 Mokhtar Abdulsattar Arif1,2†
Mokhtar Abdulsattar Arif1,2† Tuğcan Alınç1†
Tuğcan Alınç1† Salvatore Guarino3*
Salvatore Guarino3* Stefano Colazza1
Stefano Colazza1 Antonino Cusumano1
Antonino Cusumano1 Ezio Peri1
Ezio Peri1- 1Department of Agricultural, Food and Forest Sciences (SAAF), University of Palermo, Palermo, Italy
- 2Plant Protection Directorate, Ministry of Agriculture, Baghdad, Iraq
- 3Institute of Biosciences and Bioresources (IBBR), National Research Council of Italy (CNR), Palermo, Italy
Egg parasitoids foraging for suitable hosts scattered in the environment rely mainly on chemical cues. Elucidating the chemical ecology of natural enemies is important in the development of effective and successful strategies for conservation biological control. In this context, the host cuticular hydrocarbons, which are exploited by several species of egg parasitoids as contact kairomones, could be used to retain them by providing information about the presence and the sex of adults of the target species: sex is important because only females of the host species lay the eggs that can be subsequently utilized for parasitoid reproduction. However, the chemical basis of host sex discrimination in egg parasitoids is not well understood. We carried out behavioral and chemical bioassays to investigate the role played by contact chemical cues left by adults of the brown marmorated stink bug, Halyomorpha halys Stål, in host egg searching behavior and adult host sex discrimination by the egg parasitoid Trissolcus japonicus (Ashmead). A first set of bioassays showed that parasitoids spent more time exploring patches contaminated with chemicals associated with adult H. halys females compared with adult males. Similar responses were displayed by T. japonicus when hexane extracts of H. halys were tested suggesting that non-polar chemical compounds are involved in host sex discrimination. GC-MS analysis of hexane extracts revealed quantitative differences in the cuticular compounds of the two sexes, with 1-hexadecene (more abundant in males) being the most important component in determining these differences. Hexane extracts of H. halys females blended with synthetic 1-hexadecene significantly reduced the wasps’ arrestment responses compared to crude extracts.
Introduction
In recent years, several advances have been made toward the understanding of the chemical ecology of insect parasitoid foraging behavior (Wajnberg and Colazza, 2013). While foraging for hosts, insect parasitoids can exploit volatile chemical signals (e.g., herbivore-induced plant volatiles, oviposition-induced plant volatiles, host sex pheromones) and contact chemical cues (e.g., host-by products such as wing scales, walking traces, silk, frass, honeydew) (Colazza et al., 2010; Hilker and Fatouros, 2015; Turlings and Erb, 2018; Aartsma et al., 2019; Aspin et al., 2021). These cues are particularly relevant for an applied perspective as they have often been proposed to be efficient tools to manipulate wasp behavior in crop protection. Volatile and contact cues have promising potential in conservation biological control (CBC) as they can help to attract and retain natural enemies in cropping systems and also enhance their activity (Khan et al., 2008; Rodriguez-Saona et al., 2013). Studies aimed at using semiochemicals in CBC have mainly focused on volatile cues in order to attract parasitoid wasps from the areas surrounding the focal crop (Van Poecke et al., 2001; James and Grasswitz, 2005; Conti et al., 2020). The use of contact chemical cues has received relatively little attention by biological control practitioners (Meiners and Peri, 2013; Conti et al., 2020) and yet they could, for example, be used to retain resident parasitoids within a focal crop and thus reduce dispersion into surrounding areas.
Contact semiochemicals are well-explored in egg parasitoids in which they activate “biased random search,” i.e., a non-directional movement which is characterized by ortho- and klino-kinetic reactions, allowing wasps to explore intensively the area contaminated with the chemical stimulus (Bell, 1990; Fatouros et al., 2008; Colazza et al., 2014). Examples of contact chemical cues are wing scales of butterflies exploited by trichogrammatid egg parasitoids, and walking traces left by stink bugs perceived by scelionid egg parasitoids (Colazza et al., 2010). In the latter case these contact cues can give information about both the presence of the adults of the target host species as well as their sex (Conti and Colazza, 2012). Host sex is important because only the females lay the eggs which are parasitized by the scelionids, and indeed foraging scelionids respond more strongly to, and spend more time examining, chemical cues associated with adult stink bug females compared with cues associated with males and immature nymphs (Colazza et al., 1999; Salerno et al., 2009; Peri et al., 2016).
The source of the active compound eliciting egg parasitoid searching behavior is often reported to be the host insect cuticle (Colazza et al., 2014). The cuticular layer of the insect body comprises several chemicals as lipids, fatty acids, alcohols, alkyl esters, glycerides, sterols and aldehydes (Lockey, 1985), but in several cases non-polar compounds elicit parasitoid responses (Padmavathi and Paul, 1998; Paul et al., 2002; Wang et al., 2019). For example, Colazza et al. (2007) found that n-nonadecane (n-C19), a linear cuticular hydrocarbon present in Nezara viridula (L.) (Heteroptera: Pentatomidae), mediates host gender discrimination in the egg parasitoid Trissolcus basalis (Wollaston) (Hymenoptera: Scelionidae). Other scelionids have been reported to use similar strategies (Borges et al., 2003; Conti et al., 2004; Salerno et al., 2009; Gomes Lagôa et al., 2020); however, the chemical basis of host sex discrimination has not been investigated.
Here we explore the chemical ecology of host sex discrimination in the samurai wasp, Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae), the main egg parasitoid of the brown marmorated stink bug, Halyomorpha halys Stål (Heteroptera: Pentatomidae). Halyomorpha halys is a highly polyphagous species native to Asia that has become a serious invasive agricultural and nuisance pest across North America and Europe (Rice et al., 2014; Leskey and Nielsen, 2018). Trissolcus japonicus was introduced adventively into Europe and North America but is now well established in both continents (Kaser et al., 2018; Stahl et al., 2019) and is considered to have the potential to control the stink bug pest population density to below economic thresholds (Abram et al., 2017). In this context, a better understanding of the host searching mechanisms used by T. japonicus could help to develop effective and successful CBC programs (Conti et al., 2020). The few prior studies on behavioral interactions between T. japonicus and H. halys indicate that the parasitoid spends more time on substrates contaminated with H. halys walking traces rather than on untreated substrates, suggesting the presence of a contact kairomone (Hedstrom et al., 2017; Boyle et al., 2020; Malek, 2020), but the chemical bases mediating host sex discrimination were not explored. Our objective was thus to evaluate the use of cuticular compounds of H. halys as contact chemical cues by T. japonicus during short-range searching behavior. Specifically, we analyze the chemical compounds present in the cuticula of male and female H. halys and evaluate their role in mediating host sex discrimination by T. japonicus.
Materials and Methods
Insect Rearing
The colony of H. halys was established from field-collected individuals around Palermo, Italy, and reared in cages (47.5 × 47.5 × 47.5 cm; BugDorm-44545, Mega View Science Co., Ltd. Taichung, Taiwan) under controlled conditions (24 ± 2 °C; 70 ± 5% RH; 16L:8D photoperiod). The colony was fed with seasonal fresh vegetables and sunflower seeds. Food was renewed every 2 days, and water was provided with soaked cotton wool in small containers. Paper towels were hung inside each adult cage as oviposition substrates. Egg masses were collected daily and used to maintain both stink bugs and wasp colonies. Every 2–3 days, newly emerged H. halys adults were collected and placed in fresh cages so that individuals of known age were continuously available for experiments.
The colony of T. japonicus was established from wasps emerging from H. halys sentinel egg masses located in fields around Turin, Italy, and was reared on H. halys egg masses. Wasps were maintained in 85-ml glass tubes, fed with a honey–water solution (80:20 v/v) and kept in a controlled environmental room (24 ± 2°C; 70 ± 5% RH; 16L:8D photoperiod). Halyomorpha halys egg masses were singly glued onto a strip of paper and exposed to 2 female wasps for 24 h, and, after emergence, male and female wasps were kept together to allow mating. Before the bioassays, 3–5 days old female wasps were individually isolated in a 2 mL vial with a drop of honey-water for 24 h and then transferred to the bioassay room in order to be acclimatized 1 h before the tests.
Stimulus Preparation Procedure
The following stimuli were prepared.
Freshly Killed H. halys Adults
Females in pre-ovipositional state (i.e., which displayed enlarged abdomen) and virgin males were individually isolated for 3 days and then killed at −18°C for 1 h.
Hexane Extracts of H. halys Cuticular Compounds
Freshly killed pre-ovipositional females and virgin males were dissected under a binocular stereo microscope (Leica MZ12, Leica Microsystems GmbH, Wetzlar, Germany) with fine scissors to separate legs at coxal level, wings, pronotum and scutellum. The residual internal tissue remained inside of the pronotum was thoroughly cleaned by forceps to reduce contamination. Mesonotum, metanotum, and abdomen were discarded due to possible contamination by defensive chemicals from the glandular structures in these segments. For each adult, dissected body parts were weighed, placed in 4 ml glass vials and extracted with 2 ml of hexane at 30°C for 1 h. After removal of the body parts, the resulting extract was evaporated under a gentle nitrogen stream, and re-dissolved in 100 μl hexane (>99% pure, Sigma-Aldrich, Milan, Italy). Extracts were individually stored at –18°C until used in bioassays and analyzed by GC-MS (see below). Ten extracts were carried out for pre-ovipositional females and 10 for virgin males.
Hexane Extracts of H. halys Females Blended With Synthetic 1-Hexadecene
In consideration of the results obtained from the multivariate analysis of chemical profiles (see section “Results”) the role of synthetic 1-hexadecene (>99% pure, Sigma-Aldrich, Milan, Italy) was further investigated in the host gender discrimination displayed by T. japonicus females. Dissected body parts of 6 pre-ovipositional females were split into two groups, and each group was extracted with 6 ml of hexane at 30°C for 1 h. After removal of the body parts, the resulting extracts were evaporated under a gentle nitrogen stream. One group was re-dissolved in 300 μl of a hexane solution of 1-hexadecene at 0.66 ppm (test), while the other group was re-dissolved in 300 μl of hexane (control). The 1-hexadecene amount was determined by the quantitative differences observed in the cuticular extracts of individual H. halys males and females (see section “Chemical Analyses”). Extracts were individually stored at −18°C until assayed.
Open Arena Bioassay
Bioassays were conducted in an open arena consisting of a square sheet of filter paper Whatman No. 1 (25 × 25 cm; wasp/arena ratio: 0.003%). In the center of the arena, a circular area (6 cm diameter; 28.26 cm2, about 4.5% of the entire arena; wasp/arena ratio: 0.07%) was contaminated with stimuli described below. One wasp female was gently released in the middle of the circular area and tracked by a CCD video camera (Sony SSC M370 CE) fitted with a 12.5–75 mm F-1 1.8 zoom lens. The arena was illuminated from above by two 22-W cool white fluorescent tubes (Full spectrum 5,900 K, 11 W; Lival, Italy). The analogic signal from the camera was digitalized by a video grabber, and analyzed with XBug, a video tracking and motion analysis software (Colazza et al., 2007). Data recording was defined as the time between the wasp was gently released on the patch until had left the arena either by walking or by flying off. The time spent on the entire arena (25 × 25 cm) was scored as the residence time and used for statistical analyses. This value was used, rather than only the time spent on the treated area, because more representative of the active searching behavior elicited by the kairomonal stimuli (Colazza et al., 2007).
For each experiment, the following bioassay procedure was used.
Freshly Killed H. halys Adults
The dorsal part of a female or male bug (pronotum, scutellum and wings) was gently rubbed with the use of featherweight forceps (BioQuip©) in the circular area of the arena continuously for about 5 min in order to contaminate the area homogenously with cuticular compounds. A total of 30 wasp females was assayed for each treatment.
Hexane Extracts of H. halys Cuticular Compounds
Hexane extracts of H. halys male and female were tested by pipetting 100 μl of extracts separately onto the circular area. As control treatment the same amount of hexane was pipetted onto the circular area of the arena. Wasps were assayed after solvent evaporation. A total of 30 wasp females was used for each treatment.
Hexane Extracts of H. halys Females Blended With Synthetic 1-Hexadecene
The response of the wasps toward H. halys female extract blended with synthetic 1-hexadecene was compared to crude H. halys female extract. The test and control extracts were tested by pipetting 100μl onto the circular area of the arena. A total of 30 wasp females was tested for each treatment.
All the bioassays were carried out in a controlled environmental room at 26 ± 1°C from 10:00 a.m. to 3:00 p.m. The treated arena was changed for each replication. The bioassays were performed in three blocks and trials randomized within each block. The wasps were used only once and discarded at the end of each replication.
Chemical Analysis
Coupled gas chromatography-mass spectrometry (GC-MS) analyses of cuticular extracts from H. halys males and females were performed on an Agilent 6890 GC system interfaced with an MS5973 quadruple mass spectrometer. For each sample, 1 μl of extract was injected in a DB5-MS column in splitless mode with helium used carrier gas. Injector and detector temperatures were 260 and 280°C, respectively. The GC oven temperature was set at 40°C for 5 min, and then increased by 10°C/min to 250°C. Electron impact ionization spectra were obtained at 70 eV, recording mass spectra from 40 to 550 amu. For quantification purposes, peak area of each detected compound was calculated. Compounds were tentatively identified based on comparison of RI and mass spectra with those reported in literature (Adams et al., 2007; www.pherobase.com) and the NIST 2011 library. For dodecane, tridecane, tetradecane, pentadecane, 1-hexadecene, pentacosane, and nonacosane identification was confirmed by injection of authentic standards, obtained from Sigma-Aldrich (Milan, Italy). For the compound contributing the most (according to multivariate statistics) in explaining a chemical difference between the hexane extracts of male and female bugs, we carried out additional analyses. For this compound, a calibration curve with the standard at the concentration of 0.5, 1, 2, 10, and 20 ng/μl diluted in hexane (linearity R2 = 0.99) was carried out to calculate the amount present in H. halys extracts from males or females.
Statistical Analyses
Residence time data were not normally distributed (which is typical of time-to-event data) and thus were analyzed with General Linear Models (GLMs) fitting gamma error distribution and a reciprocal link function (Crawley, 2007). For each of the three experiments, the different stink bug treatments were modeled as fixed factors while wasp residence time was fitted in the GLMs as response variable. Significance of the factor in each GLM was tested using Likelihood Ratio Tests (LRTs) comparing the full model with and without the factor in question (Crawley, 2007). Model fit was assessed by inspecting residual plots. For the experiment with hexane extracts of H. halys cuticular compounds, significant differences amongst factor levels were determined using the glht function of the multcomp package of the R software (Bretz et al., 2010).
Peak areas of chemical compounds were analyzed through multivariate data analysis using projection to latent structures discriminant analysis (PSL-DA). The results of the analysis are visualized in score plots, which reveal the sample structure according to model components, and loading plots, which display the contribution of the variables to these components as well as the relationships among the variables, with the ranking based on the variable importance in the projection (VIP values) (Wold et al., 2001). The measured peak areas were first divided by the fresh stink bug weight and, subsequently, mean-centered and scaled to unit variance before they were subjected to the analysis using the software metaboanalyst (Xia et al., 2009).
Results
Open Arena Bioassay
Freshly Killed H. halys Adults
The residence time of T. japonicus females was significantly affected by the gender of the stink bugs (GLM, χ2 = 8.88, df = 1, P = 0.003). Parasitoids spent more time exploring patches contaminated with chemical residues of host females compared with males (Figure 1).
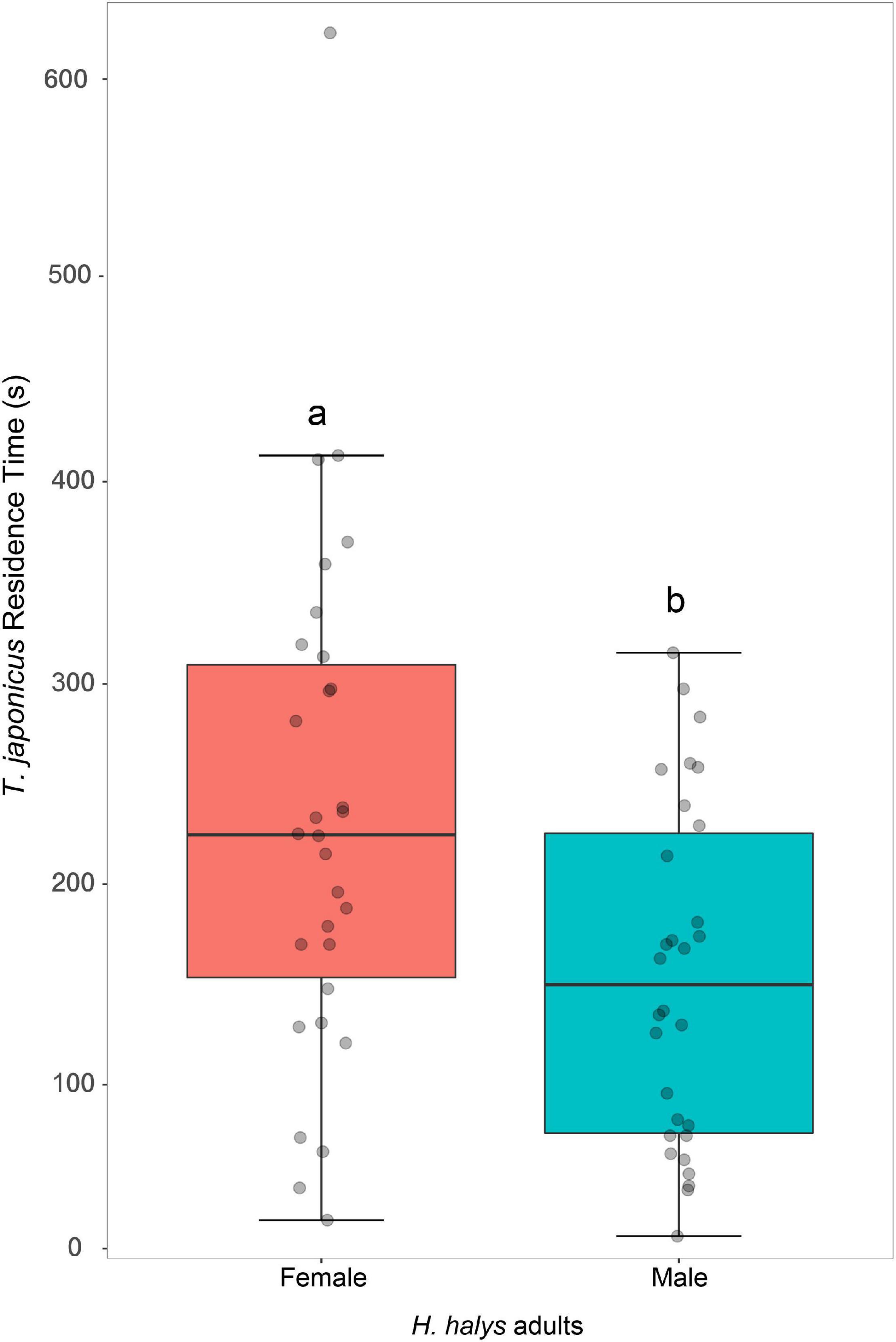
Figure 1. Response of Trissolcus japonicus females to substrate treated with traces of freshly killed Halyomorpha halys females (red box) or males (blue box). Bold horizontal lines show medians, boxes contain the 25th–50th percentiles, whiskers show the upper and lower quartiles and points represent individual replicates. Different letters indicate statistically significant difference (P < 0.05, GLM).
Hexane Extracts of H. halys Cuticular Compounds
The residence time of T. japonicus females was significantly affected by the extract treatments (GLM, χ2 = 43.12, df = 2, P < 0.001). Parasitoids displayed host sex discrimination (Z = 2.84, P = 0.012), spending, on average, the highest amount of time on crude extract from H. halys females, intermediate levels on H. halys males and the lowest amount of time on control (solvent only) (Figure 2).
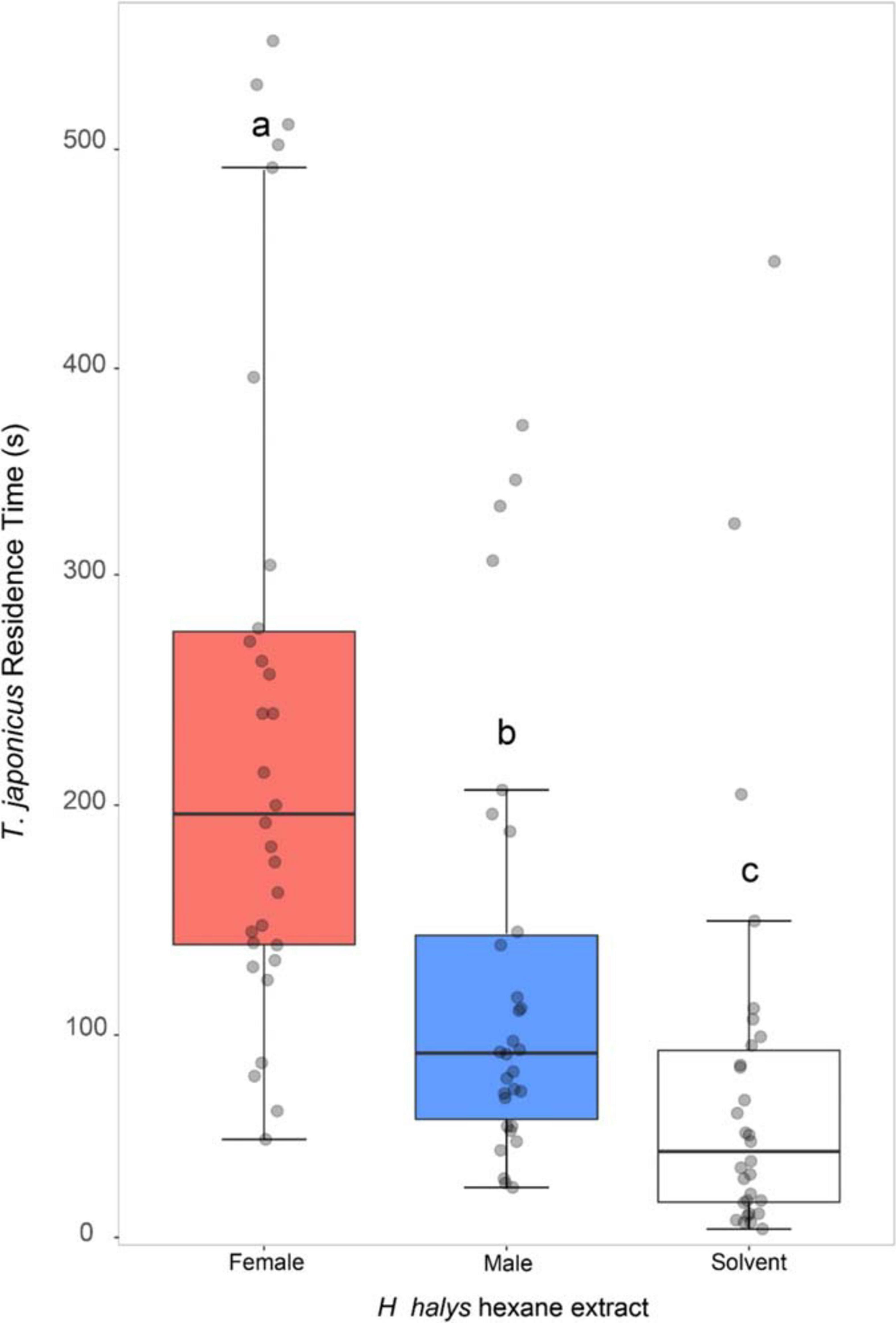
Figure 2. Response of Trissolcus japonicus females to substrate treated with hexane extracts of Halyomorpha halys females (red box) or males (blue box) or with hexane only (white box). Bold horizontal lines show medians, boxes contain the 25th–50th percentiles, whiskers show the upper and lower quartiles and points represent individual replicates. Different letters indicate statistically significant difference (P < 0.05, GLM).
Hexane Extracts of H. halys Females Blended With Synthetic 1-Hexadecene
The residence time of T. japonicus females was significantly affected by the addition of 1-hexadecene to the crude hexane extract (GLM, χ2 = 35.40, df = 1, P < 0.001). Parasitoids spent significantly less time on patches contaminated with hexane extract of H. halys females blended with 1-hexadecene compared with crude extract of H. halys females (Figure 3).
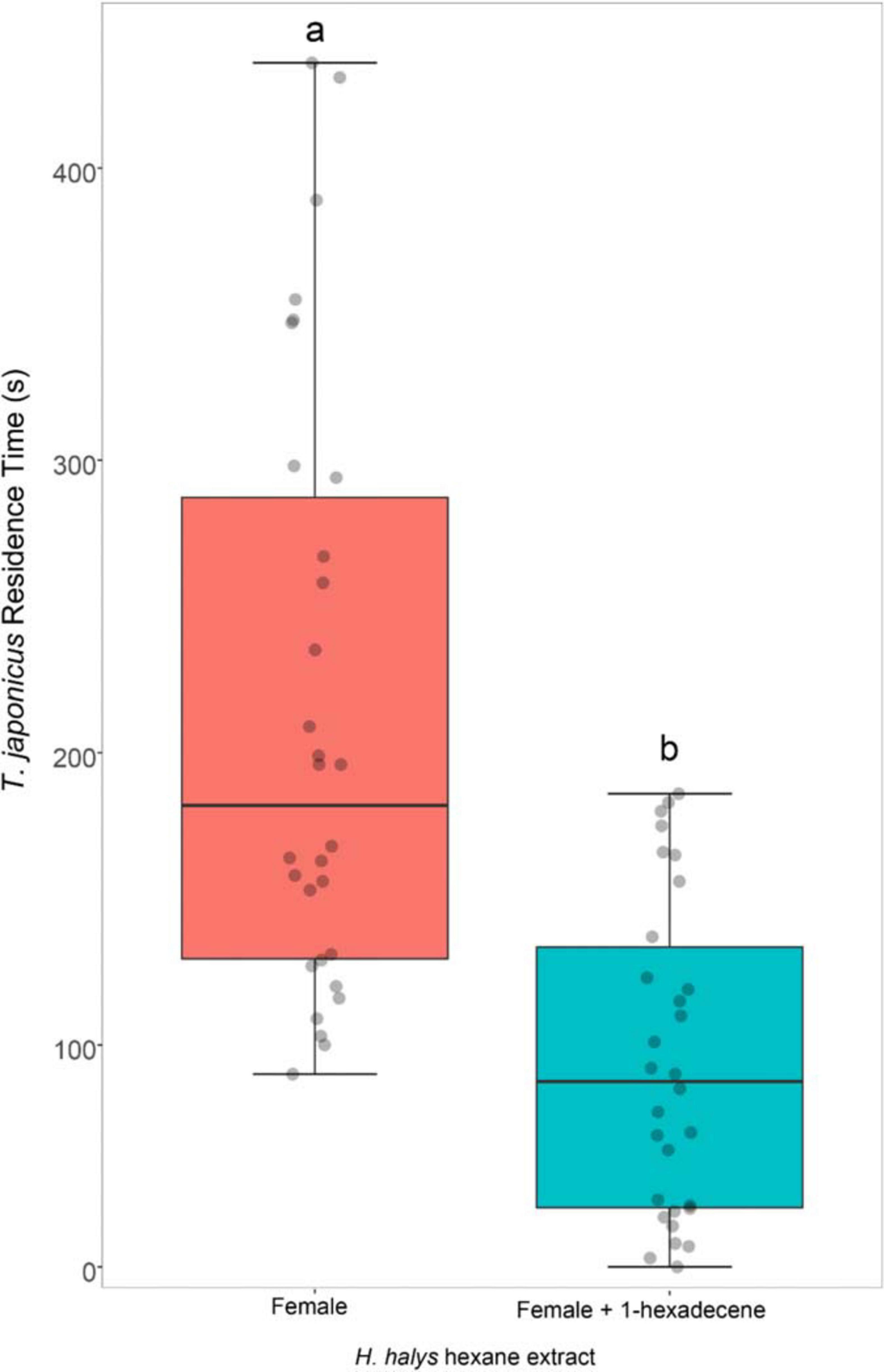
Figure 3. Response of Trissolcus japonicus females to substrate treated with hexane extract of Halyomorpha halys females (red box) or with hexane extracts of H. halys females blended with 1-hexadecene (blue box). Bold horizontal lines show medians, boxes contain the 25th–50th percentiles, whiskers show the upper and lower quartiles and points represent individual replicates. Different letters indicate statistically significant difference (P < 0.05, GLM).
Chemical Analysis
Chemical analysis of hexane extracts of dissected body parts from H. halys females or males showed the presence of 21 compounds (Table 1), Among them, 11 linear hydrocarbons (5 saturated and 6 mono-unsaturated), 3 aldehydes, 2 alcohols, 2 esters and 1 carboxylic acid were detected, while 2 compounds remained unidentified. Overall, the same compounds were detected in male and female extracts but in different proportions. Thus, the chemical composition of the extracts varied according to the stink bug sex (Figure 4A). In fact, the pairwise comparison by PLS-DA between males and females resulted in a model with a significant principal component (PCs) (Figure 4B). For this model, 9 compounds had VIP (Variable Important for the Projection) values of ≥ 1.0, which means that they strongly contributed to explaining the differences between stink bug extracts (Figure 4C). The compound with the highest VIP value (=1.995) was 1-hexadecene.
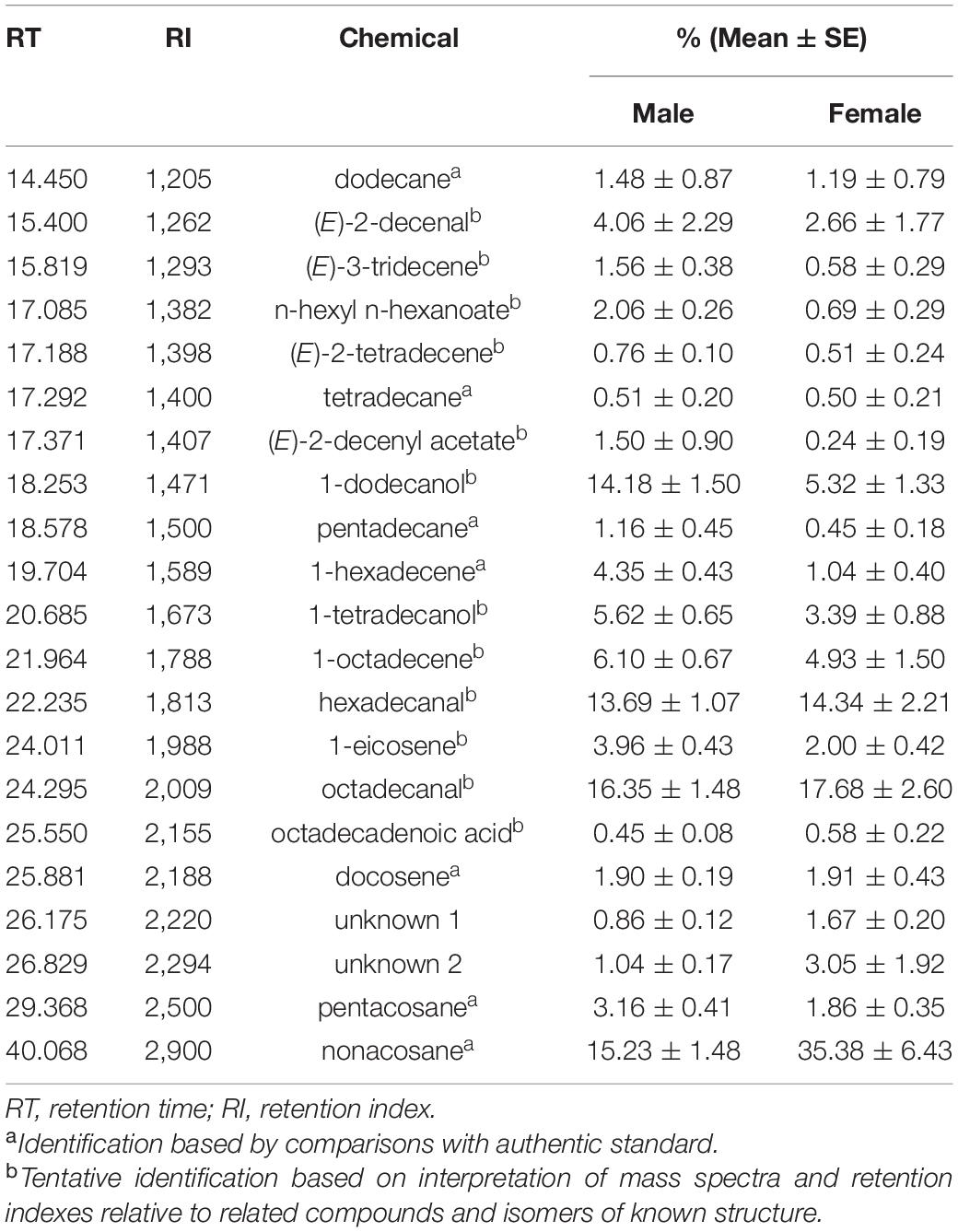
Table 1. Compounds detected in hexane extracts of dissected body parts from Halyomorpha halys adults.
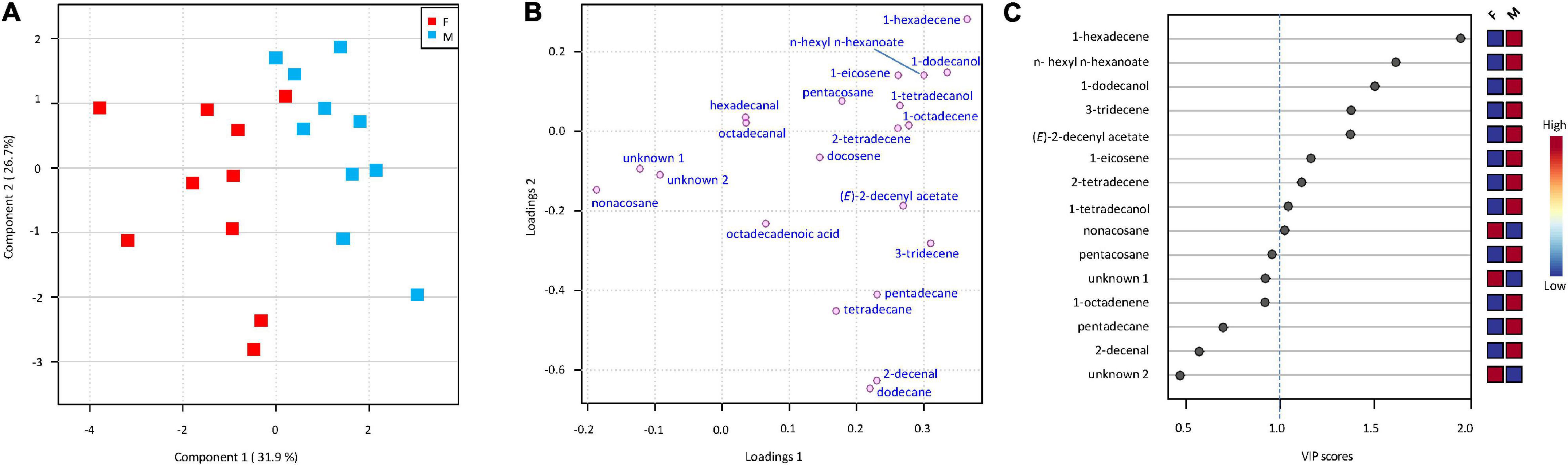
Figure 4. Projection to latent structures discriminant analysis (PLS-DA) of cuticular compounds extracted from Halyomorpha halys females (F) and males (M). (A) Score plot visualizing the grouping pattern of the samples according to the first two principal components (PCs) with the explained variance in parenthesis. The PLS-DA resulted in a model with a significant component (R2X = 0.319, R2Y = 0.628, Q2 = 0.470), but only the first two PCs are shown for representational purposes. (B) Loading plot of the first two PCs shows the contribution of each compound to the two PLS-DA components. (C) Compounds ranked according to values of Variable Important for the Projection (VIP). Compounds with VIP value < 0.5 are not shown. Red squares indicate higher relative amounts between males and females, blue squares indicate lower relative amounts.
Discussion
We have provided behavioral and chemical evidence on the role of specific cuticular compounds left on the substrate by H. halys which are subsequently exploited as contact chemical cue by its egg parasitoid T. japonicus while foraging for host eggs. Contact chemicals left by the insects during walking activity can act as intraspecific and interspecific signals (Waage, 1979; Klomp, 1981; Rostás et al., 2008; Wilms and Eltz, 2008). In the case of egg parasitoids of the family Scelionidae, chemical traces play a role as interspecific signals exploited during host searching behavior; for example this is the cases of T. basalis and Trissolcus brochymenae Ashmed (Hymenoptera: Scelionidae) the main egg parasitoids of N. viridula and Murgantia histrionica Hahn (Heteroptera: Pentatomidae), respectively (Colazza et al., 1999; Salerno et al., 2009). Our results showed that T. japonicus responded to the substrates contaminated with H. halys chemical traces, exploring more intensively and spending more time on female-associated cues over male-associated cues. Moreover, the preference of the T. japonicus for H. halys pre-ovipositional females observed when hexane extracts were used, suggests that non-polar cuticular compounds are responsible for host sex discrimination. This behavioral response is adaptive because host females are more likely to be linked with egg presence, thus a wide exploration of the area contaminated with female chemical traces left by females may increase the chances of finding reproductive opportunities for egg parasitoids (Peri et al., 2006, 2016). Other studies also reported that non-polar compounds associated with traces left by walking and crawling insects can act as kairomones for parasitoids (González et al., 2011; Colazza et al., 2014; Fürstenau and Hilker, 2017; Gomes Lagôa et al., 2020). For example, cuticular hydrocarbons of Tribolium confusum Jacquelin du Val (Coleoptera: Tenebrionidae) larvae mediate trail following and host recognition in the ectoparasitoid Holepyris sylvanidis Brethes (Hymenoptera: Bethylidae) (Fürstenau and Hilker, 2017).
The chemical analysis of the cuticular extracts of H. halys adults indicated the presence of non-polar compounds such as hydrocarbons as also reported by Malek (2020) and as also observed in other stink bugs (De Pasquale et al., 2007; Guarino et al., 2008). Our chemical investigations showed that the whole chemical profiles of H. halys males and pre-ovipositional females are quantitatively different. Our multivariate statistical analysis showed that this difference is mainly due to quantitative changes in linear hydrocarbons such as 1-hexadecene which was more abundant in males, suggesting a possible role in mediating the parasitoid’s host discrimination. In fact, the hexane extracts of H. halys females blended with synthetic 1-hexadecene mimic the chemical profile of H. halys males and induced a behavioral response in T. japonicus with intensity comparable to what we observed when male extracts were tested. In a similar study carried out by Colazza et al. (2007), it was found that another cuticular hydrocarbon, n-nonadecane, mediates analogous effects in the interaction between T. basalis and N. viridula, whereas Gomes Lagôa et al. (2020) showed that hydrocarbons left by stink bugs on the walking substrate play a role in host species discrimination by T. basalis and Telenomus podisi (Ashmead) (Hymenoptera: Scelionidae). We can thus speculate that the use of linear hydrocarbons to discriminate the sex of adults of the host species might be a common strategy that has evolved in members of the Scelionidae.
In addition to being important components of the external layers of the insect body, hydrocarbons are abundant in the epicuticular waxes of foliage and other plant tissues (Eigenbrode and Espelie, 1995; Müller and Riederer, 2005). Despite this chemical similarity between hosts and leaves (Lo Giudice et al., 2010), Trissolcus females are able to find hosts. In the epicuticular wax layer of stink bugs there are other components in addition to linear hydrocarbons, such as branched hydrocarbons or more polar components, the roles of which remain to be investigated in the context of the host searching behavior. Further, although the stink bug’s cuticle appears to be the source of the contact kairomones that mediate the foraging response of egg parasitoids to walking traces, it cannot be excluded that cues may derive from adhesive substances produced by tarsal glands that facilitate the bugs’ walking activity on different substrates, as recently observed in other Heteropteran species (Rebora et al., 2021). Although we tried to minimize the presence of defensive compounds, it cannot be excluded that a few chemicals observed in our analyses, such as dodecane and (E)-2-decenal were originated from methatoracic glands. However, these compounds did not appear to play an important role in explaining the differences between the chemical profiles of H. halys sexes, based on our multivariate statistical analysis.
The study of the semiochemicals involved in the foraging behavior of insect parasitoids is not only important from a basic ecological perspective but also in the optimization of CBC. To date, the manipulation of parasitoid behavior in order to improve the effectiveness of biological control programs has been mainly focused in the application to crops of exogenous forms of herbivore induced plant volatiles that can draw natural enemies from nearby donor habitats into crops when pests arrive (James and Price, 2004; James and Grasswitz, 2005; Gurr and You, 2016). In this context, there is also now the possibility of applying by-products of the target host species that act as contact kairomones and retain parasitoids in the focal crop. These methods could be integrated with well-known CBC practices such as planting flower strips alongside crops to provide additional food resources for natural enemies (Gurr et al., 2017). Such combined strategy, called “attract-and-reward” (Simpson et al., 2011) has been shown to mitigate the negative effects of aversion learning in parasitoid wasps, which can occur when parasitoids are lured in the focal crop with synthetic semiochemicals without finding resources such as hosts or food. Yet future studies under field conditions are needed to test the possibility to maximize T. japonicus performance by combining both volatile (Zhong et al., 2017; Bertoldi et al., 2019) and contact kairomones with floral resources. Integrating chemical ecology with habitat manipulation could be of particular importance given the large and negative economic impact that H. halys has in both North American and European cropping systems that adventive populations of T. japonicus have been found in areas previously invaded by H. halys and that T. japonicus is currently the most promising biocontrol agent in both continents.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
MA and TA carried out the behavioral experiments. SG carried out the chemical analysis. AC carried out the statistical analysis. EP and SC conceived and designed the research All authors wrote and revised the manuscript.
Funding
This research was partially funded by the University of Palermo (FFR_D13-2018/2021).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Prof. Ian Hardy (University of Nottingham) for his insightful critical comments on a previous version of the manuscript. We thank Prof. Luciana Tavella and Dr. Silvia Moraglio (University of Turin) for kindly providing a colony of Trissolcus japonicus.
References
Aartsma, Y., Cusumano, A., Fernández de Bobadilla, M., Rusman, Q., Vosteen, I., and Poelman, E. H. (2019). Understanding insect foraging in complex habitats by comparing trophic levels: insights from specialist host-parasitoid-hyperparasitoid systems. Curr. Opin. Insect Sci. 32, 54–60. doi: 10.1016/j.cois.2018.11.001
Abram, P. K., Hoelmer, K. A., Acebes-Doria, A., Andrews, H., Beers, E. H., Bergh, J. C., et al. (2017). Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 90, 1009–1020. doi: 10.1007/s10340-017-0891-7
Adams, R. G., Lohmann, R., Fernandez, L. A., MacFarlane, J. K., and Gschwend, P. M. (2007). Polyethylene devices: passive samplers for measuring dissolved hydrophobic organic compounds in aquatic environments. Environ. Sci. Technol. 41, 1317–1323. doi: 10.1021/es0621593
Aspin, E., Keller, M. A., Yazdani, M., and Hardy, I. C. W. (2021). Walk this way, fly that way: Goniozus jacintae attunes flight and foraging behaviour to leaf-roller host instar. Entomol. Exp. Appl. 169, 350–361. doi: 10.1111/eea.13028
Bell, W. J. (1990). Searching behavior patterns in insects. Annu. Rev. Entomol. 35, 447–467. doi: 10.1146/annurev.en.35.010190.002311
Bertoldi, V., Rondoni, G., Brodeur, J., and Conti, E. (2019). An egg parasitoid efficiently exploits cues from a coevolved host but not those from a novel host. Front. physiol. 10:746. doi: 10.3389/fphys.2019.00746
Borges, M., Colazza, S., Ramirez-Lucas, P., Chauhan, K. R., Blassioli Moraes, M. C., and Aldrich, J. R. (2003). Kairomonal effect of walking traces from Euschistus heros (Heteroptera: Pentatomidae) on two strains of Telenomus podisi (Hymenoptera: Scelionidae). Physiol. Entomol. 28, 349–355. doi: 10.1111/j.1365-3032.2003.00350.x
Boyle, S. M., Weber, D. C., Hough-Goldstein, J., and Hoelmer, K. A. (2020). Host kairomones influence searching behavior of Trissolcus japonicus (Hymenoptera: Scelionidae), a parasitoid of Halyomorpha halys (Heteroptera: Pentatomidae). Environ. Entomol. 49, 15–20. doi: 10.1093/ee/nvz155
Bretz, F., Hothorn, T., and Westfall, P. (2010). Multiple Comparisons Using R, 1st Edn. Boca Raton, FL: Chapman and Hall/CRC, doi: 10.1201/9781420010909
Colazza, S., Aquila, G., De Pasquale, C., Peri, E., and Millar, J. G. (2007). The egg parasitoid Trissolcus basalis uses n-nonadecane, a cuticular hydrocarbon from its stink bug host Nezara viridula, to discriminate between female and male hosts. J. Chem. Ecol. 33, 1405–1420. doi: 10.1007/s10886-007-9300-7
Colazza, S., Cusumano, A., Lo Giudice, D., and Peri, E. (2014). Chemo-orientation responses in hymenopteran parasitoids induced by substrate-borne semiochemicals. BioControl. 59, 1–17. doi: 10.1007/s10526-013-9546-7
Colazza, S., Peri, E., Salerno, G., and Conti, E. (2010). “Host searching by egg parasitoids: Exploitation of host chemical cues,” in Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma, eds F. Consoli, J. Parra, and R. Zucchi (Dordrecht: Springer), 97–147. doi: 10.1007/978-1-4020-9110-0_4
Colazza, S., Salerno, G., and Wajnberg, E. (1999). Volatile and contact chemicals released by Nezara viridula (Heteroptera:Pentatomidae) have a kairomonal effect on the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Biol. Control. 16, 310–317. doi: 10.1006/bcon.1999.0763
Conti, E., Avila, G., Barratt, B., Cingolani, F., Colazza, S., Guarino, S., et al. (2020). Biological control of invasive stink bugs: review of global state and future prospects. Entomol. Exp. Appl. 169, 28–51. doi: 10.1111/eea.12967
Conti, E., and Colazza, S. (2012). Chemical ecology of egg parasitoids associated with true bugs. Psyche J. Entomol. 2012:651015. doi: 10.1155/2012/651015
Conti, E., Salerno, G., Bin, F., and Vinson, S. B. (2004). The role of host semiochemicals in parasitoid specificity: a case study with Trissolcus brochymenae and Trissolcus simoni on pentatomid bugs. Biol. Control. 29, 435–444. doi: 10.1016/j.biocontrol.2003.08.009
De Pasquale, C., Guarino, S., Peri, E., Alonzo, G., and Colazza, S. (2007). Investigation of cuticular hydrocarbons from Bagrada hilaris genders by SPME/GC-MS. Anal. Bioanal. Chem. 389, 1259–1265. doi: 10.1007/s00216-007-1503-9
Eigenbrode, S. D., and Espelie, K. E. (1995). Effects of plant epicuticular lipids on insect herbivores. Annu. Rev. Entomol. 40, 171–194. doi: 10.1146/annurev.en.40.010195.001131
Fatouros, N. E., Dicke, M., Mumm, R., Meiners, T., and Hilker, M. (2008). Foraging behavior of egg parasitoids exploiting chemical information. Behav. Ecol. 19, 677–689. doi: 10.1093/beheco/arn011
Fürstenau, B., and Hilker, M. (2017). Cuticular hydrocarbons of Tribolium confusum larvae mediate trail following and host recognition in the ectoparasitoid Holepyris sylvanidis. J. Chem. Ecol. 43, 858–868. doi: 10.1007/s10886-017-0885-1
Gomes Lagôa, A. C., Blassioli Moraes, M. C., Borges, M., and Laumann, R. A. (2020). Selective responses of Trissolcus basalis and Telenomus podisi to chemical footprints of preferred hosts. Physiol. Entomol. 45, 60–71. doi: 10.1111/phen.12316
González, J. M., Cusumano, A., Williams, H. J., Colazza, S., and Vinson, S. B. (2011). Behavioral and chemical investigations of contact kairomones released by the mud dauber wasp Trypoxylon politum, a host of the parasitoid Melittobia digitata. J. Chem. Ecol. 37, 629–639. doi: 10.1007/s10886-011-9960-1
Guarino, S., De Pasquale, C., Peri, E., Alonzo, G., and Colazza, S. (2008). Role of volatile and contact pheromones in the mating behaviour of Bagrada hilaris (Heteroptera: Pentatomidae). Eur. J. Entomol. 105, 613–617. doi: 10.14411/eje.2008.082
Gurr, G. M., Wratten, S. D., Landis, D. A., and You, M. (2017). Habitat management to suppress pest populations: progress and prospects. Annu. Rev. Entomol. 62, 91–109. doi: 10.1146/annurev-ento-031616-035050
Gurr, G. M., and You, M. (2016). Conservation biological control of pests in the molecular era: new opportunities to address old constraints. Front. Plant Sci. 6:1255. doi: 10.3389/fpls.2015.01255
Hedstrom, C., Lowenstein, D., Andrews, H., Bai, B., and Wiman, N. (2017). Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J. Pest Sci. (2004). 90, 1169–1179. doi: 10.1007/s10340-017-0892-6
Hilker, M., and Fatouros, N. E. (2015). Plant responses to insect egg deposition. Annu. Rev. Entomol. 60, 493–515. doi: 10.1146/annurev-ento-010814-020620
James, D. G., and Grasswitz, T. R. (2005). Synthetic herbivore-induced plant volatiles increase field captures of parasitic wasps. BioControl 50, 871–880. doi: 10.1007/s10526-005-3313-3
James, D. G., and Price, T. S. (2004). Field-testing of methyl salicylate for recruitment and retention of beneficial insects in grapes and hops. J. Chem. Ecol. 30, 1613–1628. doi: 10.1023/B:JOEC.0000042072.18151.6f
Kaser, J. M., Akotsen-Mensah, C., Talamas, E. J., and Nielsen, A. L. (2018). First report of Trissolcus japonicus parasitizing Halyomorpha halys in North American agriculture. Florida Entomol. 101, 680–683. doi: 10.1653/024.101.0406
Khan, Z. R., James, D. G., Midega, C. A. O., and Pickett, J. A. (2008). Chemical ecology and conservation biological control. Biol. Control 45, 210–224. doi: 10.1016/j.biocontrol.2007.11.009
Klomp, H. (1981). Parasitic wasps as sleuth hounds—response of an ichneumon wasp to the trail of its host. Neth. J. Zool. 31, 762–772. doi: 10.1163/002829681X00275
Leskey, T. C., and Nielsen, A. L. (2018). Impact of the invasive brown marmorated stink bug in North America and Europe: history, biology, ecology, and management. Annu. Rev. Entomol. 63, 599–618. doi: 10.1146/annurev-ento-020117-043226
Lo Giudice, D., Peri, E., Lo Bue, M., and Colazza, S. (2010). Plant surfaces of vegetable crops mediate interactions between chemical footprints of true bugs and their egg parasitoids. Commun. Integr. Biol. 3, 70–74. doi: 10.4161/cib.3.1.10300
Lockey, K. H. (1985). Insect cuticular lipids. Comp. Biochem. Physiol. Part B Biochem. 81, 263–273. doi: 10.1016/0305-0491(85)90311-6
Malek, R. (2020). Novel Monitoring and Biological Control of Invasive Insect Pests. Trento: University of Trento. Doctoral thesis.
Meiners, T., and Peri, E. (2013). “Chemical ecology of insect parasitoids: essential elements for developing effective biological control programmes,” in Chemical Ecology of Insect Parasitoids, eds E. Wajnberg and S. Colazza (Hoboken, NJ: John Wiley & Sons, Ltd), 191–224. doi: 10.1002/9781118409589.ch9
Müller, C., and Riederer, M. (2005). Plant surface properties in chemical ecology. J. Chem. Ecol. 31, 2621–2651. doi: 10.1007/s10886-005-7617-7
Padmavathi, C., and Paul, A. V. N. (1998). Saturated hydrocarbons as kairomonal source for the egg parasitoid, Trichogramma chilonis Ishii (Hym., Trichogrammatidae). J. Appl. Entomol. 122, 29–32. doi: 10.1111/j.1439-0418.1998.tb01456.x
Paul, A. V. N., Singh, S., and Singh, A. K. (2002). Kairomonal effect of some saturated hydrocarbons on the egg parasitoids, Trichogramma brasiliensis (Ashmead) and Trichogramma exiguum, Pinto, Platner and Oatman (Hym., Trichogrammatidae). J. Appl. Entomol. 126, 409–416. doi: 10.1046/j.1439-0418.2002.00661.x
Peri, E., Salerno, G., Slimani, T., Frati, F., Conti, E., Colazza, S., et al. (2016). The response of an egg parasitoid to substrate-borne semiochemicals is affected by previous experience. Sci. Rep. 6:27098. doi: 10.1038/srep27098
Peri, E., Sole, M. A., Wajnberg, E., and Colazza, S. (2006). Effect of host kairomones and oviposition experience on the arrestment behavior of an egg parasitoid. J. Exp. Biol. 209, 3629–3635. doi: 10.1242/jeb.02416
Rebora, M., Salerno, G., Piersanti, S., Gorb, E., and Gorb, S. (2021). Attachment devices and the tarsal gland of the bug Coreus marginatus (Hemiptera: Coreidae). Zoomorphology 140, 85–102. doi: 10.1007/s00435-020-00515-z
Rice, K. B., Bergh, C. J., Bergmann, E. J., Biddinger, D. J., Dieckhoff, C., Dively, G., et al. (2014). Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 5, 1–13. doi: 10.1603/ipm14002
Rodriguez-Saona, C. R., Mescher, M. C., and De Moraes, C. M. (2013). “The role of volatiles in plant–plant Interactions,” in Long-Distance Systemic Signaling and Communication in Plants, ed. F. Baluška (Berlin: Springer), 393–412. doi: 10.1007/978-3-642-36470-9_19
Rostás, M., Ruf, D., Zabka, V., and Hildebrandt, U. (2008). Plant surface wax affects parasitoid’s response to host footprints. Naturwissenschaften 95, 997–1002. doi: 10.1007/s00114-008-0411-y
Salerno, G., Frati, F., Conti, E., De Pasquale, C., Peri, E., and Colazza, S. (2009). A finely tuned strategy adopted by an egg parasitoid to exploit chemical traces from host adults. J. Exp. Biol. 212, 1825–1831. doi: 10.1242/jeb.028308
Simpson, M., Gurr, G. M., Simmons, A. T., Wratten, S. D., James, D. G., Leeson, G., et al. (2011). Attract and reward: combining chemical ecology and habitat manipulation to enhance biological control in field crops. J. Appl. Entomol. 48, 580–590. doi: 10.1111/j.1365-2664.2010.01946.x
Stahl, J., Tortorici, F., Pontini, M., Bon, M. C., Hoelmer, K., Marazzi, C., et al. (2019). First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest Sci. 92, 371–379. doi: 10.1007/s10340-018-1061-2
Turlings, T. C. J., and Erb, M. (2018). Tritrophic interactions mediated by herbivore-induced plant volatiles: mechanisms, ecological relevance, and application potential. Annu. Rev. Entomol. 63, 433–452. doi: 10.1146/annurev-ento-020117-043507
Van Poecke, R. M. P., Posthumus, M. A., and Dicke, M. (2001). Herbivore-induced volatile production by Arabidopsis thaliana leads to attraction of the parasitoid Cotesia rubecula: Chemical, behavioral, and gene-expression analysis. J. Chem. Ecol. 27, 1911–1928. doi: 10.1023/A:1012213116515
Waage, J. K. (1979). Foraging for patchily-distributed hosts by the parasitoid, Nemeritis canescens. J. Animal Ecol. 49, 353–371. doi: 10.2307/4166
Wajnberg, E., and Colazza, S. (2013). Chemical Ecology of Insect Parasitoids. West Sussex: John Wiley & Sons, Ltd, doi: 10.1002/9781118409589
Wang, S. Y., Hackney Price, J., and Zhang, D. (2019). Hydrocarbons catalysed by TmCYP4G122 and TmCYP4G123 in Tenebrio molitor modulate the olfactory response of the parasitoid Scleroderma guani. Insect Mol. Biol. 28, 637–648. doi: 10.1111/imb.12581
Wilms, J., and Eltz, T. (2008). Foraging scent marks of bumblebees: footprint cues rather than pheromone signals. Naturwissenschaften 95, 149–153. doi: 10.1007/s00114-007-0298-z
Wold, S., Sjöström, M., and Eriksson, L. (2001). PLS-regression: a basic tool of chemometrics. Chemom. Intell. Lab. Syst. 58, 109–130. doi: 10.1016/S0169-7439(01)00155-1
Xia, J., Psychogios, N., Young, N., and Wishart, D. S. (2009). MetaboAnalyst: a web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 37, W652–W660. doi: 10.1093/nar/gkp356
Keywords: host searching behavior, Halyomorpha halys, cuticular hydrocarbons, kairomone, Scelionidae, 1-hexadecene, samurai wasp
Citation: Arif MA, Alınç T, Guarino S, Colazza S, Cusumano A and Peri E (2021) Only Females Oviposit: Chemical Discrimination of Adult Stink Bug Sex by the Egg Parasitoid Trissolcus japonicus. Front. Ecol. Evol. 9:652561. doi: 10.3389/fevo.2021.652561
Received: 12 January 2021; Accepted: 22 March 2021;
Published: 13 April 2021.
Edited by:
Lukasz Lech Stelinski, University of Florida, United StatesReviewed by:
Nik G. Wiman, Oregon State University, United StatesLara Maistrello, University of Modena and Reggio Emilia, Italy
Copyright © 2021 Arif, Alınç, Guarino, Colazza, Cusumano and Peri. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Salvatore Guarino, c2FsdmF0b3JlLmd1YXJpbm9AaWJici5jbnIuaXQ=
†These authors have contributed equally to this work