 Thaiane Weinert da Silva
Thaiane Weinert da Silva Carla Suertegaray Fontana
Carla Suertegaray Fontana- Laboratório de Ornitologia, Museu de Ciências e Tecnologia, Programa de Pós-Graduação em Ecologia e Evolução da Biodiversidade, Pontifícia Universidade Católica do Rio Grande do Sul (PUCRS), Porto Alegre, Brazil
Since in general the surrounding landscape influences the species diversity and abundance in fragments of native vegetation, an amount of native-vegetation cover nearby may also positively affect communities in restored areas, regardless of the sizes of individual habitat patches. We investigated for the first time whether the species richness, total abundance, and density of grassland birds in restoration sites are influenced by the amount of native grassland in the surrounding landscape in the Brazilian Pampa. We sampled birds by point counts in five restoration sites in the most representative area of grasslands in Brazil. We established an outer buffer zone with a 1 km-radius around the point-count areas in each site, and calculated the percentage of native grassland vegetation in the surrounding landscape. Bird species richness and abundance did not show a significant response to the amount of neighboring native grassland in restored areas. Individual analyses of the density of seven bird species associated to grassland also showed similar pattern. We believe the vegetation structure in these restoration sites may already been sufficiently re-established to provide necessary resources and a suitable habitat for the birds. Even so, we assume that previously existing landscape features were important for recovery of the vegetation structure, as continuous native grassland in the surroundings. Thus, we recommend consider the landscape context as an additional issue in studies dealing with conservation strategies for recovery of grasslands in Brazil.
Introduction
Global biodiversity has been continually impacted, with most species now living in fragmented patches resulting from land-use changes and habitat destruction (Haddad et al., 2015; Fletcher et al., 2018). The surrounding landscape influences the species abundance and diversity in fragments, since the landscape may include connecting corridors, influence dispersal between habitat fragments, and depending on the nature of land use, can alter conditions in habitat patches negatively or positively (Öckinger et al., 2012). Landscapes that retain substantial amounts of native vegetation cover should generate large positive ecological responses at the local scale (Kroll et al., 2014).
Community structure is influenced by the landscape configuration, and diversity within a patch depends on the structure of the surrounding landscape (Dauber et al., 2003), i.e., a community in a restored habitat may depend to some degree on the surroundings. Species richness can be shaped by the physical environment, which includes several characteristics of habitat patch area, e.g., quality, size, configuration, and connectivity (Aggemyr et al., 2018). In the case of birds, where individuals can occur across a variety of habitat patches (Whitaker and Warkentin, 2010; Lee and Carroll, 2014), the proportion of native-grassland patches remaining in the landscape can affect the presence of bird species in grasslands (Cerezo et al., 2011). The larger the amount of native grassland in the patches, the greater the richness and abundance of birds (Silva et al., 2015). Therefore, bird species distribution and occurrence can be strongly influenced by landscape characteristics (Lee and Carroll, 2014).
Grasslands have been replaced and fragmented due to changes in land use, mainly agricultural expansion (Pretelli et al., 2018). Restoration of degraded habitats, i.e., recovery of an ecosystem, is still not widely used for tropical and subtropical grasslands (Buisson et al., 2019). Conservation strategies to preserve and restore habitats should consider the quality of the landscape as a whole (Fahrig, 2001). In grassland restoration, spontaneous vegetation recovery depends on the persistence of seed banks and on input of seed from external sources such as native grasslands near the restoration sites (Favreto and de Medeiros, 2006; Andrade et al., 2015; Vieira et al., 2015). Hence, well-conserved landscape patches are important, since recovery is affected by the surrounding land-use matrix that serves as a vital source of propagules (Holl and Aide, 2011).
The total number of species in a given habitat type within a landscape increases with the total amount of that habitat in the landscape, regardless of the size of individual habitat patches (Fahrig, 2013). In view of this, and considering the high proportion of degraded grasslands in the Brazilian Pampa biome in southeastern South America (SESA Grasslands; Azpiroz et al., 2012), we have compared the structure of bird communities of restoration sites with those of native grasslands. We found similarity in the species richness and composition between sites under passive restoration and sites in native grassland (Silva et al., 2019), but these variables differed between sites under active restoration and sites in native grassland (Silva and Fontana, 2020). These findings suggested that the similarities might be due to effects from fragments of native grassland in the surroundings of grassland under restoration. Here, our objective was to examine whether the species richness, total abundance, and density of grassland birds in the same restoration sites are influenced by the amount of native grassland available in the landscape. We expected that restoration sites with large areas of native grassland vegetation nearby would have higher diversity and density of grassland bird species. The landscape matrix can facilitate the dispersal and movement of organisms between habitat patches, which can provide additional habitat for them (Haynes et al., 2007; Lindenmayer et al., 2010).
Materials and Methods
Study Area
We carried out the study at five restoration sites located in the Brazilian Pampa grasslands, state of Rio Grande do Sul, southern Brazil, which have been used in previous studies (Silva et al., 2019; Silva and Fontana, 2020; Supplementary Figure 1). This was the maximum number of restoration sites found after more than 6 months of search during previous study design. The region is characterized by the presence of native grasslands used mainly for extensive livestock and grain cultivation, especially rice and soybeans (better description of the vegetation, fauna, soil characterization and use can be found in Roesch et al., 2009). Four of the studied sites were undergoing passive restoration, i.e., unassisted recovery following abandonment of fields that had been used to grow soybeans and/or rice for more than 10 years. The sizes of these sites ranged from 65 to 600 ha, and three of them were on private land. The fifth site, on the Brazilian Army reserve, has been undergoing active restoration since 2015 and was previously planted with soybeans for at least 10 years. Several restoration techniques have been used in this 400-hectare site, including fallowing, mechanical mowing, controlled cattle grazing, cattle-exclusion periods, and cattle as transport and dispersal agents for native-plant seeds. Most of the grasslands in this region have been converted to agriculture and afforestation, but few remnants of native grassland and forest persist. This study is part of the first university-government initiative to evaluate bird communities in restoration habitats in grasslands of South America. All five sites had similar relief, soil types, and climates, besides a low cattle stocking rate (≤1 animal unit per ha). They were at least 2.5 km apart, and had a restoration time ranging from 5 to 35 years (the location map and details of each study site are in Silva et al., 2019 and Silva and Fontana, 2020).
Bird Sampling
We sampled birds during the breeding season, i.e., between November and February, in 2015–2016 and 2016–2017, totaling two sampling in each site. We surveyed birds in point counts of 5 min and a 100-m radius, totaling 50 point counts for all sites, all completed by TWS. The sampling occurred soon after sunrise on days of favorable weather (see Silva et al., 2019 and Silva and Fontana, 2020 for details). The distance from the observer to the birds was measured with a rangefinder, and birds in flight were not considered. We recorded a total of 50 species. From that we considered 30 species of birds that are restricted to or that make extensive use of grassland habitats (sensu Azpiroz et al., 2012). For the analysis, we selected 11 species with five or more occurrences in point counts, i.e., observed in at least 10% of the point counts (Lockhart and Koper, 2018).
Landscape Data
We obtained satellite images of Bing Aerial Layer, using Quantum GIS 2.18 (Qgis Development Team, 2016). For each site, first we marked a 250-m inner buffer zone around the bird point counts (with a 100-m radius and 150 m from the edges). We then marked an outer buffer zone with a 1-km radius, surrounding the smaller buffer zone. We determined this buffer proportion because it encompasses the home ranges of most Neotropical songbirds (Lee and Carroll, 2014), and is sufficiently large for the birds to perceive as a landscape (Rodewald and Yahner, 2001), covering all environmental features (Alexandrino et al., 2019). We drew polygons of all land uses except native grassland inside the outer buffer zone for each site, using a 1:20,000-scale screen. Land-use types were monocultures, native forest, water bodies, and human-impacted areas. We calculated the percentages of land uses for each outer buffer zone and subtracted them from the total area of this buffer zone, to obtain the percentage of native grassland area (Table 1).

Table 1. Relative number of grassland-bird species and individuals per point count, and area of native grassland in passive (PR) and active (AR) restoration sites in Brazilian Pampa grasslands.
Statistical Analysis
To determine if there was a relationship in species richness and abundance of grassland birds with the amount of native grassland habitat available in the landscape, we performed a Hierarchical Linear Model (HLM) using the function “lmer” in the “lmerTest” package in R software (Kuznetsova et al., 2017; R Core Team, 2020). Our models included the richness and abundance for each point-count (response variables), the percentage of native grassland (independent variable) and a null model, and site as random effect to control for non-independence of point-counts. Our full model was “x = lmer (response variable ∼ independent variable + (1|site).” We selected the best model comparing the full and null model using ANOVA comands (Zuur et al., 2009). To density, we performed a Tweedie compound Poisson generalized linear model, since the Tweedie distribution accept non-integer and zeros values. We used the “cpglm” function of the “cplm” package in R (Zhang, 2013). We compared the full and null model using the second-order Akaike’s Information Criterion (AIC) corrected for small sample sizes (AICc). The model with the lowest AICc value was selected as the best model. The significance level was α = 0.05.
We estimated the density for the seven most associated grassland species at each site using distance-sampling analysis of our point-count data and the multiple covariates distance sampling (MCDS) engine in Distance 7.1 Release 1 (Buckland et al., 2001; Thomas et al., 2010). Grassland species with >30 observations were analyzed individually (Grassland Sparrow Ammodramus humeralis, Grassland Yellow Finch Sicalis luteola, and Blue-black Grassquit Volatinia jacarina), and we post-stratified analysis by sample. For other four species, to reach the minimum number of observations required to produce a reliable detection function with Distance (Buckland et al., 2001), they were combined into a single group (Wedge-tailed Grass Finch Emberizoides herbicola, Pampa Finch Embernagra platensis, Rusty-collared Seedeater Sporophila collaris, and Pearly-billied Seedeater Sporophila pileata). We grouped these species according to their use of habitat for breeding and feeding in southern Brazil, e.g., similar types of vegetation structure, grass height and foraging strategy (Azpiroz et al., 2012; TWS and CSF, personal observation). For species analyzed as a group, we used the group detection probability function and post-stratified the model by species to obtain each species’ density in each site. We compared the following models for each species and group: half-normal and hazard-rate key functions with cosine, simple polynomial, and hermite polynomial series expansion adjustments. We chose the model based on the Kolmogorov–Smirnov probability test for goodness of fit and on Cramer–von-Mises uniform and cosine probability tests for plausibility, and then compared AIC values to select the model with the lowest AIC.
Results
We found no significant difference among the species richness (HLM, t-value = 3.22, P = 0.06) and abundance (HLM, t-value = 2.20, P = 0.13) of grassland birds and the amount of native habitat in the landscape (Figure 1). We found the same for the densities of seven grasslands bird species (Figure 2). For all species, the best model was null model, i.e., the amount of native grassland had not influence on grassland birds density in restoration areas. However, the density of Grassland Sparrow increased with the amount of native grassland, up to four individuals per hectare in the site with 67% native grassland, and for Grassland Yellow Finch up to three individuals per hectare in the site with 85% native grassland in the surroundings (Figure 2), showing a potential pattern to be explored in the light of additional data (more sample sites).

Figure 1. Species richness (A) and abundance (B) of grassland birds relative to the amount of native grassland in the landscapes.
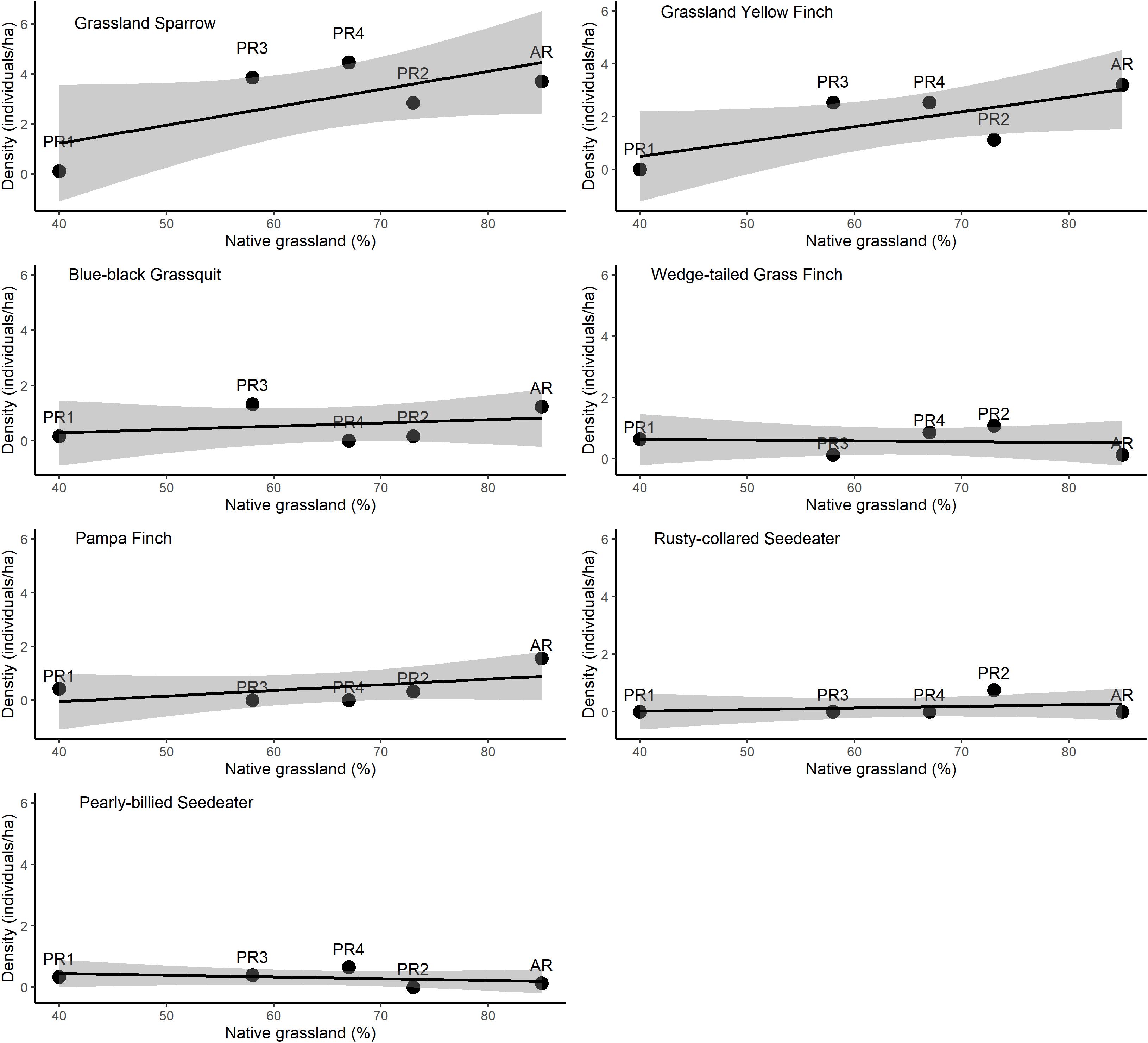
Figure 2. Proportion of grassland-bird density (individuals/ha) per 6 to 12 point counts within each restoration site, to the amount of native grassland vegetation in the landscapes. Confidence intervals are in gray.
Discussion
The surrounding landscape matrix influences the responses of species in habitat fragments (Pretelli et al., 2018), and responses to landscape attributes provide information about improvements in habitat management (Kroll et al., 2014). Increasing the amount of native vegetation may provide additional habitat and can be considered a key driver of species richness (Lindenmayer et al., 2010). Previous studies have found a significant positive relationship between bird species diversity and the amount of native vegetation in the landscape surrounding the patch of habitat where they occur (e.g., Haire et al., 2000; Lindenmayer et al., 2010; Wentworth et al., 2010). Landscapes with large areas of continuous grasslands significantly enhance the richness and abundance of grassland-specialist birds (Codesido et al., 2013; Pretelli et al., 2018). Therefore, the surrounding context has been considered a more important issue than assessing the patch size and degree of isolation (Collinge et al., 2003; Lindenmayer et al., 2010). However, we did not find a strong evidence of this association between the landscape and the species richness, total abundance, and density of grassland bird species. Similarly, a study in Canadian prairies found only weak effects of habitat amount on grassland-songbird relative abundance and richness, although the habitat configuration did strongly influence these parameters on the birds (Lockhart and Koper, 2018). However, our non-significance is weak and can be due to the small number of replicates. We believe that more replicates could provide more-robust information on the effect of the surroundings on birds in restoration areas of Brazilian grasslands. However, in these grasslands we encountered enormous difficulty in locating enough restoration areas and permissions for sampling, which limited our current study. Moreover, in our previous studies evaluating the bird species richness, abundance and composition in these same five restoration sites, we found a recovery potential of the bird community comparable to native grasslands (Silva et al., 2019; Silva and Fontana, 2020). The continuous native grassland surrounding the restoration sites may have influenced this similarity, i.e., the habitat configuration (Lockhart and Koper, 2018). Therefore, the response that we found does not preclude the existence of benefits from remnant grasslands, and there could have been reduction of habitat quality even though native vegetation covered over half of the surrounding landscape.
A high proportion of native grasslands in the landscape can further the recovery of grasslands in the process of restoration (Waldén et al., 2017). Therefore, the habitat structure (vegetation) of the restoration sites that we evaluated may already be sufficiently re-established to provide necessary resources and a suitable habitat for the birds. Furthermore, because at the sampling sites the areas of grassland under restoration were large, the amount of neighboring grassland may have little impact on grassland birds (Lockhart and Koper, 2018). In addition, when less than 30% of native habitat remains in the landscape (which was not the case for our areas, which had at least 40% native grassland), the effects of fragmentation begin to be greater (With and Crist, 1995; Fahrig, 2003). This aspect may be another reason for the observed low influence of neighboring native grassland on the grassland birds. Moreover, our first results for restored grasslands agree with another study conducted in Brazilian remaining native grasslands (Camilotti, 2009), and this pattern may be specific for the Brazilian Pampa. Even so, further studies are needed to confirm if the native habitat of the surroundings does not influence the bird community of sites under restoration.
The lack of a significant correlation between the densities of the seven species evaluated and the amount of native grassland nearby may be associated with the factors described above. Each species may respond differently to habitat and landscape transformation, because of the nature of its particular specialization for foraging and reproduction (Manning et al., 2004; Fischer and Lindenmayer, 2007; Shahan et al., 2017). Species responses can also be influenced by dispersal, movement, and the spatial scale at which species-landscape interactions manifest (Shahan et al., 2017). Although Grassland Sparrow and Grassland Yellow Finch use alternative habitats and occupy a range of grass heights, both species make extensive use of grassland habitats and showed increases in density with increased percentages of neighboring native grassland, a pattern previously observed in Brazilian grasslands (Silva et al., 2015).
This is the first landscape-matrix analysis of birds in grassland habitats under restoration in South America. In view of the small number of study sites and the level of significant found, caution is needed in extrapolating these results. However, even though we failed to find a relationship between the surrounding landscape and the richness and abundance of grassland birds, the previously existing landscape features were important for recovery of the vegetation structure (Overbeck et al., 2013). In addition, it is known that the amount of native habitat in the surroundings can significantly influence the diversity of bird species. We stress the need to consider the landscape context as a complementary approach to guide future decision-making on habitat management in restoration projects and to determine conservation strategies.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because there was not collected and/or manipulation of birds, only observation in the wild.
Author Contributions
TWS and CSF conceived and designed the research, discussed ideas, and improved all previous versions of this manuscript. TWS performed the experiments, conducted the analyses, and wrote the manuscript. CSF edited the manuscript. Both authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Coordination for Improvement of Higher Education Personnel (Capes) (grant numbers 88887.169260/2018-00 and 88881.132869/2016-1), the Brazilian National Council for Scientific and Technological Development (CNPq) (grant numbers 303318/2013-9, 309438/2016-0, and 402083/2016-4), and the Neotropical Grassland Conservancy.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the landowners for permitting us to conduct the surveys, particularly Daniel Gressler and Vanisse Paiva. We are grateful for support from the Brazilian Army (Campo de Instrução Barão de São Borja; Lieutenant Marcelo Mentges), the Agricultural Experimental Station (EEA/UFRGS; Marcelo Porto Nicola and Renato Levien), and the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA; Cibele Indrusiak and Marcelo Madeira). We also thank Janet W. Reid for reviewing the English version of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.576705/full#supplementary-material
Supplementary Figure 1 | Restoration sites studied in the Brazilian Pampa grasslands: PR1 to PR4 – passive restoration, AR – active restoration. Photos: TWS.
References
Aggemyr, E., Auffret, A. G., Jädergård, L., and Cousins, S. A. O. (2018). Species richness and composition differ in response to landscape and biogeography. Landsc. Ecol. 33, 2273–2284. doi: 10.1007/s10980-018-0742-9
Alexandrino, E. R., Buechley, E. R., Forte, Y. A., Cassiano, C. C., Ferraz, K. M. P. M. B., Ferraz, S. F. B., et al. (2019). Highly disparate bird assemblages in sugarcane and pastures: implications for bird conservation in agricultural landscapes. Neotrop. Biol. Conserv. 14, 169–194. doi: 10.3897/neotropical.14.e37602
Andrade, B. O., Koch, C., Boldrini, I. I., Vélez-Martin, E., Hasenack, H., Hermann, J.-M., et al. (2015). Grassland degradation and restoration: a conceptual framework of stages and thresholds illustrated by southern Brazilian grasslands. Nat. Conserv. 13, 95–104. doi: 10.1016/j.ncon.2015.08.002
Azpiroz, A. B., Isacch, J. P., Dias, R. A., Di Giacomo, A. S., Fontana, C. S., and Palarea, C. M. (2012). Ecology and conservation of grassland birds in southeastern South America: a review. J. F. Ornithol. 83, 217–246. doi: 10.1111/j.1557-9263.2012.00372.x
Buckland, S. T., Anderson, D. R., Burnham, K. P., Laake, J. L., Borchers, D. L., Thomas, L., et al. (2001). Introduction to Distance Sampling: Estimating Abundance of Biological Populations. New York, NY: Oxford University Press.
Buisson, E., Le Stradic, S., Silveira, F. A. O., Durigan, G., Overbeck, G. E., Fidelis, A., et al. (2019). Resilience and restoration of tropical and subtropical grasslands, savannas, and grassy woodlands. Biol. Rev. 94, 590–609. doi: 10.1111/brv.12470
Camilotti, V. L. (2009). Influência da Estrutura Espacial e Da Vegetação Sobre a Assembléia de Aves em Remanescentes Campestres no sul do Brasil. master’s thesis, Universidade Federal do Rio Grande do Sul Porto Alegre, RS.
Cerezo, A., Conde, M. C., and Poggio, S. L. (2011). Pasture area and landscape heterogeneity are key determinants of bird diversity in intensively managed farmland. Biodivers. Conserv. 20:2649. doi: 10.1007/s10531-011-0096-y
Codesido, M., González-Fischer, C., and Bilenca, D. (2013). Landbird assemblages in different agricultural landscapes: a case study in the Pampas of central Argentina. Condor 115, 8–16. doi: 10.1525/cond.2012.120011
Collinge, S. K., Prudic, K. L., and Oliver, J. C. (2003). Effects of local habitat characteristics and landscape context on grassland butterfly diversity. Conserv. Biol. 17, 178–187. doi: 10.1046/j.1523-1739.2003.01315.x
Dauber, J., Hirsch, M., Simmering, D., Waldhardt, R., Otte, A., and Wolters, V. (2003). Landscape structure as an indicator of biodiversity: matrix effects on species richness. Agric. Ecosyst. Environ. 98, 321–329. doi: 10.1016/S0167-8809(03)00092-6
Fahrig, L. (2001). How much habitat is enough? Biol. Conserv. 100, 65–74. doi: 10.1016/S0006-3207(00)00208-1
Fahrig, L. (2003). Effects of habitat fragmentation on biodiveristy. Annu. Rev. Ecol. Evol. Syst. 34, 487–515. doi: 10.1146/annurev.ecolsys.34.011802.132419
Fahrig, L. (2013). Rethinking patch size and isolation effects: the habitat amount hypothesis. J. Biogeogr. 40, 1649–1663. doi: 10.1111/jbi.12130
Favreto, R., and de Medeiros, R. B. (2006). Banco de sementes do solo em área agrícola sob diferentes sistemas de manejo estabelecida sobre campo natural. Rev. Bras. Sementes 28, 34–44. doi: 10.1590/S0101-31222006000200005
Fischer, J., and Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: a synthesis. Glob. Ecol. Biogeogr. 16, 265–280. doi: 10.1111/j.1466-8238.2007.00287.x
Fletcher, R. J., Didham, R. K., Banks-Leite, C., Barlow, J., Ewers, R. M., Rosindell, J., et al. (2018). Is habitat fragmentation good for biodiversity? Biol. Conserv. 226, 9–15. doi: 10.1016/j.biocon.2018.07.022
Haddad, N. M., Brudvig, L. A., Clobert, J., Davies, K. F., Gonzalez, A., Holt, R. D., et al. (2015). Applied ecology: habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1:e1500052. doi: 10.1126/sciadv.1500052
Haire, S. L., Bock, C. E., Cade, B. S., and Bennett, B. C. (2000). The role of landscape and habitat characteristics in limiting abundance of grassland nesting songbirds in an urban open space. Landsc. Urban Plan. 48, 65–82. doi: 10.1016/S0169-2046(00)00044-X
Haynes, K. J., Dillemuth, F. P., Anderson, B. J., Hakes, A. S., Jackson, H. B., Jackson, S. E., et al. (2007). Landscape context outweighs local habitat quality in its effects on herbivore dispersal and distribution. Oecologia 151, 431–441. doi: 10.1007/s00442-006-0600-3
Holl, K. D., and Aide, T. M. (2011). When and where to actively restore ecosystems? For. Ecol. Manage. 261, 1558–1563. doi: 10.1016/j.foreco.2010.07.004
Kroll, A. J., Ren, Y., Jones, J. E., Giovanini, J., Perry, R. W., Thill, R. E., et al. (2014). Avian community composition associated with interactions between local and landscape habitat attributes. For. Ecol. Manage. 326, 46–57. doi: 10.1016/j.foreco.2014.04.011
Kuznetsova, A., Brockhoff, P. B., and Christensen, R. H. B. (2017). {{lmerTest}} package: tests in linear mixed effects models. J. Stat. Softw. 82:13. doi: 10.18637/jss.v082.i13
Lee, M. B., and Carroll, J. P. (2014). Relative importance of local and landscape variables on site occupancy by avian species in a pine forest, urban, and agriculture matrix. For. Ecol. Manage. 320, 161–170. doi: 10.1016/j.foreco.2014.03.017
Lindenmayer, D. B., Knight, E. J., Crane, M. J., Montague-Drake, R., Michael, D. R., and MacGregor, C. I. (2010). What makes an effective restoration planting for woodland birds? Biol. Conserv. 143, 289–301. doi: 10.1016/j.biocon.2009.10.010
Lockhart, J., and Koper, N. (2018). Northern prairie songbirds are more strongly influenced by grassland configuration than grassland amount. Landsc. Ecol. 33, 1543–1558. doi: 10.1007/s10980-018-0681-5
Manning, A. D., Lindenmayer, D. B., and Nix, H. A. (2004). Continua and Umwelt: novel perspectives on viewing landscapes. Oikos 104, 621–628. doi: 10.1111/j.0030-1299.2004.12813.x
Öckinger, E., Lindborg, R., Sjödin, N. E., and Bommarco, R. (2012). Landscape matrix modifies richness of plants and insects in grassland fragments. Ecography 35:9. doi: 10.1111/j.1600-0587.2011.06870.x
Overbeck, G. E., Hermann, J.-M., Andrade, B. O., Boldrini, I. I., Kiehl, K., Kirmer, A., et al. (2013). Restoration ecology in Brazil — Time to step out of the forest. Nat. Conserv. 11:4. doi: 10.4322/natcon.2013.015
Pretelli, M. G., Isacch, J. P., and Cardoni, D. A. (2018). Species-Area relationships of specialist versus opportunistic Pampas grassland birds depend on the surrounding landscape matrix. Ardeola 65, 3–23. doi: 10.13157/arla.65.1.2018.ra1
Qgis Development Team (2016). QGIS Geographic Information System. Open Source Geospatial Foundation Project. London: QGIS Development Team.
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rio Grande do Sul (2014). Lista das espécies da fauna ameaçadas de extinção no estado do Rio Grande do Sul. Instrução Normativa no. 3. Decreto no. 51.797. Porto Alegre: Diário Oficial do Estado do Rio Grande do Sul, Secretaria do Meio Ambiente.
Rodewald, A. D., and Yahner, R. H. (2001). Influence of landscape composition on avian community structure and associated mechanisms. Ecology 82:12. doi: 10.2307/2680167
Roesch, L. F. W., Vieira, F. C. B., Pereira, V. A., Schünemann, A. L., Teixeira, I. F., Senna, A. J. T., et al. (2009). The Brazilian Pampa: a fragile biome. Diversity 1, 182–198. doi: 10.3390/dl020182
Shahan, J. L., Goodwin, B. J., and Rundquist, B. C. (2017). Grassland songbird occurrence on remnant prairie patches is primarily determined by landscape characteristics. Landsc. Ecol. 32:18. doi: 10.1007/s10980-017-0500-4
Silva, T. W., Dotta, G., Gressler, D. T., and Fontana, C. S. (2015). Habitat use by grassland birds in natural areas and soybean fields in southern Brazil and Uruguay. Wilson J. Ornithol. 127:10. doi: 10.1676/wils-127-02-212-221.1
Silva, T. W., and Fontana, C. S. (2020). Success of active restoration in grasslands: a case study of birds in southern Brazil. Restor. Ecol. 28, 512–518. doi: 10.1111/rec.13111
Silva, T. W., Lindenmayer, D. B., and Fontana, C. S. (2019). Passive restoration contributes to bird conservation in Brazilian Pampa grasslands. J. Field Ornithol. 90:14. doi: 10.1111/jofo.12316
Thomas, L., Buckland, S. T., Rexstad, E. A., Laake, J. L., Strindberg, S., Hedley, S. L., et al. (2010). Distance software: design and analysis of distance sampling surveys for estimating population size. J. Appl. Ecol. 47, 5–14. doi: 10.1111/j.1365-2664.2009.01737.x
Vieira, M., de, S., Bonilha, C. L., Boldrini, I. I., and Overbeck, G. E. (2015). The seed bank of subtropical grasslands with contrasting land-use history in southern Brazil. Acta Bot. Brasilica 29:10. doi: 10.1590/0102-33062015abb0026
Waldén, E., Öckinger, E., Winsa, M., and Lindborg, R. (2017). Effects of landscape composition, species pool and time on grassland specialists in restored semi-natural grasslands. Biol. Conserv. 214, 176–183. doi: 10.1016/j.biocon.2017.07.037
Wentworth, K. L., Brittingham, M. C., and Wilson, A. M. (2010). Conservation reserve enhancement program fields: benefits for grassland and shrub-scrub species. J. Soil Water Conserv. 65, 50–60. doi: 10.2489/jswc.65.1.50
Whitaker, D. M., and Warkentin, I. G. (2010). Spatial ecology of migratory passerines on temperate and boreal forest breeding grounds. Auk 127:14. doi: 10.1525/auk.2010.127.3.471
With, K. A., and Crist, T. O. (1995). Critical thresholds in species’ responses to landscape structure. Ecology 76:14. doi: 10.2307/2265819
Zhang, Y. (2013). Likelihood-based and bayesian methods for tweedie compound poisson linear mixed models. Stat. Comput. 23, 743–757. doi: 10.1007/s11222-012-9343-7
Keywords: active restoration, grassland birds, landscape, passive restoration, SESA Grasslands
Citation: Silva TW and Fontana CS (2021) Remnants of Native Vegetation Surrounding Do Not Affect the Diversity and Density of Birds in Brazilian Grassland-Restoration Sites. Front. Ecol. Evol. 8:576705. doi: 10.3389/fevo.2020.576705
Received: 26 June 2020; Accepted: 11 December 2020;
Published: 07 January 2021.
Edited by:
Manuel B. Morales, Autonomous University of Madrid, SpainReviewed by:
Denis Kupsch, University of Göttingen, GermanyJuliana Hipólito, National Institute of Amazonian Research (INPA), Brazil
Copyright © 2021 Silva and Fontana. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thaiane Weinert da Silva, dGhhaWFuZXdzQGdtYWlsLmNvbQ==