 Giorgos Maneas
Giorgos Maneas Dimitris Bousbouras
Dimitris Bousbouras Viggo Norrby
Viggo Norrby Håkan Berg
Håkan Berg
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 22 December 2020
Sec. Conservation and Restoration Ecology
Volume 8 - 2020 | https://doi.org/10.3389/fevo.2020.501548
This article is part of the Research Topic Ecosystem Services, Biodiversity and Water Quality in Transitional Ecosystems View all 11 articles
Located at the south-western most part of the Balkan peninsula, along an important migration route (the Mediterranean/Black Sea Flyway), the Gialova Lagoon wetland is one of the few remaining Important Bird Areas (IBAs) along the south-west coast of Greece, also designated as a Special Protection Area. The wetland serves as the first suitable stopover for many spring migrants who have flown non-stop over the Mediterranean Sea, and the last before their journey back to Africa in the autumn. In this study, we conducted monthly field visits during the period October 2016 to January 2019 with the aim to complement existing information about the site, to evaluate the current status and distribution of waterbirds, to provide insights for the management of the area and to re-assess the IBA/Ramsar criteria. A total of 149 bird species representing 43 families and 15 orders were recorded, including 36 threatened species at an International, European or/and national level, and 40 species listed in the Annex I of the EUs Birds Directive (21 species were listed as both threatened and under Annex I). 81 species were identified as wetland related species, of which 66 species were identified as waterbirds (7 orders, 11 families). Waterbirds richness and abundance were higher during the Wet season and corresponding periods (Wintering and Spring migration). All parts of the wetland supported waterbirds and threatened species, with the S. Wetland sub-area being the most diverse during the Breeding/Nesting, and both migration periods. The abundance of most waterbirds and IBA species have declined over the last 20 years, but this does not necessarily mean that the area no longer fulfills Ramsar criterion 6 (and equivalent IBA criterion A4i). However, this outcome should not be overlooked by the site managers and conservation actions, such as the restoration of fresh water inflows which could improve habitats and water conditions for IUCN and IBA species, should be implemented with high priority. In addition, our results indicate that the area meets Ramsar criterion 4 and criterion 2, and thus we suggest that it should be further investigated and evaluated to potentially become the eleventh Greek Ramsar site.
Waterbirds, are defined as “species of bird that are ecologically dependent on wetlands” (Ramsar convention, 1994, art. 1.2) including all waterfowl, seabirds and waders. Wetlands are used both as wintering areas—to stay in for longer periods—but also as important “stopover” areas where migrating birds make briefer stops when they are migrating to or from their breeding grounds (Warnock, 2010). Waterbirds migrate along broadly similar, well-established routes known, as flyways. The Mediterranean/Black Sea Flyway is one of the eight major pathways around the globe and one of the three connecting Europe with Africa (BirdLife International, 2017). When wetlands along a bird’s predetermined migration route disappear, the likelihood of the birds completing their migration is strongly impoverished (Moore et al., 2005).
To that end, more than 2,400 sites with international importance are protected by the Ramsar Convention (Ramsar convention, 1994; Ramsar Sites Information Service [RSIS], 2020). Nonetheless, the Ramsar site characterization criteria for waterbirds (criterion 5: A wetland should be considered internationally important if it regularly supports 20,000 or more waterbirds, and criterion 6: A wetland should be considered internationally important if it regularly supports 1% of the individuals in a population of one species or subspecies of waterbirds), cannot be easily met by all wetlands on the migration route of birds (Ramsar Sites Criteria, 2020). For example, Greece has about 400 wetlands (Greek Biotope Wetland Centre [GBWC], 2020), but only ten are protected under the Ramsar Convention (Greek Ramsar Sites, 2020).
To further enlarge the protection of bird species, BirdLife International, in collaboration with national NGOs working with birds, have established a network of Important Bird Areas (IBAs), including sites critical for the conservation of birds worldwide (BirdLife International, 2001). Depending on different numerical thresholds, the international importance of a site for a species may be categorized at three distinct geographical levels, from global (criteria A), to European (criteria B) and to European Union (criteria C) (IBA Criteria, 2001). Up to present, more than 13,000 sites at a global scale have been included in the IBA network (BirdLife International, 2020). The identification of IBAs in Europe is based on a site’s international importance for: (a) threatened species, (b) congregatory bird species, (c) assemblages of restricted-range bird species, and (d) assemblages of biome-restricted bird species (IBA Criteria, 2001). The criteria build upon existing legal instruments, like the EUs Birds Directive which was adopted in 1979, and it took its current form in 2009 (Birds Directive 2009/147/EC, 2009). The European Union Birds Directive obliges all member countries to protect habitats supporting birds listed in the Annex I of the Directive, also regularly occurring migratory species not listed in Annex I, and to designate Special Protection Areas (SPA). The SPAs, together with the Sites of Community Importance (SCI) and the Special Areas of Conservation (SAC), which are based on the Habitats Directive (Habitats Directive 92/43/EEC, 1992), form the Natura 2000 network of protected areas in Europe (European Commission, 2016).
For birds crossing into Africa, the Mediterranean Sea constitutes a significant obstacle and migration over the sea is concentrated at a number of narrow straits and “land bridges” like those formed by the Italian and the Greek peninsula. The high mountainous morphology encountered in Greece, separates the peninsula into a western and eastern route. With the aim of creating a network of sites to ensure that migratory species find suitable breeding, stop-over and wintering places along their respective flyways, Greece has identified 196 IBAs (Hellenic Ornithological Society [HOS], 2019a). The eastern route, hosts most of the Greek Ramsar sites and marine IBAs (Important Bird Areas for Seabirds) and many IBAs are distributed across the islands of the Aegean Sea and the east coastline of mainland Greece on different latitudes (Fric et al., 2012; Ramsar, 2017; Hellenic Ornithological Society [HOS], 2019a). Most IBAs along the west coast of Greece are mainly located at higher latitudes. In the northern part, the most important wetlands exist in the form of large lagoon and delta complexes and they attract high numbers of wintering and staging birds during migration (Maragou and Mantziou, 2000; Liordos et al., 2014).
Along the western route, at lower latitudes, wetlands are scarcer, and the Gialova Lagoon wetland (GLw) is one of the few remaining IBA wetlands (Gialova, GR119) along the south-west coast of Greece (Heath et al., 2000). While few waterbirds use the wetland as a nesting ground, the area is important as a wintering and a stopover bird area (Kardakari, 2000; Bousbouras et al., 2011), and for some waterbirds the area meets IBA criteria A4i (The site is known or thought to hold, on a regular basis, ≥ 1% of a biogeographic population of a congregatory waterbird species) and B1i (The site is known or thought to hold ≥ 1% of a flyway or other distinct population of a waterbird species) (Table 1). To that end, it has been already classified as fulfilling the Ramsar criterion 4 (A wetland should be considered internationally important if it supports plant and/or animal species at a critical stage in their life cycles, or provides refuge during adverse conditions) and criterion 6 (BirdLife International, 2001), but is it not designated as a Ramsar site (Greek Ramsar Sites, 2020). The GLw is part of a wider Natura 2000 area (SPA site: GR2550008, 2001; Birds Directive 2009/147/EC, 2009, and SCI/SAC site: Habitats Directive 92/43/EEC, 1992; GR2550004, 1995). In addition to birds, the lagoon supports several fish species with commercial value (Koutsoubas et al., 2000; Zoulias et al., 2017), and it has a rich benthic diversity (Arvanitidis et al., 1999; Koutsoubas et al., 2000; McArthur et al., 2000). Moreover, the surrounding coastal area is the basic nesting habitat for the only European population of the critically endangered African Chameleon (Chamaeleo africanus) (Legakis and Maragou, 2009).
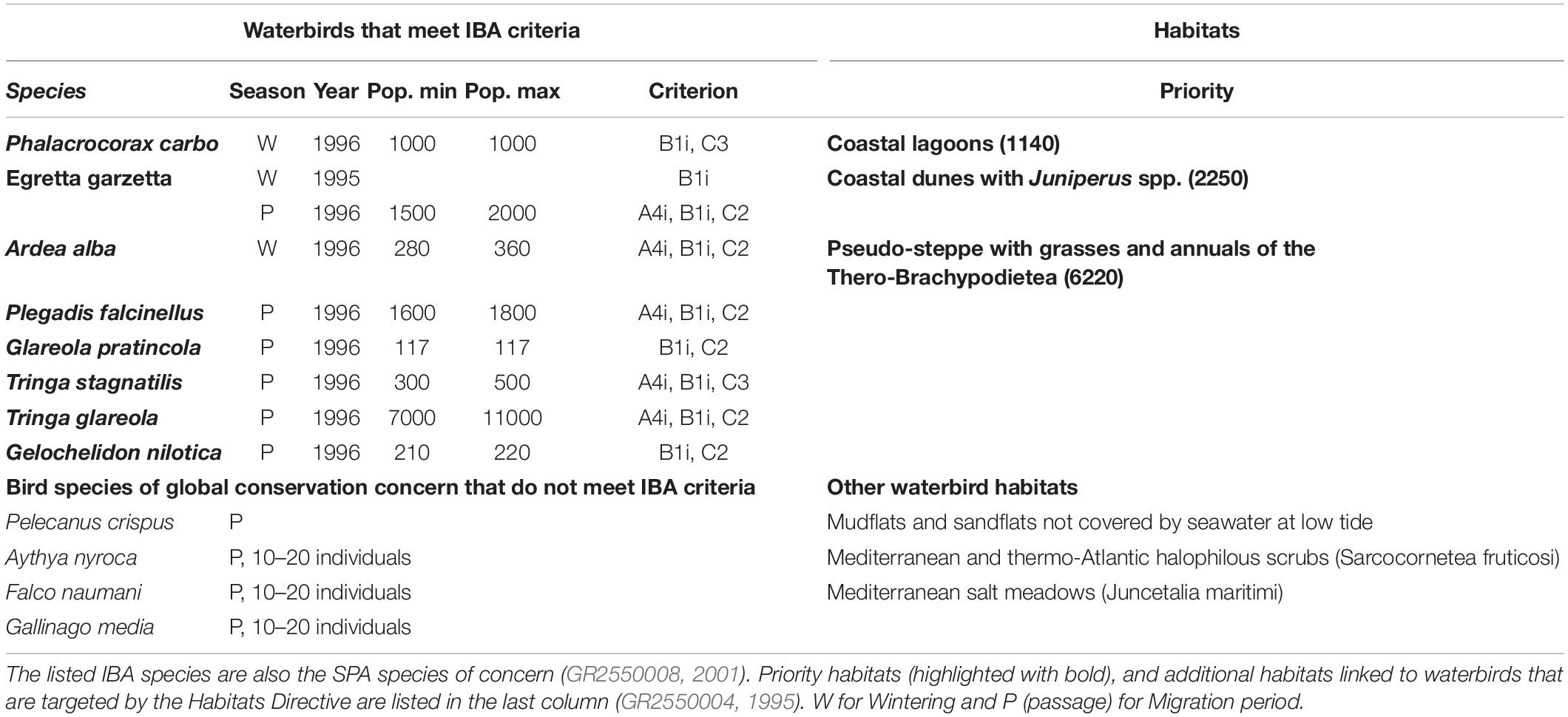
Table 1. Waterbirds in GLw-Natura2000 that meet the IBA criteria A4i, B1i, C2, C3 (Heath et al., 2000; BirdLife International, 2001), and species of global conservation concern that do not meet IBA criteria (Hellenic Ornithological Society [HOS], 2019b).
Despite its rich biodiversity, the area has been under unstable management for several years, and due to the lack of a clear management scheme1 the functions of the wetland continue to degrade (Maneas et al., 2019). Over the years, the combined effects of increased salinity and limitation in water circulation, due to anthropogenic interventions, have led to extensive mortality of emergent aquatic macrophytes (reed and cattail), which are typical bird habitats (ibid). Even today, several parts of the wetland are gradually transformed to agricultural land, or parking lots reducing the size of the natural habitats (Maneas et al., 2019). Another major threat for aquatic species and habitats, is the problem of salinity which has already been linked to waterbirds conservation in the area (Kardakari, 2000). Recent studies have shown that at present the lagoon is characterized as saline with hypersaline conditions for nearly 30% of the year, a percentage which is expected to increase under future warmer and drier climatic conditions (Manzoni et al., 2020).
An improved water management, which will ensure increased fresh water inputs to the wetland, is already demanded by the local fishers for improving fish stocks in the lagoon (personal communication with local fishers), but such management plan should also aim to create favorable conditions for birds. However, the available knowledge on the current GLw birds’ status and distribution (Standard Data Forms for site GR2550008, 2001) is based on old data reported in Kardakari (2000). Given the high salinity values, the degradation of habitats and the anthropogenic pressures mentioned above, there is a high probability that there have been changes in species numbers and the overall importance and use of the site by birds, which will need to be considered in future management strategies for the area.
Greece is currently validating the existing Environmental Assessments for all the Natura 2000 sites, including the GLw site, but in the absence of updated information, it is very likely that any management plan for the GLw will be based on previous data, which are quite detailed and important, but outdated. To that end, under this study we have contacted monthly field visits during the period October 2016 to January 2019, with the aim to complement existing information about the site and to evaluate the current status and distribution of waterbirds in the GLw, and to also provide a basis for comparison with previous studies. Since the wetland is divided in several sub-areas (Maneas et al., 2019), a sub aim of the study was to understand which parts of the wetland are mostly used by birds, identify conservation needs and potential management implications. An additional and explicit aim of our study was to re-assess the IBA/Ramsar criteria and the status of the site, as the last evaluation was in 2001 (BirdLife International, 2001). Our open data, results and suggestions could be used as insights for the sustainable management of the area and relevant decision making at local/national level, but also as updates for the waterbirds’ status in a Greek IBA at a European/International level.
The GLw is a coastal wetland located in south-west Messinia, Greece (latitude: 36°58′, longitude: 21°39′), along the Mediterranean/Black Sea Flyway migration route (BirdLife International, 2017), at the most southern-western side of the Balkan peninsula (Figure 1). It is the first wetland on the west side of the Balkan peninsula that migratory birds come across when they return from Africa in spring or the last before crossing the Mediterranean during the autumn migration (Hellenic Ornithological Society [HOS], 2019b). The boundaries of our study area, are defined at the north and east by two canals constructed during the 1960s (Figure 1). At the west, a rocky hill (also known as the Palaiokastro) and the semi-enclosed Voidokilia Bay separate the area from the Ionian Sea. A 3.8 km long and about 150m wide natural sand formation, separates the area from the semi-enclosed Navarino Bay to the south. In this study, we will refer to this area as the GLw-Natura2000 area (purple line in Figure 1).
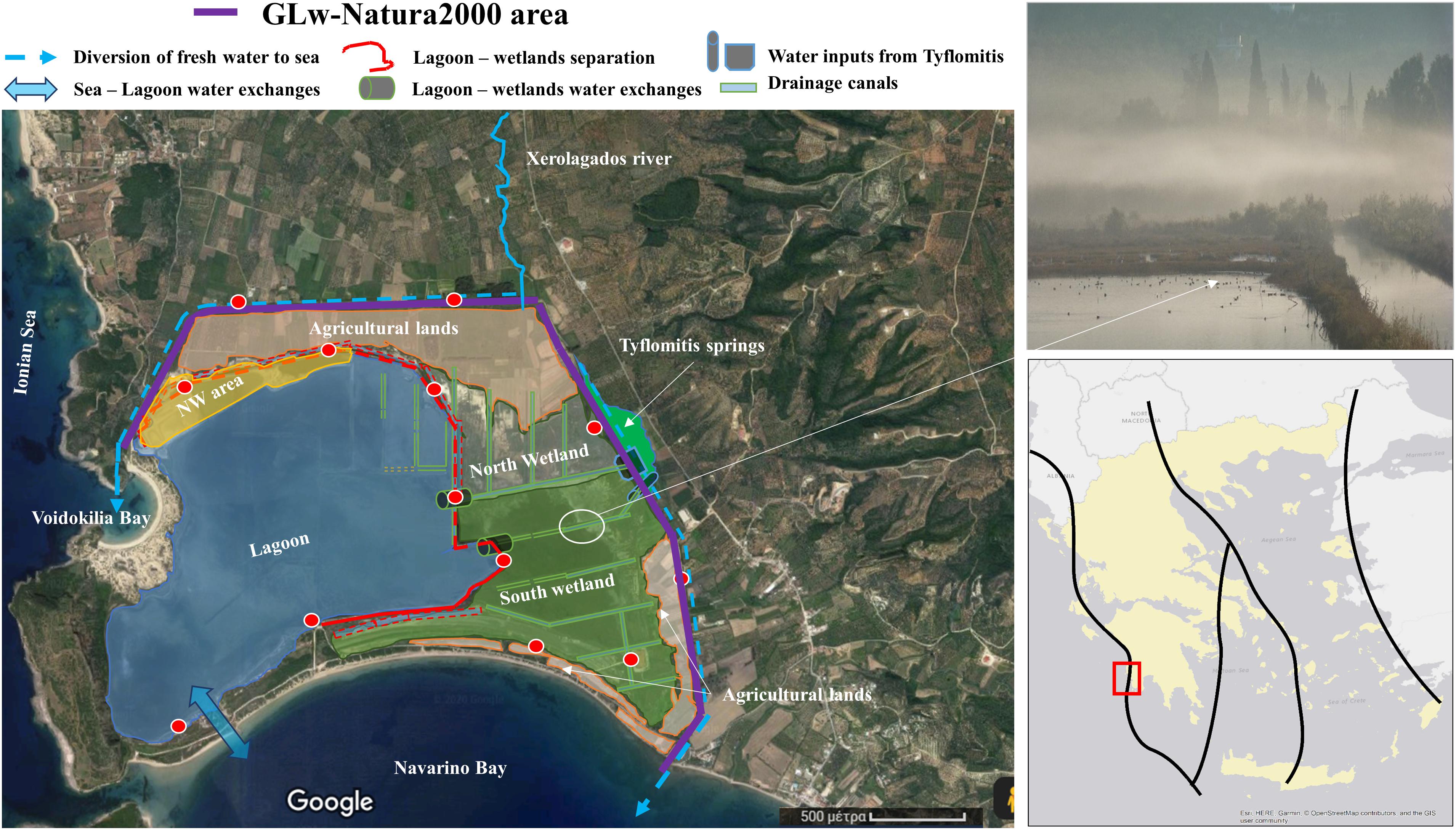
Figure 1. Left side: The map of the study area located in SW Greece (36° 58′ N, 21° 40′ E), showing the different observation points (red dots), and the major sub-areas. It also contains information about the water connections. Right side: Birds’ migration “national” routes in Greece (Hellenic forestry, 2020) along the Mediterranean/Black Sea Flyway (BirdLife International, 2017), and a picture of the site which captures one of the drainage canals in the form that it looks today.
The selected case study is part of a wider area which is characterized as an IBA, a Wild Life Refuge, and it is included in the Natura 2000 network as a Special Protection Area (SPA), under the Birds Directive (site: GR2550008, 2001; Birds Directive 2009/147/EC, 2009), and as a Site of Community Importance (SCI) and as a Special Area of Conservation (SAC), under the Habitats Directive (site: Habitats Directive 92/43/EEC, 1992; GR2550004, 1995; Table 1).
Due to man-made constructions over a period of seventy years, the GLw-Natura2000 area has been divided into sub-areas, with different characteristics (Table 2). At the east of GLw-Natura2000 area, the Tyflomitis artesian springs (a 0.072 Km2 aquatic habitat covered with reeds) provide freshwater inputs in this area (Manzoni et al., 2020). The Tyflomitis area was once connected to the wetland, but at present it is separated from it via a dike which diverts most of the up-welling groundwater to the sea (Maneas et al., 2019).

Table 2. Description of the sub-areas inside the GLw-Natura2000 area, based on Maneas et al. (2019).
Overall, the area is characterized by Mediterranean climate, with mild wet winters and dry summers (Figure 2). Evaporation and temperature trends, exhibit a maximum during the summer months, when precipitation is at its minimum (Maneas et al., 2019). The mean annual temperature is 18°C and the mean annual precipitation is approximately 695 mm/y (measured from 1956 to 2011 at the Helenic National Meteorological Service’s station of Methoni, 15.6 km South of Gialova).
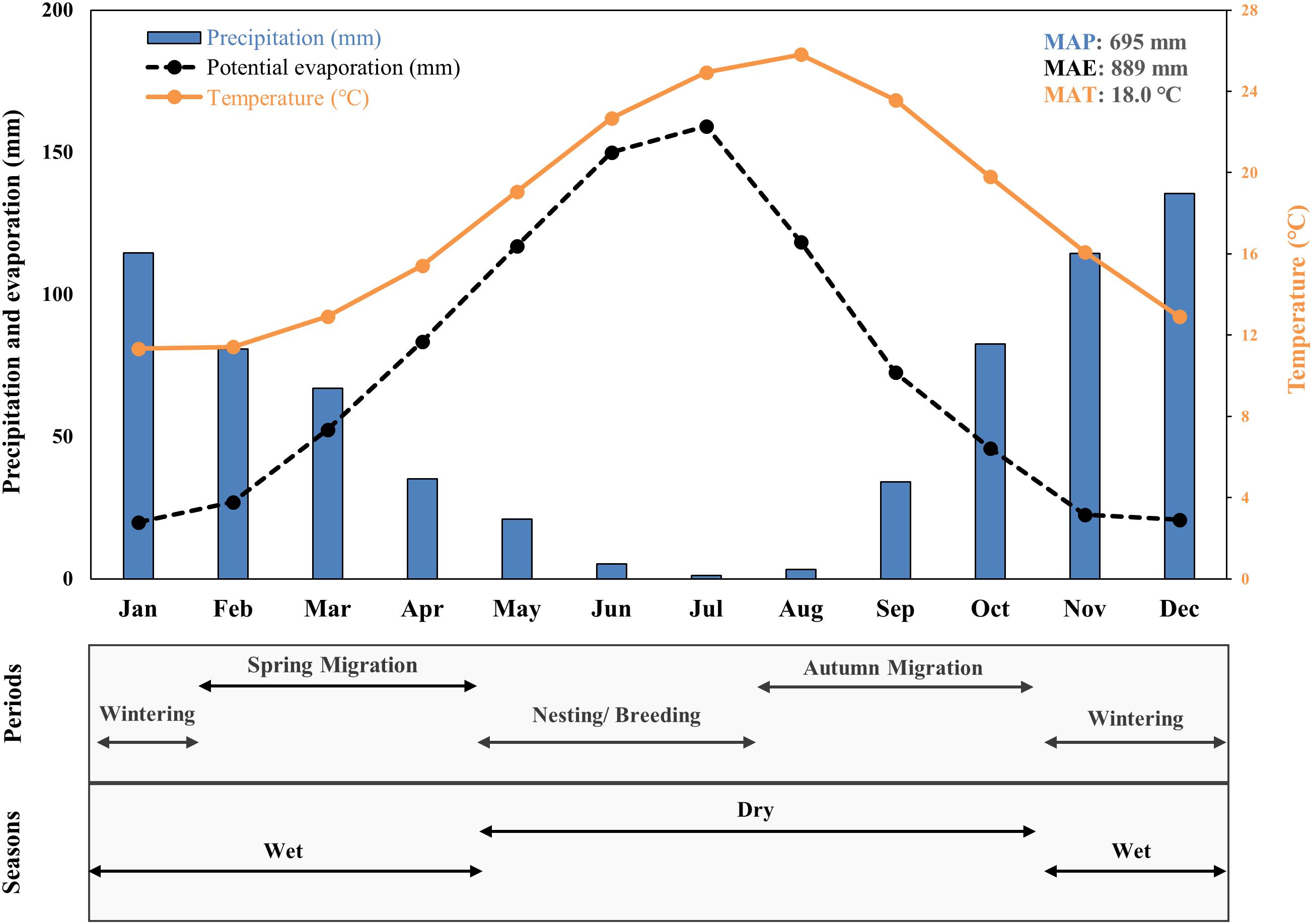
Figure 2. Graph by Maneas et al. (2019), showing monthly mean precipitation (MAP), evaporation rate (MAE) and temperature (MAT) at the study area for the period 1956–2011. At the bottom, a graph showing the birds’ periods and seasons used for the analyses.
Bird monitoring was based on monthly field visits during the period October 2016 to January 2019. During the first year (October 2016–October 2017), field monitoring was not performed in December 2016 and July 2017. In March 2017, the censuses were more frequent (every second day from March 9 to 24) to better cover the spring migration. The second year of monitoring, started in January 2018 and was completed in January 2019. During the second year, field monitoring was not performed in December 2018.
Bird counts were based on a protocol proposed by the Hellenic Ornithological Society (Bousbouras et al., 2011) following a methodology with predetermined observation points with good view, to extensively survey the GLw-Natura2000 area. The protocol consisted of 13 points covering all the sub-areas of the wetland (Figure 1). The observations started with the first light spending maximum 30 minutes at each point. During the breeding period, extra observation time was spent in specific points to thoroughly look for nests and juveniles. The field equipment consisted of binoculars (10x magnification), a scope (20-60x85) and a digital camera. During fieldwork, the Collins birds guide (Svensson et al., 2010) was used when help in bird identification was needed. Apart from the intense March 2017 counts, the rest of the counts were conducted by the same two observers.
During the censuses, all observed birds (seen and heard) were recorded, i.e., a complete census (Gregory et al., 2004), and they were listed according to BirdLife taxonomic basis (BirdLife International, 2019). The primary focus of the monitoring was to record waterbirds richness and abundance (Anatidae, Charadriidae, Laridae, Phalacrocoracidae, Phoenicopteridae, Podicipedidae, Rallidae, Recurvirostridae, Scolopacidae, Sternidae, and Threskiornithidae). Waterbirds were noted and counted with high precision (richness and abundance at exact location) into the different sub-areas (Table 2).
The rest of the species, were only noted as extra species, but their abundance in the wetland was not counted. They were grouped as wetland-dependent species and as non-wetland species. Under wetland-dependent species, birds of prey which use the wetland as a feeding area (e.g., Circus aeruginosus, Circus cyaneus, and Pandion haliaetus), the Alcedo atthis, and species from the order of Passeriformes (Acrocephalus arundinaceus, Acrocephalus melanopogon, Acrocephalus scirpaceus, Acrocephalus schoenobaenus, Riparia riparia, Emberiza schoeniclus, Cettia cetti, Cisticola juncidis, Remiz pendulinus, Panurus biarmicus) which also depend on wetland habitats, were added.
Species richness (S), as the number of observed species per month, was estimated for all the observed species, and for the different categories (waterbirds, wetland-dependent, non-wetland, all observed species).
Waterbirds species abundance (N), as the number of observed waterbirds during each count, was estimated for the whole GLw-Natura2000 area, and for each of the different sub-areas (as described in Table 2).
Waterbirds relative abundance (RN), as the number of observed waterbirds during each count per unit area (km2), was estimated for each sub-area to allow comparisons during seasons and during periods.
Waterbirds Shannon-Weiner Index (H) was calculated based on species abundance using the Shannon and Weaver (1949) formula (Bibi and Ali, 2013; Issa, 2019):
where (H) is the Shannon Index, Pi is the proportion of each species in the sample, and ln(Pi) is the natural logarithm of this proportion. The index was estimated for each sub-area to allow comparisons during seasons and during periods.
Our analysis, was divided into two seasons following weather patterns as described in section “Study Area” (Wet: November–April and Dry: May–October), and four periods based on birds’ patterns (Wintering, Breeding/Nesting, Spring and Autumn migration) (Kardakari, 2000; Figure 2). Since the four periods overlap with each other (Kardakari, 2000), to avoid double counts in our statistical analysis, we followed the below month selection:
| Wintering period (W): | November to January |
| Spring migration (sM): | February to April |
| Breeding/Nesting (B): | May to July |
| Autumn migration (aM): | August to October |
To assess the temporal distribution of wetland-dependent, non-wetland and of all the observed birds, the index of species richness was estimated for the two seasons and the four periods.
To assess the temporal distribution of waterbirds, apart from species richness we also used the index of species abundance. Both indices were estimated for the two seasons and the four periods. Since the different sub-areas are of different size, to assess the spatial distribution of waterbirds per season and per period, we used the indices (RN) and (H). As described above, relative abundance is the number of birds per unit area, thus it is independent of the size of each area and suitable for site comparisons. The Shannon-Weaver index is commonly used for site comparisons, and it was calculated in order to interpret differences in the species diversity (e.g., the index increases as both the richness and the evenness of the community increase) (Bibi and Ali, 2013; Issa, 2019). Nonetheless, we also present our (S) and (N) results, to provide a broader view of how waterbirds use the aquatic habitats of the GLw_Natura2000 area.
The count data were analyzed with GLMs (Generalized Linear Models) assuming Poisson or Negative Binomial (for overdispersed data) distribution of the dependent variable, using a log link function, as it is suggested for count data analysis (Seavy et al., 2005; O’Hara and Kotze, 2010; Warton et al., 2016).
To test if there were statistically significant differences in species richness and abundance among the seasons, and among the periods, our interpretation was based on pairwise comparisons of the EM Means (Estimated Marginal Means) produced by the GLMs, applying the Bonferroni adjustment for multiple comparisons. The analysis of (S) was conducted with Poisson GLMs considering the individual effects of season and period. The corresponding analysis of (N) was conducted with Negative Binomial GLMs since its distribution was found to be overdispersed.
To identify statistically significant differences in relative abundance across the different sub-areas per season and per period, our interpretation was based on pairwise comparisons of the EM Means produced by the GLMs with Bonferroni adjustment for multiple comparisons. The analysis of (RN) for each season and each period was conducted with Negative Binomial GLMs, using sub-areas as the factor with the main effects. For comparing the Shannon diversity indices, we relied on the Hutcheson t-test, which is developed as a method to compare the diversity of two community samples using the Shannon diversity index (Hutcheson, 1970).
In all the analysis the statistical significance was set at α ≤ 0.05. The analysis and processing of the results was conducted in Microsoft Excel, and the statistical analysis was conducted in IBM SPSS Statistics Data Editor.
The conservation status of each species at International level was retrieved from the databases of the “International Union for the Conservation of Nature” (IUCN, 2020) and Wetlands International (Wetlands International, 2020). The conservation status of each species at European level was retrieved from the “European Red List of birds” (BirdLife International, 2015) and the Annex I of EU Birds Directive (Birds Directive 2009/147/EC, 2009). The national conservation status was retrieved from the “The Red Book of Endangered Species in Greece” (Legakis and Maragou, 2009). For the observed endangered species, data of their regional population (1% threshold) were retrieved from the database of Wetlands International (Wetlands International, 2020). For these species, the estimates of their Greek population were retrieved from several sources (Legakis and Maragou, 2009; Handrinos et al., 2015; IUCN, 2020).
A total of 149 bird species representing 43 families and 15 orders were recorded during the period October 2016–January 2019, in the GLw-Natura2000 (Supplementary Table A1). Out of the total number of species, 81 (54 %) species were identified as wetland related species, of which 66 (44%) species were waterbirds representing 7 orders and 11 families (Supplementary Table A1). On a monthly basis, an average of 38 species were detected at the GLw-Natura2000 area (min: 25, max: 83) (Figure 3). The highest number of bird species was recorded during the Spring migration and the lowest during the Breeding/Nesting period (Figure 3 and Supplementary Table A2).
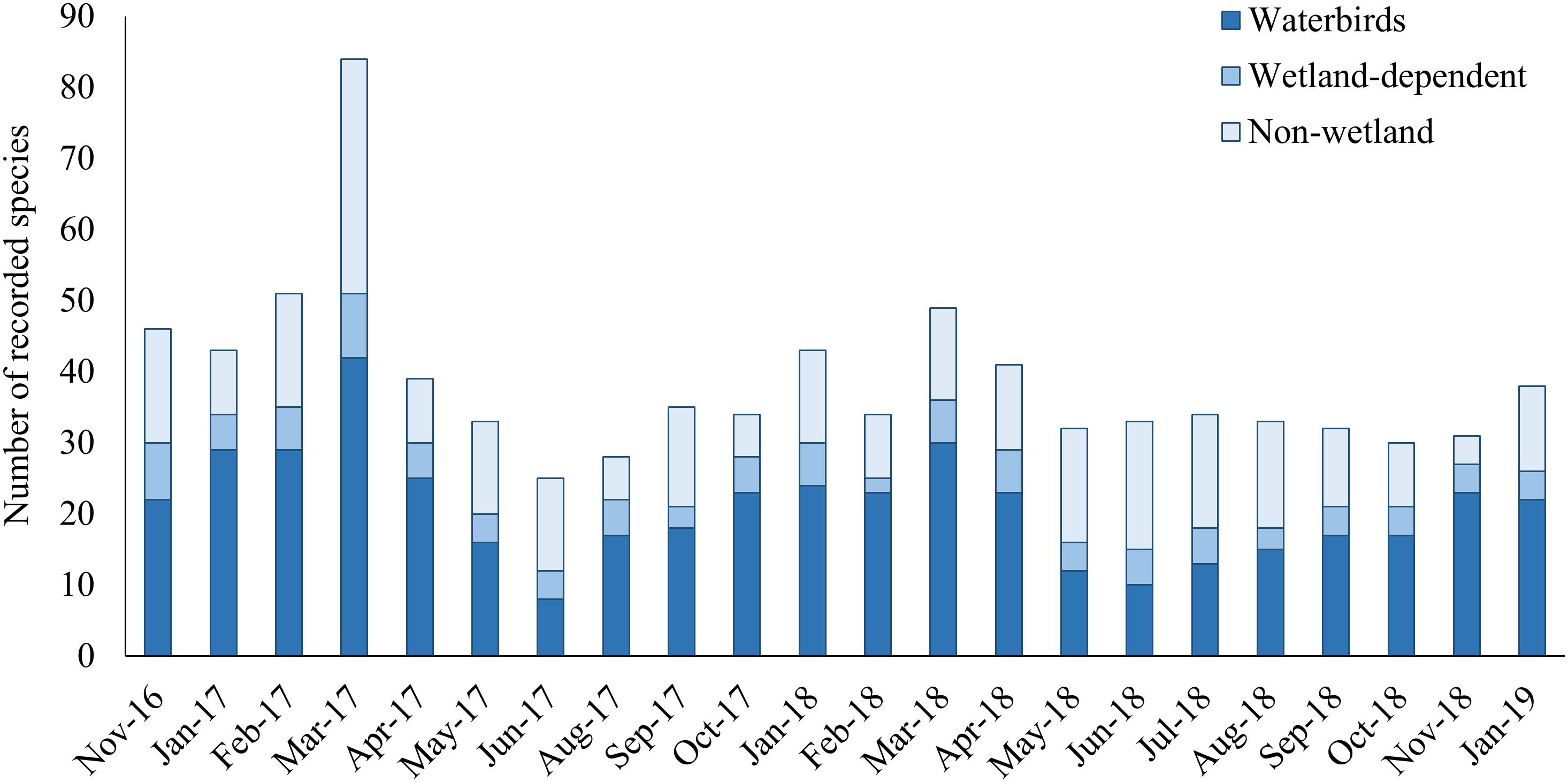
Figure 3. Number of monthly recorded species in the GLw-Natura2000 based on counts during the period November 2016—January 2019.
For all the observed birds (waterbirds, wetland-dependent and non-wetland species), species richness during the Wet season (45 ± 2.03) (mean ± standard error of mean) was higher (p < 0.001) than the Dry season (32 ± 1.7). When comparing between periods, Spring migration period was the most diverse. Species richness during the Spring migration (50 ± 2.8) was higher (p < 0.001) when compared to the Breeding/Nesting (31 ± 2.5) and the Autumn migration periods (33 ± 2.3), but not when compared to the Wintering period (40 ± 2.8). For wetland-dependent and non-wetland species, the estimates of the GLM showed no statistically significant difference of species richness per season or per period, indicating that the wetland is used by birds all year around.
Waterbirds richness during the Wet season was around 75% higher when compared to the Dry season (p < 0.001) (Table 3). When comparing between periods, the Spring migration period was the most diverse. The GLM showed that species richness during the Spring migration period was higher than the Breeding/Nesting period by almost 140% (p < 0.001), and by almost 60% (p = 0.001) when compared to the Autumn migration period. Species richness during the Spring migration was also higher when compared to the Wintering period, but this difference was not statistically significant (Table 3). The Wintering and the Autumn migration periods were also more diverse when compared to the Breeding/Nesting period, but not when compared to each other (Table 3). According to the model, during the Wintering period the species richness was almost twice as high (p < 0.001), and during the Autumn migration around 50% higher.
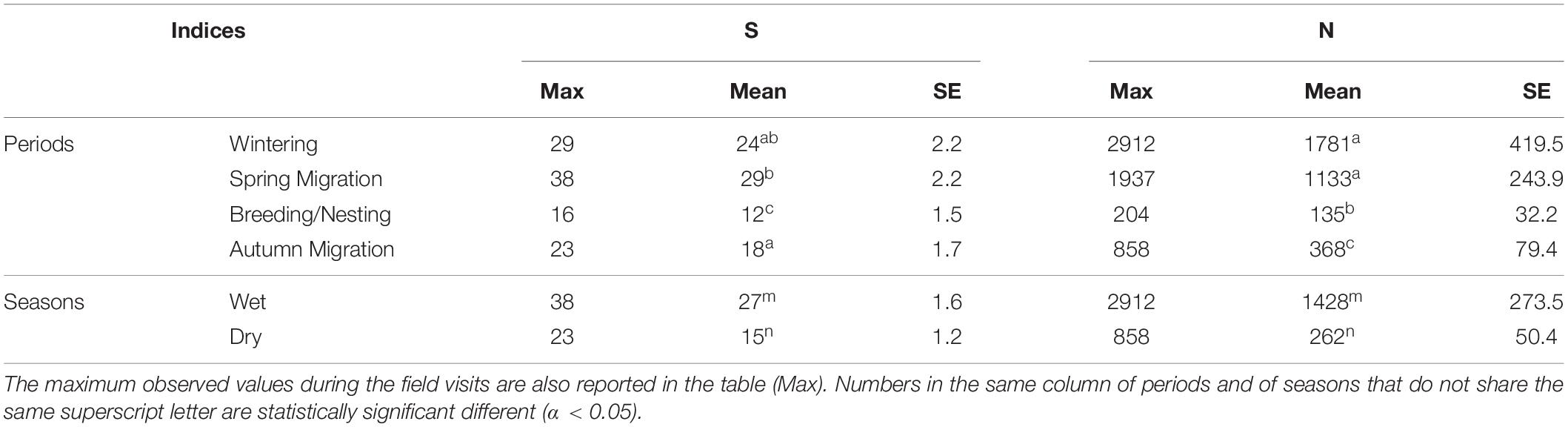
Table 3. Estimated marginal means (Mean) and standard errors (SE) for waterbirds’ richness (S) and abundance (N) during the different seasons and periods, based on GLM analysis (GLM analysis for S was based on a Poisson distribution, and GLM analysis for N was based on a Negative Binomial distribution, using log as a link function in both analyses).
Waterbirds abundance per season and per period were analyzed using a GLM with Negative Binomial distribution choosing the log link function. The GLM showed that waterbirds abundance between the two seasons was statistically significant different (p < 0.001), with estimated abundance during the Wet season being more than 400% higher (Table 3). The Wintering period had almost 57% more birds compared to the Spring migration period, but the difference was not statistically different. Waterbirds abundance during the Wintering period was higher when compared to the Breeding/Nesting and Autumn migration periods, by around 1,200% (p = 0.001), and 380% (p = 0.006), respectively. Similar to the Wintering period, waterbirds abundance during the Spring migration period, was higher when compared to the Breeding/Nesting and Autumn migration periods by around 740% (p < 0.001) and 200% (p = 0.017), respectively. In fact, waterbirds abundance during the Breeding/Nesting period was statistically significant lower compared to the Autumn migration period as well, with the abundance being 65% lower (p = 0.040) (Table 3).
Taken together, the above results suggest that waterbirds’ diversity varied significantly between seasons, with higher richness and abundance during the wet season compared to the dry season. In particular, during the Nesting/Breeding period the wetland had the lowest species richness and abundance. On the contrary, during the Wintering and the Spring migration periods, the wetland held the highest abundance. Our results also suggest that compared to the Autumn migration, the Spring migration period was more diverse in terms of both species richness and abundance.
Waterbirds variations across the aquatic habitats of the GLw-Natura2000 area varied within the different seasons and periods. Variations in waterbirds richness and total abundance provided a qualitative approach of how each sub-area was used by waterbirds per season and per period (Table 4). The S. Wetland sub-area held the highest number of species per season and per period. During the Wet season, the lagoon had the highest abundance during both periods. On the contrary, during the Dry season, waterbirds abundance was higher at the S. Wetland sub-area. Waterbirds richness at the NW area was high during the Wintering and both migration periods, but low during the Breeding/Nesting period. The N. Wetland sub-area held the lowest numbers of waterbirds richness and abundance during most of the periods, except for the Breeding/Nesting period.
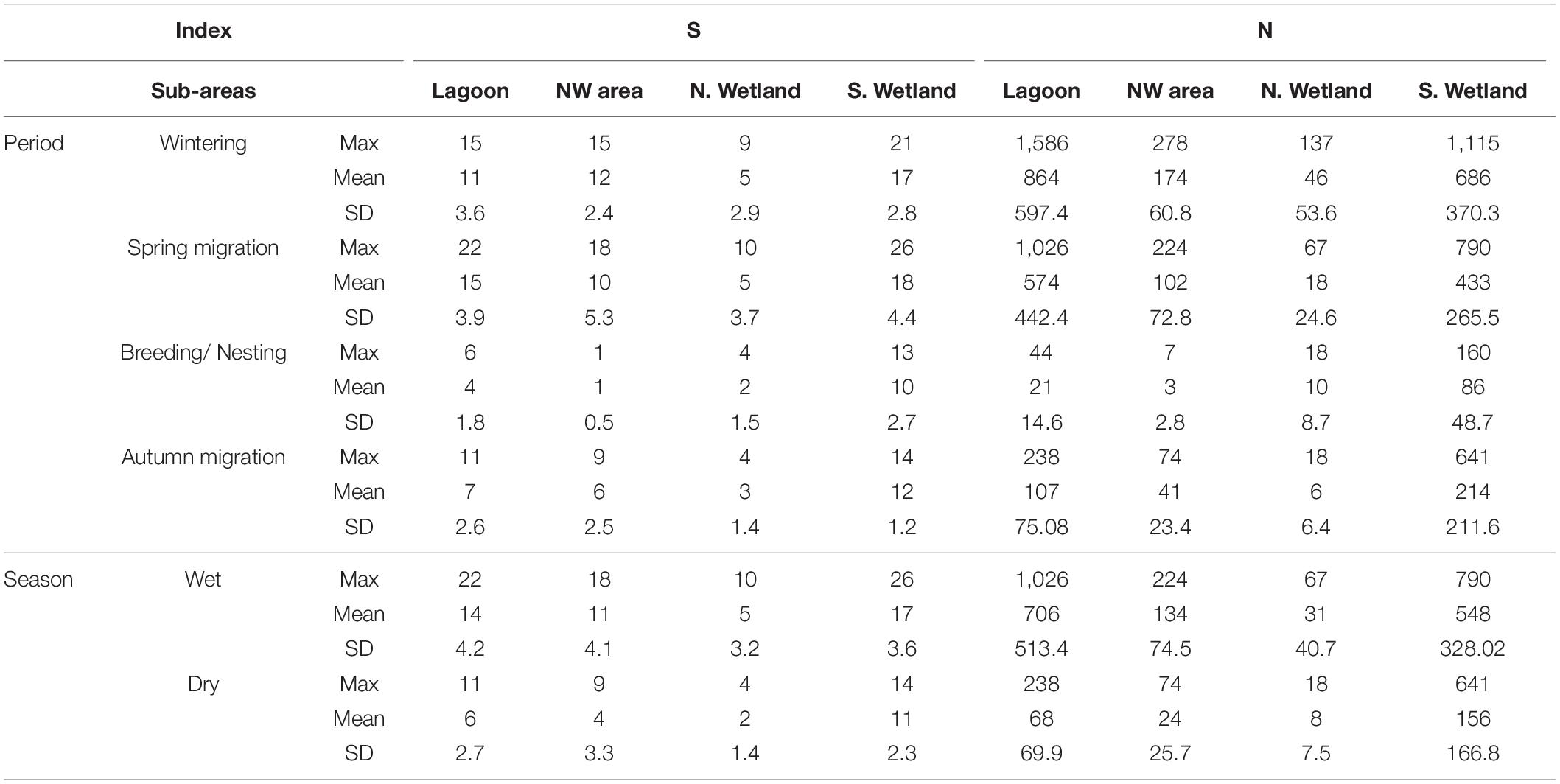
Table 4. Maximum recorded value (Max), average and standard deviation (Mean and SD) of waterbirds’ richness (S) and abundance (N) across the aquatic habitats of the GLw_Natura2000 area per season and per period.
Our quantitative approach was based on the variations of (RN) and (H) indices across the four sub-areas per season and per period (Table 5).
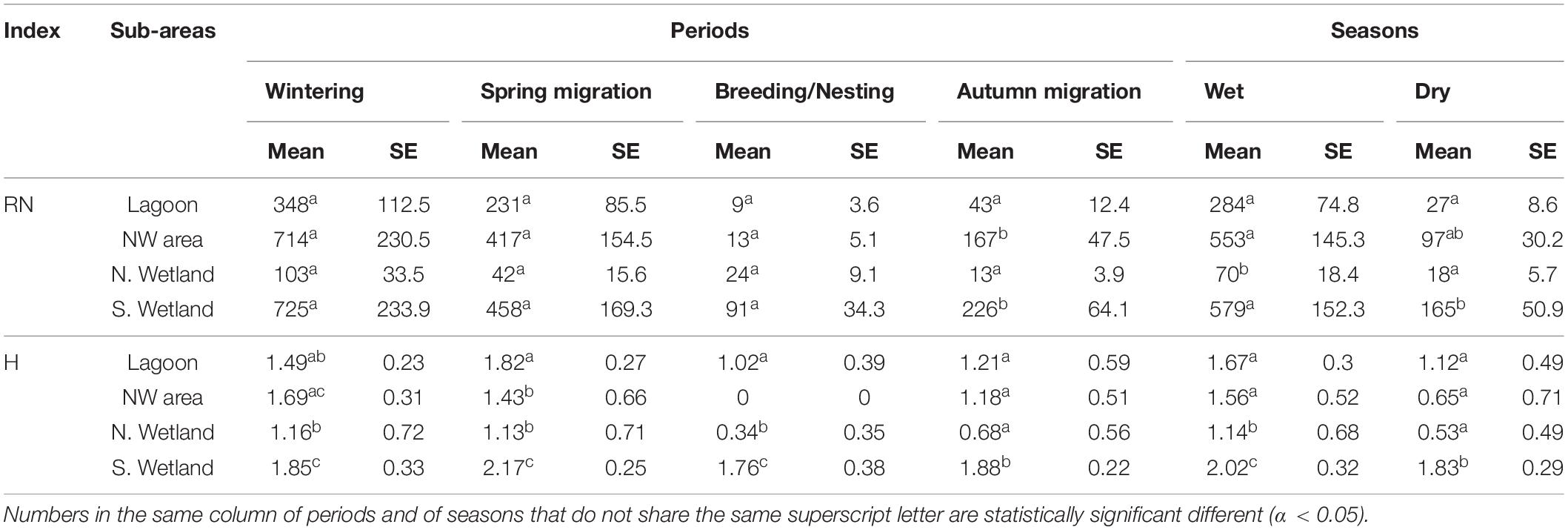
Table 5. Estimated marginal means (Mean) and standard errors (SE) for waterbirds’ relative abundance (RN), and corresponding mean values of the Shannon-Weaver index in the different sub-areas per season and per period.
During the Wet season, waterbirds relative abundance at the N. Wetland was lower from all the other sub-areas, and in particular by 88% (p = 0.005), 75% (p = 0.032) and 87% (p = 0.006) when compared to the S. Wetland, the Lagoon and the NW area, respectively. The RN in the Lagoon was 48 and 51% less when compared to the NW area and the S. Wetland, but these differences were not statistically different (Table 5). The Hutcheson t-tests, showed that the estimated value of the Shannon-Weaver index at the S. Wetland area, was statistically significant higher (p < 0.001) compared to all the corresponding (H) values in the other areas. The N. Wetland, was less diverse when compared to the Lagoon (p = 0.003) and the NW area (p = 0.030), which were the two sub-areas with similar values of H and thus similar species diversity during the Wet season (Table 5).
During the Dry season, waterbirds relative abundance at the S. Wetland was approximately five (p = 0.048) and eight (p = 0.025) times higher when compared to the Lagoon and the N. Wetland, respectively (Table 5). The (RN) at the NW area was also higher by approximately 250 and 430% when compared to the Lagoon and the N. Wetland, respectively, but these differences were not statistically different. During that time, the Shannon-Weaver index was the highest at the S. Wetland, and it was statistically different from all the other sub-areas (p ≤ 0.001). However, there were no other significant differences across the other sub-areas.
During the Wintering period, most of the waterbirds were observed at the Lagoon (Table 4), but in terms of relative abundance there were no statistically significant differences with the other sub-areas (Table 5). The total abundance of the S. Wetland was higher than of the NW area, but the estimated means of the relative abundance between these two areas were not statistically different, indicating that they were both important for waterbirds. The (RN) at the S. Wetland and the NW area was approximately double when compared to the Lagoon and six times higher when compared to the N. Wetland, but these differences were statistically insignificant. The S. Wetland not only held the highest species richness, but it was also the most diverse area as indicated by the value of the Shannon-Weaver index. The difference was statistically significant higher when compared to the Lagoon and the N. Wetland (p < 0.001), but not when compared to the NW area, complementing the above results.
During the Breeding/Nesting period, the estimated mean of (RN) at the S. Wetland was about six, nine and almost three times higher when compared to the NW area, the Lagoon and the N. Wetland, respectively, but the differences were not statistically different (Table 5). The Shannon-Weaver index at the S. Wetland was significant higher from both the Lagoon and the N. Wetland (p < 0.001), which suggests that the S. Wetland was the sub-area with the higher waterbirds diversity during the Breeding/Nesting period. The NW area was the sub-area with the lowest observations in terms of waterbirds richness and abundance, and thus the (H) was estimated at zero.
During the two migration periods, waterbirds patterns were different. During the Spring migration period, similar to the Wintering period, most of the waterbirds were observed at the Lagoon, and in high numbers at the S. Wetland (Table 4), but in terms of relative abundance there were no statistically significant differences with the other sub-areas (Table 5). On the other hand, during the Autumn migration period, the relative abundance at the Lagoon, the N. Wetland and the NW area were 80% (p = 0.030), 92% (p = 0.007), and 94% (p = 0.005) less when compared to the corresponding value at the S. Wetland (Table 5). The Shannon-Weaver index at the S. Wetland sub-area, was the highest in both migration periods, and different from all the other sub-areas (p ≤ 0.009). During the Spring migration, the (H) values at the Lagoon were higher when compared to the NW area (p = 0.002) and to the N. Wetland (p = 0.004), a difference which was not evident during the Autumn migration (Table 5).
Summarizing the above, our results suggest that all the sub-areas supported waterbirds, but their occupation by waterbirds varied between seasons and periods. During the Wet season, waterbirds were more evenly distributed around the different sub-areas of the GLW-Natura2000 wetland, indicating that the whole area was used as a feeding or/and resting area. This pattern gradually started to change during the Dry season and became evident during autumn.
The S. Wetland was the sub-area with the highest species diversity during all year, and the highest relative abundance during the Dry season and corresponding periods (Breeding/Nesting and Autumn migration). During our field visits, into this area we recorded most of the nests, and taken together these results suggest that the S. Wetland sub-area apart for suitable area for feeding and resting, it is also used as a nesting ground. Similar to the S. Wetland, the Lagoon was also used by waterbirds all year around, but mainly during the Wet season and the relevant periods (Wintering and Spring migration). The high numbers of (RN) and (H) at the NW area, during the Wintering and both migration periods, suggest that this area was important as a feeding and resting area. On the other hand, the limited observations during the Breeding/Nesting period, and the absence of recorded nests suggest that waterbirds avoid to use this area as a nesting ground. The N. Wetland, was the sub-area which had the lowest (RN) and (H) during both seasons and most periods. However, during the Breeding/Nesting period, both indices were higher than those at the Lagoon and at the NW area, which highlights the importance of that area during that period. This finding is further supported by the fact that we recorded the N. Wetland as the basic nesting habitat of the Cygnus olor during both field visits (summer 2017 and summer 2018).
Wildfowl observed at the wetland were dabbling and diving ducks, geese and a swan (Supplementary Table A1). Out of the 13 recorded wildfowl, 7 are listed in the IUCN lists (GR: one CR, four VU; Europe: two VU, International: one VU, one NT). Two species (Anas platyrynchos and Cygnus olor) were recorded as resident and breeding species, 6 as wintering species (Anas acuta, Anas crecca, Anas clypeata, Anas penelope, Aythya nyroca, and Tadorna tadorna) while the rest 5 Anatidae (Anas strepera, Anser anser, Aythya ferina, Aythya marila, and Spatula querquedula) were recorded during the Spring migration and marked as migrating species. During the Wintering and Spring Migration periods, wildfowl were observed in all the aquatic habitats of the wetland, but almost 90% of their population was observed at the Lagoon and S. Wetland sub-areas (Figure 4). During the dry season, when their numbers in the area were low, they were mainly observed at the east part of the wetland and in particular at the S. Wetland sub-area (Figure 4).
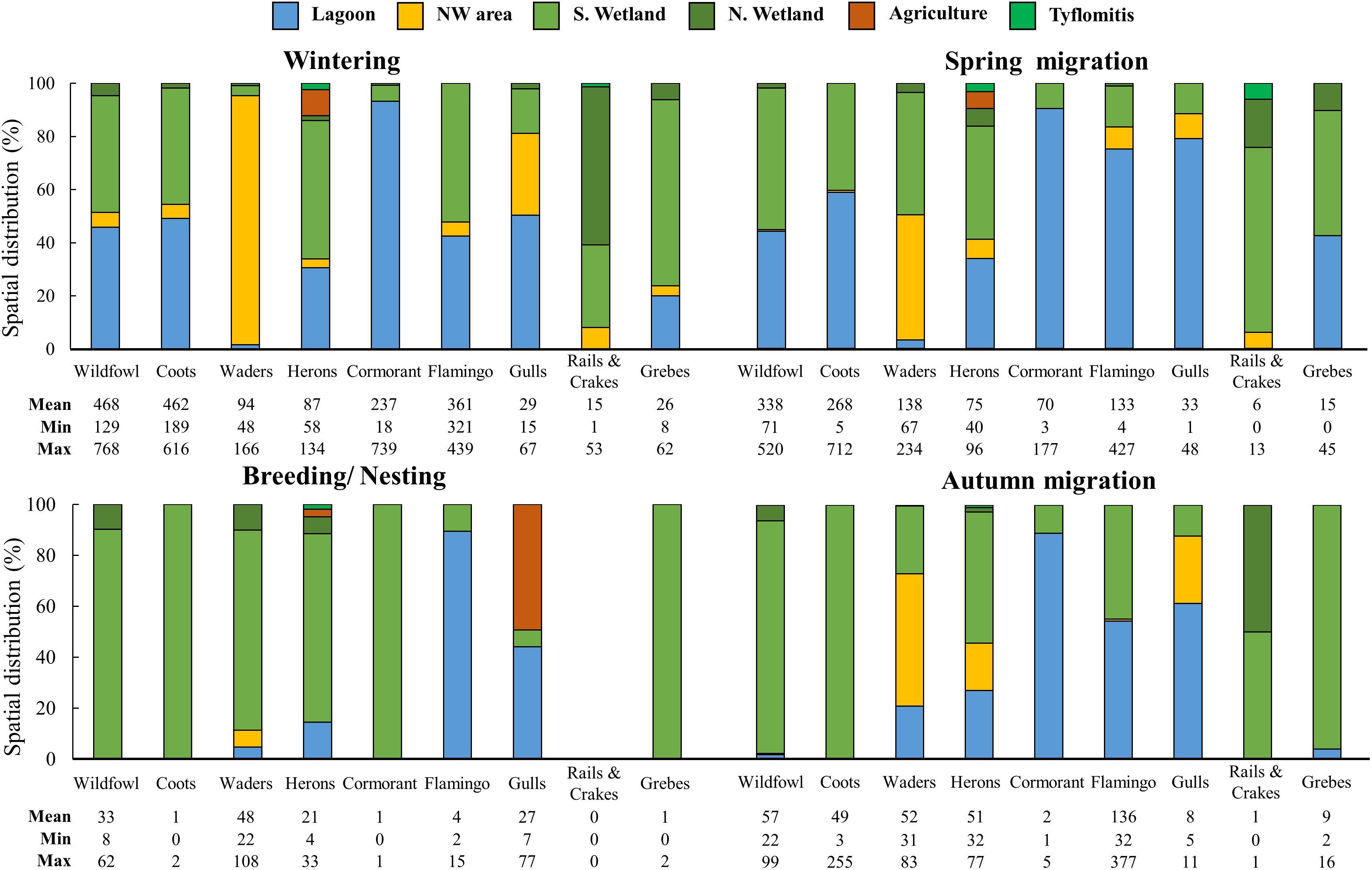
Figure 4. Stacked column graph showing the spatial distribution (percentage) of waterbirds’ categories in the different sub-areas of the GLw-Natura2000 area, based on their average population distribution during the different periods. The abundance (mean, minimum and maximum values) of each waterbirds category during the same period is presented in the table at the bottom of each graph.
Coots were found in higher numbers during the wet season, and similar to wildfowl more than 90% of their population was observed at the Lagoon and S. Wetland sub-areas (Figure 4). During the dry season, the few coots were recorded at the S. Wetland side. One pair of coots was observed nesting in the area in 2018.
In total 25 different species of waders, of which 6 are threatened (GR: three VU, Europe: four VU, International: four NT), belonging to 3 different families were recorded (Supplementary Table A1). Charadrius alexandrinus was the only wader recorded in almost all counts and also nesting. Apart from the Kentish plover, Himantopus himantopus was also observed nesting in the area. However, the Black-winged stilt arrived in the area in spring and departed in autumn. Compared to waterfowl, during the Wintering period, waders were mainly observed at the NW-area side of the GLw-Natura2000 (Figure 4). During the Spring migration, their population was divided between the NW-area and the S. Wetland sub-areas (Figure 4). On the contrary, during the Nesting/Breeding period, almost 90% of all waders were recorded at the east side of the wetland, in the S. Wetland sub-area (Figure 4). During the Autumn migration, their population was more spread, but still more than 75% of the observations were made in the NW-area and S. Wetland sub-areas (Figure 4).
In total 8 different species of herons were counted in GLw-Natura2000 (Supplementary Table A1). Egretta garzetta and Ardea cinerea were recorded in all counts and marked as resident species. However even been present during the Breeding/Nesting period no sign of nesting was recorded. The highest numbers of Ardea alba were recorded during the Wintering period, and they were present during the Spring and the Autumn migration as well. Small numbers of the other heron species were recorded during migration: Ardea purpurea and Ardeola ralloides in spring and Botaurus stellaris, Ixobrychus minutus, and Nycticorax nycticorax in autumn. During most of the periods, herons were observed all around the wetland with most frequent observations at the Lagoon and the S. Wetland sub-areas (Figure 4). Herons were among the waterbirds which were observed in the agricultural lands around the wetland, mainly during the wet season when the farmlands were wet or flooded (Figure 4). During the Breeding/Nesting period most of their population was recorded at the S. Wetland sub-area (Figure 4). Plegadis falcinellus and Platalea leucorodia were observed in low numbers at the N. Wetland area during the spring migration.
Phalacrocorax carbo was the only cormorant observed in the area. The species was recorded wintering in the area and was present during the Spring migration period. The majority of their population, was recorded at the Lagoon side of the wetland (Figure 4).
Flamingos (Phoenicopterus roseus), arrived in high numbers during the Autumn migration and departed in February. They spent most of their time at the Lagoon and the S. Wetland sub-areas of the GLw-Natura2000, and they were occasionally observed at the NW-area (Figure 4).
Larus michahellis was recorded during most counts and was the most common gull in the area. Larus ridibundus was recorded during the Wintering and both migration periods, and Larus genei only during the migration periods. Out of the 5 observed tern species, Thalasseus sandvicensis was present during the Wintering and the Spring migration periods. The other four terns (Chlidonias hybrida, Chlidonias leucopterus, Hydroprogne caspia, and Sterna hirundo), were only observed during the migration periods. During the wet season, gulls, and terns were observed in all the aquatic habitats of the wetland, but more than 50% at the Lagoon sub-area (Figure 4). During the Breeding/Nesting period, gulls were also observed at the agricultural lands (Figure 4). Their population during the Autumn migration was more spread, but still more than 50% of the observations were made at the Lagoon sub-area of the wetland (Figure 4).
Rallus aquaticus and Gallinula chloropus were recorded in few numbers in the S. Wetland and N. Wetland sub-areas, mainly during the wet period (Figure 4).
Podiceps cristatus and Podiceps nigricollis were observed only during the Wintering and the Spring migration periods. On the other hand, two families with juveniles of Tachybaptus ruficollis were observed in August 2017, indicating breeding success of the species. Similar to coots and wildfowl, during the wet season more than 90% of their population was observed at the Lagoon and S. Wetland sub-areas (Figure 4). During the dry season, the majority of the observations were made at the S. Wetland sub-area (Figure 4).
Three species of raptors (Circus aeruginosus, Circus cyaneus, and Pandion haliaetus) were observed hunting in the area during the Wintering period (Supplementary Table A1). Small numbers of Grus grus were recorded at the agricultural lands surrounding the wetland during Wintering and both migration periods. Kingfisher was also common in the area during the Wintering (at least 10) and both migration periods (at least 20) and was usually observed all around the wetland. The rest of the wetland-dependent species, belong to the order of Passeriformes, and were mainly recorded at the reeds zone of Tyflomitis artesian springs (Supplementary Table A1). Acrocephalus arundinaceus, Acrocephalus scirpaceus, and Cisticola juncidis, were recorded nesting at Tyflomitis sub-area, while Cettia cetti in areas surrounding the wetland.
Almost a quarter (40 species) of the total observed species are listed in the Annex I of the EU’s Birds Directive (Tables 6, 7).
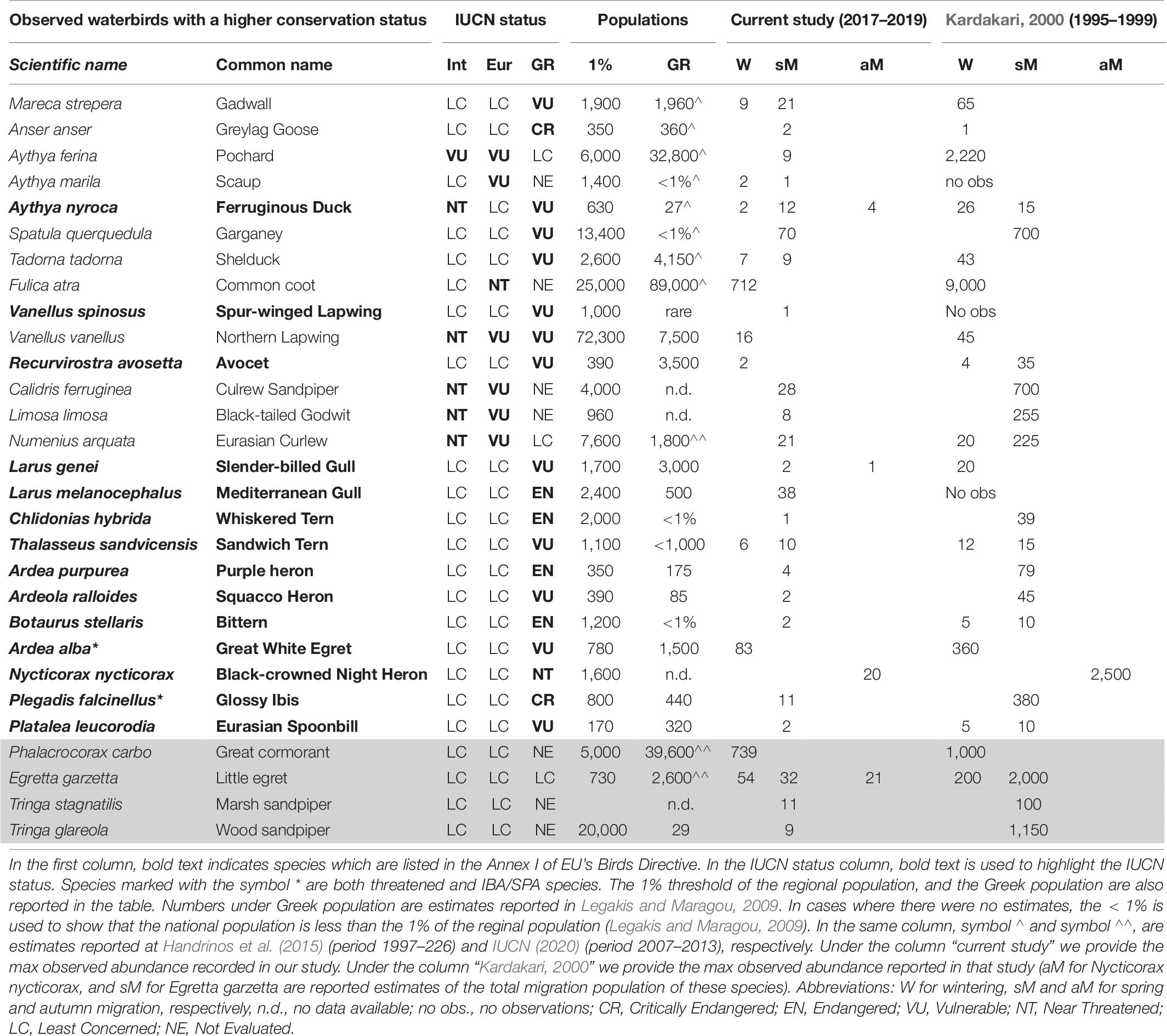
Table 6. Observed waterbirds in GLw-Natura2000, which are threatened at International (Int), European (EU) and Greek (GR) level, and additional IBA/SPA species of concern (gray part of the table).

Table 7. Observed birds in GLw-Natura2000 which are threatened at International, European and Greek level.
As evident in Table 6, our case study, did not support big wintering populations (all counts were less than 1% of the regional population), but it supported many Nearly Threatened (NT), Vulnerable (VU), Endangered (EN) or Critically Endangered (CR) species, especially during the Spring migration. At an International level, two species are listed as VU and six as NT. At a European level, eight species are listed as VU, three as NT, and all the rest (139) as Least Concern (LC). At a national level, two species (Anser anser and Plegadis falcinellus) are listed as CR, six as EN, fourteen as VU, four as NT, and fourteen as LC (Tables 3, 4). The rest 107 observed species have not been evaluated yet under the national IUCN criteria (Supplementary Table A1). From the species listed in Table 6, only the population of Phalacrocorax carbo was similar to the one reported in Kardakari (2000), and our recordings suggest declining numbers for most waterbirds including all the species with higher conservation value (Table 3 and Supplementary Table A1).
Several of the threatened bird species with an IUCN status, and those listed in the Annex I of the EU Birds Directive are connected to wetland loss and degradation due to intensification of agriculture, drainage of wetlands, disturbance from tourism activities, illegal hunting and climate change (European Commission, 2016; BirdLife International, 2018). In this context, the Glw-Natura2000 area is a representative IBA/SPA wetland (Heath et al., 2000; Birds Directive 2009/147/EC, 2009), which has been altered by past drainage efforts and agriculture expansion (Maneas et al., 2019). Furthermore, it is located in an area which is fast becoming a tourist destination, an economic activity which also poses a threat for coastal wetlands in Greece (Maragou and Mantziou, 2000; Maneas et al., 2019). At present, the area suffers from a lack of management, and conservation actions for achieving a Favorable Conservation Status (FCS) for the avian communities is urgently needed. However, before drafting any management plan it is important to understand the current status and distribution of waterbirds, also in relation to previous studies in this area (Kardakari, 2000; Bousbouras et al., 2011).
During the current study 149 species (average: 38 species per month) were recorded in the area, including 36 threatened species at an International, European or/and national level, and 40 species listed in the Annex I of the EU Birds Directive (21 species are listed as both threatened and under Annex I). 54% (81 species) of the total observed species have been identified as wetland-dependent birds, out of which 66 species (44%) were waterbirds. The results show that the wetland is mostly used by ducks, coots, herons, waders, flamingos and cormorants. Gulls, grebes, rails, and crakes were found in relatively low numbers. The higher waterbirds abundance was recorded during the Wintering period, and the higher waterbirds richness during the Spring migration period.
As reported in previous studies (Kardakari, 2000; Bousbouras et al., 2011), our results suggest the species richness and abundance were higher during the Spring migration compared to the Autumn migration. The location of the area along the Balkan peninsula, combined with better habitat during spring, should be the main reasons why the wetland has higher species richness in spring. Indeed, located at the most south-western part of the Balkan peninsula, the wetland is used as a suitable stopover for many exhausted spring migrants who have flown non-stop over the Mediterranean Sea, or even non-stop from south of Sahara (Bortels et al., 2011). The situation in the autumn is very different. When migrating from north to south over the Balkan peninsula birds have the possibility of staying in the larger wetlands at the northern part of Greece, which are very suitable stopovers for waterbirds (Hellenic Ornithological Society [HOS], 2019a). This may make a stop at GLw not as important as it may be in spring, when the wetland may be the first wetland that the birds encounter. In addition, during spring, increased volumes of freshwater due to precipitation and inflows from the catchment, result in lower salinity values and wider expansion of aquatic habitats compared to autumn conditions (Manzoni et al., 2020), making the whole area a more suitable habitat for resting and feeding.
The results of this study also confirm the importance of the site as a Wintering area (Kardakari, 2000). Each winter the GLw-Natura2000 held a high number of waterbirds; mainly wildfowl, coots, herons, cormorants and flamingos. The fluctuations of wintering populations from year to year (2017, 2018, and 2019) were expected and could be linked to reasons outside the site’s condition. Adam et al. (2015) found that very cold winters in northern and central Europe forced the birds to move further south to find open water, making concentrations of birds to appear higher than usual in places such as our case study. In the same way, a warmer winter makes it possible to avoid flying further south than necessary, lowering the numbers in southern wetlands, as in 2018 in the GLw-Natura2000.
During the Nesting/Breeding period the wetland had the lowest species richness and abundance, but some of the observed species (e.g., Rallus aquaticus, Gallinula chloropus, and Tachybaptus ruficollis) were cryptic species, and their abundance in the area might be higher. Nevertheless, some species nest in the area (e.g., Anas platyrhynchos, Cygnus olor, Himantopus himantopus, Charadrius alexandrinus), and it is important to keep their habitats untouched. Even though, large areas are needed by many species to maintain their populations, it is important to recognize the complementary value of smaller remnants such as smaller wetlands to their successful conservation (Fischer and Lindenmayer, 2002).
When compared to the other sub-areas, the higher values of the Shannon-Weaver index at the S. Wetland indicated that this part of the wetland had the highest species diversity all year around, and regularly supported high numbers of birds as shown by the comparisons of the relative abundance. Counter wise, the N. Wetland sub-area, held the lowest species diversity, abundance and relative abundance. Nevertheless, this sub-area was the basic habitat for the critically endangered (CR) Glossy Ibis. During the wet season, the Lagoon and the NW area had high species richness and abundance. The S. Wetland and the NW area were identified to be the most important sub-areas for waders during the Wintering and both migration periods. On the contrary, during the Dry season, and especially during the Breeding/Nesting period, bird diversity was much lower in most of the aquatic sub-areas. In fact, our results suggest that during the Dry season, the majority of species and individuals (including most breeding species) were summoned at the S. Wetland sub-area.
Our observations in the agricultural lands, and the Tyflomitis artesian springs sub-areas suggest that these areas are less used by waterbirds. Nevertheless, during the Wet season, herons and in particular the Ardea alba (a species which is listed as CR in the Greek IUCN red list) were frequently observed in the agricultural areas. At the Tyflomitis artesian springs sub-area, the observations were also few, but our recordings suggest that this sub-area is important as a nesting area for small wetland-dependent species.
Out of the eight IBA/SPA species of concern, Phalacrocorax carbo has been recorded in numbers similar to Kardakari, 2000, but for the other two wintering IBA waterbirds our results suggest that their abundance has been dramatically decreased since the last IBA evaluation in 2001 (BirdLife International, 2001). The maximum observed wintering abundance for Egretta garzetta (54 individuals) and Ardea alba (83 individuals) during this study (based on three winter counts), was about 75 and 80%, respectively, less when compared to corresponding values given in Kardakari (2000) (Table 6). Regarding the migrant species, although our observations during the spring migration period gave a picture of the species abundance on the day of the census, and the total number that use the wetland is multiple, they indicate that migrant species abundance has been largely decreased since the last IBA evaluation in 2001 (BirdLife International, 2001). Plegadis falcinellus was also recorded in low numbers during the Spring migration period (max: 11 individuals). This species has fluctuating numbers, but in the past, it was recorded in higher numbers (391 in 1996, and 366 in 1999) (Kardakari, 2000). Tringa glareola was referenced as the second most frequent wader in the wetland with maximum counts up to 1,150 individuals (Kardakari, 2000), but under this study we never encountered so many individuals (spring max: 8 individuals). In the same study, Tringa stagnatilis was recorded as a common wader during the Spring migration (Spring max: 100 in 1996), but under this study we recorded in total five individuals. Finally, Glareola pratincola and Gelochelidon nilotica were not observed during the study period, even though expected, since they were reported as “regular passage migrant often in large numbers” and “fairly common passage migrant,” respectively (Kardakari, 2000).
During the monitoring period of this study we have recorded one additional wildfowl species (Aythya marila—single observation) during the Spring migration period. On the contrary, our list contains 36 species less. The absence of twenty-six species from our list, which are reported as rare and very rare species in Kardakari, 2000, could be explained by the frequency and the type of our monitoring efforts. Porzana porzana, Porzana parva, and Lymnocryptes minimus are cryptic species and difficult to monitor (Stanley et al., 1977–1994; Kardakari, 2000), and Scolopax rusticola uses the wetland during the night (Kardakari, 2000), which could explain why we have not recorded these species. The absence of other migratory species from our list (Ciconia Ciconia, Ciconia negra, Gelochelidon nilotica, Glareola pratincole, Arenaria interpres), could be partly explained by the low frequency of our observations. Even though, we implemented frequent observations during March 2017, this only occurred for 2 weeks, and it is very likely that we have missed species which passed by the wetland during migration and stayed for only a short period of time. However, the absence of Hydrocoloeus minutus and the decline in the abundance of the six species of concern cannot be explained by the frequency of our observations.
A noticeable decline in the wintering population of several wildfowl species (except Anas platyrhynchos), herons, and the coot, as well as in the migrating populations of waders was also evident when compared to Kardakari (2000). Another noticeable difference was the status of flamingos. In Kardakari (2000), the species is referenced as “regular passage spring migrant and winter visitor in small numbers,” but our observations show that at present it is a common winter visitor in moderate numbers. Possible explanations such as declines and shifts due to climate change (Lehikoinen et al., 2013; Adam et al., 2015; Ramírez et al., 2018) or/and habitat degradation (Wang et al., 2011; Tavares et al., 2015; Brandis et al., 2018) which have been observed in other areas worldwide, should not be ruled out and further investigation will be needed to better understand these differences, which is out of the scope of this study.
The characterization of the area as an IBA and SPA (under the EUs Birds Directive)—also as an SCI and SAC (under the EUs Habitats Directive)—in the early 2000s, provided an adequate framework for site managers to take concrete actions for nature conservation and waterbirds protection, but few actions have been taken so far (Maneas et al., 2019). Instead, the area has been under unstable management for several years (Maneas et al., 2019), and the lack of conservation actions has added pressure to waterbirds survival at a national, European and International level. As analyzed above, compared to previous IBA evaluation (BirdLife International, 2001), our results suggest a dramatic decline in species abundance for 66.6% of the wintering IBA species, and for 100% of the migrant IBA species in Gialova Lagoon wetland, an outcome which should not be overlooked by the site managers.
Despite being an IBA and part of the Natura 2000 network, the protection status of the GLw could be further enhanced. Similar to previous evaluation (BirdLife International, 2001), our results indicate that the area cannot meet Ramsar criterion 5, which is to “regularly support 20,000 or more waterbirds” (Ramsar Sites Criteria, 2020), as the maximum abundance of waterbirds observed under this study was 2,912 in January 2017. The observed decrease of waterbirds abundance during the period of this study could imply that the area may no longer fulfill criterion 6, which is to “regularly support 1% of the individuals in a population of one species or subspecies of waterbirds” (Ramsar Sites Criteria, 2020), and equivalent IBA criterion A4i (BirdLife International, 2001). However, as mentioned above, our observations during the migration periods gave a picture of the species abundance on the day of the census, and the total abundance is multiple. Thus, we cannot make estimates based on the type of our monitoring approach, and further investigation is needed.
On the other hand, our results have confirmed the importance of the GLw-Natura2000 as an important stopover area for many waterbirds (including several threatened species), and thus we suggest that the area continue to meet Ramsar criterion 4, which is to “support animal species at a critical stage in their life cycle, or provide refuge during adverse conditions” (Ramsar Sites Criteria, 2020). Apart from an important stopover area, the GLw_Natura2000 site most likely provides a refuge during adverse conditions. As previously discussed, very cold winters in higher latitudes could force birds to move further south to find open water (Adam et al., 2015), and there was a strong indication that the area was increasingly used by waterbirds in January 2017, when exceptionally cold and snowy conditions occurred in Eastern Europe and the Balkan peninsula (European Cold Wave, 2017). Furthermore, our monitoring revealed that during the Spring and Autumn migration periods, the GLw-Natura2000 area supported several bird species which are threatened at an International (2 VU, 6 NT), European (8 VU, 3 NT) and national (2 CR, 6 EN, 14 VU, 4 NT) level, and it is already known of supporting the only European population of the CR African Chameleon (Legakis and Maragou, 2009). To that end, the area meets Ramsar criterion 2, which is to “support vulnerable, endangered or critically endangered species” (Ramsar Sites Criteria, 2020), and we suggest that it should be re-evaluated as to become a “Site of international importance for conserving biological diversity,” under the Ramsar Convention.
Moreover, placing it in a larger regional context, it can be stated to be one of the few remaining wetland IBA’s in south-western Greece (Hellenic Ornithological Society [HOS], 2019a), along the Mediterranean/Black Sea Flyway, an internationally important migration route (BirdLife International, 2017). Recent studies reveal that due to climate change, the distances that long-distance migrants will need to travel between suitable breeding and non-breeding habitats will significantly increase, and this increase in distance will require increase in refueling stopovers (Howard et al., 2018). It should be noted that the total abundance of waterbirds that use the wetland during migration is not known as their arrivals and departures are frequent and irregular. Our observations during the migration periods gave a picture of the number of individuals that have stopped in the wetland on the day of the census, but the total abundance during each migration period is multiple. If this wetland is to be further degraded or completely dried, the next closest wetland along this route is 110 km further north (Hellenic Ornithological Society [HOS], 2019a), making the GLw-Natura2000 area a very important node in the connectivity of wetlands (Smith and Chow-Fraser, 2010).
During the last 70 years, an annual water deficit of 200 mm per year and limited fresh water inputs, due to man-made constructions and increased irrigation needs, have led to increased salinity values and extensive vegetation mortality of habitats suitable for waterbirds especially at the S. Wetland side (Maneas et al., 2019). Apart from limitations in vegetation expansion, salinity values are critical for benthic communities (Newton et al., 2014) and fish (Zoulias et al., 2017), which are vital in the food chain for several waterbirds. At present, fresh water availability is affected by inland human activities, and the wetland is currently lacking fresh water inputs (Maneas et al., 2019; Manzoni et al., 2020). Waste waters from the olive-oil industry pollute surface water bodies (which could flow into the wetland, but currently flow into the sea), while increased irrigation needs in agriculture, and water demand for domestic use limit the amount of available groundwater resources during summer (Maneas et al., 2019). Unless freshwater inputs are enhanced by restoring hydrologic connectivity between the lagoon and the surrounding freshwater bodies, under future drier and warmer conditions, salinity in the lagoon is expected to increase (Manzoni et al., 2020).
The restoration of fresh water flows, could improve the status of nesting and feeding habitats enhancing FCS for several species (Stanley et al., 1977–1994; Kardakari, 2000; Bousbouras et al., 2011). For instance, if the water level at the S. Wetland sub-area could be kept at high levels until April/May (followed by level decreasing trends to avoid flooding of the nests), this could improve nesting habitats of existing nesting species such as the Mallard (Anas platyrynchos), and the Black winged stilt (Himantopus himantopus). Such management efforts could re-establish favorable nesting conditions for threatened species such as the vulnerable Garganey (Spatula querquedula), which used to nest at fresh water habitats in the past (Kardakari, 2000), and create favorable conditions for several waders including the Wood sandpiper (Tringa glareola) and the Marsh sandpiper (Tringa stagnatilis), two out of eight IBA characterization species. Less saline water conditions could lead to reed expansion (Álvarez-Rogel et al., 2007), which is the basic nesting habitat for other threatened species such as the nationally EN Purple Heron (Ardea purpurea) and the Ferruginous Duck (Aythya nyroca), which is considered VU at a European and NT at an International level (Legakis and Maragou, 2009; BirdLife International, 2015; IUCN, 2020). Increased water level during migration, could help to preserve important habitats for Collared pratincole (Glareola pratincola), Wood and Marsh sandpipers, and the critically endangered Glossy ibis (Plegadis falcinellus).
Our field observations revealed that the areas located at the wetland’s fringe support several endangered waterbirds, but at present these areas are the ones most threatened by human activities. In the existing agricultural areas, plots with horticulture—which were used during the Wintering period by the nationally VU Ardea alba—are gradually replaced by olive cultivations (Maneas et al., 2019). In the past, management efforts have led to conflicts between farmers and Natura 2000 managers (Hellenic Ornithological Society, 2000), and due to poor enforcement of guidelines inside the GLw-Natura2000 area, parts of the N. Wetland sub-area—which hosts the nationally CR Glossy ibis (Plegadis falcinellus)—have been gradually transformed to cultivated land (Maneas et al., 2019). Unless the degradation of these sub-areas is halted, by engaging the local farmers to strategic decisions about the management of the GLW-Natura2000 area, it is very likely that the numbers of these species will continue to decline.
Human disturbance has been found to be negatively linked to waders’ conservation as well (Tavares et al., 2015). In our case study, during late spring and summer we noticed that human disturbance from uncontrolled parking and motorcycle activity at the NW-area, the basic nesting habitat for the Kentish plover (Kardakari, 2000), has forced the species to search for other nesting areas, which were not ideal breeding areas. In fact, the species tried to nest in a similar soil habitat at the S. Wetland sub-area, but the nests were destroyed when the area was flooded after a rain event. Information signs and fencing for protecting the NW-area habitat at the north-western side of the wetland, could improve the conditions for Kentish plover, create favorable conditions for other waders to nest, and at the same time increase visitors’ awareness.
Another critical issue for the conservation of waterbirds is the delineation of the protected zone (Hellenic Ornithological Society, 2000). The perimetrical ditches (man-made constructions from the 60s) act well as physical boarders for the delineation of the protected wetland area. However, the Tyflomitis artesian springs area is outside the boarders of the protected area zone. Since this area, is at present the only provider of surface freshwater to the wetland (Maneas et al., 2019), our suggestion is that it should be added as a protected wetland area.
Apart from a hot-spot for birds (Kardakari, 2000), the GLw-Natura 2000 area has an important fisheries value (Koutsoubas et al., 2000; Zoulias et al., 2017), and almost 20% of the area is used for cultivations (Maneas et al., 2019). Water related issues are perceived in different ways by the different stakeholders (Maniatakou, 2020), and water management should consider these needs as well. Agriculture is a basic economic activity at the surrounding areas, and tourism is growing fast (Collaborative Land-Sea Integration Platform [COASTAL], 2019).
The development of alternative forms of tourism on site and around it (e.g., eco-tourism) could enhance the bonds of the diverse socio-ecological system and support conservation actions. For instance, in the surroundings, farms which are under organic cultivation support many bird species which could complement the bird-watching activity inside the wetland (Myers et al., 2019). Bird-monitoring projects have been among the most successful at integrating citizens in collecting data (McCaffrey, 2005), leading to successful long-term monitoring projects in Greece (Hellenic Ornithological Society [HOS], 2019c) and worldwide (Chandler et al., 2017). However, these activities could also disturb wildlife (Cardoni et al., 2008; McFadden et al., 2017), and they need to be organized carefully (for example not during Breeding/Nesting period). The example from a similar wetland area in Israel, Collins-Kreiner et al. (2013) has shown that as the number of visitors increased, the number of birds decreased. To that end, buffer zones need to be carefully designed and be appropriate both in terms of social and ecological perspectives (Glover et al., 2011).
The GLw-Natura2000 area has good accessibility, a road taking visitors around and into the lagoon, and the relatively small sized area makes it possible to see many habitats and a large diversity of birds at close distance, without too much effort. Such outdoor activities can increase support for wildlife conservation, and enhance awareness among locals and visitors. Spatial and temporal data from this study could be used as a basis for organizing sustainable bird-watching activities and organized school visits on site. The fact that species richness and abundance is higher from October to April, could prolong the touristic season and attract visitors outside the high touristic season (May–September), adding to the local economy. Income from eco-tourism could both be an income for the local community creating a positive attitude to conservation, and also for funding some of the conservation efforts.
A sustainable management strategy for the GLw area should aim to improve and enlarge waterbirds’ habitats, considering nonetheless the existing human activities and social needs at a broader scale (Habitats Directive 92/43/EEC, 1992). Such management requires not only interdisciplinary research, but also engagement of stakeholders at a broader scale (Tsianou et al., 2013). An Ecosystem Services approach could provide the links between nature and people (Díaz et al., 2015), which in turn could reduce conflicts and enhance the benefits for the local economy and ecology, by improving the mutual understanding between the stakeholders. Bringing together farmers, fishermen, tourism operators, Natura 2000 managers and policy makers could lead to improved land-sea interactions and widely accepted management strategies (Collaborative Land-Sea Integration Platform [COASTAL], 2019).
The Gialova Lagoon wetland supports many different bird species (149 species including 66 waterbirds and 15 wetland-dependent species), especially during the Wintering and both migration periods. The distribution of species richness and abundance varied significantly in the different sub-areas (formed after past anthropogenic interventions). The S. Wetland sub-area was the area mostly used by waterbirds, followed by the Lagoon and the NW area. Our results indicate that the populations of most IBA species have declined over the last 20 years. The current lack of management makes it difficult to implement efficient and inclusive conservation strategies. The restoration of fresh water inflows, could improve habitats and water conditions for the IUCN and the IBA/SPA species and gradually enhance their conservation status. With careful steps and management decisions the area could become a good example for sustainable management of multifunctional coastal wetlands, favoring both nature conservation and societal well-being. The area cannot meet Ramsar criterion 5 linked to waterbirds, and the decline in abundance of several species may imply that no longer fulfills criterion 6 (and equivalent IBA criterion A4i). However, our results indicate that the area meets criterion 4 and criterion 2, and thus we suggest that it should be further investigated and evaluated to potentially become the eleventh Greek Ramsar site.
The primary dataset (monthly observations of waterbirds) is archived in the open-access database of the Bolin Centre for Climate Research. Available at: https://bolin.su.se/data/maneas-2020.
GM, DB, and HB: conceptualization, design of the study, database, data processing, and methodology. GM, DB, and VN: fieldwork and investigation. HB: supervision. GM: writing—original draft preparation. All authors wrote sections of the manuscript, contributed to manuscript revision, read and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors acknowledge support from the Navarino Environmental Observatory (NEO), a partnership between Stockholm University, the Biomedical Research Foundation of the Academy of Athens (BRFAA) and TEMES S.A. (www.navarinoneo.gr). GM and HB acknowledge COASTAL-Collaborative Land-Sea Integration Platform. European Union’s H2020 Research and Innovation Programme under Grant Agreement No 773782.
We would like to thank Jan Sundberg, Javier Martínez-López, Stefano Manzoni, and Anna Scaini for their valuable comments and fruitful discussions, and Eirini Makopoulou for preparing Figure 1. Valuable comments were provided by two reviewers.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.501548/full#supplementary-material
Adam, M., Musilová, Z., Musil, P., Zouhar, J., and Romportl, D. (2015). Long-term changes in habitat selection of wintering waterbirds: high importance of cold weather refuge sites. Acta Ornithol. 50, 127–138. doi: 10.3161/00016454ao2015.50.2.001
Álvarez-Rogel, J., Jiménez-Cárceles, F. J., Roca, M. J., and Ortiz, R. (2007). Changes in soils and vegetation in a Mediterranean coastal salt marsh impacted by human activities. Estuar. Coast. Shelf Sci. 73, 510–526. doi: 10.1016/j.ecss.2007.02.018
Arvanitidis, C., Koutsoubas, D., Dounas, C., and Eleftheriou, A (1999). Annelid fauna of a Mediterranean lagoon (Gialova Lagoon, south-west Greece): community structure in a severely fluctuating environment. J. Mar. Biol. Assoc. U. K. 79, 849–856. doi: 10.1017/S0025315499001010
Bibi, F., and Ali, Z. (2013). Measurement of diversity indices of avian communities at Taunsa Barrage Wildlife Sanctuary, Pakistan. J. Anim. Plant Sci. 23, 469–474.
BirdLife International (2001). Important Bird Areas and Potential Ramsar Sites in Europe. Wageningen: BirdLife International.
BirdLife International (2015). European Red List of Birds. Luxembourg: Publications Office of the European Union. Available online at: https://ec.europa.eu/environment/nature/conservation/species/redlist/downloads/European_birds.pdf (accessed September 10, 2019).
BirdLife International (2017). Mediterranean/Black Sea Flyway. Available online at: http://datazone.birdlife.org/userfiles/file/sowb/flyways/5_Mediterranean_Black_Sea_Factsheet.pdf (accessed 04 November, 2017).
BirdLife International (2018). The Top Five Threats to Birds may Surprise You. Available online at: https://www.birdlife.org/worldwide/news/top-five-threats-birds-may-surprise-you (accessed 22 July, 2019).
BirdLife International (2019). BirdLife Taxonomic Basis. Available online at: http://datazone.birdlife.org/species/taxonomy (accessed 27 September, 2019).
BirdLife International (2020). Available online at: http://datazone.birdlife.org/site/ibacriteria (accessed October, 2020).
Birds Directive 2009/147/EC (2009). The European Union Birds Directive. Available online at: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32009L0147 (accessed November 2, 2020).
Bortels, L., Chan, J. C. W., Merken, R., and Koedam, N. (2011). Long-term monitoring of wetlands along the Western-Greek bird migration route using Landsat and ASTER satellite images: Amvrakikos Gulf (Greece). J. Nat. Conserv. 19, 215–223. doi: 10.1016/j.jnc.2011.01.004
Bousbouras, D., Maneas, G., Bilionis, S., and Euaggelopoulos, A. (2011). Gialova Lagoon Hydrological Study. Determining the Necessary Management Measures for Achieving a Favourable Conservation Status for Birds, Athens: Hellenic Ornithological Society, 212. (Available only in Greek).
Brandis, K. J., Bino, G., Spencer, J. A., Ramp, D., and Kingsford, R. T. (2018). Decline in colonial waterbird breeding highlights loss of Ramsar wetland function. Biol. Conserv. 225, 22–30. doi: 10.1016/J.BIOCON.2018.06.022
Cardoni, D. A., Favero, M., and Isacch, J. P. (2008). Recreational activities affecting the habitat use by birds in Pampa’s wetlands, Argentina: implications for waterbird conservation. Biol. Conserv. 141, 797–806. doi: 10.1016/j.biocon.2007.12.024
Chandler, M., See, L., Copas, K., Bonde, A. M. Z., López, B. C., Danielsen, F., et al. (2017). Contribution of citizen science towards international biodiversity monitoring. Biol. Conserv. 213(Pt B), 280–294. doi: 10.1016/j.biocon.2016.09.004
Collaborative Land-Sea Integration Platform [COASTAL], 2019 (2019). European Union’s H2020 Research and Innovation Programme Under Grant Agreement No. 773782. Available online at: https://h2020-coastal.eu/ (accessed 03 February, 2019).
Collins-Kreiner, N., Malkinson, D., Labinger, Z., and Shtainvarz, R. (2013). Are birders good for birds? Bird conservation through tourism management in the Hula Valley, Israel. Tour. Manag. 38, 31–42. doi: 10.1016/j.tourman.2013.01.009
Díaz, S., Demissew, S., Carabias, J., Joly, C., Lonsdale, M., Ash, N., et al. (2015). The IPBES conceptual framework – connecting nature and people. Curr. Opin. Environ. Sustain. 14, 1–16. doi: 10.1016/j.cosust.2014.11.002
European Cold Wave (2017). Wikipedia Results on the European Cold Wave Occurred in 2017. Available online at: https://en.wikipedia.org/wiki/January_2017_European_cold_wave (accessed 24 November, 2020).
European Commission (2016). Wild Birds: Threatened Bird Species in Annex I. Environment, European Commission. Available online at: http://ec.europa.eu/environment/nature/conservation/wildbirds/threatened/index_en.htm (accessed 19 June, 2019).
Fischer, J., and Lindenmayer, D. B. (2002). Small patches can be valuable for biodiversity conservation: two case studies on birds in southeastern Australia. Biol. Conserv. 106, 129–136. doi: 10.1016/S0006-3207(01)00241-5
Fric, J., Portolou, D., Manolopoulos, A., and Kastritis, T. (2012). Important Areas for Seabirds in Greece. Technical Report No. LIFE07 NAT/GR000285. Athens: Hellenic Ornithological Society (HOS / BirdLife Greece).
Glover, H. K., Weston, M. A., Maguire, G. S., Miller, K. K., and Christie, B. A. (2011). Towards ecologically meaningful and socially acceptable buffers: response distances of shorebirds in Victoria, Australia, to human disturbance. Landsc. Urban Plan. 103, 326–334. doi: 10.1016/j.landurbplan.2011.08.006
GR2550004 (1995). NATURA 2000 – Standard Data Form for Special Protection Areas (SPA), Proposed Sites for Community Importance (pSCI), Sites of Community Importance (SCI) and for Special Areas of Conservation (SAC). Available online at: https://natura2000.eea.europa.eu/natura2000/SDF.aspx?site=GR2550004 (accessed November 2, 2020).
GR2550008 (2001). NATURA 2000 – Standard Data Form for Special Protection Areas (SPA), Proposed Sites for Community Importance (pSCI), Sites of Community Importance (SCI) and for Special Areas of Conservation (SAC). Available online at: https://natura2000.eea.europa.eu/Natura2000/SDF.aspx?site=GR2550008 (accessed November 22, 2018).
Greek Biotope Wetland Centre (GBWC) (2020). Available online at: http://www.ekby.gr/ekby/en/EKBY_Greek_Wetlands_en.html (accessed November 2, 2020).
Greek Ramsar Sites (2020). Available online at: https://www.ramsar.org/wetland/greece (accessed November 2, 2020).
Gregory, R. D., Gibbons, D. W., and Donald, P. F. (2004). “Bird census and survey techniques,” in Bird Ecology and Conservation: A Handbook of Techniques, Vol. 6, eds W. J. Sutherland, I. Newton, and R. Green (Oxford: Oxford University Press), 17–55. doi: 10.1093/acprof:oso/9780198520863.001.0001
Habitats Directive 92/43/EEC (1992). The European Union Habitats Directive. Available online at: https://ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm (accessed September 2, 2019).
Handrinos, G., Kazantzidis, S., Alivizatos, C., Akriotis, T., and Portolou, D. (2015). International Waterbird Census in Greece (1968 – 2006). Analysis of the Populations of Wildfowl (Anseriformes) and the Coot (Fulica atra). Athens: Hellenic Ornithological Society – Hellenic Bird Ringing Centre.
Heath, M. F., Evans, M. I., Hoccom, D. G., Payne, A. J., and Peet, N. B. (eds) (2000). Important Bird Areas in Europe: Priority Sites for Conservation. Volume 1: Northern Europe, Volume 2: Southern Europe. Cambridge: BirdLife International.
Hellenic forestry (2020). Birds’ Migratory Routes in Greece. Available online at: https://dasarxeio.com/2013/11/24/9770/ (accessed March 02, 2020).
Hellenic Ornithological Society (2000). Special Environmental Study of Gialova Lagoon and Sfaktiria Island, Implemented Under by EU Project (B 4-3200/97/244): LIFE-Nature Implementation of Management Plans for Pylos Lagoon and Evrotas Delta, Natura 2000 Sites, Greece. Athens: Hellenic Ornithological Society. (Available in Greek).
Hellenic Ornithological Society (HOS) (2019b). GR119 Divari Pilou Lagoon (Gialova). Available online at: http://www.ornithologiki.gr/page_iba.php?aID=119&loc=en (accessed June 16, 2019).
Hellenic Ornithological Society (HOS) (2019c). Voluntary Projects. Available online at: http://www.ornithologiki.gr/volunteer.php (accessed June 16, 2019).
Hellenic Ornithological Society (HOS) (2019a). List of the Important Bird Areas of Greece. Available online at: http://www.ornithologiki.gr/page_cn.php?tID=1866&aID=898 (accessed June 16, 2019).
Howard, C., Stephens, P. A., Tobias, J. A., Sheard, C., Butchart, S. H. M., and Willis, S. G. (2018). Flight range, fuel load and the impact of climate change on the journeys of migrant birds. Proc. R. Soc. B Biol. Sci. 285:20172329. doi: 10.1098/rspb.2017.2329
Hutcheson, K. (1970). A test for comparing diversities based on the Shannon formula. J. Theor. Biol. 29, 151–154. doi: 10.1016/0022-5193(70)90124-4
IBA Criteria (2001). Identifying Important Bird Areas. Available online at: http://datazone.birdlife.org/userfiles/file/IBAs/EuCntryPDFs/IBA2000Vol1pp011-020_IdentifyingIBAs.pdf (accessed November 23, 2020).
Issa, M. A. A. (2019). Diversity and abundance of wild birds species’ in two different habitats at Sharkia Governorate, Egypt. J. Basic Appl. Zool. 80:34. doi: 10.1186/s41936-019-0103-5
IUCN (2020). The IUCN Red List of Threatened Species. Available online at: https://www.iucnredlist.org/ (accessed November 2, 2020).
Kardakari, N. (2000). The Birds of Gialova Lagoon: A Report on the Avifauna of the Important Bird Area: Divari Pilou Lagoon (Gialova). Athens: Hellenic Ornithological Society. (Available only in Greek).
Koutsoubas, D., Dounas, C., Arvanitidis, C., Kornilios, S., Petihakis, G., Triantafyllou, G., et al. (2000). Macrobenthic community structure and disturbance assessment in Gialova Lagoon, Ionian Sea. ICES J. Mar. Sci. 57, 1472–1480. doi: 10.1006/jmsc.2000.0905
Legakis, A., and Maragou, P. (2009). The Red Book of Endangered Species in Greece (in Greek). Athens: Hellenic Zoological Society, 1–528. Available in Greek.
Lehikoinen, A., Jaatinen, K., Vähätalo, A. V., Clausen, P., Crowe, O., Deceuninck, B., et al. (2013). Rapid climate driven shifts in wintering distributions of three common waterbird species. Glob. Chang. Biol. 19, 2071–2081. doi: 10.1111/gcb.12200
Liordos, V., Pergantis, F., Perganti, I., and Roussopoulos, Y. (2014). Long-term population trends reveal increasing importance of a Mediterranean wetland complex (Messolonghi lagoons, Greece) for wintering waterbirds. Zool. Stud. 53:12. doi: 10.1186/1810-522X-53-12
Maneas, G., Makopoulou, E., Bousbouras, D., Berg, H., and Manzoni, S. (2019). Anthropogenic changes in a Mediterranean coastal wetland during the last century-the case of Gialova Lagoon, Messinia, Greece. Water 11:350. doi: 10.3390/w11020350
Maniatakou, S., Berg, H., Maneas, G., and Daw, T. M. (2020). Unravelling diverse values of ecosystem services: a socio-cultural valuation using Q methodology in Messenia, Greece. Sustainability 12:10320. doi: 10.3390/su122410320
Manzoni, S., Maneas, G., Scaini, A., Psiloglou, B. E., Destouni, G., and Lyon, S. W. (2020). Understanding coastal wetland conditions and futures by closing their hydrologic balance: the case of Gialova Lagoon, Greece. Hydrol. Earth Syst. Sci. 24, 3557–3571. doi: 10.5194/hess-24-3557-2020
Maragou, P., and Mantziou, D. (2000). Assessment of the Greek Ramsar Wetlands. Lempesi: WWF-Greece, 59 + Answered questionnaires pp 118.
McArthur, V. E., Koutsoubas, D., Lampadariou, N., and Dounas, C. (2000). The meiofaunal community structure of a Mediterranean lagoon (Gialova lagoon, Ionian Sea). Helgoland Mar. Res. 54, 7–17. doi: 10.1007/s101520050031
McFadden, T. N., Alejandro, G. H., and Navedo, J. G. (2017). Waterbird responses to regular passage of a birdwatching tour boat: implications for wetland management. J. Nat. Conserv. 40, 42–48. doi: 10.1016/j.jnc.2017.09.004
Moore, F. R., Woodrey, M. S., Buler, J. J., Woltmann, S., and Simons, T. R. (2005). “Understanding the stopover of migratory birds: a scale dependent approach,” in Bird Conservation Implementation and Integration in the Americas: Proceedings of the 3rd International Partners in Flight Conference: USDA Forest Service Gen. Tech. Rep. No., Vol. 2013, eds C. J. Ralph and T. Rich (Asilomar, CA: USDA Forest Service), 684–689.
Myers, D., Berg, H., and Maneas, G. (2019). Comparing the soundscapes of organic and conventional olive groves: a potential method for bird diversity monitoring. Ecol. Indic. 103, 642–649. doi: 10.1016/j.ecolind.2019.04.030
Newton, A., Icely, J., Cristina, S., Brito, A., Cardoso, A. C., Colijn, F., et al. (2014). An overview of ecological status, vulnerability and future perspectives of European large shallow, semi-enclosed coastal systems, lagoons and transitional waters. Estuar. Coast. Shelf Sci. 140, 95–122. doi: 10.1016/j.ecss.2013.05.023
O’Hara, R. B., and Kotze, D. J. (2010). Do not log-transform count data. Method. Ecol. Evol. 1, 118–122. doi: 10.1111/j.2041-210x.2010.00021.x
Ramírez, F., Rodríguez, C., Seoane, J., Figuerola, J., and Bustamante, J. (2018). How will climate change affect endangered Mediterranean waterbirds? PLoS One 13:e0192702. doi: 10.1371/journal.pone.0192702
Ramsar (2017). Ramsar Sites Information Service. Available online at: https://rsis.ramsar.org (accessed November 16, 2017).
Ramsar convention (1994). Convention on Wetlands of International Importance Especially as Waterfowl Habitat 1971. Available online at: http://portal.unesco.org/en/ev.php-URL_ID=15398&URL_DO=DO_TOPIC&URL_SECTION=201.html (accessed June 19, 2019).
Ramsar Sites Criteria (2020). The Ramsar Sites Criteria: The Nine Criteria for Identifying Wetlands of International Importance. Available online at: https://www.ramsar.org/sites/default/files/documents/library/ramsarsites_criteria_eng.pdf (accessed November 2, 2020).
Ramsar Sites Information Service (RSIS) (2020). Available online at: https://rsis.ramsar.org/?pagetab=2 (accessed November 2, 2020).
Seavy, N. E., Quader, S., Alexander, J. D., and Ralph, C. J. (2005). “Generalized linear models and point count data: statistical considerations for the design and analysis of monitoring studies,” in Proceedings of the 3rd International Partners in Flight Conference. 2002 March 20-24; Asilomar, California: Bird Conservation Implementation and Integration in the Americas. USDA For. Ser. Gen. Tech. Rep. PSW-GTR-191, Vol. 2, eds C. J. Ralph and T. D. Rich (Albany, CA: United States Department of Agriculture), 744–753.
Smith, L. A., and Chow-Fraser, P. (2010). Implications of the species-area relationship on sampling effort for marsh birds in southern Ontario. Wetlands 30, 553–563. doi: 10.1007/s13157-010-0048-4
Stanley, C., Simmons, K. E. L., and Perrins, C. M. (1977–1994). Handbook of the Birds of Europe the Middle East and North Africa, The Birds of the Western Palearctic, Vols. I–IX. Oxford: Oxford University Press.
Svensson, L., Mullarney, K., and Zetterström, D. (2010). Collins Bird Guide, 2nd Edn. London: HarperCollins Publishers Ltd.
Tavares, D. C., Guadagnin, D. L., de Moura, J. F., Siciliano, S., and Merico, A. (2015). Environmental and anthropogenic factors structuring waterbird habitats of tropical coastal lagoons: implications for management. Biol. Conserv. 186: 12–21. doi: 10.1016/j.biocon.2015.02.027
Tsianou, M. A., Mazaris, A. D., Kallimanis, A. S., Deligioridi, P. S. K., Apostolopoulou, E., and Pantis, J. D. (2013). identifying the criteria underlying the political decision for the prioritization of the Greek Natura 2000 conservation network. Biol. Conserv. 166, 103–110. doi: 10.1016/j.biocon.2013.06.021
Wang, Z. Q., Wang, F., and Baiyan, W. (2011). Changes of the wetland landscape and the consequent impacts on the waterbirds in Western Songnen Plain. Procedia Environ. Sci. 10(Pt C), 2608–2613. doi: 10.1016/j.proenv.2011.09.405
Warnock, N. (2010). Stopping vs. Staging: the difference between a hop and a jump. J. Avian Biol. 41, 621–626. doi: 10.1111/j.1600-048X.2010.05155.x
Warton, D. I, Lyons, M., Stoklosa, J., and Ives, A. R. (2016). Three points to consider when choosing a LM or GLM test for count data. Method. Ecol. Evol. 7, 882–890. doi: 10.1111/2041-210X.12552
Wetlands International (2020). Waterbird Population Estimates. Available online at: wpe.wetlands.org (accessed November 08, 2020).
Keywords: waterbirds, coastal wetland, Natura 2000, management suggestions, conservation needs
Citation: Maneas G, Bousbouras D, Norrby V and Berg H (2020) Status and Distribution of Waterbirds in a Natura 2000 Area: The Case of Gialova Lagoon, Messinia, Greece. Front. Ecol. Evol. 8:501548. doi: 10.3389/fevo.2020.501548
Received: 30 September 2019; Accepted: 30 November 2020;
Published: 22 December 2020.
Edited by:
Angel Pérez-Ruzafa, University of Murcia, SpainReviewed by:
Stamatis Zogaris, Hellenic Centre for Marine Research (HCMR), GreeceCopyright © 2020 Maneas, Bousbouras, Norrby and Berg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giorgos Maneas, Z2lvcmdvcy5tYW5lYXNAbmF0Z2VvLnN1LnNl
†ORCID: Giorgos Maneas, orcid.org/0000-0001-7152-669X; Håkan Berg, orcid.org/0000-0003-3260-9710
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.