 Jen Nightingale
Jen Nightingale- 1School of Biological Sciences, University of Bristol, Bristol, United Kingdom
- 2Bristol Zoological Society, Bristol, United Kingdom
- 3Cefas, Weymouth, United Kingdom
- 4APEM Ltd., Stockport, United Kingdom
Developing an optimal diet for rearing endangered white-clawed crayfish Austropotamobius pallipes is important for captive breeding success prior to wild release. Four ex situ, 40-day experiments assessed survival and growth of crayfish fed different treatment diets. Two experiments (A and B) were undertaken with hatchlings, to determine if live food was an essential dietary component during the first few weeks after hatching. The second set of experiments (C and D) were undertaken with juvenile (60-day-old) A. pallipes, to determine an optimal diet after the initial critical feeding stage. In experiment A, we fed hatchlings: i) live Artemia nauplii + plankton (Live + P); ii) decapsulated Artemia cysts + plankton (Cyst + P) or iii) decapsulated Artemia cysts + plankton encapsulated in agar gel (Gel + CP). Survival and growth was significantly greater with Live + P than with the other two diets. In experiment B we compared Live + P with commercially available feeds by feeding hatchlings: i) live Artemia nauplii + Australian pellet (Live + Aus); ii) live Artemia nauplii plus New Zealand pellet (Live + NZ); iii) live Artemia nauplii + plankton (Live + P); or (iv) practical Spanish crayfish pellet diet (Spain). Under these experimental conditions crayfish survival was significantly higher with Live + P diet than with Live + Aus or Spain. Growth was also significantly greater with Live + P than with the Live + NZ or Spanish treatment diets. In experiment C, 60-day-old juvenile A. pallipes were fed: i) defrosted plankton plus vegetables (Standard) or (ii) defrosted plankton plus vegetables encapsulated in agar gel (Gel + PV). Survival was not significantly different between the diets; however, growth was significantly greater with the Standard diet rather than Gel + PV. In experiment D, juveniles were fed four different diets: i) Australian pellet (Australia); ii) New Zealand pellet (New Zealand); iii) plankton and vegetables (Standard); or iv) practical Spanish diet (Spain). Survival was significantly lower in crayfish fed the New Zealand diet. Crayfish growth was significantly greater with the Standard diet of plankton and vegetables than all three pellet diets. Our results showed that live food is optimal for high survival and growth in A. pallipes hatchlings and a plankton, plus vegetable, diet produces higher growth in juveniles compared to pellet diets.
Introduction
The white-clawed crayfish Austropotamobius pallipes is endangered throughout its native range in the United Kingdom and mainland Europe (Sibley et al., 2011). The loss of this species is attributed to the spread of the invasive American signal crayfish Pacifastacus leniusculus and associated crayfish plague, caused by the pathogen/oomycete Aphanomyces astaci, along with habitat degradation and pollution (Sibley et al., 2011). In response to this decline, white-clawed crayfish aquaculture is increasing in the United Kingdom and mainland Europe (Nightingale et al., 2017).
A major issue with the mass-rearing of animals in captivity is the provision of a well-balanced and nutritional diet, to ensure good survival, growth and development, which is particularly important for juvenile life stages. With crayfish, a critical period of survival occurs during the first weeks, post-hatching, when high mortality rates can occur due to a lack of adequate nutrition (González et al., 2011; Celada et al., 2012). When crayfish hatch, they initially feed on their egg yolk and remain attached to the female’s pleopods. When they have undergone two moults and are free-living, their exogenous mouth parts have formed and feeding begins (Reynolds, 2002). Observations from the wild can provide key information on suitable diets for captive-bred animals. Wild A. pallipes are opportunistic omnivores, feeding on invertebrates, carrion, vegetable matter, and organic and inorganic detritus (Gherardi et al., 2004). Scalici and Gibertini (2007) found stomach contents of wild-caught A. pallipes differing with age and sex. Insect larvae were found to be a key component of juvenile and adult female diets, in contrast, adult males mainly fed on vegetable matter. These findings are also supported by in situ analysis of the gut contents of A. pallipes juveniles, which were shown to be feeding predominantly on aquatic invertebrates (Paglianti and Gherardi, 2004). This ontogenetic diet shift may be a result of the digestive enzymes within the hepatopancreas altering as crayfish mature and this corresponds to changes in diets (Hammer et al., 2000; Figueiredo and Anderson, 2003).
Several studies have examined feeding and nutrition requirements of captive-bred crayfish, including P. leniusculus (Carral et al., 2011; González et al., 2012), red swamp crayfish Procambarus clarkii (Hua et al., 2015) and common yabby Cherax destructor (Austin et al., 1997). Commercially available fish-feed pellets were historically fed to all age-classes resulting in low survival rates of hatchlings in both A. pallipes (Sáez-Royuela et al., 2001) and P. leniusculus (Ulikowski et al., 2006; Sáez-Royuela et al., 2007). Artemia nauplii have also been used as feed for captive-bred juvenile crayfish, as these are a readily available and easily produced substitute for insect larvae found in the diet of wild juvenile crayfish. Artemia nauplii are a popular first feed within aquaculture as they are high in protein and lipids and contain proteolytic enzymes, which can aid the digestive abilities of young animals (Bengtson et al., 1991). As Artemia will readily feed on a wide variety of food items, they provide a useful vessel for enrichment products, such as lipids and algae, to be easily incorporated (Léger et al., 1986).
Artemia spp. is deficient in some nutrients, such as poly unsaturated fatty acids and therefore is often enriched prior to feeding juveniles (Coutteau and Mourente, 1997). When hatchling P. leniusculus were reared using enriched live Artemia nauplii, high survival rates of up to 80% were achieved (González et al., 2008). After day 20, however, there was no significant difference in survival between P. leniusculus fed on a pellet diet and those fed live Artemia nauplii, although growth was significantly greater if live Artemia were fed up to day 50 (González et al., 2011). Following on from this research, live Artemia nauplii were replaced with Artemia cysts and fed to free-living hatchling P. lenisuculus, also resulting in high survival rates (81%) (González et al., 2009). Subsequently, a practical pellet feed, incorporating decapsulated Artemia cysts, was developed and high survival rates (86%) were achieved with hatchling P. leniusculus (Carral et al., 2011).
Despite the ecological importance of A. pallipes, and likely due to the previous lack of aquaculture of the species, commercially available diets have not been tested or developed for the species. This study presents results from a series of experiments testing different diet formulations on the growth and survival of hatchling and juvenile A. pallipes.
Materials and Methods
Study Animals and Site
The crayfish used within the experiments were captive-born juvenile A. pallipes, hatched from 21, wild-caught, ovigerous females (collected from a local river population in South Gloucestershire, England, under Natural England licence). The females were brought into an indoor, closed-circuit, aquaculture facility (in Somerset, England), two-months prior to the experiment commencing. They were removed once the juveniles had undergone two moults and were free-living. The experiments took place within the same aquaculture facility. This comprised 24 glass tanks (0.12 m2 bottom area; 45 L3 total tank volume), on a closed-circuit, recirculating system including a filtration sump, with a de-gassing chamber, filled with bio-balls and a fluidised sand bed. Water returned from the lidded tanks to the sump via 2 mm meshed tanks outlets (to prevent escape) and was fed back to the tanks from the sump via an ultraviolet filtration unit. Turnover rate was four times per hour and total system water volume was 1,200 L3. Water temperature varied between 12 and 18°C over the course of the experiments. The temperature range was controlled with coolers, to ensure there was a maximum temperature variation of <3°C, over a 24 h period. The photoperiod was ambient and averaged 12 h light and 12 h dark. Each experimental tank had a 30 mm substrate base layer of coral sand and fine gravel (0.4–1.0 mm diameter). Polycarbonate 10 mm sheeting, held down with substrate, provided refuges (two per crayfish), together with 15 mm internal diameter pipe for the larger animals. All tanks were gravel-siphoned weekly and 20% of the water was replaced with rainwater, collected within a water reservoir adjacent to the aquaculture facility. Water quality was measured weekly using a Colombo Testlab water testing kit (Aquadistri UK Ltd., Cambridgeshire, United Kingdom). Chemical levels remained consistent throughout the experiment: ammonia <0.1 mg/L, nitrite <0.1 mg/L, nitrate <15 mg/L, phosphate 0.2 mg/L, pH 7.8, calcium ≥35 mg/L, general hardness 10 KH, potassium hardness 8 KH and a level of dissolved oxygen ≥90%.
Experimental Procedure
Four feeding experiments (A–D) took place over two breeding seasons. Experiments A and B were with juvenile (20-day-old), hatchling A. pallipes with an initial mean carapace length (mm ± SD) of 5.3 ± 0.14 mm. All hatchlings had undergone two moults, having fully formed mouth parts and uropods. Experiments C and D used 60-day-old crayfish, with an initial mean carapace length of 7.9 ± 0.23 mm. For each experiment, the juvenile A. pallipes were randomly selected and put into different treatment tanks, with six replicates of each, at varying densities for each food treatment to be trialled (Table 1). All experiments ran for 40 days.
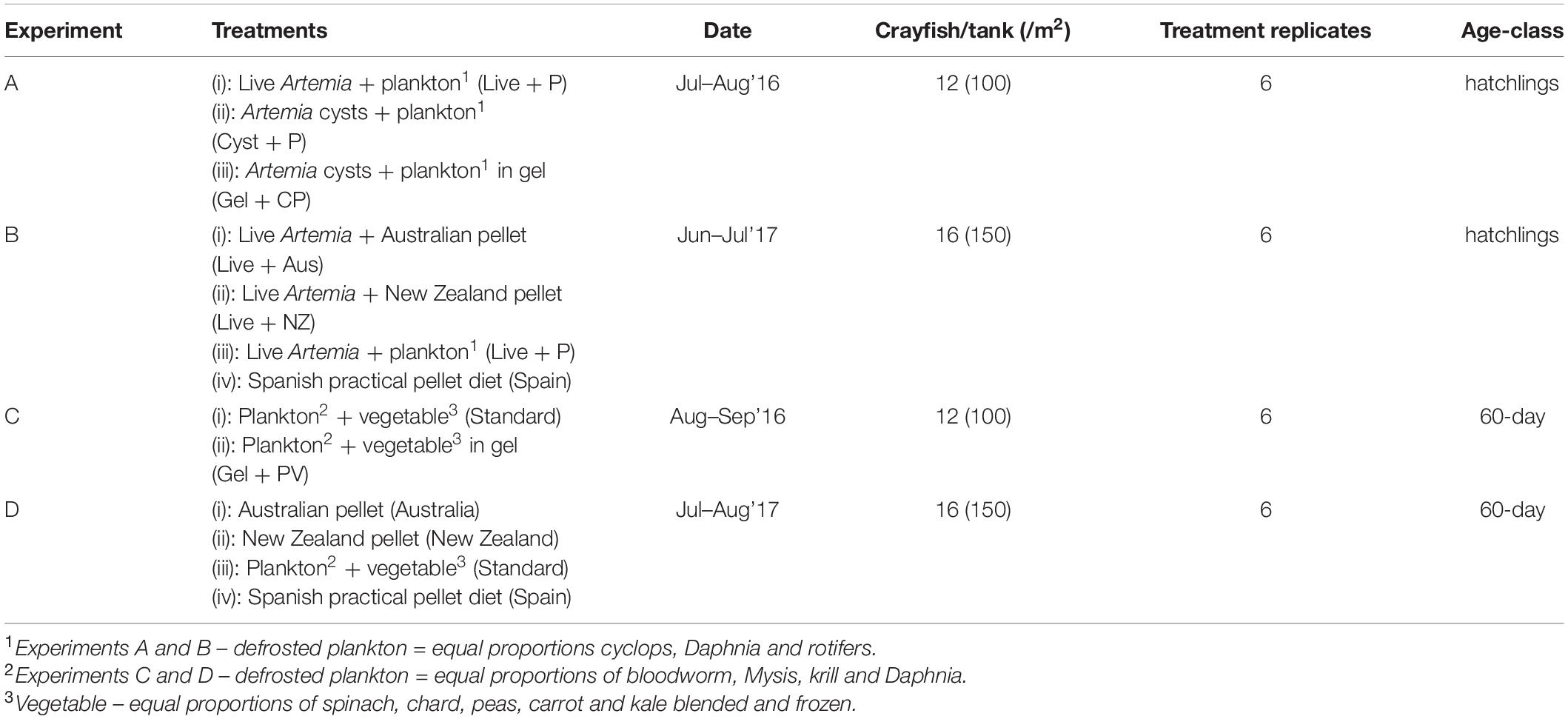
Table 1. Four feeding experiments A–D, on A. pallipes including dietary treatments, density equivalent, treatment replicates, duration of experiment, and age-class of animals.
Experiment C used juvenile crayfish that had not been used in experimental trials and had been fed an enriched diet of live Artemia fransicana nauplii and defrosted plankton since hatching. In contrast, experiment D used the same experimental crayfish that were in B as an extension of this experiment. Therefore, in experiment D, the live food component was removed after day-40 for treatments i, ii and iii, whereas treatment iv remained the same throughout both experiments B and D.
All diets, (except the pellet-only Spanish practical diet treatment, tested in experiments B and D), were enriched with 1 mL multivitamins, 1 g Spirulina, 1 mL lipids, plus 1 g of the carotenoid Astaxanthin, which was added to prevent the cray fishturning blue (Menasveta et al., 1993; Lorenz and Cysewski, 2000). As these enrichments were already included in the Spanish practical diet they were excluded. Additionally, the Spanish practical diet was not presented with live Artemia as the pellet already contained Artemia cysts. The defrosted plankton used in all four experiments was gamma-irradiated, prior to freezing. The A. fransicana nauplii were hatched from A. fransicana eggs and 24 h, post-hatching, were then gut-loaded with 0.1 mL Nannochloropsis spp., and enriched with highly unsaturated fatty acids (HUFA), for 8 h, prior to feeding to crayfish (Supplementary Table 2). The gel diet (for experiments A and C) was made by mixing 1.5 g of potato dextrose agar powder and 0.8 g of locust bean gum, which was added to help binding. To this mixture, 50 mL of water was added and then boiled for 1 min. The mixture was allowed to cool to 35°C before the other food items were added, to ensure that the mixture did not solidify but was cool enough to prevent the protein within the food items denaturing (Lepock, 1997). The mixture was then put in a fridge to set. For experiment A, we added 9 g of enriched plankton and 2 g of enriched decapsulated A. fransicana cysts to the gel. For experiment C, we added 8 g of enriched plankton and 3 g of vegetable mix (Supplementary Table 1), plus 1g of Spirulina. The pellet diets were commercially available crayfish-specific pellets (Supplementary Table 2). As these are commercially available diets not all content of the formulas was available. For experiments B and D, the crayfish the standard diets were made up at a ratio of three parts plankton to one part vegetable mix.
All crayfish were fed to excess, at a rate of approximately 4% of bodyweight of food per individual, presented at 18:00 daily. For hatchlings (experiments A and B), live A. fransicana or decapsulated A. fransicana cysts were fed at a rate of 500/crayfish/day; plankton, pellet or gel diets were fed at a rate of 0.01 g per day. For juveniles (experiments C and D), crayfish were fed at a rate of 0.02 g of food per animal per day. In both hatchling experiments, plankton or pellet was offered, in addition to the live food element, recognising that a combination diet may be important for crayfish growth and survival.
All four experiments were approved by the University of Bristol’s Animal Welfare and Ethical Review Board (AWERB) and the Bristol Zoological Society’s Conservation, Ethics and Sustainability Committee (CESC). Wild collection of animals was carried out under Natural England crayfish survey licence and an Environment Agency trapping licence. Crayfish were maintained in captivity under a Natural England scientific handling licence. The Crayfish Research Unit was inspected and certified as a hatchery facility by the Centre for the Environment, Fisheries and Aquaculture Science (Cefas).
Data Analysis
All crayfish were counted and individually measured on day-1 and day-40 (mm ± SD) of the experiments, to determine survival and growth. Growth was quantified by the carapace length, which was measured from the anterior edge of the rostrum to the posterior edge of the cephalothorax to the nearest 0.1 mm using Vernier callipers (Moore and Wright, Sheffield). Crayfish growth in each treatment group was calculated by subtracting the starting average carapace length from the final average carapace length.
To determine if there were any significant differences between the survival (% ± SD) with the different dietary treatments, data were examined using binomial generalized linear models (function glm, R package lme4) (Bates et al., 2015). To determine if there was any difference in growth among the treatments, data were examined with an ANOVA. Goodness-of-fit to normal distributions was checked by running the Shapiro - Wilk test on residuals (Shapiro and Wilk, 1965), prior to using an ANOVA. To determine if there were differences in growth among the treatments, data were log-transformed and examined with linear mixed models (function lmer, R package lme4) (Bates et al., 2015) or ANOVA if tested at tank level. The treatments were considered as fixed effects, and tanks were considered a random effect. The alpha level was set at p < 0.05. Only variables that had a significant effect were retained in models. Statistical analyses were performed using R 3.2.5 (R Development Core Team, 2006).
Results
Experiment A
Survival for those fed with the Live + P diet (91.7 ± 6.4%) was significantly higher than for those fed the Cyst + P (43.7 ± 23.2%) or Gel + CP (43.7 ± 8.9%) diets (z285 = 6.28, p < 0.001). There was no significant difference in crayfish survival between the Gel + CP and Cyst + P diet (z285 = 0.06, p = 0.95) (Figure 1).
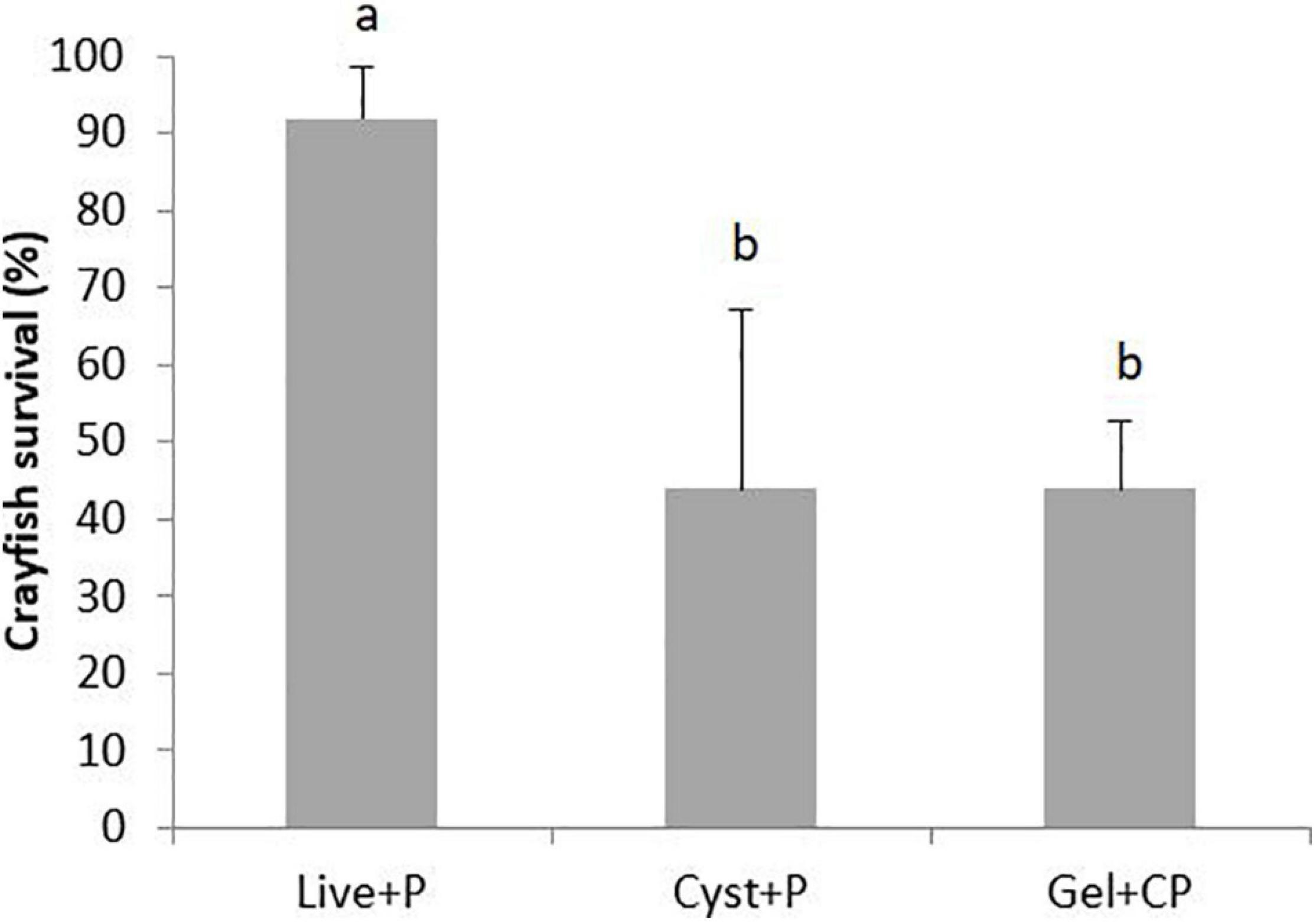
Figure 1. Percentage survival of hatchling A. palliipes, within the different treatment diets: live Artemia nauplii plus plankton (Live + P); Artemia cysts plus plankton (Cyst + P); or Artemia cysts plus plankton incorporated into agar gel (Gel + CP), at day-40. A different letter denotes significance between treatments. Error bars represent standard deviations.
Crayfish growth with the Live + P was significantly greater (2.4 ± 0.1 mm), than with the Cyst + P (2.0 ± 0.3 mm), or Gel + CP (2.0 ± 0.2 mm) diets (F2,169 = 14.94, p < 0.001). There was no significant difference in growth between the Gel + CP and Cyst + P diet (Figure 2).

Figure 2. Mean growth (final carapace length – start carapace length mm) of hatchling A. pallipes, within the different treatment diets: live Artemia nauplii plus plankton (Live + P); Artemia cysts plus plankton (Cyst + P); or Artemia cysts plus plankton incorporated into agar gel (Gel + CP), at day-40. A different letter above a treatment denotes significance. Error bars represent standard deviations.
Experiment B
Hatchling crayfish survival from day-1 to day-40 was significantly higher (z383 = 2.3, p = 0.02) with the Live + P diet (95.8 ± 5.1%) than with the Live + Aus (85.4 ± 12.3%) and Spain treatment diets (80.2 ± 6.1%) (z383 = −3.0, p = 0.002). Crayfish survival with the Live + NZ diet (91.7 ± 7.6%) was significantly higher than with the Spain diet (z383 = 5.5, p = 0.03) but was not significantly different from the Live + Aus diet (Figure 3).
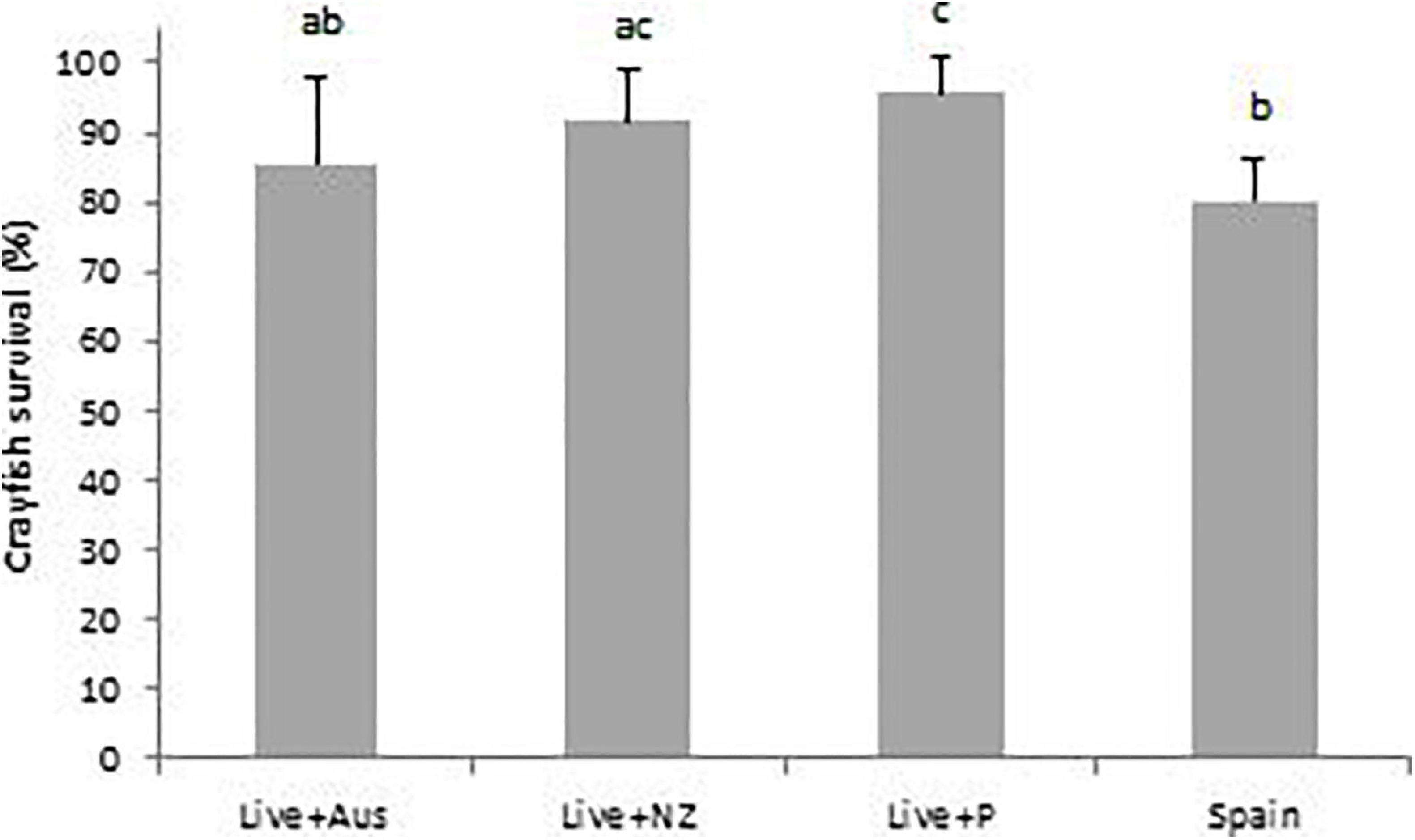
Figure 3. Percentage survival of hatchling A. pallipes for the four different treatment diets: Live Artemia nauplii plus Australian pellet (Live + Aus); Live Artemia nauplii plus plankton (Live + P); Live Artemia nauplii plus New Zealand pellet (Live + NZ) and the Spanish practical crayfish pellet diet (Spain), from day-1 to day-40. A different letter above a treatment denotes significance. Error bars represent standard deviations.
From day-1 to day-40, crayfish growth was significantly greater with the Live + P treatment diet (2.5 ± 0.1 mm) than with the Live + NZ and Spain diet (F3,335 = 7.1, p < 0.001). There was no significant difference between growth with the other three diet treatments (mean 2.24 ± 0.06 mm) (Figure 4).
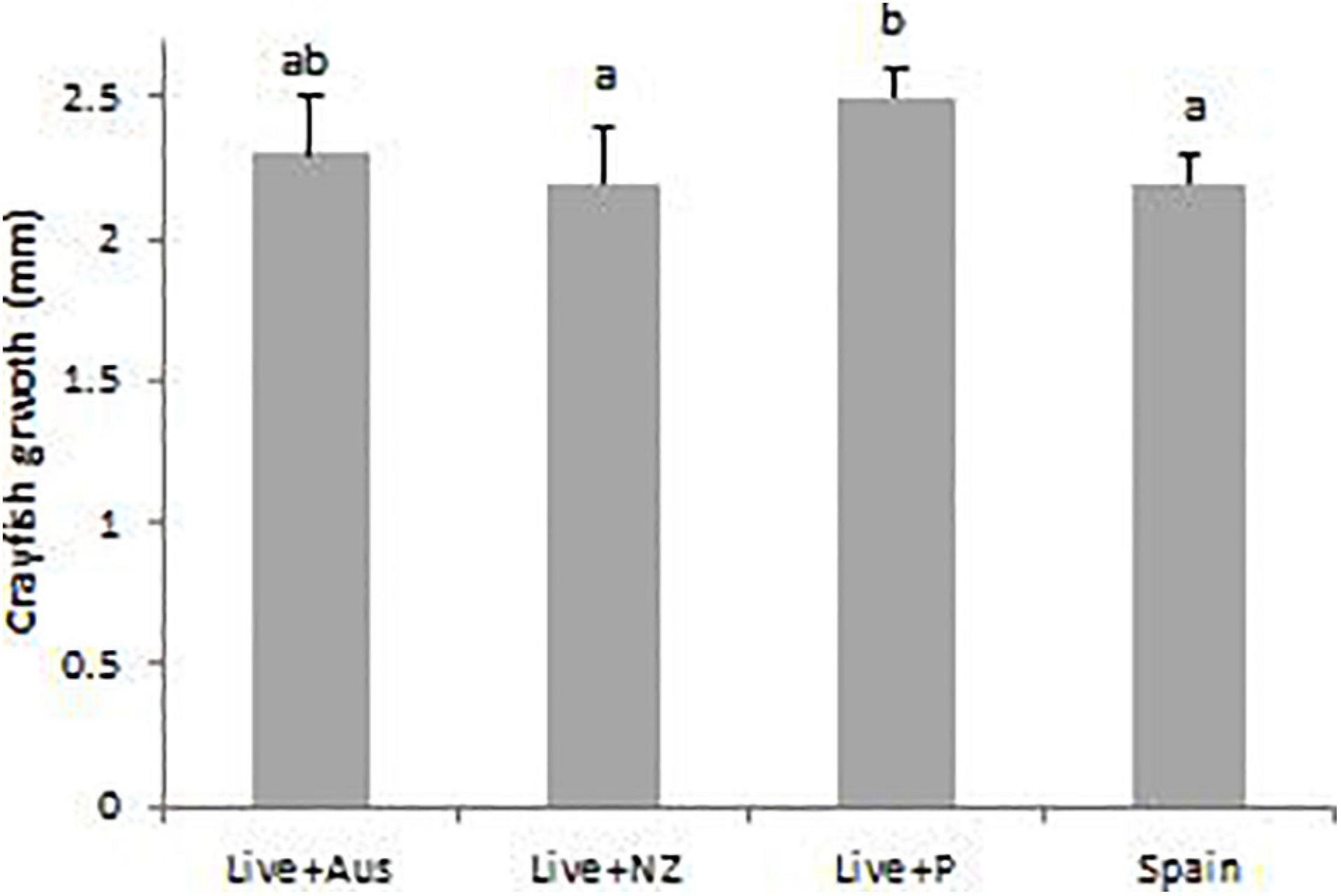
Figure 4. Mean growth (final CL – start CL mm) of hatchling A. pallipes for the four different treatment diets: Live Artemia nauplii plus Australian pellet (Live + Aus); Live Artemia nauplii plus plankton (Live + P); Live Artemia nauplii plus New Zealand pellet (Live + NZ) and the Spanish practical crayfish pellet diet (Spain), from day-1 to day-40. A different letter above a treatment denotes significance. Error bars represent standard deviations.
Experiment C
For the juvenile A. pallipes, there was no significant difference in crayfish survival between Standard (97.9 ± 5.9%) and Gel + PV diet (96.9 ± 6.2%) (t187 = 0.45, p = 0.65) (Figure 5).
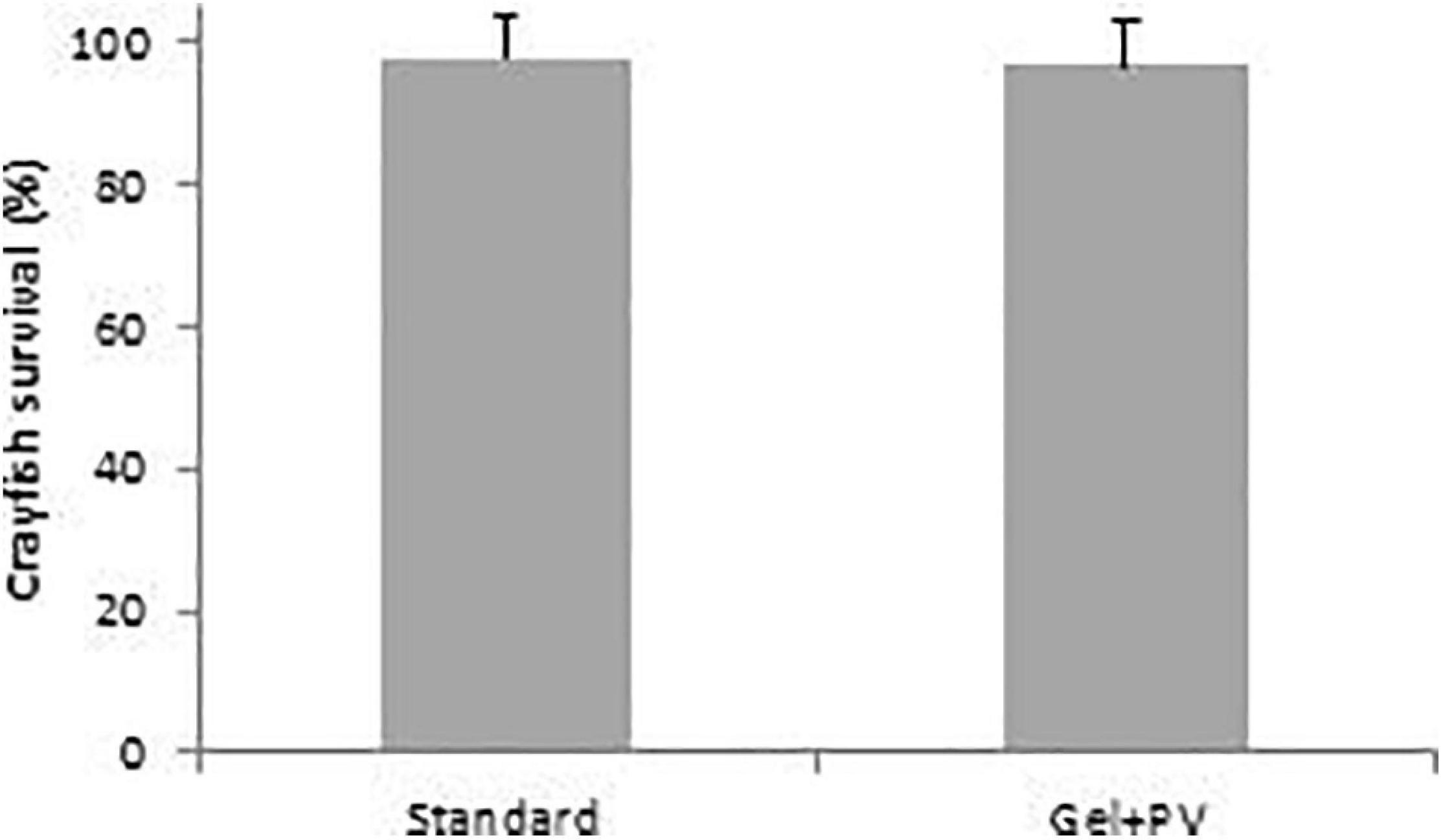
Figure 5. Percentage survival of juvenile A. pallipes, within the different treatment diet, plankton plus vegetable (Standard) or plankton plus vegetables incorporated into agar gel (Gel + PV), between day-1 to day-40. Error bars represent standard deviations.
Crayfish growth (mm ± SD) on the Standard diet (3.1 ± 0.3 mm) was significantly greater than those on the Gel + PV diet (2.4 ± 0.2 mm), (t187 = 4.38, p < 0.001) (Figure 6).
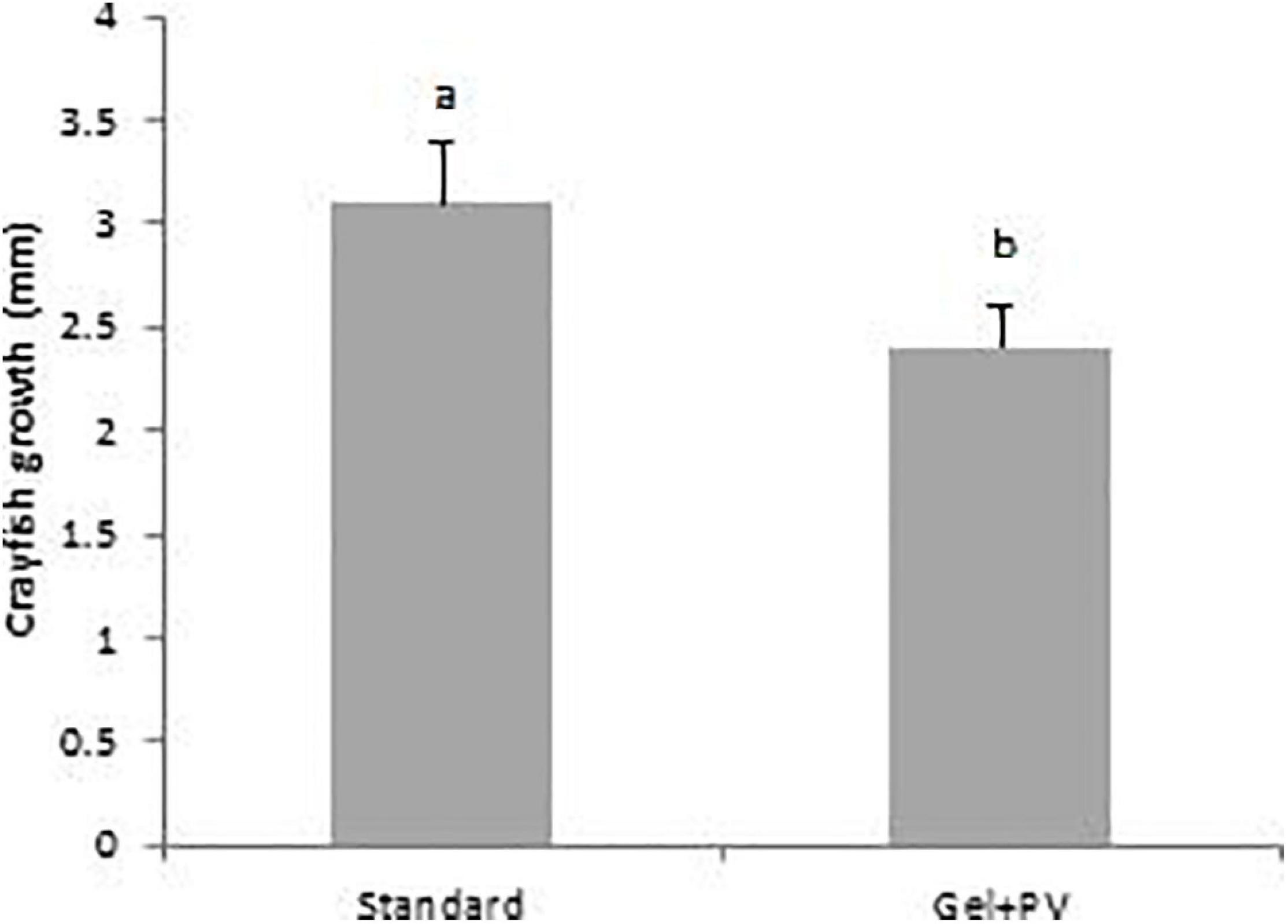
Figure 6. Mean growth (final CL – start CL mm) of juvenile A. pallipes, within the different treatment diets, plankton plus vegetable (Standard) or plankton plus vegetable incorporated into agar gel (Gel + PV), between day-1 to day-40. A different letter above a treatment denotes significance. Error bars represent standard deviations.
Experiment D
For the juvenile crayfish, survival from day-40 to day-80 was significantly lower in the New Zealand treatment diet (78.4 ± 10.3%) than all the other three treatments: Australia (97.4 ± 4.1%); (z383 = 3.2, p < 0.001), Standard (92.5 ± 11.6%); (z383 = 2.2, p = 0.03) and Spain (96.3 ± 4.0%); (z383 = 3.2, p < 0.001). Crayfish survival was not significantly different between the other three treatment diets (Figure 7).
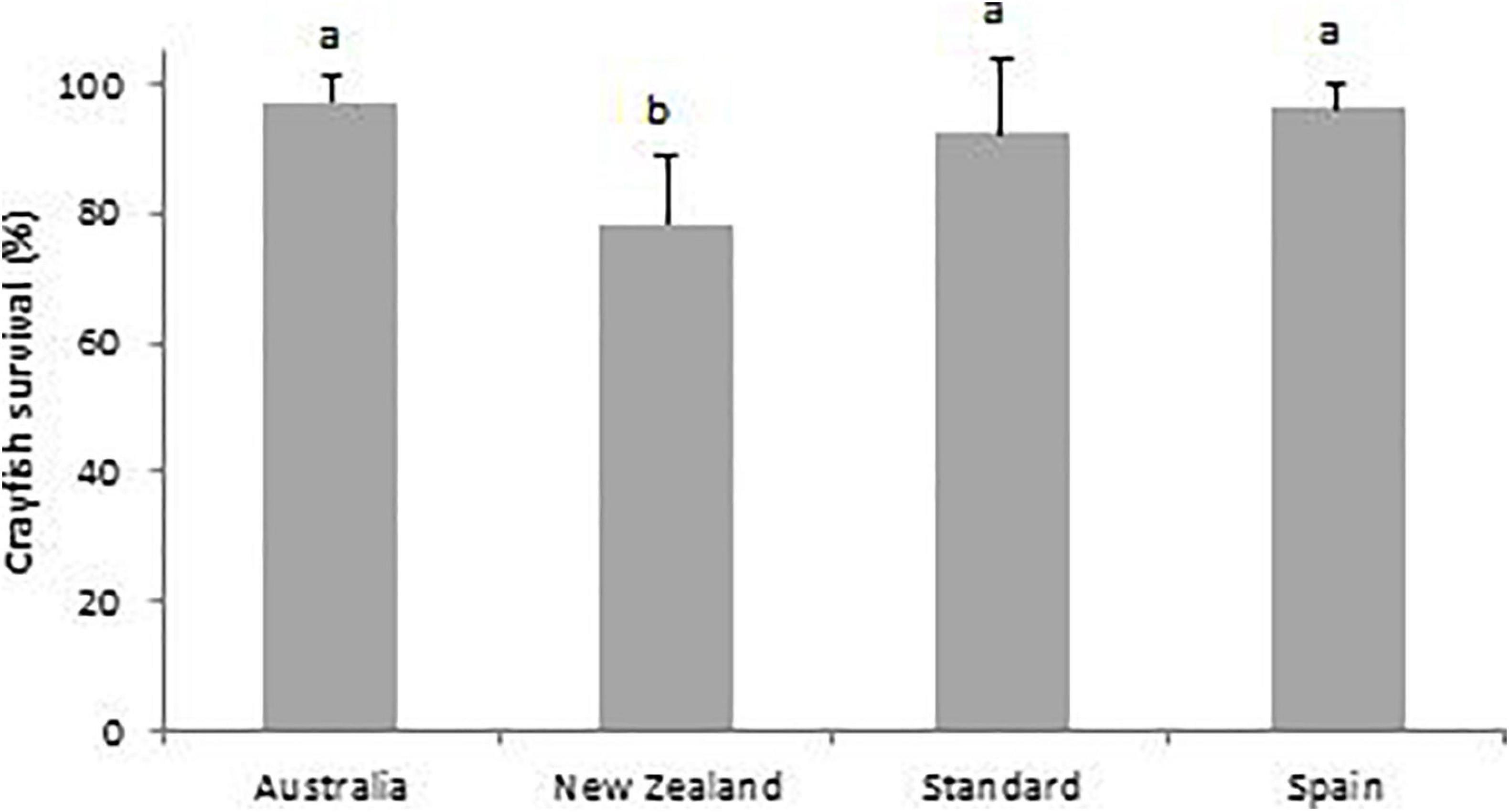
Figure 7. Percentage survival of juvenile A. pallipes for the four different treatment diets: Australian pellet (Australia); New Zealand pellet (New Zealand); plankton plus vegetable (Standard); and the Spanish practical crayfish diet (Spain); from day-40 to day-80. A different letter above a treatment denotes significance. Error bars represent standard deviations.
From day-40 to day-80, crayfish growth was significantly greater within the Standard treatment diet (2.5 ± 0.4 mm) than with all three other treatment diets: Australian (1.4 ± 0.3 mm, p < 0.001); New Zealand (2.0 ± 0.2 mm, p = 0.01); and Spanish diets (1.1 ± 0.3 mm, F3,20 = 28.52, p < 0.001). Crayfish growth with the New Zealand pellet was significantly greater than with the Australian pellet (p = 0.01) and Spanish practical diet (F3,20 = 28.52, p < 0.001) (Figure 8).
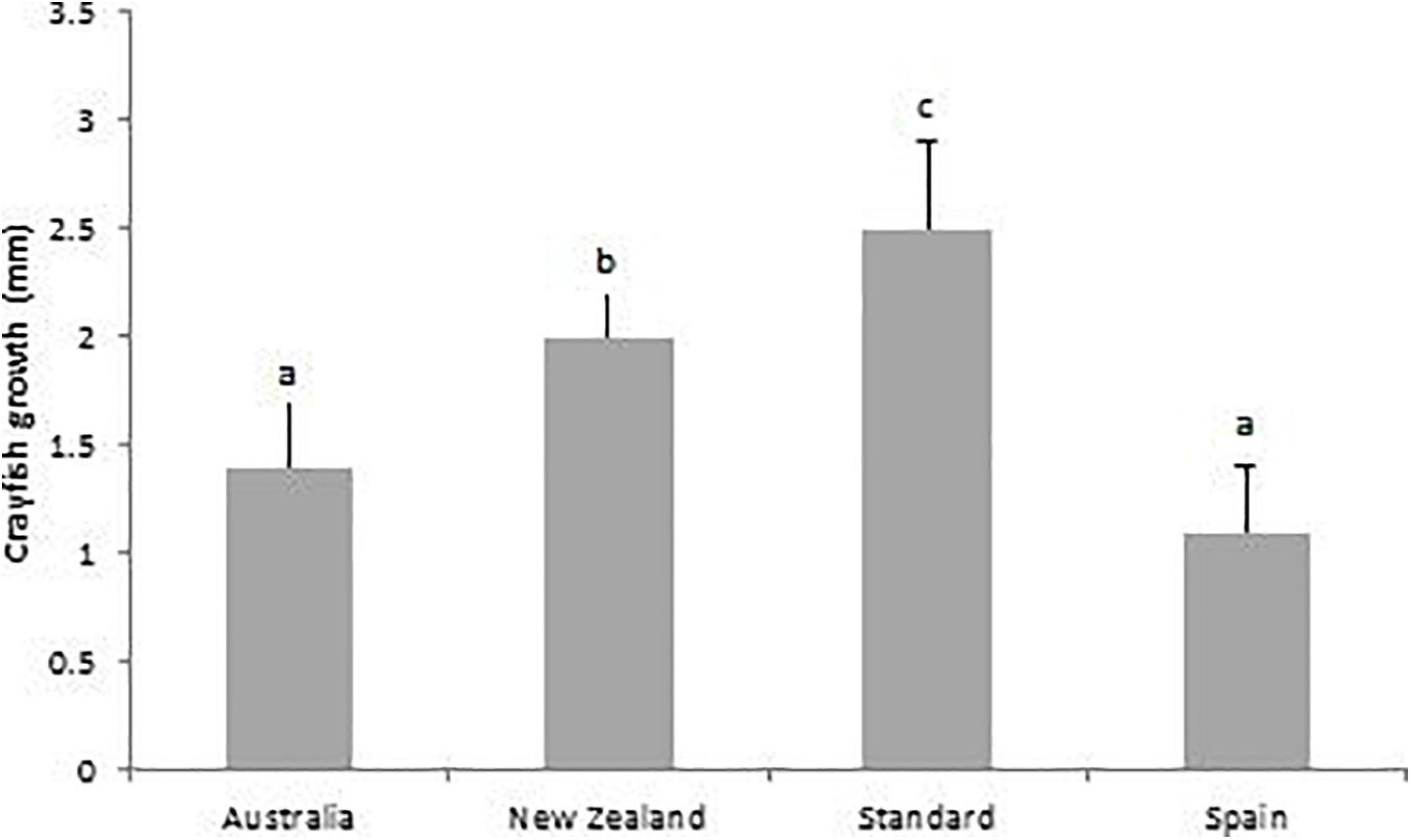
Figure 8. Mean growth (final CL – start CL mm) of juvenile A. pallipes for the four different treatment diets: Australian pellet (Australia); New Zealand pellet (New Zealand); plankton plus vegetables (Standard); and the Spanish practical crayfish diet (Spain), from day-40 to day-80. A different letter above a treatment denotes significance. Error bars represent standard deviations.
Discussion
Hatchling Survival
In both experiments with hatchling A. pallipes (A and B), consistently high survival rates (>85%) were achieved with crayfish fed with enriched live Artemia (with additional food sources). This is thought to be due to its nutritional components, having a high protein content (>50%) and levels of lipids and fatty acids (Treece, 2000). González et al. (2012), demonstrated that a level of 55% protein was optimal for survival and growth of hatchling P. leniusculus; with levels of over 33% required, to not compromise survival. However, the essential nutritional elements that hatchling crayfish require still remain unknown (González et al., 2011). During the first 4–6 weeks of life, the levels of digestive enzymes steadily increase as the hepatopancreas matures (Hammer et al., 2000). The proteolytic enzymes that Artemia nauplii contain may make digestion of dietary items easier for crayfish hatchlings and contribute to the maturation process of the hepatopancreas (Léger et al., 1986; Bengtson et al., 1991).
Lower survival rates occurred with Artemia cysts plus plankton (experiment A), than with live Artemia. This suggests that the nutritional content was not optimal and live Artemia nauplii are a more suitable first food item than Artemia cysts. This is potentially because prey motility is important for A. pallipes hatchlings with the movement of the nauplii stimulating them to feed. This is in contrast to a study on hatchling P. leniusculus, where no significant difference in survival between Artemia cysts and live Artemia nauplii was observed (González et al., 2009). When four age classes of yabby Cherax destructor were tested with live plankton versus pellet food, they spent 85% of their time feeding on live food and 15% feeding on the inert pellets (Meakin et al., 2008), demonstrating a preference for live food items. This was also observed in a study where juvenile hairy marron Cherax tenuimanus, when presented with both live Daphnia and pellet, showed a significant preference for feeding on the live food (Meakin et al., 2009). While observations on feeding preferences were not made as part of this study, a preference for live food items may be a result of their greater nutritional value. A stronger feeding response might have also been triggered by the presence of the live A. fransicana and therefore more food was ingested in total by the hatchlings in comparison to alternative diets.
The practical Spanish diet (experiment B), which contained Artemia cysts, within the pellet mix, achieved higher survival rates in hatchling crayfish than the rehydrated, decapsulated Artemia cysts, with or without agar gel. The higher survival rate in trials, which contained live elements, suggests there were some nutritional elements lacking from the non-live treatment diets, which could be due to the quality of the original food elements (such as the cysts) or the preparation. For example, in a study by Kouba et al. (2011), industrially decapsulated Artemia cysts produced lower growth and survival rates than freshly decapsulated cysts fed to six-month-old noble crayfish Astacus astacus.
Artemia cysts like Artemia nauplii also contain proteolytic enzymes. In a study by Garcıa-Ortega et al. (1998) the composition of both Artemia cysts and newly hatched Artemia nauplii was investigated. There was little difference in protease enzyme activity at the time of hatching; however, the enzyme levels increased as the nauplii developed. In our experiments, we fed the Artemia nauplii at 30 + hours post-hatching. When newly hatched, the Artemia nauplii lack exogenous mouth parts (Sorgeloos et al., 2001). We therefore grew on the nauplii until they had sufficiently developed mouth parts, to enable them to feed on the food items offered. From Garcıa-Ortega et al. (1998), protease activity was elevated and increasing up until the end of their study (7-hours post-hatching), suggesting that this proteolytic enzyme levels would increase with older nauplii, which could help to explain the increase in growth and survival that we experienced in our experiments.
Hatchling survival was lowest in the Spanish diet compared to the other treatment diets and survival was highest in the Live + P diet (experiment B). Due to the potential for increased food consumption in the presence of live feed, the exclusion of live Artemia when animals were fed the Spanish practical diet may have limited consumption in comparison to other pellet diets tested.
Hatchling Growth
Growth of the hatchlings (experiment A) was significantly greater when fed live Artemia nauplii plus plankton diet (Live + P) in comparison to decapsulated Artemia cysts plus plankton (Cyst + P). This was despite significantly lower numbers of crayfish within the cyst treatment groups during this time period due to mortalities. Previous studies have shown there is an increase in growth with a reduction in crayfish density (Nightingale et al., 2018), which was not observed in this case, and therefore suggests that diet was a limiting factor. Growth of the hatchlings (experiment B) was also significantly greater when fed live Artemia nauplii plus plankton diet (Live + P) than when they were fed the Live + NZ or Spanish diets.
The survival and growth of hatchling A. pallipes fed live food is consistent with previous studies on P. leniusculus (Sáez-Royuela et al., 2007) and C. destructor (Austin et al., 1997) where growth and survival of hatchling crayfish was significantly higher when fed live food rather than other treatment diets. The fact that hatchling survival and growth was improved with live food and plankton is supported by studies of A. pallipes analysing gut contents, which found that juvenile crayfish ingest a high proportion of invertebrates in their diet (Gherardi et al., 2004; Reynolds and O’Keeffe, 2005).
Juvenile Survival
There was no difference in survival between the treatments in experiment B, indicating that the critical life phase was over. However, in experiment D, crayfish fed the New Zealand pellet diet had significantly lower survival rates than the three other treatment diets, suggesting there may be some nutritional element lacking within this particular diet for A. pallipes juveniles. There are lower levels of protein in the New Zealand diet, compared to the other pellet feeds, which may result in lower survival rates. Mortalities were often observed during the moulting stage, where a lack of adequate nutrition can cause crayfish to die whilst moulting due to a lack of sufficient energy (Bowser and Rosemark, 1981).
Juvenile Growth
Crayfish growth was significantly higher in the Standard diet than in the Standard diet encapsulated in agar diet (experiment C), suggesting that the crayfish were not ingesting the same food items in the same quantities. In experiment D, crayfish growth in the Standard diet was significantly greater than in the all three other pellet treatment diets, suggesting that the crayfish were either eating more or better quality food items. In contrast to juvenile survival rates, the New Zealand pellet diet elicited better growth when compared to the Australian and Spanish diets; although this may have been due to cannibalism in the New Zealand pellet trial tanks, as this corresponded to a reduction in crayfish numbers within the tanks and not all the mortalities were accounted for. The crayfish may have been attacking moulting crayfish due to a nutritional deficiency within the diet (synchronised moulting was not occurring and therefore some crayfish were susceptible to cannibalism at this time). Alternatively, the reduced numbers of crayfish in the treatment tanks, due to the lower survival rate, caused an increase in growth of the remaining individuals (Savolainen et al., 2004; Nightingale et al., 2018).
The reduction in A. pallipes growth experienced with several of the treatment diets from day 40-80 is typical for this species. In a recent paper by Nightingale et al. (2018), a 240-day study of A. pallipes hatchlings, found growth was highest in the first 60-days (47.1% ± 6.6%). Growth then dropped to an average 14.1% ± 5.8% increase with every 60-day increment. This corresponds to other crayfish studies, which found that smaller crayfish grow faster than larger crayfish (Evans and Jussila, 1997; Jones and Ruscoe, 2000).
Agar Treatments
Crayfish graze periodically; therefore, if their food source is encapsulated within a gel, it should not degrade as quickly in water and should retain both its palatability and nutritional value for longer. This is supported by a study investigating survival and growth in juvenile (10 g) white yabby Cherax albidus. The crayfish were either fed fish and potatoes or this fresh food was encapsulated within pectin, alginate, agar or chitosan. There was a significant increase in growth when using the gel diets in comparison to a fresh food diet without gel (Coccia et al., 2010). However, this was not the case in our experiments (A and C). The agar may not be suitable as an early food item for A. pallipes hatchlings and juveniles, performing poorly in contrast to the plankton diets in both experiments A and C. Crayfish may be consuming too much agar and not enough of the other dietary, protein-based items at a life-stage where protein is an important dietary component.
Palatability
Studies on the palatability of diets have shown crustaceans will increase their feeding when particular stimulants are added to their diet (Harpaz et al., 1987; Hua et al., 2015). Crustacean species within aquaculture have preferences for specific textures and softness of food items (Cox and Johnston, 2003). In the case of both the agar-encapsulated (experiments A and C) and the pellet diets (experiments B and D), versus the plankton diets (experiments A–D), the plankton may be more palatable to the crayfish, in terms of both taste and texture, and therefore more is ingested, which increases growth.
Ontogenetic Dietary Changes
It has been suggested that as crayfish species mature they become predominantly detritivores (Paglianti and Gherardi, 2004); however, isotopic analysis of tissue suggests that aquatic invertebrates form a significant part of the diet throughout all age classes as found in koura Paranephrops zealandicus (Hollows et al., 2002) and P. leniusculus (Stenroth et al., 2006). A recent study investigated reproductive ability and growth in P. clarkii showed there was a significant increase in both fecundity and specific growth rate when zooplankton diets were offered rather than a commercial pellet feed. It indicates that the importance of zooplankton within crayfish diets at all age classes should not be underestimated (Sonsupharp and Dahms, 2017). This could explain why even in the older crayfish growth was still improved with the Standard diet, which was still predominantly consisting of plankton.
Conclusion
Enriched live diets are important for high survival and growth of A. pallipes from when the hatchlings are free-living, up until day-60, after which time the hepatopancreas and associated proteolytic enzymes are more mature. The hatchling may be using the proteolytic enzymes from the A. fransicanca to aid digestion at this early life stage. The enriched plankton diet consistently produced high rates of survival and growth in A. pallipes in both the hatchling and juvenile experiments (A–D), in comparison to all other diets offered, which suggests that nutritional quality and palatability is optimal. This is supported by studies of wild-caught A. pallipes juveniles, which consume a diet of predominantly aquatic invertebrates (Scalici and Gibertini, 2007). Therefore, it is possible that pellets do not give optimal nutrition in comparison to a more natural diet and the A. pallipes juveniles are showing a natural tendency to consume plankton rather than the artificial feeds offered. Evidence suggests that zooplankton may be important within crayfish diets at all age classes Hollows et al., 2002; Reynolds and O’Keeffe, 2005; Stenroth et al., 2006; Sonsupharp and Dahms, 2017). Therefore further research is required, to assess the long-term effects on growth, survival and fecundity of A. pallipes offered a solely pellet-based diet, rather than more natural food items.
The pellet diets used in this study were commercially available crayfish-specific pellets. As these are commercially produced diets, not all content and quantities of the formulas was available from the suppliers. However, the percentage values for proteins and lipids were provided for all diets; these are the components of the diets that have been discussed.
Pellet diets are a cheaper and more convenient option than producing bio-secure live food or gamma-irradiated frozen plankton. Where time and financial constraints are not an issue, feeding live Artemia plus enriched plankton for the first 40-days and then moving on to an enriched plankton diet is optimal for high survival and growth in A. pallipes. However, if A. pallipes are to be produced on a larger-scale, it may be more efficient to offer the practical crayfish pellet diet for all life-stages, but growth, survival and potentially fecundity may be compromised. All the diets trialled in these experiments have been specifically designed for particular species of freshwater crayfish, which may have different nutritional requirements, and therefore there may not be a specific diet that will work for all the species that are bred and reared. Therefore, for juvenile A. pallipes it is suggested that an enriched plankton and vegetable diet, with a live food element during the first weeks, will produce the best survival and growth.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by University of Bristol’s Animal Welfare and Ethical Review Board (AWERB) and the Bristol Zoological Society’s Conservation, Ethics and Sustainability Committee (CESC).
Author Contributions
JN designed and orchestrated the research and wrote up the findings. GM, PS, and GJ provided advice regarding the experimental design and analysis and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was funded by the Mohamed Bin Zayed Species Conservation Fund and Bristol Zoological Society.
Conflict of Interest
PS was employed by the company APEM Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Thanks to Jose Carral, John Hollows, and Damian Rigg for allowing your crayfish diets to be trialled in the experiments. Special thanks to Jonathan Williams and Timothy Clements for assisting with husbandry care, during the course of the four experiments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.607100/full#supplementary-material
References
Austin, C. M., Jones, P. L., Stagnitti, F., and Mitchell, B. D. (1997). Response of the yabby, cherax destructor clark, to natural and artificial diets: phenotypic variation in juvenile growth. Aquaculture 149, 39–46. doi: 10.1016/s0044-8486(96)01429-9
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using Lme4. J. Stat. Softw. 67, 1–48.
Bengtson, D. A., Léger, P., and Sorgeloos, P. (1991). Use of Artemia as a food source for aquaculture. in Artemia Biology, eds R. A. Browne, P. Sorgeloos, and C. N. A. Trotman. (Boca Raton, FL: CRC Press)11, 255–285. doi: 10.1201/9781351069892-11
Bowser, P. R., and Rosemark, R. (1981). Mortalities of cultured lobsters, Homarus, associated with a moult death syndrome. Aquaculture 23, 11–18. doi: 10.1016/0044-8486(81)90003-x
Carral, J. M., Gonzalez, A., Celada, J. D., Saez-Royuela, M., Garcia, V., and Gonazalez, R. (2011). Proposal of a practical diet for juvenile astacid crayfish studies from the onset of exogenous feeding under controlled conditions. Knowl. Manag. Aquat. Ecosyst. 401, 20–28. doi: 10.1051/kmae/2011034
Celada, J. D., Fuertes, J. B., Carral, J. M., Sáez-Royuela, M., and González, Á, and González-Rodríguez, Á (2012). Effects of vitamin C inclusion in practical diets on survival and growth of juvenile crayfish (Pacifastacus leniusculus Dana, Astacidae) from the onset of exogenous feeding. Aquac. Nutr. 19, 110-116.
Coccia, E., Santagata, G., Malinconico, M., Volpe, M. G., Di Stasio, M., and Paolucci, M. (2010). Cherax albidus juveniles fed polysaccharide-based pellets: rheological behaviour effect on growth. Freshw. Crayfish 17, 13–18.
Coutteau, P., and Mourente, G. (1997). Lipid classes and their content of n-3 highly unsaturated fatty acids (HUFA) in Artemia franciscana after hatching, HUFA-enrichment and subsequent starvation. Mar. Biol. 130, 81–91. doi: 10.1007/s002270050227
Cox, S., and Johnston, D. (2003). Feeding biology of spiny lobster larvae and implications for culture. Rev. Fish. Sci. 11, 89–106. doi: 10.1080/713610924
Evans, L., and Jussila, J. (1997). Freshwater crayfish growth under culture conditions: proposition for a standard reporting approach. J. World Aquac. Soc. 28, 11–19. doi: 10.1111/j.1749-7345.1997.tb00956.x
Figueiredo, M., and Anderson, A. (2003). Ontogenetic changes in digestive proteases and carbohydrases from the Australian freshwater crayfish, redclaw Cherax quadricarinatus (Crustacea, Decapoda, Parastacidae). Aquac. Res. 34, 1235–1239. doi: 10.1046/j.1365-2109.2003.00929.x
Garcıa-Ortega, A., Verreth, J. A. J., Coutteau, P., Segner, H., Huisman, E. A., and Sorgeloos, P. (1998). Biochemical and enzymatic characterization of decapsulated cysts and nauplii of the brine shrimp Artemia at different developmental stages. Aquaculture 161, 501–514. doi: 10.1016/s0044-8486(97)00297-4
Gherardi, F., Acquistapace, P., and Santini, G. (2004). Food selection in freshwater omnivores: a case study of crayfish Austropotamobius pallipes. Arch. Hydrobiol. 159, 357–376. doi: 10.1127/0003-9136/2004/0159-0357
González, A., Celada, J. D., González, R., García, V., Carral, J. M., and Sáez-Royuela, M. (2008). Artemia nauplii and two commercial replacements as dietary supplement for juvenile signal crayfish, Pacifastacus leniusculus (Astacidae), from the onset of exogenous feeding under controlled conditions. Aquaculture 281, 83–86. doi: 10.1016/j.aquaculture.2008.06.015
González, Á, Celada, J. D., Sáez-Royuela, M., González, R., Carral, J. M., and García, V. (2012). Response of juvenile astacid crayfish (Pacifastacus leniusculus) to three commercial dry diets with different protein levels during the first 6 months of intensive rearing. Aquac. Res. 43, 99–105. doi: 10.1111/j.1365-2109.2011.02808.x
González, R., Celada, J., Carral, J., García, V., Sáez-Royuela, M., and González, A. (2011). Intensive rearing of juvenile crayfish (Pacifastacus leniusculus, Astacidae) during the first 6 months: effects of size grading. Aquac. Res. 42, 1385–1392. doi: 10.1111/j.1365-2109.2010.02732.x
González, R., Celada, J., Carral, J., González, A., Sáez-Royuela, M., and García, V. (2009). Decapsulated Artemia cysts as dietary supplement for juvenile crayfish (Pacifastacus leniusculus, Astacidae) at different food supply frequencies from the onset of exogenous feeding under controlled conditions. Aquaculture 295, 200–204. doi: 10.1016/j.aquaculture.2009.07.009
Hammer, H. S., Bishop, C. D., and Watts, S. A. (2000). Activities of three digestive enzymes during development in the crayfish Procambarus clarkii (Decapoda). J. Crustac. Biol. 20, 614–620. doi: 10.1651/0278-0372(2000)020[0614:aotded]2.0.co;2
Harpaz, S., Kahan, D., Galun, R., and Moore, I. (1987). Response of freshwater prawn Macrobrachuim rosenbergii to chemical attractants. J. Chem. Ecol. 13, 1957–1965. doi: 10.1007/bf01014678
Hollows, J., Townsend, C. R., and Collier, K. (2002). Diet of the crayfish Paranephrops zealandicus in bush and pasture streams: insights from stable isotopes and stomach analysis. N. Z. J. Mar. Freshwater Res. 36, 129–142. doi: 10.1080/00288330.2002.9517076
Hua, X. M., Shui, C., He, Y. D., Xing, S. H., Yu, N., Zhu, Z. Y., et al. (2015). Effects of different feed stimulants on freshwater crayfish (Procambarus clarkii), fed diets with or without partial replacement of fish meal by biofeed. Aquac. Nutr. 21, 113–120. doi: 10.1111/anu.12148
Jones, C., and Ruscoe, I. (2000). Assessment of stocking size and density in the production of redclaw crayfish, Cherax quadricarinatus (von Martens) (Decapoda: Parastacidae), cultured under earthen pond conditions. Aquaculture 189, 63–71. doi: 10.1016/s0044-8486(00)00359-8
Kouba, A., Hamáčkov, J., Buřič, M., Policar, T., and Kozák, P. (2011). Use of three forms of decapsulated Artemia cysts as food for juvenile noble crayfish (Astacus astacus). Czech J. Anim. Sci. 3, 114–118. doi: 10.17221/1301-cjas
Léger, P. H., Bengtson, D., Simpson, K., and Sorgeloos, P. (1986). The use and nutritional value of Artemia as a food source. Oceanogr. Mar. Biol. Annu Rev. 24, 521–623.
Lepock, J. R. (1997). Protein denaturation during heat shock. Adv. Mol. Cell Biol. 19, 223–259. doi: 10.1016/s1569-2558(08)60079-x
Lorenz, T., and Cysewski, G. (2000). Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 4, 160–167. doi: 10.1016/s0167-7799(00)01433-5
Meakin, C., Qin, J., and Mair, G. (2008). Feeding behaviour, efficiency and food preference in yabbies Cherax destructor. Hydrobiologia 605, 29–35. doi: 10.1007/s10750-008-9297-0
Meakin, C., Qin, J., and Mair, G. (2009). Zooplankton predation and consumption by freshwater crayfish, Cherax tenuimanus, at different sizes and light conditions. J. Freshwater Ecol. 24, 159–167. doi: 10.1080/02705060.2009.9664276
Menasveta, P., Worawattanamateekul, W., Latscha, T., and Clark, J. (1993). Correction of black tiger prawn (Penaeus monodon Fabricius) colouration by Astaxanthin. Aquac. Eng. 12, 203–213. doi: 10.1016/0144-8609(93)90012-z
Nightingale, J., Stebbing, P., McCabe, G., and Jones, G. (2018). Determining an effective density regime for rearing juvenile Austropotamobius pallipes in a small-scale closed system hatchery. Aquac. Res. 49, 3055–3062. doi: 10.1111/are.13766
Nightingale, J., Stebbing, P., Sibley, P., Brown, O., Rushbrook, B., and Jones, G. (2017). The use of ark sites and associated conservation measures to secure the long-term survival of white-clawed crayfish in the UK. Int. Zoo Year J. 53, 50–68. doi: 10.1111/izy.12161
Paglianti, A., and Gherardi, F. (2004). Combined effects of temperature and diet on growth and survival of young of year crayfish: a comparison between indigenous and invasive species. J. Crustac. Biol. 24, 140–148. doi: 10.1651/c-2374
R Development Core Team (2006). R: A language And Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. ISBN 3-900051-07-0, URL http://www.R-project.org
Reynolds, J. D. (2002). “Growth and reproduction,” in In Biology and Freshwater Crayfish, ed. D. M. Holdich (Oxford: Blackwell Science), 152–191.
Reynolds, J. D., and O’Keeffe, C. (2005). Dietary patterns in stream-and lake-dwelling populations of Austropotamobius pallipes. Bull. Fr. Pêche Piscic. 376-377, 715–730. doi: 10.1051/kmae:2005028
Sáez-Royuela, M., Carral, J., Celada, J., and Pérez, J. (2001). Effects of shelter type and food supply frequency on survival and growth of stage-2 juvenile white-clawed crayfish (Austropotamobius pallipes Lereboullet) under laboratory conditions. Aquac. Int. 9, 489–497.
Sáez-Royuela, M., Carral, J., Celada, J., Pérez, J., and González, A. (2007). Live feed as a supplement from the onset of external feeding of juvenile signal crayfish (Pacifastacus leniusculus Dana Astacidae) under controlled conditions. Aquaculture 269, 321–327. doi: 10.1016/j.aquaculture.2007.04.053
Savolainen, R., Ruohonen, K., and Tulonen, J. (2004). Effect of stocking density on growth, survival and cheliped injuries of stage 2 juvenile signal crayfish Pacifastacus leniusculus Dana. Aquac. Res. 231, 237–248. doi: 10.1016/j.aquaculture.2003.09.045
Scalici, M., and Gibertini, G. (2007). Feeding habits of the crayfish Austropotamobius pallipes (Decapoda, Astacidae) in a brook in Latium (central Italy). Ital. J. Zool. 74, 157–168. doi: 10.1080/11250000701248688
Shapiro, S., and Wilk, M. (1965). An analysis of variance test for normality (complete samples). Biometrika 52, 591–611. doi: 10.1093/biomet/52.3-4.591
Sibley, P. J., Holdich, D. M., and Richman, N. (2011). “Monitoring the global status of crayfish, with particular reference to the white-clawed crayfish,” in Proceedings of the Species Survival: Securing White-Clawed Crayfish in a Changing Environment, Conference Held on 16th and 17th November 2010, (Bristol), 45–54.
Sonsupharp, S., and Dahms, H. U. (2017). Effect of frozen zooplankton feed on growth and reproductive performance of crayfish (Procambarus clarkii). Int. J. Agric. Technol. 13, 2317–2324.
Sorgeloos, P., Dhert, P., and Candreva, P. (2001). Use of the brine shrimp, Artemia spp., in marine fish larviculture. Aquaculture 200, 147–159. doi: 10.1016/s0044-8486(01)00698-6
Stenroth, P., Holmqvist, N., Nyström, P., Berglund, O., Larsson, P., and Granéli, W. (2006). Stable isotopes as an indicator of diet in omnivorous crayfish (Pacifastacus leniusculus): the influence of tissue, sample treatment, and season. Can. J. Fish. Aquat. Sci. 63, 821–831. doi: 10.1139/f05-265
Treece, G. (2000). Artemia production for marine larval fish culture. South. Reg. Aquac. Centre 702, 1–8. doi: 10.1007/978-1-4615-4911-6_1
Keywords: aquaculture, crayfish, conservation, nutrition, ecology
Citation: Nightingale J, Jones G, McCabe G and Stebbing P (2021) Effects of Different Diet Types on Growth and Survival of White-Clawed Crayfish Austropotamobius pallipes in Hatcheries. Front. Ecol. Evol. 9:607100. doi: 10.3389/fevo.2021.607100
Received: 16 September 2020; Accepted: 11 February 2021;
Published: 04 March 2021.
Edited by:
Ivana Maguire, University of Zagreb, CroatiaReviewed by:
Omid Safari, Ferdowsi University of Mashhad, IranJulian Reynolds, Trinity College Dublin, Ireland
Stephanie Peay, Aecom (United Kingdom), United Kingdom
Copyright © 2021 Nightingale, Jones, McCabe and Stebbing. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jen Nightingale, am5pZ2h0aW5nYWxlQGJyaXN0b2x6b28ub3JnLnVr