 W. Douglas Robinson
W. Douglas Robinson Tyler A. Hallman
Tyler A. Hallman Rebecca A. Hutchinson
Rebecca A. Hutchinson
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 10 February 2021
Sec. Conservation and Restoration Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fevo.2021.568278
This article is part of the Research Topic Benchmarking Biodiversity in an Era of Rapid Change View all 9 articles
The growth of biodiversity data sets generated by citizen scientists continues to accelerate. The availability of such data has greatly expanded the scale of questions researchers can address. Yet, error, bias, and noise continue to be serious concerns for analysts, particularly when data being contributed to these giant online data sets are difficult to verify. Counts of birds contributed to eBird, the world’s largest biodiversity online database, present a potentially useful resource for tracking trends over time and space in species’ abundances. We quantified counting accuracy in a sample of 1,406 eBird checklists by comparing numbers contributed by birders (N = 246) who visited a popular birding location in Oregon, USA, with numbers generated by a professional ornithologist engaged in a long-term study creating benchmark (reference) measurements of daily bird counts. We focused on waterbirds, which are easily visible at this site. We evaluated potential predictors of count differences, including characteristics of contributed checklists, of each species, and of time of day and year. Count differences were biased toward undercounts, with more than 75% of counts being below the daily benchmark value. Median count discrepancies were −29.1% (range: 0 to −42.8%; N = 20 species). Model sets revealed an important influence of each species’ reference count, which varied seasonally as waterbird numbers fluctuated, and of percent of species known to be present each day that were included on each checklist. That is, checklists indicating a more thorough survey of the species richness at the site also had, on average, smaller count differences. However, even on checklists with the most thorough species lists, counts were biased low and exceptionally variable in their accuracy. To improve utility of such bird count data, we suggest three strategies to pursue in the future. (1) Assess additional options for analytically determining how to select checklists that include less biased count data, as well as exploring options for correcting bias during the analysis stage. (2) Add options for users to provide additional information that helps analysts choose checklists, such as an option for users to tag checklists where they focused on obtaining accurate counts. (3) Explore opportunities to effectively calibrate citizen-science bird count data by establishing a formalized network of marquis sites where dedicated observers regularly contribute carefully collected benchmark data.
Contributions of volunteers to scientific databases are increasing as the popularity of citizen science continues to grow (Miller-Rushing et al., 2012; Chandler et al., 2017). Many citizen science projects are open-access and anyone can contribute observations without required training in best data collection practices (Cohn, 2008). eBird is an open online database with more than 560,000 users (eBirders) contributing millions of bird observations annually via checklists (Sullivan et al., 2009). Each checklist contains a list of bird species identified on a particular date and, ideally, counts of each species, as well as information on location visited, basic protocol used while birding (traveling, staying stationary, etc.), and duration of effort (Wood et al., 2011). The huge spatial extent of presence-absence data in eBird has facilitated efforts to model species distributions across continental and global spatial scales once data have been filtered to exclude potentially problematic checklists (Fink et al., 2013). The degree to which the count data may reliably inform scientific and management objectives remains unclear.
Although efforts to quantify issues associated with bird species detection have been studied and continue to be developed, both in citizen science databases and in structured scientific surveys (Buckland et al., 2008; Hutto, 2016; Walker and Taylor, 2017), less is known about potential counting errors and biases leading to noisy data. Counting birds is difficult, even by the most proficient observers (Robbins and Stallcup, 1981; Robinson et al., 2018). Methods to account for detection issues in bird counting studies continue to expand with development of new data collection and analytical methods (Buckland et al., 2008; Barker et al., 2018). Nearly all the methods, however, require a sophisticated sampling protocol that would exclude most volunteer birder contributions and therefore limit the advantages of gathering data at massive geographic scales. Yet, the potential windfall from large quantities of data can quickly be eroded if a lack of structured protocols leads to data quality concerns (Kelling et al., 2019). Given that abundance is one of the fundamental influences on population dynamics, functional roles in ecosystems, and even extinction risk (Brown, 1984), a better understanding of the potential value of count data contributed to massive online databases by untrained volunteers is needed (Greenwood, 2007). For example, species count errors in eBird data could limit our abilities to observe important abundance trends (Horns et al., 2018). Effective processes for evaluating and handling such errors need further development, owing to the potentially huge value of tracking population changes at continental and even global scales during this era of rapid environmental change (Bird et al., 2014; Fink et al., 2020).
Among the primary concerns are errors, bias and noise. Errors, for our purposes here, are differences in counts between a reference (benchmark) value and values included in eBird checklists for the same species on the same date. Errors are comprised of both bias and noise. Bias is the tendency for the errors to be consistently higher or lower than the reference value. Noise is the additional random counting error that increases variance of the counts. All three impede efforts to determine true count values, and are challenges common to many branches of biology (West, 1999; Guillery, 2002). We acknowledge that labeling such count differences as errors risks offending some eBird contributors. Errors, bias and noise all have objective statistical definitions. Our applications of the terms here are intended to improve understanding of the sources of variability in eBird count data. To acknowledge that there are sources of error in all measurements, however, we often refer to such deviations as count differences. We consider the terms “error” and “count differences” to be synonymous.
Robust comparisons of count differences are improved when data are collected in situations where detectability challenges are expected to be low. Such situations are rare but uniquely valuable. We used an extensive data set focused on benchmarking the richness and abundances of birds at a water treatment site in Oregon, USA. We compared count data gathered by a professional ornithologist focused specifically on creating an accurate benchmark measurement of daily fluctuations in waterbird counts with counts submitted by birders to eBird. We quantified the magnitude and directionality of count differences. Our data span 10 years and include 1,406 eBird checklists contributed by 246 observers, as well as 2,038 checklists in the benchmark data. The site is well suited for rigorous comparisons because all waterbirds are in the open, largely tolerant of human activity, and so provide a best-case scenario for detection, identification, and counting of birds. No adjustments for detectability or availability issues should be needed because all parts of the ponds are visible. Thus, discrepancies in counts between a professional observer focused on obtaining accurate numbers and data reported to eBird should be attributable to counting errors instead of availability and detectability issues. While there could be very minor detectability issues, like some diving waterbirds being under water briefly, the vast majority of error in this setting should be attributable to counting error.
We first quantified count differences then sought to understand potential factors explaining the magnitude and directionality of count differences. We hypothesized that count differences would be influenced by traits associated with the species being counted, with an index of observer experience (percent of species detected), and with seasonal changes in numbers of birds present. For example, we expected count differences might be slightly greater for diving ducks, which are sometimes briefly under water while foraging, and lower for dabbling species, which sit in the open continuously. We expected smaller count differences in checklists that included a higher proportion of the species present each day. We also hypothesized that count differences would be greater when overall total number of waterbirds present was high, potentially causing observers to be overwhelmed and therefore more prone to counting errors. Finally, we explored the possibility that, even if count data were biased on individual checklists, the waterbird community might be adequately characterized as a whole by combining count data from multiple observers and checklists. We conclude by proposing additional approaches that may reveal the extent to which citizen-science bird count data may be used to estimate abundances reliably.
Bird count data were gathered from 2010 to 2019 at the Philomath Wastewater Treatment facility, in Philomath, Oregon USA. The site contained two 35-ha ponds until 2011 when two additional 35-ha ponds were added. Each pond is rectangular and enclosed by a berm with a single-lane road. Birders circumnavigate the ponds typically by vehicle, rarely by walking or bicycling; WDR drove. Vegetation does not obscure the view at any pond. All shores are covered by large rocks (riprap). Birders circle all four ponds during a visit, very rarely restricting visits to fewer ponds. We found that the distribution of visit durations was unimodal (median = 60 min; Median Average Deviation (MAD) = 37; skew = 1.161; N = 1,646 checklists) suggesting that birders use similar methods while at the ponds.
We included 20 species we refer to as “waterbirds,” species that swim in the open while on the ponds and should be easily seen (Table 1). The species are primarily ducks and geese, but also include grebes, American Coot (Fulica americana), and gulls. These are species birders identify by sight, not by sound. We excluded species that occurred primarily as fly-overs, such as Cackling Goose (Branta hutchinsi), species whose counts rarely exceeded two per day, and species whose numbers varied strongly within a day. The number of waterbirds present at the site varied seasonally from a few dozen during mid-summer (June) to 5,000 or more during fall migration (October-November).
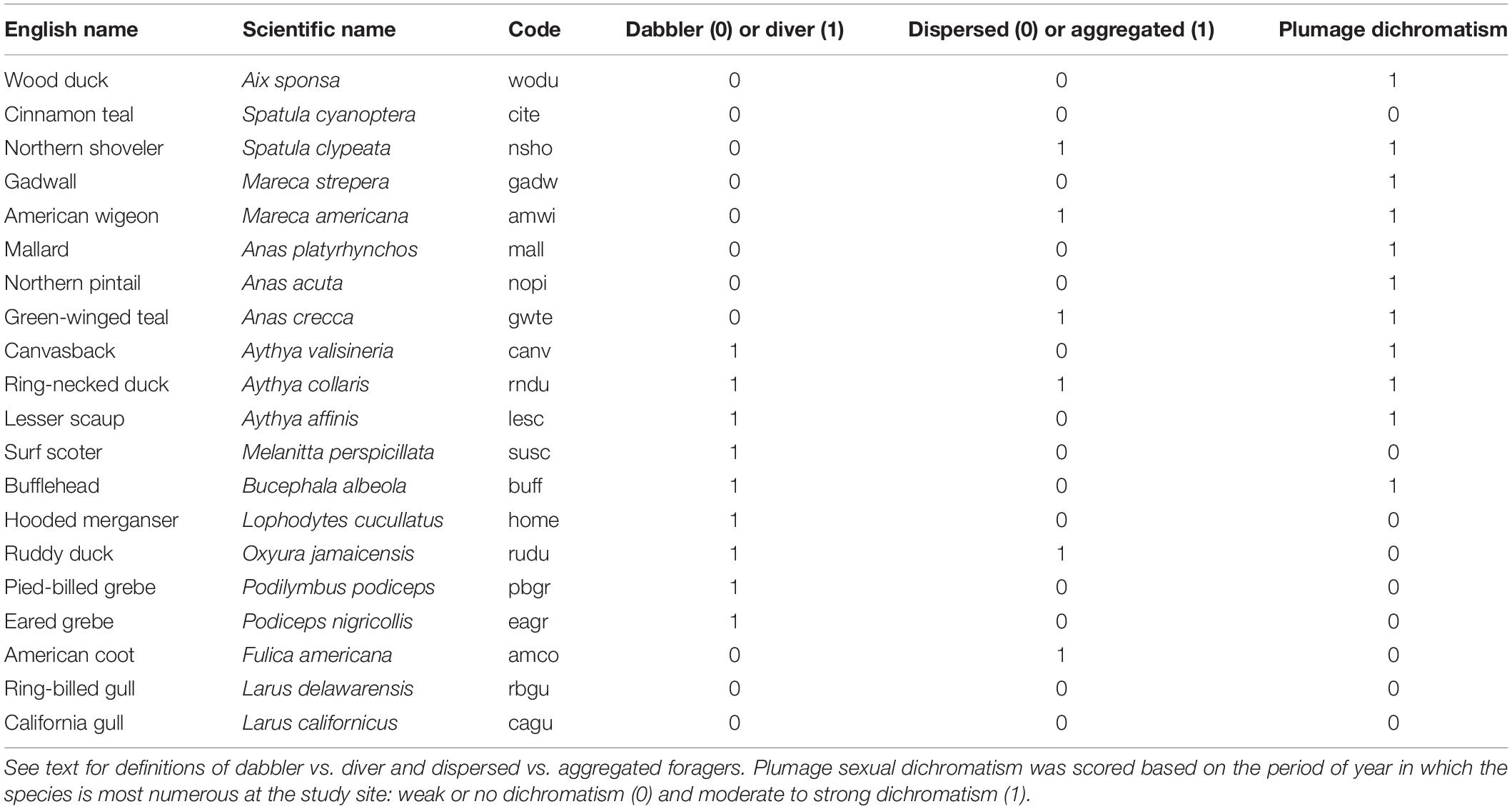
Table 1 | Twenty species were included in the study. Scientific names, sequence, and short-hand codes follow American Ornithological Society (http://checklist.aou.org/taxa).
All birds of all species were counted during each site visit by WDR. We call these our benchmark counts (R∗) and they serve as the reference values against which all other count data are compared. Waterbird counts were made to plus or minus one individual except for Northern Shoveler (Spatula clypeata), which were plus or minus 10 because they forage in constantly moving dense aggregations rendering more precise counts problematic, and Bufflehead (Bucephala albeola), which were counted to plus or minus 5 because they dive so frequently while foraging in the early morning period surveyed by WDR that more accurate counts were difficult. Counts were tallied separately for each species on each pond then aggregated later. On average, except for shovelers and coots, the two most numerous species at the site, the number of individuals of the remaining 18 species was less than 15 individuals per pond on 90% of dates. Numbers of several species were greater for 5 weeks in fall but the same method of individually counting birds was employed. In the time frame of the daily counts, movements between ponds were normally minimal. Duration of counting time was recorded separately for each pond.
On some days (N = 84), WDR counted birds more than once. These second-visit data, which we call Ref2 counts, were also complete counts of the study species and averaged 13% shorter in duration. Ref2 counts were used to characterize within-day variability in numbers across the 10-year study period. We consider them to provide a conservative estimate of variability in R∗ counting accuracy because they were largely conducted on days with exceptional levels of migratory movements. Thus, they estimate a probable upper bound on the expected amount of within-day variability in waterbird numbers and R∗ count accuracy (averaging 0 to −8% across the 20 species). We also used these Ref2 data to evaluate time-of-day effects when comparing WDR counts with data from the ten observers contributing the most study site data to eBird, because eBirders tended to count birds later in the day than did WDR. The times of day eBird checklists were initiated as well as the difference in start times of eBird and benchmark checklists were unimportant in predicting percent error in our across-species and species-specific model sets. Therefore, we concluded that comparisons of count differences between R∗ and eBird checklists were appropriate and that possible time-of-day effects could be ignored.
Our R∗ counts are from one expert observer. R∗ counts are not without error. Aside from comparisons with Ref2 counts made by the same observer, our data lack contributions from other experts as independent quantifications of accuracy and potential error of R∗ counts. To estimate the error in R∗ counts, we compared counts made in the field with counts in photos taken within 2 min of count completion. Comparisons were made in November and December, 2020, and included a range of count values from 1 to 1,050 for 17 of our 20 waterbird species (2 gulls and the scoter are not normally detected in November and December).
We downloaded eBird checklists from the Philomath Sewage Ponds eBird hotspot as well as eBirder personal locations within 1 km from 2010 to 2019. Only data obviously restricted to the ponds were included. No other waterbird sites are present within 4 km of the site. Most eBirders used the pre-established hotspot as the checklist location but some created new personal locations each time. We included eBird checklists following the stationary, traveling, and area protocols. We removed checklists with greater than ten observers or durations of over 5 h. We included only complete checklists with all birds reported and removed any checklists where observers reported no waterbirds. From each complete eBird checklist, we collected data on date, start time, observer, duration of count, identity of waterbird species reported (to allow calculation of percent richness; see below), and count data for our 20 focal species. When species were recorded as present but not counted (X noted instead of a number), those data were excluded because no count difference could be calculated.
We restricted our comparisons to dates where WDR counted birds and at least one eBird checklist was contributed on the same day (N = 767 dates). Our questions were about counting differences and not detection rates of rare species, so we further restricted our comparisons to counts of greater than three for each species detected on WDR’s first visit (R∗). We calculated the Count Difference for each species by subtracting R∗ from eBird counts on each checklist. Count differences were positive when eBird checklists reported higher numbers than R∗ or negative when eBird checklists reported fewer birds than R∗. Numeric values of count differences spanned three orders of magnitude, so we focus on reporting Percent Error, which we calculated by converting each difference to a proportion of R∗.
To evaluate factors hypothesized to be associated with percent error, we included variables associated with species, checklists, time of year and observer experience. Species characteristics included categorization as dabbler vs. diver, degree to which species form dense aggregations, and the degree of sexual dimorphism. Checklist characteristics included start time, duration and number of observers. Time-of-year characteristics were associated with daily numbers of waterbirds (R∗, Ref2 and their sums for all 20 species) and waterbird species richness present at the study site [measured as the richness detected by the professional (proRichness) as well as the aggregate of species listed in eBird checklists and proRichness]. Because observer experience at the site might also influence counting accuracy, we compared data from the 10 observers who contributed the most checklists with the R∗ and Ref2 benchmark data. Additional details on each variable are explained below.
To explore patterns of species-specific variability in count data, we created categorical variables for species traits that might impact counts (Table 1). We categorized birds as dabblers vs. divers. Dabblers were any species that foraged primarily by swimming on the surface of the water, which included gulls, American Coot, and Aix, Anas, Mareca, and Spatula ducks. Divers foraged below water regularly and included scoters, grebes, and Aythya and Bucephala ducks.
We also included an index of spatial aggregation on the ponds. Some species, for example Northern Shoveler, often forage in densely packed groups, creating challenging circumstances to accurately count birds, while other species forage singly or as spatially-distanced groups where enumeration should be much easier. The aggregation index was simply a subjective binary classification (0 for foraging alone or in loose aggregations vs. 1 for foraging in aggregations that might render counting difficult) based on our years of experience at the site.
The degree of plumage dimorphism and similarity to other species could influence error and bias in counts because of species misidentification. We categorized species as those with weak or no obvious plumage dichromatism during most of the period of time when each species was present (e.g., geese, coots) vs. strong dichromatism (males and females distinctly visually different).
To evaluate the possibility that species identification of similar species might influence count differences, we used another subjective binary category called “Doppelganger;” 1 indicated the species co-occurred with a similar species whereas 0 indicated the species was unique in appearance and unlikely to be confused with other species. The categorization may vary seasonally, especially in late summer when many waterbirds molt to eclipse plumage. Because total waterbird numbers were low during late summer, we utilized one value for each species.
Daily start time among eBird checklists was highly variable, covering all daylight hours. The mean start time was 4 h later than the mean start time for WDR visits. Although we only compared counts conducted on the same day, we wanted to evaluate potential effects of time-of-day and temporal lag between the eBird checklist counts and R∗. To do so, we converted checklist start time to minutes since midnight then calculated the difference in start time between eBird checklists and WDR first visits.
Because our Ref2 counts occurred later in the day when more eBird checklists were initiated, we included Ref2 as an “additional observer” in some comparisons to provide an important check on within-day variability in counts as a possible explanation for count differences between R∗ and eBird checklists. Because Ref2 counts were generated on days with high levels of migratory movement, we consider the count differences between R∗ and Ref2 to represent an upper bound on expected levels of within-day variability in waterbird numbers.
Additional factors associated with each checklist could influence count differences. We reasoned that duration of time spent at the site should be positively related to count accuracy. All complete eBird checklists are required to have a measurement of event duration.
Number of observers might also influence counting accuracy, so we included the reported number of observers for each eBird checklist. The R∗ and Ref2 counts were made when WDR was alone more than 99% of all dates.
Date influences the number of species present as well as the abundances of each species. Both richness and abundance could influence counting accuracy so we included day of year in our models. Because we hypothesized that total number of all waterbirds combined may influence counting accuracy, we included R∗ counts of all 20 study species and the combined daily total of all waterbirds in our model sets. In that way, we established the baseline numbers of waterbirds known to be present as a function of date. In calculating total waterbird abundance, we used data limited to the 20 study species and excluded a subset of species known to have high daily variability in counts, such as geese, which occurred primarily as fly-overs. The other species excluded from our focal group of 20 species were numerically rare. Further, to determine if percent error was influenced by the number of each particular species as opposed to overall waterbird abundance, we included R∗ of each relevant species in our model sets.
We hypothesized overall waterbird species richness present at the site on a given date may influence counting accuracy. A higher number of species to identify could reduce focus for achieving accurate counts, particularly for the more regularly-occurring and common species (e.g., Mallards, Northern Shovelers). Therefore, we included in our models the total waterbird richness detected by WDR each day. Our analyses indicated that richness observed by WDR and total waterbird richness detected by all eBird contributors were highly correlated. We calculated daily Percent Richness based on the 35 possible waterbird species at the site and included that richness in our models (see Supplementary Text for a list of species). The other 15 species that formed our set of 35 waterbird species included: Snow Goose (Anser caerulescens), Greater White-fronted Goose (Anser albifrons), Cackling Goose (Branta hutchinsii), Canada Goose (Branta canadensis), Blue-winged Teal (Spatula discors), Eurasian Wigeon (Mareca penelope), Redhead (Aythya americana), Tufted Duck (Aythya fuligula), Greater Scaup (Aythya marila), White-winged Scoter (Melanitta deglandi), Black Scoter (Melanitta americana), Long-tailed Duck (Clangula hyemalis), Common Goldeneye (Bucephala clangula), Barrow’s Goldeneye (Bucephala islandica), and Common Merganser (Mergus merganser).
Observer experience at the site could also be influential, so we compared percent error in counts from the ten observers contributing the most eBird checklists at our study site with the R∗ and Ref2 counts.
We used the “lmer” package in R (R Core Team, 2020) to run mixed-effects models. Our overarching goal was to identify factors informative for explaining variation in Percent Error, our dependent variable in all models. We included observer ID and species as random effects to account for observer- and species-specific error when appropriate. We included four categorical species characteristics as fixed effects in our model sets: Dabbler or Diver; Sexually Dichromatic or not; Doppelganger or not; and Aggregated or not. Five checklist-related characteristics were included as fixed effects: start time (minutes since midnight), difference in start time between WDR’s first count of a day and each eBird checklist, duration (minutes), number of observers, and day of year. Four fixed-effects related to time-of-year were also included: R∗ (WDR’s reference count of each species, which varied seasonally), waterbird abundance (aggregated across all species), total waterbird species richness and percent richness, our index of observer skill at species identification. We included models with the quadratic effects of species-specific abundance, waterbird abundance, waterbird richness, duration, number of observers, day of year, and percent richness to examine potential non-linear shapes of their effects.
Before running mixed effects models, we scaled and centered all numeric variables. We assessed model performance through BIC and propagated best-performing shapes for each variable to multi-variable models. We used a forward stepwise approach and added additional potentially influential variables to the best-performing model until a stable (i.e., model remained the top model after the inclusion of additional variables) top-performing BIC model was identified.
Although count difference was normally distributed, percent error was not. Non-detections of species that were detected by WDR (eBird counts of zero) equal negative 100 percent error. Non-detections caused a bimodal distribution of percent error with a second peak at negative 100 percent. We removed non-detections to create a unimodal distribution of percent error. When non-detections were removed, percent error was heavily right-skewed due to the high number of negative percent errors and the few very large positive percent errors. To adjust skew, we added a constant to make all values positive and log (base 10) transformed percent error. In addition to adjusting skew, removal of non-detections improved the focus of our analyses on count errors, reducing chances that inclusion of zero counts of species might actually be species detection or identification problems instead of counting errors. Our restriction of counting error analyses to species detected in numbers of 3 or greater probably limited most effects of zero counts. In this paper we focus on analyses of data excluding non-detections but report some analyses in Supplementary Materials to show the effects of including non-detections (zero counts) on results. It is possible that an unknown number of zero counts were a result of reporting errors (data entry mistakes), but we assume this type of error is relatively rare.
To understand the (in)consistency of variables influencing species-specific percent error, we ran standardized linear model sets of the effects of the explanatory variables described above on transformed percent error for each species. As above, we included models with quadratic effects of species abundance, waterbird abundance, waterbird richness, duration, number of observers, day of year, and percent richness. As each model set was species-specific, we excluded variables of species characteristics from these model sets. We included observer ID as an explanatory variable to examine its comparative influence. In these standardized model sets, we included separate models of the main effect of each variable and propagated the best shape for each variable into more complex models. Since start time and difference in start time were highly correlated, we use the top-performing of the two in subsequent models. We used a forward step-wise approach to determine the top-performing model of checklist covariates. We then ran models with pairs of all non-checklist explanatory variables with and without the variables in the top checklist covariate model. We used BIC to compare model performance and select top models.
To compare the overall communities described in eBird checklists, we conducted ordination in species space with NMDS on count data. We grouped checklists by observers to simplify the analysis. To visualize differences in community characterization, we chose to contrast January and October because January represents a time of year when waterbird migration is minimal and so daily numbers are relatively stable, whereas migration is at its peak during October, so richness is high and volatility in numbers can be high. To evaluate how characterization of waterbird abundance at these times varied with respect to eBirder checklists, we first removed all checklists that included an “X” for the count of any of our 20 study species. We then calculated the mean and median values of species counts across checklists for each observer during each month. To evaluate the idea that group collective contributions of multiple eBird checklists might characterize the waterbird community more similarly to R∗, we calculated mean counts of species across observers in January and October to create combined count values, which we call the Borg number (). We similarly aggregated WDR’s first-visit species counts as a Reference community. To ensure that our NMDS positions in species space were not driven overwhelmingly by an eBirder with the largest number of checklists, we reran the NMDS without checklists from the top-contributing observer included in . We used two dimensions and a maximum of 20 iterations to run NMDS with the “vegan” package in R (version 3.6.1).
We compared benchmark counts of waterbirds (R∗) and at least one eBirder on 672 dates, representing a total of 1,406 comparisons (checklists). eBird checklist contributions varied seasonally with lows during winter and summer and highs during migration periods (Supplementary Figure 1). Our analyses included 246 different eBirders who contributed from 1 to 321 checklists.
Comparisons of R∗ counts with photographic evidence indicated a mean percent error across 17 species of −0.4% (SD = 2.1%; N = 222 comparisons) indicating that R∗ counts were lower, on average, than numbers revealed in photos. The median differences varied from 0% for multiple species with counts below 200 to −1.2% for Northern Shoveler. We assume temporal consistency in counting errors for the duration of the study because the R∗ count data were gathered by the same observer using the same methods. Another estimate of R∗ count errors can be inferred from comparisons with Ref2 counts, which averaged −8%. Ref2 counts occurred throughout the 10-year duration of the study.
Across all twenty species, 76 percent of all counts were less than R∗ (Figure 1 and Supplementary Figure 2), indicating that count data in the eBird checklists regularly contained apparent counting errors. eBird checklists with species non-detections excluded (that is, no counts of zero included, even if the species was known to be present that day) had counts below R∗ values by a median of 29.1% but count differences were quite variable across species (Figure 1A), with median absolute deviations of percent error averaging 44.6% (Supplementary Table 1). At the extremes, count differences across waterbird species ranged from negative 99% for severe under-counts to more than 3,788% too large. In real numbers, count differences ranged from being too low by 1,443 to too high by 1,048 (both for Northern Shoveler; Figure 1B). Median percent error was negative, indicative of undercounting, for all waterbird species except the uncommon Surf Scoter (0%; R∗ was at most 11).
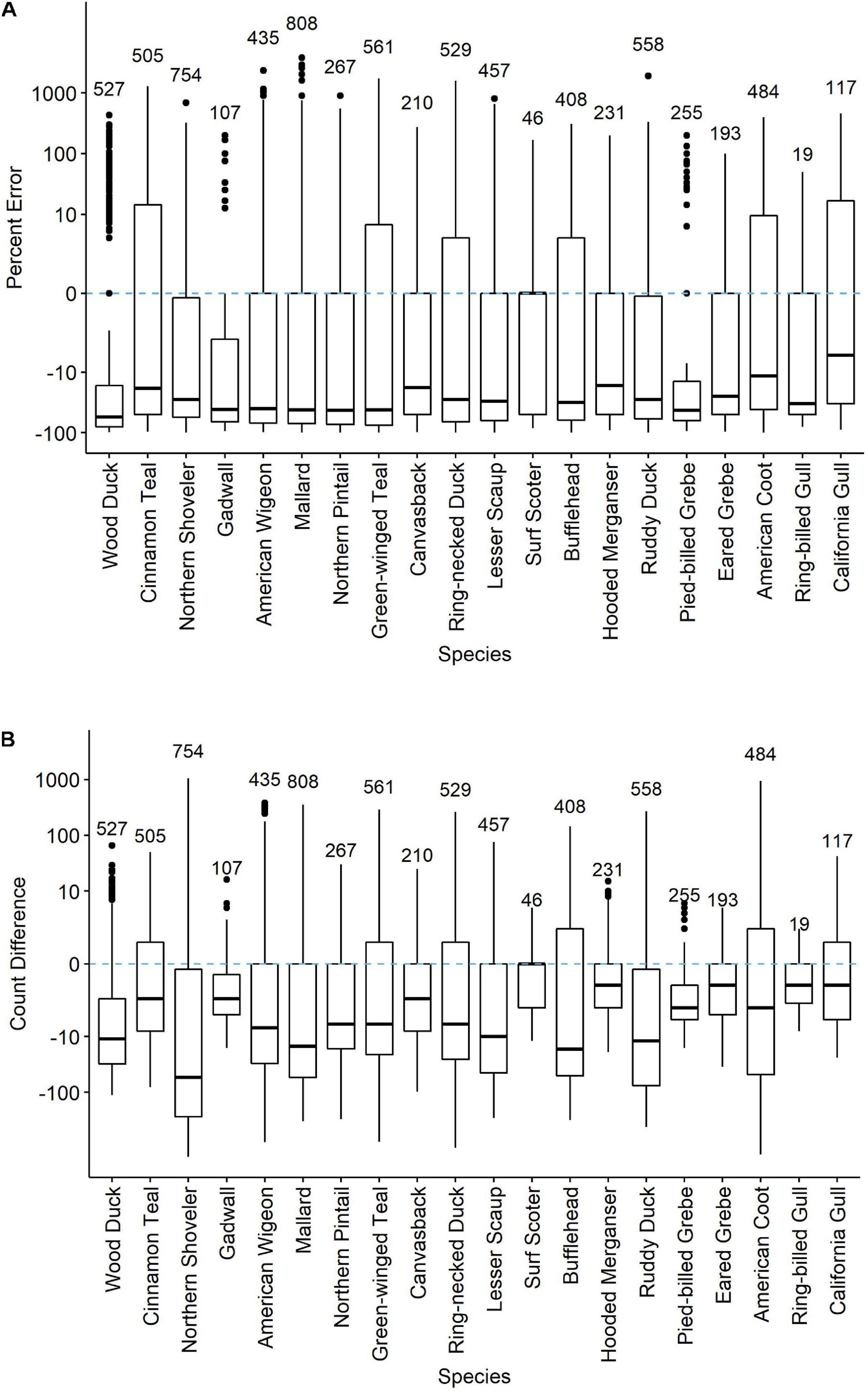
Figure 1 | Percent error (A) and count differences (B) in counts of 20 waterbird species reported on eBird checklists at the Philomath Ponds, Oregon USA, 2010–2019. Medians, quantile plots and outliers are indicated, as well as number of checklists reporting counts of each species. Only checklists reporting counts greater than zero were included. For checklists including counts of zero on dates when R* counts were non-zero, see Supplementary Figure 2.
Percent error, when averaged across species and all observers, was fairly consistent at 30% when counts were 30 or greater. Below 30, counts were more accurate, being closest to zero error when counts were of 8–10 birds (Figure 2A). Percent error was related to the percent richness (our index of observer skill where higher percentages indicated an observer included more of the species known to be present that day on their checklists) in a curvilinear fashion. Checklists including the lowest richness tended to overcount (Figure 2B). Those including 50% of the expected species undercounted by 50% on average, while checklists including 90% or more of the species reported on R∗ checklists averaged deviations of 15% or less in count.
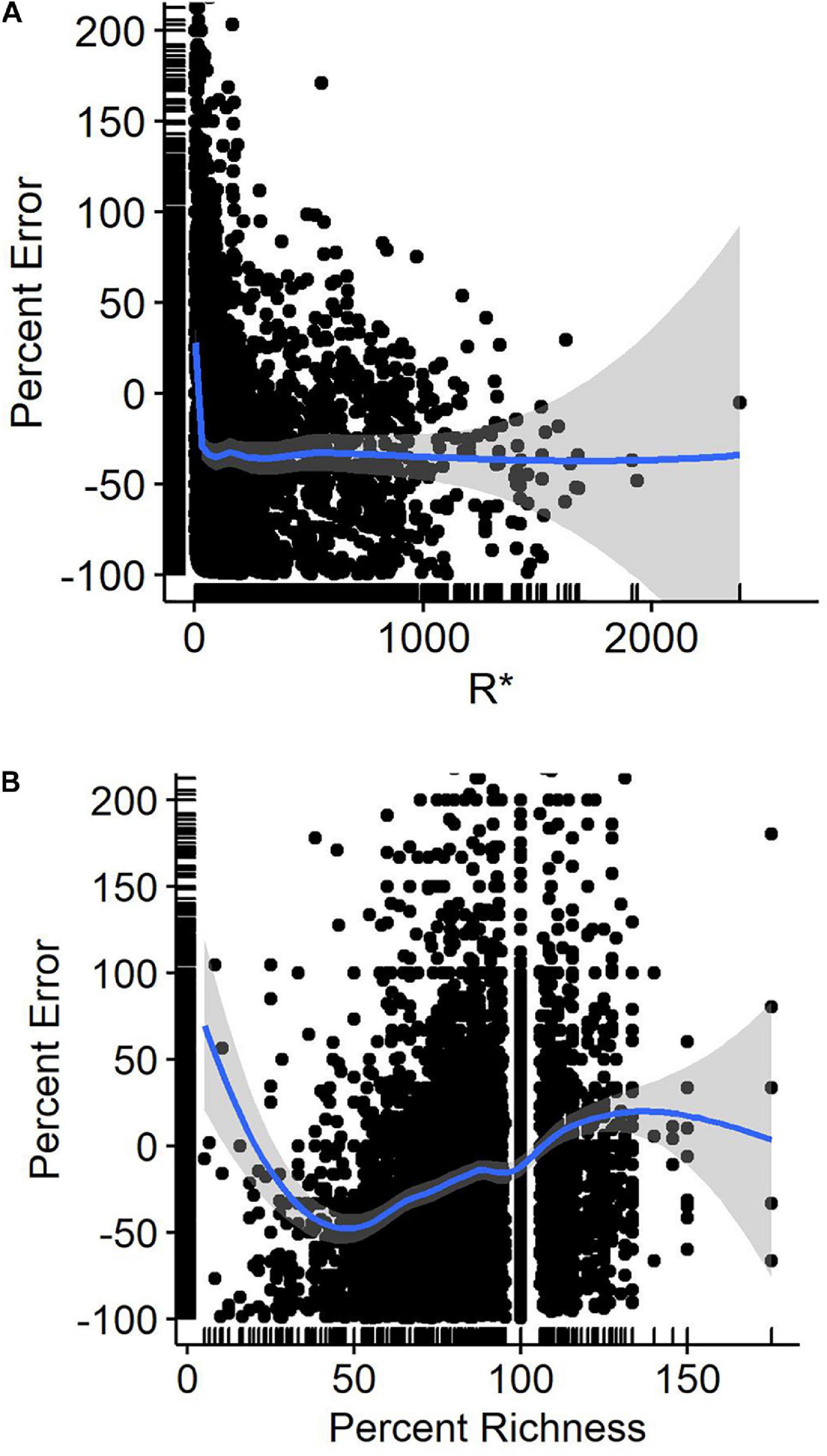
Figure 2 | Percent error in eBird waterbird counts as a function of (A) reference (benchmark) counts (R*), and (B) percent richness of waterbird species detected at Philomath ponds. Lines are Loess regression lines with 95% confidence intervals.
In our multi-species mixed-effects model set, our top model garnered 70 percent of the model weight and was over four BIC from the next most competitive model (Table 2). Our BIC top model indicated that a quadratic effect of R∗ and a linear effect of percent richness best explained variation in percent error.

Table 2 | Model results of variables most influential on percent error.
Seasonality in bird numbers was also captured when the second-order R∗ was included as the most informative variable predicting percent error. Numbers of all species varied considerably across each year (Figure 3). Likewise, total waterbird abundance varied several-fold from its nadir in June to a maximum in October and November (Supplementary Figure 3). Yet, total waterbird abundance was rarely an informative variable in our model sets. Only in counts of American Coot did it appear in the most parsimonious models (in combination with percent richness). In California Gull, waterbird abundance appeared as an informative variable but only in a weakly competitive model (19% of the model weight).
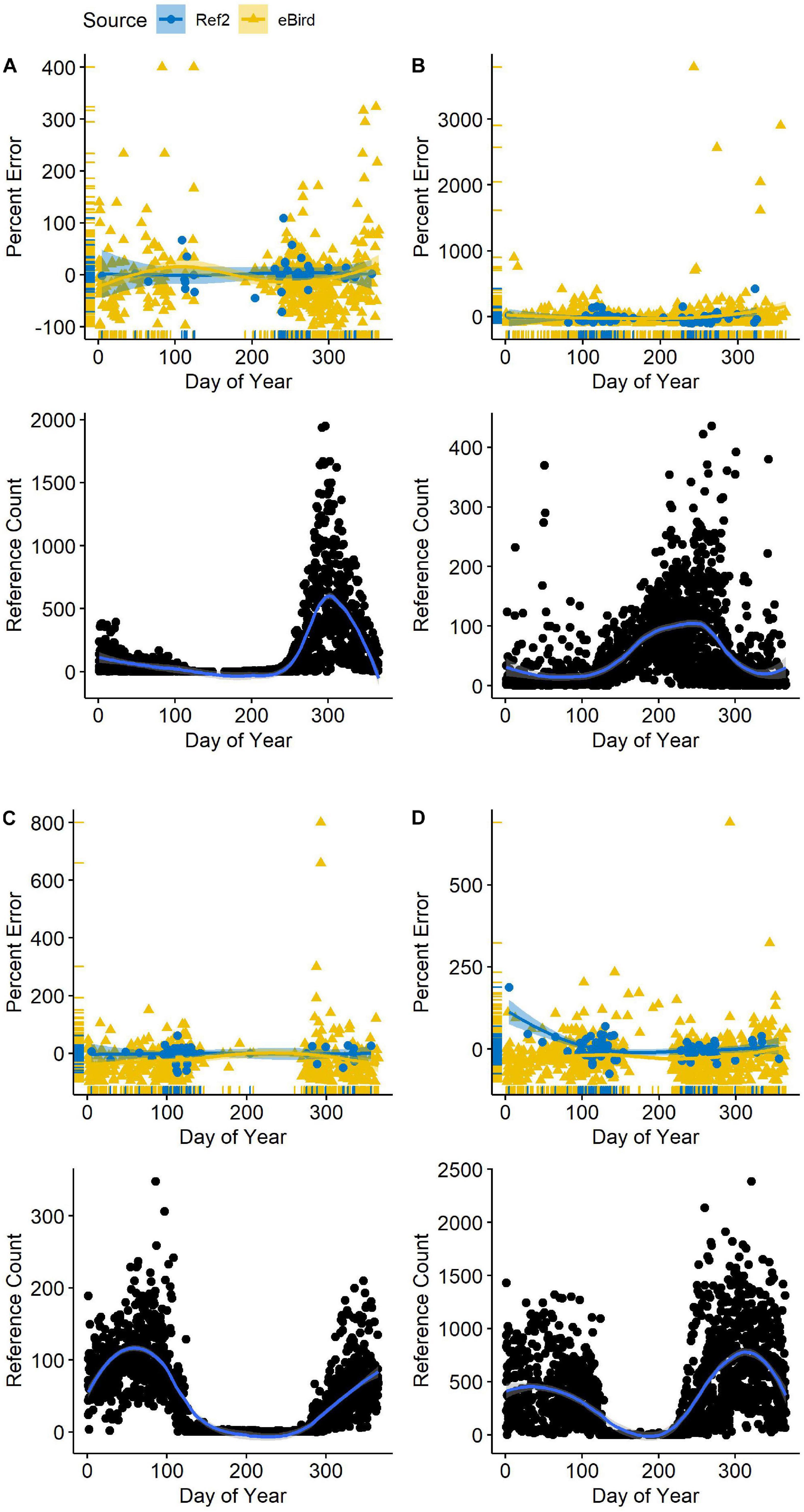
Figure 3 | Variation in reference (benchmark) counts (R*) as a function of date (lower panel) and counts reported in eBird (gold triangles in upper panel) alongside second-visit counts (Ref2; blue circles) at Philomath ponds, Oregon USA, 2010–2019. Counts in the upper panels are indicated with respect to the R* count (zero line) each day. Loess regression lines with 95% confidence intervals are included. (A) American Coot; (B) Mallard; (C) Lesser Scaup; (D) Northern Shoveler.
Within the species-specific model sets, the combination of R∗ and percent richness carried most of the model weight (mean = 0.83, SD = 0.18) in 13 of our 18 non-gull species (Supplementary Table 2). For gulls, top models struggled to outcompete the null. Altogether, R∗ and/or percent richness were in the top model sets for 17 of 18 non-gull waterbirds.
Within our full model, bird characteristics were rarely influential on percent error (Table 2). Similarly, species-specific models rarely discovered bird traits to be informative variables (Supplementary Table 2).
Our models often identified percent richness as an influential variable on percent error, so we related percent richness to percent error as means across all checklists contributed by each observer (Figure 4A). The two were positively related, yet only six of the 246 observers averaged percent errors of less than 10%. The range in percent error for observers detecting 90% or more of waterbird species was actually greater than the range for observers who detected less than 60% of species, indicating that percent error alone is an unreliable predictor of counting accuracy. The relationship was not necessarily driven by site experience because four of the six observers with the most accurate counts were contributing very few checklists (Figure 4B).
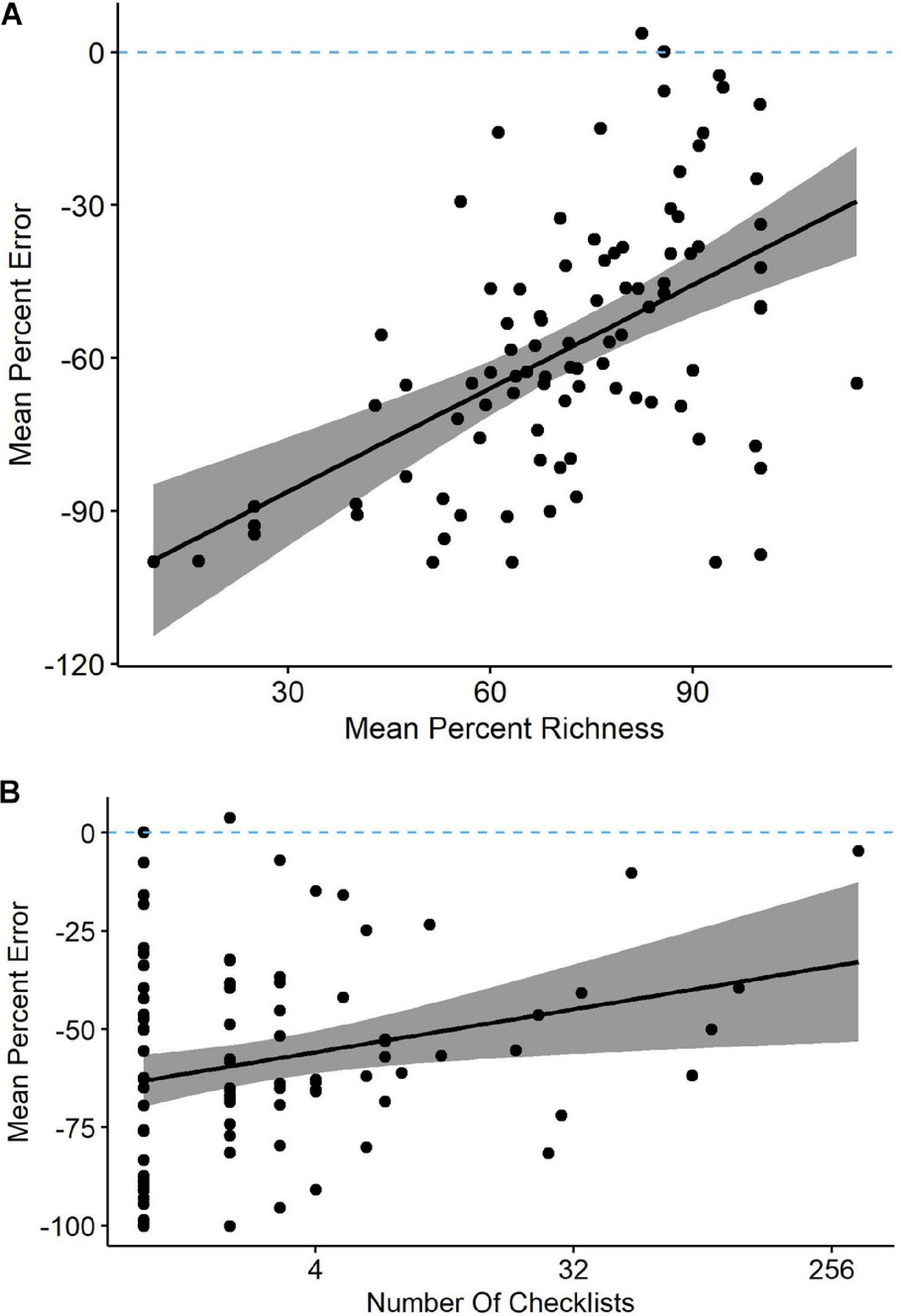
Figure 4 | (A) Observers reporting a greater percentage of waterbird species present at Philomath ponds, Oregon USA, tended to have lower percent counting errors in their eBird checklists (linear regression and 95% confidence intervals; y = −110 + 0.68x). (B) Observers submitting more total checklists tended to have lower counting errors (y = −60 + 0.17x). Note that these are means of all applicable checklists for each observer, so each point represents a unique observer.
We then selected checklists from the ten observers who contributed the most. Those checklists also showed evidence of undercounting. In nearly all 20 species, percent error was 10–60% greater than even the Ref2 counts (Figure 5). Percent error was highly variable across species. In some species, such as American Coot, three of the 10 observers reported counts averaging very near the Ref2 counts, whereas in others, such as Pied-billed Grebe, all observers undercounted by at least an average of 20%. Again, percent error was highly variable in all species even when median percent error did not deviate far from zero.
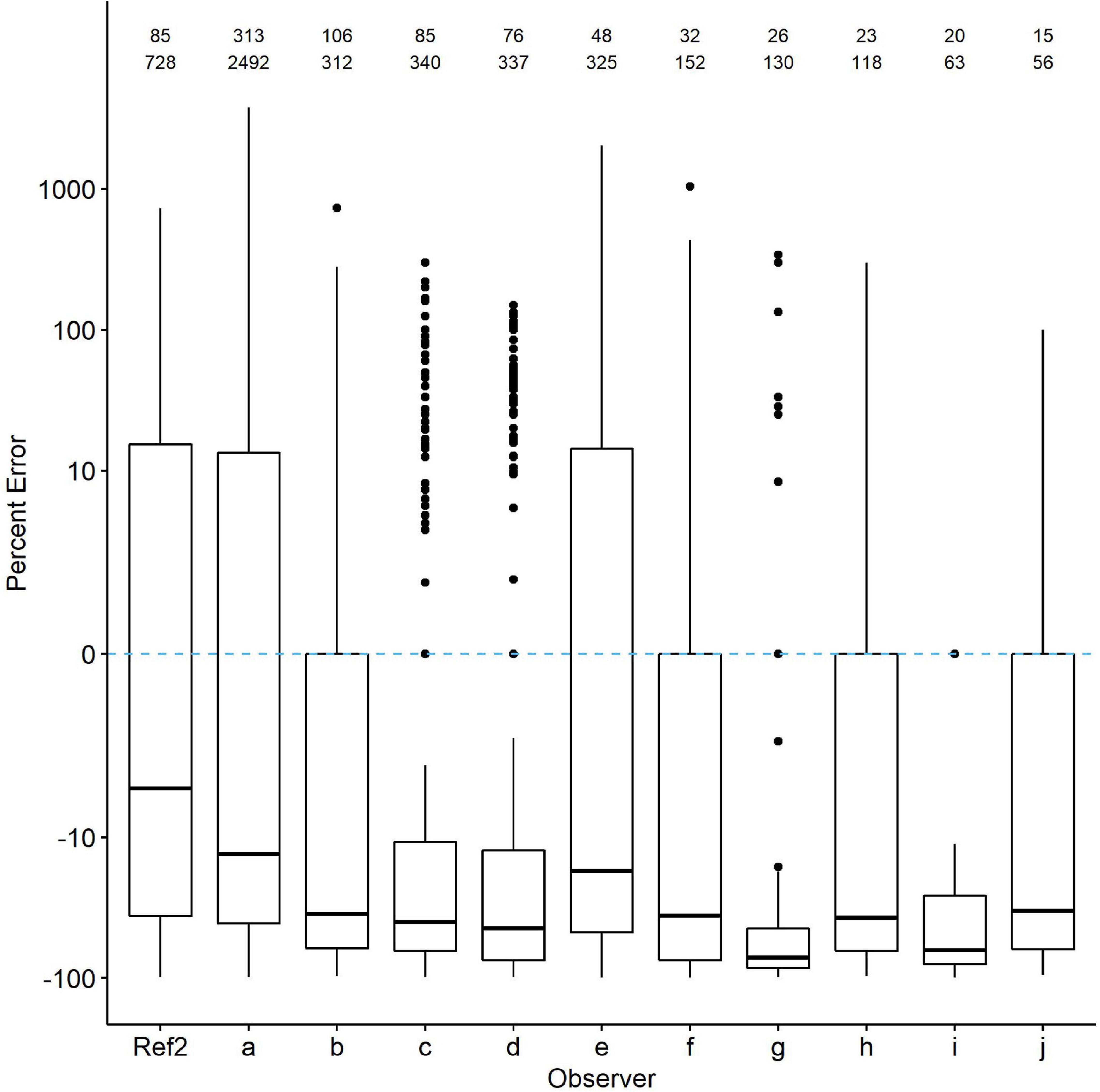
Figure 5 | Comparison of percent count errors in eBird checklists contributed by the 10 observers with the most checklists (top row of numbers) and waterbird observations (second row of numbers; each checklist includes multiple species). The zero line is R*. Ref2 is the second-visit data from WDR. Quantile plots show the median, 25th percentiles as boxes and whiskers, plus outliers. Species-specific plots are available from the authors upon request.
We visualized characterization of the richness and abundance of the daily waterbird community with NMDS through ordination of checklists (grouped by observer) in species space. Observers characterizing the community and its species abundance patterns similarly to R∗ fell nearer to R∗ whereas those positioned increasingly further from R∗ described the community in increasingly dissimilar details. In both January (Figure 6A) and October (Figure 6B) high inter-observer variability in how their checklists characterized the waterbird community led to a general lack of clustering near R∗. In both months, observers reporting more species, contributing more checklists, and surveying for more time tended to group nearer R∗. The collective average, , was nearer R∗ than any individual observer during January but one observer was closely positioned near during October. Removal of checklists from the observer contributing the most data had minimal effects on results.
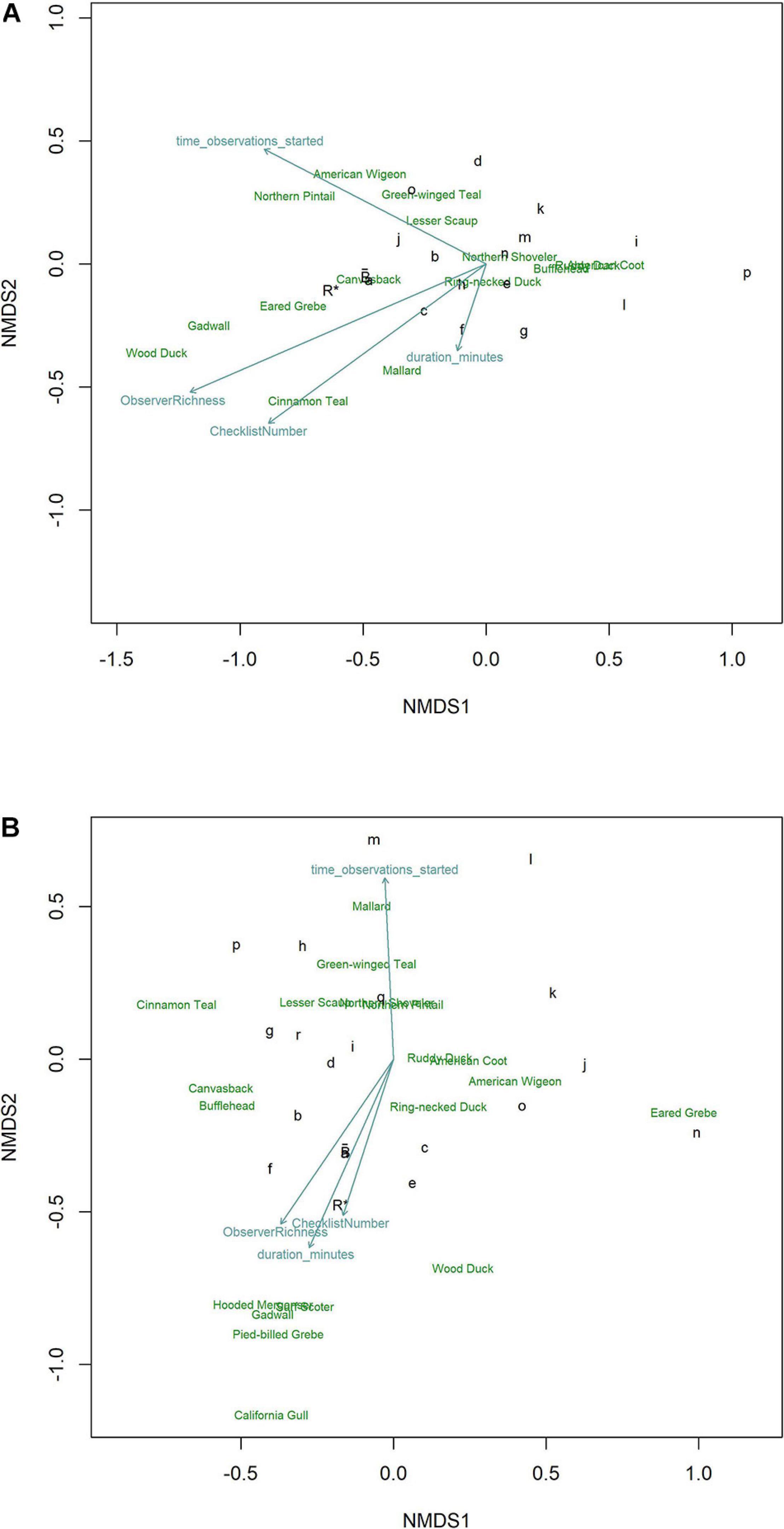
Figure 6 | Ordinations using NMDS of eBird checklists’s characterization of the waterbird community during (A) January and (B) October at Philomath ponds, Oregon USA, 2010–2019. The most influential vectors included Observer Richness (percent of known richness reported on each checklist), Checklist Number (total number of checklists per observer), observation start time each day, and the duration of each observation period. Relative positions of species in species space are noted by species English names. Benchmark counts are noted by R*. Individual observers are noted by lower case letters; those nearest to R* produced characterizations of the waterbird community most like R*. is the collective average of eBird checklists, showing that from the perspective of generally characterizing the community, averaging across checklists contributed by many observers aligns more closely with R* than do checklists from most individual observers, although observer a occupies nearly the same location in species space.
Benchmark data are often designed to understand temporal change in biodiversity (Curtis and Robinson, 2015; Curtis et al., 2016; Robinson and Curtis, 2020). Here, we show that they can also be used to establish standards that aid in quantification of count accuracy in citizen-science data. Through comparisons with such a standard, we discovered that bird count data contributed to eBird from our study site were consistently biased toward undercounting. Counts averaged approximately 30% too low whenever benchmark counts were of 30 or more birds. By comparison, estimates of errors in the R∗ data averaged −0.4 (SD = 2.1%) based on comparisons with photos. Importantly, however, eBird count data exhibited high variability across species and observers. Because of the magnitude of count deviations and the high variability, standards like our benchmark data are needed to inform decisions regarding what subsets of abundance data should be selected to most rigorously address particular scientific questions or management decisions, analogous to how checklist calibration indices help researchers choose suitable eBird checklists based on site- and time-specific expectations of species richness (Yu et al., 2010; Kelling et al., 2015; Johnston et al., 2018). Yet, situations in which such informative standards may be developed and compared appear to be rare currently.
Our study site presented a unique opportunity to compare bird count data contributed to a citizen science database (eBird) with benchmark reference data collected by a professional observer focused on generating accurate daily counts. Characteristics of the site, where all birds were in the open and identified by sight, minimized issues of availability and therefore the need for detectability adjustments to compare counts. Data were contributed by 246 observers and included 676 dates across 10 years, providing an unusual opportunity to explore patterns and potential sources of error. Although the extent to which our results may be generalized to other sites remains unclear given the rarity of opportunities like this one, the situation probably represents a best-case scenario because birds were in the open and easy to observe. Despite the advantages, count differences in 20 species of waterbird were highly variable across the calendar year, species, and observer. Coefficients of variation were high, averaging 6.6 across the 20 species and ranging from 1 to 35.6. For comparison, in an experimental study of observer counting errors of singing birds, which should have been much harder to detect and identify but had a lower range of abundances than our waterbird community, coefficients of variation averaged 0.1 (Bart, 1985).
An assessment of count differences between benchmark data and citizen science contributions will be most robust when estimates of count variability exist for both sets of counts. Estimates of variability in counts from citizen science data are easier to generate because of the large number of visits by multiple observers. Our benchmark (R∗) data were gathered by one professional ornithologist beginning in 2006 prior to widespread eBird use by the birding community (only data since 2010 are included here). The goal was to use those waterbird count data to track population trends and to be able to detect annual changes as small as 2%, thus a high degree of count accuracy was required. No internal check of R∗ counting errors was implemented consistently, in part because of the unique circumstances of the study site where all birds were in the open and easy to detect. On 90% of the days, the number of birds of 18 of our 20 species present averaged less than 15 individuals on each pond, increasing the likelihood of accurate counts. Average numbers were higher for Northern Shoveler and American Coot and during 5 weeks of peak fall migration. Our commonest measure of potential variability in the benchmark data derives from same-day counts (Ref2) by the same observer. Those counts from a later time on the same day averaged 8% lower. However, use of the same observer’s second counts addresses repeatability of count data, not necessarily count accuracy. To assist in quantification of errors, our implementation of comparisons of R∗ counts with photographic evidence revealed average counting errors of −0.4%. Involvement of additional experts counting independently could be an alternative solution to use of photos to effectively quantify counting errors in benchmark data, which we encourage when circumstances allow such opportunities. Overall, our efforts to estimate error in R∗ data reveal that those counts may be 0.4–8% under-counts, on average, compared with eBird average count deviations of approximately 30% under-counts.
Our quantification of count differences in eBird data is conservative because we excluded counts of zero on eBird checklists, even for species known to be present. We did so to minimize the potential confound of misidentifications and reporting errors (failing to enter a count for a species that was actually observed) from our analysis of counting errors. Yet, it is possible that some fraction of 100% undercounts were indeed counting errors in the sense that the species was one that observers were knowledgeable enough to identify but failed to count or report. The median percent error across the 20 species was −48.6 plus or minus 50.9% (MAD) when zero counts were included vs. −29.1 plus or minus 44.6% when zero counts were excluded. Inclusion of zero counts, therefore, has a large influence on the median, but percent errors were highly variable regardless.
Our top overall mixed-effects model carried nearly 70% of the model weight and contained only two variables. The species-specific R∗ count as a quadratic, which captured the seasonality in numbers present at the site, was the most informative variable when combined with a linear effect of percent richness. The inclusion of R∗ indicates that eBird count data were related to the benchmark numbers but that other factors were also influential. Checklists with a more complete list of the species known to be present each day had lower counting errors. Yet, checklists including 100% of expected species still undercounted by an average of 15%. Count differences on checklists from the ten observers who most often visited the site were still exhibiting undercounts even compared to the Ref2 values, which were benchmark counts made later each day during weeks with high levels of migratory movements.
We documented strong directional bias toward undercounts and also a smaller percentage of large overcounts, leading to inconsistent patterns in count differences across species. Our comparisons revealed that undercounting was pervasive, yet very large numbers of a species being present sometimes led to severe overcounting as well. Interestingly, the influence of number of birds appeared to be species-specific. The total number of waterbirds of all species present on a given day was not an influential variable in our overall model explaining percent error, except for one species, American Coot. This pattern suggests that count differences were unlikely to have been caused by observers being overwhelmed by the total number of birds to observe, identify and count. Instead, it appears that each species presented different challenges to observers. Given that our models rarely identified species’ traits as being informative, it remains unclear what species-specific factors are responsible.
The degree of variability across species in count differences should influence potential decisions regarding use of eBird count data. Our analyses clearly reveal that off-the-shelf acceptance of count data for assessments of absolute abundance should be done with great care and thoughtfulness. In addition, if researchers wish to avoid focus on absolute abundance by instead evaluating relative abundance, our results suggest further caution is warranted. We found great interspecific variability in count differences. That is, although bias was nearly uniformly directional toward undercounting, the magnitude of undercounts varied substantially across species indicating that processes generating errors are inequivalent across species. Therefore, judging differences in one species’ abundance relative to others requires careful thought. If explorations of relative abundance are focused on within-species changes across sites, care is also warranted because we found substantial differences among observers in count accuracy. If different sites have different observers, then error/bias processes will be expected to be different as well. Effective use of relative abundance data depends on assumptions of consistent errors across species and sites, which appears to be largely untrue in our data. Further exploration of techniques to determine the degree to which assumptions of similar counting errors across species might be relaxed to preserve the utility of relative abundance analyses are warranted. The use of abundance categories could be explored to maximize the information content gleaned from count data.
What role might species misidentifications have played in counting errors? Count differences were regularly so large that we conclude species misidentification was unlikely to be an important factor. Probably the most challenging identifications involved female or eclipse-plumaged ducks, which observers might ignore and exclude from checklists if identification is uncertain. We consider such omissions to be unlikely for at least three reasons. First, degree of dichromatism was uninformative in our models explaining percent error. Second, assuming that females represent approximately half of each species present during most months of a year, count differences might be expected to average 50% if males were counted accurately but females were not. Instead, percent error varied widely across species. Finally, count differences of monochromatic vs. dichromatic species were not obviously different. However, it is possible that observers were more accurate for some species than others because of paying greater attention to unusual or favorite species (Schuetz and Johnston, 2019). At our site, most charismatic species of great interest to birders are rarities and so were not included in our analyses. Counts of Surf Scoter, a species that occurs during a narrow window of time in fall, were generally accurate, but we cannot attribute the accuracy to celebrity alone given its occurrence in such small numbers.
Aside from a predominantly directional bias toward undercounts, we found few consistent species-specific patterns in percent error. Errors differed in magnitude across species, observers, and time of year. Therefore, development of some type of calibration effort, where checklist numbers are adjusted to more closely approximate species-specific abundances poses an interesting challenge. The variability in raw count data suggests that tracking trends across time without additional steps to filter data or analytically adjust for noise could be especially problematic. Depending on the particular scientific question of interest, needs for precision might decline, so other analytic approaches could be implemented. For example, if abundances can be binned into categories and approaches such as ordinal or quantile regression used (Ananth and Kleinbaum, 1997; Koenker and Hallock, 2001; Howard et al., 2014), less precisely defined trends over time might be identified. Furthermore, our observation that percent richness, which we assume to be a correlate of observer experience, was often an informative variable, suggests that additional exploration of count calibration approaches for data contributed by the most experienced observers might be informative.
If questions about patterns in abundances among species in the waterbird community are of interest, our NMDS ordination results suggest that combining checklists across multiple observers rather than selecting data from any single contributor may produce results closer to those generated by professional benchmark data. The vectors in NMDS results may also inform decisions about which criteria to use when filtering data to maximize inclusion of checklists with the greatest value for specific scientific questions. For example, the waterbird community at our site was better characterized by observers who included more species on their checklists, invested more time searching the site each time, and contributed more checklists overall. Although species-specific numbers remained inconsistently related to the R∗ counts, the level of general characterization of the entire community was improved. In a detailed comparison of eBird data with structured survey results near Sydney, Australia, overall characterization of the bird communities was similar as well, but the collectively greater effort expended by eBirders resulted in discovery of a greater number of uncommon species (Callaghan et al., 2018).
Determining the extent to which results from our site and observers may be generalized more widely will require identification of other sites with benchmark data sets. We also recommend further investigation of approaches for identifying checklists with higher probability of having the most accurate count data. New approaches for categorizing checklists based on expected numbers of species have recently been developed but it remains unclear if these same criteria also apply to bird counting accuracy (Callaghan et al., 2018). Our index of checklist quality was based solely on the percent of species reported on checklists that were also detected that day by the professional observer. Percent richness was regularly in top models, so it does have explanatory influence on count differences. Yet, direct comparisons of data from those observers and the R∗ and Ref2 numbers still showed substantial differences, primarily of undercounting.
If a sufficiently detailed benchmark data set is available, however, adjustments for seasonal fluctuations in numbers of each species could conceivably be implemented. Such calibrations might be conducted more effectively if individual observers exhibited consistency in counting errors, an issue we have not explored here. It is unknown if observers improve their counting skills over time in the same way that observers are expected to improve abilities to detect species or if temporal stochasticity drives counting errors. A goal could be to develop a count calibration metric for each observer so that it can be extended and applied to counts from sites lacking benchmark data if those sites are likely to have similar species composition and relative abundances. However, given the high level of variability in count data we quantified across observers, species and time, such calibration metrics may be quite challenging to develop. Complex models such as the Bayesian hierarchical models using Markov chain Monte Carlo approaches implemented with Christmas Bird Count data (Link et al., 2006), might be helpful in the absence of additional information on checklist accuracy and reliability. Our community ordination results suggested that combining data across multiple checklists from multiple observers (the group collective effort) might more closely approximate the community characterization than most single contributors did. Further exploration of similar approaches and sensitivities to checklist characteristics could identify necessary checklist quality criteria that must be met prior to use in such analyses. In the end, use of any checklist count data will be influenced strongly by each project’s specific objectives (Isaac and Pocock, 2015).
We hypothesize that the high variability in species count information on eBird checklists could be influenced by common aspects of birder behavior. Prior to the advent of eBird, most birders, in North America at least, focused their efforts on listing species and watching behavior (Eubanks et al., 2004). Intentional counting was done by a small percentage of particularly avid observers, while most others only counted during organized activities such as Christmas Bird Counts (Boxall and McFarlane, 1993). A much smaller percentage contributed count data to scientific projects with structured protocols such as the North American Breeding Bird Survey. eBird has revolutionized the degree of attention birders pay to numbers of birds around them (Wood et al., 2011). It has pushed birders to value data beyond the day’s species list. The novelty of this effort to count all birds every time one goes birding, may contribute to the variability in quality of the count data. Contributors are largely untrained about best practices for counting, especially when birds are present in large numbers, flying, or inconspicuous because they are secretive or available only by sound. We encourage development of additional training opportunities for eBird contributors to improve their knowledge of the value of accurate count data as well as their counting skills. Training improves data quality even for professional observers (Kepler and Scott, 1981).
An indication on checklists in the eBird database that such training had been accomplished might facilitate selection of checklists by researchers who wish to use count data only from trained observers. Furthermore, the addition of a qualitative categorization of counting accuracy for each checklist, designated by the observer at time of checklist submission to eBird, might be useful. Currently, users may code species using presence-absence information instead of counts or select a checklist protocol (incidental) indicating that not all species detected were included in the list. A count accuracy designation could allow observers to rate their own level of confidence in the accuracy of their counts or the level of attention they paid to counting accurately, which could serve as additional criteria by which researchers might choose checklists for their particular scientific question. Given that many contributors may not focus on producing accurate counts but have a variety of other motivations (Boakes et al., 2016), allowing observers to categorize quickly and easily their personal confidence in their count data would be useful.
Finally, exploration of the sources of variation in count data needs additional attention (Dickinson et al., 2010). The potential value of the vast quantities of information from citizen science databases is great. Such data have the potential to be effective at informing conservation and management decisions (McKinley et al., 2017; Young et al., 2019), but a thorough understanding of sources of error should be a priority before their use (Lewandowski and Specht, 2015). An additional strategy that may contribute to refinement of information on count data quality in citizen science databases could be development of a network of sites with trained counters. These marquis sites could be chosen to represent major habitat types where citizen science data are often gathered or where researchers specifically need high-quality information. Creating a network of high-quality benchmark sites would have the added advantage of leaving a legacy of reliable abundance data for future generations.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Ethical review and approval was not required for the animal study because purely observational.
WR conceived the project, gathered the data, helped analyze the data and wrote the manuscript. TH contributed to data collection and management, analyzed the data, and contributed to writing the manuscript. RH contributed to data interpretation and to writing the manuscript. All authors contributed to the article and approved the submitted version.
WR and TH were supported by the Bob and Phyllis Mace Professorship and the College of Agricultural Sciences. RH was supported by the NSF Grant 1910118.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the many birders who contributed their data to eBird and the many scientists who created, maintain, and continue to improve eBird. Dennis Lewis and Philomath Public Works permitted access to the site, not only to us but to more than 250 birders. We thank our Reconfiguration Grant Group for helpful discussions.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.568278/full#supplementary-material
Supplementary Figure 1 | Number of eBird checklists contributed for the study site at Philomath Ponds, Oregon USA, 2010–2019, as a function of day of year.
Supplementary Figure 2 | Counts of waterbirds in eBird checklists included in our analyses as a function of their percent error.
Supplementary Figure 3 | Relationship between mean percent error on eBird checklists (blue line) and mean waterbird abundance (green line) as a function of day of year at Philomath ponds, Oregon USA, 2010–2019. Waterbird abundance is the mean of all the counts (R∗) of all of the possible 20 study species present each day across the 10 years.
Supplementary Table 1 | Species-specific measurements of central tendency and variation in percent counting errors. (A) excluding species non-detections from checklists; (B) including species non-detections (zero counts) in checklists.
Supplementary Table 2 | Species-specific BIC model results. Full model results are presented for each species alphabetically.
Supplementary Table 3 | Full mixed-effects model results supplementing the abbreviated results presented in Table 2.
Ananth, C. V., and Kleinbaum, D. G. (1997). Regression models for ordinal responses: a review of methods and applications. Int. J. Epidemiol. 26, 1323–1333. doi: 10.1093/ije/26.6.1323
Barker, R. J., Schofield, M. R., Link, W. A., and Sauer, J. R. (2018). On the reliability of N-mixture models for count data. Biometrics 74, 369–377. doi: 10.1111/biom.12734
Bird, T. J., Bates, A. E., Lefcheck, J. S., Hill, N. A., Thomson, R. J., Edgar, G. J., et al. (2014). Statistical solutions for error and bias in global citizen science datasets. Biol. Conserv. 173, 144–154. doi: 10.1016/j.biocon.2013.07.037
Boakes, E. H., Gliozzo, G., Seymour, V., Harvey, M., Smith, C., Roy, D. B., et al. (2016). Patterns of contribution to citizen science biodiversity projects increase understanding of volunteers’ recording behaviour. Sci. Rep. 6: 33051.
Boxall, P. C., and McFarlane, B. L. (1993). Human dimensions of christmas bird counts: implications for nonconsumptive wildlife recreation programs. Wildl. Soc. Bull. (1973-2006) 21, 390–396.
Brown, J. H. (1984). On the Relationship between abundance and distribution of species. Am. Nat. 124, 255–279. doi: 10.1086/284267
Buckland, S. T., Marsden, S. J., and Green, R. E. (2008). Estimating bird abundance: making methods work. Bird Conserv. Int. 18, S91–S108. doi: 10.1017/S0959270908000294
Callaghan, C. T., Martin, J. M., Major, R. E., and Kingsford, R. T. (2018). Avian monitoring–comparing structured and unstructured citizen science. Wildl. Res. 45, 176–184. doi: 10.1071/WR17141
Chandler, M., See, L., Copas, K., Bonde, A. M., López, B. C., Danielsen, F., et al. (2017). Contribution of citizen science towards international biodiversity monitoring. Biol. Conserv. 213, 280–294.
Cohn, J. P. (2008). Citizen science: can volunteers do real research? BioScience 58, 192–197. doi: 10.1641/B580303
Curtis, J. R., and Robinson, W. D. (2015). Sixty years of change in avian communities of the Pacific Northwest. PeerJ 3:e1152. doi: 10.7717/peerj.1152
Curtis, J. R., Robinson, W. D., and McCune, B. (2016). Time trumps habitat in the dynamics of an avian community. Ecosphere 7:e01575. doi: 10.1002/ecs2.1575
Dickinson, J. L., Zuckerberg, B., and Bonter, D. N. (2010). Citizen science as an ecological research tool: challenges and benefits. Annu. Rev. Ecol. Evol. Syst. 41, 149–172. doi: 10.1146/annurev-ecolsys-102209-144636
Eubanks, T. L. Jr., Stoll, J. R., and Ditton, R. B. (2004). Understanding the diversity of eight birder sub-populations: socio-demographic characteristics, motivations, expenditures and net benefits. J. Ecotourism 3, 151–172. doi: 10.1080/14664200508668430
Fink, D., Auer, T., Johnston, A., Ruiz-Gutierrez, V., Hochachka, W. M., and Kelling, S. (2020). Modeling avian full annual cycle distribution and population trends with citizen science data. Ecol. Appl. 30:e02056. doi: 10.1002/eap.2056
Fink, D., Damoulas, T., and Dave, J. (2013). “Adaptive Spatio-Temporal Exploratory Models: Hemisphere-wide species distributions from massively crowdsourced eBird data,” in Proceedings of the Twenty-Seventh AAAI Conference on Artificial Intelligence. Available online at: https://www.aaai.org/ocs/index.php/AAAI/AAAI13/paper/view/6417 (accessed May 21, 2020)
Greenwood, J. J. D. (2007). Citizens, science and bird conservation. J. Ornithol. 148, 77–124. doi: 10.1007/s10336-007-0239-9
Guillery, R. W. (2002). On counting and counting errors. J. Comp. Neurol. 447, 1–7. doi: 10.1002/cne.10221
Horns, J. J., Adler, F. R., and Şekercioğlu, ÇH. (2018). Using opportunistic citizen science data to estimate avian population trends. Biol. Conserv. 221, 151–159. doi: 10.1016/j.biocon.2018.02.027
Howard, C., Stephens, P. A., Pearce-Higgins, J. W., Gregory, R. D., and Willis, S. G. (2014). Improving species distribution models: the value of data on abundance. Methods Ecol. Evol. 5, 506–513. doi: 10.1111/2041-210X.12184
Hutto, R. L. (2016). Should scientists be required to use a model-based solution to adjust for possible distance-based detectability bias? Ecol. Appl. 26, 1287–1294. doi: 10.1002/eap.1385
Isaac, N. J. B., and Pocock, M. J. O. (2015). Bias and information in biological records. Biol. J. Linn. Soc. 115, 522–531. doi: 10.1111/bij.12532
Johnston, A., Fink, D., Hochachka, W. M., and Kelling, S. (2018). Estimates of observer expertise improve species distributions from citizen science data. Methods Ecol. Evol. 9, 88–97.
Kelling, S., Johnston, A., Bonn, A., Fink, D., Ruiz-Gutierrez, V., Bonney, R., et al. (2019). Using semistructured surveys to improve citizen science data for monitoring biodiversity. BioScience 69, 170–179. doi: 10.1093/biosci/biz010
Kelling, S., Johnston, A., Hochachka, W. M., Iliff, M., Fink, D., Gerbracht, J., et al. (2015). Can observation skills of citizen scientists be estimated using species accumulation curves? PLoS One 10:e0139600. doi: 10.1371/journal.pone.0139600
Kepler, C. B., and Scott, J. M. (1981). Reducing bird count variability by training observers. Stud. Avian Biol. 6, 366–371.
Koenker, R., and Hallock, K. F. (2001). Quantile regression. J. Econ. Perspect. 15, 143–156. doi: 10.1257/jep.15.4.143
Lewandowski, E., and Specht, H. (2015). Influence of volunteer and project characteristics on data quality of biological surveys. Conserv. Biol. 29, 713–723. doi: 10.1111/cobi.12481
Link, W. A., Sauer, J. R., and Niven, D. K. (2006). A hierarchical model for regional analysis of population change using Christmas Bird Count data, with application to the American Black Duck. Condor 108, 13–24.
McKinley, D. C., Miller-Rushing, A. J., Ballard, H. L., Bonney, R., Brown, H., Cook-Patton, S. C., et al. (2017). Citizen science can improve conservation science, natural resource management, and environmental protection. Biol. Conserv. 208, 15–28. doi: 10.1016/j.biocon.2016.05.015
Miller-Rushing, A., Primack, R., and Bonney, R. (2012). The history of public participation in ecological research. Front. Ecol. Environ. 10:285–290. doi: 10.1890/110278
R Core Team (2020). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Robbins, C. S., and Stallcup, R. W. (1981). Problems in separating species with similar habits and vocalizations. Stud. Avian Biol. 6, 362–365.
Robinson, W. D., and Curtis, J. R. (2020). Creating benchmark measurements of tropical forest bird communities in large plots. Condor 122, 1–15. doi: 10.1093/condor/duaa015
Robinson, W. D., Lees, A. C., and Blake, J. G. (2018). Surveying tropical birds is much harder than you think: a primer of best practices. Biotropica 50, 846–849. doi: 10.1111/btp.12608
Schuetz, J. G., and Johnston, A. (2019). Characterizing the cultural niches of North American birds. Proc. Natl. Acad. Sci. U.S.A. 116, 10868–10873. doi: 10.1073/pnas.1820670116
Sullivan, B. L., Wood, C. L., Iliff, M. J., Bonney, R. E., Fink, D., and Kelling, S. (2009). eBird: a citizen-based bird observation network in the biological sciences. Biol. Conserv. 142, 2282–2292. doi: 10.1016/j.biocon.2009.05.006
Walker, J., and Taylor, P. (2017). Using eBird data to model population change of migratory bird species. Avian Conserv. Ecol. 12:4. doi: 10.5751/ACE-00960-120104
West, M. J. (1999). Stereological methods for estimating the total number of neurons and synapses: issues of precision and bias. Trends Neurosci. 22, 51–61. doi: 10.1016/S0166-2236(98)01362-9
Wood, C., Sullivan, B., Iliff, M., Fink, D., and Kelling, S. (2011). eBird: engaging birders in science and conservation. PLoS Biol. 9:e1001220. doi: 10.1371/journal.pbio.1001220
Young, B. E., Dodge, N., Hunt, P. D., Ormes, M., Schlesinger, M. D., and Shaw, H. Y. (2019). Using citizen science data to support conservation in environmental regulatory contexts. Biol. Conserv. 237, 57–62. doi: 10.1016/j.biocon.2019.06.016
Keywords: biodiversity benchmarks, birder behavior, citizen science, eBird, statistical bias, statistical error, wildlife counts
Citation: Robinson WD, Hallman TA and Hutchinson RA (2021) Benchmark Bird Surveys Help Quantify Counting Accuracy in a Citizen-Science Database. Front. Ecol. Evol. 9:568278. doi: 10.3389/fevo.2021.568278
Received: 31 May 2020; Accepted: 20 January 2021;
Published: 10 February 2021.
Edited by:
Orsolya Valkó, Hungarian Academy of Science, HungaryReviewed by:
Daniel Jay Twedt, United States Geological Survey (USGS), United StatesCopyright © 2021 Robinson, Hallman and Hutchinson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: W. Douglas Robinson, ZG91Z2xhcy5yb2JpbnNvbkBvcmVnb25zdGF0ZS5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.