 Birgit Szabo
Birgit Szabo Isabel Damas-Moreira
Isabel Damas-Moreira Martin J. Whiting
Martin J. Whiting- Department of Biological Sciences, Macquarie University, Sydney, NSW, Australia
Invasive species are a global conservation problem that have an enormous economic cost. Understanding the attributes of invasive species and what makes them successful at colonizing and flourishing in novel environments is therefore essential for preventing and ameliorating their negative impact. Learning ability and behavioral flexibility—the ability to adjust behavior flexibly when conditions change including to learn to solve novel problems or existing problems in a novel way, are thought to play a key role during invasions although cognitive ability is rarely considered in studies of invasive species. We begin by reviewing the evidence that flexible learning and problem solving can influence invasion success in both invertebrates and vertebrates. We also review brain size as an index of cognitive ability with respect to invasion success. We then focus on the specific attributes of cognition that are likely to be important for species entering novel environments as they learn the location of resources (e.g., food, shelter), and as they encounter and interact with conspecifics, heterospecifics, and potential predators. We suggest that enhanced spatial learning ability in conjunction with behavioral flexibility are likely to be adaptive. Furthermore, good memory retention and the ability to learn from others (both conspecifics and heterospecifics) are beneficial. Finally, we suggest future directions for studying the link between cognition, fitness, and invasion success. Studies of closely related “invasive” and “non-invasive” species, as well as invasive populations and their source, should provide important baseline information about the potential role of cognitive ability in determining invasion success. We also advocate an experimental approach. In particular, we borrow methods from experimental evolutionary ecology. We suggest that experimental studies in which potential invasive species can be assayed for behavior and their cognitive ability measured prior to population-level release on small islands will help inform us about the potential role of cognitive ability in determining the fitness of invasive species. The idea that cognitive ability may determine invasion success is only now starting to gain traction. This is a rich field worthy of further study that will help us better understand what makes a successful invasive species.
Introduction
Human society is changing the world at an alarming rate. Overpopulation, globalization, and climate change have led to unprecedented levels of habitat transformation resulting in negative effects on species, populations, and entire ecosystems. Consequently, many species living in altered environments either need to rapidly adapt or potentially face local extirpation (WWF, 2018). Conversely, some organisms may benefit from anthropogenic environments and human activity. For example, some species can take advantage of the existing vast global transport networks and use it to hitchhike and colonize new locations and thereby expand their range (Chapple et al., 2012; Lockwood et al., 2013; Lewis et al., 2016). This invasion process is complex, and encompasses multiple stages. Each stage poses a different set of obstacles that animals need to overcome in order to become invasive (Blackburn et al., 2011). A species that fails at any stage of the process will thus not become invasive (Figure 1). Nevertheless, biological invasions have been steadily increasing in the last century (Essl et al., 2011; Lewis et al., 2016), and therefore, understanding why some species are successful invaders is of great interest.
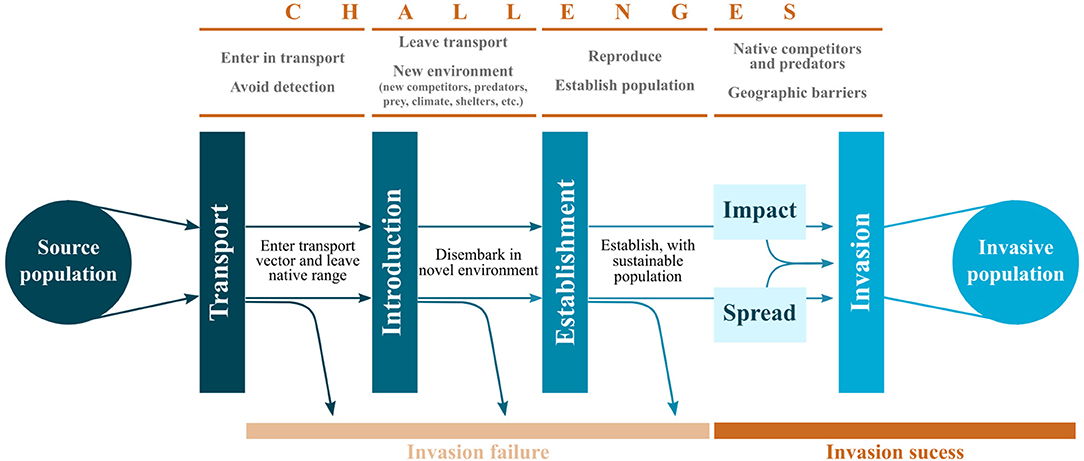
Figure 1. The resulting invasive population consists of the filtered individuals that were able to overcome all obstacles in all stages successfully. Based on Colautti and MacIsaac (2004), Blackburn et al. (2011), and Chapple et al. (2012).
Whether an organism needs to adapt to changes in their current environment, or whether they are introduced into a completely novel environment, they need to be able to adapt or change their behavior quickly in order to survive (Tuomainen and Candolin, 2011). While selection may favor particular phenotypes and behavioral types, the degree to which an organism can plasticly adjust behavior (behavioral plasticity) may also be crucial in determining the outcome of an invasion (Chapple et al., 2012; Griffin et al., 2016). In addition to behavioral plasticity, cognitive ability can plausibly give some species an advantage in the invasion process because an ability to use resources and to learn and retain the location of resources and threats should be adaptive in novel environments (Shettleworth, 2001, 2010). It is thus not surprising that cognitive ability is increasingly being recognized for the potentially important role it may play in selection affecting fitness (Cole et al., 2012; Thornton et al., 2014; Budaev et al., 2019), which highlights the importance it can have on the success of invaders.
Behavioral flexibility–the ability of an individual to adapt its behavior to changes in the environment (Brown and Tait, 2015) by, e.g., stopping current behavior and initiating new behavior (Brown and Tait, 2015), solving a novel problem (problem-solving) or solving an existing problem in a novel way (innovation) is likely associated with invasion success (Reader and Laland, 2002; Sol et al., 2002; Shettleworth, 2010). This is because being more flexible can help an individual to better cope with novel habitats, predators, or resources (Sol et al., 2002; Wright et al., 2010; Chapple et al., 2012), which can ultimately increase their fitness (Sol et al., 2002). Measuring behavioral flexibility is challenging, but some metrics include the ability and speed with which animals conduct reversal learning (e.g., Brown and Tait, 2015; Lea et al., 2020) and their ability to deal with response inhibition (e.g., Diamond, 2013; Daniels et al., 2019), proficient problem solving across multiple domains (e.g., Logan, 2016a,b; Chow et al., 2018; Daniels et al., 2019; Lea et al., 2020), and the frequency of learning innovations (e.g., Sol and Lefebvre, 2000; Sol et al., 2002; Lea et al., 2020). A link between brain size (as a proxy of behavioral flexibility) and invasion success has been reported for invasive birds (Sol et al., 2002, 2005), mammals (Sol et al., 2008), amphibians and reptiles (Amiel et al., 2011). These reviews and comparative studies are powerful because they give us the big picture of how widely distributed a trait or relationship may be and sets the stage for later hypothesis testing. Another approach is to compare the cognitive ability of closely related species that may compete for the same resources, particularly when one species is native and the other is invasive (e.g., Roudez et al., 2008; Bezzina et al., 2014; Griffin and Diquelou, 2015; Chow et al., 2018). These studies typically address how species compete for ecological resources but differences in their ability to exploit resources may be independent of direct competition and may instead be a consequence of behavior and/or cognitive ability (e.g., Foucaud et al., 2016). Because cognitive abilities are often correlated with other factors such as motivation, personality, feeding ecology, sociality and life history (e.g., Tebbich et al., 2010; Carere and Locurto, 2011; Titulaer et al., 2012; van Horik and Madden, 2016; Dougherty and Guillette, 2018), disentangling the role of cognition in determining invasion success is challenging.
In this review, we have the following aims: (1) to review the evidence that cognition can influence invasion success; (2) characterize the features of cognitive ability that are likely to give individuals an advantage during the invasion process; and (3) propose future directions for studying the link between cognitive ability, fitness, and invasive success. To this end, we first conduct a systematic review of the literature on cognitive abilities that likely relate to invasion success (behavioral flexibility, problem solving, learning and memory) in both invertebrates and vertebrates. We also review the literature on brain size (as a proxy for cognitive ability) as it relates to invasive species. We then examine cognitive traits that could give species an advantage in a competitive, novel environment. Last, we describe an experimental approach to studying the role of cognition in determining invasive success and outline ways in which we can properly link invasive ability and cognition in a fitness framework.
Systematic Literature Compilation
We searched Web of Knowledge, Scopus and ProQuest for publications on biological invasions (using the keywords “invasi*,” “invad*,” “establish*,” and “introdu*”), cognition (using the keywords “cogni*,” “learning,” “flexi*,” and “innovat*”) and fitness (using the keywords “fitness,” “surviv*,” and “reprod*”) focusing on animals (using the keyword “animal*”). To refine our very broad search, we excluded publications based on the keywords “educat*,” “child*,” “physic*,” and “gluco*” (Figure 2). To get the largest possible number of hits with these search terms, we made sure that we included all words starting with these terms by applying *. Additionally, we used the built in options each database provided to further refine our searches (for more details see electronic Supplementary Material).
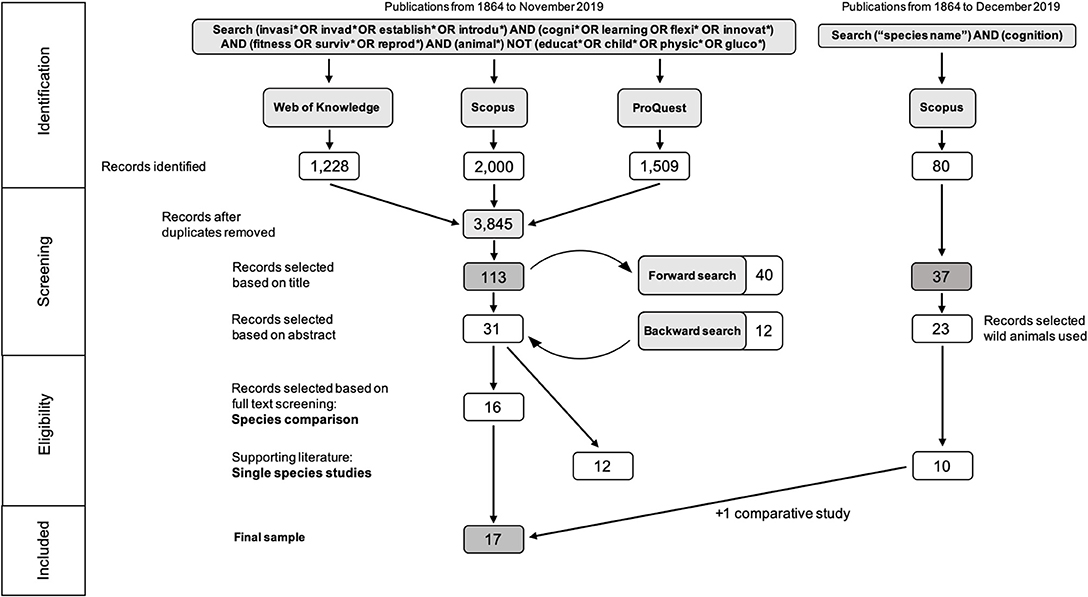
Figure 2. Graphical representation of the systematic literature compilation (PRISMA diagram) including details on search terms, databases searched, number of entries found, and number of entries selected at each step.
We downloaded 4,737 references as RIS files to import into EndNote (version X9.3.2) of which 892 were removed as duplicates. Only 2,000 of the 8,254 entries identified in Scopus could be downloaded (sorted by relevance). Of the remaining 3,845 we selected 61 based on the title. To be included in our sample, the title had to include any mention of cognitive ability such as learning, cognitive or behavioral flexibility, memory, inhibition, etc. To ensure no publications were missed, studies on behavior (which might encompass some cognitive ability) were included as well. Furthermore, the title had to mention either that the behavior or cognitive ability has a fitness or adaptive value (or increases reproductive success) and that it was tested in an invasive species or a species that successfully invaded an urban habitat. This ensured a broad selection of publications. We then conducted a forward search on these 61 entries resulting in an additional 40 entries (selection based on title). After full text download we looked at the abstracts of the 101 articles (61 + 40 = 101) and selected 27 as being relevant for our review. We included articles if the abstract (1) described a cognitive ability (studies solely looking at behavior were excluded at this stage); and (2) focused on invasiveness (an invasive species, comparing performance between invasive and native species or comparing performance between individuals from the source population and invasive population). We then conducted a backwards search (based on title) on the 27 selected articles resulting in an additional 11 publications. A second forward search on these 11 articles resulted in no new entries. Finally, we conducted a second backward search (resulting in one additional entry) and a final forward search (resulting in no new publications). We identified 113 articles based on title and selected 31 of those based on their abstract (Figure 2).
We then proceeded to read the full text of these 31 articles. Based on content, they were then divided into articles comparing species cognitive ability either directly (N = 10) or through a proxy such as brain size (N = 6) and articles looking at cognitive performance of a single invasive species (N = 12). The other three articles were reviews. Although studying the cognition of a single invasive species is interesting, without a comparison, it is unclear if the tested ability is associated with enhanced invasion success, consequently, some were used as examples to help identify possible useful cognitive abilities associated with invasion success (N = 10) but were not part of the main literature review.
Our review, therefore, focuses on those studies that make a comparison. To get a comprehensive picture of the cognitive abilities of the invasive species on which these comparative studies focus, we did an additional search for studies testing further cognitive abilities in these invasive species. We conducted these additional searches in Scopus using the species name (common or scientific) and “cognition” as the search terms (see electronic Supplementary Material for details). Studies were included if they were conducted on wild animals (either in the wild or testing wild caught individuals) and on a task that measured problem solving, innovation, behavioral flexibility, social learning, associative learning of ecologically relevant stimuli (food or threat) or spatial learning. We identified an additional 10 studies to those found in our initial search (N = 12). One of these studies was a species comparison which our initial search had missed and was included in our final sample. Overall, we identified 17 studies comparing performance between species and 21 studies testing cognitive performance on a single species (supporting literature). All searchers were conducted in November and December 2019 (Figure 2).
Invader Smarts vs. Native Know-How: a Comparison
To investigate if successful invaders possess enhanced cognition, a comparison of their cognitive ability to that of their congeneric non-invasive counterparts can give some indication of how cognition aids biological invasions. Although this seems a straightforward way to tackle the question if cognition benefits invasive species, only a small fraction of research has focused on a direct comparison between native and invasive congeners and even fewer have compared performance of the source population to the invasive population. Luckily, these few studies span a wide range of taxa from invertebrates such as crabs (Roudez et al., 2008; Ramey et al., 2009), crayfish (Hazlett et al., 2002) and flies (Foucaud et al., 2016), to vertebrates including mammals (MacDonald, 1997; Sol et al., 2008; Chow et al., 2018), birds (Sol et al., 2002, 2005; Diquelou et al., 2015; Griffin and Diquelou, 2015), fishes (Drake, 2007), reptiles (Amiel et al., 2011; Bezzina et al., 2014) and amphibians (Amiel et al., 2011) and provide a good first insight into how cognition may enhance invasive species success (Supplementary Table 1) in competing with the resident species assemblage already adapted to the invaded environment.
Comparison of Cognitive Performance Among Invasive vs. Native Species
Invertebrates
Invasive green crabs (or shore crabs, Carcinus maenas) learnt to locate a prey item (a ribbed mussel buried in the sand) faster over five test days compared to native blue crabs (Callinectes sapidus). Additionally, a larger proportion of green crabs were successful on the last three test days but both species showed similar memory of the trained location after a 10-day break (Roudez et al., 2008). Green crabs are a highly successful invader (Roudez et al., 2008). They have a high salinity and temperature tolerance, a high reproductive rate, and a long larval stage (Roman and Palumbi, 2004). They were able to transfer learnt prey handling techniques to novel prey items (Hughes and O'Brien, 2001) and were good spatial learners when tested in a complex maze (Davies et al., 2019). Invasive green crabs were also better at spontaneous alternation behavior compared to native blue crabs. Spontaneous alternation is the tendency of an individual to visit a location not previously visited or which has not been recently visited (Hughes, 2004) and can facilitate the discovery of new habitats, resources, or mates (Ramey et al., 2009). This was tested in a submerged plus-shaped maze. Only if an individual entered a novel arm four out of five times in a row was a successful spontaneous alteration recorded; otherwise it was unsuccessful. Invasive crabs made significantly more spontaneous alterations than native crabs, and only the green crabs' performed significantly above chance (Ramey et al., 2009). Spatial learning and memory, besides other traits (see above), therefore appears to play some role in invasive success in green crabs.
Apart from remembering the location of food in a new place, learning about novel predators and remembering cues associated with the threat also improves individual survival. Hazlett et al. (2002) investigated the memory for learnt predator odor in two pairs of invasive-native crayfish; the North American Orconectes virilis and its invasive competitor O. rusticus and the Italian Austropotamobius pallipes and its invasive competitor Procambarus clarkii. Individuals were either exposed to goldfish odor (an unfamiliar, herbivorous fish species unlikely to be recognized as a predator) combined with conspecific alarm chemicals (created by crushing a conspecific) for 2 h (short-term treatment) or 24 h (long-term treatment) or to a control treatment in which no goldfish odor or alarm chemicals were presented. The reaction of each crayfish to (1) water, (2) food odor (thawed cod), and (3) goldfish odor was then recorded for several weeks. Control animals did not react to goldfish odor indicating that goldfish are not innately recognized as predators, whereas trained crayfish showed a predator avoidance reaction when experiencing goldfish odor. Invasive O. rusticus ceased responding to goldfish odor after 4 weeks, whereas native O. virilis stopped responding after 1 week (2 h treatment) and 4 weeks (24 h treatment). Similarly, the native Italian A. pallipes stopped showing predator avoidance behavior 2 weeks after training no matter the treatment, whereas the invasive P. clarkii still recognized goldfish as a threat 3 weeks later when initially exposed for 24 h. Moreover, the two invasive species (O. rusticus and P. clarkii) responded as strongly to heterospecific as to conspecific alarm odors (Hazlett, 2000; Hazlett et al., 2003) while the native Italian A. pallipes did not (Hazlett et al., 2003). Further studies on O. rusticus showed that they have good spatial learning ability when escaping a T-maze, performing few wrong turns and with a low latency to exit (Tierney and Lee, 2011). Crayfish used both place learning by relying on maze cues and a response strategy when visual cues were provided within the maze. Furthermore, animals remembered the way out of the maze for 1 week (Tierney and Andrews, 2013). Finally, crayfish had a harder time learning to find the exit if conflicting cues were present and showed flexibility in learning during a spatial reversal (Tierney et al., 2019). These studies reveal that invasive crayfish species are more effective in using alarm cues provided by congeneric species and remember a predator odor for longer than native competitors. These results again suggest that better cognitive performance (here the memory of a predator odor) could provide an advantage in novel environments, consistent with conditions during invasion.
Comparing invasive species abilities to those of the native species they displace is one way to demonstrate which traits may be important for successful invasion. However, such studies are constrained by the fact that they focus on an invaders ability after it has already established itself and may have limited insight into abilities that might inherently make a species a good candidate to invade a new habitat. A comparison between female fruit flies (Drosophila subobscura) from their native range in Europe and from an area where they were introduced (in Chile) in the 1970s revealed that learning to select the correct oviposition site (medium without quinine, a bitter tasting compound) was not enhanced in the invasive population, but fecundity (number of eggs laid overall) was greater in females from Chile (Foucaud et al., 2016). Although invasive fruit flies were not better at selecting an appropriate oviposition site, tests looking at other cognitive abilities could give insight into if and how a range of cognitive abilities could be involved in predicting invasion success.
Vertebrates
Similar to invertebrates, a number of studies have used species comparisons to look for enhancements in learning and problem solving in invasive species. The Australian delicate skink (Lamprohpolis delicata) is a successful invader while their related congener, the common garden skink (Lampropholis guichenoti), is not. Both have frequently been discovered by biosecurity authorities within cargo but only the delicate skink has, so far, managed to establish viable populations outside their native range. Both species show very similar phenotype and behavior; however, delicate skinks are more exploratory and hide more (Chapple et al., 2011). Both species were tested on their ability to learn the location of food within a Y-maze. Lizards were provided with visual cues such as arm color and pattern as well as spatial intra-maze cues such as left/right position of the correct arm and distal extra-maze cues. Both species decreased the time taken to locate the hidden food but no difference was found between species. Additionally, they did not progressively take a more direct route within the maze indicating no learning had taken place (Bezzina et al., 2014). Based on these data, delicate skinks do not possess better cognitive ability than common garden skinks; however, learning ability was compared on a single test in this study. Additional work in delicate skinks showed that these lizards can learn to find a “safe” refuge to escape a simulated predator attack within a similar Y-maze using color cues (Kang et al., 2018), and that urbanized skinks are not better learners than individuals from natural areas (national park; Kang et al., 2018). Finally, behavioral phenotype affected learning performance: fast-type lizards (i.e., more active and exploratory) made more errors during learning (Chung et al., 2017; Goulet et al., 2018). Importantly, lizards were sourced within their native range to be tested in these different studies. Without data from the invasive population of the delicate skink it is still unclear if learning ability is a trait selected for during the establishment process.
Another Australian study compared the problem-solving ability between the native noisy miner (Manorina melanocephala) and the introduced common myna (Acridotheres tristis) in a foraging context. Problem solving may involve cognitive mechanisms such as associative learning, inhibitory control and cognitive flexibility (Diamond, 2013; van Horik and Madden, 2016); however, it can also involve a range of non-cognitive abilities such as motivation, persistence and motor skill (Diquelou et al., 2015; van Horik and Madden, 2016; Lermite et al., 2017). Although noisy miners are a very successful native honeyeater species, able to outcompete other small avian species when occurring in large numbers, common mynas are very opportunistic and able to occupy niches that native species cannot (Lowe et al., 2011; Sol et al., 2012a). Indeed, mynas are listed as one of the “100 World's Worst Invasive Alien species” (by the International Union for the Conservation of Nature; Boudjelas et al., 2000). To study problem-solving ability in these birds, three tasks were given in a random order in which birds had to use different techniques to open transparent food containers (task 1 and 2) or extract food from within a Styrofoam cup which was transparent on top (task 3). Overall, the invasive common myna outperformed the native noisy miner. Common mynas were faster to solve these tasks and showed greater motor diversity and flexibility, which were highly repeatable. Common mynas also showed repeatability in their persistence when solving the tasks compared to the noisy miners (Griffin and Diquelou, 2015). These result indicate that mynas show consistent, high levels of persistence, motor diversity, and flexibility, while miners do not. Furthermore, in a later study, task directed motivation (making more contact with an apparatus) and exploratory behavior (moving less within an open space) predicted task success in mynas (Lermite et al., 2017) confirming these traits to be important during problem solving. Moreover, invasive mynas can learn to remove lids from food wells (motor innovation) and motivation and neophobia were the best predictors of successful motor innovation (Sol et al., 2012b). Individuals from highly urbanized habitats were faster problem solvers and were also less neophobic and more exploratory (Sol et al., 2011), behavioral traits that covary with individual cognitive styles (Carere and Locurto, 2011; Sih and Del Giudice, 2012). Mynas were also good observational learners. They remembered a “dangerous” human after observing a staged catching event (Diquelou and Griffin, 2019) and became more wary (increase in locomotion) in the location of such an aversive event (Griffin and Boyce, 2009). Observing the behavior of an alarmed conspecific when the source of the threat was not visible, however, did not affect the observers behavior (Griffin et al., 2010).
Following Griffin and Diquelou (2015), transparent food containers (same as task 1) were presented to seven different Australian bird species: Australian ravens (Corvus coronoides), Australian magpies (Cracticus tibicen), magpie larks (Grallina cyanoleuca), common mynas (A. tristis), noisy miners (M. melanocephala), European starlings (Sturnus vulgaris), and crested pigeons (Ocyphaps lophotes) in the wild. As with common mynas, European starlings are highly invasive and should perform well in problem-solving tasks if problem solving indeed is a trait enhanced in invasive animals. Although this was true, Australian ravens were the best problem solvers closely followed by common mynas and European starlings. Except for the Australian magpie (another native species like the Australian raven), none of the other species showed much skill in solving the presented task. Motor flexibility (range of motor actions) predicted problem-solving probability across all species (Diquelou et al., 2015). These studies are great examples of how simple tests on wild birds can reveal striking difference in a range of abilities (cognitive and non-cognitive) and can give some indication of how learning, memory and flexibility are helping invaders colonize novel habitats.
Another species amongst the “100 World's Worst Invasive Alien species” (Boudjelas et al., 2000) is the gray squirrel (Sciurus carolinensis). In the UK, they outcompete and replace the native red squirrel (Sciurus vulgaris) with which they share a number of ecological traits (Koprowski, 1994; Gurnell et al., 2004; Lurz et al., 2005). Wild gray squirrels were better problem solvers compared to wild red squirrels. Animals were given an easy task in which they had to displace lids to gain access to a reward (nut), and a hard task in which they had to push or pull a bar to make a nut drop down to the bottom of a box, making it accessible. In the easy task, more gray squirrels were first-time solvers than red squirrels, and invasive gray squirrels were also faster solvers. In the hard task, more gray squirrels were solvers but both species showed the same performance, were similarly persistent (number of attempts), and used similar proportions of effective behaviors; red squirrels were even more efficient (sum of the duration of all attempts to solve the task until it was successfully solved; Chow et al., 2018). These results indicate that although both species exhibit similar levels of persistence and motor diversity (non-cognitive factors important for successful problem solving) other factors such as cognition might explain why more gray squirrels solved the tasks. Invasive wild gray squirrels also had longer lasting spatial memory of caches made by a human experimenter compared to native wild red squirrels (MacDonald, 1997). However, low sample sizes reduced statistical power and results need to be interpreted with caution. Furthermore, the difference could have been caused by the fact that gray squirrels rely more on caches (MacDonald, 1997) and could have a more specialized hippocampus, an essential brain area for spatial memory. The results of these studies indicate that learning and memory could help explain why gray squirrels have been such successful invaders although both species have good problem-solving abilities. Future studies could investigate if gray squirrels are innately better learners and problem solvers by comparing individuals from the native and introduced range. Moreover, gray squirrels have been introduced to Italy more recently (20th century; Bertolino et al., 2008) and a comparison to the UK population (introduced in the 19th century; Chow et al., 2018) could provide new insights into how selection shapes cognitive (and non-cognitive) traits during the invasion process.
In summary, comparative research in both invertebrates and vertebrates suggests that cognitive abilities such as learning and memory might help favor invasion success. However, not all studies have found differences between the tested species (Bezzina et al., 2014) or populations (Foucaud et al., 2016). Furthermore, non-cognitive factors are clearly causing differences between species in task performance (Diquelou et al., 2015; Griffin and Diquelou, 2015). It is therefore crucial to take such factors into account to avoid drawing the wrong conclusions about the role of cognition in invasion success. Studies in birds and mammals mainly test problem-solving ability focusing on non-cognitive factors such as motivation and persistence but less on cognitive factors such as inhibition and flexibility. Studies focusing on species or population comparisons are rare, which underscores how we are only just starting to understand how cognition might aid the invasion process. Moreover, if species innately possess beneficial abilities that help them invade novel habitats or if these abilities emerge as a result of selection is unclear. The degree to which cognitive abilities are beneficial in the invasion process and if these are general across different invasive species is also unknown.
Comparison Based on Relative Brain Size in Vertebrates
A link between cognitive ability, innovative behavior (feeding and problem solving) and brain size has emerged in a range of taxa (e.g., Lefebvre et al., 1997, 2004; Bouchard et al., 2007; Güntürkün, 2012; Griffin et al., 2013). Recently, comparative studies have examined the relationship between behavior, cognition, and brain size with a focus on how innovative behavior and cognition might benefit invasive species (Sol and Lefebvre, 2000; Sol et al., 2002, 2005, 2008; Drake, 2007; Amiel et al., 2011). By using data on relative brain size and invasion success after human translocation, Amiel et al. (2011) suggested that those species of amphibians and reptiles with larger relative brains are more successful in establishing a population in a novel environment. This trend was consistent geographically except for Australasia. Here, the opposite trend was observed, species with smaller brains were more successful (Amiel et al., 2011). In birds, there is a strong link between relative size of the hyperstriatum ventral (which serves a similar function to the mammalian neocortex) and cognitive flexibility (Timmermans et al., 2000) but it is still unclear if larger brain size confers similar cognitive enhancements in amphibians and reptiles. A large-scale analysis of relative brain size, fecundity, parental investment and invasion success in fishes revealed no correlation between brain size and establishment success in a novel environment (Drake, 2007). Empirical work on fishes has demonstrated a positive link between brain size and cognition (numerical associative learning; Kotrschal et al., 2013a,b). In these studies, however, fishes were artificially bred for larger brain size; if selection acts in a similar manner in natural populations is unclear, and if so, a correlation between brain size and cognitive ability might be much weaker and harder to detect. In birds, the first study looking at how establishment/invasion success relates to brain size and foraging behavior was restricted to species introduced to New Zealand. It revealed, that species with larger relative brain size were more likely to successfully invade New Zealand and that successful invaders also showed a higher number of foraging innovations. Other traits were also associated with invasiveness: the number of individuals introduced (introduction effort), migration (migratory, partially migratory, or sedentary) and developmental mode (nidicolous vs. nidifugous) (Sol and Lefebvre, 2000). Following this study, the inclusion of a much wider range of bird species revealed a similar trend. Again, species that were successful invaders had larger relative brain size and showed more foraging innovations. As before, additional variables were associated with invasion success: order a species belongs to, plumage (monochromatic or dichromatic), nest location (ground, bush/tree or hole) and if a species was also found in urbanized habitats (Sol et al., 2002). Anthropogenic environments differ greatly from natural environments and are subject to greater change and disturbance (Lowry et al., 2013). It is not surprising to find species that establish themselves in a human dominated environment to also have larger relative brains possibly to deal with these challenging environmental conditions. Sol et al. (2005) confirmed the relationship between brain size and successful adaptation to environmental change (brain size—environmental change hypothesis) first in birds. They found that birds with larger relative brains were more successful at establishing a population in novel environments and this invasion propensity was also associated with innovativeness; invasive species had more feeding innovations. Additional analyses showed the success of invasive species in coping with novel environments was due to an increase in their cognitive ability and no other mechanisms (Sol et al., 2005). The brain size-environmental change hypothesis was also confirmed in mammals. Establishment success was higher in mammals with relatively larger brains and was also associated with introduction effort and habitat generalism (Sol et al., 2008). Contrary to birds, however, no proxy for cognitive ability (e.g., feeding innovations) was included in the analysis. Nevertheless, a link between relative brain size and cognitive ability has already been established in some mammalian groups (e.g., Lefebvre et al., 2004; Güntürkün, 2012).
Across taxa, the analysis of large multi-species datasets has been useful for investigating the generality of different hypotheses about how cognition could help species become successful invaders. While a link between larger relative brain size and enhanced cognition has been established for some taxa, more effort is still needed to test for the existence of a similar relationship in other taxa. Importantly, studies using brain size as a proxy for cognitive ability run into a number of constraints and results should be interpreted with caution. Evidence suggests that brains are heterogeneous across taxa and the degree of enlargement and neuron densities of specific brain areas can differ greatly (e.g., Olkowicz et al., 2016 cited by Logan et al., 2018). Furthermore, selection may change specific brain areas without an increase in brain size (see Does Selection Act on Brain Size? in Logan et al., 2018). Moreover, analyses are hampered because the relationship between brain size and body size is not linear in all taxa [e.g., Fitzpatrick et al. (2012) cited by Montgomery et al. (2013), Logan et al. (2018) cited by Logan et al. (2018)] and often, brain measures used in comparative studies are based on a few individuals (Logan et al., 2018). Results of such comparative studies using brain size are, therefore, better used as a starting point for more detailed investigations looking at how individual differences in cognitive ability translate to brain anatomy within species (e.g., Audet et al., 2018) followed by investigations into differences between closely related species (e.g., Aamodt et al., 2020) to better understand how changes in brain size and structure are related to cognition and behavior (Logan et al., 2018).
Cognitive Abilities That Can Increase Invasion Success
During invasion into a new habitat, animals will inevitably face environmental conditions which are novel and to which they are not optimally adapted. The faster way to adapt to changing conditions is through behavioral plasticity and flexible behavior. Shifts in behavior most likely follow after experience with a certain situation allowing the individual to learn an appropriate response. Animals might innovate solutions to problems by modifying existing behavior, come up with new behaviors, or learn socially from conspecifics or even native congeneric heterospecifics (Wright et al., 2010; Damas-Moreira et al., 2018). Cognitive ability can play an important role during the different stages of the invasion process, but which abilities increase invasion success within or across stages? Below, we provide some examples of cognitive abilities which are likely to be involved in increasing the likelihood that a species becomes established in a novel environment (based on work presented in the previous section).
Behavioral Flexibility, Innovation, and Problem Solving
Although we can assume that a cognitive ability might not carry the same advantage when facing novel or changing conditions across species, behavioral flexibility, the ability to adapt flexibly to changes in the environment (Brown and Tait, 2015), is a trait likely to be advantageous to most, if not all, invaders. Behavioral flexibility is often described together with innovation and problem solving such that species or individuals that are innovators or problem solvers are behaviourally flexible (Lea et al., 2020). A key component of behavioral flexibility is that it can be adaptive in a given situation (Brown and Tait, 2015). Inventing new behaviors or techniques to solve problems such as being confronted with novel foods or having to extract edible foods from within inedible casings (e.g., shells, soil, wood) will improve survival. Furthermore, being behaviourally flexible in the traditional sense, by flexibly adjusting behavior in situations when old behavior becomes unsuccessful, also carries fitness advantages. Therefore, our definition of behavioral flexibility encompasses any change in behavior that helps individuals to adapt to changing conditions and can be measured using innovation rates, different techniques to solve puzzle box tasks (i.e., problem-solving techniques), reversal learning and similar tasks (e.g., serial reversal learning or set-shifting) that cover multiple cognitive domains. Wright et al. (2010) proposed the “Adaptive Flexibility Hypothesis” in which they relate behavioral flexibility and its usefulness (adaptive value) to the different stages of the invasion process and propose that behavioral flexibility will be most beneficial during the initial stages of an invasion in which novel problems are most likely to be encountered. For example, wild North American racoons (Procyon lotor) caught in their native range, possessed flexible problem-solving skills. They were tested on a multi-access box which could be opened in three different ways. Across nights, access points were blocked to force animals to abandon old solutions and find new ways to access the reward. Seven of the 20 racoons tested solved the box using all three ways showing high levels of problem-solving ability and flexibility in their use of behaviors to open the box when an old solution became ineffective. Exploratory behavior and persistence were robust predictors of solving success. Inhibitory control was also assessed by looking at the time spent interacting with a newly locked access point but it did not predict differences in solving success between night two and three. This shows that this species possess an innate ability to solve problems which might be the reason why it is such a successful invader (Daniels et al., 2019). Not all successful invaders, however, are great problem solvers. Great-tailed grackles (Quiscalus mexicanus) showed behavioral flexibility during reversal learning and varying degrees of problem-solving ability. Birds learnt to reverse their preference for finding food in a golden tube to choosing the previously unrewarded silver tube during a reversal stage. Fast learners were, however, not more flexible (Logan, 2016a). Problem solving was tested using the Aesop's fable paradigm (dropping stones into water to raise the water level and bring a reward closer; Logan, 2016a), a stick tool-use task (to extract a piece of bred from between two transparent walls) and a horizontal and vertical string pulling task (in which a string has to be pulled to bring a reward close enough to reach it; Logan, 2016b). Although grackles were unable to spontaneously drop stones, use sticks or pull strings to access a reward, birds could learn to drop stones to reach a reward (Logan, 2016a,b). However, they could not learn to use stick tools (Logan, 2016b). They were also flexible in learning which strategy to use during the Aesop's fable paradigm. Initially birds preferred heavier stones but when these became ineffective, they switched strategy to use lighter stones in higher numbers Logan, 2016a. Individuals tested in these tasks were wild caught from within their introduced range. If great-tailed grackles are innately good problem solvers has still to be investigated. So far, no direct link between cognition, its fitness value, and invasion success has been demonstrated. Nonetheless, increasing numbers of studies show how behavioral flexibility is enhanced in successful invaders as compared to species that did not establish a viable population in a novel environment (e.g., Sol and Lefebvre, 2000; Sol et al., 2002, 2005, 2008) and how problem-solving ability, innovation, and behavioral flexibility influence reproductive success (e.g., Cole et al., 2012; Cauchard et al., 2013, 2017; Ashton et al., 2018) and survival (e.g., Dayananda and Webb, 2017; Madden et al., 2018). Conflating these areas of research will further our understanding of if and how behavioral flexibility aids biological invaders.
The Fast Acquisition of Information Through Learning
Learning enables an individual to acquire new information and compensate for shortcomings that might arise from entering a new environment to which it is not adapted, or even maladapted. Theoretical models predict that, if the initial founder population is small, adaptation (through genetic change) alone is not fast enough to result in a sustainable population; however, learning can help a population persist even if it is initially maladapted to the novel environment (Sutter and Kawecki, 2009). Importantly, the benefit of learning to buffer differences in fitness between habitats and to increase mean fitness in the novel habitat is higher when the difference between the native and novel habitat is large (Sutter and Kawecki, 2009). This means that learning could be more beneficial for introduced species than it is during a gradual range expansion. Learning about the characteristics of a novel habitat has the potential to increase the probability that even a small founder population can persist in a new environment. Fast learning is present in invasive species, as compared to native competitors (see earlier discussion of green crabs, common mynahs and gray squirrels). In addition, invasive American bullfrogs (Lithobates catesbeianus), which experienced threat (scent of the novel largemouth bass, Micropterus salmoides, combined with conspecific alarm cues) as embryos within the egg, hid more inside a refuge when experiencing the threat later in life (as larvae after hatching). No effect could be detected in untrained control larvae (Garcia et al., 2017). These studies give some indication that enhanced learning ability may facilitate invasion success; however, we do not know if these learning abilities have actual fitness consequences (increased survival and reproductive success) or if these species are innately better learners.
Social Learning and the Spread of Knowledge
When entering a novel habitat that is dissimilar to the environment in which an individual developed and has adapted to, collecting information about predators, what to eat and where to hide, can be crucial for survival. Trial-and-error learning can be associated with high costs when sampling noxious foods or encountering dangerous predators, possibly leading to death after first contact. It can therefore be beneficial to rely on the knowledge that other individuals have already accumulated. Social learning is considered a shortcut to accessing important information that has already been vetted and is beneficial (Galef and Laland, 2005). During the early stages of the invasion process, the only individuals with relevant information might be congeneric native individuals. Some invasive species are known to use the behavior and alarm signals of native species to quickly learn about food (e.g., Camacho-Cervantes et al., 2015) and novel threats (e.g., Hazlett, 2000; Hazlett et al., 2002, 2003). For example, lab-reared female guppies that watched the behavior of either a small group of conspecifics or heterospecifics (butterfly splitfins, Ameca splendens), when food scent was introduced, changed their behavior accordingly. Observer fish showed similar behavior to demonstrators by spending more time in the upper layer of their aquarium. When no demonstrators were present or water was added instead of the food odor, observers did not change their behavior showing that their behavior was guided by that of the demonstrators irrespective of whether demonstrators were conspecifics or heterospecifics (Camacho-Cervantes et al., 2015). Furthermore, a recent study showed that invasive Italian wall lizards, Podarcis sicula, readily learn stimuli associated with a food reward (in an artificial foraging task using differently colored food wells) from both conspecifics and heterospecifics and made fewer errors when learning socially as opposed to individually (Damas-Moreira et al., 2018). Social learning has, however, limitations. It is only really beneficial if the environment is largely stable and information stays relevant for extended periods of time and if animals are selective about what information they learn (Galef and Laland, 2005). Nonetheless, it might be a powerful tool to spread important information rapidly through an invasive founder population and increase the likelihood that a large enough number of individuals survive and can reproduce.
Choosing “Smart” Mates
In some species, smart individuals are preferred as mates (e.g., Keagy et al., 2009, 2011; Chen et al., 2019) although cognitive abilities do not always provide direct benefits to the choosing sex (e.g., Keagy et al., 2011). If a cognitive ability or innovative behavior enhances survival within a novel environment and if these abilities are heritable then choosing a “smart” mate would increase offspring survival and fitness, and in the long run, help establish a population in the new location. There is evidence that cognitive ability (e.g., Galsworthy et al., 2005; Hopkins et al., 2014) and learnt behavior (e.g., Kelly and Phillips, 2017, 2018) are heritable, but this is not always the case (e.g., Quinn et al., 2016; for a review see Coston et al., 2015). Mate choice for individuals with enhanced cognitive ability has received little attention (for a review see Boogert et al., 2011) and none in relation to invasion success. It has also been shown that a number of species have to learn about intraspecific sexual traits to successfully mate and reproduce (Ryan et al., 2009). During the invasion process, animals might come in contact with congeners that are not dissimilar to themselves. On the one hand, if courting and mating with congeneric species are costly, enhanced learning for intraspecific traits that help distinguish between species could help avoid such costs and help an establishing population to grow to a sustainable size. On the other hand, hybridization can enhance cognition and adaptability to novel environments through backcrossing of favorable genes acquired from the native species into the parent lineage or through the development of extreme phenotypes that increase fitness in a novel habitat (Rice, 2020).
Linking Cognition, Fitness and Invasion Success—Future Directions
The way forward is multi-faceted, but a good starting point is studies that compare invasive and non-invasive species, source and invasive populations, and experimental studies which link or test for links between fitness, cognitive ability, and invasive ability. Ten years ago, no longitudinal studies of invasive populations were available (Wright et al., 2010), a big gap that has not been filled. Additionally, we found only one study comparing learning (female fruit flies choosing between oviposition sites) between individuals from the source and invasive population (Foucaud et al., 2016), an approach also listed by Wright et al. (2010). We need more baseline data on differences between invasive and non-invasive species pairs (i.e., congeneric species) and between invasive and source populations. The challenge will be to conduct direct tests of how cognitive ability affects fitness with links to either survival or reproductive success in the future as manipulation of cognitive ability will likely also change associated abilities (cognitive and non-cognitive; e.g., Cauchard et al., 2017). This will create a clearer picture of what attributes make a species a more successful invader than less invasive species.
Simple cognitive tests that can be used in the wild and on a wide range of species would be especially valuable, although bringing wild caught individuals to the lab and testing an array of cognitive abilities also has merit. Careful consideration should also be given to which cognitive abilities to investigate, based on a species' ecology and sociobiology. We need to think in terms of aspects of cognition that are most likely to be beneficial during an invasion event. At the same time, cognition often correlates with non-cognitive traits including motivation or neophobia (e.g., Sol et al., 2011, 2012b) which should be measured simultaneously. Moreover, correlation between cognitive traits might occur warranting additional controls for such co-variation. We also would like to highlight that, so far, fitness benefits of enhanced cognitive ability have often been implied, but less often directly studied (e.g., Thornton et al., 2014; Dayananda and Webb, 2017; Ashton et al., 2018). Studying the value of cognitive ability during biological invasions is not an easy task but certainly one that has a substantial pay-off.
Future research could look to studies of Anolis lizards (anoles) for insight on how to experimentally study the role of cognition in invasions. Anoles have become the poster child of experimental vertebrate evolution because large numbers of known individuals can be introduced on to small islands and they have sufficiently short generation time to study selection in “ecological” time. Their survival and fitness can then be measured for a range of traits and island conditions (e.g., predator vs. predator-free, different habitats, etc.), thereby creating a picture of how selection acts on traits in novel environments. As it turns out, the brown anole (Anolis sagrei) is now a common invasive species documented in far flung locations from Florida (USA) to Taiwan, including regions such as Hawai'i. Florida is also home to a large number of invasive species including many species of lizards because of its tropical climate and an historically lax control of animal imports. We highlight this because Florida is a potential epicenter for a range of studies on how cognitive ability may link to invasive ability because the spread of these species can be easily monitored. This state also has a large number of artificial islands in their inter-coastal waterways which are used for selection studies (e.g., Fargevieille et al., 2019). We suggest using these same islands for experimental studies involving the release of anoles of known cognitive ability to monitor their survival and replicate it among islands. Likewise, both invasive brown anoles and native green anoles of known cognitive ability could be released on to the same islands. These islands are small and can be easily manipulated. For example, in addition to introducing pairs of native and invasive anoles, the native green anole could first be introduced and established before subsequently releasing brown anoles. This would replicate real-world conditions where invasive and native species come into contact. Likewise, common garden experiments in which native and invasive anoles are first raised under the same conditions before being released onto islands would control for any variance contributed by the developmental environment. We also suggest assaying all individuals for behavior, such as boldness, exploratory behavior, and propensity to disperse (this can be done in large outdoor arenas). Furthermore, traits such as motivation and persistence should be measured during cognitive testing to ensure that these traits are not causing differences between species rather than cognition. It is important to be able to disentangle the effects of cognitive ability and behavior on invasive ability. And with respect to cognitive ability, although time consuming, we suggest multiple tests that focus on spatial learning and give a measure of behavioral flexibility because these traits are likely to be crucial to survival in novel environments.
In summary, there is increasing evidence that cognition influences fitness (e.g., Cole et al., 2012; Cauchard et al., 2013, 2017; Dayananda and Webb, 2017; Ashton et al., 2018; Madden et al., 2018). In the case of biological invasions, when organisms are exposed to new environments, cognitive ability is predicted to improve the probability of successful establishment. In this scenario, individuals with better cognitive ability are therefore more likely to be selected for. This hypothesis requires testing and we have suggested a potential experimental paradigm using Anolis lizards, but there are many others. We also encourage more empirical work comparing pairs of invasive and non-invasive species that are closely related and more studies comparing populations at the invasion source and the invasion front in order to better understand invasions and the role of selection. Understanding invasions in light of a species' cognitive ability and the relative fitness of potential behavioral and cognitive styles is a field with great potential. We hope this review will help stimulate innovative research in this direction.
Author Contributions
BS did the integrative review and wrote the bulk of the paper, ID-M wrote the bulk of the introduction and MW wrote the bulk of the future directions section. All authors contributed to writing and revising all sections of the manuscript. All authors conceived the framework for the paper.
Funding
Our work on cognition has been supported by Macquarie University and the Australian Research Council.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the editors of this special issue for inviting us to contribute.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.00187/full#supplementary-material
References
Aamodt, C. M., Farias-Virgens, M., and White, S. A. (2020). Birdsong as a window into language origins and evolutionary neuroscience. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375:20190060. doi: 10.1098/rstb.2019.0060
Amiel, J. J., Tingley, R., and Shine, R. (2011). Smart moves: effects of relative brain size on establishment success of invasive amphibians and reptiles. PLoS ONE 6:e18277. doi: 10.1371/journal.pone.0018277
Ashton, B. J., Ridley, A. R., Edwards, E. K., and Thornton, A. (2018). Cognitive performance is linked to group size and affects fitness in Australian magpies. Nature 554, 364–370. doi: 10.1038/nature25503
Audet, J. N., Kayello, L., Ducatez, S., Perillo, S., Cauchard, L., Howard, J. T., et al. (2018). Divergence in problem-solving skills is associated with differential expression of glutamate receptors in wild finches. Sci. Adv. 4:eaao6369. doi: 10.1126/sciadv.aao6369
Bertolino, S., Lurz, P. W. W., Sanderson, R., and Rushton, S. P. (2008). Predicting the spread of the American grey squirrel (Sciurus carolinensis) in Europe: a call for a coordinated European approach. Biol. Conserv. 141, 2564–2575. doi: 10.1016/j.biocon.2008.07.017
Bezzina, C. N., Amiel, J. J., and Shine, R. (2014). Does invasion success reflect superior cognitive ability? A case study of two congeneric lizard species (Lampropholis, Scincidae). PLoS ONE 9:e86271. doi: 10.1371/journal.pone.0086271
Blackburn, T. M., Pyšek, P., Bacher, S., Carlton, J. T., Duncan, R. P., Jarošík, V., et al. (2011). A proposed unified framework for biological invasions. Trends Ecol. Evol. 26, 333–339. doi: 10.1016/j.tree.2011.03.023
Boogert, N. J., Fawcett, T. W., and Lefebvre, L. (2011). Mate choice for cognitive traits: a review of the evidence in nonhuman vertebrates. Behav. Ecol. 22, 447–459. doi: 10.1093/beheco/arq173
Bouchard, J., Goodyer, W., and Lefebvre, L. (2007). Social learning and innovation are positively correlated in pigeons (Columba livia). Anim. Cognit. 10, 259–266. doi: 10.1007/s10071-006-0064-1
Boudjelas, S., Browne, M., De Pooter, M., and Lowe, S. (2000). 100 of the World's Worst Invasive Alien Species: A Selection From the Global Invasive Species Database. Invasive Species Specialist Group. Available online at: https://www.iucn.org/content/100-worlds-worst-invasive-alien-species-a-selection-global-invasive-species-database
Brown, V. J., and Tait, D. S. (2015). “Behavioral flexibility: attentional shifting, rule switching and response reversal” in Encyclopedia of Psychopharmacology, eds I. P. Stolerman, and L. H. Price (Germany, Berlin: Springer-Verlag), 264–269.
Budaev, S., Jørgensen, C., Mangel, M., Eliassen, S., and Giske, J. (2019). Decision-making from the animal perspective: bridging ecology and subjective cognition. Front. Ecol. Evol. 7:164. doi: 10.3389/fevo.2019.00164
Camacho-Cervantes, M., Ojanguren, A. F., and Magurran, A. E. (2015). Exploratory behaviour and transmission of information between the invasive guppy and native Mexican topminnows. Anim. Behav. 106, 115–120. doi: 10.1016/j.anbehav.2015.05.012
Carere, C., and Locurto, C. (2011). Interaction between animal personality and animal cognition. Cur. Zool. 57, 491–498. doi: 10.1093/czoolo/57.4.491
Cauchard, L., Angers, B., Boogert, N. J., Lenarth, M., Bize, P., and Doligez, B. (2017). An experimental test of a causal link between problem-solving performance and reproductive success in wild great tits. Front. Ecol. Evol. 5:107. doi: 10.3389/fevo.2017.00107
Cauchard, L., Boogert, N. J., Lefebvre, L., Dubois, F., and Doligez, B. (2013). Problem solving performance is correlated with reproductive success in a wild bird population. Anim. Behav. 85, 19–26. doi: 10.1016/j.anbehav.2012.10.005
Chapple, D. G., Simmonds, S. M., and Wong, B. (2011). Know when to run, know when to hide: can behavioural differences explain the divergent invasion success of two sympatric lizards? Ecol. Evol. 1, 278–289. doi: 10.1002/ece3.22
Chapple, D. G., Simmonds, S. M., and Wong, B. B. (2012). Can behavioral and personality traits influence the success of unintentional species introductions? Trends Ecol. Evol. 27, 57–64. doi: 10.1016/j.tree.2011.09.010
Chen, J., Zou, Y., Sun, Y. H., and Ten Cate, C. (2019). Problem-solving males become more attractive to female budgerigars. Science 363, 166–167. doi: 10.1126/science.aau8181
Chow, P. K. Y., Lurz, P. W. W., and Lea, S. E. G. (2018). A battle of wits? problem-solving abilities in invasive eastern grey squirrels and native Eurasian red squirrels. Anim. Behav. 137, 11–20. doi: 10.1016/j.anbehav.2017.12.022
Chung, M., Goulet, C. T., Michelangeli, M., Melki-Wegner, B., Wong, B. B. M., and Chapple, D. G. (2017). Does personality influence learning? A case study in an invasive lizard. Oecologia 185, 641–651. doi: 10.1007/s00442-017-3975-4
Colautti, R. I., and MacIsaac, H. J. (2004). A neutral terminology to define “invasive” species. Divers. Distrib. 10, 135–141. doi: 10.1111/j.1366-9516.2004.00061.x
Cole, E. F., Morand-Ferron, J., Hinks, A. E., and Quinn, J. L. (2012). Cognitive ability influences reproductive life history variation in the wild. Curr. Biol. 22, 1808–1812. doi: 10.1016/j.cub.2012.07.051
Coston, R., Branch, C. L., Kozlovsky, D. Y., Dukas, R., and Pravosudov, V. V. (2015). Heritability and the evolution of cognitive traits. Behav. Ecol. 26, 1447–1459. doi: 10.1093/beheco/arv088
Damas-Moreira, I., Oliveira, D., Santos, J. L., Riley, J. L., Harris, D. J., and Whiting, M. J. (2018). Learning from others: an invasive lizard uses social information from both conspecifics and heterospecifics. Biol. Lett. 14:20180532. doi: 10.1098/rsbl.2018.0532
Daniels, S. E., Fanelli, R. E., Gilbert, A., and Benson-Amram, S. (2019). Behavioral flexibility of a generalist carnivore. Anim. Cognit. 22, 387–396. doi: 10.1007/s10071-019-01252-7
Davies, R., Gagen, M. H., Bull, J. C., and Pope, E. C. (2019). Maze learning and memory in a decapod crustacean. Biol. Lett. 15:20190407. doi: 10.1098/rsbl.2019.0407
Dayananda, B., and Webb, J. K. (2017). Incubation under climate warming affects learning ability and survival in hatchling lizards. Biol. Lett. 13:20170002. doi: 10.1098/rsbl.2017.0002
Diamond, A. (2013). Executive functions. Annu. Rev. Psychol. 64, 135–168. doi: 10.1146/annurev-psych-113011-143750
Diquelou, M. C., and Griffin, A. S. (2019). It's a trap! invasive common mynas learn socially about control-related cues. Behav. Ecol. 30, 1314–1323. doi: 10.1093/beheco/arz079
Diquelou, M. C., Griffin, A. S., and Sol, D. (2015). The role of motor diversity in foraging innovations: a cross-species comparison in urban birds. Behav. Ecol. 27, 584–591. doi: 10.1093/beheco/arv190
Dougherty, L. R., and Guillette, L. M. (2018). Linking personality and cognition: a meta-analysis. Phil. Trans. R. Soc. B 373:20170282. doi: 10.1098/rstb.2017.0282
Drake, J. M. (2007). Parental investment and fecundity, but not brain size, are associated with establishment success in introduced fishes. Func. Ecol. 21, 963–968. doi: 10.1111/j.1365-2435.2007.01318.x
Essl, F., Dullinger, S., Rabitsch, W., Hulme, P. E., Hülber, K., Jarošik, V., et al. (2011). Socioeconomic legacy yields an invasion debt. Proc. Nat. Acad. Sci. U.S.A. 108, 203–207. doi: 10.1073/pnas.1011728108
Fargevieille, A., Reedy, A. M., Mitchell, T. S., Durso, A. M., Delaney, D. M., Pearson, P. R., et al. (2019). Population demographics of an invasive lizard following experimental introduction on small islands. Integ. Comp. Biol. 59:E66. doi: 10.1093/icb/icz003
Fitzpatrick, J. L., Almbro, M., Gonzalez-Voyer, A., Hamada, S., Pennington, C., Scanlan, J., et al. (2012). Sexual selection uncouples the evolution of brain and body size in pinnipeds. J. Evol. Biol. 25, 1321–1330. doi: 10.1111/j.1420-9101.2012.02520.x
Foucaud, J., Moreno, C., Pascual, M., Rezende, E. L., Castañeda, L. E., Gibert, P., et al. (2016). Introduced Drosophila subobscura populations perform better than native populations during an oviposition choice task due to increased fecundity but similar learning ability. Ecol. Evol. 6, 1725–1736. doi: 10.1002/ece3.2015
Galef, B. G. J., and Laland, K. N. (2005). Social learning in animals: empirical studies and theoretical models. Bioscience 55, 489–499. doi: 10.1641/0006-3568(2005)055[0489:SLIAES]2.0.CO;2
Galsworthy, M. J., Paya-Cano, J. L., Liu, L., Monleon, S., Gregoryan, G., Fernandes, C., et al. (2005). Assessing reliability, heritability and general cognitive ability in a battery of cognitive tasks for laboratory mice. Behav. Genet. 35, 675–692. doi: 10.1007/s10519-005-3423-9
Garcia, T. S., Urbina, J. C., Bredeweg, E. M., and Ferrari, M. C. O. (2017). Embryonic learning and developmental carry-over effects in an invasive anuran. Oecologia 184, 623–631. doi: 10.1007/s00442-017-3905-5
Goulet, C. T., Michelangeli, M., Chung, M., Riley, J. L., Wong, B. B. M., Thompson, M. B., et al. (2018). Evaluating cognition and thermal physiology as components of the pace-of-life syndrome. Evol. Ecol. 32, 469–488. doi: 10.1007/s10682-018-9948-1
Griffin, A. S., and Boyce, H. M. (2009). Indian mynahs, Acridotheres tristis, learn about dangerous places by observing the fate of others. Anim. Behav. 78, 79–84. doi: 10.1016/j.anbehav.2009.03.012
Griffin, A. S., Boyce, H. M., and MacFarlane, G. R. (2010). Social learning about places: observers may need to detect both social alarm and its cause to learn. Anim. Behav. 79, 459–465. doi: 10.1016/j.anbehav.2009.11.029
Griffin, A. S., and Diquelou, M. C. (2015). Innovative problem solving in birds: A cross-species comparison of two highly successful passerines. Anim. Behav. 100, 84–94. doi: 10.1016/j.anbehav.2014.11.012
Griffin, A. S., Guez, D., Federspiel, I., Diquelou, M., and Lermite, F. (2016). “Invading new environments: a mechanistic framework linking motor diversity and cognition to establishment success” in Biological Invasions and Animal Behaviour, ed. J. S. Weis and D. Sol (Cambridge: Cambridge University Press), 26–46.
Griffin, A. S., Guez, D., Lermite, F., and Patience, M. (2013). Tracking changing environments: innovators are fast, but not flexible learners. PLoS One 8:e84907. doi: 10.1371/journal.pone.0084907
Güntürkün, O. (2012). The convergent evolution of neural substrates for cognition. Psychol. Res. 76:212219. doi: 10.1007/s00426-011-0377-9
Gurnell, J., Wauters, L. A., Lurz, P. W. W., and Tosi, G. (2004). Alien species and interspecific competition: effects of introduced eastern grey squirrels on red squirrel population dynamics. J. Anim. Ecol. 73, 26–35. doi: 10.1111/j.1365-2656.2004.00791.x
Hazlett, B. A. (2000). Information use by an invading species: do invaders respond more to alarm odors than native species? Biol. Invas. 2, 289–294. doi: 10.1023/A:1011450702514
Hazlett, B. A., Acquistapace, P., and Gherardi, F. (2002). Differences in memory capabilities in invasive and native crayfish. J. Crustacean Biol. 22, 439–448. doi: 10.1163/20021975-99990251
Hazlett, B. A., Burba, A., Gherardi, F., and Acquistapace, P. (2003). Invasive species of crayfish use a broader range of predation-risk cues than native species. Biol. Invas. 5, 223–228. doi: 10.1023/A:1026114623612
Hopkins, W. D., Russell, J. L., and Schaeffer, J. (2014). Chimpanzee intelligence is heritable. Curr. Biol. 24, 1649–1652. doi: 10.1016/j.cub.2014.05.076
Hughes, R. N. (2004). The value of spontaneous alternation behaviour (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 28, 497–505. doi: 10.1016/j.neubiorev.2004.06.006
Hughes, R. N., and O'Brien, N. (2001). Shore crabs are able to transfer learned handling skills to novel prey. Anim. Behav. 61, 711–714. doi: 10.1006/anbe.2000.1640
Kang, F., Goulet, C. T., and Chapple, D. G. (2018). The impact of urbanization on learning ability in an invasive lizard. Biol. J. Linn. Soc. 123, 55–62. doi: 10.1093/biolinnean/blx131
Keagy, J., Savard, J. F., and Borgia, G. (2009). Male satin bowerbird problem-solving ability predicts mating success. Anim. Behav. 78, 809–817. doi: 10.1016/j.anbehav.2009.07.011
Keagy, J., Savard, J. F., and Borgia, G. (2011). Complex relationship between multiple measures of cognitive ability and male mating success in satin bowerbirds, Ptilonorhynchus violaceus. Anim. Behav. 81, 1063–1070. doi: 10.1016/j.anbehav.2011.02.018
Kelly, E., and Phillips, B. (2017). Get smart: native mammal develops toad-smart behavior in response to a toxic invader. Behav. Ecol. 28, 854–858. doi: 10.1093/beheco/arx045
Kelly, E., and Phillips, B. L. (2018). Targeted gene flow and rapid adaptation in an endangered marsupial. Conservation Biol. 33, 112–121. doi: 10.1111/cobi.13149
Kotrschal, A., Rogell, B., Bundsen, A., Svensson, B., Zajitschek, S., Brännström, I., et al. (2013a). The benefit of evolving a larger brain: big-brained guppies perform better in a cognitive task. Anim. Behav. 86:e4–e6. doi: 10.1016/j.anbehav.2013.07.011
Kotrschal, A., Rogell, B., Bundsen, A., Svensson, B., Zajitschek, S., Immler, S., et al. (2013b). Experimental evidence for costs and benefits of evolving a larger brain. Curr. Biol. 23, 168–171. doi: 10.1016/j.cub.2012.11.058
Lea, S. E. G., Chow, P. K. Y., Leaver, L. A., and McLaren, I. P. L. (2020). Behavioral flexibility: a review, a model, and some exploratory tests. Learn. Behav. 48, 173–187. doi: 10.3758/s13420-020-00421-w
Lefebvre, L., Reader, S. M., and Sol, D. (2004). Brains, innovations and evolution in birds and primates. Brain Behav. Evol. 63, 233–246. doi: 10.1159/000076784
Lefebvre, L., Whittle, P., Lascaris, E., and Finkelstein, A. (1997). Feeding innovations and forebrain size in birds. Anim. Behav. 53, 549–560. doi: 10.1006/anbe.1996.0330
Lermite, F., Peneaux, C., and Griffin, A. S. (2017). Personality and problem-solving in common mynas (Acridotheres tristis). Behav. Processes 134, 87–94. doi: 10.1016/j.beproc.2016.09.013
Lewis, M. A., Petrovskii, S. V., and Potts, J. R. (2016). The Mathematics Behind Biological Invasions. (Switzerland: Springer).
Lockwood, J. L., Hoopes, M. F., and Marchetti, M. P. (2013). Invasion Ecology. 2nd ed. (London, UK: Blackwell Publishing).
Logan, C. J. (2016a). Behavioral flexibility and problem solving in an invasive bird. PeerJ 4:e1975. doi: 10.7717/peerj.1975
Logan, C. J. (2016b). How far will a behaviourally flexible invasive bird go to innovate? R. Soc. Open Sci. 3:160247. doi: 10.1098/rsos.160247
Logan, C. J., Avin, S., Boogert, N., Buskell, A., Cross, F. R., Currie, A., et al. (2018). Beyond brain size: uncovering the neural correlates of behavioral and cognitive specialization. Comp. Cognit. Behav. Rev. 13, 55–89. doi: 10.3819/CCBR.2018.130008
Lowe, K. A., Taylor, C. E., and Major, R. E. (2011). Do common mynas significantly compete with native birds in urban environments? J. Ornithol. 152, 909–921. doi: 10.1007/s10336-011-0674-5
Lowry, H., Lill, A., and Wong, B. B. (2013). Behavioural responses of wildlife to urban environments. Biol. Rev. Cambridge Phil. Soc. 88, 537–549. doi: 10.1111/brv.12012
Lurz, P. W. W., Gurnell, J., and Magris, L. (2005). Sciurus vulgaris. Mammal. Species 769, 1–10. doi: 10.1644/1545-1410(2005)769[0001:SV]2.0.CO;2
MacDonald, I. M. V. (1997). Field experiments on duration and precision of grey and red squirrel spatial memory. Anim. Behav. 54, 879–891. doi: 10.1006/anbe.1996.0528
Madden, J. R., Langley, E. J. G., Whiteside, M. A., Beardsworth, C. E., and Van Horik, J. O. (2018). The quick are the dead: Pheasants that are slow to reverse a learned association survive for longer in the wild. Phil. Trans. R. Soc. B Biol. Sci. 373:20170297. doi: 10.1098/rstb.2017.0297
Montgomery, S. H., Geisler, J. H., McGowen, M. R., Fox, C., Marino, L., and Gatesy, J. (2013). The evolutionary history of cetacean brain and body size. Evol. 67, 3339–3353. doi: 10.1111/evo.12197
Olkowicz, S., Kocourek, M., Lučan, R. K., Porteš, M., Fitch, W. T., Herculano-Houzel, S., et al. (2016). Birds have primate-like numbers of neurons in the forebrain. Proc. Nat. Acad. Sci. U.S.A. 113, 7255–7260. doi: 10.1073/pnas.1517131113
Quinn, J. L., Cole, E. F., Reed, T. E., and Morand-Ferron, J. (2016). Environmental and genetic determinants of innovativeness in a natural population of birds. Phil. Trans. R. Soc. B. 371:20150184. doi: 10.1098/rstb.2015.0184
Ramey, P. A., Teichman, E., Oleksiak, J., and Balci, F. (2009). Spontaneous alternation in marine crabs: invasive versus native species. Behav. Processes 82, 51–55. doi: 10.1016/j.beproc.2009.04.006
Reader, S. M., and Laland, K. N. (2002). Social intelligence, innovation and enhanced brain size in primates. Proc. Natl. Acad. Sci. U.S.A. 99, 4436–4441. doi: 10.1073/pnas.062041299
Rice, A. M. (2020). The overlooked influence of hybridization on cognition. Front. Ecol. Evol. 8:39. doi: 10.3389/fevo.2020.00039
Roman, J., and Palumbi, S. R. (2004). A global invader at home: population structure of the green crab, Carcinus maenas, in Europe. Mol. Ecol. 13, 2891–2898. doi: 10.1111/j.1365-294X.2004.02255.x
Roudez, R. J., Glover, T., and Weis, J. S. (2008). Learning in an invasive and a native predatory crab. Biol. Invas. 10, 1191–1196. doi: 10.1007/s10530-007-9195-9
Ryan, M., Akre, K. L., and Kirkpatrick, M. (2009). “Cognitive mate choice” in Cognitive Ecology II, eds. R. Dukas, and J. M. Ratcliffe (Chicago: The University of Chicago Press), 137–155.
Shettleworth, S. J. (2001). Animal cognition and animal behaviour. Anim. Behav. 61, 277–286. doi: 10.1006/anbe.2000.1606
Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior. New York, NY: Oxford University Press.
Sih, A., and Del Giudice, M. (2012). Linking behavioural syndromes and cognition: a behavioural ecology perspective. Phil. Trans. R. Soc. B. 367, 2762–2772. doi: 10.1098/rstb.2012.0216
Sol, D., Bacher, S., Reader, S. M., and Lefebvre, L. (2008). Brain size predicts the success of mammal species introduced into novel environments. Am. Nat. 172, S63–S71. doi: 10.1086/588304
Sol, D., Bartomeus, I., and Griffin, A. S. (2012a). The paradox of invasion in birds: competitive superiority or ecological opportunism? Oecologia 169, 553–564. doi: 10.1007/s00442-011-2203-x
Sol, D., Duncan, R. P., Blackburn, T. M., Cassey, P., and Lefebvre, L. (2005). Big brains, enhanced cognition, and response of birds to novel environments. Proc. Nat. Academy Sci. U.S.A. 102, 5460–5465. doi: 10.1073/pnas.0408145102
Sol, D., Griffin, A. S., and Bartomeus, I. (2012b). Consumer and motor innovation in the common myna: the role of motivation and emotional responses. Anim. Behav. 83, 179–188. doi: 10.1016/j.anbehav.2011.10.024
Sol, D., Griffin, A. S., Bartomeus, I., and Boyce, H. (2011). Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS ONE 6:e19535. doi: 10.1371/journal.pone.0019535
Sol, D., and Lefebvre, L. (2000). Behavioural flexibility predicts invasion success in birds introduced to New Zealand. Oikos 90, 599–605. doi: 10.1034/j.1600-0706.2000.900317.x
Sol, D., Timmermans, S., and Lefebvre, L. (2002). Behavioural flexibility and invasion success in birds. Anim. Behav. 63, 495–502. doi: 10.1006/anbe.2001.1953
Sutter, M., and Kawecki, T. J. (2009). Influence of learning on range expansion and adaptation to novel habitats. J. Evol. Biol. 22, 2201–2214. doi: 10.1111/j.1420-9101.2009.01836.x
Tebbich, S., Sterelny, K., and Teschke, I. (2010). The tale of the finch: adaptive radiation and behavioural flexibility. Philos. Trans. R. Soc. Lond. B Biol. Sci. 365, 1099–1109. doi: 10.1098/rstb.2009.0291
Thornton, A., Isden, J., and Madden, J. R. (2014). Toward wild psychometrics: Linking individual cognitive differences to fitness. Behav. Ecol. 25, 1299–1301. doi: 10.1093/beheco/aru095
Tierney, A. J., and Andrews, K. (2013). Spatial behavior in male and female crayfish (Orconectes rusticus): learning strategies and memory duration. Anim. Cognit. 16, 23–34. doi: 10.1007/s10071-012-0547-1
Tierney, A. J., Baker, A., Forward, J., Slight, C., and Yilma, H. (2019). Response and place learning in crayfish spatial behavior. Learn. Behav. 47, 80–90. doi: 10.3758/s13420-018-0345-y
Tierney, A. J., and Lee, J. (2011). Spatial learning in a T-maze by the crayfish orconectes rusticus. J. Comp. Psychol. 125, 31–39. doi: 10.1037/a0020866
Timmermans, S., Lefebvre, L., Boire, D., and Basu, P. (2000). Relative size of the hyperstriatum ventrale is the best predictor of feeding innovation rate in birds. Brain Behav. Evol. 56, 196–203. doi: 10.1159/000047204
Titulaer, M., van Oers, K., and Naguib, M. (2012). Personality affects learning performance in difficult tasks in a sex-dependent way. Anim. Behav. 83, 723–730. doi: 10.1016/j.anbehav.2011.12.020
Tuomainen, U., and Candolin, U. (2011). Behavioural responses to human-induced environmental change. Biol. Rev. 86, 640–657. doi: 10.1111/j.1469-185X.2010.00164.x
van Horik, J. O., and Madden, J. R. (2016). A problem with problem solving: motivational traits, but not cognition, predict success on novel operant foraging tasks. Anim. Behav. 114, 189–198. doi: 10.1016/j.anbehav.2016.02.006
Wright, T. F., Eberhard, J. R., Hobson, E. A., Avery, M. L., and Russello, M. A. (2010). Behavioral flexibility and species invasions: the adaptive flexibility hypothesis. Ethol. Ecol. Evol. 22, 393–404. doi: 10.1080/03949370.2010.505580
Keywords: biological invasions, behavioral flexibility, mammal, bird, fish, amphibian, reptile, integrative review
Citation: Szabo B, Damas-Moreira I and Whiting MJ (2020) Can Cognitive Ability Give Invasive Species the Means to Succeed? A Review of the Evidence. Front. Ecol. Evol. 8:187. doi: 10.3389/fevo.2020.00187
Received: 28 January 2020; Accepted: 26 May 2020;
Published: 30 June 2020.
Edited by:
Laure Cauchard, University of Aberdeen, United KingdomReviewed by:
David Guez, The University of Newcastle, AustraliaGabrielle Davidson, University of Cambridge, United Kingdom
Copyright © 2020 Szabo, Damas-Moreira and Whiting. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin J. Whiting, bWFydGluLndoaXRpbmdAbXEuZWR1LmF1
†ORCID: Birgit Szabo orcid.org/0000-0002-3226-8621
Isabel Damas-Moreira orcid.org/0000-0003-4630-3202
Martin J. Whiting orcid.org/0000-0002-4662-0227