 Francesca Alderotti1
Francesca Alderotti1 Cecilia Brunetti1,2*
Cecilia Brunetti1,2* Giovanni Marino2
Giovanni Marino2 Mauro Centritto2
Mauro Centritto2 Francesco Ferrini1,2
Francesco Ferrini1,2 Cristiana Giordano3
Cristiana Giordano3 Massimiliano Tattini2
Massimiliano Tattini2 Bárbara Baêsso Moura1,4
Bárbara Baêsso Moura1,4 Antonella Gori1,2
Antonella Gori1,2- 1Section Woody Plants, Department of Agriculture, Food, Environment and Forestry, University of Florence, Florence, Italy
- 2Institute for Sustainable Plant Protection, National Research Council of Italy, Sesto Fiorentino, Italy
- 3Institute for BioEconomy, National Research Council of Italy, Sesto Fiorentino, Italy
- 4Research Institute on Terrestrial Ecosystems, National Research Council of Italy, Sesto Fiorentino, Italy
The shrub Cistus incanus L. is well-adapted to Mediterranean conditions thanks to its morpho-anatomical, physiological and biochemical traits. However, its distribution and survival in coastal dunes will be likely threatened by ongoing runaway climate change. We investigated how the harsh climatic conditions generated by the 2015 summer heat wave triggered specific anatomical, physiological, and biochemical responses of this species in its natural environment. These adjustments were compared to those measured in summer 2014. The drier and hotter conditions of summer 2015 determined an increment in leaf lamina thickness, due to a greater palisade parenchyma, thus leading to an increase in the whole leaf mass per area. These morphoanatomical adjustments enhanced leaf resistance against dehydration, optimized carbon assimilation, and delayed leaf senescence. In addition, the higher amount of secondary metabolites detected for 2015, in particular tannins and monoterpenes, contributed to the maintenance of physiological performances even under hotter and drier conditions, preventing oxidative stress through the consumption of excess reducing power. In conclusion, our study offers new evidence on the integration of morphophysiological and metabolic adjustments of this species growing in its natural habitat to cope with ongoing climate change.
Introduction
Mediterranean climate is characterized by mild and humid winters and hot and dry summers, with most of the rainfalls concentrated in the cold season (Lionello et al., 2006). Summer drought is considered the primary constraint affecting both productivity and distribution of Mediterranean vegetation due to a combination of high irradiance, high temperatures and water scarcity (Gratani et al., 2013; Flexas et al., 2014).
Mediterranean maquis vegetation mostly consists of evergreen sclerophylls with a deep root system and semi-deciduous shrubs, which reduce transpiring surface during stressful periods (Galmés et al., 2007; Medrano et al., 2009). Shrub species, in particular, are a widespread life form in the coastal ecosystem and their potential resilience to environmental change deserves special attentions (Arena et al., 2011; Galle et al., 2011; Puglielli et al., 2017a). Mediterranean shrubs display a vast array of strategies to cope with severe climatic constraints (Lefi et al., 2004; Sardans and Peñuelas, 2013), which may in part correlate with the large biological diversity encountered in Mediterranean areas suffering from most severe environmental pressures (Van Der Plas et al., 2016). Evergreen species usually display a conservative resource use strategy (David et al., 2007), characterized by low net photosynthesis and relative growth rate, and the utilization of the fresh assimilated carbon for leaf construction (Sardans and Peñuelas, 2013). By contrast, semi-deciduous species display a higher physiological plasticity compared to the evergreen counterparts, with a decline in photosynthetic performances during central hours in summer season, when high solar irradiance occurs in concomitance with severe reduction in soil water availability and excessive air temperature (Di Ferdinando et al., 2014).
Among these metabolic adjustments, the increase in polyphenol contents in stressed plants has been linked to improve photoinhibition tolerance and, in general, to the protection of photosynthetic organs from oxidative damages (Brunetti et al., 2015). Mediterranean shrubs also emit large amounts of biogenic volatile organic compounds (BVOCs), among which mono- and sesqui-terpenes are the most abundant. These compounds play prominent functions against biotic and abiotic stresses as well as in mediating ecological interactions (Loreto and Schnitzler, 2010; Niinemets et al., 2013; Loreto et al., 2014; Mu et al., 2018). Specifically, monoterpenes have been shown to protect leaves against oxidative and thermal damages, decreasing the rate of formation of reactive oxygen species (ROS) that can damage the photosynthetic apparatus (Loreto et al., 1998, 2014). It is known that water stress may result in increased monoterpene emission (Ormeño et al., 2007) or in a significant change in the blend composition of stored monoterpenes (Brilli et al., 2013).
Cistus L. is a genus of dicotyledonous plants colonizing open areas of stony and infertile soils (Papaefthimiou et al., 2014). Among Cistus spp., Cistus × incanus L. (pink rock-rose, hereby C. incanus) belongs to the autochthonous Tertiary Mediterranean flora (Quézel, 1985). This species is classified as a summer drought semi-deciduous species, shedding a high percentage of leaves and branches and increasing the mass investments per leaf area unit during drought (LMA, leaf mass per area) (Catoni et al., 2012; De Dato et al., 2013; Grant et al., 2015). The semi-deciduousness allows this species to be well-adapted to summer drought (Correia and Ascensao, 2016; Puglielli, 2019). In addition, also seasonal dimorphism, expressed both in the leaf morphoanatomical traits as well as in the wood properties, may be considered another important adaptation of this species to the Mediterranean harsh climate (Aronne and De Micco, 2001; De Micco and Aronne, 2009; Arena et al., 2013). Indeed, summer leaf anatomy is characterized by a crimped lamina partially rolled to form crypts in the lower surface and a thick palisade parenchyma in the mesophyll tissue (Aronne and De Micco, 2001; Rotondi et al., 2003).
Despite being well-adapted to multiple stress factors, C. incanus distribution and survival in coastal dune areas will be likely negatively affected by the ongoing climatic change conditions (Nogués et al., 2015; Gori et al., 2019). In this respect, very few field investigations have been conducted on the effects of drought and heat stress on this Mediterranean shrub (Gratani et al., 2008; De Micco and Aronne, 2009; Gratani et al., 2018; Parra and Moreno, 2018; Gori et al., 2019). Furthermore, morpho-physiological and metabolic responses of this species to extreme drought remain poorly investigated despite the increasing frequencies of heat waves hitting the Mediterranean region as a consequence of climate change.
In this context, the contrasting climatic conditions of the summers 2014 and 2015 provided an optimum opportunity to study the modifications in this maquis shrub behavior in response to ongoing climate change in its natural environment. In fact, the summer 2014 showed irregular rainfalls and moderate drought, while the summer 2015 was one of the driest and hottest in Europe in the last 70 years as a result of an extreme heatwave that lasted about 30 days until the end of July (Russo et al., 2015).
We took advantage of contrasting climatic conditions of summers 2014 and 2015 to test if changes in morpho-anatomical traits allow to maintain physiological performances and enhance the biosynthesis of secondary metabolites in C. incanus. We hypothesized that semi-deciduous Mediterranean shrubs species adapted to harsh environment, such as C. incanus, are able to face drought and heat stress thank to a coordination of morpho-anatomical, physiological and biochemical traits. We expected that higher temperature in 2015 may have promoted a major investment in leaf construction and boosted the production of secondary metabolites, thus reducing water loss and damages to photosynthetic apparatus caused by oxidative stress. Overall, this study provides the substantial contribution of secondary metabolites, both condensed tannins and monoterpenes, to the suite of traits conferring high plasticity to C. incanus and, consequently to its high resilience against extreme environmental pressures associated to climate change.
Materials and Methods
Plant Material, Study Area, Sampling Design and Meteorological Data
In May 2014, eight healthy individuals of C. incanus were selected in the population previously described in Gori et al. (2019) and screened for size and uniformity. C. incanus plants were 0.6 ± 0.07 m height with a canopy area of around 0.6 ± 0.08 m2. The study was performed over 2 consecutive years on cloudless days in summer 2014 and 2015 (8–9 July 2014 and 14–15 July 2015) on the coastal sand dunes located in Castiglione della Pescaia (GR, Italy, 42° 46′ N, 10° 53′ E).
The diurnal time courses of leaf water potential was monitored at five different sampling hours, at 4:00 (predawn, PD), 8:00, 12:00 (midday, MD), 15:00, and 18:00, while gas exchanges, chlorophyll fluorescence parameters, chlorophyll index and epidermal flavonol index at 8:00, 12:00, 15:00, and 18:00. Samples for polyphenol analysis and Biogenic Volatile Organic Compounds (BVOCs) were collected between 12:30 and 14:00. Air temperature (T), precipitation (P) and global irradiance (Gi) were recorded every hour by the weather station “Grosseto” (Italy) and provided from the regional archive of the LaMMA Consortium1.
Water Relations
Two leaves per plant were collected in the field and transported to the laboratory in sealed zip-lock, tared plastic bags stored in a fridge bag. The leaves were weighed to determine fresh mass (FW) and then allowed to hydrate until saturation (constant weight) for 48 h in darkness to determine turgid mass (TW). Then, leaves were dried in an oven at 80°C for 48 h to determine dry mass (DW). Finally, Relative Water Content (RWC) was calculated using the following equation:
Leaf water potentials (Ψw) were directly measured in field, after the cut of two twigs per plant, using a Scholander-type pressure chamber (PMS Instruments, Corvallis, OR). The measurements of RWC and Ψw obtained from the same plant were combined to make an individual replicate.
Morphological and Anatomical Measurements
From the upper part of the canopy 10 adult leaves per plant were sampled at midday to measure LMA by the calculation of the dry mass (drying the leaves in oven at 80°C for 48 h) to leaf area ratio (mg cm–2). One twig per plant was collected and transported to the laboratory for anatomical measurements. Light microscopy segments of leaf tissue were taken from the basal-mid-lamina regions of eight adult leaves and fixed for 2 h at room temperature in 0.2 M phosphate buffer (pH 7.2) with 2.5% glutaraldehyde (Sigma Aldrich). Then, they were washed twice in the same buffer and post-fixed in 2% osmium tetroxide (Sigma Aldrich) for additional 2 h. Following dehydration in a graded ethanol series (30, 40, 50, 60, 70, 80, 95, and 100%), the samples were gradually embedded in Spurr resin (Sigma Aldrich) and polymerized at 70°C for 24 h (Spurr, 1969). Semi-thin sections, 1–2 μm, were affixed to glass slides and observations were carried out in a Leica DM LB2 Light Microscope (Leica Microsystem, Germany).
Gas Exchanges and Chlorophyll Fluorescence
Measurements of gas exchanges and photosystem II maximum efficiency (ΦPSII) were performed on two fully expanded and healthy leaves per plant, selected in the most illuminated part of the canopy, utilizing a LI-6400 portable photosynthesis system (Li-Cor, Lincoln, NE, United States) equipped with a cuvette of 2 cm2. Photosynthesis (Pn), stomatal conductance (gs), intercellular CO2 concentration (Ci) and ΦPSII were measured at (CO2) of 400 μmol mol–1 and at the photosynthetic photon flux density recorded in the environment. The integral of daily photosynthesis (Pn), calculated using Sigma Plot 11.0 (Systat Software, Inc., San Jose, CA), has been employed to quantify the amount of carbon fixed during the day. Then, mesophyll conductance to CO2 (gm) was calculated using the variable J method (Harley et al., 1992) as follows:
where Γ, representing the CO2 compensation point to photorespiration, was obtained from data reported in Galle et al. (2011), while light respiration (Rd) was calculated using the Kok method at PPFD steps of 150, 100, 80, 60, and 30 μmol m–2s–1 (Kok, 1948).
Electron transport rate (JF) was calculated from chlorophyll fluorescence:
where ΦPSII is the actual photochemical efficiency of Photosystem II (Genty et al., 1989), the partitioning factor (β) between photosystems I and II was considered to be 0.5 and leaf absorbance (α) 0.84.
Moreover, we use the ratio JF/Pn as an indicator of the excess of electron transport which can be diverted to other alternative electron sinks rather than photosynthesis (Arena and Vitale, 2018; Brunetti et al., 2018).
After gas exchanges, Fv/Fm was measured on the same leaves using a portable chlorophyll fluorescence system (Handy Pea, Hansatech, Norflolk, United Kingdom). Before measurements, leaves were adapted to the dark for 30 min. Gas exchange and chlorophyll fluorescence measurements obtained from the same plant were combined to make an individual replicate.
Biochemical Parameters
Non-destructive Determination of Chlorophyll and Epidermal Flavonols
At each daily sampling hour, four leaves per plant were measured with a Dualex® device (Dualex® Research, FORCE-A®, Orsay, France) on adaxial and abaxial side. Chlorophyll index (Chli) and epidermal Flavonol index (Flavi) were obtained as described in Agati et al. (2016). In particular, the Chli was calculated as the average of the adaxial and abaxial measurements, while the Flavi was calculated as the sum of both measurements. All measurements obtained from the same plant were combined to make an individual replicate.
HPLC Quantification of Polyphenols
Two leaves per plant were collected, immediately frozen in liquid nitrogen, stored at −80°C and then lyophilized. In detail, lyophilized samples (150 mg of DW) were extracted twice with 5 mL of ethanol/water (75/25) adjusted at pH2.5 with formic acid and the supernatant partitioned with 3 × 5 mL of n-hexane. The ethanol fraction was reduced to dryness, and the residue was rinsed with 1 mL of methanol/water (90/10). Aliquots of 10μL were injected into the Flexar liquid chromatography equipped with a quaternary 200Q/410 pump and an LC 200 diode array detector (DAD) (all from Perkin Elmer®, Bradford®, CT, United States). Identification and quantification of these metabolites was carried out using retention times and UV spectral characteristics of authentic standards, as well as based on literature data (Gori et al., 2016). The total polyphenols content (PolTot) consists in the sum of individual polyphenols detected, namely tannins (Tan) and flavonols (Flav).
GC-MS Quantification of Biogenic Volatile Organic Compounds (BVOCs)
The emission of BVOCs was measured on the same leaves used for gas exchange analyses by partially diverting the outlet of the cuvette of the Li-Cor system described above into a silico-steel cartridge packed with 200 mg of Tenax (Agilent, Cernusco sul Naviglio, Italy). A volume of 4 dm3 of air was pumped through the trap at a rate of 200 cm3 min–1 utilizing an environmental air sampling equipment (A.P. BUCK, Inc., Orlando, FL). Prior to the first enclosure of the leaves, blank samples from the empty cuvettes were taken every sampling day. The cartridge was analyzed using a Perkin Elmer Clarus 580 gas chromatograph coupled with a Clarus 560 Mass-Selective-Detector and a thermal desorber TurboMatrix (Perkin Elmer Inc., Waltham, MA, United States) operating for 25 min at 250°C. The desorbed compounds were separated in a 30-m Elite-5-MS capillary column. The temperature of the column was first maintained at 40°C (5 min), then increased with a 5°C min–1 175 ramp to 250°C, and maintained at 250°C for 2 min. BVOCs were identified using the NIST library provided with the GC/MS Turbomass software. The authentic standards of α-pinene, β-pinene and myrcene (Sigma Aldrich, Milan, Italy) were used to prepare calibration curves as well as to compare the peak retention time and the peak fragmentation of monoterpenes found in the samples. Authentic standards of other compounds collected in the traps (tujene, α-phellandrene, and β-phellandrene) were not available, therefore the amount of these compounds was calculated using the calibration curve of α-pinene. Total BVOC emission (BVOCsTot) consists in the sum of individual monoterpenes detected in the samples.
Statistical Analysis
Statistical analysis was performed using the SPSS software program (SPSS, Inc., Chicago, IL, United States). To test the daily trend of parameters, we performed a two way ANOVA followed by Tukey’s post-hoc test. When a mean value was reported for each summer, Student’s t-tests were performed to evaluate differences between the two years. Linear regression analyses were performed to determine the effect of air temperature on gas exchanges and chlorophyll fluorescence parameters (Pn, gs, Fv/Fm and ΦPSII) and water relations (Ψw and RWC) considering the data collected in four different sampling hours (8:00, 12:00, 15:00, and 18:00) and on total polyphenols content (PolTot) and BVOCs emission considering the data collected at 12:00. The linear regression analysis was also applied to determine the differential contribution of the morpho-anatomical traits (upper and lower epidermis, palisade and spongy parenchyma) to the total leaf thickness (LT) and to the gas exchange and chlorophyll fluorescence parameters (Pn, gs, Fv/Fm, and ΦPSII). With the same set of data, a principal component analysis (PCA) was applied to gas exchange and chlorophyll fluorescence parameters (Pn, gs, Fv/Fm, and ΦPSII), water relations (RWC and Ψw) and morpho-anatomical (leaf thickness) or total polyphenols content (PolTot), biogenic organic compounds (BVOCs) emission and morpho-anatomical (leaf thickness) in order to distinguish the trait syndromes between the year of sampling. The PCA was calculated based on a correlation matrix using OriginPro 2020 software.
Results
Meteorological Data
In July 2014, the average maximum temperature (Tmax) was 29°C, almost 5°C lower than that recorded in July 2015. Average minimum temperatures (Tmin) were 17.7 and 21.1°C in July 2014 and 2015, respectively (Table 1). Hence, summer 2015 was unusually dry and hot as a consequence of a severe heat wave that hit the Mediterranean regions for about 30 days between the end of June and July (Russo et al., 2015). During the 2 months prior measurements cumulative rainfall was 84 mm in 2014 and 29 mm in 2015 (Table 1). During the days of measurements, temperatures (Figure 1A) and global irradiance (Figure 1B) had a typically daily trend, showing higher values during the central hours. The daily irradiance did not change between the two summers (Figure 1B). Whereas, in 2014 the daily time course of temperature was consistently lower than in 2015 (Figure 1A), with minimum and maximum temperatures of 18°C and 30°C in 2014, and 22°C, and 33°C in 2015, respectively (Figure 1A).

Table 1. Meteorological data of the two study years.
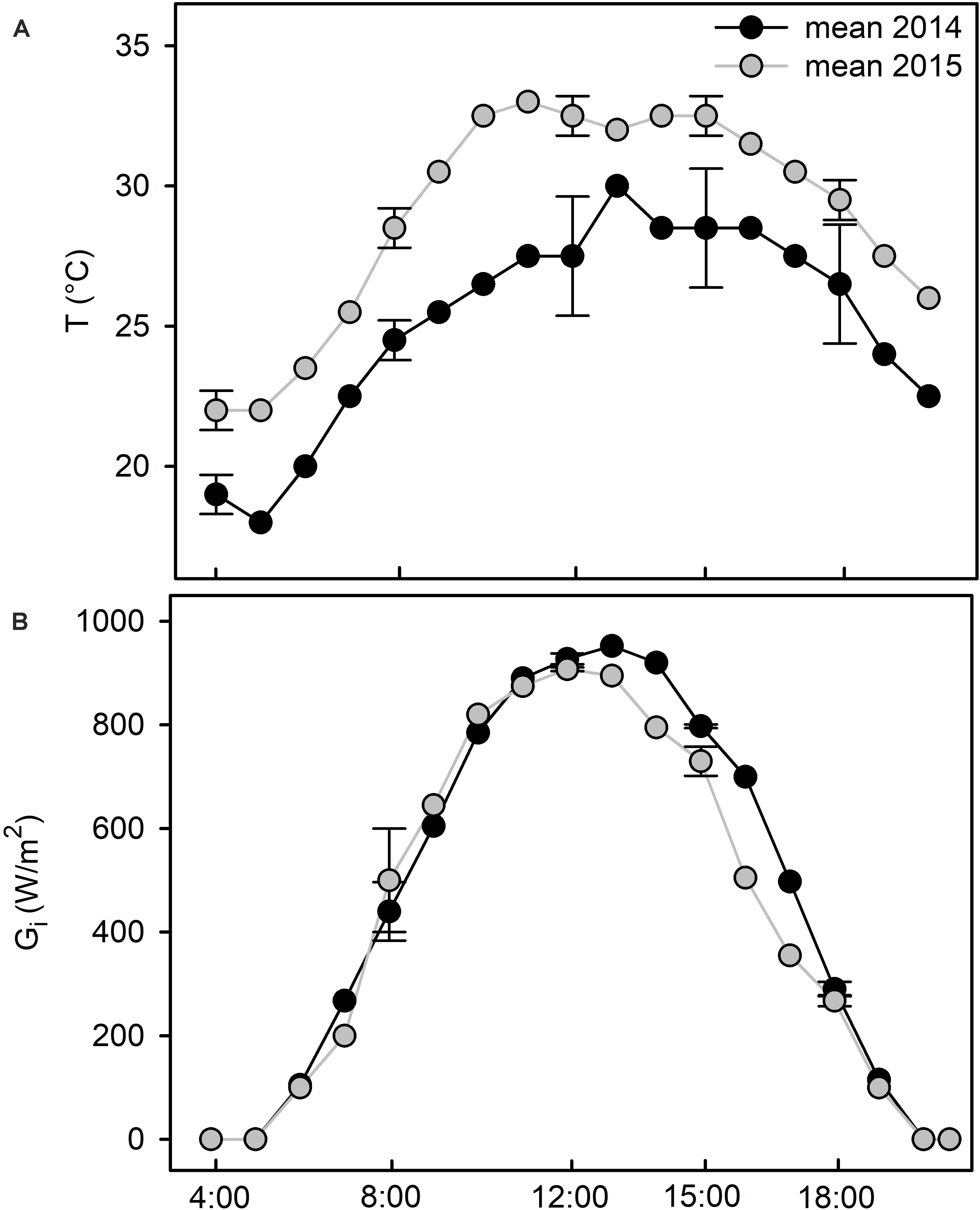
Figure 1. Time course of average air temperatures (T,°C) (A) and of global irradiance (Gi, Wm–2) (B) of the sampling days in July 2014 (black line) and July 2015 (gray line). Data of the weather station “Grosseto” (Italy) from the regional archive of the LaMMA Consortium (http://lamma.rete.toscana.it).
Morpho-Anatomical Leaf Traits
The microscopy analysis revealed significant variations among the samples collected in summer 2014 and 2015 (Figure 2A and Table 2). In both years, leaves showed a typical structure of a heliophylous plant: a dorso-ventral orientation, a high mesophyll density, with a well-developed palisade consisting of two/three layers of elongate-prismatic cells and a spongy parenchyma with a few airspaces (Figure 2A). In the leaves collected in 2015, the vacuolar accumulation of tannins, as evidenced through osmium staining, as spherical deposits or adjacent to the internal surface of the vacuolar membrane; condensed tannins fill almost completely the vacuoles of adaxial epidermis and palisade parenchyma cells, whereas occur in fewer cells or in smaller quantities in spongy and abaxial epidermis cells (Figure 2A). In addition, leaves collected in 2015 showed a significant increase in the thickness of almost all leaf tissues compared to those collected in 2014, i.e., increases of 44.2% (t-test; p ≤ 0.001) in the palisade and of 28.2% (t-test; p = 0.024) in the lower epidermis (Table 2). A slight, but not significant increment, was observed also in the spongy parenchyma. The palisade parenchyma was the leaf tissue that mostly contributed to increase the leaf thickness in 2015 presenting a higher coefficient of determination (R2 = 0.66) when compared to the other tissues (spongy mesophyll R2 = 0.35, upper epidermis R2 = 0.44 and lower epidermis = regression not significant) (Figure 2B and Supplementary Table S1B). A strong, significant increase in LMA (+34.5%) (t-test; p = 0.002) was observed in 2015, as LMA ranged from 11.2 mg cm–2 in July 2014 to 17.1 mg cm–2 in July 2015 (Table 2).
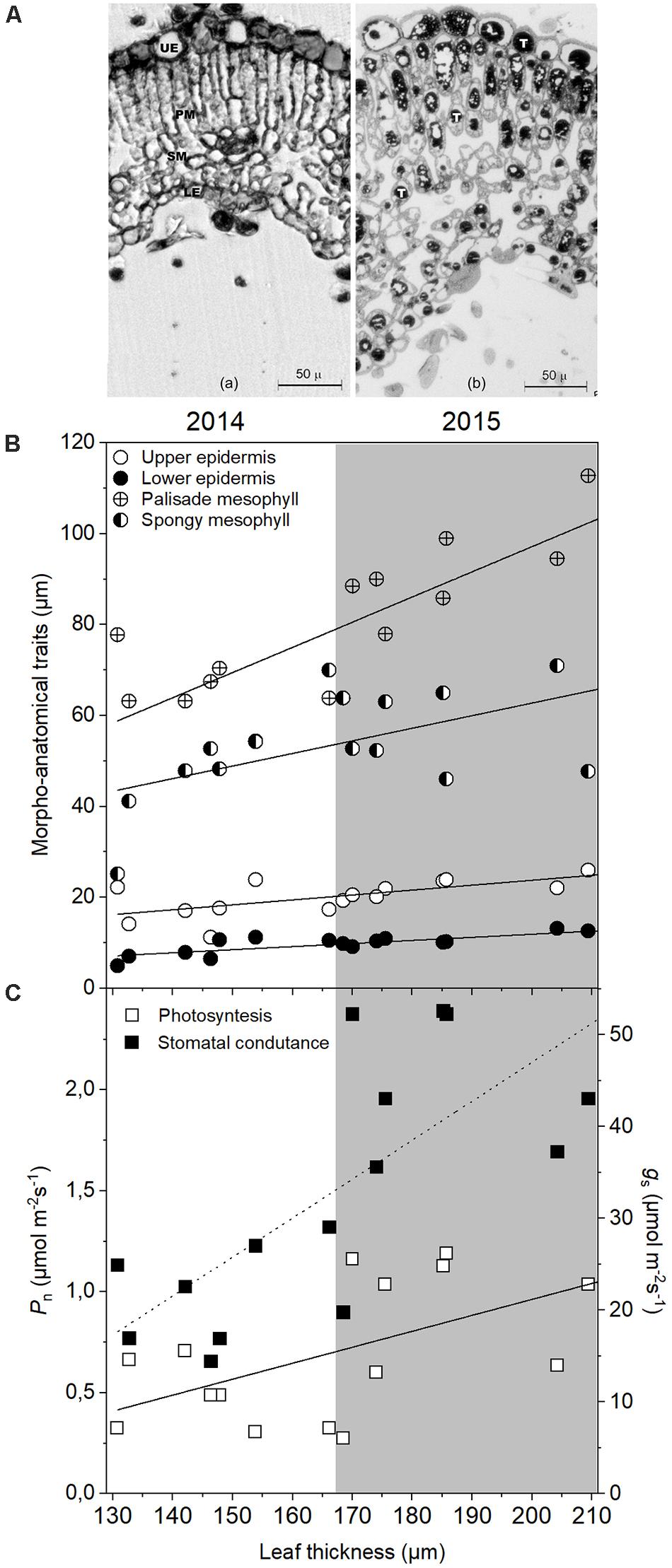
Figure 2. Transverse cross sections of osmium-stained C. incanus leaves collected in July 2014 (a) and 2015 (b) illustrating the difference in in thickness of palisade parenchyma and epidermal cells between the plants collected in the two different years (A). Condensed tannin deposits are shown in the vacuoles of cells. UE, upper epidermis; PM, palisade mesophyll; SM, spongy mesophyll; LE, lower epidermis; T, tannin. Bars = 50 μm. Linear regressions between leaf thickness and the other morpho-anatomical traits (B). Linear regressions between leaf thickness and photosynthesis (Pn) and stomatal conductance (gs) (C). Detailed analysis is presented in Supplementary Table S1.
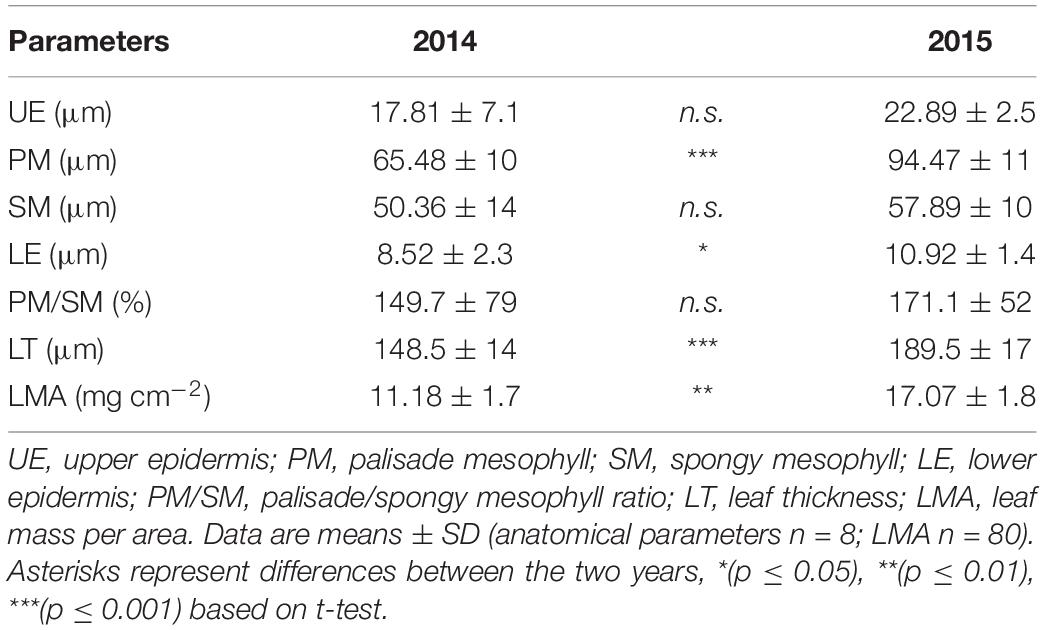
Table 2. Morpho-anatomical traits of C. incanus leaves in July 2014 and 2015.
Water Relations and Gas Exchanges
Plants had a better water status in 2014 than in 2015, as shown by the significantly higher values of RWC recorded at all sampling hours (two way ANOVA; p < 0.001) (Table 3). As expected, RWC showed the maximum values at PD (∼70% in 2014 vs. ∼60% in 2015) in both years. In 2014, after 8:00 RWC decreased significantly (two way ANOVA; p < 0.001) never recovering to the morning value during the day. Similarly, in 2015, RWC declined significantly after 8:00 and, following the depression registered during the hottest hours of the day, showed a significant recovery to the morning values. A significant year-sampling time interaction was revealed for water potential (Ψw) (two way ANOVA; p < 0.001). Ψw was significant lower in 2015 than in 2014 (two way ANOVA; p < 0.001), except for the late-afternoon value 2014 (two way ANOVA; p = 0.463) (Table 3). This parameter followed a typically daily pattern, showing marked reduction during the central hours of the day in both years. It is worth noting that midday Ψw fell to a minimum of −4.8 MPa in 2015, compared to the 2014 value which reached −3.2 MPa (Table 3). Both parameters of water status (RWC and Ψw) presented a negative relation with the air temperatures (Supplementary Table S1A). A significant year-sampling time interaction was revealed for net photosynthetic rate (Pn) (two way ANOVA; p < 0.001). Pn dropped from morning to midday in 2015, ranging from 2.92 ± 0.80 μmol m–2 s–1 to 0.99 ± 0.24 μmol m–2 s–1 (-66% two way ANOVA; p < 0.001), while in 2014 it did not change in the same time slot (two way ANOVA; p = 0.064). Differences in Pn diurnal time courses emerged between the two summers after 12:00. In fact, while in 2014 Pn values increased from 12:00 to 15:00 (+85% two way ANOVA; p < 0.001) and subsequent decreased at 18:00 (-65% two way ANOVA; p < 0.001), in 2015, Pn did not change from 12:00 to late afternoon (two way ANOVA; p > 0.05) (Figure 3A). The daily integral of photosynthesis (Pn) did not show significant differences between the two summers (0.12 ± 0.03 g CO2 d–1m–2 in 2014 and 0.11 ± 0.01 g CO2 d–1m–2 in 2015) (t-test; p = 0.55) (Figure 3B). Stomatal conductance (gs) was significantly lower in 2014 than in 2015 at all sampling hours (two way ANOVA; p < 0.001), except for values recorded at 18:00 (two way ANOVA; p = 0.324) (Figure 4A). However, gs had a very similar trend in the two summers, with the highest values recorded at 8:00 and 15:00 and the lowest at 12:00 and 18:00 (Figure 4A). A significant linear regression between the leaf thickness and the Pn and gs was observed (Figure 2C and Supplementary Table S1A) with gs presenting the higher coefficient of determination (R2 = 0.59) when comparing to Pn (R2 = 0.28). However, the relationship between those parameters with the air temperature was only significant for the gs (Supplementary Table S1A). Similarly to gs, mesophyll conductance calculated at 12:00 (gmMD), showed significantly higher values in 2015 than in 2014 (t-test; p = 0.05) (Figure 4A). In 2014, Ci did not change from morning to 12:00 in 2014, while a significant increment was observed for this parameter in 2015 (Figure 4B). Then while in 2014 Ci strongly dropped from 12:00 to 15:00 (-24% two way ANOVA; p < 0.001) and partially recovered in the late afternoon, in 2015 this parameter slowly returned to the morning value at 18:00 (Figure 4B). In addition, significant year-sampling time interaction was revealed for both Ci and JF/Pn (two way ANOVA; p < 0.001). The morning and midday values of the JF/Pn ratio were similar and did not change between the 2 years (two way ANOVA; p > 0.05) (Figure 4C). However, in 2015, the JF/Pn ratio increased significantly during the afternoon and resulted 86 and 68% higher than in 2014 at 15:00 (two way ANOVA; p < 0.001) and at 18:00 (two way ANOVA; p < 0.001), respectively.
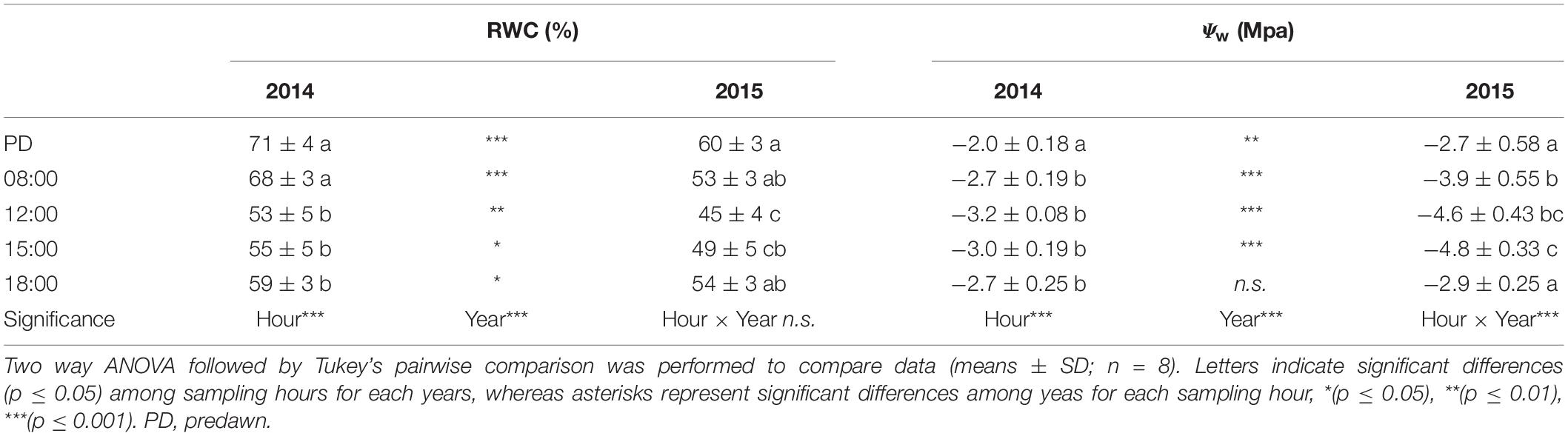
Table 3. Time course of relative water content (RWC) and of water potential (Ψw) of C. incanus plants in July 2014 and 2015.
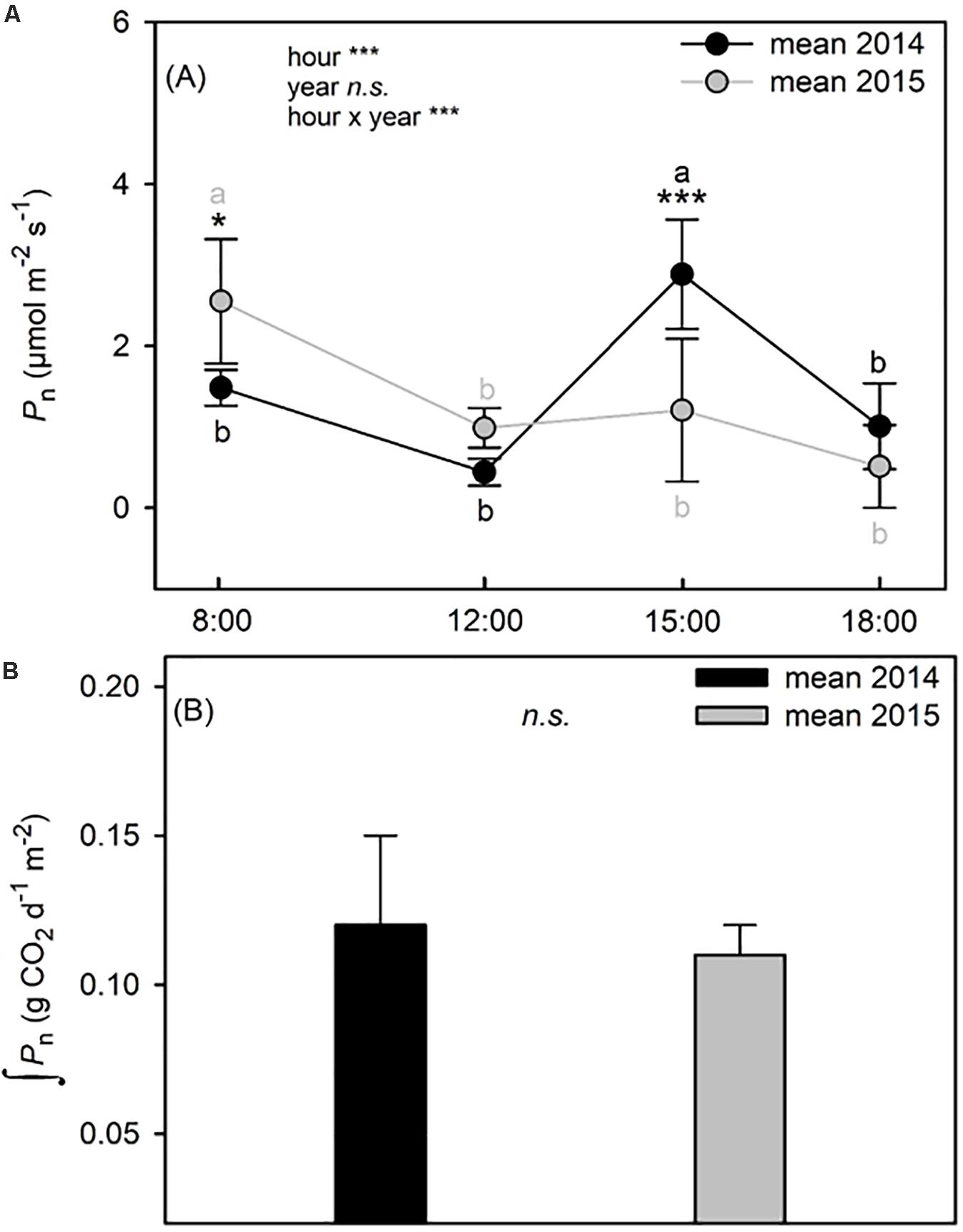
Figure 3. Time course of net photosynthetic rate (Pn) (A) and integration of daily net photosynthetic rate (Pn) (B) of C. incanus leaves in July 2014 and 2015. Two way ANOVA followed by Tukey’s pairwise comparison was performed to compare data (means ± SD; n = 8) in (A). Letters indicate significant differences (p ≤ 0.05) among sampling hour for each year, whereas asterisks represent significant differences among years for each sampling hour, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001). The statistic adopted in panel B was a Student’s t-tests, asterisks represent differences between the two years, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001).
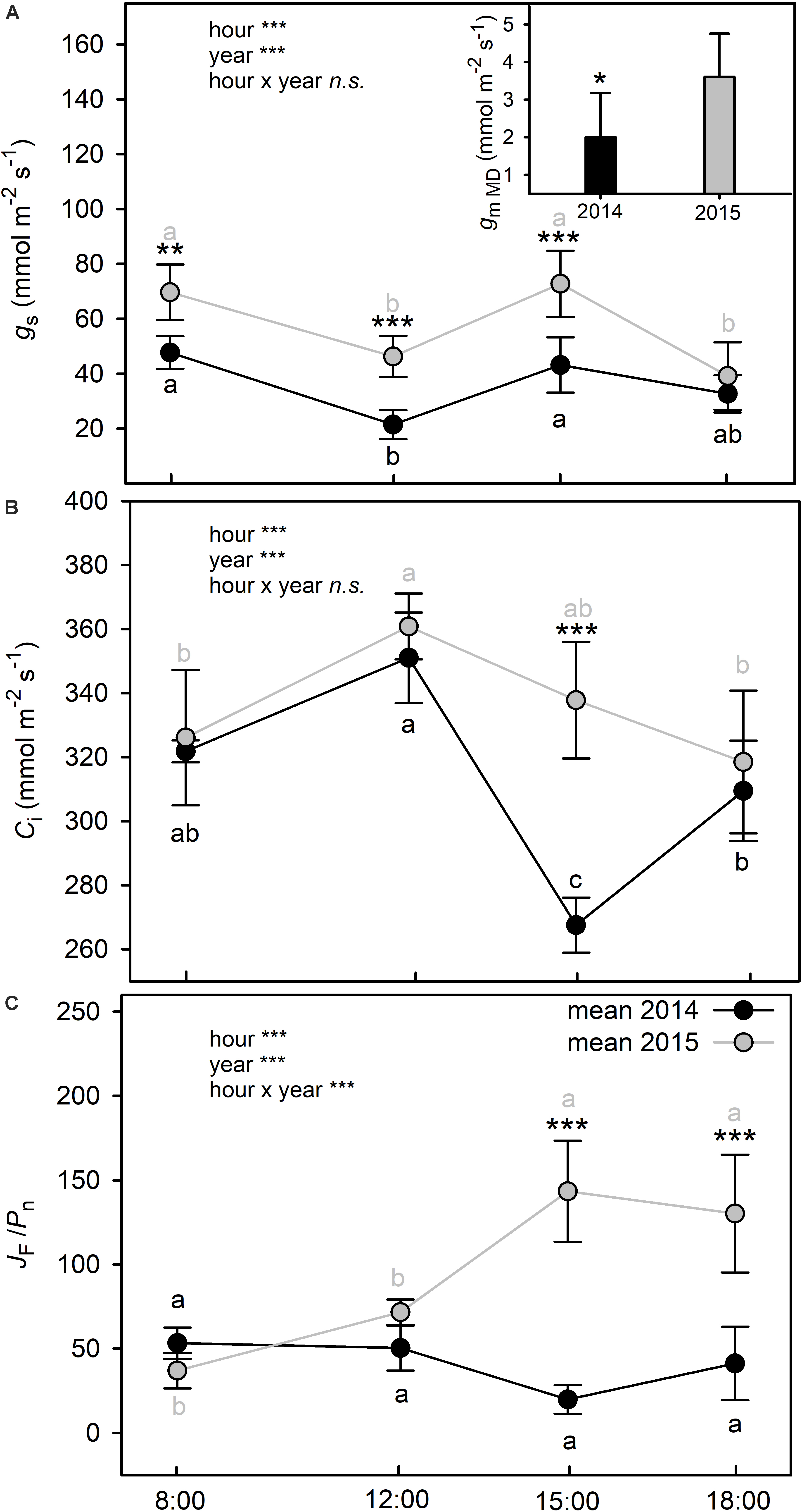
Figure 4. Time course of stomatal conductance (gs) (A), intercellular CO2 concentration (Ci) (B), and electron transport rate (JF) to net photosynthetic rate (Pn) ratio (C) of C. incanus leaves in July 2014 and 2015. Insert graph in (A) shows mesophyll conductance at midday (gmMD) in July 2014 and 2015. Two way ANOVA followed by Tukey’s pairwise comparison was performed to compare data (means ± SD; n = 8). Letters indicate significant differences (p ≤ 0.05) among sampling hour for each year, whereas asterisks represent significant differences among years for each sampling hour, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001). The statistic adopted for the inclusion in (A) was a Student’s t-test, asterisks represent differences between the two years, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001).
The PCA identified groups of plants with similar water relations, gas exchange and morpho-anatomical features, with the first two components of the PCA explaining 41.58 and 27.74% of the variances. In the biplot (Supplementary Figure S1A) samples collected in different years were segregated in distinct groups.
The samples collected in 2015 were mainly grouped at the negative score of PC1 presenting lower values of Ψw and RWC than samples collected in 2014 and grouped at the positive score of PC2 for gs and leaf thickness (LT). The Pn vector contributed less for distinguishing both years.
Chlorophyll Fluorescence and Chlorophyll Index
Photosystem II maximum efficiency (Fv/Fm) showed a typical midday depression at 12:00 in both years (Figure 5A) (two way ANOVA; p < 0.001) with a negative correlation with air temperature along the day (Supplementary Table S1A). However, while in summer 2014 Fv/Fm recovered optimal values at 15:00, in 2015, this parameter fully recovered only at 18:00. In addition, Fv/Fm was significantly higher in 2014 than in 2015 at 8:00 (two way ANOVA; p = 0.024) and at 15:00 (two way ANOVA; p < 0.001) (Figure 5A). Photosystem II actual efficiency (ΦPSII) differed between the 2 years, with significant lower values in 2014 than in 2015 (two way ANOVA; p < 0.001). The minimum values of ΦPSII were observed from 12:00 to 15:00 in both summers (Figure 5B). The chlorophyll index (Chli) was significantly higher in 2014 than in 2015 at all sampling hours (two way ANOVA; p < 0.001) (Figure 5C). However, the daily trends of this parameter were similar in both years, with a reduction during the central hours of the day followed by a recovery at 18:00 (Figure 5C). The PCA identified groups of plant species with similar chlorophyll fluorescence and chlorophyll index as previously described for gas exchanges and water relations, with samples collected in different years segregated in distinct groups (Supplementary Figure S1A). According to the PCA analysis the samples collected in 2015 were mainly grouped at the negative score of PC1 presenting the lower values of Fv/Fm however, ΦPSII vector contributed less for distinguishing both years.
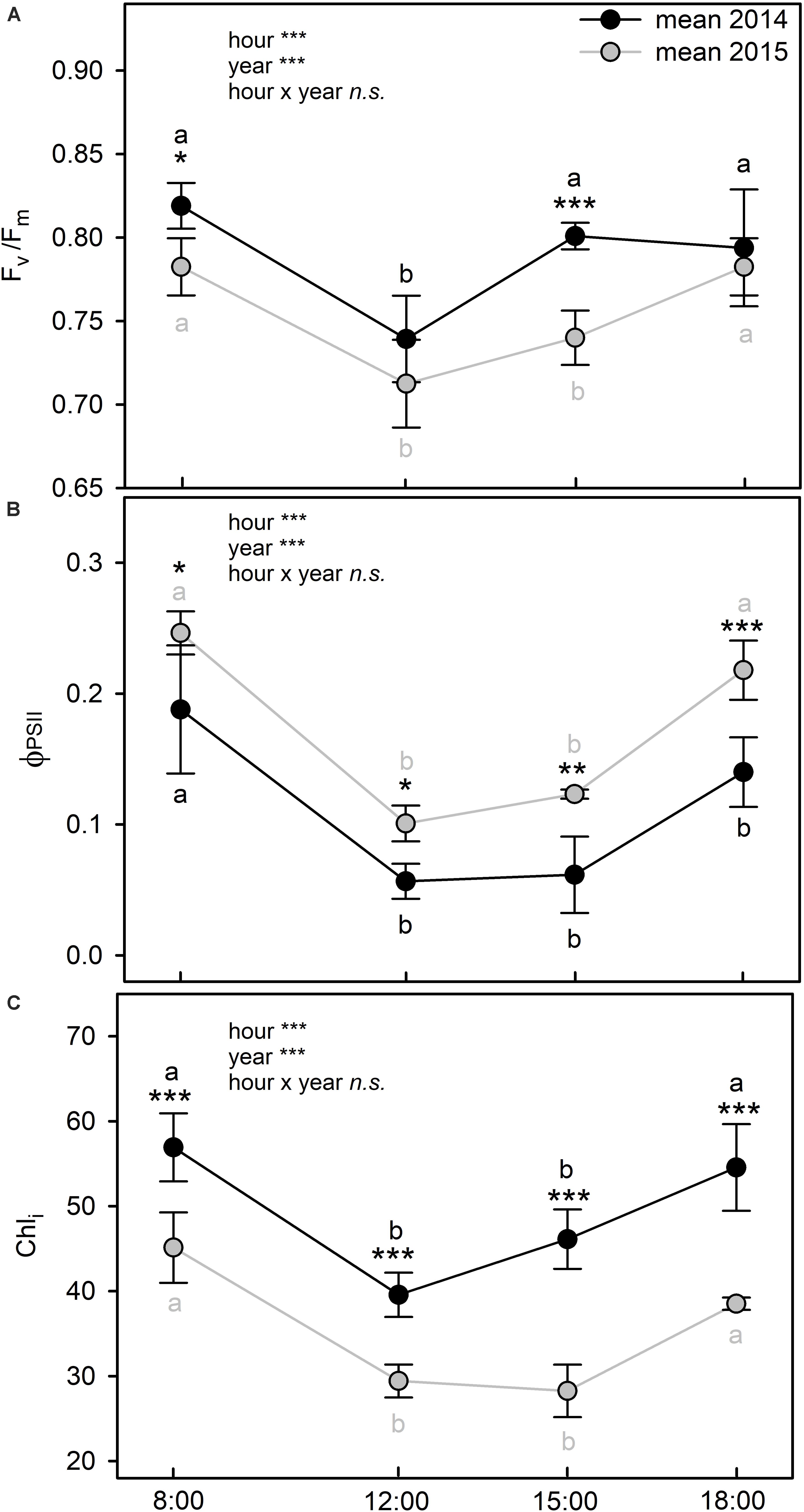
Figure 5. Diurnal trends of maximal photochemical efficiency of PSII (Fv/Fm) (A), actual efficiency of PSII (ΦPSII) (B), and Chlorophyll index (Chli) (C) of C. incanus leaves in July 2014 and 2015. Two way ANOVA followed by Tukey’s pairwise comparison was performed to compare data (means ± SD; n = 8). Letters indicate significant differences (p ≤ 0.05) among sampling hour for each year, whereas asterisks represent significant differences among years for each sampling hour, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001).
Flavonol Index and Polyphenols
The Flavonol index (Flavi) exhibited very similar values and daily trends in the two summers, increasing from 8:00 to 12:00 and then declining from 12:00 to 15:00 until 18:00 (Figure 6A). Moreover, at 12:00, the value of Flavi was significantly higher in summer 2014 than in 2015 (two way ANOVA; p = 0.026) (Figure 6A). Total polyphenols (PolTot) were higher in 2015 than in 2014 (t-test; p = 0.022) (Figure 6B) with a positive relation with air temperature (Supplementary Table S1), because of the significant difference in the total content of tannins, passing from ∼ 51 μmol g–1 dw–1 in 2014 to ∼70 μmol g–1 dw–1 in 2015 (t-test; p = 0.002) (Figure 6C). In contrast, the total content of flavonols did not change from 2014 to 2015 (t-test; p = 0.183) (Figure 6C).
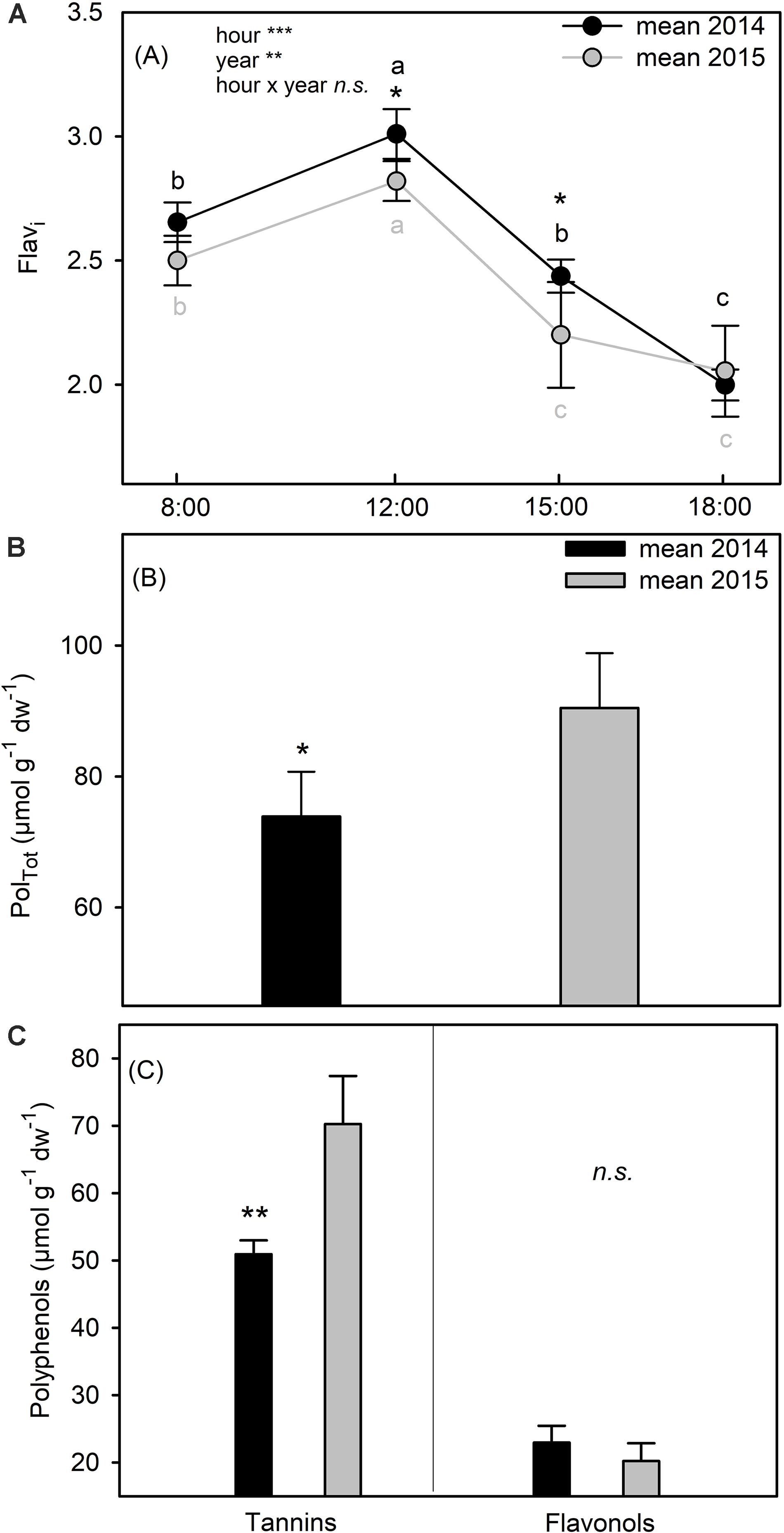
Figure 6. Epidermal Flavonol index (Flavi) (A), total polyphenols content (PolTot) (B), condensed tannins (Tannins) and flavonols leaves content (Flavonols) (C) of C. incanus leaves in July 2014 and 2015. Two way ANOVA followed by Tukey’s pairwise comparison was performed to compare data (means ± SD; n = 8). Letters indicate significant differences (p ≤ 0.05) among sampling hour for each year, whereas asterisks represent significant differences among years for each sampling hour, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001).
Biogenic Volatile Organic Compounds
The total emission of BVOCs increased about fivefold in 2015 compared to 2014 (t-test; p < 0.001) (Figure 7A) presenting a positive relation with air temperature (Supplementary Table S1). All BVOCs detected in the study were monoterpenes. The most important monoterpenes emitted by C. incanus leaves were α- and β-pinene, their sum accounted for about 69% of the total emitted compounds in 2014 and for 60% in 2015 (Figure 7B). Other monoterpenes detected were: α-, β- phellandrene, thujene, and myrcene. In summer 2015, the emission of α-pinene, β-pinene and myrcene were about five times higher compared to summer 2014 (t-test; p < 0.001; p = 0.013; p = 0.011). The emissions of β-phellandrene and thujene increased three and nine times, respectively (t-test; p = 0.005; and p < 0.001), while α-phellandrene emission was about 15 times higher in July 2015 than July 2014 2014 (t-test; p < 0.001) (Figure 7B). The PCA identified groups of plant species with similar BVOCs (and also for PolTot), with samples collected in different years segregated in distinct groups (Supplementary Figure S1B). The samples collected in 2015 were grouped at the positive score of PC1 (84.16% of the total variation), presenting the higher values for PolTot and BVOCs as well as for the leaf thickness (LT).
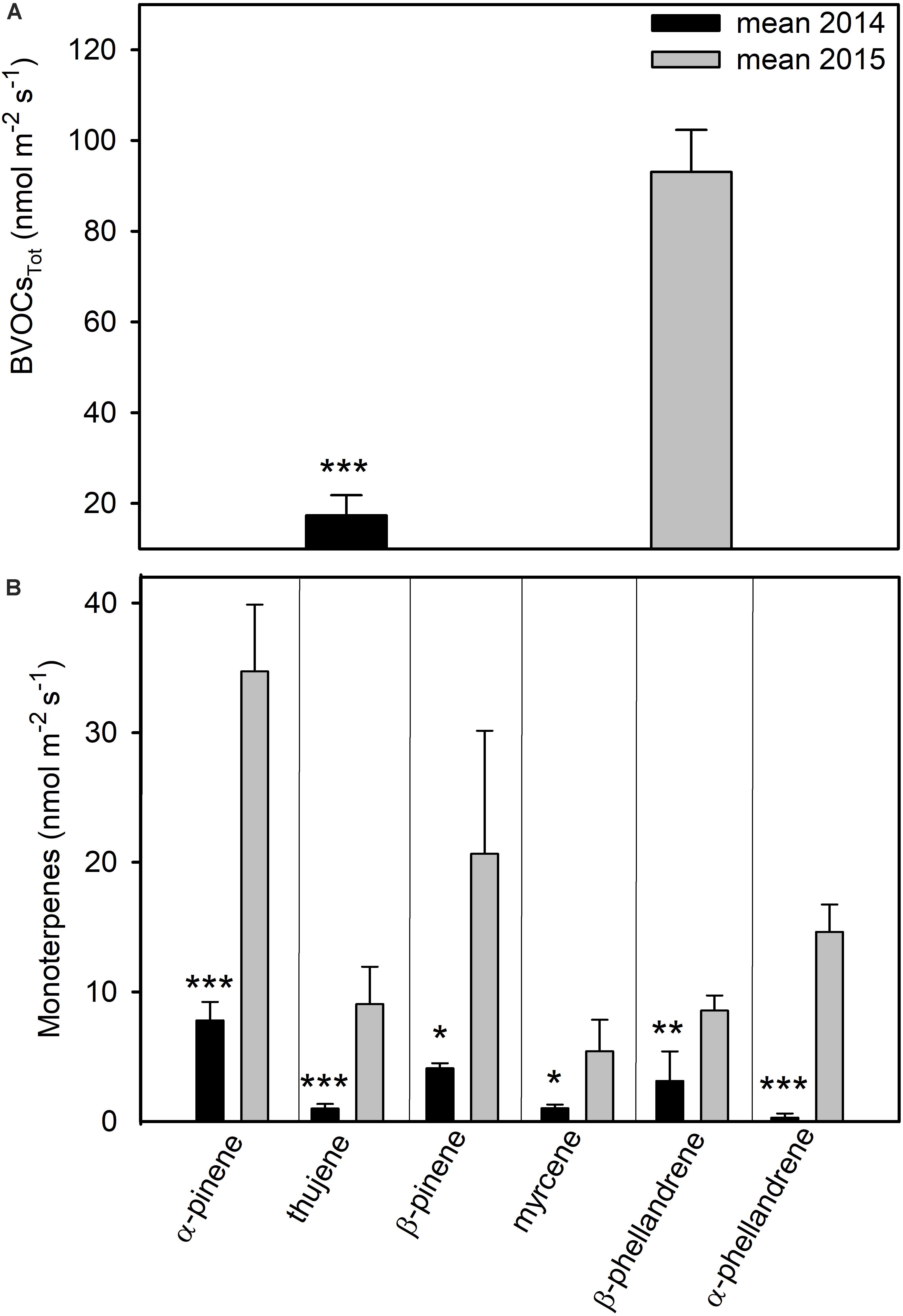
Figure 7. Total biogenic volatile organic compound emission (BVOCTot) (A) and monoterpenes emitted (B) by C. incanus leaves in July 2014 and 2015. Data are means ± SD (n = 8). Asterisks represent differences between the two years, *(p ≤ 0.05), **(p ≤ 0.01), ***(p ≤ 0.001) based on t-test.
Discussion
Leaves of summer 2014 maintained a better water status compared to the drier and hotter summer 2015 (Table 3). In particular, the higher leaf dehydration observed during the heat wave in July 2015, as estimated by the lower RWC, led to a consequent drop in Ψw (Table 3). Leaf morpho-anatomical traits may have played an important role to avoid leaf shrinkage associated to dehydration during drought. Indeed, when water is a limiting factor, plants undergo anatomical alterations, particularly in their leaves, in which the transpiration flow is regulated by stomatal closure (Bosabalidis and Kofidis, 2002; Scoffoni et al., 2014). It has already been reported that the dimorphic C. incanus develops xerophytic leaves during summer (Puglielli and Varone, 2018; Puglielli, 2019). In our study, total lamina thickness was increased by 28% in 2015, as compared to 2014, mainly due to a higher palisade parenchyma (+44%) and to a greater epidermal thickness (+28%) (Table 2 and Supplementary Table S1B). The smaller leaf size (Supplementary Figure S2) and the greater contribution of upper epidermis and of mesophyll tissues, in particular palisade parenchyma, to the whole leaf structure (Figure 2B and Supplementary Table S1B), could have been important to increase resistance against dehydration and cell collapsing induced by the drier conditions of summer 2015 (Bosabalidis and Kofidis, 2002; De Micco and Aronne, 2012; Mansoor et al., 2019). We hypothesize that accumulation of leaf condensed tannins, both in upper epidermis and mesophyll cells (Figure 2A), may have contributed to strengthen the leaf structure of C. incanus in 2015 as previously reported for other species grown in xeric environments (Grossoni et al., 1998; Gravano et al., 2000; Bussotti, 2008). We suggest that the increment in leaf condensed tannins and the higher LT observed in the drier and hotter summer 2015 resulted in a significant increase in LMA (+36%). Indeed, previous studies have clearly shown a positive relationship between LMA and LT (Puglielli et al., 2017c, 2019b; Gratani et al., 2018) and a possible contribution of soluble phenolics to LMA (Poorter et al., 2009). The higher LMA may have provided both a lower evaporative surface area and a lower light harvesting capability per unit of leaf mass (Table 2; Boughalleb and Hajlaoui, 2011; De Micco et al., 2011; Perez-Martin et al., 2014; De la Riva et al., 2016), enabling plants to tolerate lower Ψw without significant losses in leaf turgor (Munné-Bosch et al., 2003). LMA may have also an influence on leaf physiology by affecting the diffusion of CO2 from substomatal cavities through intercellular air space to sites of carboxylation in the mesophyll tissues (Loreto and Centritto, 2008; Flexas et al., 2014; Peguero-Pina et al., 2017). In particular, several studies on non-stressed tree species have shown that a higher LMA constrains leaf mesophyll conductance (gm), thus limiting photosynthetic assimilation rates (Syvertsen et al., 1995; Hassiotou et al., 2009). Nevertheless, recent evidences have shown that, when a higher LMA is accompanied to an increment in thickness of palisade parenchyma, this anatomical adaptation may be accompanied by a higher diffusion and availability of CO2 across the leaf, facilitating the assimilation process (Evans, 1999; Milla-Moreno et al., 2016; Peguero-Pina et al., 2017). Similarly, in our study, the increment in LT and LMA in July 2015 may have led to a higher gm and, in turn, resulted in a higher CO2 uptake at midday compared to July 2014 (Figure 3A and Table 3). This result is in accordance with previous research on drought-acclimated plants, in which a positive relationship between gm and mesophyll thickness was found, supporting the hypothesis that, under water stress, a thicker leaf increases the surface area of chloroplasts exposed to intercellular airspace per unit leaf area (Hanba et al., 1999; Terashima et al., 2001; De Micco et al., 2011; Galmés et al., 2013; Flexas et al., 2014). The higher LMA may have also contributed to lower leaf temperature in summer 2015 (Gratani and Varone, 2004), as similar daily leaf temperatures were observed between the 2 years (Supplementary Figure S3), despite the higher air temperatures caused by the heatwave in July 2015 (Figure 1A; Russo et al., 2015). In addition, the peculiar location of stomata inside epidermal crypts may have allowed the higher gs values in 2015, which contributed to the regulation of leaf temperature and the maintenance of similar Pn between the 2 years (Figure 3B and Supplementary Figure S1A; Aronne and De Micco, 2001). We cannot exclude that other leaf morpho-anatomical adjustments (e.g., leaf rolling, variations in leaf angles and density in leaf pubescence) may have contributed to protect the photosynthetic apparatus under the drier and hotter conditions of July 2015 (Figures 3A, 4A and Supplementary Figure S2; De Micco and Aronne, 2012; Arena et al., 2019). Particularly, paraheliotropism is an adaptative strategy of C. incanus to counteract the deleterious impact of high irradiance to protect leaves under stress condition, especially in summer, thus avoiding photoinhibition (Puglielli et al., 2017b; Pérez-Llorca et al., 2019). However, the Pn time course in 2015 (Figure 3A) showed the typical trend of a severe water-stressed plants, with a higher value in the morning followed by significant drop as the day progressed (Marino et al., 2014). In contrast, in 2014 at 15:00, the peak in Pn coupled with the drop of Ci (Figures 3A, 4B) indicated the lack of significant biochemical limitations to photosynthesis (Lawlor and Cornic, 2002). Conversely, in summer 2015, Ci did not change significantly throughout the day with respect to the morning value, despite the significant drop in Pn. This result might be associated to leaf rolling and suggests that, despite C. incanus leaf photochemistry is tolerant to extreme drought as well as different environmental conditions (Arena et al., 2011; Gori et al., 2019; Puglielli et al., 2019a), RUBP (ribulose 1, 5-bisphosphate) regeneration was partially impaired in July 2015, particularly during the hottest hours of the day, thus determining a Pn decline in 2015 when temperature was higher (Gratani et al., 2018). It is noteworthy that many studies have shown that enzymes involved in RUBP regeneration are impaired by water stress at gs lower than 0.1 mol H2O m–2 s–1 (Sharkey and Seemann, 1989; Sánchez-Rodríguez et al., 1997; Grassi and Magnani, 2005). In our case, even if gs values were below this threshold in both years, we can hypothesize a significant metabolic impairment of Pn only in July 2015, especially during the hottest hours of the day (Figure 4). However, in C. incanus leaves, RUBP regeneration is likely not limited by a decrease in Jf and NADPH (nicotinamide adenine dinucleotide phosphate) synthesis (Flexas et al., 2004). In fact, the observed increase of Jf/Pn at 15:00 in July 2015 (Figure 4C) indicates an excess of reducing power along the electron transport chain and a lower ability to use electrons for carbon assimilation (Sebastiani et al., 2019). In 2015, the excess of reducing power may have resulted in suboptimal Fv/Fm values (Figure 5A; Demmig and Björkman, 1987), which together with higher Ci (Figure 4B) and ΦPSII (Figure 5B), may suggest higher utilization of alternative electron sinks (Grant et al., 2015; Puglielli et al., 2017b; Brunetti et al., 2018). At the same time, the daily down-regulation of ΦPSII and the reduction of leaf chlorophyll content (Figure 5C) observed in both years may have worked as a photoprotection mechanism to dissipate the high amount of light intercepted during the central hours of the day (Figure 1B; Baquedano and Castillo, 2007; Gori et al., 2019). When the capacity of plants to use the radiant energy is severely constrained, the excess of reducing power may be utilized for the biosynthesis of secondary metabolites, such as isoprenoids and polyphenols (Niinemets et al., 2002a,b; Harrison et al., 2013; Tattini et al., 2015). Indeed, secondary metabolites have been suggested to act as alternative electron sinks by consuming trioses phosphate, ATP (adenosine triphosphate) and NADPH, thus playing a role as an energy escape valve (Hernández and Van Breusegem, 2010; Morfopoulos et al., 2013). Among secondary metabolites, polyphenols perform different functions depending on their location in the leaf, acting as both radiation screeners, quenchers of ROS and key molecules involved in the acclimation processes to diurnal changes in UV-B radiation (Agati and Tattini, 2010; Agati et al., 2012; Jansen et al., 2012; Barnes et al., 2016; Tohge and Fernie, 2017; Neugart and Schreiner, 2018). Indeed, the comparable trends of flavonol index (Flavi) observed in both years (Figure 6A) could be related to similar diurnal changes in UV radiations (Stephanou and Manetas, 1997; Brossa et al., 2009). Furthermore, the reduction of chlorophyll content (Figure 5C), concomitantly with the increase in flavonols in the epidermis (Figure 6A) observed at 12:00 in both summers, may indicate that reversible changes in leaf biochemistry have helped C. incanus overcome stressful conditions on a daily basis (Mittler, 2006). Polyphenols of C. incanus leaves mainly belong to the classes of flavonols (quercetin and myricetin derivatives) and condensed tannins (Gori et al., 2016, 2020). Both classes of molecules possess antioxidant function and may have played a role in the inhibition of radical formation under severe stress conditions (Figure 6C; Hernández et al., 2004; Barbehenn et al., 2006; Sebastiani, 2019). In addition, flavonols may have contributed to both UV-B shielding and improve photo-protection at 12:00 (Burchard et al., 2000). As above mentioned, tannins may have helped increase leaf structure under the drier and hotter conditions of summer 2015 (Figure 2 and Supplementary Figure S1B; Ishida et al., 2008; Tharayil et al., 2011; Sumbele et al., 2012; Top et al., 2017). In support of this hypothesis, we found a statistically significant correlation between total polyphenols, mainly constituted by tannins, and air temperature (Table 2 and Supplementary Table S1A). The increase in tannin production observed in July 2015 could be also interpreted as a defense trait of C. incanus to protect the resources acquired during the growing season, thus contributing to retard leaf senescence and abscission and optimizing resource acquisition in nutrient-poor soils conditions (Mediavilla et al., 2001; Wright and Westoby, 2002; Poorter et al., 2009; Massad et al., 2014; Gratani et al., 2016; Puglielli et al., 2017a). Finally, the increment of the fiber-bound proportion of tannins in the leaf cell walls during leaf maturation may have contributed to the lignification process, thus protecting against pathogens and herbivores attack (Zucker, 1983; Bussotti et al., 1998). Another mechanism for counteracting leaf photo-oxidative damage under drought is the biosynthesis of BVOCs (Loreto and Schnitzler, 2010). The emission of monoterpenes is largely widespread among Mediterranean shrub vegetation exposed to concomitant stresses. In fact, these BOVCs have multiple protective functions (Rivoal et al., 2010; Fares et al., 2013; Fineschi et al., 2013; Llusià et al., 2016), ranging from antioxidant (Loreto et al., 2004; Llusià et al., 2006), protection against high temperatures (Owen et al., 2002; Haberstroh et al., 2018), defense against pathogens and herbivores to wound sealing after damage (Pichersky and Gershenzon, 2002). We observed a strong increment in monoterpene emissions in July 2015 compared to July 2014 (Figure 7), likely as a consequence of the higher air temperatures due to the 2015 heat wave (Figure 1A and Supplementary Figure S1B). This is because, being highly volatile, the higher air temperature in 2015 increased the monoterpene partial pressure gradient between leaf and the atmosphere (Niinemets et al., 2010), resulting in their higher emissions than in 2014 (Supplementary Table S1A). However, no qualitative differences in the blend of monoterpenes emerged between the two years. Our results are consistent with those of Owen et al. (2001, 2002) who found α-pinene as the major monoterpene produced by this species. In addition, α-pinene and β-pinene, β-myrcene and α-phellandrene emissions were already detected in others Cistus spp. (Ormeño et al., 2007; Rivoal et al., 2010; Haberstroh et al., 2018). A high production of these compounds has been shown to help the leaves to withstand drought, which supports the assumption that they improve membrane stability, thus conferring thermal protection (Loreto et al., 1998; Copolovici et al., 2005). Hence, the diverse blend of terpenes detected in the emissions of this species may likely contributes to the drought tolerance of C. incanus under field conditions.
Conclusion
In conclusion, the results of our study indicate that C. incanus is a climate resilient species able to tolerate different environmental conditions, including threats caused by extreme weather events, as revealed from the new evidence on its capacity to adjust morpho-physiological and biochemical traits. In particular, the major investment in leaf construction observed in the drier and hotter conditions caused by the summer 2015 heat wave helped avoid leaf shrinkage associated to dehydration. Interestingly, the stimulation of the biosynthesis of secondary metabolites, in particular of monoterpenes and tannins, may have increased leaf thermo-tolerance and may have counteracted photooxidative damage, thus protecting C. incanus photosynthetic apparatus and allowing the maintenance of carbon assimilation despite the severe water and temperature stresses. Therefore, our results suggest that C. incanus might have greater potential to acclimate to climate change in its natural environment.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
AG and CB: conceptualization and validation. GM, CG, BM, and AG: data curation. GM and BM: formal analysis. MC and FF: funding acquisition. AG: investigation. CB: methodology. MC: supervision. FA, CB, and AG: writing – original draft. MC, FF, and MT: writing – review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by funds from the Italian MIUR to CNR, project Economia Circolare (Green & Circular Economy-GECE) (FOE-2019, 436 DBA.AD003.139).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.576296/full#supplementary-material
Footnotes
References
Agati, G., Azzarello, E., Pollastri, S., and Tattini, M. (2012). Flavonoids as antioxidants in plants: location and functional significance. Plant Sci. 196, 67–76. doi: 10.1016/j.plantsci.2012.07.014
Agati, G., and Tattini, M. (2010). Multiple functional roles of flavonoids in photoprotection. New Phytol. 186, 786–793. doi: 10.1111/j.1469-8137.2010.03269.x
Agati, G., Tuccio, L., Kusznierewicz, B., Chmiel, T., Bartoszek, A., Kowalski, A., et al. (2016). Nondestructive optical sensing of flavonols and chlorophyll in white head cabbage (Brassica oleracea L. var. capitata subvar. alba) grown under different nitrogen regimens. J. Agric. Food Chem. 64, 85–94. doi: 10.1021/acs.jafc.5b04962
Arena, C., De Micco, V., De Maio, A., Mistretta, C., Aronne, G., and Vitale, L. (2013). Winter and summer leaves of Cistus incanus: differences in leaf morphofunctional traits, photosynthetic energy partitioning, and poly (ADP-ribose) polymerase (PARP) activity. Botany 91l, 805–813.
Arena, C., Mistretta, C., Di Natale, E., Mennella, M. R. F., De Santo, A. V., and De Maio, A. (2011). Characterization and role of poly (ADP-ribosyl) ation in the Mediterranean species Cistus incanus L. under different temperature conditions. Plant Physiol. Biochem. 49, 435–440. doi: 10.1016/j.plaphy.2011.02.004
Arena, C., and Vitale, L. (2018). Chilling-induced reduction of photosynthesis is mitigated by exposure to elevated CO2 concentrations. Photosynthetica 56, 1259–1267. doi: 10.1007/s11099-018-0843-3
Arena, C., Vitale, L., Bianchi, A. R., Mistretta, C., Vitale, E., Parisi, C., et al. (2019). The ageing process affects the antioxidant defences and the poly (ADPribosyl) ation activity in Cistus incanus L. leaves. Antioxidants. 8:528. doi: 10.3390/antiox8110528
Aronne, G., and De Micco, V. (2001). Seasonal dimorphism in the Mediterranean Cistus incanus L. subsp. incanus. Ann. Bot. 87, 789–794. doi: 10.1006/anbo.2001.1407
Baquedano, F. J., and Castillo, F. J. (2007). Drought tolerance in the Mediterranean species Quercus coccifera, Quercus ilex, Pinus halepensis, and Juniperus phoenicea. Photosynthetica 45, 229–238. doi: 10.1007/s11099-007-0037-x
Barbehenn, R. V., Jones, C. P., Hagerman, A. E., Karonen, M., and Salminen, J. P. (2006). Ellagitannins have greater oxidative activities than condensed tannins and galloyl glucoses at high pH: potential impact on caterpillars. J. Chem. Ecol. 32, 2253–2267. doi: 10.1007/s10886-006-9143-7
Barnes, P. W., Tobler, M. A., Keefover-Ring, K., Flint, S. D., Barkley, A. E., Ryel, R. J., et al. (2016). Rapid modulation of ultraviolet shielding in plants is influenced by solar ultraviolet radiation and linked to alterations in flavonoids. Plant Cell Environ. 39, 222–230. doi: 10.1111/pce.12609
Bosabalidis, A. M., and Kofidis, G. (2002). Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Sci. 163, 375–379. doi: 10.1016/S0168-9452(02)00135-8
Boughalleb, F., and Hajlaoui, H. (2011). Physiological and anatomical changes induced by drought in two olive cultivars (cv Zalmati and Chemlali). Acta Physiol Plant 33, 53–65. doi: 10.1007/s11738-010-0516-8
Brilli, F., Tsonev, T., Mahmood, T., Velikova, V., Loreto, F., and Centritto, M. (2013). Ultradian variation of isoprene emission, photosynthesis, mesophyll conductance, and optimum temperature sensitivity for isoprene emission in water-stressed Eucalyptus citriodora saplings. J. Exp. Bot. 64, 519–528. doi: 10.1093/jxb/ers353
Brossa, R., Casals, I., Pintó-Marijuan, M., and Fleck, I. (2009). Leaf flavonoid content in Quercus ilex L. resprouts and its seasonal variation. Trees 23, 401–408. doi: 10.1007/s00468-008-0289-5
Brunetti, C., Guidi, L., Sebastiani, F., and Tattini, M. (2015). Isoprenoids and phenylpropanoids are key components of the antioxidant defense system of plants facing severe excess light stress. Environ. Exp. Bot. 119, 54–62. doi: 10.1016/j.envexpbot.2015.04.007
Brunetti, C., Loreto, F., Ferrini, F., Gori, A., Guidi, L., Remorini, D., et al. (2018). Metabolic plasticity in the hygrophyte Moringa oleifera exposed to water stress. Tree Physiol. 38, 1640–1654. doi: 10.1093/treephys/tpy089
Burchard, P., Bilger, W., and Weissenböck, G. (2000). Contribution of hydroxycinnamates and flavonoids to epidermal shielding of UV-A and UV-B radiation in developing rye primary leaves as assessed by ultraviolet-induced chlorophyll fluorescence measurements. Plant Cell Environ. 23, 1373–1380. doi: 10.1046/j.1365-3040.2000.00633.x
Bussotti, F. (2008). Functional leaf traits, plant communities and acclimation processes in relation to oxidative stress in trees: a critical overview. Glob Chang Biol. 14, 2727–2739. doi: 10.1111/j.1365-2486.2008.01677.x
Bussotti, F., Gravano, E., Grossoni, P., and Tani, C. (1998). Occurrence of tannins in leaves of beech trees (Fagus sylvatica) along an ecological gradient, detected by histochemical and ultrastructural analyses. New Phytol. 138, 469–479. doi: 10.1046/j.1469-8137.1998.00121.x
Catoni, R., Gratani, L., and Varone, L. (2012). Physiological, morphological and anatomical trait variations between winter and summer leaves of Cistus species. Flora 207, 442–449. doi: 10.1016/j.flora.2012.02.007
Copolovici, L. O., Filella, I., Llusià, J., Niinemets, Ü., and Peñuelas, J. (2005). The capacity for thermal protection of photosynthetic electron transport varies for different monoterpenes in Quercus ilex. Plant Physiol. 139, 485–496. doi: 10.1104/pp.105.065995
Correia, O., and Ascensao, L. (2016). “Summer semi-deciduous species of the Mediterranean landscape: a winning strategy of cistus species to face the predicted changes of the Mediterranean climate,” in Plant Biodiversity. Monitoring, Assessment and Conservation, eds A. A. Ansari, S. S. Gill, Z. K. Abbas, and M. Naeem (Wallingford: CAB International), 195–217.
David, T. S., Henriques, M. O., Kurz-Besson, C., Nunes, J., Valente, F., Vaz, M., et al. (2007). Water-use strategies in two co-occurring Mediterranean evergreen oaks: surviving the summer drought. Tree Physiol. 27, 793–803. doi: 10.1093/treephys/27.6.793
De Dato, G. D., Micali, M., Jaoudé, R. A., Liberati, D., and De Angelis, P. (year>2013). Earlier summer drought affects leaf functioning of the Mediterranean species Cistus monspeliensis L. Environ. Exp. Bot. 93, 13–19. doi: 10.1016/j.envexpbot.2013.03.007
De la Riva, E. G., Olmo, M., Poorter, H., Ubera, J. L., and Villar, R. (2016). Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 Mediterranean woody species along a water availability gradient. PLoS One 11:e0148788. doi: 10.1371/journal.pone.0148788
De Micco, V., Arena, C., Vitale, L., Aronne, G., and Virzo De Santo, A. (2011). Anatomy and photochemical behaviour of Mediterranean Cistus incanus winter leaves under natural outdoor and warmer indoor conditions. Botany 89, 677–688. doi: 10.1139/b11-059
De Micco, V., and Aronne, G. (2009). Seasonal dimorphism in wood anatomy of the Mediterranean Cistus incanus L. subsp. incanus. Trees 23, 981–989. doi: 10.1007/s00468-009-0340-1
De Micco, V., and Aronne, G. (2012). “Morpho-Anatomical traits for plant adaptation to drought,” in Plant Responses to Drought Stress: From Morphological to Molecular Features, eds A. Ricardo (New York, NY: Springer), 37–61.
Demmig, B., and Björkman, O. (1987). Comparison of the effect of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 171, 171–184. doi: 10.1007/BF00391092
Di Ferdinando, M., Brunetti, C., Agati, G., and Tattini, M. (2014). Multiple functions of polyphenols in plants inhabiting unfavorable Mediterranean areas. Environ. Exp. Bot. 103, 107–116. doi: 10.1016/j.envexpbot.2013.09.012
Evans, J. R. (1999). Leaf anatomy enables more equal access to light and CO2 between chloroplasts. New Phytol. 143, 93–104. doi: 10.1046/j.1469-8137.1999.00440.x
Fares, S., Schnitzhofer, R., Jiang, X., Guenther, A., Hansel, A., and Loreto, F. (2013). Observations of diurnal to weekly variations of monoterpene-dominated fluxes of volatile organic compounds from mediterranean forests: implications for regional modeling. Environ. Sci. Technol. 47, 11073–11082. doi: 10.1021/es4022156
Fineschi, S., Loreto, F., Staudt, M., and Peñuelas, J. (2013). “Diversification of volatile isoprenoid emissions from trees: evolutionary and ecological perspectives” in Biology, Controls and Models of Tree Volatile Organic Compound Emissions, eds Ü. Niinemets and R. K. Monson (Dordrecht, NL: Springer) 1–20.
Flexas, J., Bota, J., Loreto, F., Cornic, G., and Sharkey, T. D. (2004). Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 6, 269–279. doi: 10.1055/s-2004-820867
Flexas, J., Diaz-Espejo, A., Gago, J., Gallé, A., Galmés, J., Gulías, J., et al. (2014). Photosynthetic limitations in Mediterranean plants: a review. Environ. Exp. Bot. 103, 12–23. doi: 10.1016/j.envexpbot.2013.09.002
Galle, A., Florez-Sarasa, I., Aououad, H. E., and Flexas, J. (2011). The Mediterranean evergreen Quercus ilex and the semi-deciduous Cistus albidus differ in their leaf gas exchange regulation and acclimation to repeated drought and re-watering cycles. J. Exp. Bot. 62, 5207–5216. doi: 10.1093/jxb/err233
Galmés, J., Abadía, A., Cifre, J., Medrano, H., and Flexas, J. (2007). Photoprotection processes under water stress and recovery in Mediterranean plants with different growth forms and leaf habits. Physiol. Plant. 130, 495–510. doi: 10.1111/j.1399-3054.2007.00919.x
Galmés, J., Ochogavía, J. M., Gago, J., Roldán, E. J., Cifre, J., and Conesa, M. À. (2013). Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: anatomical adaptations in relation to gas exchange parameters. Plant Cell Environ. 36, 920–935. doi: 10.1111/pce.12022
Genty, B., Briantais, J. M., and Baker, N. R. (1989). The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 990, 87–92. doi: 10.1016/S0304-4165(89)80016-9
Gori, A., Ferrini, F., Marzano, M. C., Tattini, M., Centritto, M., Baratto, M. C., et al. (2016). Characterisation and antioxidant activity of crude extract and polyphenolic rich fractions from C. incanus leaves. Int. J. Mol. Sci. 17:1344. doi: 10.3390/ijms17081344
Gori, A., Nascimento, L. B., Ferrini, F., Centritto, M., and Brunetti, C. (2020). Seasonal and diurnal variation in leaf phenolics of three medicinal mediterranean wild species: what is the best harvesting moment to obtain the richest and the most antioxidant extracts? Molecules 25:956. doi: 10.3390/molecules25040956
Gori, A., Tattini, M., Centritto, M., Ferrini, F., Marino, G., Mori, J., et al. (2019). Seasonal and daily variations in primary and secondary metabolism of three maquis shrubs unveil different adaptive responses to Mediterranean climate. Conserv. Physiol. 7:coz070. doi: 10.1093/conphys/coz070
Grant, O. M., Tronina, Ł., García-Plazaola, J. I., Esteban, R., Pereira, J. S., and Chaves, M. M. (2015). Resilience of a semi-deciduous shrub, Cistus salvifolius, to severe summer drought and heat stress. Funct. Plant Biol. 42, 219–228. doi: 10.1071/FP14081
Grassi, G., and Magnani, F. (2005). Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 28, 834–849. doi: 10.1111/j.1365-3040.2005.01333.x
Gratani, L., Catoni, R., and Varone, L. (2013). Morphological, anatomical and physiological leaf traits of Q. ilex, P. latifolia, P. lentiscus, and M. communis and their response to Mediterranean climate stress factors. Bot Stud. 54, 35–46. doi: 10.1186/1999-3110-54-35
Gratani, L., Catoni, R., and Varone, L. (2016). Evergreen species response to Mediterranean climate stress factors. iForest 9, 946–953. doi: 10.3832/ifor1848-009
Gratani, L., and Varone, L. (2004). Adaptive photosynthetic strategies of the Mediterranean maquis species according to their origin. Photosynthetica 42, 551–558. doi: 10.1007/S11099-005-0012-3
Gratani, L., Varone, L., and Catoni, R. (2008). Relationship between net photosynthesis and leaf respiration in Mediterranean evergreen species. Photosynthetica 46, 567–573. doi: 10.1007/s11099-008-0095-8
Gratani, L., Varone, L., Crescente, M. F., Catoni, R., Ricotta, C., and Puglielli, G. (2018). Leaf thickness and density drive the responsiveness of photosynthesis to air temperature in Mediterranean species according to their leaf habitus. J. Arid Environ. 150, 9–14. doi: 10.1016/j.jaridenv.2017.12.007
Gravano, E., Desotgiu, R., Tani, C., Bussotti, F., and Grossoni, P. (2000). Structural adaptations in leaves of two Mediterranean evergreen shrubs under. JME 1, 165–170.
Grossoni, P., Bussotti, F., Tani, C., Gravano, E., Antarelli, S., Bottacci, A., et al. (1998). Morpho-anatomical aterations in leaves of Gagus vatica L. and Quercus ilex L. in different environmental stress condition. Chemosphere 36, 919–924. doi: 10.1016/S0045-6535(97)10148-5
Haberstroh, S., Kreuzwieser, J., Lobo-do-Vale, R., Caldeira, M. C., Dubbert, M., and Werner, C. (2018). Terpenoid emissions of two Mediterranean woody species in response to drought stress. Front. Plant Sci. 9, 1071–1087. doi: 10.3389/fpls.2018.01071
Hanba, Y. T., Miyazawa, S. I., and Terashima, I. (1999). The influence of leaf thickness on the CO2 transfer conductance and leaf stable carbon isotope ratio for some evergreen tree species in Japanese warm-temperate forests. Funct. Ecol. 13, 632–639. doi: 10.1046/j.1365-2435.1999.00364.x
Harley, P. C., Loreto, F., Di Marco, G., and Sharkey, T. D. (1992). Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2. Plant Physiol. 98, 1429–1436. doi: 10.1104/pp.98.4.1429
Harrison, S. P., Morfopoulos, C., Dani, K. S., Prentice, I. C., Arneth, A., Atwell, B. J., et al. (2013). Volatile isoprenoid emissions from plastid to planet. New Phytol. 197, 49–57. doi: 10.1111/nph.12021
Hassiotou, F., Ludwig, M., Renton, M., Veneklaas, E. J., and Evans, J. R. (2009). Influence of leaf dry mass per area, CO2, and irradiance on mesophyll conductance in sclerophylls. J. Exp Bot. 60, 2303–2314. doi: 10.1093/jxb/erp021
Hernández, I., Alegre, L., and Munné-Bosch, S. (2004). Drought-induced changes in flavonoids and other low molecular weight antioxidants in Cistus clusii grown under Mediterranean field conditions. Tree Physiol. 24, 1303–1311. doi: 10.1093/treephys/24.11.1303
Hernández, I., and Van Breusegem, F. (2010). Opinion on the possible role of flavonoids as energy escape valves: novel tools for nature’s Swiss army knife? Plant Sci. 179, 297–301. doi: 10.1016/j.plantsci.2010.06.001
Ishida, A., Nakano, T., Yazaki, K., Matsuki, S., Koike, N., Lauenstein, D. L., et al. (2008). Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms. Oecologia 156, 193–202. doi: 10.1007/s00442-008-0965-6
Jansen, M. A. K., Hideg, E., and Lidon, F. J. C. (2012). UV-B radiation: when does the stressor cause stress? Emir. J. Food Agric. 24, 1–3. doi: 10.9755/ejfa.v24i6.14663
Kok, B. (1948). A critical consideration of the quantum yield of Chlorella photosynthesis. Enzymologia 13, 1–56.
Lawlor, D. W., and Cornic, G. (2002). Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 25, 275–294. doi: 10.1046/j.0016-8025.2001.00814.x
Lefi, E., Medrano, H., and Cifre, J. (2004). Water uptake dynamics, photosynthesis and water use efficiency in field-grown Medicago arborea and Medicago citrina under prolonged Mediterranean drought conditions. Ann. Appl. Biol. 144, 299–307. doi: 10.1111/j.1744-7348.2004.tb00345.x
Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Alpert, P., Artale, V., Li, L., et al. (2006). “The Mediterranean climate: an overview of the main characteristics and issues,” in Mediterranean Climate Variability, eds P. Lionello, P. Malanotte-Rizzoli, and R. Boscolo (Amsterdam, NL: Elviser), 1–26. doi: 10.1016/S1571-9197(06)80003-0
Llusià, J., Peñuelas, J., Alessio, G. A., and Estiarte, M. (2006). Seasonal contrasting changes of foliar concentrations of terpenes and other volatile organic compound in four dominant species of a Mediterranean shrubland submitted to a field experimental drought and warming. Physiol Plant. 127, 632–649. doi: 10.1111/j.1399-3054.2006.00693.x
Llusià, J., Roahtyn, S., Yakir, D., Rotenberg, E., Seco, R., Guenther, A., et al. (2016). Photosynthesis, stomatal conductance and terpene emission response to water availability in dry and mesic Mediterranean forests. Trees 30, 749–759. doi: 10.1007/s00468-015-1317-x
Loreto, F., Förster, A., Dürr, M., Csiky, O., and Seufert, G. (1998). On the monoterpene emission under heat stress and on the increased thermotolerance of leaves of Quercus ilex L. fumigated with selected monoterpenes. Plant Cell Environ. 21, 101–107. doi: 10.1046/j.1365-3040.1998.00268.x
Loreto, F., Pinelli, P., Manes, F., and Kollist, H. (2004). Impact of ozone on monoterpene emissions and evidence for an isoprene-like antioxidant action of monoterpenes emitted by Quercus ilex leaves. Tree Physiol. 24, 361–367. doi: 10.1093/treephys/24.4.361
Loreto, F., Pollastri, S., Fineschi, S., and Velikova, V. (2014). Volatile isoprenoids and their importance for protection against environmental constraints in the Mediterranean area. Environ. Exp. Bot. 103, 99–106. doi: 10.1016/j.envexpbot.2013.09.005
Loreto, F., and Schnitzler, J. P. (2010). Abiotic stresses and induced BVOCs. Trends Plant Sci. 15, 154–166. doi: 10.1016/j.tplants.2009.12.006
Loreto, F., and Centritto, M. (2008). Leaf carbon assimilation in a water-limited world. Plant Biosyst. 142, 154–161. doi: 10.1080/11263500701872937
Mansoor, U., Fatima, S., Hameed, M., Naseer, M., Ahmad, M. S. A., Ashraf, M., et al. (2019). Structural modifications for drought tolerance in stem and leaves of Cenchrus ciliaris L. ecotypes from the Cholistan Desert. Flora 261:151485. doi: 10.1016/j.flora.2019.151485
Marino, G., Pallozzi, E., Cocozza, C., Tognetti, R., Giovannelli, A., Cantini, C., et al. (2014). Assessing gas exchange, sap flow and water relations using tree canopy spectral reflectance indices in irrigated and rainfed Olea europaea L. Environ. Exp. Bot. 99, 43–52. doi: 10.1016/j.envexpbot.2013.10.008
Massad, T. J., Trumbore, S. E., Ganbat, G., Reichelt, M., Unsicker, S., Boeckler, A., et al. (2014). An optimal defense strategy for phenolic glycoside production in Populus trichocarpa–isotope labeling demonstrates secondary metabolite production in growing leaves. New Phytol. 203, 607–619. doi: 10.1111/nph.12811
Mediavilla, S., Escudero, A., and Heilmeier, H. (2001). Internal leaf anatomy and photosynthetic resource-use efficiency: interspecific and intraspecific comparisons. Tree Physiol. 21, 251–259. doi: 10.1093/treephys/21.4.251
Medrano, H., Flexas, J., and Galmés, J. (2009). Variability in water use efficiency at the leaf level among Mediterranean plants with different growth forms. Plant Soil 317, 17–29. doi: 10.1007/s11104-008-9785-z
Milla-Moreno, E. A., McKown, A. D., Guy, R. D., and Soolanayakanahally, R. Y. (2016). Leaf mass per area predicts palisade structural properties linked to mesophyll conductance in balsam poplar (Populus balsamifera L.). Botany 94, 225–239. doi: 10.1139/cjb-2015-0219
Mittler, R. (2006). Abiotic stress, the field environment and stress combination. Trends Plant Sci. 11, 15–19. doi: 10.1016/j.tplants.2005.11.002
Morfopoulos, C., Prentice, I. C., Keenan, T. F., Friedlingstein, P., Medlyn, B. E., Peñuelas, J., et al. (2013). Unifying conceptual model for the environmental responses of isoprene emissions from plants. Ann. Bot. 112, 1223–1238. doi: 10.1093/aob/mct206
Mu, Z., Llusià, J., Liu, D., Ogaya, R., Asensio, D., Zhang, C., et al. (2018). Seasonal and diurnal variations of plant isoprenoid emissions from two dominant species in Mediterranean shrubland and forest submitted to experimental drought. Atmos Environ. 191, 105–115. doi: 10.1016/j.atmosenv.2018.08.010
Munné-Bosch, S., Jubany-Marí, T., and Alegre, L. (2003). Enhanced photo-and antioxidative protection, and hydrogen peroxide accumulation in drought-stressed Cistus clusii and Cistus albidus plants. Tree Physiol. 23, 1–12. doi: 10.1093/treephys/23.1.1
Neugart, S., and Schreiner, M. (2018). UVB and UVA as eustressors in horticultural and agricultural crops. Sci. Hortic. 234, 370–381. doi: 10.1016/j.scienta.2018.02.021
Niinemets, Ü., Arneth, A., Kuhn, U., Monson, R. K., Peñuelas, J., and Staudt, M. (2010). The emission factor of volatile isoprenoids: stress, acclimation, and developmental responses. Biogeosciences 7, 2203–2223. doi: 10.5194/bg-7-2203-2010
Niinemets, Ü., Hauff, K., Bertin, N., Tenhunen, J. D., Steinbrecher, R., and Seufert, G. (2002a). Monoterpene emissions in relation to foliar photosynthetic and structural variables in Mediterranean evergreen Quercus species. New Phytol. 153, 243–256. doi: 10.1046/j.0028-646X.2001.00323.x
Niinemets, Ü., Kännaste, A., and Copolovici, L. (2013). Quantitative patterns between plant volatile emissions induced by biotic stresses and the degree of damage. Front. Plant Sci. 4:276. doi: 10.3389/fpls.2013.00262
Niinemets, Ü., Seufert, G., Steinbrecher, R., and Tenhunen, J. D. (2002b). A model coupling foliar monoterpene emissions to leaf photosynthetic characteristics in Mediterranean evergreen Quercus species. New Phytol. 153, 257–275. doi: 10.1046/j.0028-646X.2001.00324.x
Nogués, I., Medori, M., and Calfapietra, C. (2015). Limitations of monoterpene emissions and their antioxidant role in Cistus sp. under mild and severe treatments of drought and warming. Environ. Exp. Bot. 119, 76–86. doi: 10.1016/j.envexpbot.2015.06.001
Ormeño, E., Mevy, J. P., Vila, B., Bousquet-Mélou, A., Greff, S., Bonin, G., et al. (2007). Water deficit stress induces different monoterpene and sesquiterpene emission changes in Mediterranean species. Relationship between terpene emissions and plant water potential. Chemosphere 67, 276–284. doi: 10.1016/j.chemosphere.2006.10.029
Owen, S. M., Boissard, C., and Hewitt, C. N. (2001). Volatile organic compounds (VOCs) emitted from 40 Mediterranean plant species: VOC speciation and extrapolation to habitat scale. Atmos. Environ. 35, 5393–5409. doi: 10.1016/S1352-2310(01)00302-8
Owen, S. M., Harley, P., Guenther, A., and Hewitt, C. N. (2002). Light dependency of VOC emissions from selected Mediterranean plant species. Atmos Environ. 36, 3147–3159. doi: 10.1016/S1352-2310(02)00235-2
Papaefthimiou, D., Papanikolaou, A., Falara, V., Givanoudi, S., Kostas, S., and Kanellis, A. K. (2014). Genus Cistus: a model for exploring labdane-type diterpenes’ biosynthesis and a natural source of high value products with biological, aromatic, and pharmacological properties. Front. Chem. 2:35. doi: 10.3389/fchem.2014.00035
Parra, A., and Moreno, J. M. (2018). Drought differentially affects the post-fire dynamics of seeders and resprouters in a Mediterranean shrubland. Sci. Total Environ. 626, 1219–1229. doi: 10.1016/j.scitotenv.2018.01.174
Peguero-Pina, J. J., Sisó, S., Flexas, J., Galmés, J., García-Nogales, A., Niinemets, Ü., et al. (2017). Cell-level anatomical characteristics explain high mesophyll conductance and photosynthetic capacity in sclerophyllous Mediterranean oaks. New Phytol. 214, 585–596. doi: 10.1111/nph.14406
Pérez-Llorca, M., Casadesús, A., Müller, M., and Munné-Bosch, S. (2019). Leaf orientation as part of the leaf developmental program in the semi-deciduous shrub, Cistus albidus L.: diurnal, positional and photoprotective effects during winter. Front. Plant Sci. 10:767. doi: 10.3389/fpls.2019.00767
Perez-Martin, A., Michelazzo, C., Torres-Ruiz, J. M., Flexas, J., Fernández, J. E., Sebastiani, L., et al. (2014). Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: correlation with gene expression of carbonic anhydrase and aquaporins. J. Exp. Bot. 65, 3143–3156. doi: 10.1093/jxb/eru160
Pichersky, E., and Gershenzon, J. (2002). The formation and function of plant volatiles: perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 5, 237–243. doi: 10.1016/S1369-5266(02)00251-0
Poorter, H., Niinemets, Ü., Poorter, L., Wright, I. J., and Villar, R. (2009). Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol. 182, 565–588. doi: 10.1111/j.1469-8137.2009.02830.x
Puglielli, G. (2019). Beyond the concept of winter-summer leaves of mediterranean seasonal dimorphic species. Front. Plant Sci. 10:696. doi: 10.3389/fpls.2019.00696
Puglielli, G., Catoni, R., Spoletini, A., Varone, L., and Gratani, L. (2017a). Short-term physiological plasticity: trade-off between drought and recovery responses in three Mediterranean Cistus species. Ecol. Evol. 7, 10880–10889. doi: 10.1002/ece3.3484
Puglielli, G., Gratani, L., and Varone, L. (2019a). Leaf rolling as indicator of water stress in Cistus incanus from different provenances. Photosynthetica 57, 202–208. doi: 10.32615/ps.2019.014
Puglielli, G., Redondo-Gómez, S., Gratani, L., and Mateos-Naranjo, E. (2017b). Highlighting the differential role of leaf paraheliotropism in two Mediterranean Cistus species under drought stress and well-watered conditions. J. Plant Physiol. 213, 199–208. doi: 10.1016/j.jplph.2017.02.015
Puglielli, G., and Varone, L. (2018). Inherent variation of functional traits in winter and summer leaves of Mediterranean seasonal dimorphic species: evidence of a ‘within leaf cohort’spectrum. AoB Plants 10:ply027. doi: 10.1093/aobpla/ply027
Puglielli, G., Varone, L., and Gratani, L. (2019b). Diachronic adjustments of functional traits scaling relationships to track environmental changes: revisiting Cistus species leaf cohort classification. Flora 254, 173–180. doi: 10.1016/j.flora.2018.08.010
Puglielli, G., Varone, L., Gratani, L., and Catoni, R. (2017c). Specific leaf area variations drive acclimation of Cistus salvifolius in different light environments. Photosynthetica 55, 31–40. doi: 10.1007/s11099-016-0235-5
Quézel, P. (1985). “Definition of the Mediterranean region and the origin of its flora,” in Plant Conservation in the Mediterranean Area, eds C. Gòmez Campo (Dordrecht,NL: Spinger), 9–24.
Rivoal, A., Fernandez, C., Lavoir, A. V., Olivier, R., Lecareux, C., Greff, S., et al. (2010). Environmental control of terpene emissions from Cistus monspeliensis L. in natural Mediterranean shrublands. Chemosphere 78, 942–949. doi: 10.1016/j.chemosphere.2009.12.047
Rotondi, A., Rossi, F., Asunis, C., and Cesaraccio, C. (2003). Leaf xeromorphic adaptations of some plants of a coastal Mediterranean macchia ecosystem. JME 4, 25–36.
Russo, S., Sillmann, J., and Fischer, E. M. (2015). Top ten European heatwaves since 1950 and their occurrence in the coming decades. Environ. Res. Lett. 10:124003.
Sánchez-Rodríguez, J., Martinez-Carrasco, R., and Pérez, P. (1997). Photosynthetic electron transport and carbon-reduction-cycle enzyme activities under long-term drought stress in Casuarina equisetifolia Forst. & Forst. Photosyn. Res. 52, 255–262. doi: 10.1023/A:1005878307607
Sardans, J., and Peñuelas, J. (2013). Plant-soil interactions in Mediterranean forest and shrublands: impacts of climatic change. Plant Soil 365, 1–33. doi: 10.1007/s11104-013-1591-6
Scoffoni, C., Vuong, C., Diep, S., Cochard, H., and Sack, L. (2014). Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance. Plant Physiol. 164, 1772–1788. doi: 10.1104/pp.113.221424
Sebastiani, F., Torre, S., Gori, A., Brunetti, C., Centritto, M., Ferrini, F., et al. (2019). Dissecting adaptation mechanisms to contrasting solar irradiance in the mediterranean shrub Cistus incanus. Int. J. Mol. Sci. 20:3599. doi: 10.3390/ijms20143599
Sharkey, T. D., and Seemann, J. R. (1989). Mild water stress effects on carbon-reduction-cycle intermediates, ribulose bisphosphate carboxylase activity, and spatial homogeneity of photosynthesis in intact leaves. Plant Physiol. 89, 1060–1065. doi: 10.1104/pp.89.4.1060
Spurr, A. R. (1969). A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 26, 31–43.
Stephanou, M., and Manetas, Y. (1997). The effects of seasons, exposure, enhanced UV-B radiation, and water stress on leaf epicuticular and internal UV-B absorbing capacity of Cistus creticus: a Mediterranean field study. J. Exp. Bot. 48, 1977–1985. doi: 10.1093/jxb/48.11.1977
Sumbele, S., Fotelli, M. N., Nikolopoulos, D., Tooulakou, G., Liakoura, V., Liakopoulos, G., et al. (2012). Photosynthetic capacity is negatively correlated with the concentration of leaf phenolic compounds across a range of different species. AoB Plants. 2012:pls25. doi: 10.1093/aobpla/pls025
Syvertsen, J. P., Lloyd, J., McConchie, C., Kriedemann, P. E., and Farquhar, G. D. (1995). On the relationship between leaf anatomy and CO2 diffusion through the mesophyll of hypostomatous leaves. Plant Cell Environ. 18, 149–157. doi: 10.1111/j.1365-3040.1995.tb00348.x
Tattini, M., Loreto, F., Fini, A., Guidi, L., Brunetti, C., Velikova, V., et al. (2015). Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus× acerifolia plants during Mediterranean summers. New Phytol. 207, 613–626. doi: 10.1111/nph.13380
Terashima, I., Miyazawa, S. I., and Hanba, Y. T. (2001). Why are sun leaves thicker than shade leaves? Consideration based on analyses of CO2 diffusion in the leaf. J. Plant Res. 114, 93–105. doi: 10.1007/PL00013972
Tharayil, N., Suseela, V., Triebwasser, D. J., Preston, C. M., Gerard, P. D., and Dukes, J. S. (2011). Changes in the structural composition and reactivity of Acer rubrum leaf litter tannins exposed to warming and altered precipitation: climatic stress-induced tannins are more reactive. New Phytol. 191, 132–145. doi: 10.1111/j.1469-8137.2011.03667.x
Tohge, T., and Fernie, A. R. (2017). An overview of compounds derived from the shikimate and phenylpropanoid pathways and their medicinal importance. Mini Rev Med Chem. 17, 1013–1027. doi: 10.2174/1389557516666160624123425
Top, S. M., Preston, C. M., Dukes, J. S., and Tharayil, N. (2017). Climate influences the content and chemical composition of foliar tannins in green and senesced tissues of Quercus rubra. Front. Plant Sci. 8:423. doi: 10.3389/fpls.2017.00423
Van Der Plas, F., Manning, P., Allan, E., Scherer-Lorenzen, M., Verheyen, K., Wirth, C., et al. (2016). Jack-of-all-trades effects drive biodiversity–ecosystem multifunctionality relationships in European forests. Nat. Commun. 7, 1–11. doi: 10.1038/ncomms11109
Wright, I. J., and Westoby, M. (2002). Leaves at low versus high rainfall: coordination of structure, lifespan and physiology. New Phytol. 155, 403–416. doi: 10.1046/j.1469-8137.2002.00479.x
Keywords: drought, flavonols, heat wave, leaf morpho-anatomical traits, Mediterranean maquis, monoterpenes, photosynthesis, tannins
Citation: Alderotti F, Brunetti C, Marino G, Centritto M, Ferrini F, Giordano C, Tattini M, Moura BB and Gori A (2020) Coordination of Morpho-Physiological and Metabolic Traits of Cistus incanus L. to Overcome Heatwave-Associated Summer Drought: A Two-Year On-Site Field Study. Front. Ecol. Evol. 8:576296. doi: 10.3389/fevo.2020.576296
Received: 25 June 2020; Accepted: 12 October 2020;
Published: 06 November 2020.
Edited by:
Andrea Ghirardo, Helmholtz Zentrum München, GermanyReviewed by:
Giacomo Puglielli, Estonian University of Life Sciences, EstoniaCarmen Arena, University of Naples Federico II, Italy
Jürgen Kreuzwieser, University of Freiburg, Germany
Copyright © 2020 Alderotti, Brunetti, Marino, Centritto, Ferrini, Giordano, Tattini, Moura and Gori. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cecilia Brunetti, Y2VjaWxpYS5icnVuZXR0aUBpcHNwLmNuci5pdA==