 James A. Cox
James A. Cox R. Todd Engstrom
R. Todd Engstrom David R. Breininger
David R. Breininger Erin L. Hewett Ragheb
Erin L. Hewett Ragheb- 1Stoddard Bird Lab, Tall Timbers Research Station & Land Conservancy, Tallahassee, FL, United States
- 2Department of Biological Science, Florida State University, Tallahassee, FL, United States
- 3NASA Ecological Monitoring Program, IMSS-300, John F. Kennedy Space Center, IMSS-300, Merritt Island, FL, United States
- 4Department of Biology, University of Central Florida, Orlando, FL, United States
- 5Independent Researcher, New Market, MD, United States
Recovery of listed species requires that land managers and research biologists work together to address the factors affecting population stability and growth. In Florida, an essential factor affecting rare species habitat quality and restoration is fire management. Fire plays an essential role in restoring and maintaining almost every upland ecosystem in Florida, but fires also have negative effects (e.g., mortality and displacement) that play out today within an altered landscape where rare species are often limited to small, fragmented areas and negative effects may be accentuated. Fire effects also are complex, not well studied experimentally, and likely to change as urbanization and global temperatures increase over coming decades. These conditions can create missteps in both fire research and fire management without regular communication between scientists and practitioners. We assessed the fire-related research associated with four federally listed birds in Florida: Cape Sable seaside sparrow (Ammodramus maritimus mirabilis), Florida grasshopper sparrow (Ammodramus savannarum floridanus), Florida scrub-jay (Aphelocoma coerulescens), and red-cockaded woodpecker (Dryobates borealis). Fire research has not addressed the needs of some of these species for starkly different reasons. Land managers, in turn, have not successfully applied the recommendations of fire research in other instances. Our results point to fire frequency as an important focus for practitioners managing habitat for rare species in Florida. Our review also suggests that successful integration of research and management will be best served when (1) ecological burning practices are used, (2) local fire management goals are prioritized annually, (3) instructional products are developed for managers, (4) land manager tenure is promoted, (5) stakeholders meet regularly, and (6) creative solutions are devised to overcome staff and equipment shortages.
Introduction
Science should permeate all aspects of the Endangered Species Act (ESA) from the methods used to identify imperiled taxa to the actions taken to lower their risk of extinction. The ESA was established to recover both rare species and the ecosystems upon which they depend (16 U.S.C. §1531(b). The United States Fish and Wildlife Service (USFWS), the agency responsible for conserving freshwater and terrestrial taxa, must base ESA listing decisions on the best scientific information available and may classify species and populations as threatened or endangered if the taxon’s existence is threatened by: (1) habitat destruction/modification, (2) commercial exploitation, (3) disease/predation, (4) regulatory inadequacies, or (5) other natural or manmade factors (U. S. Fish and Wildlife Service [USFWS], 1973). Once a taxon is listed, USFWS biologists collaborate with experts from governmental agencies, non-governmental organizations, and academia to develop a recovery plan that lays out steps needed to lower the risk of extinction. Recovery plans integrate the best scientific information regarding a taxon’s population status and life-history traits into a set of measurable criteria for assessing recovery.
While science plays a key role in both the listing process and the development of ESA recovery plans, the actions needed for recovery often occur in settings where uncertainty and urgency abound. Essential research on habitat requirements often does not commence until after a taxon is listed and rarely yields quick answers to the complex questions needed to manage and restore taxa effectively (much less the ecosystems they inhabit). Uncertainty and urgency are further accentuated when dealing with rare taxa that require habitat conditions created by storms, fires, and other seemingly destructive forces. Scientists are asked to provide habitat management recommendations that mimic these disturbance events (e.g., prescribed fire, mechanical removal of shrubs), but scientists often lack the long-term data they need to predict the possible outcomes their recommendations entail. Instead, scientists base recommendations on the natural timing and frequency of the disturbance, potential variation in timing and frequency across broad geographic areas, and ultimately the threat the disturbance might pose to the viability of a rare population.
Fire is essential for maintaining and restoring the habitat conditions needed by many rare species in Florida. Florida experiences some of the highest rates of lightning strikes in the western hemisphere (Duncan et al., 2010). Brevard County in central Florida receives over 22,000 direct strikes (ca. 1.2 lightning strikes per 20 ha) annually (National Oceanic and Atmospheric Administration [NOAA], 2019). When coupled with the distinct wet and dry seasons that have characterized Florida’s weather patterns for centuries, lightning-initiated fires regularly “managed” most habitat types in the state (Florida Natural Areas Inventory [FNAI], 2010). Frequent fire (every 2–10 years) promotes the vegetation structure that many rare species require (Breininger and Schmalzer, 1990; Jones et al., 2013), recycles nutrients (Venne et al., 2016), and may improve the quality of food resources (Lashley et al., 2015). In the case of ESA-listed species, however, fire may represent a double-edged sword because fire can result in direct mortality (Engstrom, 2010) at the same time that it is essential for sustaining or restoring suitable habitat (Steen et al., 2013). Recovery plans list habitat degradation brought about by the inappropriate use of fire as a threat to 57% of the terrestrial ESA species in Florida (n = 56).
Any uncertainties about how and when to use fire in Florida must be weighed in a landscape where rapid habitat loss often increases the urgency of action. Florida contains some of the earliest European settlements in North America, but frequent storms and hot summers suppressed significant human population growth until the early 1960s (Gannon, 1993). Since that time, Florida’s human population has skyrocketed to become the third largest in the nation (ca. 21.3 million). This growth resulted in the loss of an estimated 0.5 million ha of wildlife habitat between the late 1980s and 2003 (Kautz et al., 2007). The growth has also fragmented conservation lands and severely restricted the land base available for all rare species. At the same time, Florida state ranks seventh in the nation in terms of the number of rare plant and animal species it supports (n = 4468) and sixth in terms of the percentage (14.3%) of plants and animals at risk of extinction (Stein, 2002). These totals include irreplaceable elements of global biodiversity in the form of many species (n = 441) that occur only in Florida (Florida Natural Areas Inventory [FNAI], 2019).
We focus on efforts to translate fire-management research into practical habitat management and restoration on public lands for four ESA-listed avian taxa: Cape Sable seaside sparrow (Ammodramus maritimus mirabilis), Florida grasshopper sparrow (Ammodramus savannarum floridanus), Florida scrub-jay (Aphelocoma coerulescens), and red-cockaded woodpecker (Dryobates borealis). These birds are associated with biologically diverse habitats maintained with frequent fire but also vary in life-history traits, geographic distributions (Figure 1), fire needs, and the ease with which populations can be monitored and assessed. Conservation status of our focal taxa also ranges from critically endangered (seaside and grasshopper sparrows), to threatened and declining (scrub-jay), to endangered and increasing (red-cockaded woodpecker). The four taxa also serve as potential surrogates for broader conservation efforts focused on the unique habitats they each occupy (Simberloff, 1988; Breckheimer et al., 2014).
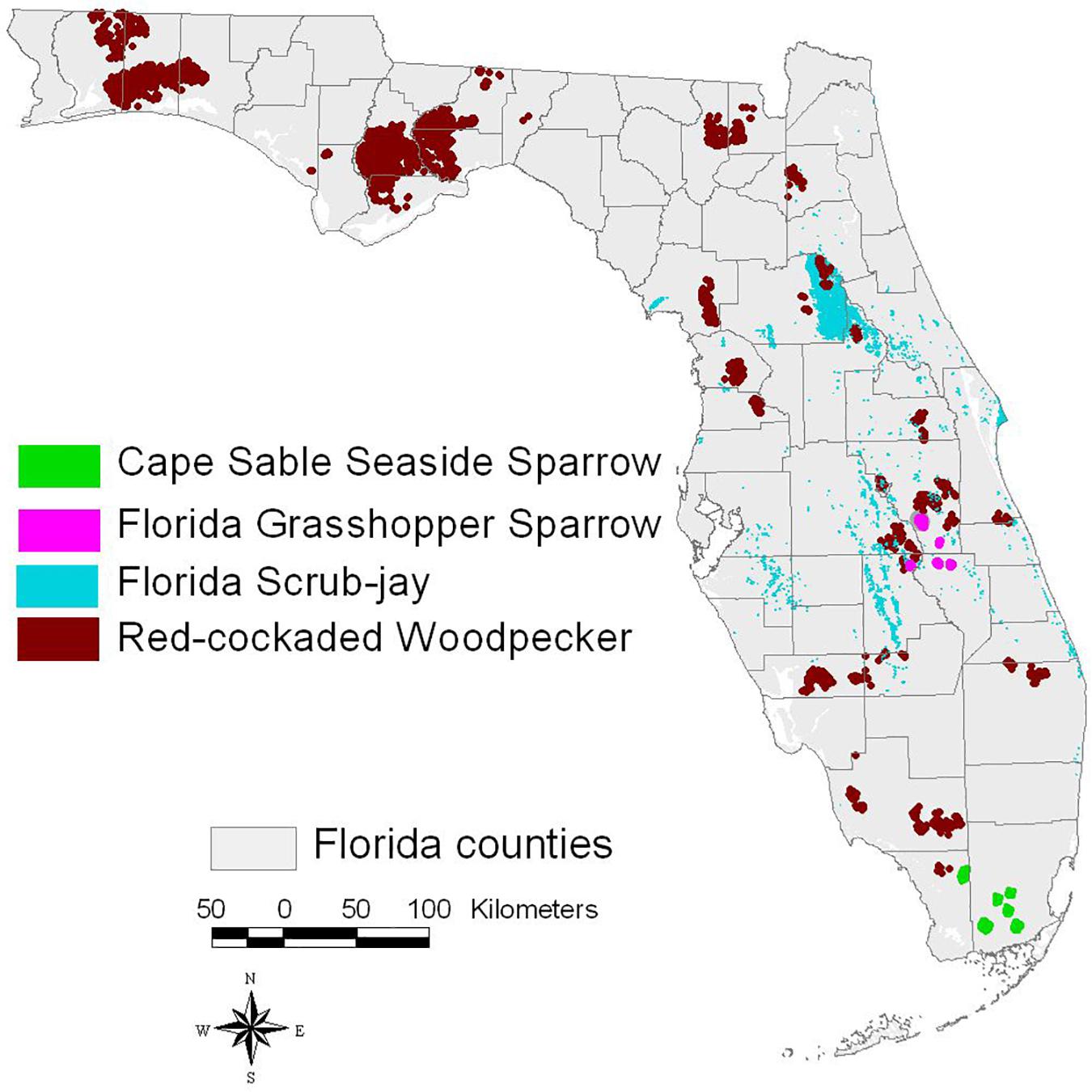
Figure 1. Distribution map for four focal taxa. The woodpecker is found in other states.
Although each species faces unique challenges, we address three common questions:
1. What research is available for promoting habitat restoration and fire management for these fire-dependent taxa?
2. How effectively have the results of fire-management research been provided to and adopted by land managers?
3. What are the major challenges for recovery, including the knowledge exchange between scientists and managers for habitat restoration using prescribed fire?
Our goals are to assess the status of fire-management research for each taxon and the role that science has played in guiding the fire management practitioners provide. We also evaluate the effects that different scientific perspectives concerning fire-management have had and conclude with recommendations for future fire research and management needs and how to improve the links between researchers and practitioners.
Materials and Methods
The fire-related information associated with each focal species was compiled using literature searches (Google Scholar and Web of Science, December 2019), personal interviews, and our many years of collective research in Florida (Breininger et al., 1991; Engstrom, 1993; Cox et al., 1994; Hewett Ragheb et al., 2019b). The woodpecker has the most extensive list of referenced studies (n = 513) followed by scrub-jay (n = 282), Florida grasshopper sparrow (n = 36), and Cape Sable seaside sparrow (n = 27). Similar queries based on the habitats associated with each species yielded similar rankings: 1,300, 815, 76, and 26 references for longleaf pine (woodpecker), Florida scrub (scrub-jay), Florida dry prairie (grasshopper sparrow), and Florida marl prairie (seaside sparrow), respectively. The red-cockaded woodpecker has the largest land base supported on public lands (Table 1) while the Florida scrub-jay is supported by the largest total number of managed areas (n = 198), including a high percentage (ca. 40%) of county and municipal managed properties.
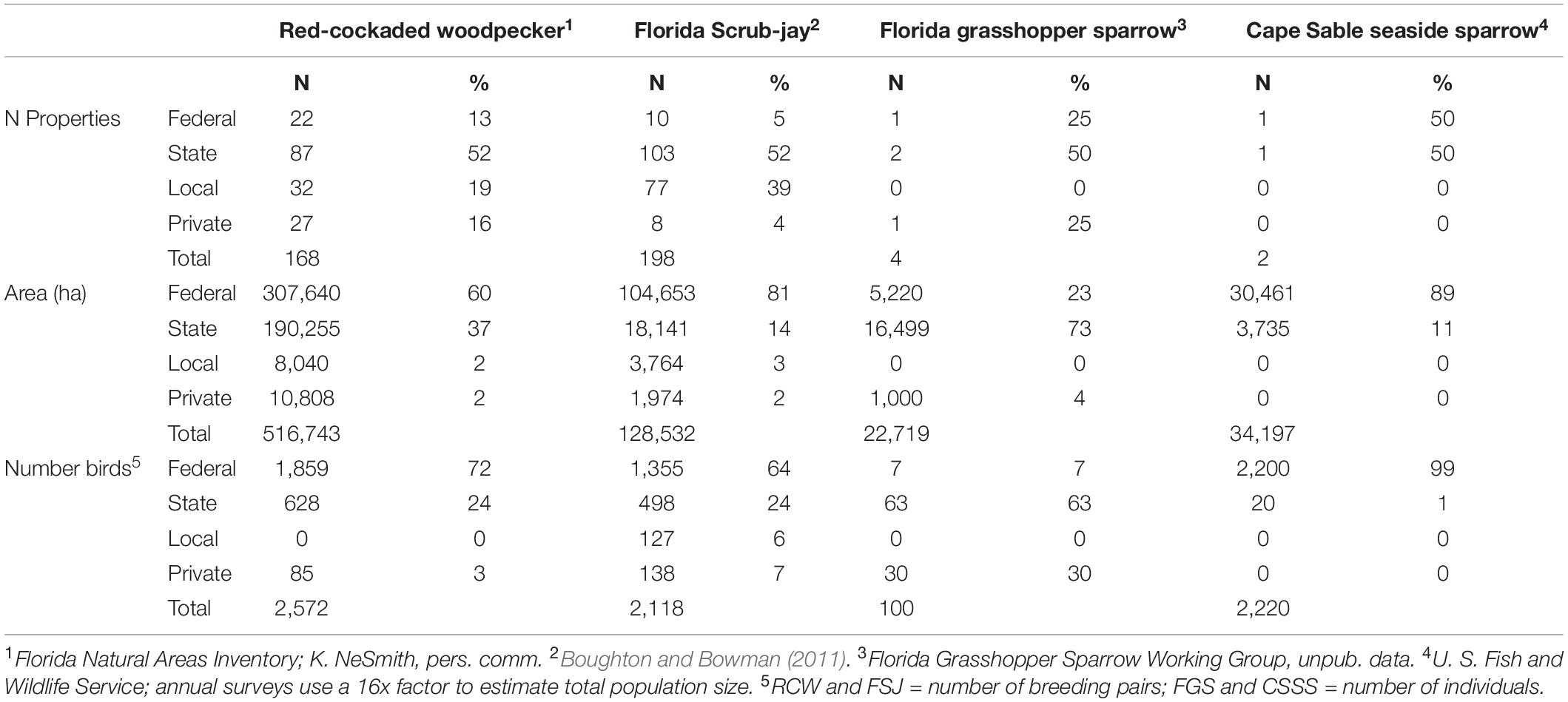
Table 1. Ownership, managed habitat area, and number of breeding pairs (or individuals) for four federally listed birds found in Florida.
The peer-reviewed literature on fire management has grown dramatically in recent years, but important information regarding fire effects and rare-species management also resides in government documents, grant reports, and other gray literature. For example, a Google search using the key words “fire management” and “Cape Sable seaside sparrow” yielded ca. 1,500 hits for our focal taxa with the fewest peer-reviewed articles. One item included was an interview with a federal fire manager that described a situation where fire helped to protect a seaside sparrow population. This interview provided a tangible benefit that fire might provide for the sparrow that was not documented in the peer-reviewed literature. Our aim is to include and summarize all available information on our focal taxa rather than restricting our search to peer-reviewed literature.
We also documented the flow of scientific information into fire management using interviews with scientists and land managers that work with our focal species coupled with results from a survey we distributed to research biologists and land managers that work closely with the Florida scrub-jay (see Supplementary Material for methods). Much like the red-cockaded woodpecker, which has been the subject of a separate survey (Weiss et al., 2019), the scrub-jay occurs on dozens of municipal, state, and federal managed properties with different management goals, administrative structures, and management resources. The scrub-jay questionnaire was distributed to scrub managers and scrub research biologists (n = 144) using a contact list maintained by the Florida Fish and Wildlife Conservation Commission (FFWCC) (A. Doyle, pers. comm.). Ten questions were presented (Supplementary Material) and answered by 47 respondents (ca. 33% of the distribution list). Land managers (55%) were the most common respondents followed by research biologists (19%), and land management administrators (15%). Most respondents (78%) were associated with municipal (30%), state (26%), or federal (21%) agencies (see Supplementary Material).
Results
Cape Sable Seaside Sparrow
The Cape Sable seaside sparrow (Figure 2) is restricted to 70 km2 (Figure 1) within Big Cypress National Preserve (BCNP; 26.05, −80.90; 2,900 km2), Everglades National Park (ENP; 25.75, −80.55; 20,201 km2), and the Southern Glades Wildlife and Environmental Area (SGWEA; 25.33, −80.52 12,300 ha). This disjunct, non-migratory subspecies occupies biologically diverse marl prairie communities where short periods of inundation (3–7 months annually) support low-statured bunch grasses (Figure 3). Interstitial patches of bare ground and wet, shallow depressions provide foraging substrates for sparrows (Hanan et al., 2010; Post and Greenlaw, 2018). Suitable vegetation structure is maintained through a complex interplay of fire, soil conditions, and hydrology that can yield markedly different plant communities with a mere 30 cm change in elevation (Sah et al., 2014).

Figure 2. Cape Sable seaside sparrow (photo by Tom Virzi).

Figure 3. Marl prairie habitat near Long Pine Key in the Florida Everglades, Monroe County, Florida (photo by Gary Knight).
The first range-wide assessment (1978–80) found ca. 6,500 sparrows distributed among six subpopulations (Bass and Kushlan, 1982). Today’s populations are much reduced (Table 1). Sparrows nest near the ground (<30 cm) with nest height changing throughout the breeding season in response to water depth (Lockwood et al., 2001). The low nesting height leaves populations susceptible to sudden changes in water levels. A large sparrow population (ca. 2,500 individuals) once found in BCNP declined to fewer than 200 individuals after the area was flooded by an emergency water release in 1981 (Jenkins et al., 2003). This population has not recovered.
Recovery goals envision a change in status from “endangered” to “threatened” when (1) six stable or increasing populations exist; (2) genetic integrity is maintained through natural interchange among populations; (3) management of habitat is performed using natural hydrologic and fire regimes; and (4) available habitat supports viable populations in advance of rising sea levels (U. S. Fish and Wildlife Service [USFWS], 2019b). The area required to support a viable population has been estimated as 1,600 ha, the smallest area occupied by seaside sparrows for an extended period (Lockwood et al., 2001). Critical habitat areas (63,273 ha) have been designated based on data from annual range-wide surveys.
Habitat Restoration and Fire Management Research
Historical data suggest fires affected large areas (>90 km2) in the Everglades every 10–15 years; smaller, more frequent fires were interspersed among more extensive fires (Gunderson and Snyder, 1994). Small, frequent fires ignited by lightning strikes likely occurred during wet periods of the annual cycle (June–August); larger fires likely occurred during drier periods earlier in the year (April–June). Lockwood et al. (2003) suggested a longer fire-return interval of 30–60 years for marl prairies. Differences in the estimated fire-return interval for this grassland community likely reflect the rate with which vegetation recovers following fire. Sites with deeper soils (>40 cm) recover more quickly and may support breeding seaside sparrows within a few years (Taylor, 1993). Sites on shallow soils recover slowly and take longer both to support sparrows and amass the grass vegetation needed to carry fires. Fire and hydroperiod also interact in ways that affect vegetation recovery (Sah et al., 2014). If flooding occurs soon after a burn, prairie grasses die and characteristic vegetation may not recover for decades (Sah et al., 2014).
Early studies considered fires to be essential for managing seaside sparrow habitat (Werner and Woolfenden, 1983; Taylor, 1993; Walters et al., 2000). Sparrow numbers increased in 1 population following a fire and supported a large population ∼10 years post-burn (Bass and Kushlan, 1982). Other studies report a peak in seaside sparrow numbers 3–5 years post-burn and declines occurring 6–10 years post-burn depending on soil conditions (Werner and Woolfenden, 1983; Taylor, 1993). Fire removes dead plant materials that obstruct foraging areas (Post et al., 1983; Taylor, 1993) and may recycle important nutrients (James et al., 1997; Venne et al., 2016). Fire also promotes the C4 grasses associated with occupied seaside sparrow habitat (Sah et al., 2010) and hinders growth of shrubs and trees. Prairies become unsuitable for sparrows once the prevalence of 1.5 m shrubs exceeds 1 ha–1 (Sah et al., 2010).
Follow-up studies of fire have stressed more negative effects. Annual breeding season counts (1992–2001) found low numbers in areas where frequent fires had occurred (ca. 5-year return intervals; Pimm et al., 2002). Frequent fires (≤5-year return interval) also appeared to limit breeding activity (Curnutt et al., 1998). Pimm et al. (2002) concluded that “…strong evidence proving a non-sparrow-related need to burn marl prairies must be presented before Everglades National Park management reinstates prescribed fires within sparrow habitat.” Another study found lower nesting success in the 2 years following a fire that burned 3,410 ha (La Puma et al., 2007). None of the sparrows marked on study area before the burn were seen on the site again. La Puma et al. (2007) urged “…caution against the assumption that occupancy of disturbance-prone habitat automatically suggests dependence on disturbance.”
Communication and Application of Fire Research
Everglades National Park staff organized annual meetings to discuss fire management with sparrow research biologists; the meetings were suspended during a contentious period where one sparrow biologist suggested ENP fire management could be doing more harm than good (R. Anderson, pers. comm.). During this time, ENP staff responded to research results using fire suppression (R. Anderson, pers. comm.). Aerial water tankers and helicopters with water buckets were regularly dispatched to suppress fires approaching occupied sparrow habitat (R. Anderson, pers. comm.). Suppression was applied even on low-intensity fires that might have yielded the patchy mix of burned and unburned areas that Taylor (1993) found sparrows using soon after a burn. ENP staff also used weed whackers and mowers to hand clear fire breaks on some sites scheduled for burns. Land managers working on state lands adjacent to ENP also hand clipped hardwood shrubs in the critical sparrow habitat they managed rather than controlling shrubs more economically with fire (J. Schuette, pers. com.). One fire-related research project relied on steel drums to contain experimental fires conducted within critical sparrow habitat (Snyder and Barrios, 2008). The drum was placed over vegetation and a 0.0002 ha fire was conducted within the cylinder.
A recent assessment points again to potentially positive effects of fire (Benscoter et al., 2019). Annual counts conducted from 1992 to 2014 found higher sparrow numbers 5–8−years after a burn; numbers also improved in relation to the proportion of the area burned within 16 ha of each census station (Benscoter et al., 2019). Error bars about the estimates were broad and point to the need for additional research, but marl prairies in ENP will burn at some point, perhaps under wildfire conditions that will be difficult control. In 2016, a wildfire ignited in a long-unburned pine-dominated area on ENP moved toward a sparrow-occupied prairie. The fire stopped before reaching the population because the fuels needed to carry the fire had been significantly reduced in 2015 using a prescribed burn (R. Anderson, pers. com.).
Challenges to Recovery
Restoring appropriate hydrology is critical to restoring the large sparrow population extirpated on BCNP, but fire has an important role to play, especially in maintaining the grass-dominance and biodiversity of marl prairies. The need for better fire-related research has been emphasized in revisions to the sparrow’s recovery plan (U. S. Fish and Wildlife Service [USFWS], 1999) and recent assessments of research needs (T. Virzi, pers. comm.). Some of the issues warranting study were succinctly outlined by Werner and Woolfenden (1983) nearly 35 years earlier: “Fires either aid or harm sparrows, depending on frequency of burning, percent of contiguous habitat burned, pattern of the burn, size and percent of the occupied area evicted, and proximity to adjacent habitat blocks.” Fortunately, ENP and BCNP have the capacity of sustaining the long-term studies needed to study these factors.
A revised fire management plan for ENP (Everglades Fire Management Plan [ENP], 2015) tries to balance the double-edged effects that fires may have on this endangered sparrow. The acreage treated annually is limited to <35% of critical habitat and <20% of occupied habitat. Within these guidelines, fire likely needs to be re-introduced carefully in an experimental framework given the uncertainties that exist. Research focused on the question of fire-return intervals is a common starting point and seems to be a key information gap for ENP given the broad range of fire-return intervals suggested [from ≤10 years (Werner and Woolfenden, 1983; Taylor, 1993) to ≤60 years (Lockwood et al., 2003)]. Fire research also needs to be replicated across different soil and hydrologic conditions given their importance to vegetation recovery (Sah et al., 2014). Other data suggest changes in hydrology may lead to results that differ from those documented earlier. Marl prairie communities in portions of ENP have become drier in recent decades as a result of hydrologic changes (Sah et al., 2013).
Very large burns (>3,000 ha) are detrimental (La Puma et al., 2007) but less is known about the effects smaller burns may have. Fire creates habitat for many ground-foraging sparrows (Jones et al., 2013; Hewett Ragheb et al., 2019b) and is considered beneficial for other subspecies of seaside sparrow (Gabrey and Afton, 2000; Kern and Shriver, 2014). In addition to small burns, fires could be applied under weather conditions likely to retain unburned patches within the burn unit in an attempt to provide the heterogeneous conditions that Taylor (1993) found sparrows using 2 years post-burn. Night burns that are lit using ground ignition techniques would spread slowly and likely create such conditions. Experimental fires also could be applied annually to 1–3 small burn blocks that are broadly distributed across years.
Suppressing fire in seaside sparrow habitat also represents a single-species approach that may not be compatible with ENP’s ecosystem management. The risks associated with fire suppression also may be higher than estimated. Marl prairies will burn at some point, perhaps catastrophically. Prairies also are likely to shift gradually toward greater shrub dominance if they are not burned (Hanan et al., 2010). Experienced fire managers can adjust the intensity and extent of burns using weather conditions (e.g., high relative humidity) and ignition patterns. Fires might also be applied strategically along ENP boundaries where threats posed by burns performed on neighboring agricultural lands exist.
Florida Grasshopper Sparrow
The grasshopper sparrow is a ground-nesting songbird found in grasslands experiencing frequent fire and other forms of disturbance. The species consists of several migratory subspecies with widespread breeding ranges (southern Canada and the United States) as well as several non-migratory subspecies existing in small isolated populations (southern United States, Mexico, and the Caribbean; Vickery, 2020). The Florida grasshopper sparrow (Figure 4) is a non-migratory subspecies endemic to dry prairie habitat (Figure 5) in central Florida (Walsh et al., 1995), ca. 400 km away from the nearest migratory subspecies population in southern Georgia.

Figure 4. Florida grasshopper sparrow.

Figure 5. Dry prairie habitat on Three Lakes Wildlife Management Area, Osceola County, Florida occupied by the largest remaining population of Florida Grasshopper Sparrows.
The Florida grasshopper sparrow is nearing extinction, having declined by 89% across the three public land populations monitored annually for 20 years (Florida Grasshopper Sparrow Working Group, unpub. data). Surveys in 2019 estimated <35 breeding pairs across all known wild populations (Florida Grasshopper Sparrow Working Group, unpub. data). If population declines continue, the Florida grasshopper sparrow may become the first bird to go extinct in the United States since the dusky seaside sparrow (Ammodramus maritimus nigrescens) in 1978 (Post and Greenlaw, 2018). Historical declines have been attributed to habitat loss, fragmentation, and degradation as a result of cattle grazing, fire suppression, and altered hydrology (Pranty and Tucker, 2006). Estimates of dry prairie coverage prior to European settlement vary from 203,730 to 830,000 ha (Bridges, 2006; Delany et al., 2007). As of 2004, only 44,993 ha of suitable habitat remained (Delany et al., 2007) and currently only 22,719 ha are actively managed for grasshopper sparrows (Table 1).
The Florida grasshopper sparrow was federally listed as endangered in 1986 (U. S. Fish and Wildlife Service [USFWS], 1999). The recovery plan states that the subspecies may be downlisted to threatened when ≥10 sites contain growing, self-sustaining populations with >50 breeding pairs (U. S. Fish and Wildlife Service [USFWS], 1999). Habitat restoration recommendations include the use of frequent prescribed fire (≤2 years), mechanical removal of woody vegetation, and discouraging cattle rangeland improvements that reduce native vegetation (U. S. Fish and Wildlife Service [USFWS], 1999). Recovery actions within the range of the sparrow also include additional surveys on private lands, creating habitat corridors, conducting reintroductions, and developing a captive-propagation plan (U. S. Fish and Wildlife Service [USFWS], 1999).
Habitat Restoration and Fire Management Research
Dry prairie habitat was historically maintained by frequent, lightning initiated fires (every 1–3 years; Huffman, 2006; Huffman and Platt, 2014) and seasonal rainfall patterns resulting in extended periods of ground water saturation (Platt et al., 2006). Existing research supports the importance of frequent fires for maintaining Florida grasshopper sparrows. Sparrow occupancy, territory density (Walsh et al., 1995; Shriver and Vickery, 2001), and annual productivity (Shriver and Vickery, 2001; Delany et al., 2002) are higher in dry prairies burned within the last 18 months than in prairies burned ≥2 years ago.
Fire seasonality has been another important topic of among researchers and practitioners. Prior to the late 1990s, prescribed fires were recommended during the dormant fire season (January–March) to avoid burning during the nesting period (April–August; Walsh et al., 1995). However, lightning-induced wildfires were more prevalent in areas occupied by sparrows historically during the transition season (April–June) when early summer thunderstorms overlapped with the dry, ground-level fuels capable of sustaining fires (Platt et al., 2006). Burning dry prairie during the transition season also reduces woody shrubs and promotes flowering of prairie grasses (Walsh et al., 1995) and for these reasons many managers shifted towards predominately transition-season burns. However, recent demographic research revealed that frequent transition-season fires did not result in higher nest survival than fires conducted earlier in the year (within a 2-year fire interval; Hewett Ragheb et al., 2019b). More important to sparrow management, transition-season fires have the potential to destroy active nests and delay renesting (Hewett Ragheb et al., 2019b). Therefore, current recommendations for habitat management of the remaining tiny populations focus on strategic planning of transition-season fires to maintain or restore habitat adjacent to occupied areas. Whereas, occupied areas can be burned during the dormant season prior to nest initiation (Hewett Ragheb et al., 2019b). Application of these recommendations are dependent on effective communication between monitoring teams and land managers.
Many research questions remain regarding the relationship between Florida grasshopper sparrows and prescribed fire. The extent of prairie burned when fires are applied varies among sites, and a suitable scale of area burned remains unknown. For example, sparrow burn units at Three Lakes Wildlife Management Area (TLWMA; 27.93, −81.13; 25,000 ha) have been relatively small (<375 ha, 2000–2013) resulting in a heterogeneous mosaic of fire interval and seasonality that provides many options for nesting sparrows, whereas several burn units at Kissimmee Prairie Preserve State Park (KPPSP; 27.53; −81.02; 21,800 ha) have been consistently larger (>1000 ha; 1998–2013). Large-scale fires may be helpful if nest predators avoid the core of the unit as a result of direct mortality or loss of cover (Jones et al., 2004). However, large-scale fires may present less room for error if the applied fire interval, seasonality, or intensity turns out to be sub-optimal. Alternatively, small-scale fires may require considerably more time and may not be feasible when staff and resources are low.
Other fire-management questions relate to the intensity or “patchiness” of the burns and finer details associated with fire-return intervals and season. Low-intensity fires yield a matrix of burned and unburned vegetation providing a heterogeneous mix of new grasses that attract insect prey while still maintaining cover for nests or young fledglings. Alternatively, high intensity fires during periods of drought may be necessary to reduce patches of unburned vegetation surrounding embedded wetlands that may provide cover for predators. Little is known about how alternation of fire season may affect vegetative response, prey availability, or sparrow demographics over time. It is also unknown if annual fires instead of biennial fires are optimal.
Communication and Application of Fire Research
The Florida Grasshopper Sparrow Working Group was established in 2002 and persists as the primary means of communication among research biologists (from state agencies, universities, and private research stations) and public land managers (see Table 1 for distribution of land ownership). The implementation of fire-related research recommendations has been relatively successful in recent years because of the small number of properties (4) and outside stakeholders (<15) involved. The working group holds biannual meetings to exchange information and has been pivotal in establishing standardized monitoring methods, communicating research on novel predation management techniques (Hewett Ragheb et al., 2019a), and implementing a captive propagation and release program (U. S. Fish and Wildlife Service [USFWS], 2019d). The working group has attended habitat management workshops (Cox, 2014) and recently assisted federal and state agencies with the development of a 5-year Strategic Vision document intended to guide future conservation and management actions for the subspecies (U. S. Fish and Wildlife Service [USFWS], 2019d).
Communication of Florida grasshopper sparrow habitat management recommendations to practitioners on private cattle ranches has been more challenging. A recent 3-year project by the USFWS provided financial incentives to four private ranches in return for access to conduct sparrow surveys, develop and implement site-specific management plans, and monitor vegetative response to habitat improvements (Castellon, 2017). Habitat management recommendations specific to cattle ranches are being prepared. These positive initiatives notwithstanding, private landowner participation in sparrow management is voluntary and often difficult to incentivize. The added expenses associated with sparrow management are high relative to more routine pasture management. There is a need to reach more private landowners and convey sparrow management recommendations through other, well-attended platforms (e.g., Florida Cattleman’s Association meetings) rather than meetings of the Florida Grasshopper Sparrow Working Group.
Challenges to Recovery
Advancements in fire management for Florida grasshopper sparrows on public lands have been hindered by a lack of research-based recommendations more so than a lack of communication between researchers and practitioners. For example, a recommendation of burning sparrow habitat every 1–3 years during the transition season (April–June; U. S. Fish and Wildlife Service [USFWS], 1999) was willingly adopted by land managers. However, this recommendation was based on a general effort to mimic natural fire regimes rather than research directly monitoring the effect of fire seasonality on sparrow nest success (which later revealed transition-season fires actually hinder reproduction for small populations; Hewett Ragheb et al., 2019b). This previous emphasis on burning only during the brief transition season in combination with local burn bans and staffing limitations may have resulted in prairies lapsing into fire-return intervals that are not preferred by grasshopper sparrows (>3 years; Walsh et al., 1995; Shriver and Vickery, 2001), a problem that may have substantial impact to populations when primarily large-scale burns are conducted at a site.
Previous gaps in habitat research for this subspecies have been driven by periods of inadequate funding. An increase in public awareness in the mid-2010s (Williams, 2013) resulted in access to more federal funding, however, researcher investment simultaneously shifted to emergency actions such as predation management and captive propagation. There is still an urgent need to resume habitat management related research objectives to generate long-term solutions to population viability.
Several additional hurdles exist for recovery beyond the development of sustainable fire-management recommendations. First, the conversion of private cattle pastures to suitable grasslands will be necessary to meet the recovery goal of 10 populations (U. S. Fish and Wildlife Service [USFWS], 1999). This is a substantial challenge because previous habitat alterations aimed to improve cattle grazing (e.g., planting non-native grasses) appear to increase threats from non-native red-imported fire ants (Solenopsis invicta; Tucker et al., 2010), a formidable nest predator (Hewett Ragheb et al., 2019a). Second, little to no research has been conducted to understand how mechanical removal of woody vegetation, exotic plant treatments, or alterations to hydrology impact sparrow demography. Third, Florida grasshopper sparrow nest predation rates are very high (Hewett Ragheb et al., 2019b), but only a few studies have addressed how current habitat management practices influence nest predators (Tucker et al., 2010; Harris et al., 2019). Finally, small populations are vulnerable to stochastic events (Caughley, 1994) beyond the control of land managers, which may reduce the effectiveness of new management actions and hinder the ability to monitor management changes.
Florida Scrub-Jay
The Florida scrub-jay (Figure 6) is the only bird species endemic to Florida (Woolfenden and Fitzpatrick, 1991). Scrub-jays are monogamous and generally occupy the same territory for life once they become breeders. Territorial groups comprise a breeding pair and non-breeding offspring that remain within natal territories for a year or more to help feed future young. Family members also defend territories from neighboring groups and scan for (and mob) predators. Helpers improve productivity and recruitment compared to territories without helpers (Mumme, 1992). This cooperative breeding system also affects efforts to monitor responses to management because helpers buffer population numbers and affect productivity and recruitment (Woolfenden and Fitzpatrick, 1984). Florida scrub-jays also are one of the most habitat restricted species in North America and often spend their entire lives within a few kilometers of natal territories (Coulon et al., 2010).

Figure 6. Florida scrub-jay.
Scrub-jay territories usually include oak scrub ridges, pine flatwoods, and ephemeral marshes (Breininger et al., 1995). Scrub ridges (Figure 7) are ancient sand dunes created by changes in sea levels. The porous sands found beneath scrub ridges drain well and make them prime sites for human development. As a result, many scrub-jay populations are essentially enclaves surrounded by the added threats posed by domestic animals and roads (U. S. Fish and Wildlife Service [USFWS], 2019c). Habitat fragmentation and fire suppression have degraded most of the large blocks of natural habitat found elsewhere and left populations vulnerable to extinction as a result of low dispersal rates, inbreeding, and continued human development (Breininger et al., 1995; Chen et al., 2016).

Figure 7. Optimal scrub-jay habitat on Valkaria Scrub Sanctuary, Brevard County, Florida. Optimal habitat has medium height (1.2–1.7 m) shrubs and young trees with open sand patches intermixed over ∼10 ha.
The scrub-jay has experienced steady declines since it was listed under ESA in 1987 (U. S. Fish and Wildlife Service [USFWS], 2019c). The scrub-jay is extirpated from 10 of the 39 Florida counties where it once occurred; only nine counties support ≥30 scrub-jays (U. S. Fish and Wildlife Service [USFWS], 2019c). Recovery goals seek to sustain scrub-jays within seven large landscapes. Recovery will be reached when each of these landscapes supports stable or increasing populations sustained through natural recruitment (U. S. Fish and Wildlife Service [USFWS], 2019c). Genetic diversity among subpopulations within the focal landscapes must also be maintained naturally, and long-term threats posed by climate change and development need to be adequately addressed.
Habitat Restoration and Fire Management Research
The importance of fire management to scrub-jays and scrub ecosystems has been established by an extensive body of research (Fitzpatrick et al., 1991; Breininger et al., 2002, 2014a,b; Florida Fish and Wildlife Conservation Commission [FFWCC], 2018). Lack of fire has catastrophic consequences for scrub-jay populations (Woolfenden and Fitzpatrick, 1984, 1991; Fitzpatrick et al., 1991), and several studies have helped to define the vegetation structure that scrub-jays seek and fires provide. Optimal habitat (Figure 7) has medium height (1.2–1.7 m) shrubs and young trees with open sand patches intermixed over ∼10 ha. Open sand patches are important for foraging and acorn caching but do not persist long after fire. Long unburned sites as well as sites where most of the vegetation has been removed by fire are not usually occupied or else experience high mortality (Breininger et al., 2009, Breckheimer et al., 2014). Family groups with large territories have an advantage in landscapes where fire management creates a more heterogeneous mix of bare sand and low shrubs.
While research has yielded state-of-the-art habitat models and management guidelines (Breininger et al., 2009, 2010; Florida Fish and Wildlife Conservation Commission [FFWCC], 2018), the process of maintaining and restoring suitable conditions with fire is complicated by geographic differences in the dominant shrubs and trees present and the response vegetation has when burned (Breininger et al., 2018). Suitable habitat conditions in areas dominated by sand pine (Pinus clausa) were maintained historically by high-intensity crown fires that are difficult to control and mimic using prescribed fires. Instead, fire managers attempt to use the shortest fire-return intervals recommended for the composition and type of scrub present with a goal of promoting a heterogenous composition of burned and unburned patches (Breininger et al., 2002).
In addition, much of the plant biomass found in scrub lies underground and is used to store the nutrients plants need to recover after a burn. As a result, growth of scrub vegetation post-fire varies greatly based on pre-burn conditions, species composition, nutrient availability, and depth to water table (Breininger et al., 2010, 2018). Scrub that has not been burned for long periods grows back faster post-burn than scrub subjected to shorter fire-return intervals. Additional variation in the rate of vegetation recovery is linked to rainfall post-burn. Edge effects created by roads, fire-breaks, and adjacent forest are additional factors that affect fire spread and cause differences in site-specific responses to fire (Duncan and Schmalzer, 2004; Breininger et al., 2010, 2018).
The variability and uncertainties associated with scrub management argue against using any single fire-return interval throughout the range of the scrub-jay. Fire return intervals should be more frequent on many areas, but Adaptive Resource Management (ARM) programs (Johnson et al., 2011; Breininger et al., 2014a) are recommended whereby local monitoring of scrub-jay populations and scrub-jay habitat are used to guide site-specific management decisions (Johnson et al., 2011). The predicted responses to management are compared with the predicted responses to alternative management actions and often begin with projections that biologists and managers provide. Monitoring is used to assess the projections and provide precision to the evaluation of management strategies (Williams et al., 2011). ARM has been developed to facilitate direct interaction between scientists and managers along Florida’s east coast (Eaton et al., submitted).
Communication and Application of Fire Research
Our survey of Florida scrub-jay researchers and practitioners (see Supplementary Material) revealed widespread belief that sufficient research exists to promote and understand fire management for scrub-jays. At the same time, both groups expressed uncertainty about the future prognoses because of the effects of fragmentation, urbanization, and other factors. A majority (87%) of the managers believed they had enough information on habitat requirements to conduct effective management, and a majority of these (67%) had received enough feedback about their management. On the other hand, a majority (77%) was unsure that Florida’s managed area supported enough scrub-jays to provide long-term viability. Survey participants suggested additional literature on management needs and training workshops were low priorities; however, recurring annual meetings to discuss fire management, ARM, and understanding demographic and genetic trends would be very useful.
Some of the variation in responses among participants related to regional differences in scrub habitat and the knowledge base available. Archbold Biological Station (27.18, −81.35; 2101 ha) has studied scrub-jays and scrub for decades along the Lake Wales Ride in Central Florida. This ridge supports some of the oldest upland habitats in peninsular Florida that sit atop deep, sandy soils. Conditions on the Lake Wales Ridge differ from conditions found on other ridge systems in Florida (Schmalzer and Hinkle, 1992; Menges and Hawkes, 1998; Schmalzer et al., 1999; Menges et al., 2008). Scrub in coastal areas of east central Florida has been especially difficult to restore. Most managers at coastal sites are more uncertain about how to maintain the mix of patches of open sand and medium height scrub needed by scrub-jays (Breininger et al., 2018). In addition, the largest extant scrub-jay population occurs on Ocala National Forest in north central Florida (29.17 N, −81.79; 175 km2) where habitat management for scrub-jays was performed for many decades using forestry practices, not fire (Franzreb and Zarnoch, 2011). This large population is now the subject of enhanced population monitoring coupled with restoration work focused on taking a large area of sand pine out of timber rotation and managing the area instead with fire (K. Miller, pers. comm.).
Despite the extensive information available on scrub-jays, reviews of the quality of scrub-jay habitat on many public lands suggest opportunities for improved management exist (Breininger et al., 2006, 2018; Boughton and Bowman, 2011). Fire is not being applied frequently enough on some sites; burns are too extensive on others and do not yield the mosaic of low-statured scrub and open patches required by scrub-jays (Figure 7). Extending the application of ARM to other scrub-jay populations should help to evaluate the habitat quality provided through fire management in a non-confrontational manner. Scrub-jay survey results also point to constraints on the application of fire because of staff limitations. Scrub-jay populations are also distributed across a large number of small managed areas (Table 1) where factors other than habitat quality will affect scrub-jay viability. Optimal habitat conditions could be provided by land management on these smaller sites, but scrub-jay populations are likely to continue to decline because of their small sizes, distance to other populations, and threats posed by surrounding development (Boughton and Bowman, 2011; Breininger et al., 2018).
Challenges to Recovery
Results of our scrub-jay survey point to many of the common challenges associated with fire management (e.g., staff limitations and too few burn days). Providing solutions to these problems will be complex because of the many different agencies involved in managing scrub-jay habitat, variation in the size of the managed units, and the landscape context within which the units exist. Agencies also work under different guidelines and rules that can seem frustratingly inane to research biologists. For example, some agencies allow their fire crews to be shared but not the equipment needed to conduct a fire – either because of maintenance issues or simply because the equipment used by agency A is not allowed on sites managed by agency B.
Florida scrub-jays have large territories for their body size, a trait that may reflect benefits provided by defending areas with a mix of burned and unburned scrub (Fitzpatrick and Bowman, 2016). Creating burn heterogeneity at the scale of individual territories is challenging because scrub is difficult to ignite except under conditions when the resulting fire may be difficult to control. Survey respondents also suggested fire effects in scrub were difficult to predict for 33% of the areas surveyed (see Supplementary Material). Heterogeneous burns yield more discontinuous fuels after a burn and pose less wildlife risk between fires (Figure 8), but heterogeneous burns also require post-burn monitoring to ensure unburned patches do not ignite hours or days after burn crews leave (Duncan et al., 2015). Alternatively, heterogeneity can be created by altering the extent of the area burned, but whether this approach provides the persistent openings scrub-jays used historically is unknown. Additional applied research integrated with fire management techniques are needed to determine how to restore persistent openings and get scrub to burn.

Figure 8. Heterogenous scrub burn conducted on Jonathan Dickinson State Park, Martin County, Florida, April 2020 (photo by Rob Rossmanith).
Another research issue lies with increases in the prevalence of sand, slash (Pinus elliottii), and longleaf pines (Pinus palustris) that have grown to heights and densities detrimental to scrub-jays. Timber operations that were once used to restore appropriate conditions for scrub-jays on some areas are no longer economical viable. Persistent openings also were often created by burning dead tree debris piles left by intentional removal of tall pines and oaks. Tree piles create hot spots that kill below-ground roots and produce more persistent openings. However, timber operations also can lead to unacceptable levels of soil disturbance, spread exotic plants transported on the equipment needed to fell trees, and lessen the ability for sites to carry fire (Menges and Hawkes, 1998; Menges and Gordon, 2010).
There also are differences regarding the importance of season of burning and the fuels needed to carry fire in scrub. Native grasses found in scrub-jay habitat burn easily within 1–2 years post fire. Saw palmetto (Serenoa repens), another common ground cover species, can take years to accumulate the dead fronds needed to carry a fire. Grasses were once a dominant ground cover component of pine flatwoods but have become much reduced at the expense of an increase in palmetto cover. Fires conducted during the transition period (April–June) when drier conditions prevail favor grasses at the expense of shrubs and palmetto (Robbins and Myers, 1992). Burning during this seasonal period could restore grass dominance in pine flatwoods and make it easier to ignite neighboring oak scrub. This could yield added benefits to scrub-jays because fires moving through pine flatwoods often cross the ecotone and then extinguish in the less flammable oak scrub. Many persistent scrub openings occur as a result along the ecotone. Scrub-jays need ecotones between oak scrub and adjacent pinelands with open ground for caching acorns.
Red-Cockaded Woodpecker
The red-cockaded woodpecker (Figure 9) requires mature (70+ years), open-structured pine forests (Figure 10) that are maintained by frequent fire (Conner et al., 2001). This woodpecker excavates its roosting and nesting cavities exclusively in old living pine trees. The cavities often take years to excavate but also persist and may be used for over a decade. This life history trait likely led to development of cooperative breeding in this species whereby young males often remain on natal territories with a surplus of cavity resources rather than attempting to disperse (Walters et al., 1988). These helper males provide care to future young, defend territories, and also excavate additional cavities. This woodpecker prefers to forage on large, living pine trees in relatively large territories (40–160 ha; U. S. Fish and Wildlife Service [USFWS], 2003).

Figure 9. Red-cockaded Woodpecker.

Figure 10. Old-growth longleaf pine on the Wade Tract, Thomas County, Georgia. The tract supports 1 of the highest densities of red-cockaded woodpeckers (ca. 1 breeding group per 50 ha).
Although it occurred in 15 southeastern states and was once considered common (Jackson, 1994; Conner et al., 2001), its population in 2009 was <3 percent of the estimated abundance at the time of European settlement (Florida Fish and Wildlife Conservation Commission [FFWCC], 2009). The extensive longleaf pine ecosystem that dominated or was a significant component of 37 million ha of the presettlement southeastern United States (Frost, 2007) provided optimal habitat for the woodpecker, but woodpeckers also use other pine species for cavities and foraging. A recent estimate for the extent of remnant longleaf pine was 1.7 million ha (Oswalt et al., 2012) – a decline of 95%.
The principal causes of woodpecker declines were loss of the original longleaf pine ecosystem, habitat degradation from fire suppression and fragmentation, and the switch to short-rotation silviculture (U. S. Fish and Wildlife Service [USFWS], 2003). The most important steps toward recovery occurred when techniques were developed to construct artificial cavities (Figure 11) in living pine trees (Copeyon, 1990; Allen, 1991) and to translocate woodpeckers to both augment small population and restore populations on formerly occupied sites (Cox and McCormick, 2016). Combining these techniques enabled land managers to increase territory densities on the land base available, enhance small populations that would otherwise unsustainable, and re-create populations where the woodpecker had been extirpated (Weiss et al., 2019).

Figure 11. Artificial cavity installation conducted on Tall Timbers Research Station, Leon County, Florida. The type being excavated here is a drilled version that typically takes about an hour to complete.
With nearly 2,600 active territories in Florida (Figure 1), the woodpecker is supported today by 500,000 ha of pine woodlands (Table 1) and another 500,000 ha of potential habitat (Endries et al., 2009). As of 2019, all 17 recovery sites identified in the recovery plan in Florida had increased in the number of active territories; 10 had exceeded the population goals outlined in the plan (U. S. Fish and Wildlife Service [USFWS], 2003; Table 1). Range-wide population increases have led to changes in the woodpecker’s status and consideration for down-listing at the federal level (W. McDearman, pers. comm.). As important as artificial cavities and translocation have been to the steady growth of woodpecker populations, frequent application of prescribed fire, transitioning from planted slash pine to longleaf pine, and implementing “silviculture that sustains” (Mitchell et al., 2010) are essential management components of sustained recovery over coming decades.
Habitat Restoration and Fire Management Research
The red-cockaded woodpecker is arguably one of the best-studied woodpeckers in the world (Jackson, 1994). In many ways management for the red-cockaded woodpecker helped spur conservation of the longleaf pine ecosystem, one of the most biologically rich forest types in North America (Kirkman et al., 2018). Interest in restoring and conserving the ecosystem generated a substantial scientific literature on both prescribed fire and silviculture (Jose et al., 2006; Kirkman and Jack, 2018) that is directly relevant to red-cockaded woodpecker management. This research – both for the species and ecosystem – was distilled into a thorough and scientifically based recovery plan (U. S. Fish and Wildlife Service [USFWS], 2003). This exemplary recovery plan provided explicit management guidelines for application of frequent prescribed fire, including the season of application, cavity tree protection, burning for habitat restoration, and burn prioritization.
The recovery plan emphasizes application of frequent fires conducted from March-June to maintain optimal red-cockaded woodpecker habitat and cautions against reliance on dormant season (October–February) fires (U. S. Fish and Wildlife Service [USFWS], 2003). Growing-season fires mimic the natural fire regime (Komarek, 1968) and are more effective than dormant-season fires in controlling hardwood trees and shrubs (Waldrop et al., 1992). Some studies suggest that the grass and herb dominated ground cover created by growing-season fires may also improve the nutritional quality and abundance of arboreal arthropods in the woodpecker’s diet (James et al., 1997), but another assessment found no strong link between ground cover characteristics and nest productivity (Garabedian et al., 2014). Infrequent fire or fire exclusion will lead to development of a hardwood midstory that causes abandonment of the territory by red-cockaded woodpeckers (Van Balen and Doerr, 1978). Mechanically removing the mid-story and re-introducing fire can restore the open structure preferred by the woodpecker and other birds that prefer open pinelands (Steen et al., 2013). Ecological reasons for use of growing-season fire must be weighed against the possibility that an overemphasis on growing-season fires could effectively reduce the number of days that fires are applied and result in fewer acres burned. Frequent fire with variable fire seasonality is most likely to enhance a diverse ground cover of grasses and forbs (Kirkman and Giencke, 2018).
Restoration of former longleaf pine sites where other pines (mostly slash pine) were once planted is an important objective in the management plan for the National Forests of Florida (U. S. Department of Agriculture and Forest Service [USDA-FS], 1999). This restoration will provide many acres of foraging habitat and eventually nesting habitat for the red-cockaded woodpecker once trees mature. Longleaf pine restoration should also improve fire management given the pyrogenic qualities of longleaf pine needles. Longleaf pines produce long, resinous, highly flammable needles that are shed annually and account for a large percentage of the fine fuels at ground level (Platt et al., 2016). These fine fuels drive prescribed fires (Platt et al., 2016) and require that consideration be given to maintaining overstory densities of longleaf pines to provide adequate needle cast for restoration efforts (Knapp et al., 2008; Hess and Tschinkel, 2017). Restoration of grass and other fine fuels is also critical in the application of prescribed burns when restoring former agricultural lands and long fire-suppressed pinelands (Loudermilk et al., 2018).
Communication and Application of Fire Research
Research on the use of prescribed fire in woodpecker management has effectively expanded for several reasons. Over 95% of the woodpecker population in Florida resided on government land (77% federal and 19% state) in 2000, including nearly 30% on Eglin Air Force Base (EAFB; 30.46, −86.55; 1850 km2), a core population for recovery (Hovis and Swan, 2004). Four of the remaining 13 core woodpecker populations designated for recovery also occur on Department of Defense lands (U. S. Fish and Wildlife Service [USFWS], 2003). A 1989 jeopardy opinion issued by the USFWS caused military leadership to reconsider the relationship between natural resource management and the military’s mission and resulted in focused funding and goals to increase the woodpecker population (Petrick and Hagedorn, 2004). Strong top-down directives within the military to comply with the ESA coupled with the large annual budgets the Department receives led to the allocation of significant funds to research and management of this species. As a result, ≥40 articles on woodpecker ecology and management have been published over the last 20 years based on the work conducted on military bases in North Carolina and Florida (T. Engstrom, unpubl. data).
Communications have also been enhanced by procedures adopted for woodpecker translocation. To coordinate movement of birds from healthy populations to developing populations, the USFWS holds three regional meetings each year that bring together land managers, woodpecker managers, and research biologists. These forums provide opportunities for research and management to discuss new field methods, population status, prescribed fire achievements, and the logistics of translocation (W. McDearman, pers. comm.). Because one of these regional meetings takes place in Florida each year, woodpecker biologists and land managers in Florida hold a separate statewide gathering on the day before the regional meeting starts.
Challenges for Recovery
A formidable challenge for sustainable recovery of the red-cockaded woodpecker in Florida and elsewhere will be maintaining appropriate fire regimes. Long-term ecosystem restoration and management goals also require a shift in focus, although traditional woodpecker management techniques should continue to be used. For example, the Apalachicola National Forest (ANF; 30.11, −84.72; 2561 km2) population and the population on EAFB have been instrumental in generating young birds that are used to augment other populations via translocation. One of the most important management goals for pine forests on the ANF is to maintain a 3-year fire return interval (U. S. Department of Agriculture and Forest Service [USDA-FS], 1999). This fire-return goal has not been achieved on approximately 80% of the acres in pine forests in the ANF (M. Keys, pers. comm.) and may be leading to steady decline in ecosystem health. Fire-maintained uplands in ANF are deteriorating as shrubs are encroaching into upland herbaceous groundcover vegetation (Hess, 2014). These changes have the potential to change fire behavior, making burns less efficient when they are conducted, and lead to lasting effects on other listed species. Meanwhile, another effect of the enhanced funds provided by the Department of Defense has been the maintenance of more frequent burning on EAFB. EAFB has approximately 2.5-times the annual fire-management budget of ANF (D. Beard and K. Hiers, pers comm.).
Another challenge to fire management is the fuel reduction used to treat areas around red-cockaded woodpecker cavity trees. Woodpeckers excavate wounds near cavity entrances that allow resin to ooze onto the trunk. The resin is an effective deterrent to climbing rat snakes (Pantherophis sp.) that may eat eggs, nestlings, and incubating adults (Rudolph et al., 1990). Active cavities (i.e., cavities currently being used by woodpeckers) can be covered in this highly flammable resin and may catch fire and burn during prescribed burns. If this happens, fire may reach into the canopy and kill the tree or lead to abandonment of the cavity by the woodpecker. To prevent this from happening, the grasses and fine fuels within 2 m of cavity tree are mowed and raked away from the base of the tree (Williams et al., 2006). It is a singularly time-consuming activity that may impede application of prescribed fire and effectively reduce the number of acres burned (K. Russell, pers. comm.). Additional applied research to improve the efficiency of pre-burn site preparation of woodpecker cavity trees could help increase the number of acres burned.
Discussion
Florida has one of the best fire administration programs in the world. Certificate coursework and annual meetings of regional working groups provide great training (Melvin, 2018). On-line tools are available to predict the site-specific smoke impacts that any burn may pose. Permits are granted on the morning a burn is scheduled to take place to guard against sudden weather changes, and, once a permit is issued, legal protection is conferred against charges of gross negligence.
While these are conducive conditions for burning, our assessments of four rare avian taxa point to outstanding issues even when attempting to sustain habitat with fire on public lands. The natural fire regimes science suggests once affected many of Florida’s ecosystems are simply very difficult to mimic today. Burn bans are imposed to protect homes and human lives when prolonged droughts occur, effectively prohibiting the use of fire when natural conditions for fires are most favorable (Nowell et al., 2018). Natural fires also burned into night when humid conditions and calm winds prevailed. Night fires meander slowly consuming only the most volatile vegetation while skirting sizeable areas with less flammable vegetation. Permits for night burns are difficult to obtain because the smoke generated blankets nearby highways, airports, shopping malls, and residential areas. The heterogenous mix of burned and unburned patches that night burns once created are less common as a result.
Research on natural fire regimes is extremely important for understanding the ecological and evolutionary roles fire plays, but efforts to explore “…the full range of fire severity, fire sizes and fire intervals…” (Driscoll et al., 2010) seem impractical and potentially dangerous, especially when attempting to manage rare species restricted to a small fraction of their former ranges. Fire managers set fires when burns can be initiated and controlled. The factors they manipulate are (1) frequency with which fire returns to an area; (2) season in which fires are applied; (3) extent of the area treated; (4) ignition techniques used to launch a fire (Figure 12), and (5) the specific weather conditions under which a fire is initiated. These factors represent the basic elements of fire management and can lead to both positive and negative outcomes for rare species. To help manage and restore habitat for rare species, each of these factors needs to be the focus of additional research.

Figure 12. Aerial spot ignition (bottom) and ground-based strip ignition (top). Aerial ignition creates multiple fire lines that typically cover a larger area and contain more complex fire convergence patterns (photos provided by Greg Titus).
Fire Frequency
Fire frequency plays a fundamental role in sustaining habitat conditions for the focal species assessed here (and many others). Substantially altered fire frequencies represent a leading factor behind the decline of many rare species (Driscoll et al., 2010). Science-based fire-frequency recommendations exist for Florida’s natural areas (Florida Natural Areas Inventory [FNAI], 2010) as well as the focal species considered here thanks to the history of fire research in Florida that includes dendrochronology studies extending back decades prior to European settlement (Huffman, 2006; Huffman and Platt, 2014). This important background information does not exist for other parts of the globe. Despite its importance, appropriate fire frequencies have not been consistently maintained on areas used by our focal species for markedly different reasons. Fire-management was suppressed for seaside sparrows even though lack of fire could lead to shifts in the structure of the grass-dominated prairies sparrows need (Hanan et al., 2010; Sah et al., 2014). Fires conducted were less frequent on some dry prairie sites used by Florida grasshopper sparrows following a narrow focus on the season when fires were applied. Fire frequencies used on many large public landholdings also have been longer than recommended for maintaining woodpecker and scrub-jay habitat.
A prime reason for not maintaining appropriate fire frequencies likely relates to the lack of resources devoted to fire management. Although a lack of suitable burn days was listed as a major impediment to burning by scrub-jay managers (56.2%), burn days and staffing are conflated. Staff need to be available and prepared to burn when suitable conditions exist, but suitable conditions are often difficult to predict ≥4 days ahead of a proposed burn, meaning that staff schedules can be filled with other responsibilities beforehand. Burn days are also site-specific and dictated by surrounding land-uses and local weather. Chiodi et al. (2018) estimated ≥60 burn days were available annually for many areas in Florida. Land managers working on large, remote public landholdings report ≥100 burn days available each year (S. Glass, pers. com. [TLWMA]; K. Hiers, pers. com. [EAFB]).
Support for regional fire-management teams that focus exclusively on conducting prescribed burns is essential for improving fire frequencies on public lands in Florida. Burn crews operating within the range of the Florida scrub-jay added ≥30 burns to the typical number conducted on properties supporting scrub-jays (Z. Prusack, pers. comm.). Another option is to involve research biologists in burns by asking them to complete the minimum training needed to assist (ca. 30 h of on-line sessions and a single field day). One Florida agency cross trained most of its staff to assist with burns; the acreage burned increased substantially (Miller et al., 2004). In addition, there is no better way for research biologists to learn about fire management than to see its application first-hand.
Research that fine-tunes fire frequencies based on local conditions is needed for scrub-jays, seaside sparrows, and certainly other listed species not assessed here. The speed with which vegetation recovers post-fire varies across the range of both species based on soil and water conditions. Recent guidelines for managing scrub-jay habitat focused on site-specific vegetation conditions to guide fire application rather than recommending a specific burn interval (Florida Fish and Wildlife Conservation Commission [FFWCC], 2018). Similar application of ARM might be used to evaluate habitat for Cape Sable seaside sparrow and devise projections on population trajectories based on the time since fire and prevalence of shrubs ≥1.5 m. The very broad range of fire frequencies recommended for the marl prairie habitats sparrows use (5–60 years; Taylor, 1993; Lockwood et al., 2003) also suggest fire-related studies can proceed cautiously because habitat suitability is not likely to degrade quickly.
Season and Extent
More studies on the seasonal effects of fire are needed, especially in light of data (Chiodi et al., 2018) suggesting the number of burn days available during the 3-month transition-season when natural fires occurred (April–June; Platt et al., 2006) is reduced compared to other times of the year. Frequent transition-season burning is more effective in controlling woody shrubs than burns conducted in other seasons (Willcox and Giuliano, 2010; Hess, 2014). However, as observed for grasshopper sparrows, a narrow focus on transition-season burning can lower habitat quality overall if this focus reduces the acreage burned annually. Burning across a broader window of time can also provide other benefits. Cox and Widener (2008) documented higher adult survival for the northern bobwhite (Colinus virginianus) when fires were extended across multiple seasons rather than a strict focus on burning within a 2–4 week period. Spreading burns out allowed vegetation to recover on areas burned early; these areas then provided cover for this ground-foraging bird when adjacent units were burned later in the year. On the other hand, the C4 grasses associated with Florida’s prairies, pinelands, and other fire-dependent habitats fare better when transition-season fires are applied. Additional work should be designed specifically to address the tradeoffs involved with burning across different seasons. Long-term effects need to be assessed by manipulating the mix of transition-season burns with burns conducted outside this window (e.g., 1:3 vs. 1:1 ratio of transition-season to burns in other seasons).
Better information on the importance of the extent of the area burned also is warranted for our focal taxa and other rare species (Hill et al., 2017). The reproductive potential of a significant proportion of the total populations of both sparrows could be negatively affected by extremely large burns (La Puma et al., 2007; Hewett Ragheb et al., 2019b). Fire-management for both sparrows needs to consider landscape-level factors such as the proximity of nearby unburned patches and their continuity with areas burned. The scrub-jay could also be negatively affected by large burns and is also more likely to remain on burned territories even when habitat quality is low (as is the woodpecker). Still other life-history traits could influence the positive or negative effects of burn extents. Given the importance of nesting success to both sparrows (Baiser et al., 2008; Hewett Ragheb et al., 2019a), attempts to manage prominent nest predators using large-extent fires might be pursued (Jones et al., 2004; Carter et al., 2007); however, managing habitat for predators will be complicated compared to managing for vegetative structure. While the woodpecker does not appear to be affected directly by burn extent, pressures to burn smaller units for other species on ANF (e.g., butterflies and quail) could lead to further reductions in the fire frequency in woodpecker habitat.
Ignition and Weather
Natural fires began as point ignitions that radiated out from a patch of dry vegetation struck by lightning. The rate of spread, flame length, intensity, and other characteristics of the spreading fire were affected by complex interactions involving weather (both past and present), topography, and the vegetation available for carrying the fire. If the weather was cool and humidity high, the flame length moving into prevailing winds likely would have gone out. If the weather was warm and conditions dry, the flame length moving with prevailing winds would have grown quickly in size and moved quickly across the landscape (Figure 12).
Fire managers study these relationships extensively as part of their early training (National Wildfire Coordinating Group, 2020). Unfortunately, the current fuel models used to predict fire behavior in Florida are based on western ecosystems and often lack the precision needed to create the burn heterogeneity recommended here for three of our focal species (as well as other rare species; Hill et al., 2017). Ground-based LiDAR imagery is being used to quantify grass biomass, hardwood stem densities, palmetto shrub cover, and other fuels with high accuracy in mature pineland ecosystems to help improve fuel models (K. Hiers, pers. comm.). Similar efforts are need for scrub and dry prairie habitat. With several years of added experience, fire managers may be able to use these models to fine-tune the outcome a prescribed fire is likely to have on ground-foraging species. Greater post-burn heterogeneity and the type and configuration of fire patches promote avian diversity in other fire-maintained systems (Docherty et al., 2020).
Improve Efficiency
While steps are being taken to improve the funding provided to agencies that burn public lands (Wildland Fire Leadership Council, 2020), better prioritization is needed now to ensure that fire management is as efficient as possible. Prioritization was stressed on EAFB to help manage these trade-offs. Research biologists and land managers developed a spatially referenced burn prioritization tool to guide the application of their limited prescribed fire resources each year (Hiers et al., 2003). Criteria evaluated included habitat requirements for woodpeckers and other rare species as well the time elapsed since the last burn. The tool evolved over time in an adaptive manner based on continued research and monitoring. Another simple method of prioritization is to focus on units within managed areas when frequency goals are not be met for many burn units. For example, St. Sebastian River Preserve State Park (9,000 ha; 27.82, −80.61) collaborated with Tall Timbers Research Station to establish a large ecological restoration effort covering a 2,000 ha area used by their small red-cockaded woodpecker population (6–10 breeding groups). Mechanical treatments were applied to reduce shrubs and fire has been applied consistently to this area using a 3-year return interval (and often with a trade-off of not burning other pinelands).
Future Challenges
Managing fire is staged to become increasingly complex under changing climatic conditions. Fire regimes that once excluded hardwood trees and shrubs may no longer perform this function in the face of higher year-round temperatures and increased CO2, a key plant nutrient. Shrub encroachment associated with this change has been noted throughout the grass-dominated biomes of North America and is not likely to abate (Knapp et al., 2008). Most fire-dependent grasslands evolved during periods with lower CO2 levels and were promoted by both fire and herbivory (Bond, 2008). Grassy biomes depend on the competitive advantages many grasses have below ground, the flammable fuels they provide for fire, and often the effects large native herbivores convey, which are nowhere to be found in Florida today (Bond, 2008).
Combinations of mechanical and fire treatments will be needed to maintain fire-dependent systems in Florida as both temperatures and humans increase. Sites supporting Florida scrub-jays (e.g., Marjorie Harris Carr Cross Florida Greenway; 29.02, −82.10; 1317 ha) are already surrounded by extensive infrastructure and housing (Figure 13). Land managers rarely burn 10 ha in a given year (L. Dolan, pers. comm.), but such small burns are less effective in controlling shrubs and providing conditions the local scrub-jay population needs. Instead, mechanical treatment of over-grown scrub is being used and led to a tripling of the population (Dolan, 2019). The cost of mechanical treatments is approximately $600 per ha compared to $60 per ha to burn an equivalent area (L. Dolan, pers. comm.), but maintaining even small scrub-jay populations is critically important to sustaining genetic diversity (Chen et al., 2018). This site also points to the increased management costs needed to manage thin corridors that contain fire-maintained habitat. Mechanical methods will also be needed to conduct burns safely in long unburned scrub (Schmalzer and Boyle, 1998). Most agree, however, that mechanical methods are not a substitute for fire and the best system responses occur when mechanical methods are primarily used in the initial phases of restoration then followed by fire. Mechanical treatments also carry risks of excessive soils disturbance with potential problems of soil compaction and spreading exotics.
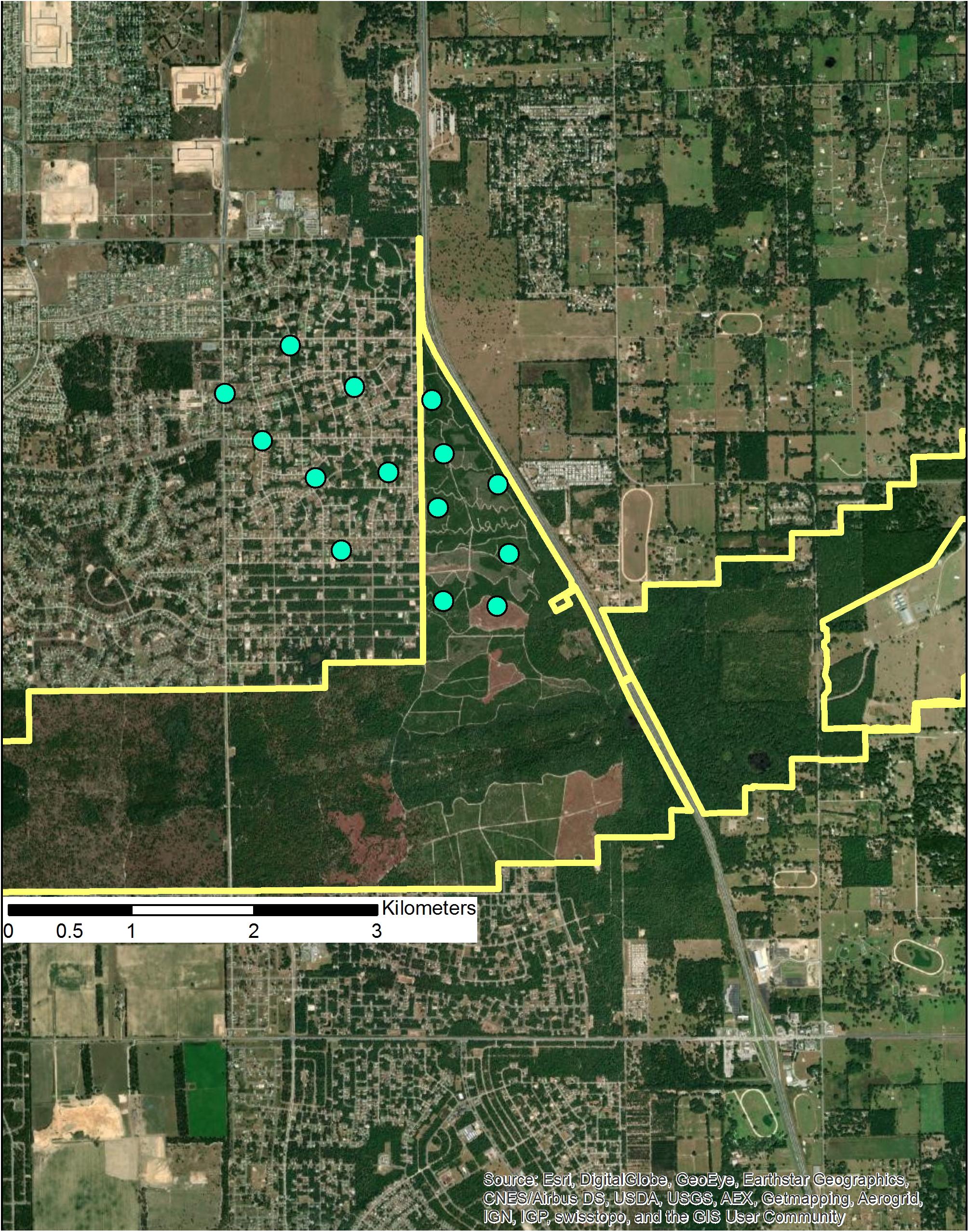
Figure 13. Florida scrub-jay habitat on the Marjorie Harris Carr Cross Florida Greenway (yellow polygon), Marion County, Florida. The centers of current and former scrub-jay territories are shown as blue dots. Residential housing and an interstate highway limit annual burns to ca. 10 ha (L. Dolan, pers. comm.).
Recommendations
Florida has provided remarkable support for wildlife conservation through the acquisition and management of natural areas (Knight et al., 2011). Under the Preservation 2000 Program, Florida invested $3 billion dollars in conservation land purchases over a 10-year period (Knight et al., 2011). To ensure these investments conserve rare fauna and flora for future generations, financial support for management is recognized as a key element of Florida’s land acquisition process. Funds are invested to manage habitat using fire, professional staff, and other tools. The end products are some of the best managed conservation lands anywhere. Regrettably, Florida’s human population also is projected to reach 33.7 million by 2070 (Carr and Zwick, 2006). The developed lands needed to support this growing population will increase from 18.7 to 33.7% of the terrestrial land base (ca. 28500 km2; Carr and Zwick, 2006) and means management tools must be developed to anticipate these future conditions. In light of this urgency, we provide six recommendations to maximize the effectiveness of fire management and information exchange on its use in Florida.
Focus on Frequency
Fire frequency affects habitat suitability for many rare species and needs to be a focus for management and research. Frequent application of fire makes future burns easier to conduct (Waldrop and Goodrick, 2012), inhibits changes in vegetation structure, and prevents wildfires. Even subtle shifts (i.e., ±1 year) in fire frequency can lead to changes in vegetation structure when carried out over several decades (Glitzenstein et al., 2003, 2012). If the fire frequencies rare species need are not being met across most of a managed area, managers should manipulate the other factors they control in an effort to increase fire frequency across as much of the area as they can (e.g., enlarge burn units, burn multiple units at once, or expand the timeframe over which are applied). These changes should be monitored and adjusted in an adaptive way.
Restore Ecological Burn Goals
Many agencies support ecological burning for rare species, but the funding provided to other agencies often is linked to hazardous fuel management to prevent wildfires. Fuel management burns are often postponed (i.e., fire frequency reduced) until vegetation becomes so dense that the entire burn unit can be cleared by a single fire. Conditions picked to burn also favor ignition patterns and weather conditions that clear the unit as thoroughly as possible. This is not beneficial to scrub-jays and other rare species but is now practiced regularly on many federal holdings (e.g., Merritt Island National Wildlife Refuge; 28.64, −80.71; 56,560 ha). Ecological burning for rare species needs to be performed more frequently than fuel-reduction burns. Consistent use of an ecologically based framework can also restore conditions for rare species when applied over many years (Steen et al., 2013). Simply establishing the ground conditions capable of carrying frequent fires can restore habitat for rare species decades later even on former agriculture lands (Cox and McCormick, 2016).
Prioritize Annually
Fire management programs face external challenges from tightened smoke and air quality regulations, increased urban-wildland interfaces, and insufficient funding. It is crucial to make burning as efficient as possible. Burn prioritization models (Hiers et al., 2003) should be applied wherever frequency goals are not being met. Prioritization ensures single-species management is applied where needed and ecosystem restoration occurs elsewhere (Hiers et al., 2003). Establishing focal areas where fire is applied regularly is another way to set priorities. Failure to prioritize may simply extend a mediocre job to a larger area.
Adaptive resource management is another method for evaluating management effectiveness based on species-specific responses to different management programs (Johnson et al., 2011). ARM also helps to bridge the research-implementation gap and help to avoid confrontation (e.g., Cape Sable seaside sparrow). Using population data to assess responses is straightforward for most of our focal species, but data for red-cockaded woodpeckers should link back to habitat management and not the population changes created by artificial cavities. A more appropriate metric for ARM would be the number of woodpecker territories not dependent on artificial cavities.
Develop Products for Practitioners
Peer-reviewed research can be difficult for practitioners to obtain and digest. These factors are often listed as a key source of information gaps between science and land managers (Habel et al., 2013). Exemplary recovery plans that distil research and provide clear management targets (e.g., U. S. Fish and Wildlife Service [USFWS], 2003) are needed for the Florida scrub-jay and other listed species that occupy multiple managed areas. Packaging research into management booklets also is imperative. Good examples are fire management guidelines for scrub (Florida Fish and Wildlife Conservation Commission [FFWCC], 2018), gopher tortoise (Florida Fish and Wildlife Conservation Commission [FFWCC], 2012), amphibian (Bailey et al., 2006), and breeding birds (Cox and Widener, 2008).
Promote Land Manager Tenure
Each burn is different; each burn provides new information. There are success stories in Florida where land managers apply fires frequently and rare species are doing well. A common pattern on these refugia is the presence of land managers with decades of site-specific experience (e.g., TLWMA, Jonathan Dickinson State Park; 27.00, −80.10 [4646 ha]). Long-tenured land managers may apply fires to individual burn units 6–10 times during their careers and amass site-specific knowledge that is impossible to transfer. Fire management crews for some agencies instead cover huge areas (sometimes multiple states) and promotion often means accepting an administrative position.
Species Working Groups
Land managers look to species-specific working groups for guidance. These forums enhance information exchange and the integration of new scientific data into decision-making processes. Increasing the frequency of in-person meetings also helps to foster relationships built on trust and respect. Working groups also have the capacity to focus on novel instruments for change, such as the strategic vision plan adopted for the Florida grasshopper sparrow (U. S. Fish and Wildlife Service [USFWS], 2019b). Coordination of action, comparison of results, and developing momentum also are more likely to occur when research biologists and land managers meet regularly.
The effectiveness of working groups is linked to the diversity and complexity of issues they address. The Florida grasshopper sparrow has a single working group (N = 15 stakeholders) focused on the management taking place on four public landholdings. In contrast, the Florida scrub-jay is supported by 198 managed areas (Table 1) that once were aligned with five different working groups (Florida Fish, and Wildlife Conservation Commission [FFWCC], 2020). Such diversity can lead to obstacles, but there may be ways to narrow the focus to encourage large, annual gatherings. For example, the All-Florida Red-cockaded Woodpecker Working Group also involves scores of different state and federal agencies, but the group has a narrow focus on woodpecker translocation rather than woodpecker habitat management. A scrub-jay group with a similarly narrow focus might convene to discuss scrub-jay translocations and other information exchange. Annual gatherings also often lead to unexpected discoveries (i.e., a site 40 km away has a grass seed harvester to loan) as well as creative solutions to bureaucratic roadblocks, lack of funding, equipment and crew needs, and staff training.
Summary
Fire represents one of the most ancient forms of land management practiced by humanity (Pyne, 1995). Fires set by ancient cultures differed from natural fires in their frequency, seasonality, and intensity (Pyne, 1995). In the absence of widespread wildfires, natural areas today must be managed using prescribed fires that also differ from natural fires in their frequency, seasonality, and intensity. Prescribed fires are critically needed to sustain habitat for fire-dependent species. They are also likely to perpetuate most, but perhaps not all, of the intricate ecological relationships that natural fire regimes once provided.
Author Contributions
Each author prepared a species account, compiled data from published and unpublished reports, edited survey questions, and edited earlier drafts of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the many scientists and land managers who shared their personal data and insights on fire management in Florida. We also appreciate the status information provided by Katy NeSmith and Mark Barrett. The pictures provided by Gary Knight, Rob Rossmanith, Greg Titus, and Tom Virzi also were extremely helpful.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.00267/full#supplementary-material
References
Allen, D. H. (1991). Constructing Artificial Red-Cockaded Woodpecker Cavities. Atlanta, GA: U. S. Department of Agriculture. doi: 10.2737/SE-GTR-73
Bailey, M. A., Holmes, J. N., Buhlmann, K. A., and Mitchell, J. C. (2006). Habitat Management Guidelines for Amphibians and Reptiles of the Southeastern United States. Montgomery, AL: Partners in Amphibian and Reptile Conservation Technical Publication HMG-2.
Baiser, B., Boulton, R. L., and Lockwood, J. L. (2008). Influence of water depth on nest success of the endangered Cape Sable seaside sparrow in the Florida Everglades. Anim. Cons. 11, 190–197. doi: 10.1111/j.1469-1795.2008.00167.x
Bass, O. L., and Kushlan, J. (1982). Status of the Cape Sable Sparrow. South Florida Research Center Report T-672. Homestead, FL: National Park Service.
Benscoter, A. M., Beerens, J. M., Pearlstine, L. G., and Romañach, S. S. (2019). Fire disturbance influences endangered Cape Sable Seaside Sparrow (Ammospiza maritima mirabilis) relative bird count. Cons. Sci. Pract. 1:e130. doi: 10.1111/csp2.130
Bond, W. J. (2008). What limits trees in C4 grasslands and savannas? Ann. Rev. Ecol. Evol. Syst. 39, 641–659. doi: 10.1146/annurev.ecolsys.39.110707.173411
Boughton, R., and Bowman, R. (2011). Statewide Assessment of Florida Scrub-Jays on MANAGED AREAS: A Comparison of Current Populations to the results of the 1992–93 Survey. Jacksonville, FL: US Fish and Wildlife Service.
Breckheimer, I., Haddad, N. M., Morris, W. F., Trainor, A. M., Fields, W. R., Jobe, R. T., et al. (2014). Defining and evaluating the umbrella species concept for conserving and restoring landscape connectivity. Cons. Biol. 28, 1584–1593. doi: 10.1111/cobi.12362
Breininger, D. R., Duncan, B., Eaton, M., Johnson, F., and Nichols, J. (2014a). Integrating land-cover modeling and adaptive management to conserve endangered species and reduce catastrophic fire risk. Land 3, 874–897. doi: 10.3390/land3030874
Breininger, D. R., Stolen, E., Carter, G., Oddy, D., and Legare, S. (2014b). Quantifying how territory quality and sociobiology affect recruitment to inform fire management. Anim. Cons. 17, 72–79. doi: 10.1111/acv.12059
Breininger, D. R., Duncan, B. W., and Dominy, N. J. (2002). Relationships between fire frequency and vegetation type in pine flatwoods of East-Central Florida. USA. Nat. Areas J. 22, 186–193.
Breininger, D. R., Foster, T. E., Carter, G. M., Duncan, B. W., Stolen, E., and Lyon, J. E. (2018). The effects of vegetative type, edges, fire history, rainfall, and management in fire-maintained habitat. Ecosphere 9:e02120. doi: 10.1002/ecs2.2120
Breininger, D. R., Larson, V. L., Duncan, B. W., Smith, R. B., Oddy, D. M., and Goodchild, M. F. (1995). Landscape patterns of Florida Scrub-jay habitat use and demographic success. Cons. Bilo. 9, 1442–1453. doi: 10.1046/j.1523-1739.1995.09061442.x
Breininger, D. R., Nichols, J. D., Carter, G. M., and Oddy, D. M. (2009). Habitat-specific breeder survival of Florida Scrub-Jays: inferences from multistate models. Ecology 90, 3180–3189. doi: 10.1890/08-1123.1
Breininger, D. R., Nichols, J. D., Duncan, B. W., Stolen, E. D., Carter, G. G., Hunt, D. K., et al. (2010). Multistate modeling of habitat dynamics: factors affecting Florida scrub transition probabilities. Ecology 91, 3354– 3364.
Breininger, D. R., Provancha, M. J., and Smith, R. B. (1991). Mapping Florida scrub jay habitat for purposes of land-use management. Photogrammetr. Eng. Remote Sens. 57, 1467–1147.
Breininger, D. R., and Schmalzer, P. A. (1990). Effects of fire and disturbance on plants and birds in a Florida oak/palmetto scrub community Am. Midl. Nat. 123, 64–74. doi: 10.2307/2425760
Breininger, D. R., Toland, B., Oddy, D. M., and Legare, M. L. (2006). Landcover characterizations and Florida scrub-jay (Aphelocoma coerulescens) population dynamics. Biol. Conserv. 128, 169–181. doi: 10.1016/j.biocon.2005.09.026
Bridges, E. L. (2006). “Landscape ecology of Florida dry prairie in the Kissimmee River region,” in The Land of Fire and Water: Proceedings of the Florida Dry Prairie Conference, ed. R. F. Noss, (DeLeon Springs, FL: E. O. Painter Printing Co), 14–42.
Carr, M. H., and Zwick, P. D. (2006). Florida 2070. Mapping Florida’s Future and Alternative Patterns of Development in 2070. Gainesville, FL: GeoPlan Center, University of Florida.
Carter, G. M., Legare, M. L., Breininger, D. R., and Oddy, D. M. (2007). Nocturnal nest predation: a potential obstacle to recovery of a Florida Scrub-jay population. J. Field Ornith. 78, 390–394. doi: 10.1111/j.1557-9263.2007.00127.x
Castellon, T. (2017). Florida Grasshopper Sparrow Management of Private Ranch Lands. Tallahassee, FL: The Wildlife Foundation of Florida, Inc.
Caughley, G. (1994). Directions in Conservation Biology. J. Anim. Ecol. 63, 215–244. doi: 10.2307/5542
Chen, N., Cosgrove, E. J., Bowman, R., Fitzpatrick, J. W., and Clark, A. G. (2016). Genomic consequences of population decline in the endangered Florida scrub-jay. Curr. Biol. 26, 2974–2979. doi: 10.1016/j.cub.2016.08.062
Chen, N., Cosgrove, E. J., Bowman, R., Fitzpatrick, J. W., and Clark, A. G. (2018). Genomic consequences of population decline in the endangered Florida scrub-jay. Curr. Biol. 26, 2974–2979.
Chiodi, A. M., Larkin, N. S., and Varner, J. M. (2018). An analysis of southeastern US prescribed burn weather windows: seasonal variability and El Niño associations. Int. J. Wildl. Fire 27, 176–189. doi: 10.1071/WF17132
Conner, R. N., Rudolph, D. C., and Walters, J. R. (2001). The Red-Cockaded Woodpecker: Surviving in a Fire-Maintained Ecosystem. Austin: University of Texas Press.
Copeyon, C. K. (1990). A technique for constructing cavities for the Red-cockaded Woodpecker. Wildl. Soc. Bull. 18, 303–311.
Coulon, A., Fitzpatrick, J. W., Bowman, R., and Lovette, I. J. (2010). Effects of habitat fragmentation on effective dispersal of Florida scrub-jays. Cons. Biol. 24, 1080–1088. doi: 10.1111/j.1523-1739.2009.01438.x
Cox, J. (2014). Notes from A Land Management Workshop Focused on the Florida Grasshopper Sparrow. Kenansville, FL: Three Lakes Wildlife Management, doi: 10.13140/RG.2.1.5187.4642
Cox, J., Kautz, R., MacLaughlin, M., and Gilbert, T. (1994). Closing the Gaps in Florida’s Wildlife Habitat Conservation System. Tallahassee: Florida Game and Fresh Water Fish Commission, doi: 10.13140/RG.2.1.1136.8800
Cox, J., and McCormick, J. K. (2016). New insights from an attempt to reintroduce Red-cockaded Woodpeckers in northern Florida. J. Field Orn. 87, 360–370. doi: 10.1111/jofo.12165
Cox, J., and Widener, B. (2008). Lightning-season burning: friend or foe of breeding birds. Tall Timbers Res. Station Misc. Publ. 17, 6–8. doi: 10.13140/RG.2.1.4282.6087
Curnutt, J. L., Mayer, A. L., Brooks, T. M., Manne, L., Bass, O. L. Jr., Fleming, D. M., et al. (1998). Population dynamics of the endangered Cape Sable seaside-sparrow. Anim. Cons. 1, 11–21. doi: 10.1111/j.1469-1795.1998.tb00221.x
Delany, M. F., Linda, S. B., Pranty, P., and Perkins, D. W. (2002). Density and reproductive success of Florida grasshopper sparrows following fire. J. Range Manag. 55, 336–340.
Delany, M. F., Matthew, B., Shumar, M. B., McDermott, M. E., Kubilis, P. S., Hatchitt, J. L., et al. (2007). Florida grasshopper sparrow distribution, abundance, and habitat availability. Southeast. Nat. 6, 15–26. doi: 10.1656/1528-7092(2007)6[15:fgsdaa]2.0.co;2
Docherty, T. D. S., Hethcoat, M. G., MacTavish, L. M., MacTavish, D., Dell, S., Stephens, P. A., et al. (2020). Burning savanna for avian species richness, and functional diversity. Ecol. Appl. 30:e02091. doi: 10.1002/eap.2091
Dolan, L. (2019). Managing Florida scrub-jay habitat. Florida Park Service. Available at: https://www.floridastateparks.org/learn/managing-florida-scrub-jay-habitat (accessed January 15, 2020).
Driscoll, D. A., Lindenmayer, D. B., Bennett, A. F., Bode, M., Bradstock, R. A., Cary, G. J., et al. (2010). Fire management for biodiversity conservation: key research questions and our capacity to answer them. Biol. Cons. 143, 1928–1939. doi: 10.1016/j.biocon.2010.05.026
Duncan, B. W., Adrian, F. W., and Stolen, E. D. (2010). Isolating the lightning-ignition regime from a contemporary background fire regime in east-central Florida. USA. Can. J. For. Res. 40, 286–297. doi: 10.1139/X09-193
Duncan, B. W., and Schmalzer, P. A. (2004). Anthropogenic influences on potential fire spread in a pyrogenic ecosystem of Florida. USA. Landscape Ecol. 19, 153–165. doi: 10.1023/B:LAND.0000021714.97148.ac
Duncan, B. W., Schmalzer, P. A., Breininger, D. R., and Stolen, E. D. (2015). Comparing fuels reduction and patch mosaic fire regimes for reducing fire spread potential: a spatial modeling approach. Ecol. Mod. 314, 90–99. doi: 10.1016/j.ecolmodel.2015.07.013
Endries, M., Stys, B., Mohr, G., Kratimenos, G., Langley, S., Root, K., et al. (2009). Wildlife Habitat Conservation Needs in Florida. Updated Recommendations for Strategic Habitat Conservation Areas. Fish and Wildlife Research Institute Technical Report TR-15. Tallahassee, FL: Fish and Wildlife Research Institute.
Engstrom, R. T. (1993). Characteristic mammals and birds of longleaf pine forests. Proc. Tall Timbers Fire Ecol. Conf. 18, 127–138.
Engstrom, R. T. (2010). First-order fire effects on animals: review and recommendations. Fire Ecol. 6, 115–130. doi: 10.4996/fireecology.0601115
Everglades Fire Management Plan [ENP] (2015). National Park Service, U. S. Department of the Interior. Washington, DC: ENP, 356.
Fitzpatrick, J. W., and Bowman, R. (2016). Florida Scrub-Jays: Oversized Territories and Group Defense in a Fire-Maintained Habitat. Cooperative Breeding in Vertebrates: Studies Of Ecology, Evolution, and Behavior. Cambridge: Cambridge University Press, 77–96.
Fitzpatrick, J. W., Woolfenden, G. E., and Kopeny, M. T. (1991). Ecology and Development-Related Habitat Requirements of the Florida Scrub Jay (Aphelocoma coerulescens coerulescens). Tallahassee, IL: Florida Fish and Wildlife Conservation Commission.
Florida Fish and Wildlife Conservation Commission [FFWCC] (2009). Red-Cockaded Woodpecker Management Plan. Available online at: https://myfwc.com/media/2046/rcw-plan-only.pdf (accessed June 2020)
Florida Fish and Wildlife Conservation Commission [FFWCC] (2012). Gopher Tortoise Management Plan. Available online at https://myfwc.com/media/1819/gt-management-plan.pdf
Florida Fish and Wildlife Conservation Commission [FFWCC] (2018). Scrub Management Guidelines for Peninsular Florida: Using the Scrub-Jay as an Umbrella Species. Available at: https://www.partnersinflight.org/wp-content/uploads/2017/03/Partners-in-Flight-Scrub-Mgmt-Guidelines-for-Peninsular-Florida.pdf (accessed January 15, 2020).
Florida Fish, and Wildlife Conservation Commission [FFWCC] (2020). Florida Scrub-jay Working Group. Available at: https://fsjconservation.wordpress.com/working-groups/ (accessed December 2019).
Florida Natural Areas Inventory [FNAI] (2010). Guide to the Natural Communities of Florida: 2010. Edition. Tallahassee, FL: FNAI.
Florida Natural Areas Inventory [FNAI] (2019). Summary of Florida Conservation Lands. Tallahassee, FL: FNAI.
Franzreb, K. E., and Zarnoch, S. J. (2011). Factors affecting Florida scrub-jay nest survival on Ocala National Forest. Florida. J. Wildl. Manag. 75, 1040–1050. doi: 10.1002/jwmg.139
Frost, C. (2007). “History and future of the longleaf pine ecosystem,” in The Longleaf Pine Ecosystem. Springer Series on Environmental Management, eds S. Jose, E. Jokela, and D. Miller, (New York, NY: Springer), 9–48. doi: 10.1007/978-0-387-30687-2_2
Gabrey, S. W., and Afton, A. D. (2000). Effects of winter marsh burning on abundance and nesting activity of Louisiana seaside sparrows in the Gulf Coast Chenier Plain. Wilson J. Ornith. 112, 365–372. doi: 10.1676/0043-5643(2000)112[0365:eowmbo]2.0.co;2
Garabedian, J. E., Moorman, C. E., Peterson, M. N., and Kilgo, J. C. (2014). Systematic review of the influence of foraging habitat on red-cockaded woodpecker reproductive success. Wild. Biol. 20, 37–46. doi: 10.2981/wlb.13004
Glitzenstein, J. S., Streng, D. R., Masters, R. E., Robertson, K. M., and Hermann, S. M. (2012). Fire-frequency effects on vegetation in north Florida pinelands: another look at the long-term Stoddard Fire Research Plots at Tall Timbers Research Station. For. Ecol. Manag. 264, 197–209. doi: 10.1016/j.foreco.2011.10.014
Glitzenstein, J. S., Streng, D. R., and Wade, D. D. (2003). Fire frequency effects on longleaf pine (Pinus palustris) vegetation in South Carolina and northeast Florida, USA. Nat. Areas J. 23, 22–37.
Gunderson, L. H., and Snyder, J. R. (1994). “Fire patterns in the southern Everglades,” in Everglades: The Ecosystem and Its Restoration, eds S. M. Davis and J. C. Ogden, (Delray Beach, FL: St. Lucie Press), 291–305.
Habel, J. C., Gossner, M. M., Meyer, S. T., Eggermont, H., Jurge, L., Weisser, D., et al. (2013). Mind the gaps when using science to address conservation concerns. Biodivers. Conserv. 22, 2413–2427. doi: 10.1007/s10531-013-0536-y
Hanan, E. J., Ross, M. S., Ruiz, P. L., and Sah, J. P. (2010). Multi-scaled grassland-woody plant dynamics in the heterogeneous marl prairies of the southern Everglades. Ecosystems 13, 1256–1274. doi: 10.1007/s10021-010-9386-6
Harris, S. N., Doonan, T. L., Hewett Ragheb, E. L., and Jachowski, D. S. (2019). Den site selection by the Florida Spotted Skunk. J. Wildl. Manag. 84, 127–137. doi: 10.1002/jwmg.21780
Hess, C. A. (2014). Restoration of longleaf pine in slash pine plantations: using fire to avoid the landscape trap. PhD thesis, Florida State University, Tallahassee, FL.
Hess, C. A., and Tschinkel, W. R. (2017). Effect of thinning and clear-cuts on the transmission of fire in slash pine plantations during restoration to longleaf pine. Ecol. Restor. 35, 33–40. doi: 10.3368/er.35.1.33
Hewett Ragheb, E. L., Miller, K. E., and Hoerl Leone, E. (2019a). Exclosure fences around nests of imperiled Florida Grasshopper Sparrows reduce rates of predation by mammals. J. Field Ornith. 90, 309–324. doi: 10.1111/jofo.12310
Hewett Ragheb, E. L., Miller, K. E., and Kiltie, R. A. (2019b). Optimizing reproductive opportunity for Florida grasshopper sparrows: when to burn? J. Wildl. Manag. 83, 544–555. doi: 10.1002/jwmg.21622
Hiers, J. K., Laine, S. C., Bachant, J. J., Furman, J. H., Greene, W. W. Jr., Compton, V., et al. (2003). Simple spatial modeling tool for prioritizing prescribed burning activities at the landscape scale. Cons. Biol. 17, 1571–1578. doi: 10.1111/j.1523-1739.2003.00381.x
Hill, K. C., Bakker, J. D., and Dunwiddie, P. W. (2017). Prescribed fire in grassland butterfly habitat: targeting weather and fuel conditions to reduce soil temperatures and burn severity. Fire Ecol. 13, 24–41. doi: 10.4996/fireecology.130302441
Hovis, J., and Swan, D. (2004). “The red-cockaded woodpecker in Florida: a 2000 update,” in Red-Cockaded Woodpecker: Road to Recovery, eds S. Daniels and R. Costa, (Blaine, WA: Hancock House), 254–262.
Huffman, J. (2006). Historical Fire Regimes In Southeastern Pine Savannas. Dissertation. Louisiana State University, Baton Rouge, LA.
Huffman, J., and Platt, W. (2014). Fire history of the Avon Park Air Force Range: evidence from tree-rings: final report to Department of Defense, Avon Park Air Force Range. Available at: https://talltimbers.org/wp-content/uploads/2016/11/Huffman-and-Platt-2014-Avon-Park-Report_reduced.pdf (accessed December 19, 2014).
Jackson, J. A. (1994). “Red-cockaded Woodpecker (Dryobates borealis), version 2.0,” in The Birds of North America, eds A. F. Poole and F. B. Gill, (Ithaca (NY): Cornell Lab of Ornithology).
James, F. C., Hess, C. A., and Kufrin, D. (1997). Species-centered environmental analysis: Indirect effects of fire history on red-cockaded woodpeckers. Ecol. Appl. 7, 118–129. doi: 10.1890/1051-0761(1997)007[0118:sceaie]2.0.co;2
Jenkins, C. N., Powell, R. D., Bass, O. L. Jr., and Pimm, S. L. (2003). Demonstrating the destruction of the habitat of the Cape Sable seaside sparrow (Ammodramus maritimus mirabilis). Anim. Conserv. 6, 29–30. doi: 10.1017/S1367943003003056
Johnson, F. A., Breininger, D. R., Duncan, B. W., Nichols, J. D., Runge, M. A., and Williams, B. K. (2011). A Markov decision process for managing habitat for Florida Scrub-Jays. J. Fish Wildl. Manag. 2, 234–246. doi: 10.3996/012011-JFWM-003
Jones, C. D., Cox, J. A., Toriani-Moura, E., and Cooper, R. J. (2013). Nest-site characteristics of Bachman’s Sparrows and their relationship to plant succession following prescribed burns. Wilson J. Ornith. 125, 293–300. doi: 10.1676/12-119.1
Jones, D. D., Conner, L. M., Storey, T. H., and Warren, R. J. (2004). Prescribed fire and raccoon use of longleaf pine forests: implications for managing nest predation? Wildl. Soc. Bull. 32, 1255–1259. doi: 10.2193/0091-7648(2004)032[1255:pfaruo]2.0.co;2
Jose, S., Jokela, J. E., and Miller, D. L. (eds) (2006). The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration. New York, NY: Springer-Verlag, doi: 10.1007/978-0-387-30687-2_1
Kautz, R., Stys, B., and Kawula, R. (2007). Florida vegetation 2003 and land use changes between 1985-89 and 2003. Fl. Sci. 70, 12–23.
Kern, R. A., and Shriver, W. G. (2014). Sea level rise and prescribed fire management: Implications for Seaside Sparrow population viability. Biol. Cons. 173, 24–31. doi: 10.1016/j.biocon.2014.03.007
Kirkman, L. K., and Giencke, L. M. (2018). “Restoring and managing a diverse ground cover,” in Ecological Restoration and Management of Longleaf Pine Forests, eds K. Kirkman and S. B. Jack, (Boca Raton, FL: CRC Press), 207–232. doi: 10.1201/9781315152141-11
Kirkman, L. K., and Jack, S. B. (eds) (2018). Ecological Restoration and Management of Longleaf Pine Forests. Boca Raton, FL: CRC Press.
Kirkman, L. K., Jack, S. B., and Melvin, R. K. (2018). “The fire forest of the past and present,” in Ecological Restoration and Management of Longleaf Pine Forests, eds K. Kirkman and S. B. Jack, (Boca Raton, FL: CRC Press), 3–13.
Knapp, A. K., Briggs, J. M., Collins, S. L., Archer, S. R., Bret-Harte, M. S., Ewers, B. E., et al. (2008). Shrub encroachment in North American grasslands: shifts in growth form dominance rapidly alters control of ecosystem carbon inputs. Glob. Change Biol. 14, 615–623. doi: 10.1111/j.1365-2486.2007.01512.x
Knight, G. R., Oetting, J. B., and Cross, L. (2011). Atlas of Florida’s Natural Heritage: Biodiversity, Landscapes, Stewardship, and Opportunities. Tallahassee, FL: Institute of Science and Public Affairs, Florida State University.
Komarek, E. V. (1968). Lightning and lightning fires as ecological forces. Proc. Tall Timbers Fire Ecol. Conf. 8, 169–197.
La Puma, D. A., Lockwood, J. L., and Davis, M. J. (2007). Endangered species management requires a new look at the benefit of fire: the Cape Sable seaside sparrow in the Everglades ecosystem. Biol. Cons. 136, 398–407. doi: 10.1016/j.biocon.2006.12.005
Lashley, M. S., Chitwood, M. C., Harper, C. A., DePerno, C. A., and Moorman, C. E. (2015). Variability in fire prescriptions to promote wildlife foods in the longleaf pine ecosystem. Fire Ecol. 11, 62–79. doi: 10.4996/fireecology.1103062
Lockwood, J. L., Fenn, K. H., Caudill, J. M., Okines, D., Bass, O. L. Jr., Duncan, J. R., et al. (2001). The implications of Cape Sable seaside sparrow demography for Everglades restoration. Anim. Cons. 4, 275–281. doi: 10.1017/S1367943001001329
Lockwood, J. L., Ross, M. S., and Sah, J. P. (2003). Smoke on the water: the interplay of fire and water flow on Everglades restoration. Front. Ecol. Env. 1:462–468. doi: 10.2307/3868113
Loudermilk, E. L., Hiers, J. K., and O’Brien, J. J. (2018). “The role of fuels for understanding fire behavior and fire effects,” in Ecological Restoration and Management of Longleaf Pine Forests, eds L. K. Kirkman and S. B. Jack, (Boca Raton, FL: CRC Press, Taylor & Francis Group), 107–122. doi: 10.1201/9781315152141-6
Melvin, M. A. (2018). National Prescribed Fire Use Survey Report. Washington, DC: Coalition of Prescribed Fire Councils, Inc.
Menges, E. S., Craddock, A., Salo, J., Zinthefer, R., and Weekley, C. W. (2008). Gap ecology in Florida scrub: species occurrence, diversity and gap properties. J. Veg. Sci. 19, 503–514. doi: 10.3170/2008-8-18399
Menges, E. S., and Gordon, D. R. (2010). Should mechanical treatments and herbicides be used as fire surrogates to manage Florida’s uplands? A review. Florida Sci. 73:147.
Menges, E. S., and Hawkes, C. V. (1998). Interactive effects of fire and microhabitat on plants of Florida scrub. Ecol. Appl. 8, 935–946. doi: 10.1890/1051-0761(1998)008[0935:ieofam]2.0.co;2
Miller, S. R., Emanuel, B. W., and Corby, M. K. (2004). “Prescribed fire at the St. Johns River Water Management District: what can be accomplished when fire is job one?,” in Proceedings of the Tall Timbers Fire Ecology Conference, (Tallahassee, FL: Tall Timbers Research Station).
Mitchell, R. J., Hiers, J. K., O’Brien, J. J., Jack, S. B., and Engstrom, R. T. (2010). Silviculture that sustains: the nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Can. J. For. 40, 596–596. doi: 10.1139/x06-100
Mumme, R. L. (1992). Do helpers increase reproductive success - an experimental-analysis in the Florida Scrub-jay. Behav. Ecol. Sociol. 31, 319–328. doi: 10.1007/BF00177772
National Oceanic and Atmospheric Administration [NOAA] (2019). Melbourne Lightning Statistics. Brevard County, FL: NOAA.
National Wildfire Coordinating Group (2020). S-130, Firefighter Training. Available at: https://www.nwcg.gov/publications/training-courses/s-130/course-materials (accessed January 2020).
Nowell, H. K., Holmes, C. D., Robertson, K., Teske, C., and Hiers, J. K. (2018). A new picture of fire extent, variability, and drought interaction in prescribed fire landscapes: insights from Florida government records. Geophy. Res. Lett. 45, 7874–7884. doi: 10.1029/2018GL078679
Oswalt, C. M., Cooper, J. A., Brockway, D. G., Brooks, H. W., Walker, J. L., Connor, K. F., et al. (2012). History and current condition of longleaf pine in the southern United States. Scotts Valley, CA: CreateSpace Independent Publishing Platform.
Petrick, C. J., and Hagedorn, B. W. (2004). “Population status and trend of red-cockaded woodpeckers on Eglin Air Force Base, Florida,” in Red-Cockaded Woodpecker: Road to Recovery, eds S. Daniels and R. Costa, (Blaine, WA: Hancock House), 254–262.
Pimm, S. L., Lockwood, J. L., Jenkins, C. N., Curnutt, J. L., Nott, M. P., Powell, E. D., et al. (2002). Sparrow in the Grass: A Report on the First Ten Years of Research on the Cape Sable Seaside Sparrow (Ammodramus maritimus mirabilis). Homestead, FL: Everglades National Park.
Platt, W. J., Ellair, D. P., Huffman, J. M., Potts, S. E., and Beckage, B. (2016). Pyrogenic fuels produced by savanna trees can engineer humid savannas. Ecol. Monogr. 86, 352–372. doi: 10.1002/ecm.1224
Platt, W. J., Huffman, J. M., Slocum, M. G., and Beckage, B. (2006). “Fire regimes and trees in Florida dry prairie landscapes,” in The Land of Fire and Water: Proc. Florida Dry Prairie Conf, ed. R. F. Noss, (DeLeon Springs, FL: E. O. Painter Printing Co), 3–13.
Post, W., and Greenlaw, J. (2018). “Seaside Sparrow (Ammodramus maritimus). version 2.1,” in The Birds of North America, ed. P. G. Rodewald, (Ithaca, NY: Cornell Lab of Ornithology).
Post, W., Greenlaw, J. S., Merriam, T., and Wood, L. (1983). “Comparative ecology of northern and southern populations of the Seaside Sparrow,” in Proceedings of the Seaside Sparrow: Its Biology and Management. Occasional Papers of the North Carolina Biological Survey, Raleigh, 123–136.
Pranty, B., and Tucker, J. W. (2006). “Ecology and management of the Florida grasshopper sparrow,” in The Land of Fire and Water: Proceedings of the Florida Dry Prairie Conferrence, ed. R. F. Noss, (DeLeon Springs, FL: E. O. Painter Printing Co), 188–200.
Pyne, S. J. (1995). World Fire: The Culture of Fire on Earth. New York, NY: Henry Holt, doi: 10.1023/A:1005371923658
Robbins, L. E., and Myers, R. L. (1992). “Seasonal effects of prescribed burning in Florida: a review,” in Miscellaneous Publication 8, (Tallahassee, FL: Tall Timbers Research Station), 95.
Rudolph, D. C., Conner, R. N., and Turner, J. (1990). Competition for red-cockaded woodpecker roost and nest cavities: effects of resin age and entrance diameter. Wilson Bull. 102, 23–36.
Sah, J. P., Ross, M., Snyder, J. R., Ruiz, P. L., Stofella, S., Colbert, N., et al. (2010). Cape Sable Seaside Sparrow habitat–vegetation monitoring: Fiscal year 2009 Final Report. Homestead, FL: S. E. R. Center, 91.
Sah, J. P., Ross, M. S., and Ruiz, P. L. (2013). Landscape Pattern–Marl Prairie/Slough Gradient: Vegetation Composition Along the Gradient and Decadal Vegetation Change Pattern in Shark Slough: Annual Report 2012. Southeast Environmental Research Center Report 101. Miami, FL: Southeast Environmental Research Center.
Sah, J. P., Ross, M. S., Saha, S., Minchin, P., and Sadle, J. (2014). Trajectories of vegetation response to water management in Taylor Slough, Everglades National Park, Florida. Wetlands 34, 65–79. doi: 10.1007/s13157-013-0390-4
Schmalzer, P. A., and Boyle, S. (1998). Restoring long-unburned oak-mesic flatwoods requires mechanical cutting and prescribed burning. Restor. Manag. Notes 16, 96–97.
Schmalzer, P. A., Boyle, S. R., and Swain, H. M. (1999). Scrub ecosystems of Brevard County, Florida: a regional characterization. Florida Sci. 62, 13–47.
Schmalzer, P. A., and Hinkle, C. R. (1992). Recovery of oak-saw palmetto scrub after fire. Castanea 57, 158–173.
Shriver, W. G., and Vickery, P. D. (2001). Response of breeding Florida grasshopper and Bachman’s sparrows to winter prescribed burning. J. Wildl. Manag 65, 470–475. doi: 10.2307/3803099
Simberloff, D. (1988). The contribution of population and community biology to conservation science. Ann. Rev. Ecol. Syst. 19, 473–511. doi: 10.1146/annurev.es.19.110188.002353
Snyder, J. R., and Barrios, B. (2008). “The use of microburns to study seasonal effects of fire on Cape Sable Seaside Sparrow habitat,” in Proceedings of the USGS Gulf Coast Science Conference and Florida Integrated Science Center Meeting Orlando, (Florida: U. S. Department of Interior).
Steen, D. A., Conner, L. M., Smith, L. L., Provencher, L., Hiers, J. K., Pokswinski, S., et al. (2013). Bird assemblage response to restoration of fire-suppressed longleaf pine sandhills. Ecol. App. 23, 134–147. doi: 10.1890/12-0197.1
Stein, B. A. (2002). States of the Union: Ranking America’s Biodiversity. Arlington, VI: NatureServe.
Taylor, D. L. (1993). “Fire management and the Cape Sable seaside sparrow,” in The Seaside Sparrow; Its Biology and Management, eds T. L. Quay, J. B. Funderburg, D. S. Lee, E. F. Potter, and C. S. Robbins, (Raleigh, NC: North Carolina Biological Survey), 147–152.
Tucker, J. W., Schrott, G. R., and Bowman, R. (2010). Fire ants, cattle grazing, and the endangered Florida grasshopper sparrow. Southeast. Nat. 9, 237–250. doi: 10.1656/058.009.0203
U. S. Department of Agriculture and Forest Service [USDA-FS] (1999). Revised Land and Resource Management Plan for the National Forests in Florida. Washington, DC: U. S. Department of Agriculture, Forest Service.
U. S. Fish and Wildlife Service [USFWS] (1973). Endangered Species Act of 1973. Washington, D.C: U. S. Fish and Wildlife Service.
U. S. Fish and Wildlife Service [USFWS] (1999). Florida Grasshopper Sparrow. Vero Beach, FL: Multi-species recovery plan for South Florida.
U. S. Fish and Wildlife Service [USFWS] (2003). Recovery Plan for the Red-Cockaded Woodpecker (Picoides borealis): Second Revision. Atlanta, GA: U. S. Fish and Wildlife Service, 296.
U. S. Fish and Wildlife Service [USFWS] (2019a). Environmental Conservation Online System. Atlanta, GA: U. S. Fish and Wildlife Service.
U. S. Fish and Wildlife Service [USFWS] (2019b). Recovery Plan for Cape Sable seaside sparrow (Ammodramus maritimus mirabilis). Atlanta, GA: U. S. Fish and Wildlife Service.
U. S. Fish and Wildlife Service [USFWS] (2019c). Draft Revised Recovery Plan for the Florida scrub-jay (Aphelocoma coerulescens). Atlanta, GA: U. S. Fish and Wildlife Service.
U. S. Fish and Wildlife Service [USFWS] (2019d). Florida Grasshopper Sparrow 5-Year Strategic Vision – A Recovery Implementation Strategy. Atlanta, GA: United States Fish and Wildlife Service.
Van Balen, J. B., and Doerr, P. D. (1978). The relationship of understory vegetation to red-cockaded woodpecker activity. Proc. Ann. Conf. Southeast. Assoc. Fish Wildl. Agencies 32, 82–92.
Venne, L. S., Trexler, J. C., and Frederick, P. C. (2016). Prescribed burn creates pulsed effects on a wetland aquatic community. Hydrobiologia 771, 281–295. doi: 10.1007/s10750-016-2640-y
Vickery, P. D. (2020). “Grasshopper Sparrow (Ammodramus savannarum), version 1.0,” in Birds of the World, eds A. F. Poole and F. B. Gill, (Ithaca, NY: Cornell Lab of Ornithology), doi: 10.2173/bow.graspa.01
Waldrop, T. A., and Goodrick, S. L. (2012). Introduction to Prescribed Fires in Southern Ecosystems. Science Update SRS-054, Vol. 54. Asheville, NC: US Department of Agriculture Forest Service, Southern Research Station, 1–80.
Waldrop, T. A., White, D. L., and Jones, S. M. (1992). Fire regimes for pine-grassland communities in the southeastern United States. Forest Ecol. Manag. 47, 195–210. doi: 10.1016/0378-1127(92)90274-d
Walsh, P. B., Darrow, D. A., and Dyess, J. G. (1995). Habitat selection by Florida grasshopper sparrows in response to fire. Proc. Ann. Conf. Southeast. Assoc. Fish Wildl. Agenc. 49, 340–347.
Walters, J. R., Beissinger, S. R., Fitzpatrick, J. W., Greenberg, R., Nichols, J. D., Pulliam, H. R., et al. (2000). The AOU Conservation Committee Review of the biology, status, and management of Cape Sable seaside sparrows: Final Report. Auk 117, 1093–1115. doi: 10.1093/auk/117.4.1093
Walters, J. R., Doerr, P. D., and Carter, J. H. III (1988). The cooperative breeding system of the red-cockaded woodpecker. Ethology 78, 275–305. doi: 10.1111/j.1439-0310.1988.tb00239.x
Weiss, S. A., Toman, E. L., and Corace, R. G. (2019). Aligning endangered species management with fire-dependent ecosystem restoration: manager perspectives on red-cockaded woodpecker and longleaf pine management actions. Fire Ecol. 15, 19. doi: 10.1186/s42408-019-0026-z
Werner, H. W., and Woolfenden, G. E. (1983). “The Cape Sable Seaside Sparrow: Its habitat, habits and history,” in The Seaside Sparrow. its Biology and Management, eds T. L. Quay, J. B. Funderberg, D. S. Lee, E. F. Potter, and C. S. Robbins, (Raleigh, NC: North Carolina Biological Survey and North Carlina State Museum), 55–75.
Wildland Fire Leadership Council (2020). Summit with Secretary of Agriculture Sonny Perdue. Salinas, CA: Wildland Fire Leadership Council.
Willcox, E. V., and Giuliano, W. M. (2010). Seasonal effects of prescribed burning and roller chopping on saw palmetto in flatwoods. For. Ecol. Manag. 259, 1580–1585. doi: 10.1016/j.foreco.2010.01.034
Williams, B. K., Eaton, M. J., and Breininger, D. R. (2011). Adaptive resource management and the value of information. Ecol. Model. 22218, 3429–3436. doi: 10.1016/j.ecolmodel.2011.07.003
Williams, B. W., Moser, E. B., Hiers, J. K., Gault, K., and Thurber, D. K. (2006). Protecting red-cockaded woodpecker cavity trees predisposed to fire-induced mortality. J. Wildl. Manag. 70, 702–707. doi: 10.2193/0022-541x(2006)70[702:prwctp]2.0.co;2
Williams, T. (2013). The Most Endangered Bird in the Continental U. S.: The fight to save the Florida Grasshopper Sparrow Inspires All Who Love Wildlife. Manhattan, NY: Audubon Magazine.
Woolfenden, G. E., and Fitzpatrick, J. W. (1984). The Florida Scrub Jay: Demography of a Cooperative-Breeding Bird. Princeton NJ: Princeton University Press.
Keywords: Cape Sable seaside sparrow, Florida grasshopper sparrow, Florida scrub-jay, prescribed fire, red-cockaded woodpecker, research-implementation gap
Citation: Cox JA, Engstrom RT, Breininger DR and Hewett Ragheb EL (2020) Interpreting Smoke Signals: Fire Ecology and Land Management for Four Federally Listed Birds. Front. Ecol. Evol. 8:267. doi: 10.3389/fevo.2020.00267
Received: 09 January 2020; Accepted: 28 July 2020;
Published: 15 September 2020.
Edited by:
Adrián Regos, Universidade de Santiago de Compostela, SpainReviewed by:
Jennifer Lesley Silcock, The University of Queensland, AustraliaKaya Lee Klop-Toker, The University of Newcastle, Australia
Copyright © 2020 Cox, Engstrom, Breininger and Hewett Ragheb. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: James A. Cox, amNveEB0YWxsdGltYmVycy5vcmc=