al., 2006; Wei et al., 2018), which suggesting the increase of Artemisia and Chenopodiaceae may indicate climate change that resulted in vegetational degradations or intensifications of the grazing activity in temperate steppe region. Therefore, when using human-related fossil pollen types to reconstruct the history of grazing activity, the location and overall vegetational background of the study area should be considered, and more importantly, comprehensively applications of a spectrum of grazing-related pollen types, charcoal and coprophilous fungal spores are needed.
Pollen Indicators of Highlands Cultivation
PCA result indicates that Brassicaceae and Cerealia Poaceae are significantly correlated with CII. In this study, the average value of Brassicaceae pollen is 67.3% in rapeseed farmland and Cerealia Poaceae pollen is averagely 14.9% in highland barley farmland, respectively, and they are swiftly drops to below 3% in non-cultivating sampling sites. Surface pollen analysis from croplands in northeastern margin of the QTP (Ma et al., 2009; Huang et al., 2011, 2018; Wei et al., 2018), as well as in central China (Wang et al., 2010; Li et al., 2019) shows similar characteristics. These distribution patterns suggesting that the aerial dispersed distances of Brassicaceae and Cerealia Poaceae pollen are very short and the content of these pollen types can be used as an important assessment method for highlands cultivation on the eastern QTP. Additionally, our previous study found that Cerealia Poaceae pollen content is relatively higher (averagely 5.1%) in domestic herbivores dung samples in this region (Duan et al., 2021), which imply herbivores are important in the dispersion of Cerealia Poaceae pollen into pastures. In this study, we have measured the morphological characteristics of these two pollen types. Most of the pollen grains of Brassica campestris-type are range between 21 and 31 μm in polar axis length, and 13–28 μm in equator axis length, with oblateness range between 0.5 and 1.0 (Figure 4A). While the diameter of Hordeum-type pollen grains are ranges between 37–48 μm in size, with pore diameter of > 2.6 μm, and ratio of pore diameter to pollen grain size ranges between 0.06 and 0.12 (Figure 4B).
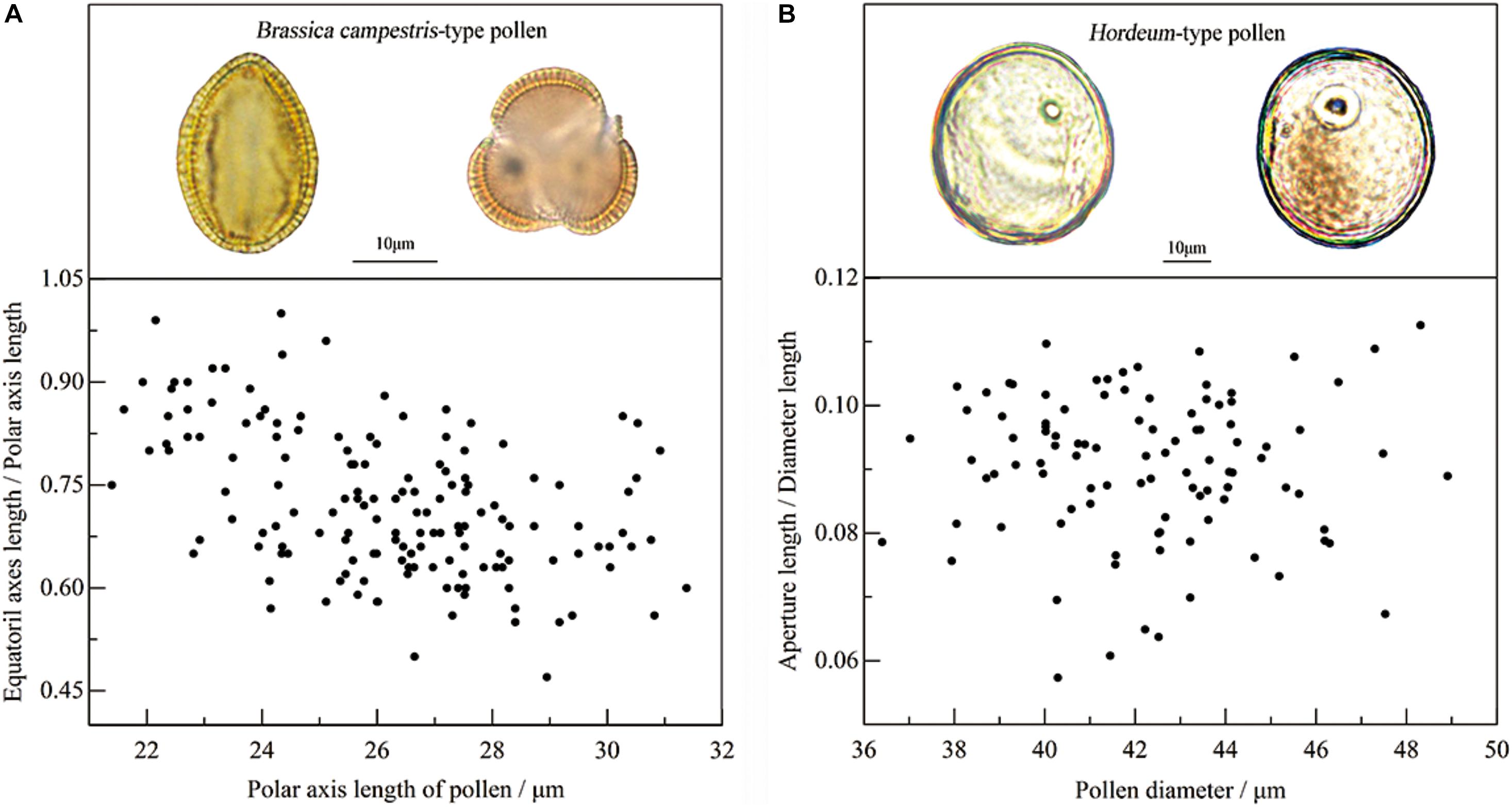
Figure 4. Light microscopic images and morphological characteristics of Brassica campestris-type and Hordeum-type pollen. (A) Brassica campestris-type pollen; (B) Hordeum-type pollen.
Conclusion
(1) Topsoil pollen assemblages could distinguish different vegetation communities in the typical agro-pastoral ecotone of the eastern Tibetan Plateau. Brassicaceae and Cerealia Poaceae pollen are the dominant types in croplands, while Cyperaceae is the most important taxon in alpine shrub and alpine meadow. Cyperaceae, Poaceae, and Artemisia are the dominant pollen types in alpine steppe, and topsoil pollen assemblage of alpine steppe in eastern QTP shows higher content of Cyperaceae and lower Artemisia compared with similar vegetation community in the northeastern QTP. The Artemisia/Cyperaceae ratio is an effective indicator of moisture estimation in the alpine meadow-steppe regions of the QTP. Arboreal pollen, including Cupressaceae, Pinus, Picea, and Betula, are important types in the mountain forest in the eastern QTP. Surface pollen assemblages can be used to distinguish the vegetation communities and the dominant types in the eastern QTP.
(2) Rosaceae, Asteraceae, Fabaceae and Ranunculaceae pollen are clearly associated with grazing, and these pollen types are effective indicators of grazing activity in the eastern QTP. Brassicaceae and Cerealia Poaceae pollen are strongly related to cultivation, which is detectable on the basis of the content of Brassicaceae and Cerealia Poaceae in pollen assemblages and related morphological characteristics.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
RD and HW finished pollen analysis and prepared manuscript. GH, JG, YD, and ZQ involved field work. All authors contributed to the article and approved the submitted version.
Funding
This work was financially supported by the National Natural Science Foundation of China (Grant #41877455), the Natural Science Foundation of Qinghai Province (Grant #2017-ZJ-931Q), and the Second Tibetan Plateau Scientific Expedition and Research Program (STEP) (Grant #2019QZKK0805).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.685942/full#supplementary-material
Supplementary Table 1 | Sites information and vegetation surveys at 84 surface pollen sites in the eastern Qinghai-Tibetan Plateau.
References
Chen, F. H., Chen, S. Q., Zhang, X., Chen, J. H., Wang, X., Gowanet, E. J., et al. (2020). Asian dust-storm activity dominated by Chinese dynasty changes since 2000 BP. Nat. Commun. 11:992. doi: 10.1038/s41467-020-14765-4
Chen, F. H., Dong, G. H., Zhang, D. J., Liu, X. Y., Jia, X., An, C. B., et al. (2015). Agriculture facilitated permanent human occupation of the Tibetan Plateau after 3600 B.P. Science 347, 248–250. doi: 10.1126/science.1259172
Chen, F. H., Fu, B. J., Xia, J., Wu, D., Wu, S., Zhang, Y. L., et al. (2019). Major advances in studies of the physical geography and living environment of China during the past 70 years and future prospects. Sci. Chin. Earth Sci. 62, 1665–1701. doi: 10.1007/s11430-019-9522-7
Diaz, S., Lavorel, S., McIntyre, S. U. E., Falczuk, V., Casanoves, F., Milchunas, D. G., et al. (2007). Plant trait responses to grazing-a global synthesis. Glob. Chang. Biolo. 13, 313–341. doi: 10.1111/j.1365-2486.2006.01288.x
Dong, G. H., Jia, X., Elston, R., Chen, F. H., Li, S. C., Wang, L., et al. (2013). Spatial and temporal variety of prehistoric human settlement and its influencing factors in the upper yellow river valley, qinghai province, china. J. Archaeol. Sci. 40, 2538–2546. doi: 10.1016/j.jas.2012.10.002
Dong, Q. M., Li, Q. Y., Ma, Y. S., Shi, J. J., and Wang, Q. J. (2004). Effects of yaks stocking pates on aboveground and belowground blomass in kobrecia parva alpine meadow. Pratacultural Sci. 21, 48–53.
Dong, Q. M., Zhao, X. Q., and Ma, Y. S. (2007). Effects of grazing intensity and time on forage nutrition contents in alpine mixed-sown grassland. Chin. J. Grassl. 29, 67–73.
Du, Y. G., Liang, D. Y., Cao, G. M., Wang, Q. L., and Wang, C. T. (2008). Effect on incrassation of mattic epipedon and pasture nutrient-moisture using by grazing in alpine Kobresia meadow. Acta Prataculturae Sin. 17, 146–150.
Duan, R. L., Hou, G. L., Wei, H. C., Xu, Q. H., and Gao, J. Y. (2021). The characteristics and environmental implications of dung pollen assemblages of grazing livestock in alpine meadow in the eastern Qinghai-Tibetan Plateau. Arid Land Geo. 44, 229–239.
Fægri, K., and Iversen, J. (1989). Textbook Of Pollen Analysis, Fourth Ed. Chichester: John Wiley and Sons.
Gao, J. Y., Hou, G. L., Wei, H. C., Chen, Y. C., E, C. Y., Chen, X. L., et al. (2020). Prehistoric human activity and its environmental background in Lake Donggi Cona basin, northeastern Tibetan Plateau. Holocene 30, 657–671. doi: 10.1177/0959683619895583
Geng, R. W., Zhao, Y., Cui, Q. Y., and Qin, F. (2019). Representation of modern pollen assemblages with respect to vegetation and climate in Northeast China. Quat. Int. 532, 126–137. doi: 10.1016/j.quaint.2019.11.003
Grimm, E. C. (2011). TILIA Software Version 1.7.16. Illinois State Museum. Springfield, IL: Research and Collection Center. Available online at: https://www.tiliait.com/
Guedes, J. A. D. A., Lu, H., and Hein, A. M. (2015). Early evidence for the use of wheat and barley as staple crops on the margins of the Tibetan Plateau. Pro. Natl. Acad. Sci. U. S. A. 112, 5625–5630. doi: 10.1073/pnas.1423708112
Herzschuh, U. (2007). Reliability of pollen ratios for environmental reconstructions on the Tibetan Plateau. J. Biogeogr. 34, 1265–1273. doi: 10.1111/j.1365-2699.2006.01680.x
Hou, G. L., Gao, J. Y., Chen, Y. C., Xu, C. J., Lancuo, Z. M., Xiao, Y. M., et al. (2020). Winter-to-summer seasonal migration of microlithic human activities on the Qinghai-Tibet Plateau. Sci. Rep. 10:18928. doi: 10.1038/s41598-020-75952-3
Huang, X. Z., Chen, X. M., Chen, C. Z., Ma, Y. L., and Zhou, G. (2011). Surface pollen assemblage characters of farmland from different altitudes in upper and middle reaches of the Heihe River, arid north-western China. J. Lanzhou Univ. 47, 14–23. doi: 10.13885/j.issn.0455-2059.2011.02.022
Huang, X. Z., Chen, X. M., and Du, X. (2018). Modern pollen assemblages from human-influenced vegetation in northwestern China and their relationship with vegetation and climate. Veget. Hist. Archaeobot. 27, 767–780. doi: 10.1007/s00334-018-0672-0
Huang, X. Z., Liu, S. S., Dong, G. H., Qiang, M. R., Bai, Z. J., Zhao, Y., et al. (2017). Early human impacts on vegetation on the northeastern Qinghai-Tibetan Plateau during the middle to late Holocene. Prog. Phys. Geogr. 41, 286–301. doi: 10.1177/0309133317703035
Li, J. H., Li, Z. Q., and Ren, J. Z. (2002). The effects of grazing on grassland plants. Acta Prataculturae Sin. 11, 4–11.
Li, Y. C., Xu, Q. H., Zhang, L. Y., Wang, X. L., Cao, X. Y., and Yang, X. L. (2009). Relationship between modern pollen and vegetation and climate in forest communities in northern China. J. Geogra. Sci. 19, 643–659.
Li, Y. C., Xu, Q. H., Zhao, Y. K., Yang, X. L., Xiao, J. L., Chen, H., et al. (2005). Pollen indication to source plants in the eastern desert of China. Chin. Sci. Bull. 50, 1632–1641. doi: 10.1360/04wd0170
Li, Y. F., Li, C. H., Xu, B., Shu, J. W., Li, Y. C., and Xu, Q. H. (2019). Modern pollen assemblages from human-influenced vegetation in Eastern China and their spatial changes. Adv. Earth Sci. 34, 333–345. doi: 10.11867/j.issn.1001-8166.2019.04.0333
Liu, H. Y., Wang, Y., and Tian, Y. H. (2006). Climatic and anthropogenic control of surface pollen assemblages in East Asian steppes. Rev. Palaeobot. Palyno. 138, 281–289. doi: 10.1016/j.revpalbo.2006.01.008
Liu, H. Y., Wei, F. L., Kan, L., Zhu, J. L., and Wang, H. Y. (2008). Determinants of pollen dispersal in the east asian steppe at different spatial scales. Rev. Palaeobot. Palyno. 149, 219–228. doi: 10.1016/j.revpalbo.2007.12.008
Lu, H. Y., Wu, N. Q., Liu, K. B., Zhu, L., Yang, X. D., Yao, T. D., et al. (2011). Modern pollen distributions in Qinghai-Tibetan Plateau and the development of transfer functions for reconstructing Holocene environmental changes. Quat. Sci. Rev. 30, 947–966. doi: 10.1016/j.quascirev.2011.01.008
Lu, J. Y. (2018). Study on Relevant Source Area of Pollen and Relative Pollen Productivity Estimates of the Alpine Meadow in Northeastern Tibetan Plateau. Ph.D. thesis. Hebei: Hebei Normal University.
Ma, T., Rolett, B. V., Zheng, Z., and Zong, Y. Q. (2020). Holocene coastal evolution preceded the expansion of paddy field rice farming. Proc. Natl. Acad. Sci. U. S. A. 117, 24138–24143. doi: 10.1073/pnas.1919217117
Ma, Y. L., Xu, Q. H., Huang, X. Z., Zhou, G., Zhang, L. Y., Tao, S. C., et al. (2009). Pollen assemblage characters of human disturbed vegetation in arid area in northwestern China. J. Palaeogeog. 11, 542–550.
Madsen, D. B., Ma, H. Z., Brantingham, P. J., Gao, X., Rhode, D., Zhang, H. Y., et al. (2006). The Late Upper Paleolithic occupation of the northern Tibetan Plateau margin. J. Archaeol. Sci. 33, 1433–1444. doi: 10.1016/j.jas.2006.01.017
Marcelo, S., Mario, G., Avi, P., Eugene, D. U., and Jaime, K. (2000). Vegetation Response to Grazing Management in a Mediterranean Herbaceous Community: a Functional Group Approach. J. Appl. Ecol. 37, 224–237. doi: 10.2307/2655905
Meyer, M. C., Aldenderfer, M. S., Wang, Z., Hoffmann, D. L., Dahl, J. A., Degering, D., et al. (2017). Permanent human occupation of the central Tibetan Plateau in the early Holocene. Science 355, 64–67. doi: 10.1126/science.aag0357
Miao, Y. F., Jin, H. L., Liu, B., Herrmann, M., Sun, Z., and Wang, Y. P. (2015). Holocene climate change on the northeastern Tibetan Plateau inferred from mountain-slope pollen and non-pollen palynomorphs. Rev. Palaeobot. Palyno. 221, 22–31. doi: 10.1016/j.revpalbo.2015.05.006
Miehe, G., Miehe, S., Böhner, J., Kaiser, K., Hensen, I., Madsen, D. B., et al. (2014). How old is the human footprint in the world’s largest alpine ecosystem? A review of multiproxy records from the Tibetan Plateau from the ecologists’ viewpoint. Quat. Sci. Rev. 86, 190–209. doi: 10.1016/j.quascirev.2013.12.004
Miehe, G., Miehe, S., Kaiser, K., Reudenbach, C., Behrendes, L., Duo, L., et al. (2009). How old is pastoralism in Tibet? An ecological approach to the making of a Tibetan landscape. Palaeogeogr. Palaeoclimatol. Palaeoecol. 276, 130–147. doi: 10.1016/j.palaeo.2009.03.005
Moore, P. D., Webb, J. A., and Collinson, M. E. (1991). Pollen Analysis, Second Ed. London: Blackwell Scientific.
Schlütz, F., and Lehmkuhl, F. (2009). Holocene climatic change and the nomadic Anthropocene in Eastern Tibet: palynological and geomorphological results from the Nianbaoyeze Mountains. Quat. Sci. Rev. 28, 1449–1471. doi: 10.1016/j.quascirev.2009.01.009
Schwalb, A., Schütt, B., and Fang, X. (2010). Climate evolution and environmental response on the Tibetan Plateau. Quat. Int. 218, 1–202. doi: 10.1016/j.quaint.2010.02.017
Su, Y. Z., Li, Y. L., Cui, J. Y., and Zhao, W. Z. (2005). Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland. Inner Mongolia, northern China. Catena 59, 267–278. doi: 10.1016/j.catena.2004.09.001
Sun, H. L., Zheng, D., Yao, T. D., and Zhang, Y. L. (2012). Protection and construction of the national ecological security shelter zone on Tibetan Plateau. Acta Geogr. Sin. 67, 3–12.
Tang, L. Y., Mao, L. M., Shu, J. W., Li, C. H., Shen, C. M., and Zhou, Z. Z. (2016). “An illustrated handbook of quaternary pollen and spores in China,” in Chinese With English Summary, First Ed, eds Meng and Hu (Beijing: Science Press Ltd).
Ter Braak, C. J. F., and Smilauer, P. (2002). CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5). Ithaca: Microcomputer Power.
Wang, F. X., Qian, N. F., and Zhang, Y.L, and Yang, H. Q. (1997). Pollen Flora of China. Second Edition. Beijing: Science Press.
Wang, J., Xia, H., Yao, J., Shen, X., Cheng, T., Wang, Q., et al. (2020). Subsistence strategies of prehistoric hunter-gatherers on the Tibetan Plateau during the Last Deglaciation. Sci. Chin. Earth Sci. 63, 395–404. doi: 10.1007/s11430-019-9519-8
Wang, T., Zhang, S. R., Xu, Q. H., Shen, W., Sun, Y. H., and Li, Y. W. (2018). Pollen morphology of summer-flowering species of Alpine Meadows in northeastern Tibetan Plateau. Acta Palaeontol. Sin. 57, 524–542.
Wang, X. L., Li, Y. C., Xu, Q. H., Cao, X. Y., Zhang, L. Y., and Tian, F. (2010). Pollen assemblages from different agricultural units and their spatial distribution in Anyang area. Chin. Sci. Bull. 55, 544–554.
Wei, H. C., Duan, R. L., Xu, Q. H., Yang, S. X., Fan, Q. S., Hou, G. L., et al. (2021). Fungal spore indicators of vegetation and highland pastoralism in modern topsoil and dung, eastern Tibetan Plateau. Catena 202:105231.
Wei, H. C., E, C. Y., Zhang, J., Sun, Y. J., Li, Q. K., Hou, G. L., et al. (2020). Climate change and anthropogenic activities in Qinghai Lake basin over the last 8500 years derived from pollen and charcoal records in an aeolian section. Catena 193:104616. doi: 10.1016/j.catena.2020.104616
Wei, H. C., Ma, H. Z., Zheng, Z., Pan, A. D., and Huang, K. Y. (2011). Modern pollen assemblages of surface samples and their relationships to vegetation and climate in the northeastern Qinghai-Tibetan Plateau, Chin. Rev. Palaeobot. Palyno. 163, 237–246. doi: 10.1016/j.revpalbo.2010.10.011
Wei, H. C., Yuan, Q., Xu, Q. H., Qin, Z. J., Wang, L. Q., Fan, Q. S., et al. (2018). Assessing the impact of human activities on surface pollen assemblages in Qinghai Lake Basin, Chin. J. Quat. Sci. 33, 702–712. doi: 10.1002/jqs.3046
Xu, Q. H., Li, Y. C., Yang, X. L., and Zheng, Z. H. (2007). Quantitative relationship between pollen and vegetation in northern China. Sci. China. Ser. D. 50, 582–599. doi: 10.1007/s11430-007-2044-y
Yao, T. D., Chen, F. H., Cui, P., Ma, Y. M., Xu, B. Q., Zhu, L. P., et al. (2017). From Tibetan Plateau to Third Pole and Pan-Third Pole. Bull. Chin. Acad. Sci. 32, 924–931. doi: 10.16418/j.issn.1000-3045.2017.09.001
Yao, T. D., Thompson, L. G., Mosbrugger, V., Zhang, F., Ma, Y. M., Luo, T. X., et al. (2012). Third pole environment (TPE). Environ. Dev. 3, 52–64. doi: 10.1016/j.envdev.2012.04.002
Zhang, D. J., Xia, H., Chen, F. H., Li, B., Slon, V., Cheng, T., et al. (2020). Denisovan DNA in Late Pleistocene sediments from Baishiya Karst Cave on the Tibetan Plateau. Science 370:584–587. doi: 10.1126/science.abb6320
Zhang, R. Y., Wang, Z. W., Han, G. D., Schellenberg, M. P., Wu, Q., and Gu, C. (2018). Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the desert steppe, Northern China. Agricu. Ecosyst. Environ. 265, 73–83. doi: 10.1016/j.agee.2018.05.014
Zhang, Y., Kong, Z., Yang, Z., Wang, L., and Duan, X. (2017). Surface pollen distribution fromalpine vegetation in eastern Tibet, China. Sci. Rep. 7:586. doi: 10.1038/s41598-017-00625-7
Zhao, Y., and Herzschuh, U. (2009). Modern pollen representation of source vegetation in the Qaidam Basin and surrounding mountains, northeastern Tibetan Plateau. Veg. Hist. Archaeobot. 18, 245–260. doi: 10.1007/s00334-008-0201-7
Keywords: modern pollen, anthropogenic activity, pastoral indicator, Cerealia Poaceae pollen, Qinghai-Tibetan Plateau
Citation: Duan R, Wei H, Hou G, Gao J, Du Y and Qin Z (2021) Modern Pollen Assemblages in Typical Agro-Pastoral Ecotone in the Eastern Tibetan Plateau and Its Implications for Anthropogenic Activities. Front. Ecol. Evol. 9:685942. doi: 10.3389/fevo.2021.685942
Received: 02 April 2021; Accepted: 09 June 2021;
Published: 09 July 2021.
Edited by:
Joel Guiot, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Zhigang Zhao, Lanzhou University, ChinaWenying Jiang, Institute of Geology and Geophysics, Chinese Academy of Sciences (CAS), China
Copyright © 2021 Duan, Wei, Hou, Gao, Du and Qin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Haicheng Wei, aGN3ZWlAaXNsLmFjLmNu