 Michael Weber1*
Michael Weber1* Théo Tacail1,2
Théo Tacail1,2 Federico Lugli3
Federico Lugli3 Marcus Clauss4Katrin Weber1Jennifer Leichliter1Daniela E. Winkler1,5Regina Mertz-Kraus1
Marcus Clauss4Katrin Weber1Jennifer Leichliter1Daniela E. Winkler1,5Regina Mertz-Kraus1 Thomas Tütken1
Thomas Tütken1- 1Institute of Geosciences, Johannes Gutenberg University, Mainz, Germany
- 2Bristol Isotope Group, School of Earth Sciences, University of Bristol, Bristol, United Kingdom
- 3Department of Cultural Heritage, University of Bologna, Ravenna, Italy
- 4Clinic for Zoo Animals, Exotic Pets and Wildlife, Vetsuisse Faculty, University of Zürich, Zurich, Switzerland
- 5Center of Natural History, University of Hamburg, Hamburg, Germany
Strontium isotopes in biogenic apatite, especially enamel, are widely employed to determine provenance and track migration in palaeontology and archaeology. Body tissues record the 87Sr/86Sr of bioavailable Sr of ingested food and water. To identify non-local individuals, knowledge of the 87Sr/86Sr of a non-migratory population is required. However, varying factors such as tissue turnover rates, feeding selectivity, Sr content, digestibility of food, and the ingestion of mineral dust can influence body tissue 87Sr/86Sr. To evaluate the Sr contribution of diet and water to mammalian hard tissues 87Sr/86Sr, controlled feeding studies are necessary. Here we present 87Sr/86Sr from controlled feeding experiments with two rodent species (Rattus norvegicus, Cavia porcellus). Due to the continuous and fast incremental growth of rat and guinea pig incisors (~0.1 – 0.5 mm/day), their enamel is expected to record isotopic dietary changes. For Experiment-1: Diet Switch, animals were switched from their respective supplier food to a pelleted experimental diet containing either insect-, plant-, or meat-meal and a staggered-sampling approach was used to monitor the 87Sr/86Sr changes in rat incisor enamel and bone over the course of the experiment. In Experiment-2: Basic Diets, separated cohorts (n = 6) of rats and guinea pigs were fed one of the three pelleted diets and received tap water for 54 days. While the rat incisors showed a complete tissue turnover, the slower-growing guinea pig incisors partially retained supplier diet-related isotopic compositions. In addition, one group of rats fed plant-meal pellets received Sr-rich mineral water, demonstrating that drinking water can be an important Sr source in addition to diet. Additionally, a leaching experiment showed that only a small fraction of diet-related Sr is bioavailable. Finally, in Experiment-3: Dust Addition, guinea pigs were fed pellets with and without addition of 4% of isotopically distinct dust (loess or kaolin). Animals that received kaolin-containing pellets displayed increased enamel 87Sr/86Sr. Intra-population 87Sr/86Sr variability within each feeding group was small and thus we conclude that it should not affect interpretations of 87Sr/86Sr in provenance studies. However, the differences between bulk food and leachate 87Sr/86Sr highlight the importance of Sr bioavailability for provenance studies and Sr isoscapes.
Introduction
The number of studies dealing with the radiogenic Sr isotope ratio (87Sr/86Sr) have substantially increased within the past decades, and it has become an important tool in many scientific fields, such as petrology, palaeoclimatology, forensics and food traceability, especially in combination with light stable isotopes (e.g., McArthur et al., 2001, 2012; Kelly et al., 2005; Kimura and Chang, 2012; Marchionni et al., 2016; Weber et al., 2018b). The 87Sr/86Sr of bones and tooth enamel is commonly used in archaeological and palaeontological applications, mainly for the reconstruction of mobility and provenance of populations and individuals, as well as for reconstructing the life histories of animals and humans (e.g., Balasse et al., 2002; Outridge et al., 2002; Bentley, 2006; Copeland et al., 2011; Gregoricka, 2014; Lugli et al., 2017a). The mineral composition of bone and tooth enamel is hydroxylapatite [Ca5(PO4)3(OH)], and the Ca2+ in the hydroxylapatite is substituted by Sr2+ in trace amounts due to their similar chemical behavior (Comar et al., 1957), making the determination of Sr isotope ratios in these materials feasible. Observed differences in 87Sr/86Sr in nature are related to differences in the relative abundance of the radiogenic 87Sr vs. the stable 86Sr, due to the decay of the radioactive parent isotope 87Rb in geological materials (Banner, 2004). Therefore, different geographical regions with distinct bedrock geologies will yield different 87Sr/86Sr ratios (Faure and Mensing, 2005). The isotopic composition of the locally bioavailable Sr enters the food chain via Sr-bearing nutrients from ingested water and plants (Carr et al., 1962; Capo et al., 1998; Blum et al., 2000). After digestive assimilation, radiogenic Sr is incorporated into body tissues without any observed isotopic variation. There are apparently no biotic or abiotic factors affecting radiogenic Sr isotopes (Flockhart et al., 2015), while non-radiogenic stable Sr isotopes (88Sr/86Sr expressed as δ88/86Sr) show a trophic level effect caused by biologically induced and mass-dependent fractionation (Knudson et al., 2010; Lewis et al., 2017). For radiogenic Sr (87Sr/86Sr), natural mass-dependent fractionation (Urey, 1947), as well as fractionation during mass spectrometric Sr isotope analysis are corrected for after normalization to a constant 88Sr/86Sr of 8.375209 during data reduction (Meija et al., 2016).
Differences in the formation time of vertebrate hard tissues allow the reconstruction of individual movements during an animal's lifetime from Sr isotope analysis of teeth and bones that form at different ontogenetic stages. While tooth enamel 87Sr/86Sr is not altered after its complete maturation (Hillson, 2005), bones undergo a continuous remodeling throughout life, depending on bone type and ontogenetic age (Jowsey et al., 1971). Therefore, tooth enamel of diphyodont mammals without ever-growing teeth reflects the 87Sr/86Sr during different phases of the animal's early life, while bones reflect an average of dietary 87Sr/86Sr ratios over the last years or the whole life of an animal, depending on the animals age and longevity (Knipper, 2004; Tütken, 2010). The differences in the timing of formation and turnover (or lack thereof in the case of teeth) of these tissues have led to extensive use of bone-tooth pairs to reconstruct the mobility and life history of animals and humans using radiogenic Sr isotopes (Müller et al., 2003; Schweissing and Grupe, 2003; Bentley, 2006; Scheeres et al., 2013). However, relatively little is known about the natural intra-population variability of Sr isotopes, which in turn may potentially influence the interpretation of differences in tooth-bone or tooth-tooth pairs, which are important to identify local vs. non-local values in comparison to Sr isoscapes (Bataille et al., 2020). Some studies evaluated intra- and/or inter-individual Sr isotope variability in archaeological sites (Gregoricka, 2014; Knudson et al., 2016). However, Sr sources and life histories are often poorly resolved in many archaeological populations, so the results of these studies potentially over-estimate true intra-population variability. Therefore, feeding experiments with controlled diet/water Sr inputs and analyses of tissues formed during the experimental period are necessary to properly quantify intra- and/or inter-individual Sr isotope variability. Lewis et al. (2017) performed such a controlled feeding experiment on domestic pigs over two generations to evaluate the influence of marine resources on Sr/Ca, 87Sr/86Sr, and δ88/86Sr. Analyzed materials included diet, tooth enamel, bones and faeces. The authors determined that the 87Sr/86Sr of the animals reflected the diet they received, in that the Sr/Ca ratio increased with increasing marine portion of the food. In addition, they showed a depletion of δ88/86Sr of 0.322 ± 0.060‰ between diet and tooth enamel. Anders et al. (2019) used 87Sr/86Sr to study the locally bioavailable Sr isotope composition and characterize variability in different tissue types of pigs (tooth enamel, bones, bristles, meat, organs and blood). They observed a high intra- and inter-individual variability, as well as tissue-specific patterns. Tissues with higher turnover rates, such as soft tissues and blood, were found to have the highest variability in 87Sr/86Sr. The authors suggested that only tissues with slow turnover, thus allowing for sufficient time averaging of the Sr isotope signal should be used for estimating local bioavailable Sr isotope ranges.
Rodent incisors are ever growing, which makes them an ideal, continuously recording archive in which to study the incorporation of isotopic signals into a hard tissue from diet and water sources. Additionally, enamel is frequently well-preserved in the archaeological and palaeontological record (Copeland et al., 2010; Kirsanow and Tuross, 2011; Gehler et al., 2012). Newly formed tooth enamel and dentine is constantly produced at the basal end of the incisor (Leblond and Warshawsky, 1979), while older enamel is worn away and lost due to regular abrasion. This allows for a continuous monitoring of the Sr isotope composition, using high-resolution sampling techniques, such as micro-milling or in-situ laser ablation. While metabolically active soft tissues (e.g., liver or kidney) which turn over rapidly, reflect only the Sr isotopic compositions of the diet most recently consumed, tooth enamel remains unchanged after its maturation, making rodent incisor enamel a well-suited focal tissue in controlled feeding studies. With a growth rate of 0.5 mm/day for rats (Law et al., 2003) and 0.1 – 0.3 mm/day for guinea pigs (Müller et al., 2015), a typical rat or guinea pig incisor (~2.6 – 3 cm total length) reflects up to the last 60 days of life (Figure 1). By measuring the 87Sr/86Sr along the rodent incisor, a time-resolved Sr isotope incorporation can be monitored. In addition, the analysis of bone material allows for a direct comparison between a fast (incisor enamel) and slow growing (bone) hard tissue with different turnover times.
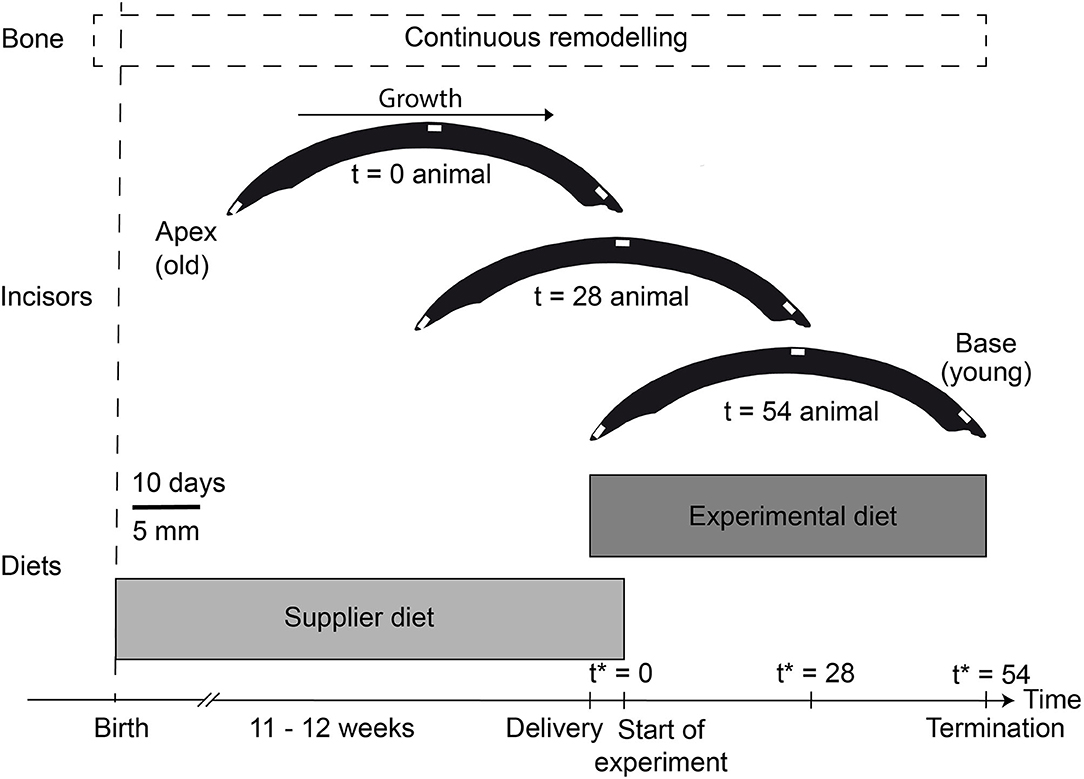
Figure 1. Experimental overview for this study with the approximate growth phases of bone (dashed box) and (rat) incisor enamel (schematic illustration). The base of the incisors consist of the youngest enamel, while the apex is the oldest part of the incisor. We assumed a growth rate of 0.3 – 0.5 mm/day for the rodent incisors (Law et al., 2003; Müller et al., 2015). Feeding duration for each diet type is indicated by the two gray boxes (note the brief overlap between the supplier diet and the experimental diet during the acclimatization phase of the animals for 5 days, where both diets were provided). The white lines within the incisors indicate the sampling locations for LA-MC-ICP-MS Sr isotope analyses (see also Supplementary Figure 1). *Timing (t) is always relative to the start of the experiment (i.e., the consumption of experimental food only).
Here we present radiogenic Sr isotope data from three controlled feeding experiments with two rodent species: the omnivorous, opportunistic feeding rat (Rattus norvegicus forma domestica) and the herbivorous guinea pig (Cavia procellus). In two of the experiments, both rats and guinea pigs received different meat-, insect-, or plant-meal based pelleted diets (25 – 56 wt%, see Supplementary Table 1 for diet formula), while in the third experiment animals received pelleted diets with and without the addition of 4 wt% dust. In a first experiment (Experiment-1: Diet Switch), a staggered-sampling approach was used to determine the timing of dietary 87Sr/86Sr incorporation into the enamel and bone of rats after a diet switch from the supplier diet. For a second experiment (Experiment-2: Basic Diets), rats and guinea pigs were fed with the three different pelleted diets described above for 54 days, resulting in a (near) complete growth of the incisors during the experimental duration (Figure 1). Due to the slower remodeling of bone, 87Sr/86Sr did not reach dietary equilibrium with the diets. Additionally, one group of rats was provided with Sr-rich mineral water. In a third experiment (Experiment-3: Dust Addition), guinea pigs were fed with a plant-based pelleted diet, supplemented with two different silt to clay sized natural and isotopically distinct mineral dusts (loess and kaolin) at a 4 wt% level, to simulate ingestion of soil components and assess its influence on the Sr isotope composition of an animal. Diet, water, teeth (all experiments), bones (Experiments-1: Diet Switch and−2: Basic Diets) as well as faeces samples (Experiment-2: Basic Diets) were analyzed for 87Sr/86Sr to evaluate the timing of the dietary overturn in the ever-growing incisors. A leaching experiment of the pelleted diets was performed to assess their differences in bioavailability of Sr from the different diets.
Materials and Methods
Controlled Feeding Experiments
The controlled feeding experiments were approved by the Cantonal Veterinary Office in Zurich, Switzerland (license no. ZH135/16) and performed at the University of Zurich between July and October 2017. Adult female WISTAR (RjHan:WI) rats (11 – 12 weeks old at the beginning of the experiment, n = 46) and Dunkin Hartley (HsdDhl:DH) guinea pigs (3 – 5 weeks old at the beginning of the experiment, n = 36) were housed in groups of six individuals in separate indoor enclosures with water and food provided for ad libitum consumption. Animals were prevented from gnawing on the enclosure or other materials during the experiment to avoid unnecessary mechanical abrasion of tooth enamel. Prior to the experiments (from birth until arrival in Zurich), the animals received a standard pelleted diet (formula 2040 Teklad Global Guinea Pig Diet, for guinea pigs and formula 2018S Teklad Global 18% Protein Rodent Diet, for rats) at the supplier facility. All animals were provided with the supplier diet together with the experimental diet for 5 days to allow acclimatization to the new diet, before switching entirely to the experimental diet. For guinea pigs (which cannot synthesize vitamin C), drinking water was supplemented with vitamin C at a concentration of 200 mg/L. After the experiments, the animals were euthanized with CO2 and dissected to sample soft- and hard-tissues for isotope analysis. Lower mandibular incisors and distal tibias were chosen for all analyses during this study. Additionally, faeces were collected during Experiment-2: Basic Diets after at least 20 days on the experimental diet.
In total, three different experimental setups were used in the framework of this study: (1) Diet Switch (three groups of rats; (2) Basic Diets (four groups of rats, including water enrichment, and three groups of guinea pigs; (3) Dust Addition (three groups of guinea pigs).
In the Diet Switch experiment, a staggered-sampling approach was performed, in which rats were euthanized after 0 – 40 days on the experimental diet (n = 7 for insect- and plant diets each, and n = 8 for meat based diet), to study Sr incorporation into enamel and bone after the switch from the supplier to the experimental diet (Table 1). Each plant-based diet also included either a plant (lucerne, Medicago sativa, 56 wt%), insect (Protix Insect Protein of soldier fly larvae, Hermetia illucens, 26 wt%), or mammalian meat (lamb, Ovis aries, 25 wt%, potentially with minor contribution of bone) meal ingredient to simulate different feeding categories (i.e., herbivore, insectivore, or carnivore, see Supplementary Table 1 for the exact ingredients for each diet).
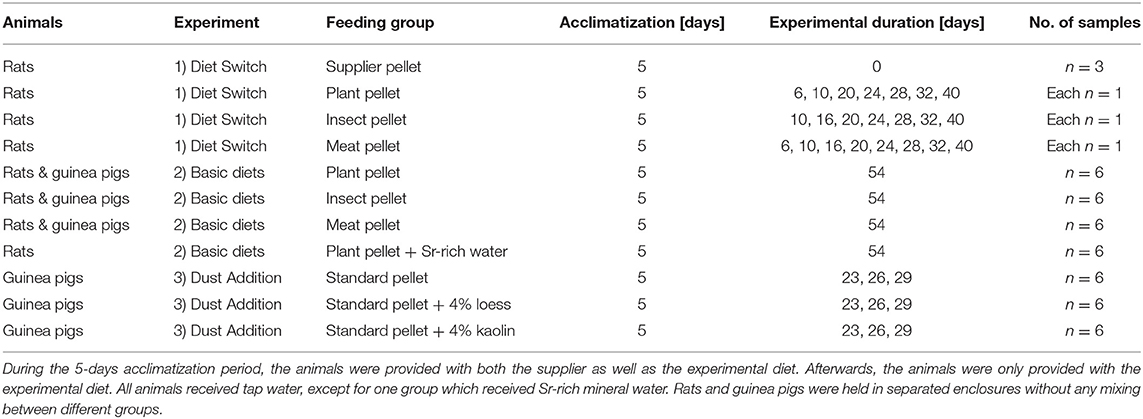
Table 1. Overview of the three controlled feeding experiments.
For the Basic Diets experiment, groups of six rats and six guinea pigs were each fed with the experimental diet for 54 days. Each feeding group received local Zurich tap water, with the exception of one group of rats, which received a commercially available Ca-Sr-rich mineral water (spring in Sersheim, Germany). Individuals from each group from the Basic Diets experiment Diets (excluding the rats provided the Sr-rich mineral water) were also kept in isolation in metabolic cages for 4 days after at least 20 days on the experimental diet to measure differences in food intake for each diet, and for individual fecal collection (≥20 g/ individual). For each diet group, one fecal sample (25 – 50 mg) was analyzed.
In the third Dust Addition experiment, three groups of guinea pigs were fed, in groups of six, for 23 – 29 days with a standard plant pelleted diet (71 wt% lucerne meal) with and without 4% dust added. Dust additives included loess and kaolin, both of which have distinctively higher 87Sr/86Sr from the standard plant pelleted diet. These pellets were produced from a ground, pelleted diet (formula 2820 Granovit AG, Kaiseraugst, Switzerland) into which 4 wt% of loess or kaolin, respectively, were mixed. The mixture was then re-pelleted and fed to the animals to assess the influence of external, clay-to-silt-sized mineral dust particles on dental wear (see Winkler et al., 2020 for further details and the influence of the external abrasives on dental microwear texture). For Experiment-3: Dust Addition, only incisor enamel was sampled using LA-MC-ICP-MS.
Leaching Experiment
A leaching protocol was applied to all pelleted diets (each 2.5 – 3 g of whole pellets, see section “Controlled feeding experiment” for detailed description of the pellets) to evaluate potential differences in Sr chemical availability of Sr bearing phases. While the usual total bulk dissolution protocol involves a treatment with concentrated nitric acid overnight on a hot plate, the leaching experiment was performed with two dilutions of ultrapure hydrochloric acid for 3 h at 38°C to mimic conditions in a rodent stomach. For rats, we assumed a stomach pH value of 3 and a volume of 6 mL (Mcconnell et al., 2008) and for guinea pigs a pH of 3 and a volume of 15 mL (Merchant et al., 2011). To mimic stomach conditions intermediate between those of both animals, we selected a volume of 10 mL and a HCl concentration of 0.001 mol/L (pH 3). In addition, a second experiment was performed with a lower pH of 1 and acid concentration (0.1 mol/L HCl) to evaluate different dissolution conditions. After the experiment, the supernatant was separated from the un-dissolved solid remains by centrifugation at 6,000 min−1 for 5 min and evaporated to dryness. Afterwards, the supernatants of the leached pellet samples were treated as described in the section “Sample preparation.”
Sample Preparation
Bone (distal tibia) and enamel (sampled from the central part of the lower mandibular incisor) samples (1 – 5 mg) were used for solution-mode MC-ICP-MS Sr isotope analyses and digested using 500 μL of distilled trace metal grade concentrated nitric acid (Slovak and Paytan, 2012). Ball-milled food and fecal samples (25 – 50 mg) were digested in 4 mL of concentrated nitric acid and 1 mL of concentrated H2O2 (trace metal grade). All samples were placed in closed PFA screw-cap vials on a hot plate at 120°C overnight. This procedure removes the organic fraction of the foods prior to chemical separation (Eisenhauer et al., 2019). After digestion, samples were evaporated to dryness and re-dissolved in 2 mol/L HNO3. Water samples were acidified using concentrated nitric acid until a concentration of 2 mol/L HNO3 was reached.
Separation of Sr
For the separation of the Sr fraction, we used a prepFAST MC (ESI Elemental Scientific). The prepFAST is a low-pressure ion exchange chromatographic system (Romaniello et al., 2015) and is housed in a clean room at the Institute of Geosciences, University of Mainz. We slightly modified the default 1 mL Sr-Ca column (CF-MC-SrCa-1000) Sr-Ca separation protocol provided by ESI, by reducing the elution volume for the Sr fraction to 5 mL of 6 mol/L HNO3. We find that this reduction prevents the potential elution of Ca into the Sr fraction during Sr elution steps. After sample purification, sample aliquots were evaporated to dryness and re-dissolved in 250 μL concentrated HNO3 and 100 μL concentrated H2O2 to digest any potential organic remains (Eisenhauer et al., 2019). The samples were again evaporated to dryness and re-dissolved in 0.8 mol/L HNO3, in preparation for Sr isotope analysis.
Solution-Based MC-ICP-MS Sr Isotope Analyses
Analysis of bulk samples for Sr isotopes was performed at the Institute of Geosciences, University of Mainz, using a Thermo Neptune Plus MC-ICP-MS systems coupled to either a CETAC Aridus 3 or an ESI Apex Omega HF desolvating system. All solutions were prepared in 0.8 mol/L HNO3 with a Sr concentration of ~10 ng/g. Sample introduction was performed using a 100 μL/min nebuliser and a Jet-X-interface. The following ion beams were monitored during analysis: 82Kr, 83Kr, 84Sr, 85Rb, 86Sr, 87Sr, and 88Sr. A static multi-collection mode was applied to record all signals simultaneously within a single block of 50 cycles with 4.2 s integration time. Samples were bracketed and normalized to NIST SRM 987 (reference 87Sr/86Sr = 0.710248, McArthur et al., 2001). Analysis of NIST SRM 987 yielded a longterm average 87Sr/86Sr of 0.71028 ± 0.00003 (2 SD, n = 452). The methods closely follow Weber et al. (2018a), where the data evaluation is described in detail. Data evaluation was performed offline, using an in-house R script (R Core Team, 2013). Krypton correction was performed iteratively using the natural 86Kr/83Kr of 1.505657. All 87Sr/86Sr ratios from solution-based analyses are mean values with their respective 2 SE (standard error, see Weber et al., 2017 for details).
In-situ LA-MC-ICP-MS Sr Isotope Analysis
In situ analysis of samples for 87Sr/86Sr was performed by LA-MC-ICP-MS (laser ablation multi-collector inductively coupled plasma mass spectrometry) at the Institute of Geosciences, University of Mainz. Incisors were cleaned with ethanol and placed on a glass slide to ablate the outer enamel surface. We chose sampling areas in the base, middle and apex of each incisor, in order to evaluate enamel that formed at different times during the experiment (Figure 1).
A Neptune Plus (Thermo Scientific) MC-ICP-MS was coupled to an ArF Excimer 193 nm laser system (ESI NWR193), equipped with a TwoVol2 ablation cell. In addition, a CETAC Aridus 3 desolvating system was coupled to the sample line to introduce N2 and Ar and to enhance sensitivity. Further details about the instrumentation are given in Supplementary Table 2. The methods generally follow Weber et al. (2017) and are only briefly described here. Nine faraday cups were used to monitor signal intensities of the following m/z: 82Kr, 83Kr, 84Sr, 85Rb, 86Sr, 87Sr, and 88Sr, as well as the doubly charged half-masses of 167Er and 171Yb. Due to their small signals, Rb, Er, and Yb were monitored using Faraday cups equipped with 1013 Ω resistors, while all other signals were monitored with Faraday cups equipped with 1011 Ω resistors. For the laser ablation analysis, line scans of 500 μm length were applied with a circular spot size of 110 μm, a translation speed of 5 μm/s, a repetition rate of 50 Hz and a resulting fluence of 5 J/m2. Data evaluation and processing followed the methods described in Weber et al. (2017) and was performed using an in-house script in the statistical software R (R Core Team, 2013). Signals were corrected for Kr using the on-peak baseline technique and measuring a 30 s background without the laser firing prior to each analysis. Instrumental mass bias was corrected by applying the exponential law and an 88Sr/86Sr ratio of 8.375209. To correct 87Sr for 87Rb, we applied the mass bias correction factor and used the constant 87Rb/85Rb of 0.3857. We used a modern marine shark tooth as reference material to monitor drift during each analytical session. The long-term average for this shark tooth during this study yielded a 87Sr/86Sr of 0.70918 ± 0.00007 (2 SD, n = 96), in agreement with the modern-day seawater value of 0.70918 (McArthur et al., 2001). In addition, we used several bioapatite in-house reference materials (horse, elephant and shark teeth, human deciduous tooth, Supplementary Table 3), as well as internationally available apatite (bone ash NIST SRM 1400 and bone meal NIST SRM 1486) and carbonate (JCp-1, JCp-1, and NanoSr) reference materials (Okai et al., 2002; Inoue et al., 2004; Galler et al., 2007; Ohno and Hirata, 2007; Weber et al., 2018a, 2020). All results for 87Sr/86Sr are presented as the mean of each analysis with its corresponding 2 SE (standard error, see Weber et al., 2017 for further details).
Quantification of Sr Concentration
The Sr concentrations of water samples were determined using a Spectro CIROS Vision ICP-OES (inductively coupled plasma optical emission spectrometry) system at the Institute of Geosciences, University of Mainz.
Three rat incisors from the Diet Switch experiment with plant diet and three guinea pig incisors from each feeding group from the Basic Diet experiment were analyzed for their Sr/Ca ratio by in-situ LA-ICP-MS at the Institute of Geosciences, Mainz. An ArF Excimer 193 nm laser system (ESI NWR193), equipped with a TwoVol2 ablation cell, was coupled to an Agilent 7500ce ICP-MS. NIST SRM 612 was used for calibration purposes, while NIST SRM 610, USGS BCR-2G and Durango Apatite were used as quality control materials (Supplementary Table 4). Spot analyses with a circular spot size of 100 μm were employed, using a repetition rate of 10 Hz and a fluence of 3.5 J/cm2. Background time was set to 15 s, followed by 60 s of ablation. Data reduction was performed using an in-house spreadsheet based on Jochum et al. (2007). Details of the applied calculations are given in Mischel et al. (2017). The calcium isotope 43Ca was used as internal standard element. Results for the quality control materials are presented in Supplementary Table 4. The investigated rat incisors include animals euthanized after 0, 32, and 40 days on the experimental plant pellet (after initial 5 days of acclimatization, see section “Controlled feeding experiments,” Table 1 and Figure 1). The guinea pig incisors include animals from the Basic Diets experiment which received the experimental diet for the complete experimental duration of 54 days.
Strontium concentrations for pellets and their leachates were determined using a Thermo Neptune Plus MC-ICP-MS at the Institute of Geosciences, Mainz. The NIST SRM 987 was diluted to three different concentrations (1, 5, and 10 ng/g) for quantification of the sample Sr concentration. Sample introduction was performed according to the methods described for the solution MC-ICP-MS Sr isotope analysis.
Statistics
Data evaluation and statistical analyses for Sr isotopes were performed using the statistical software R (R Core Team, 2013). Processing of the data followed Weber et al. (2017). After interference correction, we performed a 2 SD outlier test for the mean of all individual 87Sr/86Sr ratios within one measurement and calculated a corresponding 2 SE uncertainty (standard error, SE = SD / ), both for solution based and laser ablation analyses. For the estimation of statistical significance, we used the implemented R function “wilcox.test” (Mann-Whitney-U-Test).
Water-Food Sr Mixing Models
In order to discuss the Sr isotope compositions measured in rat and guinea pig teeth and bones in relation to their dietary intake of Sr, we built a series of mixing models, to estimate the ranges of 87Sr/86Sr ratios as assimilated by the organisms and resulting from the mixing of Sr from drinking water and solid food. The basic principle consists of writing the mass balance of the contribution of water and food to the Sr eventually assimilated by the organism, also referred to as bioavailable Sr, expressed as follows:
where , and correspond to the daily mass of bioavailable Sr in total, from water and solid food, respectively. We consider a fractional assimilation of Sr from water to the blood fwt, and from food to the blood ffd referred to as bioavailability values and defined for both water and food as the proportion of Sr actually available for assimilation by the organism. We can thus express the total mass of daily bioavailable Sr as follows:
where and correspond to the daily ingested mass of Sr from water and food, respectively. The mass balance for Sr then dictates the following relation for the isotope composition of the total daily bioavailable Sr:
where corresponds to the radiogenic isotope composition of the bioavailable Sr in total (tot), water (wt), and food (fd). This relation can also be expressed using the Sr concentration ([Sr]wt, [Sr]fd) and daily ingested mass (mwt, mfd) of water and food:
This mixing equation was then used to draw maps of the total bioavailable Sr isotope composition in the 2D space of daily intake of water and food, as these values vary with time, between individuals and from one species to another. A map was drawn for each pellet type and for each measured isotope composition of pellet leachates (which were taken as food bioavailable Sr end-member values). Two sets of maps are presented in the main text and Supplementary Information, one with distinct bioavailability values for water and food Sr and another with equal bioavailability values for both (assuming no differential assimilation of Sr from water and food). The calculations and plots were performed using the statistical software R (R Core Team, 2013).
Results
Food and Water
An overview of the Sr isotope ratios, as well as the Sr concentration of each diet and water are given in Table 2. Supplier diets for the two rodent species differed, with a 87Sr/86Sr of 0.70944 ± 0.00002 (2 SE, here and in the following if not specified otherwise) for the guinea pigs and 0.70891 ± 0.00001 for the rats. During their time at the breeder facilities, the animals received de-mineralised drinking water with Sr concentrations below the quantification limit of the ICP-OES system. Therefore, the contribution of the Sr isotope composition of supplier water is not considered to have influence on body tissues 87Sr/86Sr while the animals were at the breeding facilities.
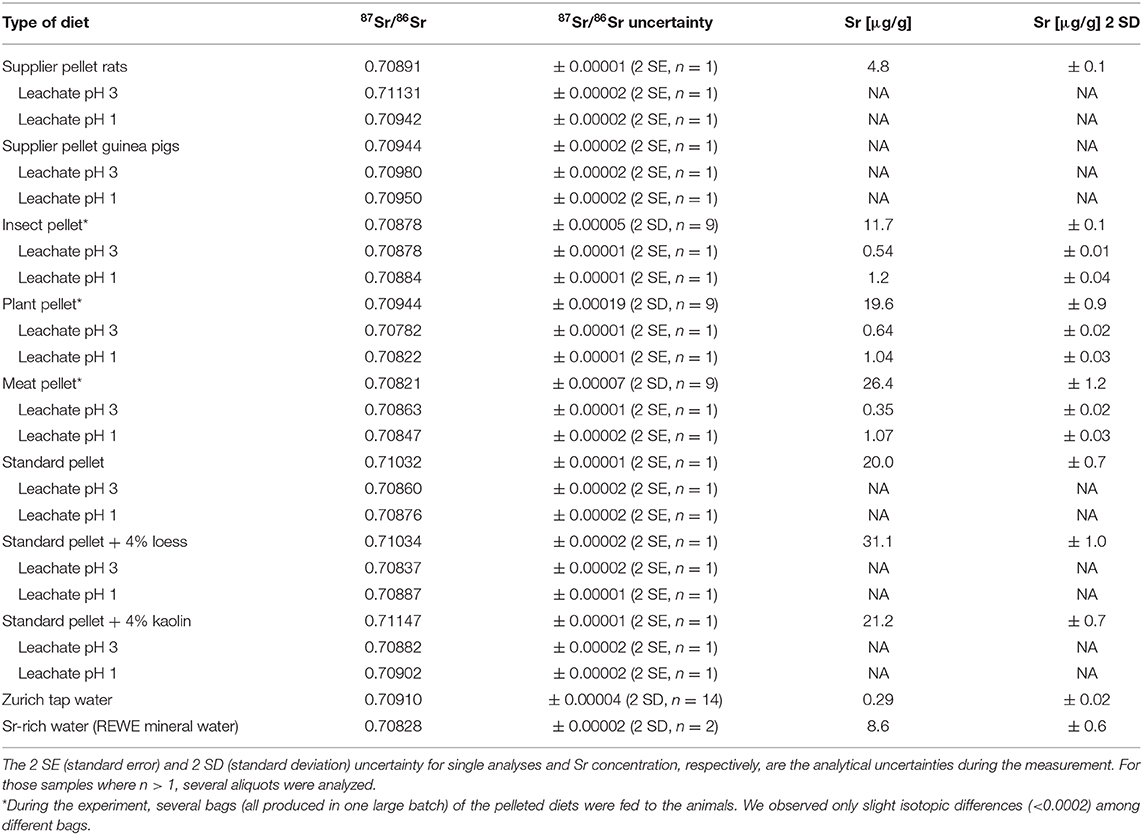
Table 2. Sr isotope composition and Sr concentration for all diets (including leachates) and waters provided to the animals in this study.
The Zurich tap water showed slight seasonal variation in 87Sr/86Sr, ranging from 0.70907 ± 0.00003 to 0.70914 ± 0.00003, with an average value of 0.70910 ± 0.00004 (2 SD). In contrast, the Sr-rich mineral water yielded a much lower 87Sr/86Sr of 0.70828 ± 0.00002 (2 SD), with a Sr concentration ca. 30 times higher than the Zurich tap water. The supplemental vitamin C received by the guinea pigs did not yield Sr concentrations above the blank level after sample preparation, and can therefore be ruled out as a factor influencing 87Sr/86Sr in these animals.
The insect, plant and meat diets differed distinctively in their Sr isotope composition (Table 2). Analyses of ball-milled, powdered samples of the pellets which were completely digested with concentrated HNO3 yielded different results compared to the leaching experiment. In HNO3 bulk digested samples, the meat pellet had the lowest 87Sr/86Sr of 0.70821 ± 0.00007 and the plant pellet the highest 87Sr/86Sr of 0.70944 ± 0.00019 (2 SD). An intermediate ratio was observed for the insect diet, with an average 87Sr/86Sr of 0.70878 ± 0.00005 (2 SD). In the leaching experiment, the plant diet leachate yielded distinctively lower 87Sr/86Sr of 0.70782 ± 0.00001 at pH 3 and 0.70822 ± 0.00001 at pH 1 than for the fully digested pellet. The meat diet leachate yielded slightly higher ratios compared to the HNO3 digested pellet (87Sr/86Sr of 0.70863 ± 0.00001 at pH 3 and 0.70847 ± 0.00001 at pH 1), while the insect diet showed similar values for the leaching experiment as for the completely dissolved pellets with 87Sr/86Sr of 0.70878 ± 0.00001 at pH 3 and 0.70884 ± 0.00001 at pH 1.
The completely dissolved rat supplier pellet yielded a much lower 87Sr/86Sr (0.70891 ± 0.00001) than both leachates at pH 3 (0.71131 ± 0.00002) and pH 1 (0.70942 ± 0.00002). In contrast, the bulk solution of the guinea pig supplier pellet yielded a lower 87Sr/86Sr (0.70944 ± 0.00002) than both leachates (0.70980 ± 0.00002 at pH 3 and 0.70950 ± 0.00002 at pH 1).
The highest bulk ratios of 87Sr/86Sr were obtained from the experiment in which mineral dust (loess and kaolin) was added into the standard pellet (0.71032 ± 0.00001). While the addition of 4 wt% loess did not change the 87Sr/86Sr notably (0.71034 ± 0.00002), the addition of 4 wt% kaolin shifted the Sr isotope ratio to a higher ratio of 0.71147 ± 0.00001 (Table 2). All leachates from these three pellets from the Dust Addition experiment yielded lower 87Sr/86Sr than their bulk solutions, where the leachates at pH 1 were generally higher in 87Sr/86Sr than at pH 3 (Table 2).
Strontium Isotope Incorporation Over Time (Experiment-1: Diet Switch)
To establish baseline values for the initial Sr isotope composition (i.e., supplier food derived 87Sr/86Sr) of the rats, three animals were euthanized after the initial 5 days acclimatization period. The Sr isotope compositions of their hard tissues are therefore only influenced by the food and the water supplied by the breeder. Two animals had very similar bone and enamel 87Sr/86Sr, however, one individual, despite a similar bone 87Sr/86Sr (between 0.70918 ± 0.00001 and 0.70924 ± 0.00001), had a higher enamel 87Sr/86Sr, especially in the apex and middle part of the incisor (up to 0.71024 ± 0.00009, Figure 2, i.e., the older parts of the incisor). It is therefore necessary to take this initial variability in 87Sr/86Sr into consideration when interpreting data from the Diet Switch experiment. However, the observed 87Sr/86Sr of those animals (independent on tissue type) fall within the range of the rat supplier pellet when considering bulk and leachate data (Figure 2 and Table 2), indicating that this variability is caused by the diet.
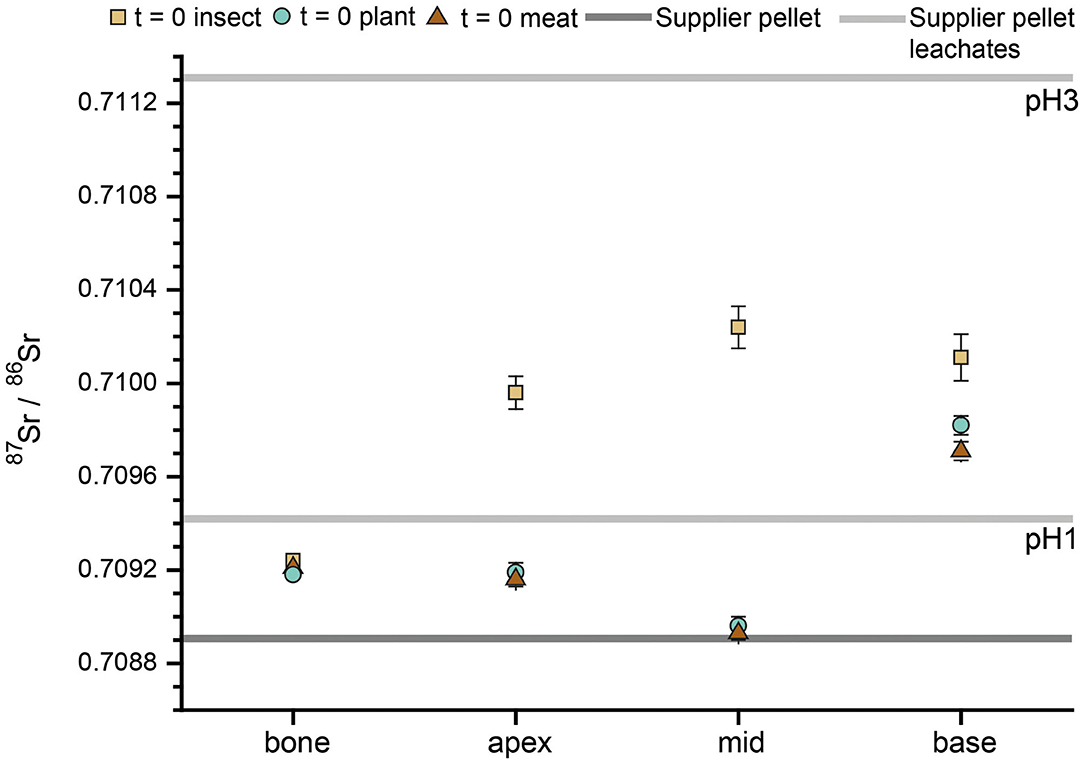
Figure 2. Incisor enamel 87Sr/86Sr of three rats from t = 0 of the Diet Switch experiment. The dark gray line represents the 87Sr/86Sr from the supplier pellet, which the animals received during their entire life, post-weaning. The leachates (pH 3 and pH 1) of the supplier pellet are shown as light gray lines. For the last 5 days before euthanization, the animals were provided with both, supplier and experimental pelleted diets. Values for apex, middle and base were obtained by laser ablation, the bone and pellets were analyzed by solution mode MC-ICP-MS.
The temporal evolution of 87Sr/86Sr in the hard tissues of rats euthanized at different time points during the experiment (n = 8 for insect and plant group, n = 9 for meat group, as well as n = 6 for each group with full experimental duration) is shown in Figure 3. Day 0 is designated as the 1st day that animals received only the experimental diet (i.e., after the initial 5 days acclimatization period). At the beginning of the experiment, all rats display similar bone 87Sr/86Sr starting values of 0.7091–0.7093. This is slightly higher than the value expected based on the bulk rat supplier diet (~0.7089), but lower than the leachate at pH 1 (~0.7094). Over the course of the experiment, the bone values show a linear trend (R > 0.9, Figure 3) toward lower ratios, which differed slightly among the diet groups (see section below for further details). In all groups, the incisor enamel 87Sr/86Sr tends to lower ratios at the base (i.e., the youngest part, grown later in the experiment, compared to the apex, which grew in the beginning of the experiment). The base usually reaches a plateau value by the middle of the tooth. Some spikes of higher 87Sr/86Sr are visible, particularly in the apex. This part of the incisor is formed earliest and this mainly reflects supplier food which had high 87Sr/86Sr values in the leachate. These spikes are most prominent in the animals that were fed the insect diet. The 87Sr/86Sr obtained for animals that received an experimental diet for the full 54 days duration of the Basic Diets experiment does not necessarily agree with the bulk diet 87Sr/86Sr (i.e., when digested with concentrated nitric acid) obtained for the pellets. For instance, the incisor enamel 87Sr/86Sr of the rats receiving the plant diet have much lower ratios than their diet and somewhat lower values than their drinking water. The enamel 87Sr/86Sr of those on the insect diet largely match the Sr isotope composition of their food. Overall, the rats fed with the meat diet have 87Sr/86Sr above their diet, but below their drinking water. The incisors show a better agreement with their respective experimental foods when compared to the diet Sr isotope data obtained in the leaching experiment (Figure 3).
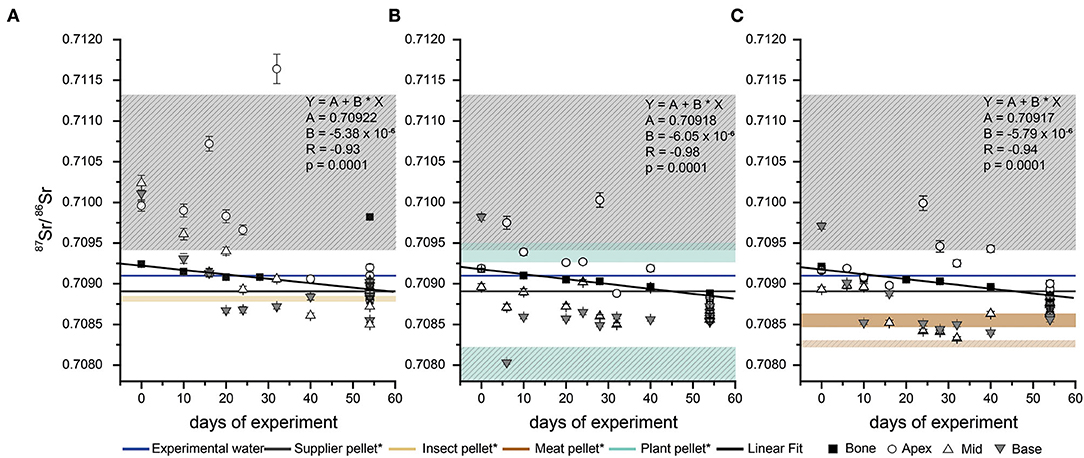
Figure 3. Temporal evolution of the 87Sr/86Sr from different parts of the rat incisors and the bone during the Diet Switch experiment. For each feeding group, supplier pellet (which animals received prior to the experiment) and Zurich tap water (supplied during the feeding experiment) values are indicated by horizontal lines. Each x-axis value represents a single individual. The coloured boxes indicate the 87Sr/86Sr range, derived from the leaching of the pelleted diets: (A) insect pellet (beige) (B) plant pellet (cyan) (C) meat pellet (brown) and (A–C) supplier pellet (gray). All values were obtained by laser ablation, except for the bone values. The error bars indicate the analytical uncertainty for each data point. *The hatched boxes for the meat, plant and supplier diet represent the 87Sr/86Sr from the completely dissolved pellets. For the insect pellets, the 87Sr/86Sr overlap for both digestion types. The bone turnover shows a linear trend (black line) with the given R and p-values for each diet group.
Evolution of the Sr/Ca Ratio From the Supplier Toward the Experimental Diet (Experiments-1: Diet Switch and 2: Basic Diets)
Rats from the Diet Switch experiment show variable trends in their Sr/Ca ratio (Figure 7). These animals received the plant diet for 0, 32, and 40 days. The rat euthanized at t = 0 (Figure 7) has the lowest Sr/Ca of 8.2 x 10−5 ± 1.3 × 10−5. The animal euthanized at t = 32 days shows increasing Sr/Ca toward the base (Figure 7), up to a ratio of ~4 × 10−5. The initial values at the apex agree with the individual on the supplier diet (Sr/Ca ~ 8 × 10−5). A plateau value (discounting the last base measurements) for Sr/Ca is reached for the most recently grown ~30% of the total incisor enamel length. The rat euthanized at t = 40 days (Figure 7) shows a pattern similar to the individual fed for 32 days. The starting value is slightly higher than the ones for day 0 and day 32 individuals (1.0 × 10−4) and a plateau of ~2.8 × 10−4 is reached at 50% of the total incisor enamel length.
In the guinea pigs from the Basic Diets experiment, the animal that received the plant diet shows the highest Sr/Ca (7.8 × 10−4) with slightly increasing values toward the base (Figure 7). The animal fed the meat diet starts with a value similar to that of the guinea pig on the plant pellet at the apex and then this value progressively decreases to a minimum of ~5 × 10−4 in the middle of the incisor. Toward the base (i.e., the ontogenetically youngest part of the incisor), the Sr/Ca for the individual on the meat pellet progressively increases again (Figure 7). The enamel of the guinea pig that received the insect diet starts at a lower Sr/Ca than the two other individuals and decreases to a value similar to that of the meat diet specimen. Afterwards, the Sr/Ca increases again toward the base to a Sr/Ca that is similar to the two other individuals (Figure 7).
Sr Isotope Composition of Bones and Teeth After the Full Experimental Duration (Experiment-2: Basic Diets)
For the 87Sr/86Sr of the rats fed with the three basic diets for the full experimental duration (5 + 54 days), the bone values are similar across and within the feeding groups, except for one individual in the insect diet group (Figure 4). The 87Sr/86Sr obtained by LA-MC-ICP-MS for the incisors display a larger variability in comparison to the bones. However, the 87Sr/86Sr generally tends to have lower ratios toward the base of the incisor. The middle and base part of the incisors usually show similar results, while the apex (oldest) showed (some) higher 87Sr/86Sr. Interestingly, the 87Sr/86Sr from the middle of incisors obtained by solution-based MC-ICP-MS are distinctively lower than the LA values by 0.0003 ± 0.0001 (1 SD) on average. As described in section “Strontium isotope incorporation over time (Experiment-1: Diet Switch),” enamel 87Sr/86Sr deviate from bulk pellet 87Sr/86Sr values and instead better agree with the data from the pellet leachates. The 87Sr/86Sr of rat fecal samples (n = 3) differ from those of the bulk pellet values (Figure 4). The rats that received the Sr-rich mineral water and the plant diet have lower 87Sr/86Sr than the rats that received the tap water. Both the bone and the incisor values are lower and shifted toward the 87Sr/86Sr of the Sr-rich mineral water (Figure 4).
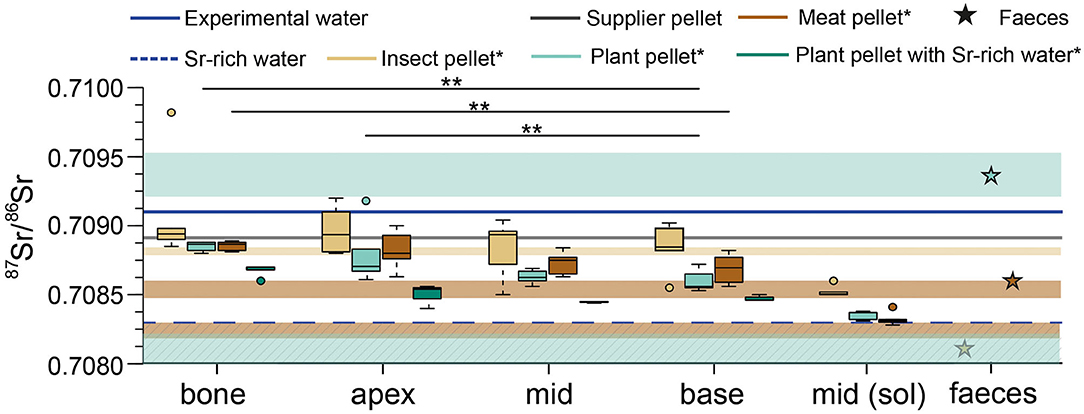
Figure 4. Strontium isotope ratios as boxplots after the full experimental duration (54 + 5 days) for the different feeding groups of rats from the Basic Diets experiment. The 87Sr/86Sr of the supplier pellet and the experimental water supply is indicated by the horizontal lines. The coloured boxes indicate the 87Sr/86Sr range of the food pellets, derived from the leaching of the pellets: insect pellet (beige), plant pellet (cyan), meat pellet (redbrown Sr isotope ratios for apex, mid and base of the incisor were obtained by laser ablation. For each basic feeding group one fecal sample was analyzed. Apex, middle and base values were obtained by laser ablation. *The hatched boxes for the meat and the plant pellet show the 87Sr/86Sr ratios for the completely dissolved pellets. For the insect pellets, the 87Sr/86Sr overlap for both types of dissolution. The leachates for the supplier pellet are not shown due to better clarity (see Figure 2). Significance levels are presented as stars between bone and base (plant and meat group), as well as for apex – base pairs of the plant group).
In general, the changes in 87Sr/86Sr described for the rats are also observed in the guinea pigs (Figure 5). All animals have higher 87Sr/86Sr in enamel in the apex of the incisor, even exceeding the values measured for bone. This is most likely related to the higher 87Sr/86Sr ratios of ~0.7094 (pH 1 and 3 leachates at ~0.7095 and ~0.7098, respectively) of the guinea pig supplier food and the slower growth rate of guinea pig incisors (0.1 – 0.3 mm/day) compared to rats (0.5 mm/day). For some individuals, especially those fed the insect diet, we observed higher values in the middle part compared to the base, which mineralised just prior to the termination of the experiment.
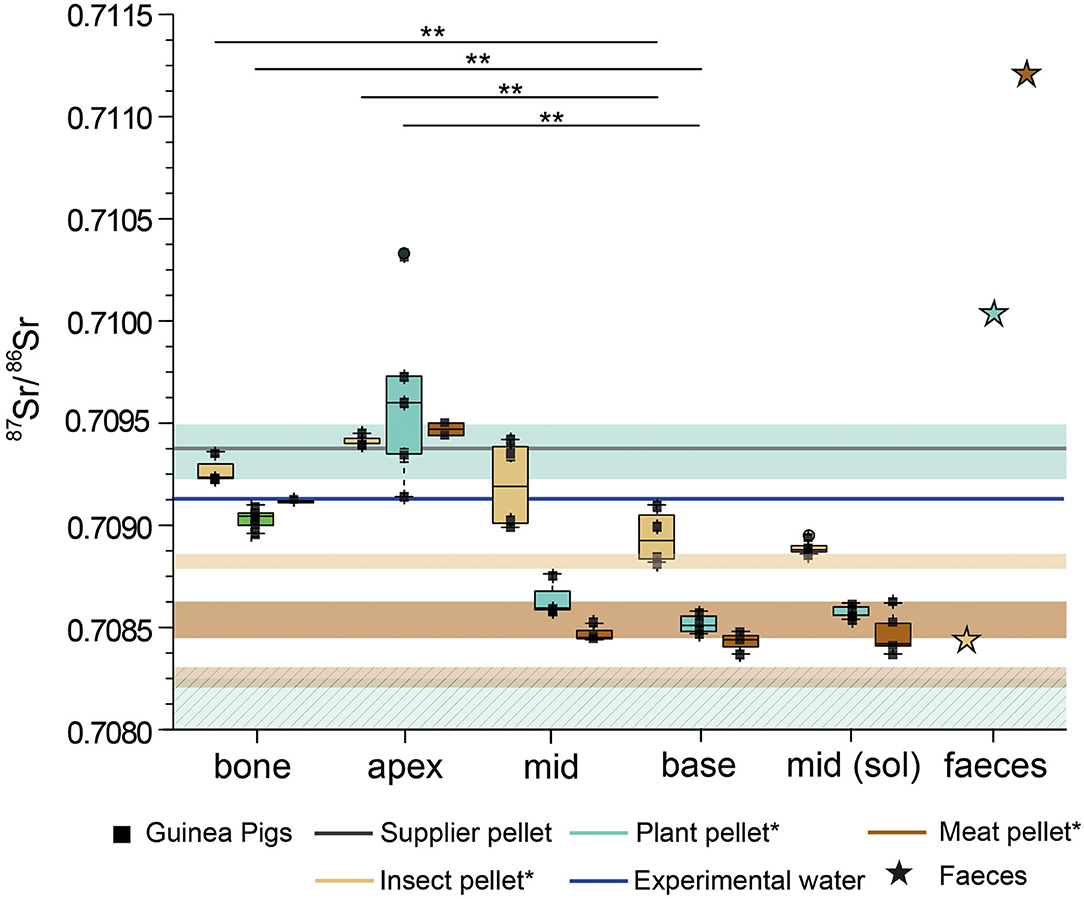
Figure 5. Strontium isotope ratios as boxplots with underlying data points after the full experimental duration (54 + 5 days) for the different feeding groups of guinea pigs from the Basic Diets experiment. The 87Sr/86Sr of the supplier pellet and the drinking water during the experiment is provided. The coloured boxes indicate the 87Sr/86Sr range of the food pellets, derived from the leaching of the pellets: insect pellet (beige), plant pellet (cyan), meat pellet (brown). Strontium isotope ratios for apex, mid and base of the incisor have been obtained by laser ablation. For each basic feeding group one fecal sample was analyzed. *The hatched boxes for the meat and the plant pellet show the 87Sr/86Sr from the completely dissolved pellets. For the insect pellets, the 87Sr/86Sr overlap for all types of dissolution. Significance levels are presented as stars between bone and base, as well as for apex – base pairs.
In summary, bone and enamel 87Sr/86Sr in the insect group are higher than those of the two other groups. Values for guinea pigs fed in the plant group do not reflect the 87Sr/86Sr of the completely dissolved pellets, but do approach the values measured in the leachate of these same pellets. Guinea pig faeces from the plant and meat diet groups show higher 87Sr/86Sr than rat faeces from animals fed the same diets, while faeces from the insect fed guinea pigs and rats are very similar (Figure 5).
Influence of External Dust Sources (Experiment-3: Dust Addition)
Guinea pigs fed the standard diet are compared with guinea pigs fed the exact same standard diet, but with the addition of 4 wt% of loess and kaolin (for 23 – 29 days, Figure 6). In the apex, the 87Sr/86Sr of the three different feeding groups largely overlap, with two outliers (2 SD outlier test) at either side of the range, both of which are animals fed the standard pellet. While the enamel toward the middle of the incisor is characterized by a larger scatter and overlap in 87Sr/86Sr, toward the incisor base the 87Sr/86Sr decreases progressively. Here, a difference between the animals that received the kaolin pellet and those that received the loess or standard pellet was observed. The guinea pigs fed the standard pellet show a large variation, covering nearly the entire 87Sr/86Sr range of all base samples. All incisor 87Sr/86Sr are distinctively lower than the 87Sr/86Sr of their respective food pellets, indicating that a fraction of (radiogenic) Sr from the pelleted diet was not bioavailable to the animals. Instead, values are similar to those of the drinking water and the leachates at pH 1.
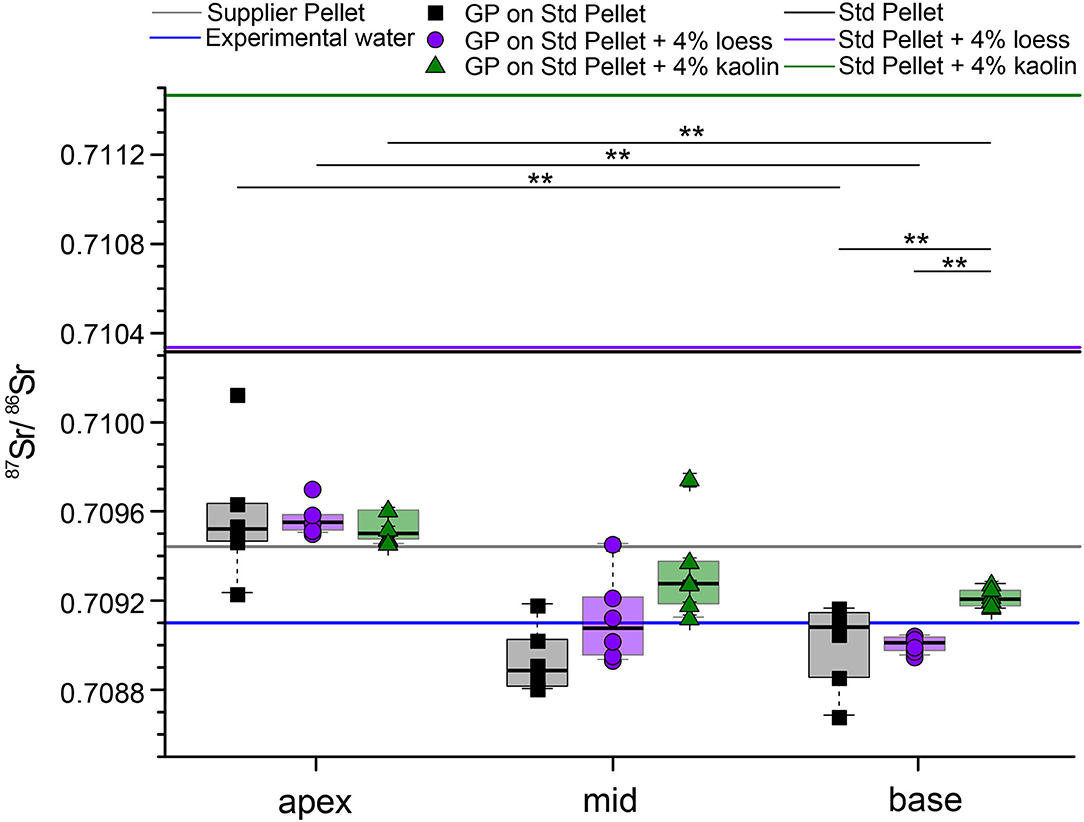
Figure 6. Strontium isotope ratios of incisors from guinea pigs fed for up to 29 days on a standard pelleted diet with and without addition of 4 wt% loess or 4 wt% kaolin, respectively (Dust Addition experiment). The 87Sr/86Sr of the supplier pellet and the Zurich tap water supply is plotted for comparison, as well as the 87Sr/86Sr of the three different pelleted diets. All enamel 87Sr/86Sr were determined by LA-MC-ICP-MS. Significance levels are presented as stars between apex and base pairs, as well as within the base among different feeding groups.
Discussion
Isotopic Homogeneity of Food and Water and Sr Bioavailability Assessed by Leaching
The variability of the 87Sr/86Sr of the pelleted diets among bags of the same batch was negligible for meat and insect pellets (2 SD = 0.00007 and 0.00005, respectively, each n = 9), when entirely dissolved in concentrated nitric acid. However, the plant pellets showed a two to four times higher variability of 2 SD = 0.00019 (n = 9). The seasonal variability of the Zurich tap water was low (2 SD = 0.00004 over a period of 1.5 years, n = 14) and is therefore also considered to be negligible since each experiment only lasted about 2 months. Therefore, the Sr isotope input from both food and water supply seem to have been mostly homogeneous and provided an isotopically constant Sr source during the controlled feeding experiment.
The 87Sr/86Sr measured in totally digested basic food pellets and the HCl leaching experiments at the two different pH levels (pH 1 and 3) yielded different 87Sr/86Sr ratios. While no distinct differences between the 87Sr/86Sr of the leachate and the whole food for the insect diet were observed, this difference seemed relevant for the meat diet, and was particularly large for the plant diet. These differences were however, in opposite directions, with the 87Sr/86Sr values of the leachates higher than the bulk meat diet, and lower for the plant diet. For the formulation of the pellets, which were produced specifically for our feeding experiments, different components were mixed, resulting in potentially different Sr bearing phases displaying varying 87Sr/86Sr end-member values and Sr bioavailabilites (see Supplementary Table 1 for pellet ingredients). The different chemical environments of the Sr bearing phases are likely to induce a differential bioavailability of Sr to the animal. While Sr from some of the ingredients is highly available during the biological processing of food (e.g., digestion, metabolization), Sr from other ingredients may not have been as bioavailable. This is supported by the leaching data. The weaker acid (0.001 mol/L HCl) used for the leaching experiment, designed to mimic rodent stomach conditions, released Sr from Sr bearing phases where it is more bioavailable to the animals during digestion. This causes the pellet leachate 87Sr/86Sr to converge on a 87Sr/86Sr value ranging between 0.7082 and 0.7088. This range matches the range observed in the incisor data well (Figures 4, 5). We therefore assume that the diet 87Sr/86Sr data obtained from the leaching experiments best reflects the Sr bioavailable to the animals.
Incorporation of Sr in the Ever-Growing Incisor Enamel (Experiment-1: Diet Switch)
The ever-growing incisors of rodents allow us to temporally constrain the incorporation of Sr into enamel. It is especially important to precisely determine which regions of the incisor were completely equilibrated with the experimental diet in order to further investigate the influence or the experimental diet on the isotopic composition of the dental hard tissues. We performed both Sr/Ca and Sr isotopic analyses on the incisors of rats which underwent a diet switch from a supplier diet to one of our custom designed diets in our controlled feeding experiment. The Sr/Ca of the rat that only received the supplier pellet (Figure 7) showed no clear trend, with only a slight increase in value toward the most proximal part at the base (i.e., the youngest part at a very early enamel mineralisation state). In comparison, the rats fed one of the experimental diets for 32 or 40 days showed an increase in Sr/Ca. While the individual that received the plant diet for 32 days had a value of Sr/Ca at the apex similar to the t = 0 animal, the individual fed for 40 days displayed a slightly higher apex Sr/Ca ratio. The transition to higher values in Sr/Ca ratios corresponds with the experimental diet thus appears to be phase shifted from t = 32 to t = 40 as expected. However, this difference is small and only based on this single individual. We therefore cannot rule out the possibility that this difference is simply the result of natural variability among individuals. Nevertheless, based on this observation, we propose that even the oldest part of the incisor (up to 60 days, see Figure 1), which grew for about 10 days prior to the start of the feeding experiment, was slightly influenced by the composition of the experimental diet. This observation can be explained by the enamel maturation process. Rodent enamel can be divided into three distinct zones of immature, maturing and mature enamel (Leblond and Warshawsky, 1979), and the chemical composition of enamel changes through the addition and maturation of bioapatite until the final maturation stage (i.e., full mineralisation of enamel) is reached. The whole process of enamel formation (~1.5 days of pre-secretion, ~7 days of secretion, and ~20 days of maturation) can take up to 30 days (Leblond and Warshawsky, 1979). After the initial secretory stage of enamel formation, the maturing enamel remains an open system and can be influenced by dietary changes occurring after the initial secretory stage, resulting in the phase-shifting of time-dependent chemical signals (Passey and Cerling, 2002). This process also explains why there is no sharp transition in chemical composition from supplier to experimental diet, but rather a smooth one toward a plateau of Sr/Ca (Figure 7). Indeed, in addition to the phase-shifting, maturation of enamel also induces a dampening of time-dependent chemical signals. This plateau was reached for the last ~30% of the incisor length for the rat fed for 32 days, while in the rat fed for 40 days this plateau comprised the full lower 50% of the incisor length (Figure 7), indicating the progression of the maturation front toward the apex because of the longer period of incisor growth and maturation.
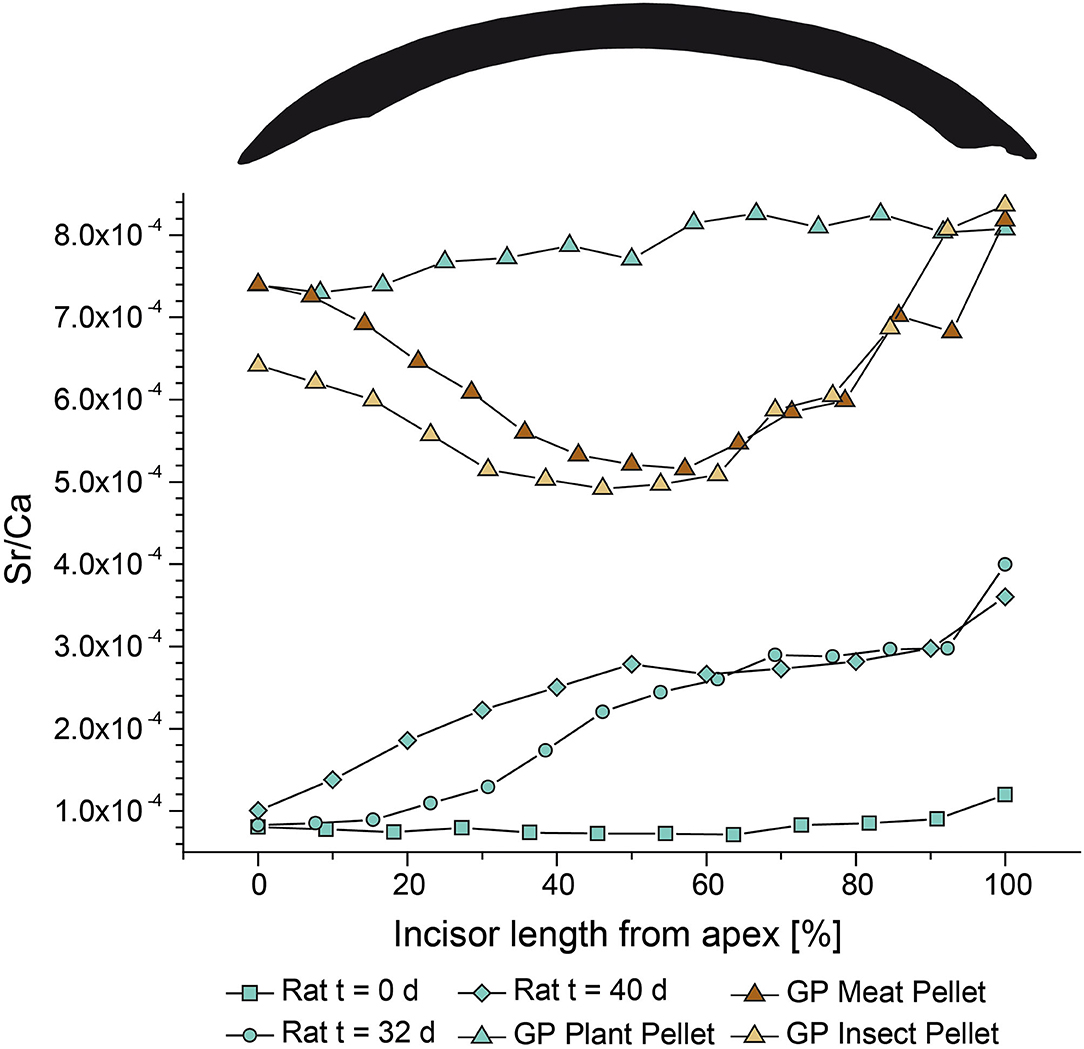
Figure 7. Enamel incisor Sr/Ca of rats (each n = 1) from different points in time during the Diet Switch experiment. Rats were fed with the plant pellet for 0 days (cyan squares), 32 days (cyan circles), or 40 days (cyan diamonds). The Sr concentrations of the supplier and experimental plant diet are given in Table 2. Enamel incisors Sr/Ca for guinea pigs (each n = 1) from the Basic Diets experiment which were fed for the full experimental duration (54 + 5 days) with the three different pelleted diets based on insect (beige triangles), plant (cyan triangles), and meat meal (brown triangles), respectively.
Similar patterns were also recorded in the Sr isotope composition of the rat incisors during the Diet Switch experiment (Figure 3). Generally, isotopic turnover induced by the diet switch is fastest at the base, since it is the youngest-forming part of the incisors and reflects the most recent Sr input incorporated into the enamel by secretion and maturation. However, the three feeding groups differed slightly in terms of when they reached equilibrium with the bioavailable 87Sr/86Sr in the experimental diet in the base part. While the animals fed the plant diet reached 87Sr/86Sr values that reflected bioavailable 87Sr/86Sr from the experimental diet after just 10 days, the rats that received the insect diet needed at least 20 days to reach this value. The rats fed the meat diet were more variable. The observed differences in the timing of equilibrium may either be caused by intra-individual variability, or by the differences in 87Sr/86Sr between supplier diet and experimental diets. This difference is largest between supplier and plant pellet (leachates), indicating that the corresponding turnover signal might be enhanced due to the offset. Interestingly, while the individual that received the meat diet for 10 days seemed to have already reached a diet-related 87Sr/86Sr, the individual fed the same diet for 16 days, had an elevated 87Sr/86Sr. Here it is important to note, that each point in time is represented by a different individual (n = 1), which may have contributed to inter-individual bias due to slight differences in individual metabolism and feeding behavior. Similarly, a distinct time point at which a diet-related 87Sr/86Sr equilibrium is reached cannot be detected in the middle part of the rat incisor. The animals fed the meat diet had lower 87Sr/86Sr in the middle of the incisor in comparison to the base between 16 and 32 days of experiment, which were both lower than the 87Sr/86Sr after the full experimental duration of 54 days. The oldest part of the incisor (apex) always lagged distinctively behind the trend visible in the younger incisor parts. In addition, pronounced radiogenic Sr spikes (i.e., 87Sr/86Sr increases up to 0.002) are detected in every feeding group, but were most prominent in the rats fed the insect diet. However, most of these spikes toward higher 87Sr/86Sr occurred in the apex, likely representing enamel that mostly mineralised before the start of the experiment. Therefore, these radiogenic inputs occurred during the supplier period, beyond the scope of the feeding experiment. As the supplier diet (the supplier water was demineralized and thus nearly Sr-free) did not yield high 87Sr/86Sr, the source of these high values can only be explained by the bioavailable leachate values of the rat supplier pellet, reaching a 87Sr/86Sr as high as ~0.7113 at pH 3.
The guinea pigs fed for the whole experimental duration show distinct Sr/Ca patterns among the feeding groups (Figure 7). While the Sr/Ca was similar in the apex region for all three individuals (still reflecting a remnant supplier food-related value), toward the middle part, the Sr/Ca clearly diverged. While the animals fed with plant pellets showed an increasing value similar to that seen in rats, guinea pigs fed the insect and meat diets had decreasing Sr/Ca ratios. This agrees with literature data showing that in general, Sr/Ca decreases with increasing trophic level due to Ca-biopurification processes (e.g., Elias et al., 1982; Burton and Wright, 1995; Burton et al., 1999; Blum et al., 2000; Balter et al., 2002). However, toward the base, the Sr/Ca of all three individuals increased and converged again, producing an U-shaped pattern for the two animals fed with animal-based (insect and meat) diets, while the plant diet individual is characterized by a similar Sr/Ca throughout the feeding period (Figure 7). This pattern could be an artifact of (i) the lower mineralisation state and lower Ca content of the base (Leblond and Warshawsky, 1979; Hillson, 2005), or (ii) an increased portion of dentin sampled during laser ablation, which typically has a higher Sr concentration in comparison to enamel (Kohn et al., 2013). However, the data seem to show that during secretion, there is more selectivity for Sr than during the maturation phase. The differences in the Sr/Ca ratio at the base of the incisor between rats and guinea pigs could also indicate that the area of progressively maturing enamel is spread across a larger portion of the incisor base in the latter compared to the former.
Our data suggests that a complete turnover of the Sr incorporated in the incisor enamel takes about 2 months for rats and a similar or longer time for guinea pigs. This difference might be related to the different incisor growth rates, as well as the difference in size of the animals. While the rats (11 – 12 weeks old) started at 197 ± 12 g (n = 24) and did not exceed a body weight of 300 g (Supplementary Figure 5), the guinea pigs (3 – 5 weeks) started at 398 ± 17 g (n = 18) and reached a final body weight of up to >700 g (Supplementary Figure 7). In the guinea pigs, therefore, one could speculate that incisors were not only growing for replacement of tissue lost by gnawing, but were also growing allometrically as a function of overall body size increase.
In contrast to the larger 87Sr/86Sr variability visible in the fast and continuously growing enamel, we observed a linear trend of 87Sr/86Sr in bone for the Basic Diets experiment (R > 0.9). Bone values at the start of the experiment reflect those of the supplier diet while those toward the end of the experiment reflect to the experimental diet. All animals from the three feeding groups on the three different diets started with a bone 87Sr/86Sr of ~0.7092 and gradually moved toward lower 87Sr/86Sr values. After the full 54 days, bones reached a final 87Sr/86Sr between 0.7088 and 0.7090. This was slightly higher than the 87Sr/86Sr of the bioavailable Sr leached from the pellets during the leaching experiments. However, the enamel 87Sr/86Sr was slightly shifted toward higher values, possibly due to the additional influence of drinking water on an animal's Sr-uptake. In contrast to the incisor enamel, bone is a tissue with a much lower tissue turnover rate (Bryant and Loutit, 1964; Dahl et al., 2001). Therefore, the Sr isotope composition of bone averages the 87Sr/86Sr of the body water pool over a much longer time period than the ever-growing incisors.
Bioavailability of Sr From the Pelleted Diet (Experiment-2: Basic Diets)
It is generally accepted that the 87Sr/86Sr recorded in teeth and bones represents the locally bioavailable Sr isotopic signature at the basis of a local food chain (e.g., Blum et al., 2000; Slovak and Paytan, 2012; Flockhart et al., 2015; Lewis et al., 2017). The hard tissues, bones and incisor enamel, from animals of all three different feeding groups that fed the same basic diets for the full experimental duration did not reflect the 87Sr/86Sr of their respective, completely dissolved pellets. For bone it is expected that, due to its slow turnover rate, equilibrium with the experimental diet is not reached after a relatively short experimental duration (i.e., 54 days, Figure 1). A full turnover would take longer, and it is reasonable to assume that most of the Sr stored in the bones still derives from food consumed during the period the animals spent at the supplier facility. However, the ever-growing enamel of the incisors was expected to record the bioavailable Sr input from the diet and drinking water entirely toward the end of the experimental duration, especially the most recently grown and mineralised parts (i.e., the base and the middle area). Nevertheless, we observed distinct differences between the 87Sr/86Sr of the diets and the enamel. In addition, the 87Sr/86Sr obtained by LA-MC-ICP-MS were higher than those obtained by solution-based MC-ICP-MS by up to 0.0005 (Figures 4, 5). This is a common observation for Sr isotope analyses by LA-MC-ICP-MS and is usually explained by unresolved interferences (Rb, Kr, REEs, Ca argides and dimers, CaPO for apatites) which influence the in-situ measurements (Copeland et al., 2008, 2010; Horstwood et al., 2008; Simonetti et al., 2008; Lewis et al., 2014). The effect was more pronounced for the rat samples, since these incisors had a much lower Sr/Ca (~3 × 10−4, Figure 7) than the guinea pig incisors (up to 8 × 10−4, Figure 7) and are thus, due to their lower Sr content, more prone to such interferences. However, we tried to minimize this effect by adding nitrogen to the sample gas flow, as N2 is known to reduce potential interferences (Zhang et al., 2018), using a desolvator system, and monitored the accuracy of the 87Sr/86Sr using several matrix-matched tooth samples with known 87Sr/86Sr (Willmes et al., 2016). By plotting the accuracy of 87Sr/86Sr vs. 1/88Sr [V] (Supplementary Figure 2), one would expect a strong linear correlation if polyatomic interferences on m/z = 87 (40Ca31P16O) will affect the 87Sr/86Sr (Horstwood et al., 2008; Lugli et al., 2017b). However, we did not observe such a correlation, which most likely excludes this explanation. Due to the low Sr concentration of the rodent incisors, it was necessary to employ a large spot size and repetition rate to increase the test portion mass (see method section) to achieve a sufficient signal intensity during LA-MC-ICP-MS acquisition for reliable Sr isotope analysis (usually >1 V for 88Sr, e.g., Müller and Anczkiewicz, 2016; Weber et al., 2018a). This may have resulted in ablation of both enamel and the underlying dentin during laser ablation (see Supplementary Figure 1). There is a time lag between enamel and dentin formation in mammals (Hillson, 2005) which could, if dietary input changes temporally, potentially lead to differences in the 87Sr/86Sr of both dental tissues. As dentin forms prior to enamel, it will therefore have incorporated higher 87Sr/86Sr (in our case Sr input from the supplier feeding period), as demonstrated by the analyses of individuals from the Diet Switch experiment. Most recently formed enamel shows lower 87Sr/86Sr, since the source of its Sr is already shifted further toward the experimental diets. This could explain the differences between the in-situ and solution-based 87Sr/86Sr analyses, since for the latter the enamel was carefully separated from the dentin prior to the digestion and purification of Sr for solution-based 87Sr/86Sr analyses. Interestingly, the average guinea pig 87Sr/86Sr are generally higher by 0.0002 when compared to the rats, although both rodent groups received the same diets and water, potentially due to an increased absorption of water-derived Sr for the guinea pigs. In addition, the rodent incisor enamel 87Sr/86Sr does not record either the bulk ingested pellet nor the leachates of the pellets 87Sr/86Sr values (although they do plot closer to the leachates values, Figures 4, 5). Every diet item, whether a designed pellet or a plant or animal food in the natural habitat, contains more or less bioavailable mineral components. The main difference between our experiment and a natural habitat is not that the individual sub-components of diet items vary in Sr availability, but rather that in natural habitats, all these different Sr-compounds have similar Sr isotope ratios. This occurs because most sub-components originate from the same habitat, and thus share the same baseline Sr values [though isotopic differences in bioavailable Sr have been observed even under such conditions (Maurer et al., 2012)]. In contrast, in our designed diets, items with variable Sr availability also differ in their individual Sr isotope ratios, because they derive from different ingredients produced in different regions and setting. In other words, whereas in the natural habitat, differences in Sr availability will not affect Sr isotope values, because the different Sr compounds all have similar Sr isotope composition, differences in Sr availability will distinctively affect the Sr isotopic signal in a pelleted diet in which the individual ingredients originate from different regions with different Sr isotope compositions. This becomes obvious by comparing the 87Sr/86Sr from the leachate of the pellets with the completely dissolved pellets (Figures 3–5 and Table 2). The composition of the pellets causes differences in the bioavailability of Sr from the different ingredients of the pellets. Some components are more easily dissolved, while others can only be accessed using stronger acids and/or higher digestion temperatures. Of note, the insect diet showed the same 87Sr/86Sr, regardless of which dissolution process was used, indicating better bioavailability of Sr in this type of pellet [i.e., in its main bioavailable component(s)], in comparison to the plant diet. These yielded much lower 87Sr/86Sr when digested in weaker acids and at lower temperatures, indicating a weak dissolution of the main plant matter and the preferential intake of Sr from a more easily accessible source within the pellet.
For the calculation of the mixing between water and diet (Figure 8, Supplementary Figures 3, 4), we used the known daily food consumption of the rats and guinea pigs, obtained from metabolic cages. Rats consumed 15 – 20 g of pellets per day on average, while guinea pigs consumed 20 – 50 g per day. Water consumption was not monitored during the experiments but was assumed to be 10 mL/day per 100 g body weight (Sharp and Villano, 2012; Suckow et al., 2012). Generally, not all Sr ingested from food and water is absorbed during digestion. Values for Sr (International Commission on Radiological Protection, 1994; Kohn et al., 2013) and Ca absorption (Guéguen and Pointillart, 2000; Rozenberg et al., 2016), which is better documented in comparison to Sr (as a non-essential trace element), but has generally similar chemical behavior, range between 15 and 45%, typically 30%. We therefore used an absorption of 30% for both food and water for the experimental data, independent of the animal model, even though there is reason to assume that guinea pig absorb a higher proportion of dietary Sr than rats, due to the similarity between Sr and Ca in biological systems, and the generally higher Ca digestibility in rabbits or guinea pigs as compared to rats (Cheeke and Amberg, 1973; Hagen et al., 2015). Note that we used the Sr concentration and 87Sr/86Sr ratios as measured in the leachates at pH 1 and 3 for the calculations. Since guinea pigs were roughly twice as heavy as the rats, their water consumption is estimated to be in the range of 50 – 100 mL/day, while we used a range of 10 – 40 mL/day for the rats. The mixing lines using the Sr isotope ratios and concentrations obtained from the completely dissolved pellet did not match the observed 87Sr/86Sr in the incisors, except for the insect pellet and the laser ablation data from the base of the rat incisors (Supplementary Figure 3). However, the leachates of the different pellets agreed much better with the incisor data, but still do not always match the observed 87Sr/86Sr incisor values (Figure 8). Nevertheless, especially for the plant pellet, we can see a much better agreement with the data from the leaching experiment, indicating a lower availability of Sr from the plant material (lucerne). The contribution of water in this feeding group appears to considerably contribute to the Sr budget, as mixing calculations agree well with the tissue values (Figure 8).

Figure 8. Mixing calculations for the three insect, plant, and meat pelleted diets and the experimental tap water for 87Sr/86Sr. We used bioavailable 87Sr/86Sr for the different pellet feeding groups, depending on the leachate values of the pellets leached at pH 1 and 3. For both food and water, a bioavailable component of 30% of the Sr was assumed as intake (International Commission on Radiological Protection, 1994; Guéguen and Pointillart, 2000; Rozenberg et al., 2016). Strontium isotope ratios and concentrations used for the mixing calculations are shown in Table 2. Rat (blue) and guinea pig (red) 87Sr/86Sr ratios for the enamel from laser ablation and solution MC-ICP-MS analyses are shown as dashed and stippled lines, respectively.
For future studies using Sr isotopes as provenance proxy, the careful assessment of 87Sr/86Sr from both the skeletal and dental remains, as well as the underlying geology and vegetation is necessary. We show that bulk food 87Sr/86Sr can largely differ from the bioavailable fraction and thus, may not necessarily reflect the 87Sr/86Sr recorded in the animal hard tissues. This effect should be considered when comparing hard tissue 87Sr/86Sr with local geology and potential Sr isotope isoscapes.
Strontium Isotope Variability Within the Feeding Groups (Experiment-2: Basic Diets)
The time-resolved analyses of the Sr isotope ratios from different time points in the experiment indicated that the full experimental duration of 54 days (and 5 days of acclimatization) was sufficient to reach diet-related 87Sr/86Sr for large parts of the ever-growing rodent incisors (entirely for rats, but not in the apex region for guinea pigs), but not for bone. However, most of the feeding groups display inter-individual 87Sr/86Sr differences among animals fed the same diet, even if only the base part of the incisor is considered (Figures 4, 5). For the rat Basic Diets experiment, differences in 87Sr/86Sr intra-feeding group variability can be observed, even within different parts of the incisor enamel. Typical inter-individual variability ranged from 0.0001 to 0.0005, and this variability tended to decrease toward the younger base of the incisor (Figure 9). This trend is due to the fact that the animals received the experimental diet for longer and thus incorporated diet-related 87Sr/86Sr for longer, whereas in the (ontogenetically oldest) apex area, high 87Sr/86Sr spikes inherited from the supplier feeding period produce more scatter in enamel 87Sr/86Sr. The main reason for this pattern is the continuous exchange between immature enamel and the bioavailable Sr pools in the body, i.e. the diet and the resorbed bone fraction (Johnson et al., 1968; Jowsey et al., 1971; Neiman and Eisenmann, 1975; Boyde and Reith, 1981; Montgomery et al., 2010). Bone (i.e., the skeleton) usually contains up to 99% of all body Sr (Schroeder et al., 1972; Sugihira and Suzuki, 1991). Thus, an important contribution of Sr in the blood originates from “old” Sr pool (incorporated into the bone at the supplier facility where animals were raised and then released from the bone during remodeling) that is incorporated in incisor enamel during the maturation process (Leblond and Warshawsky, 1979), which thus has a higher 87Sr/86Sr than the 87Sr/86Sr ingested during experimental conditions. While the bone 87Sr/86Sr slowly shifts toward the incisor value, the influence of the Sr fraction resorbed from the bones on the 87Sr/86Sr of new forming (hard)tissue is reduced. The same trend was also visible in the guinea pigs. The apex of the guinea pig incisors likely does not completely reflect the experimental diet, enhancing the inter-, as well as the intra-individual (i.e., intra-incisor) variability. For the guinea pigs fed the plant diet, the inter-individual 87Sr/86Sr variability was as high as 0.001 for the apex part and therefore an order of magnitude higher than within most other guinea pig groups, but also two to 10 times higher than for the rats (Figure 9). This variability is rather high for a population of small animals of the same species, sex, age, and feeding on the same diet and water sources. Typically, a 87Sr/86Sr cut-off value of 0.001 between bone and enamel of an individual is used in archaeological studies to differentiate between local and non-local individuals within one human population (e.g., Grupe et al., 1997; Price et al., 2004; Knudson et al., 2016). Other controlled feeding experiments, however, detected even higher inter-individual Sr isotope ranges of up to 0.002 for pigs raised on local derived diets (Anders et al., 2019). We observe a much lower variability in our study, and the high guinea pig variability is likely related to the influence of bone-resorption derived Sr. In addition, the sampling strategy using LA-MC-ICP-MS is more prone to a higher 87Sr/86Sr variability in comparison to solution-based analyses, due to the potential influence of interferences during data acquisition.
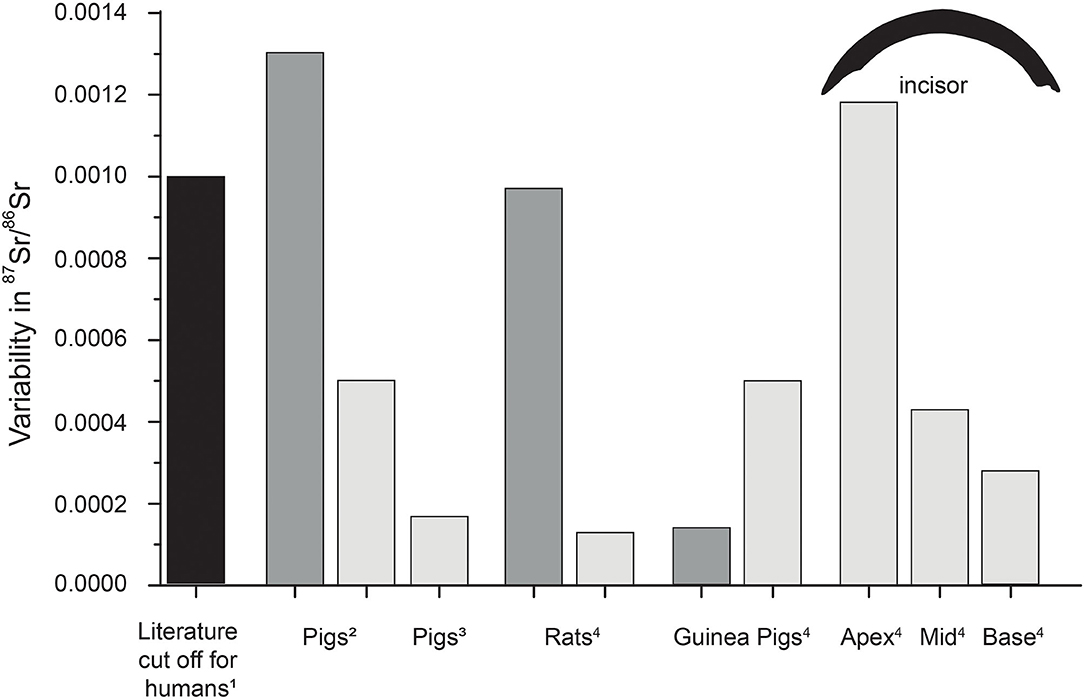
Figure 9. Typical literature 87Sr/86Sr cut-off value for humans (1, e.g., Grupe et al., 1997; Price et al., 2004; Burton and Price, 2013; Knudson et al., 2016), as well as intra-population variability of 87Sr/86Sr in bones (dark gray) and teeth (light gray) of pigs (Lewis et al., 2017; Anders et al., 2019) and rats and guinea pigs (4, this study). Incisor variability reflects the largest variability observed in the respective part for both animal groups. For humans often a difference of 0.001 in 87Sr/86Sr between bones and tooth enamel is used to distinguish between local and non-local individuals.
Besides the inter-individual variability, another important aspect for future Sr isotope studies is the observed intra-individual variability. Depending on the type of tooth or bone, differences in 87Sr/86Sr might be detected, due to differences in bone turnover rates reflecting different time averaging of isotopically distinct intake sources (Gregoricka, 2014; Knudson et al., 2016; Anders et al., 2019). Here only one type of bone (distal tibia) was investigated, and we did not compare the incisor enamel 87Sr/86Sr with those of the molar enamel of rats and/or guinea pigs. However, since molars of rats are not ever-growing, their 87Sr/86Sr will likely reflect the Sr ingested during their time at the supplier facility and therefore differ from our pelleted experiment diet. This is evident in the rat incisor 87Sr/86Sr of individuals that were euthanized prior to entering the feeding experiment (Figure 2). It highlights the importance of the adequate choice of teeth for Sr isotope analysis and the knowledge of tooth formation and maturation times to constrain accurately diet-related 87Sr/86Sr and hence modern or past mobility patterns. Interestingly, the intra-incisor variability rarely exceeds 0.0001, indicating a much lower intra-individual variability than suggested by Anders et al. (2019), who found a 87Sr/86Sr variability of 0.0005 for tooth and 0.0013 for bone samples (Figure 9).
The overall observed Sr isotope variability of both rodent species is small and does not exceed the cut off values typically employed in 87Sr/86Sr in provenance studies. Some incisor Sr isotope values from the apex (oldest part), especially for the guinea pigs, showed a higher Sr isotope variability than the other, ontogenetically younger parts of the incisor (Figure 9). This is an artifact of the feeding experiment, where the oldest part of the guinea pig incisors is still influenced by the supplier diet and thus, does not reflect a natural feeding behavior where an individual feeds only from local and similar food sources. Since the rat incisors grow faster than the guinea pig incisors (0.5 mm/day vs. 0.1 – 0.3 mm/day), even their oldest part (apex) mainly reflects the experimental diet and is suited for Sr isotope variability estimation of animals feeding constantly on the same diet.
Influence of Water (Experiment-2: Basic Diets) and External Dust Sources on Tissue Sr Isotope Composition (Experiment-3: Dust Addition)
The rats that received the plant diet and Sr-enriched mineral water showed significantly lower 87Sr/86Sr (0.0001) than the group receiving the same pelleted diet but the less Sr-rich Zurich tap water for bone (p < 0.01), as well as for all parts of the incisors (p < 0.02). This shows that the high Sr concentration of the Ca- and Sr-enriched mineral water from a limestone aquifer had a significant influence on the incisor 87Sr/86Sr. While the Sr concentration in natural freshwater water sources is often below 1 mg/L (Voerkelius et al., 2010), this Sr-enriched water had a Sr concentration of 8.6 mg/L and thus significantly influenced the 87Sr/86Sr of the hard tissues.
Although silicate minerals are usually not digested (Van Soest and Jones, 1968) and hence elements derived from them are not bioavailable, the small particle size and intra-layer bound cations (of the clay mineral kaolinite) bear the potential for some Sr uptake from the kaolin and the silt-sized carbonate fraction of loess, which makes up 20% of the 4 wt%. However, due to the relatively short experimental duration of up to 29 days, only the base of the incisor yielded the diet-related (i.e., potentially mineral dust-influenced) values (Figure 1). A significant difference, exceeding the analytical uncertainty between the animals feeding on the kaolin and the loess pellets was observed in the base part (Figure 6, p < 0.01) with the former having slightly higher values than the latter and also clearly different from the bulk diet values. While the bulk pellets yielded 87Sr/86Sr >0.710, all 87Sr/86Sr obtained from the enamel of the base area were <0.7093. Again, the Sr bioavailability of the different ingredients of the diets differ from the completely dissolved pellet. This is especially crucial for the silicate minerals, which are non-digestible. As described for the other pelleted diets, the leachates better reflect the bioavailable Sr fraction, especially those at pH 1. Furthermore, the pellet into which the mineral dust was admixed consisted of lucerne, from which Sr was shown to be less bioavailable. Nevertheless, enamel 87Sr/86Sr differences among incisors of the three feeding groups (mineral-dust-free, loess-containing, and kaolin-containing pellets) are visible.
Conclusion
Our controlled feeding experiment offered a unique possibility to evaluate the timing of Sr incorporation in tooth enamel, as well as the influence of diet-derived Sr (including water and mineral dust) on the 87Sr/86Sr of hard tissues (i.e., bone, enamel) of rodents. Intra-population variability in 87Sr/86Sr was lower (<0.001) than previously reported for other mammals, potentially due to the high degree of experimental control. Therefore, our data provide a minimum estimate of the 87Sr/86Sr variability within a (small mammal) population, and highlight the importance of proper sampling strategies in building local isotopic baselines using rodents. We also determined that the inter-individual variability is generally low and thus likely does not affect provenance studies based on radiogenic Sr isotopes. Mixing model calculations further support the importance of differential bioavailability of Sr in the diet (including both food and water). In addition, we observed differences in isotopic and elemental variability between bone and incisor enamel. While the bone showed slow and incomplete turnover over the course of the experiment, the incisors exhibited much higher variability. We demonstrated that a complete turnover of the rat incisors occurred after 54 days, while the guinea pig incisors still retained remnant isotopic signals from supplier food in the oldest part of their incisors. This indicates that we can estimate a minimum period of roughly 2 months, before the new, diet-related 87Sr/86Sr is recorded in enamel throughout the full length of the incisor. This information must be taken into consideration in human provenance studies which frequently use rodents' teeth to characterize locally bioavailable Sr isotope ranges. In addition, the leachates of the pelleted diets clearly demonstrate the effect of differences in Sr bioavailability on 87Sr/86Sr, and should also be considered in Sr isoscapes studies. Furthermore, we observed an offset in 87Sr/86Sr between rats and guinea pigs feeding on the exact same diet, indicating taxon-specific absorption coefficients for Sr, especially from the drinking water.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The animal study was reviewed and approved by Cantonal Veterinary Office Zurich, Switzerland (license no. ZH135/16).
Author Contributions
MW, TTü, and MC designed the study. DW and MC performed the animal experiments. MW performed the solution-based MC-ICP-MS analyses, evaluated the data, performed the analyses, and wrote the manuscript. MW and RMK developed the LA-MC-ICP-MS methods. KW performed the LA-ICP-MS measurements and evaluated the in-situ Sr concentration data. TTü performed the ICP-OES measurements. MW, KW, JL, and DW prepared the samples prior to analysis. TTa did the mixing calculations. TTü, MC, TTa, FL, and KW improved the manuscript. MW, TTü, TTa, FL, and MC designed the leaching experiment. JL proof-read the English language. All authors discussed the results and commented on the manuscript.
Funding
This project received funding from the European Research Council (ERC) under the European Union's Horizon 2020 research and innovation programme, grant agreement No. 681450.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank D. Codron for assistance during the setup of the study, M. Großkopf and J. Klose for their assistance in the laboratory and L. Martin, N. Schmid, K. Zbinden, D. Codron, and S. Heldstab for taking care of the animals during the experiments. The help of the student assistants in Mainz is greatly acknowledged. We also thank D. Scholz for the access to the laboratories at the Institute of Geosciences, University of Mainz. The comments and suggestions of three independent reviewers and the Editor Brooke E. Crowley helped to improve the study and are greatly acknowledged.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.569940/full#supplementary-material
References
Anders, D., Osmanovic, A., and Vohberger, M. (2019). Intra-and inter-individual variability of stable strontium isotope ratios in hard and soft body tissues of pigs. Rapid Commun. Mass Spectrometry 33, 281–290. doi: 10.1002/rcm.8350
Balasse, M., Ambrose, S. H., Smith, A. B., and Price, T. D. (2002). The seasonal mobility model for prehistoric herders in the south-western cape of south africa assessed by isotopic analysis of sheep tooth enamel. J. Archaeol. Sci. 29, 917–932. doi: 10.1006/jasc.2001.0787
Balter, V., Bocherens, H., Person, A., Labourdette, N., Renard, M., and Vandermeersch, B. (2002). Ecological and physiological variability of Sr/Ca and Ba/Ca in mammals of West European mid-Würmian food webs. Palaeogeogr. Palaeoclimatol. Palaeoecol. 186, 127–143. doi: 10.1016/S0031-0182(02)00448-0
Banner, J. L. (2004). Radiogenic isotopes: systematics and applications to earth surface processes and chemical stratigraphy. Earth Sci. Rev. 65, 141–194. doi: 10.1016/S0012-8252(03)00086-2
Bataille, C. P., Crowley, B. E., Wooller, M. J., and Bowen, G. J. (2020). Advances in global bioavailable strontium isoscapes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 555:109849. doi: 10.1016/j.palaeo.2020.109849
Bentley, R. A. (2006). Strontium isotopes from the earth to the archaeological skeleton: a review. J. Archaeol. Method Theory 13, 135–187. doi: 10.1007/s10816-006-9009-x
Blum, J. D., Taliaferro, E. H., Weisse, M. T., and Holmes, R. T. (2000). Changes in Sr/Ca, Ba/Ca and 87Sr/86Sr ratios between trophic levels in two forest ecosystems in the northeastern USA. Biogeochemistry 49, 87–101. doi: 10.1023/A:1006390707989
Boyde, A., and Reith, E. J. (1981). Display of maturation cycles in rat incisor enamel with tetracycline labelling. Histochemistry 72, 551–561. doi: 10.1007/BF00493275
Bryant, F. J., and Loutit, J. F. (1964). The entry of strontium-90 into human bone. Proc. R. Soc. Lond. Series B Biol. Sci. 159, 449–465. doi: 10.1098/rspb.1964.0013
Burton, J. H., and Price, T. D. (2013). “Seeking the local 87Sr/86Sr ratio to determine geographic origins of humans,” in Archaeological chemistry VIII, eds R. A. Armitage and J. H. Burton (Washington, DC: American Chemical Society), 309–320.
Burton, J. H., Price, T. D., and Middleton, W. D. (1999). Correlation of bone Ba/Ca and Sr/Ca due to biological purification of calcium. J. Archaeol. Sci 26, 609–616. doi: 10.1006/jasc.1998.0378
Burton, J. H., and Wright, L. E. (1995). Nonlinearity in the relationship between bone Sr/Ca and diet: paleodietary implications. Am. J. Phys. Anthropol. 96, 273–282. doi: 10.1002/ajpa.1330960305
Capo, R. C., Stewart, B. W., and Chadwick, O. A. (1998). Strontium isotopes as tracers of ecosystem processes: theory and methods. Geoderma 82, 197–225. doi: 10.1016/S0016-7061(97)00102-X
Carr, T. E. F., Harrison, G. E., Loutit, J. F., and Sutton, A. (1962). Movement of strontium in the human body. Br. Med. J. 2, 773–775. doi: 10.1136/bmj.2.5307.773
Cheeke, P. R., and Amberg, J. W. (1973). Comparative calcium excretion by rats and rabbits. J. Anim. Sci 37, 450–454. doi: 10.2527/jas1973.372450x
Comar, C. L., Russell, R. S., and Wasserman, R. H. (1957). Strontium-calcium movement from soil to man. Science 126, 485–492. doi: 10.1126/science.126.3272.485
Copeland, S. R., Sponheimer, M., De Ruiter, D. J., Lee-Thorp, J. A., Codron, D., Le Roux, P. J., et al. (2011). Strontium isotope evidence for landscape use by early hominins. Nature 474, 76–78. doi: 10.1038/nature10149
Copeland, S. R., Sponheimer, M., Le Roux, P. J., Grimes, V., Lee-Thorp, J. A., De Ruiter, D. J., et al. (2008). Strontium isotope ratios (87Sr/86Sr) of tooth enamel: a comparison of solution and laser ablation multicollector inductively coupled plasma mass spectrometry methods. Rapid Commun. Mass Spectrom. 22, 3187–3194. doi: 10.1002/rcm.3717
Copeland, S. R., Sponheimer, M., Lee-Thorp, J. A., Le Roux, P. J., De Ruiter, D. J., and Richards, M. P. (2010). Strontium isotope ratios in fossil teeth from South Africa: assessing laser ablation MC-ICP-MS analysis and the extent of diagenesis. J. Archaeol. Sci. 37, 1437–1446. doi: 10.1016/j.jas.2010.01.003
Dahl, S. G., Allain, P., Marie, P. J., Mauras, Y., Boivin, G., Ammann, P., et al. (2001). Incorporation and distribution of strontium in bone. Bone 28, 446–453. doi: 10.1016/S8756-3282(01)00419-7
Eisenhauer, A., Müller, M., Heuser, A., Kolevica, A., Glüer, C. C., Both, M., et al. (2019). Calcium isotope ratios in blood and urine: a new biomarker for the diagnosis of osteoporosis. Bone Rep. 10:100200. doi: 10.1016/j.bonr.2019.100200
Elias, R. W., Hirao, Y., and Patterson, C. C. (1982). The circumvention of the natural biopurification of calcium along nutrient pathways by atmospheric inputs of industrial lead. Geochim. Cosmochim. Acta 46, 2561–2580. doi: 10.1016/0016-7037(82)90378-7
Faure, G., and Mensing, T. M. (2005). Isotopes: Principles and Applications. (Hoboken: John Wiley & Sons Inc), 928.
Flockhart, D. T. T., Kyser, T. K., Chipley, D., Miller, N. G., and Norris, D. R. (2015). Experimental evidence shows no fractionation of strontium isotopes (87Sr/86Sr) among soil, plants, and herbivores: implications for tracking wildlife and forensic science. Isotopes Environ. Health Stud. 51, 372–381. doi: 10.1080/10256016.2015.1021345
Galler, P., Limbeck, A., Boulyga, S. F., Stingeder, G., Hirata, T., and Prohaska, T. (2007). Development of an on-line flow injection Sr/matrix separation method for accurate, high-throughput determination of Sr isotope ratios by multiple collector-inductively coupled plasma-mass spectrometry. Anal. Chem. 79, 5023–5029. doi: 10.1021/ac070307h
Gehler, A., Tütken, T., and Pack, A. (2012). Oxygen and carbon isotope variations in a modern rodent community–implications for palaeoenvironmental reconstructions. PLoS ONE 7:e49531. doi: 10.1371/journal.pone.0049531
Gregoricka, L. A. (2014). Assessing life history from commingled assemblages: the biogeochemistry of inter-tooth variability in Bronze Age Arabia. J. Archaeol. Sci. 47, 10–21. doi: 10.1016/j.jas.2014.04.004
Grupe, G., Price, T. D., Schröter, P., Söllner, F., Johnson, C. M., and Beard, B. L. (1997). Mobility of Bell Beaker people revealed by strontium isotope ratios of tooth and bone: a study of southern Bavarian skeletal remains. Appl. Geochem. 12, 517–525. doi: 10.1016/S0883-2927(97)00030-9
Guéguen, L., and Pointillart, A. (2000). The bioavailability of dietary calcium. J. Am. College Nutr. 19, 119S−136S. doi: 10.1080/07315724.2000.10718083
Hagen, K. B., Tschudin, A., Liesegang, A., Hatt, J. M., and Clauss, M. (2015). Organic matter and macromineral digestibility in domestic rabbits (Oryctolagus cuniculus) as compared to other hindgut fermenters. J. Anim. Physiol. Anim. Nutr. 99, 1197–1209. doi: 10.1111/jpn.12323
Horstwood, M. S. A., Evans, J. A., and Montgomery, J. (2008). Determination of Sr isotopes in calcium phosphates using laser ablation inductively coupled plasma mass spectrometry and their application to archaeological tooth enamel. Geochim. Cosmochim. Acta 72, 5659–5674. doi: 10.1016/j.gca.2008.08.016
Inoue, M., Nohara, M., Okai, T., Suzuki, A., and Kawahata, H. (2004). Concentrations of trace elements in carbonate reference materials coral JCp-1 and giant clam JCt-1 by inductively coupled plasma-mass spectrometry. Geostand. Geoanalytical. Res. 28, 411–416. doi: 10.1111/j.1751-908X.2004.tb00759.x
International Commission on Radiological Protection, C. (1994). ICRP Publication 67: Age-dependent Doses to Members of the Public from Intake of Radionuclides: Part 2 Ingestion Dose Coefficients (Oxford: Elsevier Health Sciences).
Jochum, K. P., Stoll, B., Herwig, K., and Willbold, M. (2007). Validation of LA-ICP-MS trace element analysis of geological glasses using a new solid-state 193 nm Nd : YAG laser and matrix-matched calibration. J. Anal. At. Spectrom. 22, 112–121. doi: 10.1039/B609547J
Johnson, A. R., Armstrong, W. D., and Singer, L. (1968). The incorporation and removal of large amounts of strontium by physiologic mechanisms in mineralized tissues of the rat. Calcif. Tissue Res 2, 242–252. doi: 10.1007/BF02279212
Jowsey, J., Gordon, G., and Bourne, G. H. (1971). Bone turnover and osteoporosis. Biochem. Physiol. Bone 3, 201–238. doi: 10.1016/B978-0-12-119203-7.50013-1
Kelly, S., Heaton, K., and Hoogewerff, J. (2005). Tracing the geographical origin of food: The application of multi-element and multi-isotope analysis. Trends Food Sci. Technol. 16, 555–567. doi: 10.1016/j.tifs.2005.08.008
Kimura, J. I., and Chang, Q. (2012). Origin of the suppressed matrix effect for improved analytical performance in determination of major and trace elements in anhydrous silicate samples using 200 nm femtosecond laser ablation sector-field inductively coupled plasma mass spectrometry. J. Anal. At. Spectrom. 27, 1549–1559. doi: 10.1039/c2ja10344c
Kirsanow, K., and Tuross, N. (2011). Oxygen and hydrogen isotopes in rodent tissues: Impact of diet, water and ontogeny. Palaeogeogr. Palaeoclimatol. Palaeoecol. 310, 9–16. doi: 10.1016/j.palaeo.2011.03.022
Knipper, C. (2004). Die Strontiumisotopenanalyse: eine naturwissenschaftliche Methode zur Erfassung von Mobilität in der Ur-und Frühgeschichte. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz 51, 589–686. doi: 10.11588/jrgzm.2004.2.21081
Knudson, K. J., Stanish, C., Cerna, M. C. L., Faull, K. F., and Tantaleán, H. (2016). Intra-individual variability and strontium isotope measurements: a methodological study using 87Sr/86Sr data from Pampa de los Gentiles, Chincha Valley, Peru. J. Archaeol. Sci. Rep. 5, 590–597. doi: 10.1016/j.jasrep.2016.01.016
Knudson, K. J., Williams, H. M., Buikstra, J. E., Tomczak, P. D., Gordon, G. W., and Anbar, A. D. (2010). Introducing δ88/86Sr analysis in archaeology: a demonstration of the utility of strontium isotope fractionation in paleodietary studies. J. Archaeol. Sci. 37, 2352–2364. doi: 10.1016/j.jas.2010.04.009
Kohn, M. J., Morris, J., and Olin, P. (2013). Trace element concentrations in teeth–a modern Idaho baseline with implications for archeometry, forensics, and palaeontology. J. Archaeol. Sci. 40, 1689–1699. doi: 10.1016/j.jas.2012.11.012
Law, K.-T., Lee, C.-K., King, N. M., and Rabie, A.-B. M. (2003). The relationship between eruption and length of mandibular incisors in young rats. Med. Sci. Monitor 9, BR47–BR53.
Leblond, C. P., and Warshawsky, H. (1979). Dynamics of enamel formation in the rat incisor tooth. J. Dent. Res. 58, 950–979. doi: 10.1177/00220345790580024901
Lewis, J., Coath, C. D., and Pike, A. W. G. (2014). An improved protocol for 87Sr/86Sr by laser ablation multi-collector inductively coupled plasma mass spectrometry using oxide reduction and a customised plasma interface. Chem. Geol. 390, 173–181. doi: 10.1016/j.chemgeo.2014.10.021
Lewis, J., Pike, A. W. G., Coath, C. D., and Evershed, R. P. (2017). Strontium concentration, radiogenic (87Sr/86Sr) and stable (δ88Sr) strontium isotope systematics in a controlled feeding study. STAR: Sci. Technol. Archaeol. Res. 3, 53–65. doi: 10.1080/20548923.2017.1303124
Lugli, F., Cipriani, A., Arnaud, J., Arzarello, M., Peretto, C., and Benazzi, S. (2017a). Suspected limited mobility of a Middle Pleistocene woman from Southern Italy: strontium isotopes of a human deciduous tooth. Sci. Rep. 7:8615. doi: 10.1038/s41598-017-09007-5
Lugli, F., Cipriani, A., Peretto, C., Mazzucchelli, M., and Brunelli, D. (2017b). In situ high spatial resolution 87Sr/86Sr ratio determination of two Middle Pleistocene (c.a. 580 ka) Stephanorhinus hundsheimensis teeth by LA–MC–ICP–MS. Int. J. Mass Spectrometry 412, 38–48. doi: 10.1016/j.ijms.2016.12.012
Marchionni, S., Buccianti, A., Bollati, A., Braschi, E., Cifelli, F., Molin, P., et al. (2016). Conservation of 87Sr/86Sr isotopic ratios during the winemaking processes of ‘Red’wines to validate their use as geographic tracer. Food Chem. 190, 777–785. doi: 10.1016/j.foodchem.2015.06.026
Maurer, A.-F., Galer, S. J. G., Knipper, C., Beierlein, L., Nunn, E. V., Peters, D., et al. (2012). Bioavailable 87Sr/86Sr in different environmental samples—effects of anthropogenic contamination and implications for isoscapes in past migration studies. Sci. Total Environ. 433, 216–229. doi: 10.1016/j.scitotenv.2012.06.046
McArthur, J. M., Howarth, R. J., and Bailey, T. R. (2001). Strontium Isotope Stratigraphy: LOWESS version 3: best fit to the marine Sr-isotope curve for 0–509 Ma and accompanying look-up table for deriving numerical age. J. Geol. 109, 155–170. doi: 10.1086/319243
McArthur, J. M., Howarth, R. J., and Shields, G. A. (2012). Strontium isotope stratigraphy. Geol. Time Scale 1, 127–144. doi: 10.1016/B978-0-444-59425-9.00007-X
Mcconnell, E. L., Basit, A. W., and Murdan, S. (2008). Measurements of rat and mouse gastrointestinal pH, fluid and lymphoid tissue, and implications for in-vivo experiments. J. Pharm. Pharmacol. 60, 63–70. doi: 10.1211/jpp.60.1.0008
Meija, J., Coplen, T. B., Berglund, M., Brand, W. A., De Bièvre, P., Gröning, M., et al. (2016). Isotopic compositions of the elements 2013 (IUPAC technical Report). Pure Appl. Chem. 88, 293–306. doi: 10.1515/pac-2015-0503
Merchant, H. A., Mcconnell, E. L., Liu, F., Ramaswamy, C., Kulkarni, R. P., Basit, A. W., et al. (2011). Assessment of gastrointestinal pH, fluid and lymphoid tissue in the guinea pig, rabbit and pig, and implications for their use in drug development. Eur. J. Pharm. Sci. 42, 3–10. doi: 10.1016/j.ejps.2010.09.019
Mischel, S. A., Mertz-Kraus, R., Jochum Klaus, P., and Scholz, D. (2017). TERMITE: An R script for fast reduction of laser ablation inductively coupled plasma mass spectrometry data and its application to trace element measurements. Rapid Commun. Mass Spectrom. 31, 1079–1087. doi: 10.1002/rcm.7895
Montgomery, J., Evans, J. A., and Horstwood, M. S. A. (2010). Evidence for long-term averaging of strontium in bovine enamel using TIMS and LA-MC-ICP-MS strontium isotope intra-molar profiles. Environ. Archaeol. 15, 32–42. doi: 10.1179/146141010X12640787648694
Müller, J., Clauss, M., Codron, D., Schulz, E., Hummel, J., Kircher, P., et al. (2015). Tooth length and incisal wear and growth in guinea pigs (Cavia porcellus) fed diets of different abrasiveness. J. Anim. Physiol. Anim. Nutr. 99, 591–604. doi: 10.1111/jpn.12226
Müller, W., and Anczkiewicz, R. (2016). Accuracy of laser-ablation (LA)-MC-ICPMS Sr isotope analysis of (bio) apatite–a problem reassessed. J. Anal. At. Spectrom. 31, 259–269. doi: 10.1039/C5JA00311C
Müller, W., Fricke, H., Halliday, A. N., Mcculloch, M. T., and Wartho, J.-A. (2003). Origin and migration of the alpine iceman. Science 302, 862–866. doi: 10.1126/science.1089837
Neiman, A., and Eisenmann, D. R. (1975). The effect of strontium, cobalt and fluoride on rat incisor enamel formation. Anat. Rec. 183, 303–321. doi: 10.1002/ar.1091830207
Ohno, T., and Hirata, T. (2007). Simultaneous determination of mass-dependent isotopic fractionation and radiogenic isotope variation of strontium in geochemical samples by multiple collector-ICP-mass spectrometry. Anal. Sci. 23, 1275–1280. doi: 10.2116/analsci.23.1275
Okai, T., Suzuki, A., Kawahata, H., Terashima, S., and Imai, N. (2002). Preparation of a new geological survey of japan geochemical reference material: coral JCp-1. Geostand. Geoanalytical. Res. 26, 95–99. doi: 10.1111/j.1751-908X.2002.tb00627.x
Outridge, P., Chenery, S., Babaluk, J., and Reist, J. (2002). Analysis of geological Sr isotope markers in fish otoliths with subannual resolution using laser ablation-multicollector-ICP-mass spectrometry. Environ. Geol. 42, 891–899. doi: 10.1007/s00254-002-0596-x
Passey, B. H., and Cerling, T. E. (2002). Tooth enamel mineralization in ungulates: implications for recovering a primary isotopic time-series. Geochim. Cosmochim. Acta 66, 3225–3234. doi: 10.1016/S0016-7037(02)00933-X
Price, T. D., Knipper, C., Grupe, G., and Smrcka, V. (2004). Strontium isotopes and prehistoric human migration: the Bell Beaker period in central Europe. Eur. J. Archaeol. 7, 9–40. doi: 10.1177/1461957104047992
Romaniello, S. J., Field, M. P., Smith, H. B., Gordon, G. W., Kim, M. H., and Anbar, A. D. (2015). Fully automated chromatographic purification of Sr and Ca for isotopic analysis. J. Anal. At. Spectrom. 30, 1906–1912. doi: 10.1039/C5JA00205B
Rozenberg, S., Body, J.-J., Bruyere, O., Bergmann, P., Brandi, M. L., Cooper, C., et al. (2016). Effects of dairy products consumption on health: benefits and beliefs—a commentary from the belgian bone club and the european society for clinical and economic aspects of osteoporosis, osteoarthritis and musculoskeletal diseases. Calcif. Tissue Int. 98, 1–17. doi: 10.1007/s00223-015-0062-x
Scheeres, M., Knipper, C., Hauschild, M., Schönfelder, M., Siebel, W., Vitali, D., et al. (2013). Evidence for “Celtic migrations”? strontium isotope analysis at the early La Tène (LT B) cemeteries of Nebringen (Germany) and Monte Bibele (Italy). J. Archaeol. Sci. 40, 3614–3625. doi: 10.1016/j.jas.2013.05.003
Schroeder, H. A., Tipton, I. H., and Nason, A. P. (1972). Trace metals in man: strontium and barium. J. Chronic Dis. 25, 491–517. doi: 10.1016/0021-9681(72)90150-6
Schweissing, M. M., and Grupe, G. (2003). Stable strontium isotopes in human teeth and bone: a key to migration events of the late Roman period in Bavaria. J. Archaeol. Sci. 30, 1373–1383. doi: 10.1016/S0305-4403(03)00025-6
Sharp, P., and Villano, J. S. (2012). The Laboratory Rat. (Boca Raton: CRC press), 393. doi: 10.1201/b13862
Simonetti, A., Buzon, M. R., and Creaser, R. A. (2008). In-situ elemental and Sr isotope investigation of human tooth enamel by Laser Ablation-(MC-)ICP-MS: Successes and pitfalls. Archaeometry 50, 371–385. doi: 10.1111/j.1475-4754.2007.00351.x
Slovak, N. M., and Paytan, A. (2012). “Applications of Sr isotopes in archaeology,” in Handbook of Environmental Isotope Geochemistry, ed M. Baskaran (Berlin: Springer), 743–768. doi: 10.1007/978-3-642-10637-8_35
Suckow, M. A., Stevens, K. A., and Wilson, R. P. (2012). The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents. (San Diego: Academic Press), 1288.
Sugihira, N., and Suzuki, K. T. (1991). Discrimination between strontium and calcium in suckling rats. Biol. Trace Elem. Res. 29, 1–10. doi: 10.1007/BF03032669
R Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/
Tütken, T. (2010). Die Isotopenanalyse fossiler Skelettreste – Bestimmung der Herkunft und Mobilität von Menschen und Tieren. Anthropologie, Isotopie und DNA -biografische Annäherung an namenlose vorgeschichtliche Skelette. Tagungsband 2, 33–51.
Urey, H. C. (1947). The thermodynamic properties of isotopic substances. J. Chem. Soc. 562–581. doi: 10.1039/jr9470000562
Van Soest, P. J., and Jones, L. H. P. (1968). Effect of silica in forages upon digestibility. J. Dairy Sci. 51, 1644–1648. doi: 10.3168/jds.S0022-0302(68)87246-7
Voerkelius, S., Lorenz, G. D., Rummel, S., Quétel, C. R., Heiss, G., Baxter, M., et al. (2010). Strontium isotopic signatures of natural mineral waters, the reference to a simple geological map and its potential for authentication of food. Food Chem. 118, 933–940. doi: 10.1016/j.foodchem.2009.04.125
Weber, M., Lugli, F., Hattendorf, B., Scholz, D., Mertz-Kraus, R., Guinoiseau, D., et al. (2020). NanoSr–a new carbonate microanalytical reference material for in situ strontium isotope analysis. Geostand. Geoanalytical. Res. 44, 69–83. doi: 10.1111/ggr.12296
Weber, M., Lugli, F., Jochum, K. P., Cipriani, A., and Scholz, D. (2018a). Calcium carbonate and phosphate reference materials for monitoring bulk and microanalytical determination of Sr isotopes. Geostand. Geoanalytical. Res. 42, 77–89. doi: 10.1111/ggr.12191
Weber, M., Scholz, D., Schröder-Ritzrau, A., Deininger, M., Spötl, C., Lugli, F., et al. (2018b). Evidence of warm and humid interstadials in central Europe during early MIS 3 revealed by a multi-proxy speleothem record. Quat. Sci. Rev. 200, 276–286. doi: 10.1016/j.quascirev.2018.09.045
Weber, M., Wassenburg, J. A., Jochum, K. P., Breitenbach, S. F. M., Oster, J., and Scholz, D. (2017). Sr-isotope analysis of speleothems by LA-MC-ICP-MS: High temporal resolution and fast data acquisition. Chem. Geol. 468, 63–74. doi: 10.1016/j.chemgeo.2017.08.012
Willmes, M., Kinsley, L., Moncel, M. H., Armstrong, R. A., Aubert, M., Eggins, S., et al. (2016). Improvement of laser ablation in situ micro-analysis to identify diagenetic alteration and measure strontium isotope ratios in fossil human teeth. J. Archaeol. Sci. 70, 102–116. doi: 10.1016/j.jas.2016.04.017
Winkler, D. E., Tütken, T., Schulz-Kornas, E., Kaiser, T. M., Müller, J., Leichliter, J., et al. (2020). Shape, size, and quantity of ingested external abrasives influence dental microwear texture formation in guinea pigs. Proc. Natl. Acad. Sci. U.S.A. 117, 22264–22273. doi: 10.1073/pnas.2008149117
Keywords: strontium isotopes, rodents, population variability, enamel, bioavailability, bone, feeding study, faeces
Citation: Weber M, Tacail T, Lugli F, Clauss M, Weber K, Leichliter J, Winkler DE, Mertz-Kraus R and Tütken T (2020) Strontium Uptake and Intra-Population 87Sr/86Sr Variability of Bones and Teeth—Controlled Feeding Experiments With Rodents (Rattus norvegicus, Cavia porcellus). Front. Ecol. Evol. 8:569940. doi: 10.3389/fevo.2020.569940
Received: 05 June 2020; Accepted: 12 November 2020;
Published: 07 December 2020.
Edited by:
Brooke Crowley, University of Cincinnati, United StatesReviewed by:
Anneke Janzen, The University of Tennessee, Knoxville, United StatesRobert S. Feranec, New York State Museum, United States
Rachel Reid, Virginia Tech, United States
Copyright © 2020 Weber, Tacail, Lugli, Clauss, Weber, Leichliter, Winkler, Mertz-Kraus and Tütken. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Weber, bWljaGFlbC53ZWJlckB1bmktbWFpbnouZGU=