 Kimberley J. Hockings
Kimberley J. Hockings Hannah Parathian2,3
Hannah Parathian2,3 Joana Bessa
Joana Bessa Amelia Frazão-Moreira
Amelia Frazão-Moreira- 1Centre for Ecology and Conservation, College of Life and Environmental Sciences, University of Exeter, Cornwall, United Kingdom
- 2Centre for Research in Anthropology (CRIA – NOVA FCSH), Lisbon, Portugal
- 3Department of Anthropology, Faculty of Social and Human Sciences, New University of Lisbon, Lisbon, Portugal
- 4Department of Zoology, University of Oxford, Oxford, United Kingdom
Understanding the capacity for humans to share resources (crops, wild foods, space) with large-bodied wildlife is vital for biodiversity conservation and human wellbeing, and requires comprehensive examination of their temporal interactions over fine spatial scales. We combined ecological (plant identification, wild fruit availability plots, animal fecal and trace sampling) and social science (free-listing, semi-structured interviews, participant observation) methods to systematically and simultaneously collect data on the availability and selection of fruits from wild plants by humans and critically endangered chimpanzees (Pan troglodytes verus), a national conservation flagship species at Cantanhez National Park, Guinea-Bissau. Within an area of 12.7 km2, we demonstrate that local people’s monthly use of wild fruits was driven by its overall availability in the habitat, whereas chimpanzees, as ripe fruit specialists, sought out fruits year-round. Humans and chimpanzees overlap in the selection of fruits from at least 27 wild plant species. The ranked use of fruits from species which were used by both chimpanzees and humans was significantly positively correlated, suggesting they preferentially target fruits of the same wild plant species. Each month, humans and chimpanzees selected three to six of the same wild fruit species. Chimpanzees fed significantly more on wild fruit species that were available for longer periods, with no effect of that plant species density. Neither plant density nor number of fruiting months impacted human selection of fruit from a plant species, suggesting people might seek out desired resources irrespective of a species’ abundance in the landscape. These findings are important for the development of a shared knowledge base to establish culturally relevant conservation management strategies. We recommend the active management of plant species that are exploited for their fruits by both humans and chimpanzees at Cantanhez National Park, including figs (Ficus spp.), oil-palm (Elaeis guineensis) and velvet tamarind (Dialium guineense). This can be achieved through supporting traditional resource management practices and the strategic replanting of shared plants in deforested areas and degraded corridors between forest fragments. This situation is representative of human-chimpanzee coexistence scenarios found across West Africa; the importance of shared resource use should be incorporated into local, national and regional conservation strategies.
Introduction
Expanding human populations have meant that wildlife increasingly inhabit landscapes in proximity to people (Woodroffe et al., 2005; Karanth et al., 2010; Hockings et al., 2015; Angelici, 2016). Understanding the capacity and mechanisms for humans to coexist with large-bodied wildlife at fine spatial scales is vital for contemporary conservation (Carter et al., 2012) and can inform larger scale conservation efforts (Nyhus, 2016). Effective local conservation strategies should be based on an intricate knowledge of complex social-ecological systems.
Humans change wildlife habitats and ecological systems in numerous ways through highly conspicuous land-use changes, such as large-scale clearing of forests for cultivation, but also through smaller-scale resource exploitation, such as harvesting of wild plants (i.e., non-cultivated) for subsistence and medicinal purposes (Winterhalder, 2001). Agricultural and forager communities rely on access to wild foods as well as foods which they cultivate or actively manage, and these wild resources are highly valued for their medicinal, cultural and subsistence importance (Bharucha and Pretty, 2010). The use of wild plants by local people is part of traditional subsistence systems and often linked with financial security (Belcher et al., 2005), particularly for highly valuable fruiting species. Ethnobotanical studies have shown that people living in rural Africa place great emphasis on the importance of fruit trees, with fruits regularly incorporated into the diet (Campbell, 1987; Malaisse, 1997). If certain wild plant resources are exploited heavily or destructively by people and wildlife, and these resources show low temporal or spatial availability, this might lead to their decline over time. However, if access to plant resources is restricted, for example through government regulations preventing the harvest of rare plant species, this can create hostility between local user groups, and between local people and authorities, which equally might compromise wildlife conservation objectives (Sheil et al., 2012; Redpath et al., 2013). Furthermore, if animals exploit resources which people value it can lead to their retaliatory persecution (Hockings and Humle, 2009). Understanding which plant resources are selected and why, by humans and wildlife within and outside protected areas, is for the most-part lacking, yet is important for integrating the needs of local people with conservation objectives.
Human and non-human primates (hereafter primates) are sympatric across much of the latter’s range (Fuentes and Wolfe, 2002; Estrada et al., 2017) and often exploit the same plant resources, including agricultural crops (Riley, 2007; McLennan et al., 2017). The degree to which humans and primates overlap in their selection of wild plant species is likely underestimated by wildlife researchers, with an assumption that farming communities, as opposed to hunter-gatherers, infrequently use a broad array of forest plants for various reasons (Bharucha and Pretty, 2010). Up to now, detailed examination of the co-utilization of wild plant resources by sympatric humans and primates has received limited attention, and approaches vary for several reasons. Firstly, obtaining data on plant selection by both humans and primates mostly requires different methodological approaches (Sugiyama and Koman, 1992; Koné et al., 2008: Parathian et al., 2018). However, for habituated primates, resource selection can be quantified through direct behavioral observations, with similar participatory observational methods possible for people if few resources are targeted by specific user groups. Secondly, data on plant selection by humans and primates are often collected over different time periods and geographical areas, making it difficult to assess the extent of shared resource use across space and time, and potential ecological drivers (e.g., plant availability) (Chepstow-Lusty et al., 2006; Riley, 2007). Thirdly, investigation can focus on a single plant species that is confirmed as important to humans and primates (Kinnaird, 1992; Reynolds et al., 2012; Sheil et al., 2012; Waller and Pruetz, 2016). Although this approach can yield important information on spatial and temporal overlap in the use of a single resource that is heavily exploited by both, it provides an incomplete picture where humans and primates select numerous plants and plant parts across different seasons. As an alternative, a first step to understand the extent of resource partitioning in sympatric humans and primates is to focus on use of the main food group consumed by the focal primate species (e.g., fruits to frugivores, leaves to foliovores, etc.). However, taking this broader approach to understanding plant selection makes quantifying precise spatial and temporal overlap in plant species selection (i.e., human and primate use of the same zones or plants at the same times) challenging. This applies in particular to sites where little research has been conducted and primates are unhabituated to researchers, precluding direct observations of feeding behavior. In such cases, human and primate plant selection and overlap can be examined over a predetermined spatial scale, for example the ranging area of the focal primate group. To examine co-utilization of multiple wild plant resources by humans and primates in shared landscapes is methodologically challenging, requiring knowledge of plants available in a habitat, and the systematic and simultaneous collection of empirical data on both humans and primates.
Here, we bridge this inter-disciplinary research gap by examining the selection of wild fruits by chimpanzees and local people that coexist in Caiquene-Cadique, Cantanhez National Park, Guinea-Bissau. Chimpanzees occur extensively in areas of anthropogenic influence throughout Africa (Hockings and McLennan, 2012; Hockings et al., 2015). Their diets can consist of several hundred different plant species (e.g., Nishida and Uehara, 1983; Sugiyama and Koman, 1992). However, chimpanzees are not generalist feeders and seek out ripe fleshy fruits which can be seasonal and patchily distributed (Goodall, 1986; Tutin et al., 1997; Hockings et al., 2009). We examine the wild fruit species that are selected by chimpanzees and humans at Caiquene-Cadique, and determine the effects of their density and temporal availability on selection. We directly compare temporal overlap in human and chimpanzee selection of wild fruits as ripe fruits form a large part of chimpanzee diets; humans living in agroforestry environments use fruits for food and medicine; using fecal sampling to examine the importance of items eaten by chimpanzees is biased toward fruits (McLennan, 2013); and the ingestion of fruits and dispersal of seeds has important implications for forest health and the sustainability of shared resource use.
We test the following null hypotheses:
1. the selection of wild fruits by chimpanzees and humans is not determined by the overall temporal availability of wild fruits in the habitat;
2. the selection of wild fruits from individual plant species by chimpanzees and humans do not overlap, including on a monthly basis;
3. the selection of wild fruits from individual plant species by humans and chimpanzees is not affected by the density of the plant species nor duration of fruiting.
Materials and Methods
Study Area
The Cantanhez National Park (CNP) is approximately 1,067 km2 and is a mosaic of subhumid forest, secondary forest, mangrove, savanna, human settlements, and agricultural fields, located in the southern Tombali administrative region of Guinea-Bissau (Figure 1). CNP is managed following the IUCN Category V model for Protected Areas (IUCN 2016), including 14 formally protected forest patches as well as zones allocated to human development. There are two main seasons: the dry season from November to mid-May (with no rainfall) and the rainy season from mid-May to October (Bessa et al., 2015; Bersacola, 2019).
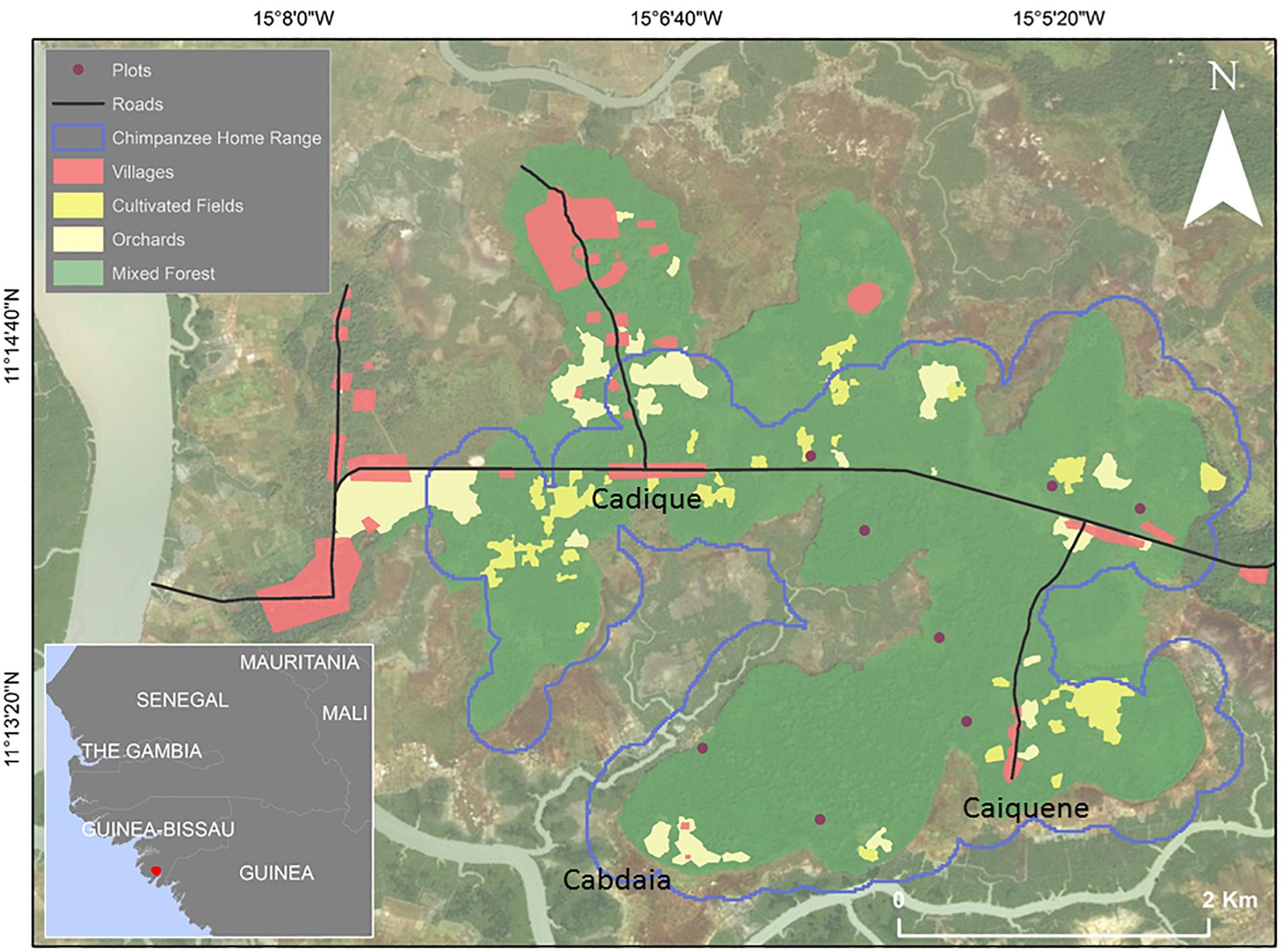
Figure 1. Maps showing the location of Cantanhez NP in West Africa, and the Caiquene-Cadique chimpanzees’ home range (also see Supplementary Figures S1, S2), different habitat types, rivers, roads, and the village locations of the three human communities involved in this study (the Nalú villages of Cadique, Caiquene, and the Balanta village of Cabdaia). Eight randomized phenology plots are shown as circles (see section “Data Collection”).
There are approximately 110 villages within CNP with an estimated human population of 22,000 individuals that are comprised of several ethnic groups (Temudo, 2009). The human communities involved in this study (two Nalú villages and one Balanta village; n = 3) were chosen because of their locations in the study chimpanzee community’s core ranging area and the areas where humans harvest wild plant resources (Figure 1). Aside from these villages, there are some individual houses along the roadside that were not included in this study. The Nalú are a farming and foraging society who exhibit guardianship over the forest. The Nalú régulo (“ruler”) traditionally implements certain rules and regulations which act to regulate plant use during periods where the availability of certain plant species are scarce. The régulo is also in charge of making decisions (with the support of the council) over land distribution and land use. Therefore any person wishing to clear a patch of forest to build a house or cultivate an agricultural area must first be granted permission (Frazão-Moreira, 2016; Parathian et al., 2018). The Balanta have been migrating to this region since the 1950s and are traditionally rice paddy farmers, but more recently farm cashew (Frazão-Moreira, 2009). The Balanta and Nalú consider the chimpanzee as similar to humans and view this species as a non-edible animal (Casanova et al., 2014; Casanova, 2016). The Nalú people’s traditional tolerance toward chimpanzees, which are not hunted, is said to stem from a belief that chimpanzees were previously humans that were punished by God (Sousa and Frazão-Moreira, 2010; Sousa et al., 2014). However, chimpanzees are sometimes killed in retaliation for foraging on crops, especially orange fruits. Increased conversion of forest to other land uses, such as cashew plantations, is putting pressure on dwindling natural resources (Parathian et al., 2018). For additional information on human-chimpanzee coexistence at this site see Parathian et al. (2018).
The western chimpanzee (Pan troglodytes verus) is classified as critically endangered by the IUCN (Humle et al., 2016), with only 17% of western chimpanzees occurring in protected areas (Heinicke et al., 2019a). Guinea-Bissau represents the westernmost limit for chimpanzees, where they are threatened by habitat loss and increasing interactions with local people including disease. Recent models estimate 1,908 chimpanzees (CI: 923–6,121) in Guinea-Bissau (Heinicke et al., 2019a). In CNP and the surrounding areas, chimpanzee population size was estimated at 376–2,632 individuals (Torres et al., 2010); although updated population surveys across Guinea-Bissau are urgently required for more accurate assessments (see Supplementary Figure S1 for map showing the location of CNP). Several chimpanzee communities reside in the central-southern forests of CNP (Hockings and Sousa, 2013; Bersacola, 2019). The Caiquene-Cadique chimpanzee community comprises at least 48 individuals as confirmed through opportunistic observations and camera trap footage (chimpanzee communities across West Africa range in size from 11 to 63 individuals; Herbinger et al., 2001) and range over an area of approximately 12.7 km2 (see Supplementary Figure S2 for analyses of chimpanzee ranging). Moreover, humans and chimpanzees at this site show extensive overlap in habitat selection, with both using areas inside and outside the main forest blocks (Hockings and Sousa, 2012; Bersacola, 2019).
Data Collection
Plant Identification and Food Availability
We collected cross-disciplinary data from 4th February to 14th November 2013, with data collection carried out during wet and dry months (see Supplementary Table S1 for an overview of methods). We collected phenology data in eight 50 m × 50 m plots (Figure 1) selected randomly across the study site. Plots incorporated different habitat types, including primary subhumid forest, secondary forest and palm groves. Plots were accessible to humans and chimpanzees and did not cross sacred areas of forest (Frazão-Moreira, 2009). Although not planted, people protect some wild tree species that are deemed useful, for example by not cutting them down and protecting young saplings. This is a feature of an anthropogenic landscape in which the human population has an intricate relationship with the environment. In order to minimize habitat disturbance and reduce the risk of creating paths for hunters, transects were not cut for this research.
We identified and marked trees and lianas with a diameter at breast height (DBH) ≥10 cm (Chapman et al., 2005) with metal numbered tags; these totaled 1,994 trees/lianas from 124 species (65 known by their scientific name, 27 by their local name only, and 32 unknown). We compiled species lists and counted the number of each plant species per hectare (Serckx et al., 2014). We monitored each tree and liana every first and third week per month for a total of 9 months (i.e., 18 phenology surveys) and collected data on the percentage cover of ripe fruits within the plots (n = 35,892 data points). The DBH provides an accurate indicator of a tree’s size and hence its ability to produce fruit, with high inter-observer reliability (Chapman et al., 1994). We used the following scores to record the presence of fruit: 0 (absent), 1 (1–25% of canopy coverage), 2 (26–50% coverage), 3 (51–75% coverage), 4 (76–100% coverage). We calculated a twice-monthly ripe fruit availability index (FAI) for plant species present in plots using the following formula, where Pi is the basal area of the tree (cm2) and Fi is the fruit score of the tree (0–4) (Hockings et al., 2009): FAI=[S(Pi × Fi)]/[S(Pi × 4)] × 100. We calculated the FAI using the availability of fruits from all species found in the plots. Our data showed that the twice-monthly availability of ripe fruits from plant species consumed by chimpanzees (n = 834 trees and lianas from 37 species) was positively correlated with availability of ripe fruits from all plant species in the plots (Spearman’s rank: Rs = 0.983, n = 18, p < 0.001).
At the start of the research period we carried out free-listing (Albuquerque et al., 2014) with 12 adult male informants to identify plant species that were potentially used by local people thus allowing us to compile a plant list for the semi-structured interviews (see section “Mixed-Methods to Examine Human Resource Use” below). This method assumes that informants list what they perceive as the most salient items. Informants included the chief from each of the three villages and men who were locally recognized for their extensive knowledge of plants for food and medicine and selected by the chief to assist. This method was carried out by the two social science researchers who spoke with informants explaining what information was required, why it was needed and how it would be used. To identify unknown plant species, we conducted botanical sampling in the phenology plots and through participating in activities with local people, such as gathering wild foods and medicinal plants. To obtain local names of unknown plants we showed them to local botanical experts. To identify them scientifically we used the botanical guides of Guinea-Bissau (Catarino et al., 2006), and were assisted by a specialist team at the Herbário LISC, Instituto de Investigação Científica Tropical in Lisbon. Voucher specimens are kept in the herbarium of the Centre for Research in Anthropology in Lisbon.
Chimpanzee Resource Use
To examine wild plant utilization by chimpanzees, we collected data on feeding behavior through indirect recordings (from fecal samples and feeding traces) and direct opportunistic observations (where chimpanzees were seen eating plants). Chimpanzees were unhabituated to human observers which precluded systematic observations of feeding behavior. The decision was made not to habituate chimpanzees at this site as humans and chimpanzees encounter each other on a daily basis, and habituation for research might be a contributing factor driving aggressive interactions (McLennan and Hockings, 2016).
Chimpanzee fecal sampling, processing and analysis followed well established methods by Tutin and Fernandez (1993), McGrew et al. (2009) and McLennan (2013) to quantify seasonal variation in the relative amount of fruit and foliage in the diet. All fecal samples (n = 377; monthly mean = 41.9 ± 7.5; n = 9 months) were less than 2 days old. Chimpanzee feces were easily distinguishable from feces of other sympatric primate species including Temminck’s red colobus (Piliocolobus badius temminckii), western black and white colobus (Colobus polykomos), Green monkey (Chlorocebus sabaeus), Guinea baboon (Papio papio), and Campbell’s monkey (Cercopithecus campbelli). To ensure independence of data, only one sample was collected if nearby fecal matter looked like it was from the same individual. We washed fecal samples through a fine-meshed sieve and laid the undigested content out on newspaper sheets for immediate analysis. We scored fecal content using volume percentage values for fruit (“fruit score” is the overall percentage of pulp and seeds), and other parts (leaves, pith, flower, and bark, and other foods), according to the total mass of the fecal sample (Kuroda et al., 1996). Fruit pulp is easily distinguishable from other non-fruit parts. Where possible we identified fruit to species level based on the seeds and undigested fruit parts, and counted the numbers of fruit species in each fecal sample. Quantitative data on fruit ingestion were only available for chimpanzees if the fruit was identified in feces via presence of seeds or pulp. We recorded chimpanzee feeding traces that were less than three days old. We attributed a trace to chimpanzees if a feeding group had been previously observed at the site, or according to the presence of species-specific signs such as knuckle prints, nests, bark stripping, or fruit wadges (chimpanzees place several fruits in the mouth and extract the juices, discarding a wadge). We collected a botanical sample for identification. Bessa et al. (2015) provided an in-depth analysis of chimpanzee feeding behavior at Caiquene-Cadique including use of agricultural crops and flowers, leaves, and pith in the diet. This paper focuses on use of wild fruits by chimpanzees and we exclude their use of other plant parts and cultivated plants from our analyses. Research with chimpanzees was reviewed and approved by the Centre for Research in Anthropology (CRIA – NOVA FCSH), Lisbon, Portugal. This research was also reviewed and approved by the Instituto da Biodiversidade e das Áreas Protegidas (IBAP) in Guinea-Bissau. All research involving wild chimpanzees was non-invasive and strictly adhered to ethics guidelines detailed by the Association for the Study of Animal Behaviour (United Kingdom).
Mixed-Methods to Examine Human Resource Use
There was a total of 490 people and 49 households in the three study villages based on our own census (Cadique, n = 311 people, n = 29 households; Caiquene, n = 145, n = 15; Cabdaia, n = 34, n = 5). We were given permission by the chief from each village to repeatedly visit families and request that someone from each household complete the survey on plant use for that household. With the help of a local Nalú guide we tried to speak to the same person each week, preferably the female head of each household deemed to be the person with the most knowledge about plant food utilization, to ensure consistency. However, on occasions where the person was out or otherwise occupied, we interviewed another family member instead.
Concurrently with chimpanzee research we carried out weekly semi-structured interviews (Bernard, 1995; Cunningham, 2001), visiting all 49 households for 36 weeks to record plant use by local people (n = 8,380 reports of plant use). The use of a plant part (e.g., fruit, flower, leaf, bark, seed, etc.) from a particular plant species was recorded once per household per week to ensure independence of data. Each household used multiple plant species per week. As data becomes increasingly unreliable when recall takes place over longer periods (Godoy et al., 1993), we asked people to recall which plants their household had used, collected and consumed in the previous seven days only. Participants were asked to provide the local name of the plant species their household used that week, which plant part they had used (i.e., fruit, flower, leaf, bark, seed, etc.) and for what purpose. Following Frazão-Moreira (2009), we recorded plant-use under five categories: consumables (i.e., food), medicine, fuel (i.e., firewood), artifacts (including tools), and construction. From data collected during these semi-structured interviews, we calculated Use Value (UV) integers for fruits of each plant species using the following calculation: UV = ∑Ui/n, where Ui = the number of times fruit of a given species is used, n = the total number of informants at the family level (Phillips and Gentry, 1993). To examine which plant species were most used by local people, we ranked the total UV fruit scores for each plant species. A plant’s relative importance is low if it is used infrequently by all informants (as it functions as a “general use” plant harvested in small amounts). While we are aware that some rarely used fruiting plant species can fill highly specialized roles for people living in forest communities (Kristensen and Balslev, 2003), we decided that assessing plant importance according to overall frequency of human use would allow us to make more accurate comparisons with chimpanzees. We obtained additional information on fruit selection by local people, such as extraction methods and the potential commercial value of different fruit species, through participant observation of fruit harvesting, processing practices and subsistence or commercial use (Spradley, 1980; Bernard, 1995). When fruit extraction was either directly observed or traces identified, we recorded the plant species and harvesting technique (Cunningham, 2001). Research with local people was reviewed and approved by the Centre for Research in Anthropology (CRIA – NOVA FCSH), Lisbon, Portugal. Research followed Ethical Guidelines for Good Research Practice set by the Association of Social Anthropologists of the United Kingdom and Commonwealth. Oral consent was obtained from people, but written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Results
Wild Plants
We took samples of 193 wild plant species of which 143 were identified by their scientific and local names, 16 were identified by local name only, and 34 remain unidentified. Ripe fruit availability was highly seasonal, and was approximately four times higher during the dry season than the rainy season (mean ripe fruit FAI: dry season 4.14 ± SD 2.16, N = 8 phenology surveys, versus rainy season 1.14 ± SD 1.65, N = 10).
Chimpanzee Plant Use
Chimpanzees consumed a minimum of 57 wild plant species, from at least 25 families, including six plant parts (fruit, leaf, pith, bark, flower, sap). Numerically, fruit dominated the chimpanzees’ diet, with at least 46 wild fruit species consumed. Wild fruit was present in 97.6% of fecal samples, with a mean monthly wild fruit score of 79% (SD ± 10.4). Fluctuations in the availability of ripe fruit (FAI) was not a predictor of chimpanzee wild fruit score and explained only 2.5% of variance (Linear regression: b ± SE: −0.742 ± 1.288, df = 1,14, t = −0.576, p = 0.575; Figure 2A), thus indicating that, chimpanzees maintained a diet dominated by wild fruit irrespective of availability.
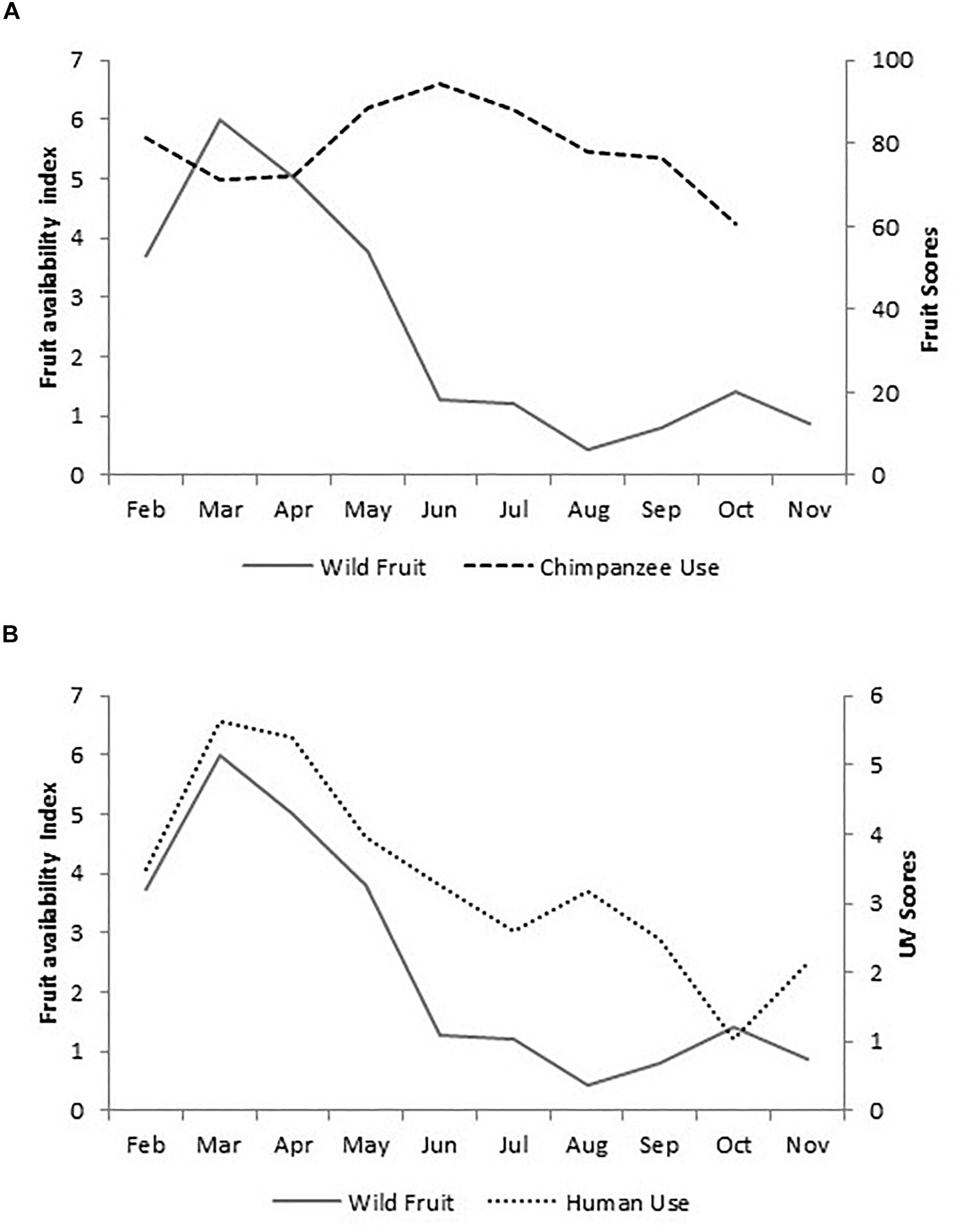
Figure 2. Mean monthly values for wild fruit availability (FAI) plotted against (A) chimpanzee fruit scores (scores for November are not available), and (B) human UV scores.
Human Plant Use
Local people used a minimum of 142 plant species (129 confirmed by their local and scientific name, 13 by their local name only). Households utilized the different parts of wild plants in multiple ways (Supplementary Table S2), with the fruits of at least 47 plant species used as consumables and medicine. Wild fruit availability strongly predicted human fruit use values (Linear regression: b ± SE: 0.567 ± 0.108, df = 1,17, t = 5.269, p < 0.001; Figure 2B), explaining 63.4% of variation, thus indicating that human use was influenced by availability.
Wild Fruit Selection by Chimpanzees and Humans
Humans and chimpanzees overlap in the selection of fruits from at least 27 wild plant species (Supplementary Table S3). Humans use the fruits of all 27 species for food and three species for medicine, with crossovers in what people consider food and medicine. The overall ranked use of fruits from species which were used by both chimpanzees (percentage of total fecal samples containing that fruit species) and humans (UV scores) was significantly positively correlated (Rs = 0.568, n = 19 wild fruit species with fruit scores confirmed through fecal samples, p = 0.011), suggesting that humans and chimpanzees preferentially target fruits of the same plant species (also see Figure 3). Some fruit species have relatively low use by both humans and chimpanzees, and others are more heavily used including figs (Ficus spp.), oil-palm (Elaeis guineensis) and the velvet tamarind (Dialium guineense), albeit with monthly variation in overlap (Figure 3). Each month, humans and chimpanzees overlapped in their use of three to six fruit species (mean ± SD: 4.33 ± 1.22), but also each fed on certain fruit species exclusively (Figure 4 and Supplementary Figure S3).
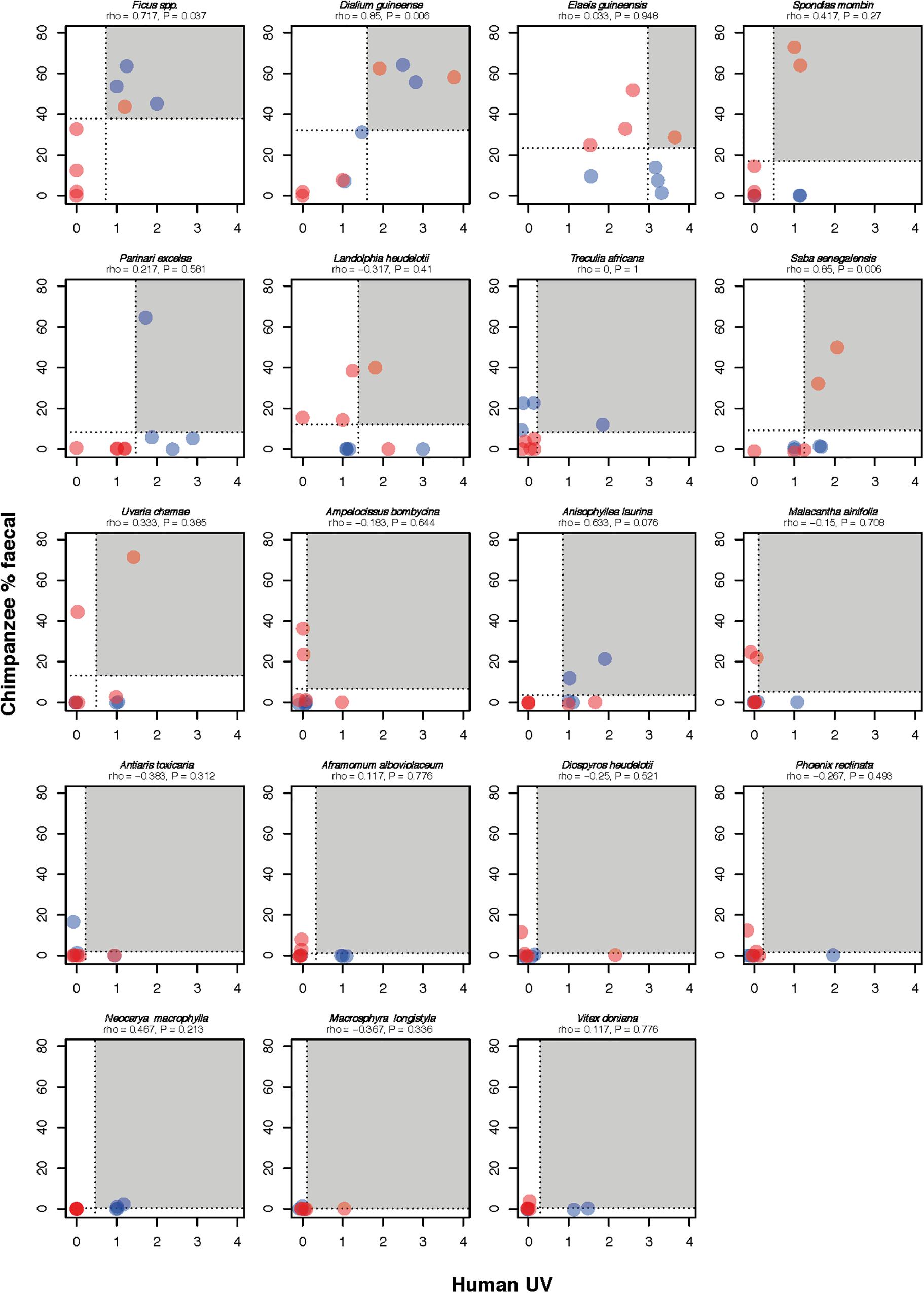
Figure 3. Monthly selection of fruits from 19 plant species used by both humans and chimpanzees (with chimpanzee selection confirmed through fecal analysis and human selection confirmed through weekly semi-structured interviews). Plant species are ordered by importance in chimpanzee diet. Months are color coded by season: red for rainy season, blue for dry season. Dashed lines indicate the mean usage for that month/plant species, and the gray area shows the values that are above average for both species. Spearman’s rank correlation coefficient is shown for fruit usage for each plant species.
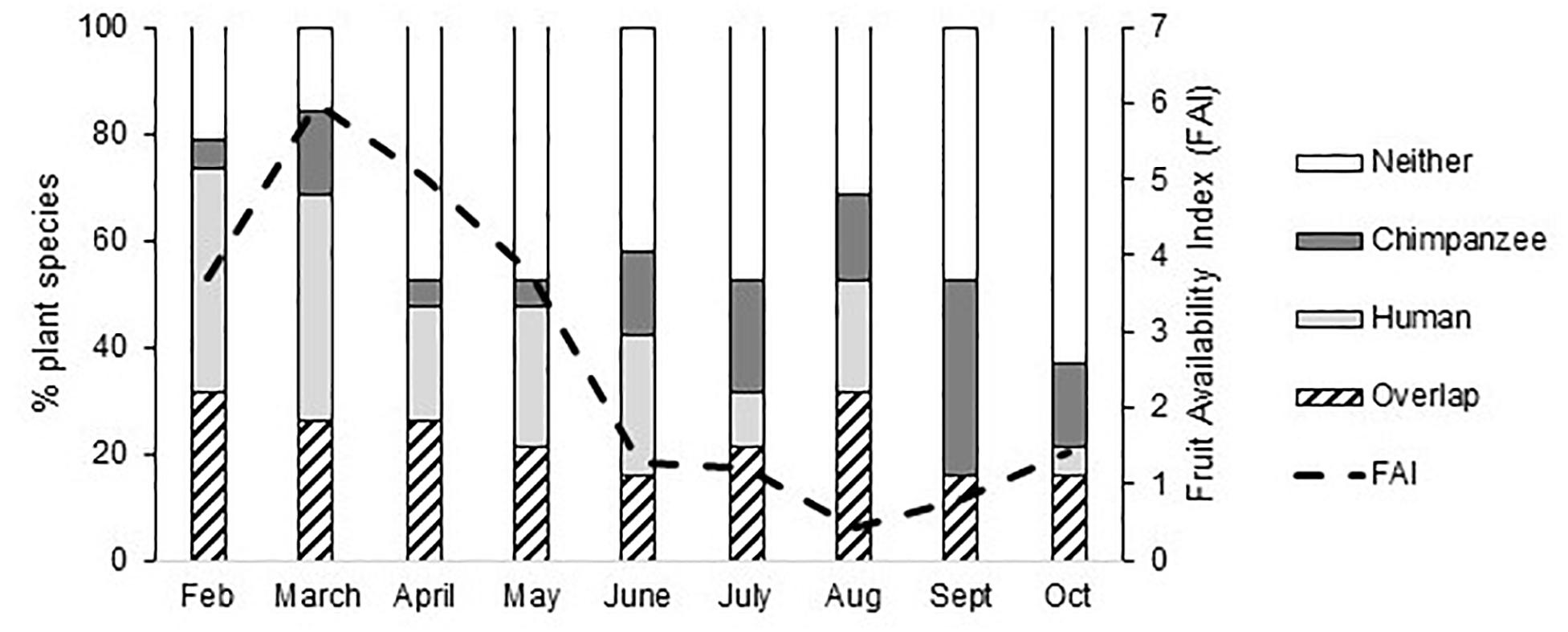
Figure 4. The monthly selection of fruits from wild plant species (n = 19) by chimpanzees and humans (Overlap), humans only (Human), chimpanzees only (Chimpanzee), and not used by either (Neither), potentially because the fruits of some species are not available during that month, and monthly Fruit Availability Index (FAI).
For chimpanzees, the density of a plant species together with the number of months it fruited accounted for 44.2% of variation in fruit consumption. The number of months a plant species was fruiting significantly impacted chimpanzee fruit consumption (Linear Regression: b ± SE: 0.492 ± 0.220, df = 2,13, t = 2.235, p = 0.047), whereas the density of a plant species alone did not have a significant effect (b ± SE: 0.021 ± 0.037, df = 2,13, t = 0.563, p = 0.585). There was no significant effect of the density of a plant species (b ± SE: 0.019 ± 0.012, df = 2,17, t = 1.651, p = 0.120) nor number of fruiting months (b ± SE: 0.099 ± 0.056, df = 2,17, t = 1.769, p = 0.097) on human fruit use.
Discussion
In depth understanding of the co-utilization of wild resources by humans and wildlife can be incorporated into landscape, regional and national conservation policy that acknowledges the needs of both. Overall, local people and chimpanzees at CNP used fruits from a large range of wild plant species. People mostly targeted wild fruits during periods of seasonal abundance, relying on cultivated resources throughout the year, hence not supporting hypothesis one. Abundant fruits might be more visible and easier to collect, but local community leaders also control periods of extraction of certain resources to ensure their continued availability (Frazão-Moreira, 2009). Chimpanzees maintained a diet predominantly comprised of wild fruit year-round, seeking out wild fruits even when availability within the habitat was low. It is likely that chimpanzees use a greater diversity of wild fruiting plants than we identified, as the number of known plant species in chimpanzee diet increases with research duration (Matsuzawa et al., 2011). Furthermore, we relied on indirect methods to quantify chimpanzee fruit selection and some fruit species that were infrequently consumed were likely missed. Our study took place over a 9-month period and although we ensured equal sampling effort across the wet and dry seasons, future dietary studies should aim to collect longer-term data. As human selection of fruits strongly followed overall monthly fruit availability, overall resource partitioning between humans and chimpanzees over wild fruits at Caiquene-Cadique is probably higher when fewer fruits are available in the habitat.
Humans and chimpanzees overlapped extensively in their use of fruits from the same wild plant species and preferentially target some of the same wild fruits at the same times, hence we reject hypothesis two. We have identified specific plant species and periods of the year where attention needs to be taken to avoid over-exploitation of fruits, to ensure chimpanzees are not detrimentally impacted by human over-harvesting. Our scientific data show which plant species should be prioritized for replanting in corridors between forest fragments and which should be afforded additional protection to ensure their persistence and long-term sustainable use by humans and chimpanzees at CNP. However, many of the fruiting plant species identified as important in this study are present throughout West Africa. Even though chimpanzee and human communities across West Africa will exhibit variations in resource use, strategies for their conservation should be prioritized and incorporated into systematic and evidence-based conservation policy (e.g., Heinicke et al., 2019b; IUCN, 2020). This may offer a lifeline for chimpanzee populations throughout the region, especially as most live in unprotected areas that are undergoing extensive habitat change. For example, the most frequently used wild plant by local people at CNP was oil-palm (Elaeis guineensis), and the fruit was used especially for making cooking oil and soap. Oil-palm is found at high densities across the study site, it was available year-round, and its fruits were an important food for chimpanzees. Mature oil palms are fire resistant and survive slash-and-burn activities, and hence this species is found frequently in cultivated and fallow areas as well as forest edges. This situation is representative of that found in many other locales in West Africa (e.g., Humle and Matsuzawa, 2004; Garriga et al., 2019) and special attention should be given to the national and regional conservation of this shared resource.
Neither human nor chimpanzee selection of fruits from individual plant species was driven by their density. Unlike humans, chimpanzees fed more on fruits from plant species that had longer fruiting periods. If human use of fruits from these plant species increases or their occurrence within the habitat decreases, it is possible that in the short-term chimpanzees can switch to alternative wild foods (Reynolds et al., 2012) and crops (Hockings et al., 2009). In the long-term overlap in the use of certain fruits might increase, possibly reducing the density of chimpanzees that can persist in this type of mosaic landscape which characterizes much of West Africa. As the drive to convert more forest to other land uses continues (e.g., forest to cashew, Anacardium occidentale, plantations in CNP; Hockings and Sousa, 2012), and cultural practices and traditional land management techniques are changed or ignored, there will be increased pressure on shared fruit resources. Additionally, risks of zoonotic disease transmission are likely higher in shared landscapes such as CNP. Effective strategies to limit the transmission and spread of emerging infectious disease should be based on an intricate knowledge of overlapping resource utilization and use of space by humans and wildlife, as well as a better understanding of how environmental stressors impact susceptibility to infectious disease.
Traditionally local people in this agroforest mosaic landscape live alongside wildlife and share with them the forest and resources (Parathian et al., 2018). During an intensive 1-year camera trap study, Bersacola (2019) found no evidence for overall site or habitat partitioning between chimpanzees and humans at Caiquene-Cadique. Due to the restricted ranging area of this chimpanzee community, it is very likely that they are targeting the same locales as local people to access fruits. To accurately quantify this spatial overlap in fruit selection would require different methods. This may not be possible for unhabituated chimpanzee populations as fecal samples (which are used to quantify the fruit species in the diet) are not reliably collected from a feeding site and camera traps will not capture all feeding events. As important shared fruit resources have now been identified, we recommend the longer-term quantification of extraction levels of fruits and other plant parts from these species, in particular oil-palm and velvet tamarind (Dialium guineense), by humans and chimpanzees at CNP and in the broader landscape. Although challenging, the goal would be to model the sustainability of human-wildlife interactions to test different resource exploitation scenarios, and to produce extraction maps to identify relative intensity of use across the landscape and region more generally, and determine how this overlaps with chimpanzee distribution inside and outside protected areas. As chimpanzees can access most places, complete separation of areas for human and wildlife use are unrealistic. However, these maps would reveal areas of shared resource use which have the potential to be over-exploited and thus where “competition” could be potentially greater, as well as identifying degraded areas for the replanting of important shared resources that encourages habitat regeneration. To answer landscape-level questions on resource overlap, there is a need to explore different methods to determine the importance of wild plants to humans and chimpanzees, for example through the use of molecular markers or isotopes in wildlife feces or hair (Loudon et al., 2016).
No research has been conducted on the ecological and spatial resilience of the shared landscape at CNP. Although we focus on the chimpanzee (a large-bodied fruit specialist that plays an important ecosystem function, especially in seed dispersal, and are a priority/flagship species for conservation in Guinea-Bissau), many other wildlife species also likely rely on these same plant resources. To guide habitat restoration efforts, data are urgently needed on the ecological role of the important plant species identified within the wider ecosystem and across habitat types (Catarino and Palminha, 2018; Catarino et al., 2020). To inform natural resource management action and maximize the likelihood of long-term conservation success, understanding of the broader ecosystem functioning in such forest-agricultural ecotones should be integrated with more local and direct information on human-wildlife interactions including resource overlap (Chambers et al., 2019).
Human population growth is a critical factor for the sustainability of human-wildlife interactions. It should also be recognized however, that the cultural beliefs and traditional land management practices of human communities play a crucial role in the resources available to wildlife, as well as forest longevity. The importance of wild fruits is recognized by local people as they too rely on these for their subsistence and medicinal needs. However, despite extensive local ecological knowledge on the properties of different wild plants, people may not be aware of specific patterns in chimpanzee foraging. Shared ecological knowledge bases that inform conservation strategy should be a priority in landscapes like CNP. The importance of low level extraction of forest resources by local communities is also an often overlooked component of human well-being. Guinea-Bissau is one of the most economically poor countries in the world, with 67% of the population living below the poverty line of $1.90 USD/day (World Bank, 2019). Most local people in CNP rely on traditional medicines extracted from local plants, and forest fruits are a crucial source of vitamins and minerals for young children. Unlike many other protected areas where resource use by local people is controlled, placing strict controls on the subsistence harvest of wild plants at CNP is neither feasible nor ethical. However, some of these same plant species used as a subsistence food by local people (e.g., the fruits of Saba senegalensis) are harvested at high levels and sold elsewhere in West Africa (Waller and Pruetz, 2016). If human exploitation of forest products changes from subsistence to commercial use, for example if infrastructural development allowed for improved access to markets, this would inherently change the dynamics of wild resource use (Belcher et al., 2005), and potentially human-chimpanzee interactions.
The “biocultural” conservation of traditional strategies to manage shared landscapes is an existing tool that supports the sustainability of valuable plant resources. However, we should also carefully monitor the impact of human activities especially if they are destructive or influence growth and fruiting of important shared plant species. With the growth of large-scale mono-crop agriculture, forest is cut down to grow cash crops and the wild plants that were once highly valued by previous generations can be devalued and replaced by new measures of prosperity such as economic wealth. This has already led to rapid land use cover change from shifting agriculture to planting permanent cashew orchards (Temudo and Abrantes, 2014). Instead of self-reliance through a complex mosaic of agricultural fields, fallows and forest patches, increased dependence on a homogenous landscape of cashew agroforests is high risk. It impacts both the natural environment and local livelihoods. For conservation efforts to succeed, we need to work with local people to support tolerance towards wildlife and merge traditional practice and knowledge with solutions to protect important plant species whilst maintaining forest cover, connectivity, and ecological resilience.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
The studies involving human participants were reviewed and approved by the Centre for Research in Anthropology (CRIA – NOVA FCSH), Lisbon, Portugal. Research followed Ethical Guidelines for Good Research Practice set by the Association of Social Anthropologists of the United Kingdom and Commonwealth. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements. The animal study was reviewed and approved by the Centre for Research in Anthropology (CRIA – NOVA FCSH), Lisbon, Portugal. This research was also reviewed and approved by the Instituto da Biodiversidade e das Áreas Protegidas (IBAP) in Guinea-Bissau. All research involving wild chimpanzees was non-invasive and strictly adhered to ethics guidelines detailed by the Association for the Study of Animal Behaviour (United Kingdom).
Author Contributions
KH conceived the ideas, led the writing of the manuscript. KH, HP, JB and AF-M designed methodology. KH, HP, JB and AF-M collected the data. KH, JB and HP analyzed the data. All authors contributed critically to the drafts and gave final approval for publication.
Funding
The authors were supported by a fellowship and research grant (IF/01128/2014 and PTDC/CS-ANT/121124/2010) from Fundação para a Ciência e a Tecnologia (FCT), Portugal to KH; a fellowship from the Centre for Research in Anthropology (CRIA – NOVA FCSH) to HP (CRIA/04038/BPD/DASE); and Darwin Initiative funding (Grant Number: 26-018), United Kingdom, to KH.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank to the chiefs and communities where we conducted our research, and our local guides Mamadu Cassamá, Djibi Indjai, and Iaia Camara. Acknowledgements are due to the Instituto da Biodiversidade e das Áreas Protegidas (IBAP) for permission to carry out this research and for support with field logistics. We thank the team at the Herbário LISC, Instituto de Investigação Científica Tropical in Lisbon, in particular Luís Catarino, for help identifying plant samples. We also thank Catherine Hill, Matthew McLennan, Kristian Metcalfe, Ruth Thurstan, and Elena Bersacola for comments, Erik Postma for advice on data analysis and the production of graphs, and Chris Kerry for chimpanzee ranging analysis. We would like to dedicate this manuscript to the memory of Cláudia Sousa who passed away after contributing so much of her time and energy to helping with this research.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.00123/full#supplementary-material
References
Albuquerque, U. P., Cunha, L., Lucena, R., and Alves, R. (2014). Methods and Techniques in Ethnobiology and Ethnoecology. New York: Springer.
Belcher, B., Ruız-Perez, M., and Achidiawan, R. (2005). Global patterns and trends in the use and management of commercial NTFPs. World Dev. 33, 1435–1452. doi: 10.1016/j.worlddev.2004.10.007
Bernard, H. R. (1995). Research Methods in Anthropology. Qualitative and Quantitative Approaches. Walnut Creek: Altamira Press.
Bersacola, E. (2019). Zooming in on Human-Wildlife Coexistence: Primate Community Responses in a Shared Agroforest Landscape in Guinea-Bissau. PhD dissertation, Oxford Brookes University, Oxford.
Bessa, J., Sousa, C., and Hockings, K. J. (2015). Feeding ecology of chimpanzees (Pan troglodytes verus) inhabiting a forest-mangrove-savanna-agricultural matrix at caiquene-cadique, cantanhez national park, guinea-bissau. Am. J. Primatol. 77, 651–665. doi: 10.1002/ajp.22388
Bharucha, Z., and Pretty, J. (2010). The roles and values of wild foods in agricultural systems. Philos. Trans. R. Soc. B 365, 2913–2926. doi: 10.1098/rstb.2010.0123
Campbell, B. M. (1987). The use of wild fruits in zimbabwe. Econ. Bot. 41, 375–385. doi: 10.1007/bf02859054
Carter, N. H., Shrestha, B. K., Karki, J. B., Pradhan, N. M. B., and Liu, J. (2012). Coexistence between wildlife and humans at fine spatial scales. PNAS 109, 15360–15365. doi: 10.1073/pnas.1210490109
Casanova, C. (2016). Serão os não humanos os ultimos “outros” na Antropologia? Representações sobre a superioridade humana. Trab. Antropol. Etnol. 56, 106–134.
Casanova, C., Sousa, C., and Costa, S. (2014). Are animals and forests forever? Perceptions of wildlife at Cantanhez Forest National Park, Guinea-Bissau. Memórias 16, 69–104.
Catarino, L., Frazao-Moreira, A., Bessa, J., Parathian, H., and Hockings, K. J. (2020). Plants Used by Chimpanzees and Humans in Cantanhez, Guinea-Bissau – Field Guide. Lisboa: LAE/CRIA.
Catarino, L., and Palminha, A. (2018). Plano de Gestão das Florestas do Parque Nacional de Cantanhez. Inventário Florestal do Parque Nacional de Cantanhez. Bissau: Instituto da Biodiversidade e das Areas Protegidas.
Catarino, L. M., Martins, E. S., Pinto-Basto, M. F., and Diniz, M. A. (2006). Plantas Vasculares e Briófitos da Guiné-Bissau. Lisboa: Instituto de Investigação Cientifica Tropical.
Chambers, J. C., Allen, C. R., and Cushman, S. A. (2019). Operationalizing ecological resilience concepts for managing species and ecosystems at risk. Front. Ecol. Evol. 7:241. doi: 10.3389/fevo.2019.00241
Chapman, C. A., Chapman, L. J., Struhsaker, T. T., Zanne, A. E., Clark, C. J., and Poulsen, J. R. (2005). A long-term evaluation of fruiting phenology: importance of climate change. J. Trop. Ecol. 21, 31–45. doi: 10.1017/s0266467404001993
Chapman, C. A., Wrangham, R., and Chapman, L. J. (1994). Indices of habitat-wide fruit abundance in tropical forest. Biotropica 26, 160–171.
Chepstow-Lusty, A., Winfield, M., Wallis, J., and Collins, A. (2006). The importance of local tree resources around Gombe National Park, western Tanzania: implications for humans and chimpanzees. AMBIO 35, 124–129. doi: 10.1579/0044-7447(2006)35[124:tioltr]2.0.co;2
Cunningham, A. B. (2001). Applied Ethnobotany: People, Wild Plant use and Conservation. London: EarthScan.
Estrada, A., Garber, P. A., Rylands, A. B., Roos, C., Fernandez-Duque, E., Di Fore, A., et al. (2017). Impending extinction crisis of the world’s primates: Why primates matter. Sci. Adv. 3:e1600946. doi: 10.1126/sciadv.1600946
Frazão-Moreira, A. (2009). Plantas e “Pecadores”. Percepções da Natureza em África. Lisboa: Livros Horizonte.
Frazão-Moreira, A. F. (2016). Challenging approaches and crossovers in anthropology and conservation in Guinea-Bissau. Etnografica 20, 663–667. doi: 10.4000/etnografica.4762
Fuentes, A., and Wolfe, L. D. (2002). Primates Face to Face: the Conservation Implications of Human-Nonhuman Primate Interconnections. Cambridge: Cambridge University Press.
Garriga, R. M., Marco, I., Casas-Díaz, E., Acevedo, P., Amarasekaran, B., Cuadrado, L., et al. (2019). Factors influencing wild chimpanzee (Pan troglodytes verus) relative abundance in an agriculture-swamp matrix outside protected areas. PLoS One 14:e0215545. doi: 10.1371/journal.pone.0215545
Godoy, R., Lubowski, R., and Markandaya, A. (1993). A method for the economic valuation of non-timber forest products. Econ. Bot. 47, 220–233. doi: 10.1007/bf02862288
Goodall, J. (1986). The Chimpanzees of Gombe: Patterns of Behaviour. Cambridge, MA: Harvard University Press.
Heinicke, S., Mundry, R., Boesch, C., Amarasekaran, B., Barrie, A., Brncic, T., et al. (2019a). Advancing conservation planning for western chimpanzees using IUCN SSC A.P.E.S. – the case of a taxon-specific database. Environ. Res. Lett. 14:064001. doi: 10.1088/1748-9326/ab1379
Heinicke, S., Mundry, R., Boesch, C., Hockings, K. J., Kormos, R., Ndiaye, P. I., et al. (2019b). Towards systematic conservation planning for western chimpanzees. Am. J. Primatol. 81:e23042. doi: 10.1002/ajp.23042
Herbinger, I., Boesch, C., and Rothe, H. (2001). Territory characteristics among three neighboring chimpanzee communities in the Taï National Park, Côte d’Ivoire. Int. J. Primatol. 22, 143–167.
Hockings, K. J., Anderson, J. R., and Matsuzawa, T. (2009). Use of wild and cultivated foods by chimpanzees at Bossou, Republic of Guinea: feeding dynamics in a human-influenced environment. Am. J. Primatol. 71, 636–646. doi: 10.1002/ajp.20698
Hockings, K. J., and Humle, T. (2009). Best Practice Guidelines for the Prevention and Mitigation of Conflict Between Humans and Great Apes. Gland: IUCN/SSC, 40.
Hockings, K. J., and McLennan, M. (2012). From forest to farm: systematic review of cultivar feeding by chimpanzees – management implications for wildlife in anthropogenic landscapes. PLoS One 7:e33391. doi: 10.1371/journal.pone.0033391
Hockings, K. J., McLennan, M. R., Carvalho, S., Ancrenaz, M., Bobe, R., Byrne, R., et al. (2015). Apes in the Anthropocene: flexibility and survival. Trends Ecol. Evol. 30, 215–222. doi: 10.1016/j.tree.2015.02.002
Hockings, K. J., and Sousa, C. (2012). Differential exploitation of cashew - a low conflict crop - by sympatric humans and chimpanzees. Oryx 46, 375–381. doi: 10.1017/s003060531100130x
Hockings, K. J., and Sousa, C. (2013). Human-chimpanzee sympatry and interactions in Cantanhez National Park, Guinea-Bissau: current research and future directions. Primate Conserv. 26, 57–65. doi: 10.1896/052.026.0104
Humle, T., and Matsuzawa, T. (2004). Oil palm use by adjacent communities of chimpanzees at Bossou and Nimba Mountains, West Africa. Int. J. Primatol. 25, 551–581. doi: 10.1023/b:ijop.0000023575.93644.f4
Humle, T., Boesch, C., Campbell, G., Junker, J., Koops, K., Kuehl, H., et al. (2016). Pan troglodytes ssp. verus (errata version published in 2016). Red List Threat. Species 2016:e.T15935A102327574.
IUCN (2020). Regional Action Plan for the Conservation of Western Chimpanzees (Pan troglodytes verus) 2020–2030. Gland: IUCN SSC.
Karanth, K. K., Nichols, J. D., Karanth, K. U., Hines, J. E., and Christensen, N. L., Jr. (2010). The shrinking ark: patterns of large mammal extinctions in India. Proc. R. Soc. B 277, 1971–1979. doi: 10.1098/rspb.2010.0171
Kinnaird, M. F. (1992). Competition for a forest palm: use of Phoenix reclinata by human and nonhuman primates. Conserv. Bio. 6, 101–107. doi: 10.1046/j.1523-1739.1992.610101.x
Koné, I., Lambert, J. E., Refisch, J., and Bakayoko, A. (2008). Primate seed dispersal and its potential role in maintaining useful tree species in the Taï region, Côte-d’Ivoire: implications for the conservation of forest fragments. Trop. Conserv. Sci. 1, 293–306.
Kristensen, M., and Balslev, H. (2003). Perceptions, use and availability of woody plants among the Gourounsi in Burkina Faso. Biodiv. Conserv. 12, 1715–1739.
Kuroda, S., Nishihara, T., Suzuki, S., and Oko, R. A. (1996). “Sympatric chimpanzees and gorillas in the Ndoki Forest, Congo,” in Great ape societies, eds W. C. McGrew, L. F. Marchant, and T. Nishida (Cambridge: Cambridge University Press), 71–81.
Loudon, J. E., Sandberg, P. A., Wrangham, R. W., Fahey, B., and Sponheimer, M. (2016). The stable isotope ecology of Pan in Uganda and beyond. Am. J. Primatol. 78, 1070–1085. doi: 10.1002/ajp.22552
Malaisse, F. (1997). Se Nourrir en Forêt Claire Africaine: Approche Écologique et Nutritionnelle. Gembloux: Les Presses Agronomic de Gembloux.
Matsuzawa, T., Humle, T., and Sugiyama, Y. (2011). The Chimpanzees of Bossou and Nimba. Tokyo: Springer.
McGrew, W. C., Marchant, L. F., and Phillips, C. A. (2009). Standardised protocol for primate faecal analysis. Primates 50, 363–366. doi: 10.1007/s10329-009-0148-z
McLennan, M. R. (2013). Diet and feeding ecology of chimpanzees (Pan troglodytes) in Bulindi, Uganda: foraging strategies at the forest–farm interface. Int. J. Primatol. 34, 585–614. doi: 10.1007/s10764-013-9683-y
McLennan, M. R., and Hockings, K. J. (2016). “The aggressive apes? Causes and contexts of great ape attacks on local persons,” in Problematic Wildlife: A Cross-Disciplinary Approach, ed. F. M. Angelici (Switzerland: Springer), 373–394. doi: 10.1007/978-3-319-22246-2_18
McLennan, M. R., Spagnoletti, N., and Hockings, K. J. (2017). The implications of primate behavioural flexibility for sustainable human-primate coexistence in anthropogenic habitats. Int. J. Primatol. 38, 105–121. doi: 10.1007/s10764-017-9962-0
Nishida, T., and Uehara, S. (1983). Natural diet of chimpanzees (Pan troglodytes schweinfurthii): long-term record from the Mahale Mountains, Tanzania. Afr. Study Monogr. 3, 109–130.
Nyhus, P. J. (2016). Human–wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 41, 143–171.
Parathian, H., McLennan, M. R., Hill, C. M., Frazão-Moreira, A., and Hockings, K. J. (2018). Breaking through disciplinary barriers: Human-wildlife interactions and multispecies ethnography. Int. J. Primatol. 39, 749–775. doi: 10.1007/s10764-018-0027-9
Phillips, O., and Gentry, A. H. (1993). The useful plants of Tambopata, Peru: I. statistical hypotheses tests with a new quantitative technique. Econ. Bot. 47, 15–32. doi: 10.1007/bf02862203
Redpath, S. M., Young, J., Evely, A., Adams, W. M., Sutherland, W. J., Whitehouse, A., et al. (2013). Understanding and managing conservation conflicts. Trends Ecol. Evol. 28, 100–109. doi: 10.1016/j.tree.2012.08.021
Reynolds, V., Lloyd, A., and English, C. (2012). Adaptation by Budongo Forest chimpanzees (Pan troglodytes schweinfurthii) to loss of a primary source of dietary sodium. Afr. Primates 7, 156–162.
Riley, E. P. (2007). The human–macaque interface: conservation implications of current and future overlap and conflict in Lore Lindu National park. Sulawesi, Indonesia. Am. Anthropol. 109, 473–484. doi: 10.1525/aa.2007.109.3.473
Serckx, A., Huynen, M.-C., Bastin, J.-F., Hambuckers, A., Beudels-Jamar, R. C., Vimond, M., et al. (2014). Nest grouping patterns of bonobos (Pan paniscus) in relation to fruit availability in a forest-savannah mosaic. PLoS One 9:e93742. doi: 10.1371/journal.pone.0093742
Sheil, D., Ducey, M., Ssali, F., Mukasa, N. J., van Heist, M., Ezuma, P., et al. (2012). Bamboo for people, Mountain gorillas, and golden monkeys: Evaluating harvest and conservation trade-offs and synergies in the Virunga Volcanoes. For. Ecol. Manag. 267, 163–171. doi: 10.1016/j.foreco.2011.11.045
Sousa, C., and Frazão-Moreira, A. (2010). “Etnoprimatologia ao serviço da conservação na Guiné-Bissau: o chimpanzé como exemplo,” in Etnoecologia em Perspectiva: Natureza, Cultura e Conservação, eds A. Alves, F. Souto, and N. Peroni (Recife: NUPEEA), 189–200.
Sousa, J., Vicente, L., Gippoliti, S., Casanova, C., and Sousa, C. (2014). Local knowledge and perceptions of chimpanzees in Cantanhez National Park. Guinea-Bissau. Am. J. Primatol. 76, 122–134. doi: 10.1002/ajp.22215
Sugiyama, Y., and Koman, J. (1992). The Flora of Bossou: its utilization by chimpanzees and humans. Afr. Study Monogr. 13, 127–169.
Temudo, M. P. (2009). A narrativa da degradação ambiental no Sul da Guiné-Bissau: uma desconstrução etnográfica. Etnográfica 13, 237–264. doi: 10.4000/etnografica.1341
Temudo, M. P., and Abrantes, M. (2014). The cashew frontier in guinea-bissau, West Africa: changing landscapes and livelihoods. Hum. Ecol. 42, 217–230. doi: 10.1007/s10745-014-9641-0
Torres, J., Brito, J. C., Vasconcelos, M. J., Catarino, L., Goncalves, J., and Honrado, J. (2010). Ensemble models of habitat suitability relate chimpanzee (Pan troglodytes) conservation to forest and landscape dynamics in Western Africa. Biol. Conserv. 143, 416–425. doi: 10.1016/j.biocon.2009.11.007
Tutin, C. E., Ham, R. M., White, L. J. T., and Harrison, M. J. S. (1997). The primate community of the Lopé reserve, Gabon: diets, responses to fruit scarcity, and effects on biomass. Am. J. Primatol. 42, 1–24. doi: 10.1002/(sici)1098-2345(1997)42:1<1::aid-ajp1>3.0.co;2-0
Tutin, C. E. G., and Fernandez, M. (1993). Faecal analysis as a method of describing diets of apes: examples from sympatric gorillas and chimpanzees at Lopé. Gabon. Trop. 2, 189–197. doi: 10.3759/tropics.2.189
Waller, M., and Pruetz, J. (2016). “Competition between chimpanzees and humans: The effects of harvesting non-timber forest products,” in Ethnoprimatology: Primate Conservation in the 21st Century, ed. M. Waller (Springer: Switzerland), 157–177. doi: 10.1007/978-3-319-30469-4_9
Winterhalder, B. (2001). “The behavioural ecology of hunter-gatherers,” in Hunter-Gatherers: An Interdisciplinary Perspective, eds C. Panter-Brick and R. H. Layton (Cambridge: Cambridge University Press), 12–38.
Woodroffe, R., Thirgood, S., and Rabinowitz, A. (2005). People and wildlife: Conflict or coexistence?. Cambridge: Cambridge University Press.
World Bank (2019). Poverty Headcount Ratio at $1.90 a day (2011 PPP) (% of population). Avaliable at: https://data.worldbank.org/indicator/SI.POV.DDAY.
Keywords: human-wildlife conflict, social-ecological systems, wild resource use, great ape, inter-disciplinary science, local ecological knowledge, shared landscapes, large mammal conservation
Citation: Hockings KJ, Parathian H, Bessa J and Frazão-Moreira A (2020) Extensive Overlap in the Selection of Wild Fruits by Chimpanzees and Humans: Implications for the Management of Complex Social-Ecological Systems. Front. Ecol. Evol. 8:123. doi: 10.3389/fevo.2020.00123
Received: 14 November 2019; Accepted: 16 April 2020;
Published: 19 May 2020.
Edited by:
J. Guy Castley, Griffith University, AustraliaReviewed by:
Giuseppe Donati, Oxford Brookes University, United KingdomDivya Karnad, Ashoka University, India
Copyright © 2020 Hockings, Parathian, Bessa and Frazão-Moreira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kimberley J. Hockings, ay5ob2NraW5nc0BleGV0ZXIuYWMudWs=