
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 17 March 2020
Sec. Behavioral and Evolutionary Ecology
Volume 8 - 2020 | https://doi.org/10.3389/fevo.2020.00052
This article is part of the Research Topic Flexibility in the Migration Strategies of Animals View all 31 articles
 Phil F. Battley1*
Phil F. Battley1* Jesse R. Conklin2
Jesse R. Conklin2 Ángela M. Parody-Merino1
Ángela M. Parody-Merino1 Peter A. Langlands3Ian Southey4Thomas Burns3
Peter A. Langlands3Ian Southey4Thomas Burns3 David S. Melville5
David S. Melville5 Rob Schuckard6Adrian C. Riegen7
Rob Schuckard6Adrian C. Riegen7 Murray A. Potter1
Murray A. Potter1While avian migration timing is clearly influenced by both breeding and non-breeding geography, it is challenging to identify the relative and interdependent roles of endogenous programs, early-life experience, and carry-over effects in the development of adult annual schedules. Bar-tailed godwits Limosa lapponica baueri migrate northward from New Zealand toward Asian stopover sites during the boreal spring, with differences in timing between individuals known to relate to their eventual breeding-ground geography in Alaska. Here, we studied the timing of northward migration of individual godwits at three sites spanning 1,100 km of New Zealand’s 1,400-km length. A lack of morphological or genetic structure among sites indicates that the Alaskan breeding population mixes freely across all sites, and larger birds (southern breeders) tended to migrate earlier than smaller birds (northern breeders) at all sites. However, we unexpectedly found that migration timing varied between the sites, with birds from southern New Zealand departing on average 9.4–11 days earlier than birds from more northerly sites, a difference consistent across 4 years of monitoring. There is no obvious adaptive reason for migration timing differences of this magnitude, and it is likely that geographic variation in timing within New Zealand represents a direct response to latitudinal variation in photoperiod. Using resightings of marked birds, we show that immature godwits explore widely around New Zealand before embarking on their first northward migration at age 2–4 years. Thus, the process by which individual migration dates are established appears to involve: (1) settlement by sub-adult godwits at non-breeding sites, to which they are highly faithful as adults; (2) a consequent response to environmental cues (i.e., photoperiod) that sets the local population’s migration window; and (3) endogenous mechanisms, driven by breeding geography, that establish and maintain the well-documented consistent differences between individuals. This implies that behavioral decisions by young godwits have long-lasting impacts on adult annual-cycle schedules, but the factors guiding non-breeding settlement are currently unknown.
In birds, breeding ground geography, or more specifically geographic variation in breeding phenology, can be a major determinant of migration timing (e.g., Both, 2010; Conklin et al., 2010; Emmenegger et al., 2014; Briedis et al., 2016; Ouwehand et al., 2016), and associated processes of molt (Conklin and Battley, 2011a) and migratory fueling (Fry et al., 1972; Scheiffarth et al., 2002). This likely occurs through a combination of inheritance (genetic and/or parental effects) and response to early-life conditions (Ciarleglio et al., 2010), resulting in individuals showing natal site-fidelity (at least at a regional scale) and having a circannual program that enables timely arrival for breeding. Although migration timing can be very consistent within individuals (reviewed in Both et al., 2016), it is also subject to annual variation based on environmental conditions (Duriez et al., 2009; Conklin and Battley, 2011b), and can be modified over time through social information and individual improvement (Mueller et al., 2013; Sergio et al., 2014).
Additionally, migration timing can vary by non-breeding site, particularly in populations with a large non-breeding range (e.g., Myers et al., 1985; Piersma et al., 2005; van Bemmelen et al., 2019). This is to be expected, as populations traveling farther ought to start migrating earlier if they require longer to reach the breeding grounds. Less clear are the mechanisms that generate such population-level differences in phenology. Migration timing in birds is thought to be controlled by an internal circannual clock that is entrained by photoperiod (Gwinner, 1996a). Population-level comparisons indicate that differences in schedules can result from differences in the underlying circannual cycles and their responses to photoperiod (Helm et al., 2009), so the timing of migration of individuals may reflect both inherited circannual cycles and the photoperiodic environment the birds experience (Helm and Gwinner, 2005; Bojarinova and Babushkina, 2015). Hence, differences in timing could simply reflect photoperiod cues that vary geographically, or they could also arise through local environmental conditions (Dawson, 2008) or differences in migration strategy (Alerstam and Lindström, 1990). This means that annual-cycle schedules are not simply a product of the natal site, but can be modified by experience and both biotic and abiotic conditions after the first southbound migration.
In many species, adults show extremely high fidelity to non-breeding sites (e.g., Lourenço et al., 2016), but we generally know little about how these sites are chosen. Non-breeding settlement may occur non-randomly, through ecological selection for certain aspects of phenotype (e.g., size, feeding morphology; Myers, 1981; Nebel, 2005) or competitive occupation of high-quality sites (Gunnarsson et al., 2005; Studds and Marra, 2005), but a large element of chance may determine where juveniles end up at the end of their first southward migration (Thorup et al., 2003; Cresswell, 2014). In short-lived species that migrate to breed in their first year of life, it may then be difficult to differentiate among endogenous programs, early-life experience, and potentially temporary carry-over effects of natal or migratory conditions (Senner et al., 2015) in the development of life-long adult migration timing patterns.
By contrast, many migratory species show delayed maturity and do not migrate to the breeding grounds for one or more years. During these immature years, birds may be highly mobile and ‘sample’ potential non-breeding sites before settling at a site to which they remain faithful as adults (Battley et al., 2011). Thus, individuals in such species have potentially several years in which to make settlement decisions that may affect their subsequent life-long migration schedules. For these species, the window in which information relevant to settlement decisions and migration timing is assimilated may be prolonged. Examination of behavior in this ‘pre-migratory’ phase of life may shed light on how routines as adults are established, with implications for site choice and timing of migration.
Bar-tailed godwits Limosa lapponica baueri provide a clear example of the relative individual timing of migration being predominantly ‘set’ by breeding ground geography on the other side of the world. Conklin et al. (2010) showed that bar-tailed godwits from a single non-breeding site on the North Island of New Zealand bred across the entire Alaskan range, from the Yukon-Kuskokwim Delta in the south to the North Slope in the north. As there is a difference of about 3 weeks in the timing of the spring thaw across that range, breeding opportunities arise much earlier for southern breeders than for northern breeders. This difference in optimal arrival date was reflected in the timing of migration of individuals across the entire northward migration, with southern breeders migrating earlier than northern breeders both from New Zealand and after a 4- to 6-week stopover in Asia (Conklin et al., 2010; Battley et al., 2012). Additionally, as godwit size also varies across Alaska (larger in the south, smaller in the north), larger birds in New Zealand tend to embark on northward migration earlier than smaller birds (Conklin et al., 2011). Monitoring of departures of marked birds, and repeat tracking of individuals by geolocators, showed that individual godwits were highly consistent in their timing of initiation and later stages of migration (Battley, 2006; Conklin et al., 2013). What we know about migration timing in baueri, however, comes almost exclusively from latitudes 37–41°S in New Zealand, while the non-breeding range extends from 34.5°S to 46.5°S in New Zealand, and extends much further north into the Tropics in eastern Australia. If migration timing varies by non-breeding latitude, then the juvenile settlement period might have important impacts on annual-cycle schedules, with early-life decisions modifying or over-riding endogenous programs derived from natal areas.
Here, we document variation in northward migration timing of bar-tailed godwits among three sites covering 1,100 km of the non-breeding range in New Zealand. Given that regional differences in migration timing could arise through population structure on the non-breeding grounds (i.e., differential settlement of southern- versus northern-breeders, which show some genetic differentiation: Parody-Merino, 2018; J. R. Conklin, unpublished data), we also test for population structure via biometrics (culmen length as a size measure) and neutral genetic variation (microsatellites). Then, we explore the pre-migratory settlement period of sub-adult godwits using resightings of marked birds to describe how extensively immature birds range around New Zealand and at what age they first migrate north. We discuss the relevance of our findings for understanding the role of early-life experience and the interaction of breeding and non-breeding geography for the development of individual annual schedules.
We studied migration timing of bar-tailed godwits in detail at three sites across New Zealand—the Firth of Thames near Auckland in the northern North Island (37.17°S, 175.32°E), Manawatu River Estuary in the southern North Island (40.47°S, 175.22°E) and the estuary of the Owaka River in the southern South Island (46.48°S, 169.70°E)—we refer to these as Auckland, Manawatu, and Otago hereafter (Figure 1). The distances between sites are approximately 365 km (3.3° latitude) between Auckland and Manawatu, and 800 km (6.0° latitude) between Manawatu and Otago. Godwits were caught by cannon-net or mist-net, aged on the basis of plumage characteristics (age 1 = juvenile, 2 or 3 = immature non-migrant, or 3+ = adult), measured (exposed culmen, mm) and weighed, and individually marked with either color-bands or a leg-flag engraved with a unique three-letter code. Juveniles were aged based on retained juvenile plumage. Age 2 and 3 birds were aged by a combination of features: presence during the boreal breeding season, stage of primary molt (starting during the late austral winter, so are more advanced than adults in the spring), retained juvenile outer primary feathers or greater primary coverts for age 2, presence of breeding plumage (suggestive of age 3) and relative primary feather wear [including the presence of replaced (unworn) primaries]. Adults could be distinguished by primary feather wear, extent of breeding plumage and later primary molt than younger birds. As aging of year 2 and 3 birds can be difficult, we group them here as immatures. Birds were sexed by culmen length (males = 70–99 mm, females 89–130 mm; Conklin et al., 2011), but ca. 10% of birds cannot be sexed by this method, due to overlapping ranges; for large males and small females, sex was confirmed by the extent of breeding plumage before departure (Conklin and Battley, 2011a). Birds were caught from 2004 onward in Auckland, 2006 onward in Manawatu and 2009 onward in Otago. As part of a wider study of movements of northern hemisphere shorebirds in New Zealand (Battley et al., 2011), we also banded godwits at a number of other sites spanning the length of New Zealand. Resightings of these birds have been compiled and we used this larger dataset to explore movements of sub-adult godwits.
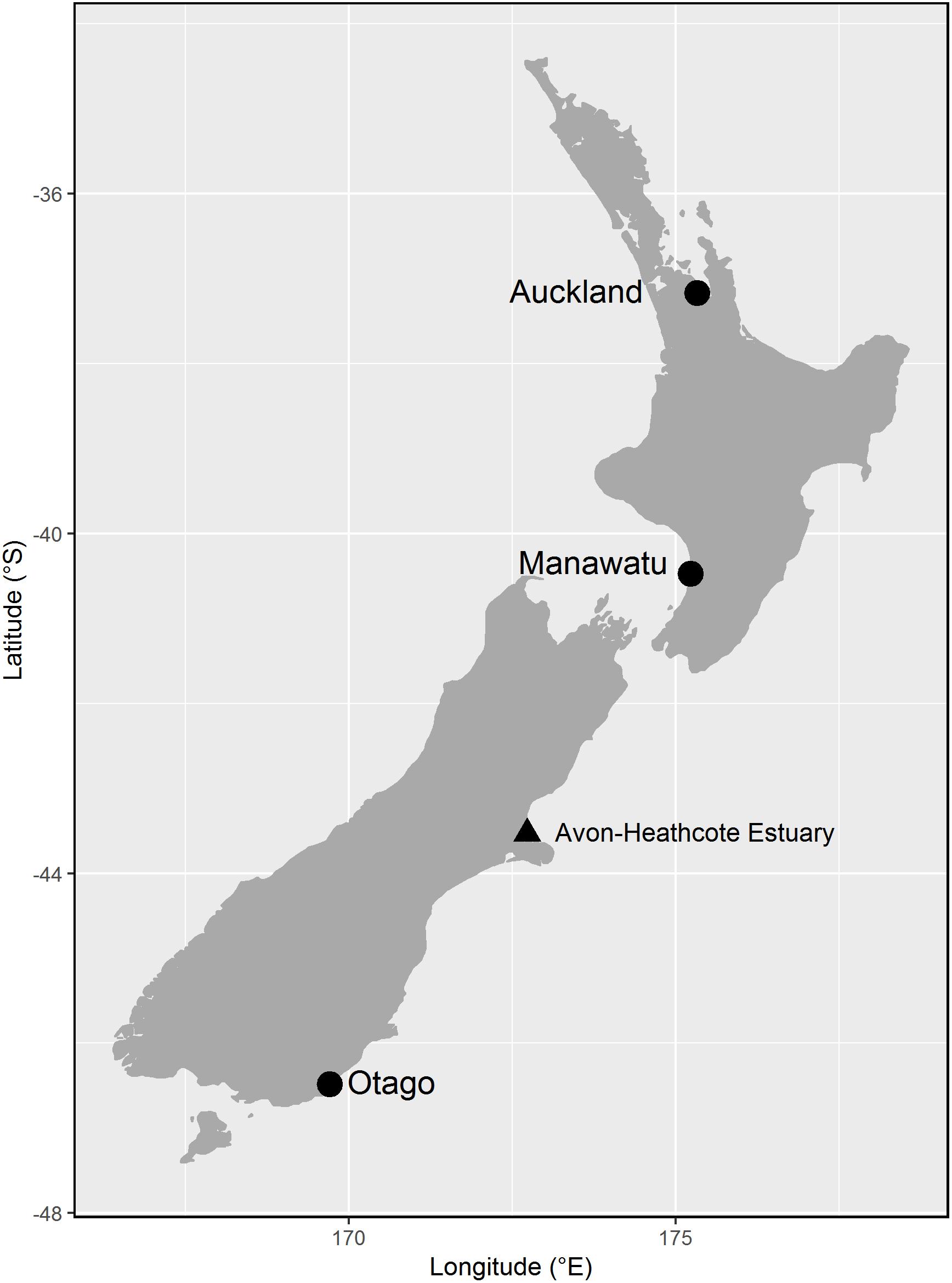
Figure 1. Map of New Zealand showing the main study sites (circles), and another site where counts during the departure period were available from one year (triangle). Distances between the study sites are approximately 365 km between Auckland and the Manawatu Estuary and 800 km between the Manawatu and Otago sites.
There is a latitudinal cline in godwit body size across Alaska (northern = smaller; Conklin et al., 2011) and evidence of slight genetic structure in the breeding range (Fst = 0.013 between northern and southern breeders, based on microsatellites; Parody-Merino, 2018) that could also be present in the non-breeding range. We looked for evidence of population structure among non-breeding sites using both morphometrics and neutral genetic markers. For biometric comparisons we also used data from birds caught at other sites around the Auckland region (Manukau and Kaipara Harbours) and Otago–Southland (Warrington, Otago, and Awarua Bay and Invercargill Estuary, Southland; see Battley et al., 2011 for site details).
A subset of birds was blood-sampled at the time of capture (95 in Auckland, 109 in Manawatu, and 19 in Otago) for genetic analyses. We genotyped 223 godwits at 27 microsatellite loci (full methodological details are provided in the Supplementary Material). For comparison with structure detected within the Alaska breeding range using the same microsatellite loci [Fst = 0.013 between northern (>65°N) and southern (<65°N) breeders; Parody-Merino, 2018], we calculated pairwise Fst among the three non-breeding sites using Arlequin v.3.5.2.2 (Excoffier and Lischer, 2010). To further explore potential non-breeding structure, we used the PRIORLOC function in STRUCTURE v.2.3.4 (Hubisz et al., 2009) to test whether birds from the three study sites formed distinguishable genetic clusters. The PRIORLOC function uses an individual’s non-breeding location in New Zealand (Auckland, Manawatu, or Otago) to estimate the most likely number of clusters in the population. K-means clustering Bayesian information criterion (BIC) indicated a single population (K = 1) or two clusters. To further explore potential subtle structure by study site, we ran STRUCTURE with PRIORLOC again with an assumption of K = 3. STRUCTURE was run with the following parameters: length of burn-in period = 1,000,000; MCMC runs = 500,000; number of iterations per run = 15. Results were visualized using Genesis v.0.2.5 (Buchmann and Hazelhurst, 2015).
At each of the key study sites, we undertook monitoring of migratory departure in late February–early April by visual observation of marked birds, recording the last day of observation or, where possible, confirming the exact day of departure when a bird was seen in a departing flock (Conklin and Battley, 2011b). At the Manawatu site (population ∼200 birds), >80% of individuals were directly observed migrating, and the remaining departures were deduced from intensive daily resighting and flock counts. Geolocator conductivity values confirm that observation-based dates at this site are exact (Battley and Conklin, 2017). In Otago (∼350 birds) migration dates were mostly deduced from daily resighting and flock counts; four dates were derived from geolocators. The number of daily records of birds ranged from 1–24 with a mean of 7.0. Low values were associated with very early migrants. In Auckland (population 3,000+), the last date of observation for individuals seen repeatedly during the observation period was taken as the migration date, although seven birds were visually confirmed departing. We restricted records to those with six or more resightings unless a departure was observed, or a record with <6 resightings was later than records in other years for that individual or was corroborated by the dates in other years. The number of daily records of birds ranged from 1 to 26 with a mean of 9.8. The distributions of resighting frequencies for Auckland and Otago are given in the Supplementary Material.
There are some subtle biases in the determination of migration timing at the three sites. Very early departures in Auckland and Otago are likely to be overlooked, as repeated sightings are necessary to evaluate a bird’s likelihood of being resighted and some birds seen only once soon after fieldwork started in Otago were not included as February departures. Last dates of observation for Auckland birds usually represent minimum estimates of departure date (as birds could not be confirmed as being absent in a large population), and the true departure dates for many birds will be later than assumed. Auckland birds might migrate on average slightly later than documented, but our dataset may under-represent the early-departing sector of the population. Intensive departure monitoring took place at Auckland in 2014–2016, at Manawatu from 2008–2017 and in Otago in 2013–2016, but we conducted analyses of migration timing on the period of greatest overlap in the datasets, 2013–2016 (4 years for Manawatu and Otago; 3 years for Auckland). This resulted in a sample of 409 birds for which we had a migration date in one or more years (range 1–4 years); for birds with multiple years of data, we used an individual’s mean migration date for analysis.
We compared migration phenology between sites using ANOVA with sex and site as factors, followed by a Tukey test for differences between levels of any significant factors. To test whether the relationship between size (culmen length) and migration date was consistent across all sites, we ran a linear model of migration date with bill nested within sex within site; this tests whether departure dates of birds within each sex varied by size, allowing for differences in migration timing between sites. Trends for each site (each sex separately) were compared via their slope estimates and 95% confidence intervals. The size-structures of the populations at the study regions were compared in ANOVAs with site as a factor but with sexes analyzed separately, with Tukey tests for differences between sites.
A subset of godwits from Manawatu and Otago were also tracked with light-level geolocators in 2013 and 2014. We retrieved 27 loggers (23 Migrate Technology Intigeo-CK65K and four Biotrack MK4093) from Manawatu birds and four from Otago (one Migrate Technology and three Biotrack). Loggers of one Manawatu and two Otago birds recorded data only as far as Asia. While there can be uncertainty about precise migration timing derived from analysis of light data, in shorebirds such as godwits, wetting of the logger during foraging or bathing means that extended dry periods clearly delineate non-stop migratory flights. This pattern is easy to identify in the conductivity data and these have been shown in godwits to give exact correspondence with observed migration departure dates (Battley and Conklin, 2017). We therefore used conductivity data to determine the departure date from New Zealand, duration of flight to Asia, length of the subsequent staging period in Asia, and the migration date toward the Alaskan breeding grounds, and compared these measures between Manawatu and Otago birds. Positional data were analyzed using Geolight (Lisovski and Hahn, 2012) and confirmed that all birds had their stopovers in the Yellow Sea region of eastern Asia. The Migrate Technology loggers also recorded min/max temperatures across each 4-h block of recording, which we use to evaluate the relative climate before and after the flight to Asia.
As the distribution of godwits around New Zealand reflects the non-breeding settlement decisions of young birds, we used our database of resightings of individually marked godwits to identify how extensively young birds explore New Zealand and over how long a period. Of 2,020 total godwits marked during 1993–2018, 327 were aged as 1–3 years when captured, including birds that could not be aged precisely but could be confirmed by a combination of date, plumage and wing molt not to be migratory adults (i.e., 3+ years old). To detect long-distance movements by sub-adults, we extracted all records of these immature birds that were seen away from their banding region (see Battley et al., 2011 for region details) before adulthood (≤3 years old). We summarized by region how many immature birds banded elsewhere had been recorded in that region. A similar analysis of adult resightings confirmed that they have virtually complete non-breeding site-fidelity between years (Battley et al., 2011).
We also use resightings to identify the age of first migration and to evaluate the period (age in months) over which young godwits were potentially still exploring New Zealand. For this we restricted the analysis to 215 individuals confirmed as age 1 (juvenile; n = 159) or age 2 (n = 56) when captured (on the basis of retained juvenile plumage). To estimate the ages of first northward migration we used records of these known-age birds either overseas (Asia or Alaska, demonstrating migration), or in New Zealand during the boreal breeding season (demonstrating non-migration). For these 215 known-age birds, we also looked at the age (in months, assuming hatch in June) of the last known long-distance (between-region) movement either southward or northward within New Zealand in the first 36 months of life, as most birds migrate by that time (see the section “Results”). For this, we recorded the bird’s age at the last known resighting of the bird before a subsequent record in a new region; this represents the minimum age at the time of the move, and may underestimate the age by weeks or months. We excluded records of birds from the migration departure and arrival periods (March–April and August–October) to exclude movements that may reflect migratory stopovers rather than true non-breeding location shifts.
There was little evidence of population structure across New Zealand. Comparisons of the size distributions between sites found only subtle differences between sites (males, ANOVA F2,660 = 8.651, P < 0.001; females, F2,772 = 4.218, P < 0.02; Figure 2, upper panels). The range of sizes was similar across sites for both sexes (Table 1), but Manawatu males were significantly smaller on average than in both Auckland and Otago (Tukey post hoc test, P < 0.001) and Otago females were slightly larger than in Auckland (Tukey post hoc test, P < 0.01; Table 1).
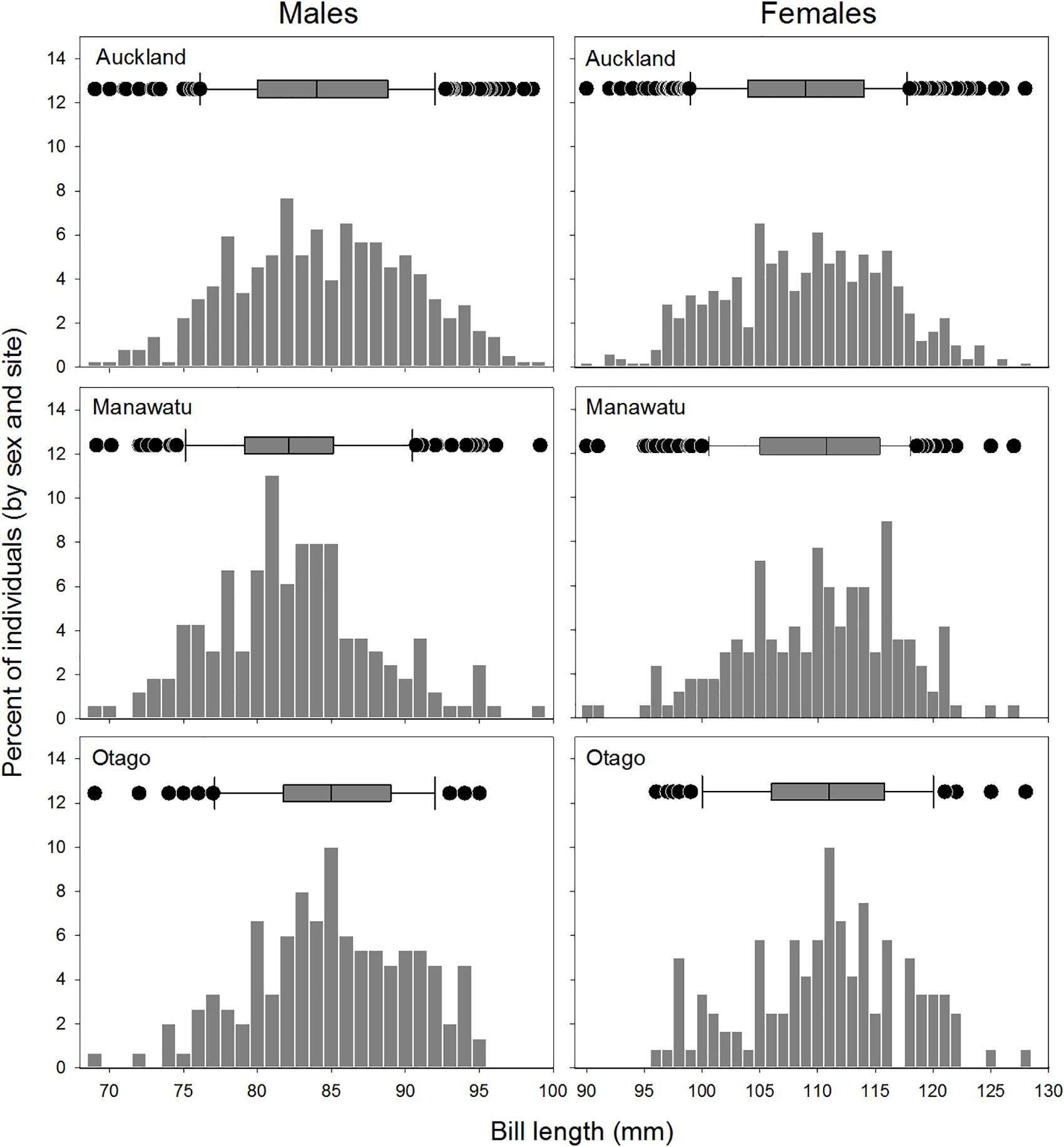
Figure 2. Size-distribution of bar-tailed godwits at the Auckland, Manawatu, and Otago sites, plotted for females and males separately. Samples sizes are (male/female): Auckland 350/487; Manawatu 163/168; Southland 150/120. Boxplots across the tops of each plot summarize the distribution of values in each group (boxes show the median and 25th to 75th percentiles, whiskers the 5th and 95th percentiles, and points show outliers).

Table 1. Summary of bar-tailed godwit bill lengths (mm, mean ± SD, range and n) by sex and region.
Analyses of neutral genetic variation in microsatellites revealed no evidence for population genetic structure among non-breeding sites. All values of among-site pairwise Fst were indistinguishable from zero: Otago vs. Manawatu, Fst = −0.0001, P = 0.50; Otago vs. Auckland, Fst = 0.0011, P = 0.41; Manawatu vs. Auckland, Fst = −0.0004, P = 0.62. Moreover, the STRUCTURE analysis indicated uniform representation of assumed genetic clusters among sites (see Supplementary Material).
Godwits departed New Zealand from late February to late March, but the timing of migration varied by site (ANOVA, F2,405 = 202.9, P < 0.001). There was a small difference between the two North Island sites (1.7 days; Tukey post hoc test, adjusted P < 0.05) but large differences between the South Island site (Otago) and both North Island sites (Otago departures being 9.3 days earlier than Manawatu and 10.9 days earlier than Auckland; Tukey post hoc test, adjusted P < 0.001 for both; Figure 3 and Table 2). There was a slight difference in the timing of migration of males and females (males were 1.1 days earlier on average; ANOVA, F1,405 = 5.343, Tukey post hoc test, adjusted P < 0.05). At each site, larger birds within each sex tended to depart earlier than small birds (Figure 4); these trends were statistically significant for all slopes (Table 3). The slopes of size vs. departure date were steepest at Manawatu, being significantly so compared to Auckland and Otago for males and compared to Otago for females, based on non-overlapping 95% confidence intervals of slope estimates.
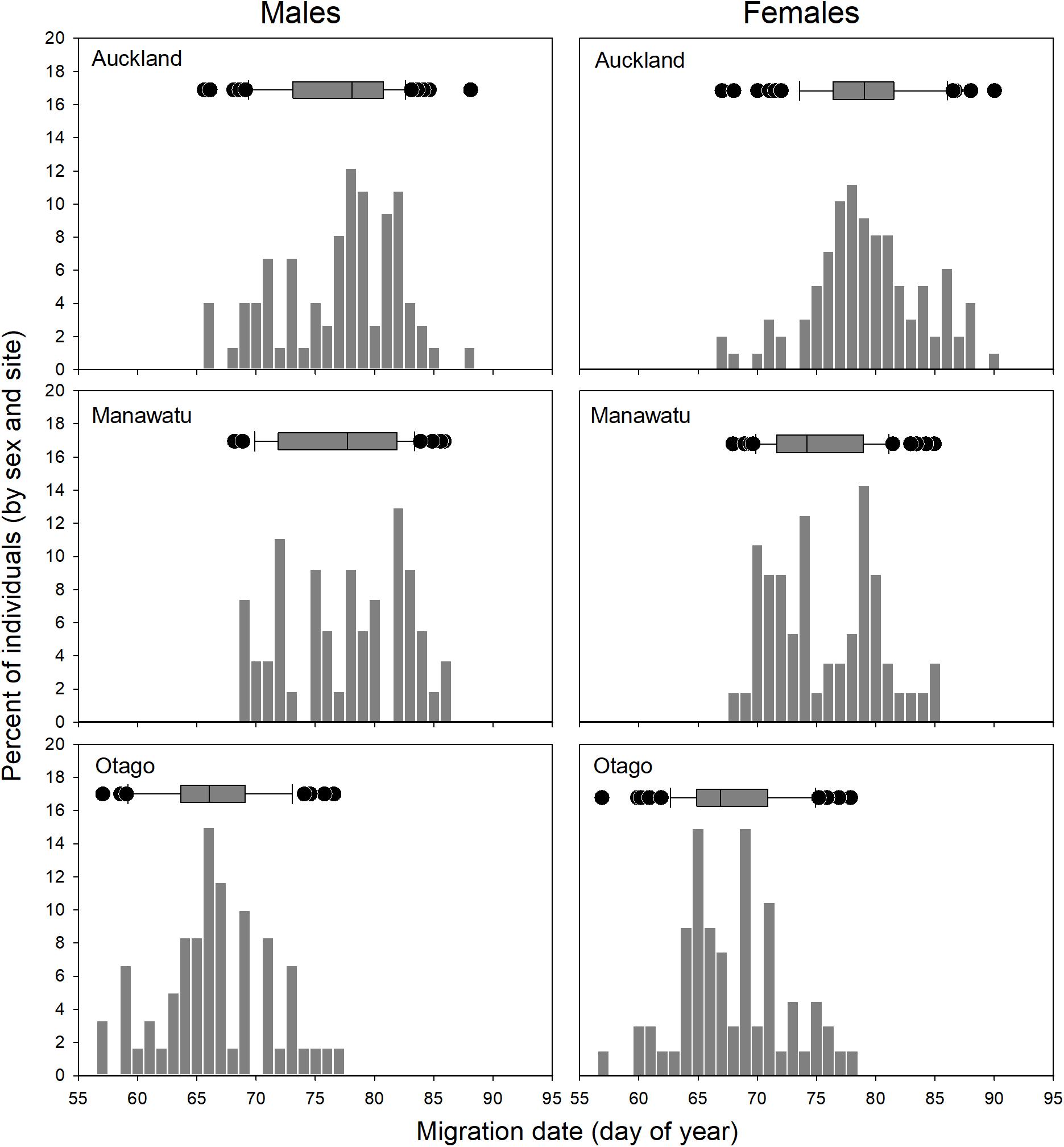
Figure 3. Frequency distributions of migration dates of bar-tailed godwits from three sites in New Zealand. Values represent individual mean departure dates of marked birds (1–4 years per individual) during 2013–2016.

Table 2. Migration dates of bar-tailed godwits from three sites in New Zealand.
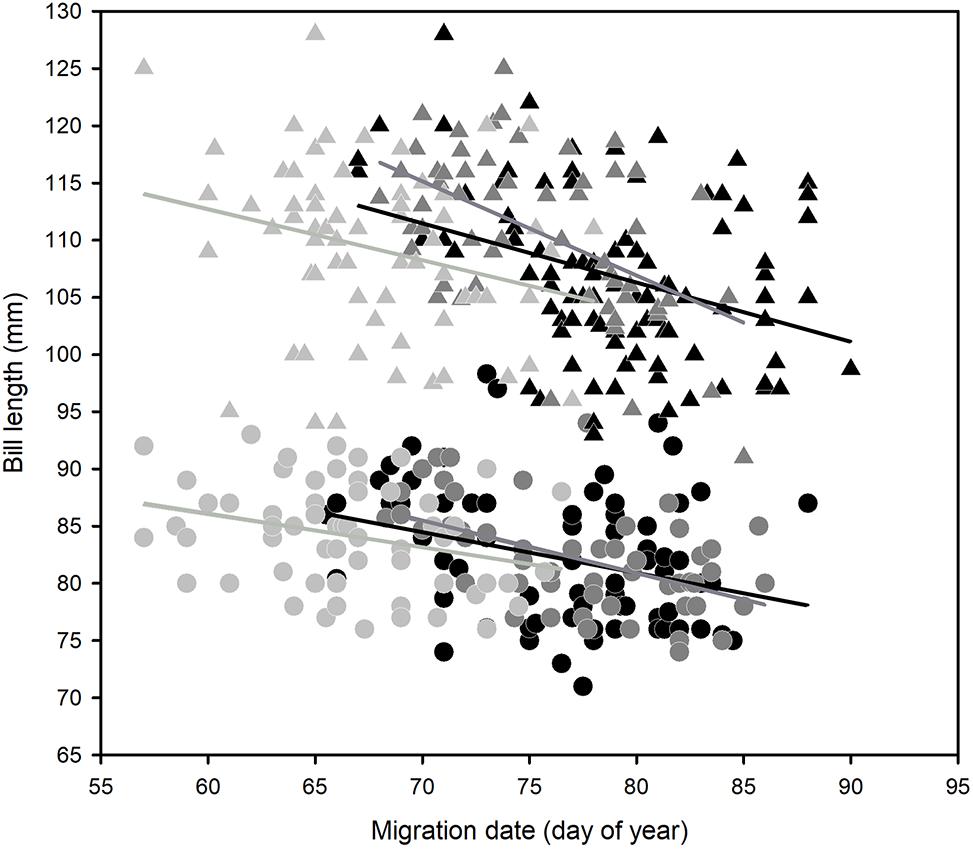
Figure 4. Relationships between body size (represented by bill length) and migration date in bar-tailed godwits in New Zealand. The sexes are shown with different symbols: males (smaller) are the circles, while females (larger) are the triangles. The symbol shading represents the three study sites: black = Auckland, dark gray = Manawatu, and light gray = Otago. Trends are shown by the fitted lines (from a linear regression of size nested within sex within site).

Table 3. Slopes of the relationships between northward migration date and bill length for bar-tailed godwits in New Zealand.
The difference in migration timing inferred from the monitoring of marked individuals is corroborated by flock counts at the two smaller sites where numbers could be monitored closely (Manawatu and Otago; Figure 5). Across all 4 years of study, godwit numbers at Otago dropped dramatically in early March (on 4–8 March). In contrast, major declines at the Manawatu Estuary occurred only in the second week of March (9–14 March). In 2017, weekly counts were also available for the Avon-Heathcote Estuary in Canterbury (see Figure 1), 400 km NE of the Otago site and 400 km SW of Manawatu (A. C. Crossland, personal communication). The migration phenology matched that of Otago (gray points in Figure 5).
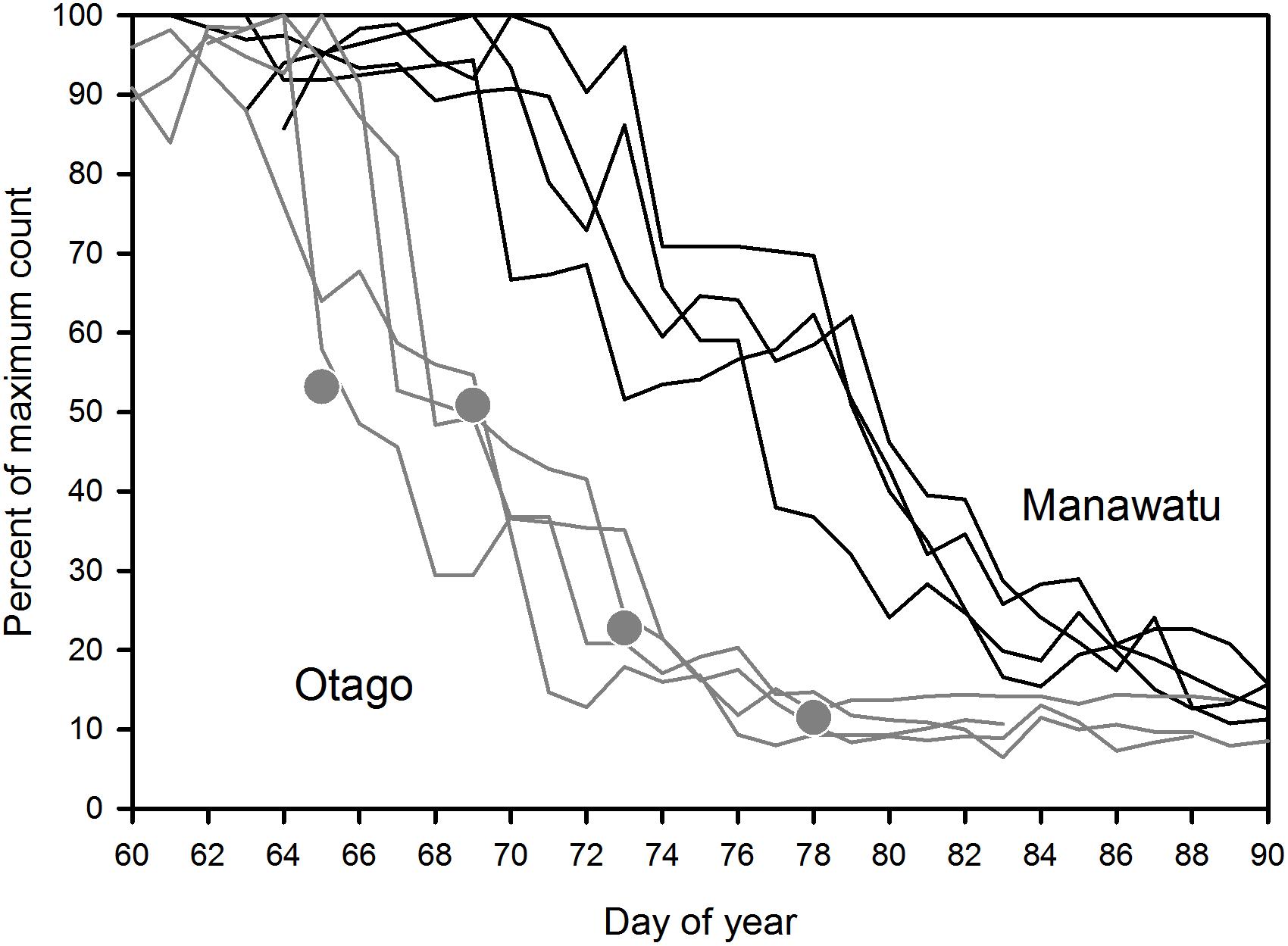
Figure 5. Flock counts through the migratory periods at the Manawatu and Otago sites, 2013–2016. March 1 is represented by day 60 or 61 (the latter in a leap year). Gray circles represent the Avon-Heathcote Estuary, Canterbury, South Island, in 2017.
While, we had only limited geolocator tracking available for the Otago birds, comparisons with birds tracked from the Manawatu in the same years (Figure 6) showed that Otago birds departed from New Zealand earlier and arrived in Asia earlier than Manawatu birds, but departed from Asia around the same time. Specifically, Otago birds left New Zealand 12 days earlier on average (Otago: day 63.5 ± SD 5.0 days, Manawatu: 77.6 ± 4.7 days; t = −5.3, P < 0.01), took a similar time to fly to Asia (8.8 ± 1.0 days vs. 7.9 ± 0.6 days; t = −0.18, n.s.) and arrived in Asia around 13 days earlier (72.3 ± 4.6 days vs. 85.4 ± 4.7 days; t = −5.4, P < 0.01, n = 4 and 27, respectively, for all comparisons). The two Otago birds tracked after staging departed Asia within the same period as Manawatu birds (days 119 and 137 vs. 131.2 ± 9.3 days, range 119–156; Figure 6) and arrived in Alaska within the same period (days 124 and 140 vs. 139.4 ± 8.9, range 121–159 days, n = 2 and 26, respectively, for both comparisons), suggesting similar timetables at this stage of the migration.
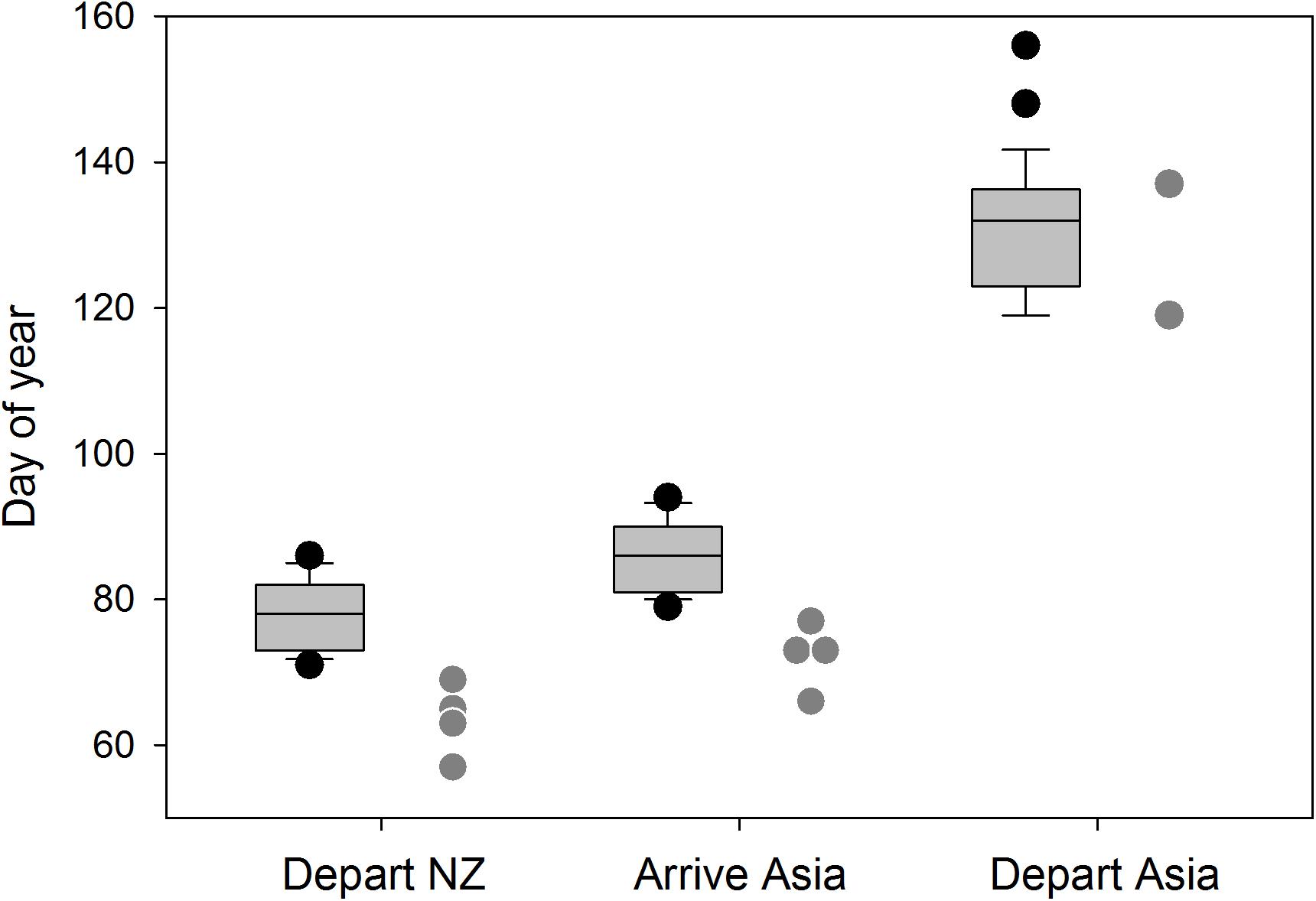
Figure 6. Northward migration timing of geolocator-tracked birds from Manawatu (boxplots) and Otago (gray points) in 2013–2014. Sample sizes are 27 and 4, respectively, for the first two comparisons and 26 and 2 for the third.
Of 327 godwits marked when 1–3 years of age, 113 individuals were recorded making 119 movements between regions (55–1,200 km from the banding site) before adulthood, showing that young birds range widely across New Zealand (Figure 7). This is necessarily an underestimate of movements made by young godwits, as it does not include: (1) movements made prior to initial capture, (2) brief stops missed by observers, and (3) temporary stops or permanent settlement at unsurveyed sites. Of 215 godwits of known age (marked at age 1–2), resightings provided information regarding age of first northward migration for 92 individuals (Figure 8); a combination of resightings unambiguously identified the age of first migration for 24 individuals, and narrowed it down to one of 2 years for an additional 68. A small number of birds migrated north at age 2, but most migrated north for the first time at age 3 or 4 (Figure 8). They therefore have a period of 2–4 years in which to settle in a non-breeding site from which they will subsequently migrate. Some young birds were still moving in their third year of residence in New Zealand, and birds were as likely to move northward within New Zealand as southward over that period (Figure 9). Compared to birds banded as adults, young birds had a much higher rate of being recorded away from the banding region (75 of 193 immatures with resighting histories (38%) versus 173 of 1,208 adults (14%); Fisher exact test, P < 0.001). The adult records include birds caught on migration and birds seen on migration in New Zealand; only 19 adults (1.5%) appear to have relocated outside their banding region (evidenced by multiple consecutive resightings at those sites).
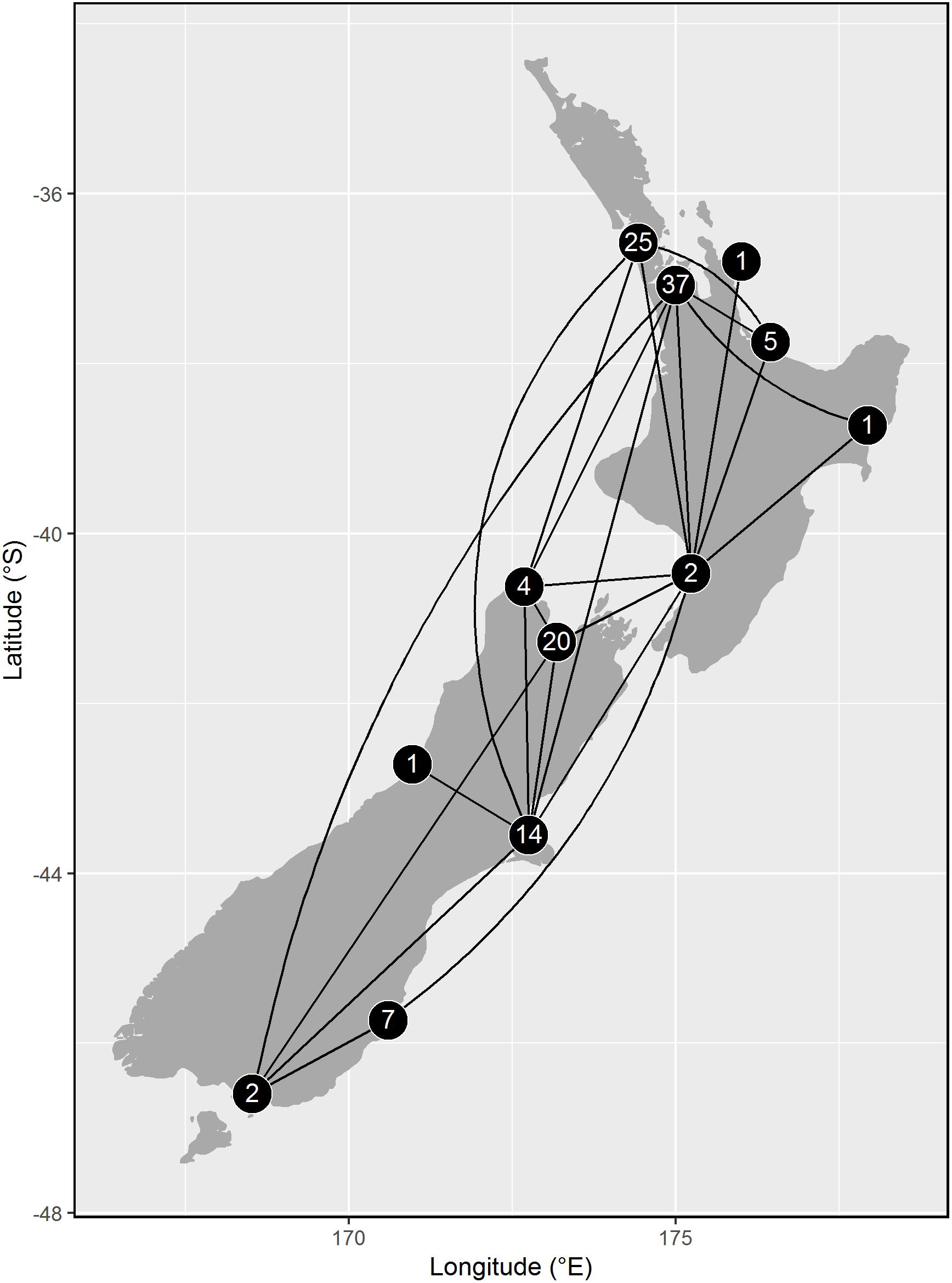
Figure 7. Long-distance movements of bar-tailed godwits caught as juveniles or immatures within New Zealand on the non-breeding grounds in the pre-migration period (age < 3 years). Lines show links between regions confirmed through observations of marked birds; numbers are the number of movements to that region from another (representing 119 movements of 113 birds, out of 327 marked).
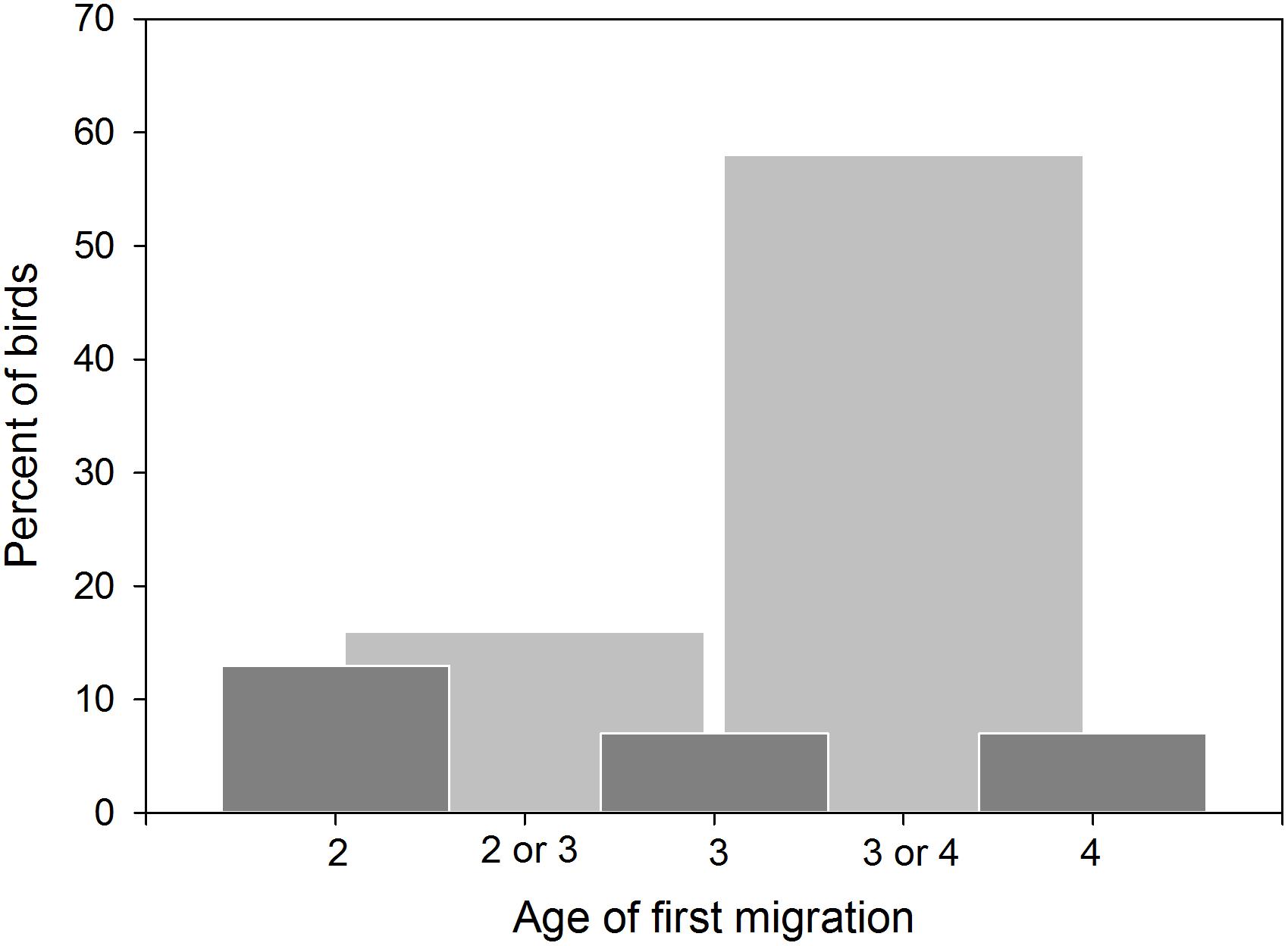
Figure 8. Age of first northward migration for bar-tailed godwits caught aged 1 or 2 in New Zealand. Dark bars represent confirmed ages of migration; pale bars indicate those where the year of migration could not be confirmed to one of the two possible ages. N = 92 birds of 225 marked.
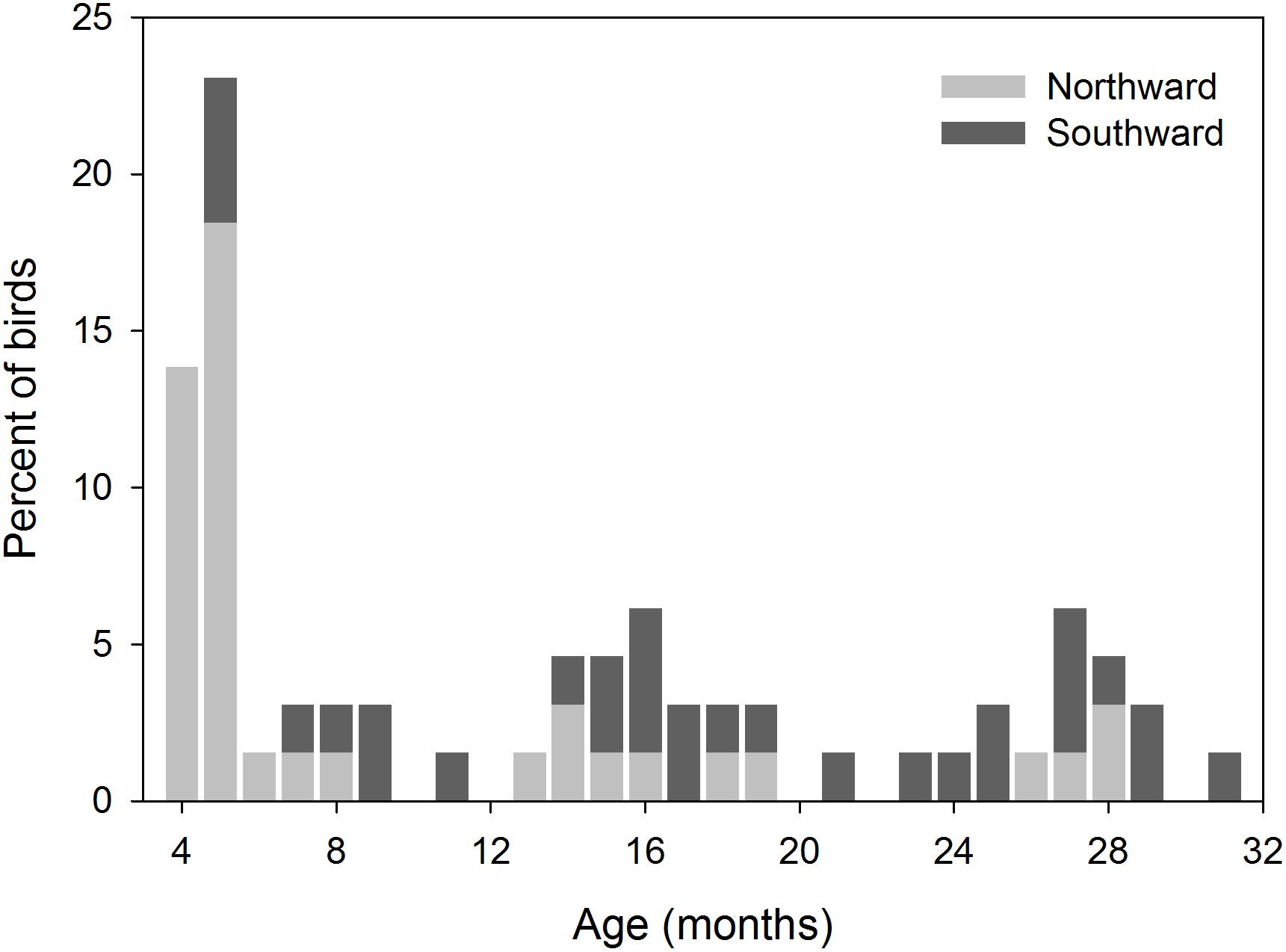
Figure 9. Minimum age at which young bar-tailed godwits (n = 65) made their last regional movements within New Zealand before becoming migratory adults.
We show that the timing of migration of bar-tailed godwits in New Zealand is more complex than realized from earlier studies, in which the only recognized driver of differences in migration timing was geographical variation on the breeding grounds that leads to consistent differences between individual birds within a non-breeding site (Battley, 2006; Conklin et al., 2010; Conklin and Battley, 2011b). We found an unexpected population-level difference in migration timing associated with latitude, with southern New Zealand birds migrating earlier than northern birds, and show that immature birds explore widely around the country before settling at a non-breeding site. This implies that the settlement decisions made by young birds set the ‘window’ within which departures may take place, and thus have life-long consequences for migration timing of individuals of this site-faithful species.
We expanded previous monitoring of bar-tailed godwits departing from the North Island of New Zealand to include birds from the southernmost extent of the non-breeding range, so that our three study sites spanned 1,100 km of the 1,400-km latitudinal ‘length’ of New Zealand. Three lines of reasoning led us to expect that migration schedules would be similar in Otago to elsewhere in New Zealand: (1) similar migration timing had been documented previously at different sites across the northern half of New Zealand (Battley, 1997; Battley, 2006; Conklin et al., 2010; Conklin and Battley, 2011b); (2) biometric analyses indicated little or no population structure in the non-breeding season, with godwits from across the Alaska breeding range mixing freely in New Zealand (Conklin et al., 2011); and (3) the flight lengths to Asia from each of our study sites were relatively similar, so that southern birds do not have appreciably farther to fly than northern birds.
Despite expectations, we found that departures from southern New Zealand were much earlier overall than those from central and northern New Zealand, a pattern that was consistent across all 4 years, and we further detected a small difference between the two North Island sites. Although godwits have a departure span of over 3 weeks at each site, the 9–11 days earlier initiation of migration in Otago meant that in some years half of the southern birds had departed before the northern birds had even begun to migrate.
Previous work from the Manawatu Estuary established that the timing of migration from New Zealand relates to an individual’s eventual breeding latitude in Alaska, with birds from the southern extent of the breeding range (the Yukon-Kuskokwim Delta) leaving in early and mid-March, and birds breeding on the Seward Peninsula and North Slope not migrating until late March (Conklin et al., 2010). Because body size also varies along this S–N axis in Alaska, a relationship between body size and migration timing exists, with larger birds migrating earlier (Battley, 2006; Conklin et al., 2011). Our larger samples reinforce this previously described pattern across all sites. There was a significant negative relationship between migration date and body size for both sexes at all sites. While the slopes of the relationship were steepest at Manawatu, for any given body size Otago godwits leave substantially earlier on migration than do northern birds. Across New Zealand it seems that within a site, individuals vary according to the same ‘rule’ that arises from breeding-ground variation, but there is additional variation at the population level between non-breeding sites varying in latitude.
The slight differences in body size distributions and a lack of genetic population structure among study sites indicate that the observed differences in migration timing are not driven by geographic structure within the non-breeding range. At Manawatu, there were relatively few males >85 mm in bill length, which might cause a slight skew toward smaller, later departing birds at that site. However, such subtle differences cannot explain the magnitude of disparity in migration times between Otago and the North Island, or the regional differences for birds of the same size.
Recent work indicates that geographic variation in body size across the Alaska breeding range is accompanied by some degree of genetic differentiation, in both microsatellites and genome-wide markers (Parody-Merino, 2018; J. R. Conklin, unpublished data). Our genetic analysis is based on the expectation that any potential genetic structure would be detectable among non-breeding sites, if it was strong enough to drive different migration timing. Compared to differences between northern and southern breeders in Alaska detected in the same microsatellite loci (Fst = 0.013; Parody-Merino, 2018), we found no differences among sites in New Zealand: pairwise Fst values were effectively zero (all P > 0.40) and STRUCTURE detected no unequal distribution of genetic clusters. This lack of structure implies that godwits from different breeding areas are distributed approximately equally among non-breeding sites. Therefore, we are confident that hidden population structure cannot explain our results.
There is no clearly adaptive reason for godwits in Otago to depart more than a week earlier from New Zealand. Migration distance alone cannot explain this: the straight-line (great circle) distance to the primary stopover area in the Yellow Sea, the Yalu Jiang National Nature Reserve in China, is ca. 10,000 km from Auckland and ca. 10,600 km from Otago. These flights differ by only 6%, a distance easily traveled by a godwit in less than 12 h of non-stop flight.
Earlier departure could potentially confer benefits of early arrival in Asia or Alaska. Although we have only two geolocator tracks from Otago birds, the limited data suggest that they do not arrive in Alaska earlier than other godwits; despite departing New Zealand earlier than all godwits tracked from Manawatu, departures from Asia and arrivals in Alaska were in the same ranges as for Manawatu birds (Figure 8). Both groups flew non-stop to the Yellow Sea region, so the earlier New Zealand departures do not reflect an alternative migration strategy, in terms of route or number of stops, but did achieve a longer staging duration in Asia. Godwits spend ca. 4–6 weeks in intertidal areas of the Yellow Sea (Conklin et al., 2010; Battley et al., 2012), during which they recover from the non-stop flight from New Zealand, complete their molt into breeding plumage (Conklin and Battley, 2011a), and fuel for the subsequent flight to Alaska. Additional stopover time, or a competitively early arrival, could therefore have benefits for a bird’s condition upon arrival in Alaska, especially if food depletion occurs during staging and early-arriving birds have access to higher food levels than later-arriving birds (Choi, 2015). However, it is not clear why early arrival would be particularly advantageous for birds from southern New Zealand.
Early arrival in Asia may also come with energetic costs, given the potentially severe conditions at latitudes 35–40°N in early March. Tidal flats in north-east China can still have substantial ice cover when the first godwits arrive (Choi, 2015), and cold conditions on arrival were confirmed by geolocators (22 Manawatu, 1 Otago). For these 23 birds, the lowest temperature experienced in the week after arrival in Asia averaged −0.8°C (range −4.4 to 3.0°C), while the coldest 4-h block (i.e., with the lowest maximum temperature) averaged 2.6°C (range −1.9 to 8.8°C). These were considerably lower than temperatures in the week before departure from New Zealand (lowest temperatures: mean 7.5°C, range 3.5–10.8°C; lowest maximum: mean 12.5°C, range 8.5–16.5°C), so godwits are flying to colder conditions than they left from.
Still, godwits in Otago might face different energetic tradeoffs (i.e., the relative advantages of being in New Zealand or Asia), if they face more steeply declining temperatures or prey resources in February–March than do northern birds. We have no data to address fine-scale temporal variation in prey availability in New Zealand, but we find this explanation unlikely, as any scenario based on deteriorating conditions in southern New Zealand would also have to explain how these birds manage to fuel sufficiently for a 10,000 km non-stop flight earlier than more northerly godwits. If Otago godwits face harsher or more unpredictable conditions in New Zealand and Asia, we would expect them to experience lower or more variable annual survival or breeding success; as we also lack data to address this question, the fitness consequences of these migration differences remain unknown.
In general terms, annual routines in birds are believed to involve an endogenous circannual cycle, which is entrained by photoperiod (Gwinner, 2003). Given that migrants experience a range of photoperiods through the year, there are complex interactions between photoperiod and circannual cycles that make birds responsive to critical daylengths at seasonally appropriate times. Differences between populations in the response to photoperiod and therefore the timing of annual cycle events can be regarded as ‘adaptive population-specific reaction norms’ (Gwinner, 2003). The influence of photoperiod has been studied most extensively in relation to the timing of breeding, but some key insights from studies of photoperiodism and the annual cycle are relevant to the timing of migration. First, a given cue-response system will show conditional plasticity, in which birds with identical photoperiod response systems will produce different, and potentially appropriate, timing of annual cycle events under different photoperiods (Hahn and MacDougall-Shackleton, 2008). Second, plastic responses to novel photoperiod conditions need not result in adaptive change (Coppack and Pulido, 2004).
A limited number of experiments have simulated, in effect, a range shift in migratory birds similar to our situation with godwits in northern and southern New Zealand. Gwinner (1996b) studied the nocturnal activity of garden warblers Sylvia borin exposed to photoperiods simulating 0° and 20°S, and showed that birds with 20°S photoperiods (outside the usual range) exhibited zugunruhe about 2 months earlier than those with equatorial photoperiods. He interpreted this advancement as being advantageous if it would allow individuals to reach the breeding grounds on time, despite a longer migration. Coppack et al. (2008) simulated a northward shift by pied flycatchers Ficedula hypoleuca from 10°N to 50°N, and found that the onset of migration was advanced by 25–33 days in all treatments (20°N to 50°N) compared with 10°N, suggesting the existence of a photoperiod threshold between 10°N and 20°N. These studies indicate that photoperiod can have a direct influence on the timing of migration in birds, and that longer photoperiods resulted in earlier migration.
In this context, it seems likely that the earlier migration of godwits in southern New Zealand represents a direct response to the longer photoperiods experienced throughout the southern summer by those birds. If true, there need not be any selective advantage to migrating earlier. The pattern of larger birds migrating earlier than smaller birds was similar across all sites, suggesting that individuals from across the breeding range respond similarly to photoperiod, regardless of the actual photoperiod experienced. This implies that the inputs to the finer-scale control of timing, derived on the breeding grounds (genetic inheritance, parental effects, and entrainment by perinatal conditions), are strong and persistent, and individuals from different parts of the breeding range respond differently to a common photoperiod environment at any given non-breeding site.
Photoperiod responses presumably evolved as adaptive systems to conditions experienced by given populations. The current timing of migration of godwits in southern New Zealand seems excessively early relative to the timing of birds further north. It could be that a general system in which birds living further from the equator leave earlier on migration is adaptive if the migration is income-fueled, with birds making multiple short flights and fueling at each stop. In contrast, the long-distance flights of shorebirds are fueled by large tissue deposits accumulated before migration starts (Piersma and Gill, 1998; Battley and Piersma, 2005), resulting in quick travel between very distant sites, changing the relative balance between active traveling time and overall migration speed. It could also be that the mechanism and response evolved under more northerly photoperiods, and result in appropriate local timing at those latitudes. If the distribution of godwits has expanded further south within the East Asian-Australasian Flyway, birds may be experiencing longer photoperiods than previously. It is not known whether the trans-Pacific migration system of Alaskan bar-tailed godwits evolved through a shift in the wintering range (from Asia to Australia and New Zealand) or from a shift in the breeding range from Russia to Alaska (Hedenström, 2010); the former would entail a shift toward increasingly long photoperiods on the wintering grounds.
If the timing of migration of godwits at the population level does respond to photoperiod, this should lead to predictable differences in migration timing across the entire non-breeding range of latitude, which extends northward into the Tropics in eastern Australia. For a preliminary look at this, we compiled all previous information about bar-tailed godwit migration timing from New Zealand and Australia (Battley, 1997; Wilson et al., 2007; this study; Figure 10). Morphometric and phenology data from eastern Australia (Wilson et al., 2007) suggest that these sites also contain individuals from across the entire Alaska breeding range. There is no published information about migration phenology of L. l. baueri north of 32°S, however, two godwits were recently tracked by satellite-telemetry from Moreton Bay (27.2°S) to breeding sites on the north slope of Alaska (Z. Ma, personal communication). Their departure dates are ca. 1–2 weeks later than northern breeding birds from Manawatu, and more than 3 weeks later than the latest observed departures from Otago (Figure 10). Although these studies include a variety of methods and time periods, and therefore are not ideally comparable, it appears that migration timing in New Zealand can be viewed as part of a cline that extends for the entire non-breeding range, as might be expected if differences are photoperiod-driven. Again, migration distance can explain very little of this variation, as a non-stop flight from Moreton Bay to the Yellow Sea is ca. 2,600 km shorter than from Otago, a difference of less than 2 days of flight.
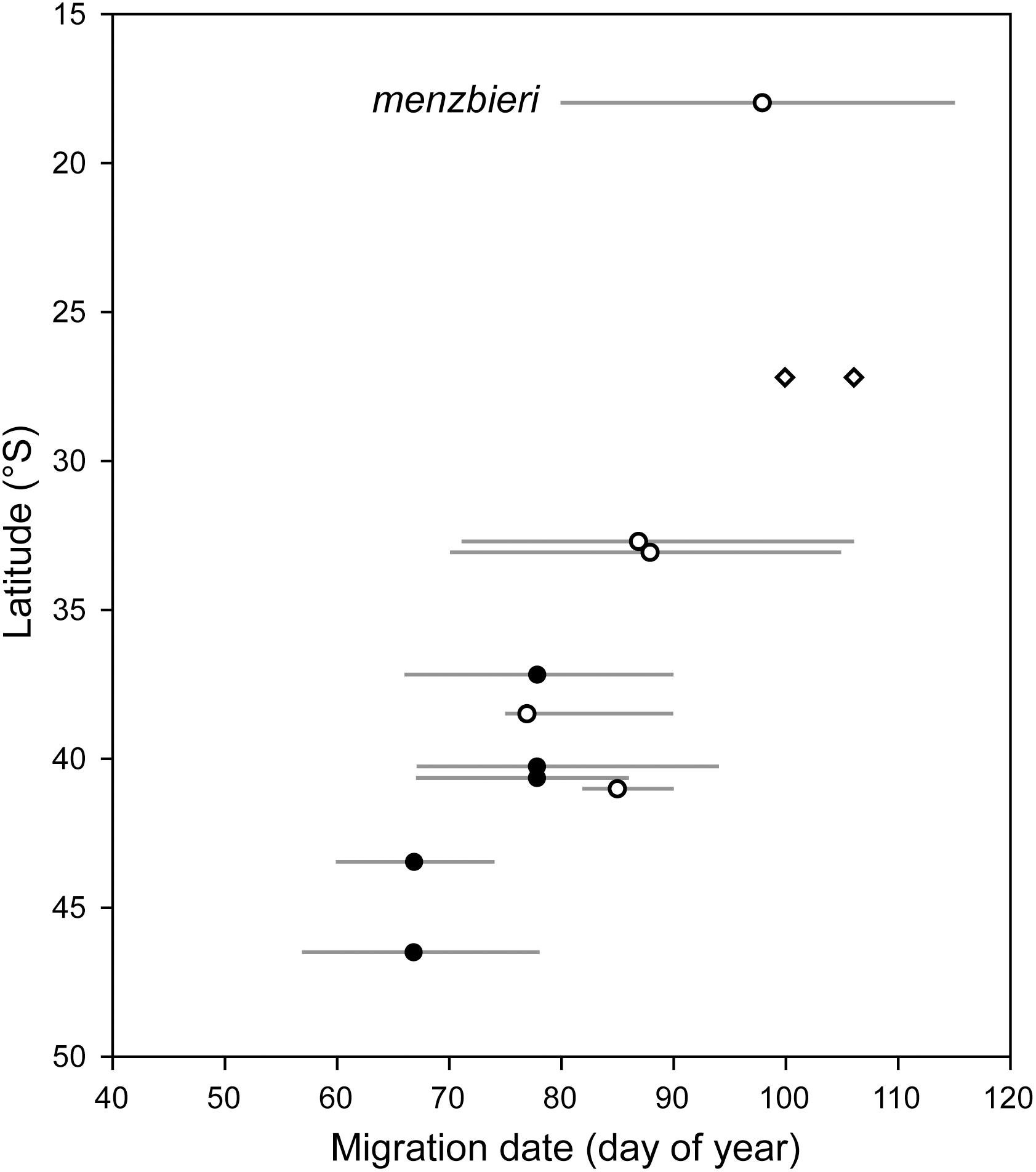
Figure 10. Variation in timing of northward departure by bar-tailed godwits across non-breeding latitudes in New Zealand (filled circles) and Australia (open circles). For each site, line indicates range of departure dates directly observed or inferred from flock counts, and circle indicates date when ca. 50% of local population had migrated. Data sources: Battley, 1997; Wilson et al., 2007; this study. Diamonds indicate departures from Moreton Bay, Australia by two PTT-tracked godwits in 2019; both were tracked to breeding sites in northernmost Alaska (Z. Ma, personal communication).
A second bar-tailed godwit subspecies (L. l. menzbieri) breeds in northeastern Russia and spends the non-breeding season in western and northern Australia. This population is known to migrate later than baueri, both on departure from northwest Australia (Figure 10; Wilson et al., 2007) and arrival at staging sites in the Yellow Sea (Choi et al., 2015), which is generally attributed to its later breeding phenology and shorter migration distance (Battley et al., 2012). However, this intuitive interpretation is subject to confounding effects of non-breeding latitude. At one site in northwest Australia, Verhoeven et al. (2016) found a surprising lack of differences in timing of fueling and migratory departure in two subspecies of red knots, Calidris canutus rogersi and C. c. piersmai. Based on their disparate phenologies on the breeding grounds (Chukotka Peninsula and the New Siberian Islands in Russia, respectively) and perceived passage times through the Yellow Sea, these populations were expected to differ by 2–4 weeks in departure timing; their indistinguishable timing leaving Australia suggests that common non-breeding geography effectively over-rides circannual schedules conferred by breeding geography, at least for the first stage of northward migration. If this similarly applies to bar-tailed godwits, we may more correctly view the migration timing of menzbieri as part of the latitudinal cline seen in baueri (Figure 10).
Other components of the annual cycle are also known to be influenced by photoperiod, and an additional question is whether photoperiod-driven differences in departure are reflected in similar differences in timing of molt and fueling, or carry through to later stages of migration and even breeding. We require more individual data on these factors, and complete northward and southward migration timing, to determine the extent to which non-breeding latitude influences phenology of the entire annual cycle.
Regardless of the physiological mechanisms involved, we have shown that some portion of between-individual variation in migration timing in bar-tailed godwits is governed by non-breeding site, and this is independent from variation associated with the natal site. This demonstrates that adult annual schedules, while guided to some degree by an endogenous program conferred by direct inheritance combined with the pre-fledging environment, are further modified according to behavioral decisions of young birds after arrival in the non-breeding range. We have also shown that, although some young godwits appear to settle at their ultimate non-breeding sites quite quickly after arrival, others do not settle until the age of 2–3 years or possibly later, providing quite an extended period for extrinsic forces to shape the highly repeatable behavior of adults. Furthermore, some young godwits arrive in Australia and subsequently shift to New Zealand as juveniles or immatures (Australasian Wader Studies Group, unpublished data), so there may be additional variation in when birds reach New Zealand resulting from differences in their initial southward migration. Once in New Zealand, young birds may move widely around the country, both northward and southward, indicating that post-banding movements are not just extensions of the first southward migration but appear to represent large-scale ‘sampling’ of habitats around the country.
It is not clear whether this suggests a prolonged ‘ontogenetic window’ (sensu Senner et al., 2015) for godwits and other avian species showing delayed maturation, in the sense of having a longer period of ‘developmental plasticity’ (sensu Piersma and Drent, 2003). If the population departure time is set by a simple response to a local photoperiod, then birds might simply need to have settled at a site for a single summer before migrating, to match other local individuals. What is more interesting is whether the internal cues for relative migration time are reinforced by repeated exposure to local photoperiod (being stronger in early-settling birds), and whether these cues are reinforced with migration to the breeding grounds.
It is intriguing that young godwits vary substantially in both when they settle at a non-breeding site and when they make their first northbound migration. Currently, we lack the data to determine whether these timings are linked. If earlier-settling birds indeed also migrate at a younger age, the causality could plausibly operate in either direction: (1) birds are somehow predisposed to migrate at different ages and then settle accordingly to ensure timely preparation for the first northward migration, or (2) the act of settling effectively initiates the adult annual cycle, including molt and fueling, after which migration naturally ensues. In the latter scenario, age of first migration could be influenced by the specific time of year that a bird settles at its final non-breeding site. For example, if godwits use an environmental cue to begin migratory preparation (e.g., photoperiod in late December), perhaps birds that have not settled by this time are insensitive to the cue and thus delay migration until the following year. Alternatively, all birds are sensitive to the cue, but birds that have not yet settled simply cannot complete migratory preparation in time. So, it is possible that age of first migration is a pre-determined strategy that varies among individuals, or a carry-over effect of circumstances experienced after arrival in New Zealand.
With its demonstrated influence on migration schedules in adult bar-tailed godwits, the processes and circumstances promoting non-breeding settlement by subadult birds may have life-long effects on behavior of individuals. Observed pre-migratory movements in New Zealand suggest a variable period of ‘sampling’ before individuals ‘choose’ a non-breeding site, to which they are extremely faithful as adults. To understand this process, the first step is to quantify the between-individual variation in duration and extent of site-sampling, and to link this with adult migratory behavior; this requires tracking individual movements from first arrival in New Zealand until the adoption of adult routines. The next step is to understand the specific processes that promote an individual’s movement or settlement, which likely include the interaction of intrinsic factors (e.g., personality, quality, circannual rhythm, condition) and extrinsic aspects of sampled sites, such as carrying capacity, prey types, and social environment.
The datasets generated for this study are available on request to the corresponding author.
All work was undertaken under appropriate Animal Ethics permits from the University of Otago and Massey University and Department of Conservation approvals.
PB and JC conceived the study and wrote the manuscript. PB, JC, and ÁP-M analyzed the data. All authors participated in bird captures. DM, AR, and RS led substantial cannon-netting efforts as part of this. JC, PL, IS, and TB conducted the departure monitoring.
Much of this work was supported by a Marsden Grant (MAU1202) from the Royal Society of New Zealand to PB. PB was also supported by the Massey University Research Fund. JC was supported by a Massey University Doctoral Scholarship, the Manawatu Estuary Trust, the Dobberke Foundation for Comparative Psychology, and the Academy Ecology Fund of the Royal Netherlands Academy of Arts and Sciences (KNAW). Early banding work was funded by a Foundation for Science, Research and Technology Postdoctoral Fellowship to PB (UOOX0231), and by the New Zealand Department of Conservation (Investigation number 3739 to PB, DM, and RS).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Thanks to B. Helm and J. Karagicheva for discussions about photoperiodic control of migration. Assistance with godwit capturing, and resightings of marked birds, were provided by numerous people—thanks to them all! Thanks to A. Crossland for the counts from the Avon-Heathcote Estuary in Christchurch. Thanks to A. Fidler and Y. Verkuil for assistance with microsatellite analyses, S. Lisovski for assistance with geolocator analyses, and Z. Ma (Fudan University) for access to godwit tracking data. We would also like to thank the various Maori iwi around New Zealand who have given their support to our kuaka research, including Ngāti Kurī and Te Aupōuri Trust Boards, Ngati Paoa, Rangitaane o Manawatū, Ngati Raukawa and Ngai Tahu.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.00052/full#supplementary-material
Alerstam, T., and Lindström, A. (1990). “Optimal bird migration: the relative importance of time, energy and safety,” in Bird Migration: The Physiology and Ecophysiology, ed. E. Gwinner (New York, NY: Springer), 331–351. doi: 10.1007/978-3-642-74542-3_22
Battley, P. F. (1997). The northward migration of arctic waders in New Zealand: departure behaviour, timing and possible migration routes of red knots and bar-tailed godwits from Farewell Spit, north-west Nelson. Emu 97, 108–120. doi: 10.1071/mu97014
Battley, P. F. (2006). Consistent annual schedules in a migratory shorebird. Biol. Lett. 2, 517–520. doi: 10.1098/rsbl.2006.0535
Battley, P. F., and Conklin, J. R. (2017). Geolocator wetness data accurately detect periods of migratory flight in two species of shorebird. Wader Study 124, 112–119. doi: 10.18194/ws.00068
Battley, P. F., and Piersma, T. (2005). Body composition and flight ranges of bar-tailed godwits (Limosa lapponica baueri) from New Zealand. Auk 122, 922–937. doi: 10.1093/auk/122.3.922
Battley, P. F., Schuckard, R., and Melville, D. S. (2011). Movements of Bar-Tailed Godwits and Red Knots within New Zealand. Science for Conservation 315. Wellington: New Zealand Department of Conservation.
Battley, P. F., Warnock, N., Tibbitts, T. L., Gill, R. E., Piersma, T., Hassell, C. J., et al. (2012). Contrasting extreme long-distance migration patterns in bar-tailed godwits Limosa lapponica. J. Avian Biol. 43, 21–32. doi: 10.1111/j.1600-048X.2011.05473.x
Bojarinova, J., and Babushkina, O. (2015). Photoperiodic conditions affect the level of locomotory activity during autumn migration in the Long-tailed Tit (Aegithalos c. caudatus). Auk 132, 370–379. doi: 10.1642/auk-14-155.1
Both, C. (2010). Flexibility of timing of avian migration to climate change masked by environmental constraints en route. Curr. Biol. 20, 243–248. doi: 10.1016/j.cub.2009.11.074
Both, C., Bijlsma, R. G., and Ouwehand, J. (2016). Repeatability in spring arrival dates in pied flycatchers varies among years and sexes. Ardea 104, 3–21. doi: 10.5253/arde.v104i1.a1
Briedis, M., Hahn, S., Gustafsson, L., Henshaw, I., Träff, J., Král, M., et al. (2016). Breeding latitude leads to different temporal but not spatial organization of the annual cycle in a long-distance migrant. J. Avian Biol. 47, 743–748. doi: 10.1111/jav.01002
Buchmann, R., and Hazelhurst, S. (2015). The “Genesis” Manual. Johannesburg: University of the Witwatersrand.
Choi, C. Y., Battley, P. F., Potter, M. A., Rogers, K. G., and Ma, Z. (2015). The importance of Yalu Jiang coastal wetland in the north Yellow Sea to Bar-tailed Godwits Limosa lapponica and Great Knots Calidris tenuirostris during northward migration. Bird Conserv. Int. 25, 53–70. doi: 10.1017/S0959270914000124
Choi, J. (2015). The Northward Migration Stopover Ecology of Bar-Tailed Godwits and Great Knots in the Yalu Jiang Estuary National Nature Reserve, China. Ph. D. thesis. Massey University, Palmerston North.
Ciarleglio, C. M., Axley, J. C., Strauss, B. R., Gamble, K. L., and McMahon, D. G. (2010). Perinatal photoperiod imprints the circadian clock. Nat. Neurosci. 14, 25–27. doi: 10.1038/nn.2699
Conklin, J. R., and Battley, P. F. (2011a). Contour-feather moult of Bar-tailed Godwits (Limosa lapponica baueri) in New Zealand and the Northern Hemisphere reveals multiple strategies by sex and breeding region. Emu 111, 330–340. doi: 10.1071/MU11011
Conklin, J. R., and Battley, P. F. (2011b). Impacts of wind on individual migration schedules of New Zealand bar-tailed godwits. Behav. Ecol. 22, 854–861. doi: 10.1093/beheco/arr054
Conklin, J. R., Battley, P. F., and Potter, M. A. (2013). Absolute consistency: individual versus population variation in annual-cycle schedules of a long-distance migrant bird. PLoS One 8:e54535. doi: 10.1371/journal.pone.0054535
Conklin, J. R., Battley, P. F., Potter, M. A., and Fox, J. W. (2010). Breeding latitude drives individual schedules in a trans-hemispheric migrant bird. Nat. Commun. 1:67. doi: 10.1038/ncomms1072
Conklin, J. R., Battley, P. F., Potter, M. A., and Ruthrauff, D. R. (2011). Geographic variation in morphology of Alaska-breeding bar-tailed godwits (Limosa lapponica) is not maintained on their nonbreeding grounds in New Zealand. Auk 128, 363–373. doi: 10.1525/auk.2011.10231
Coppack, T., and Pulido, F. (2004). Photoperiodic response and the adaptability of avian life cycles to environmental change. Adv. Ecol. Res. 35, 131–150. doi: 10.1016/s0065-2504(04)35007-5
Coppack, T., Tindemans, I., Czisch, M., Van der Linden, A., Berthold, P., and Pulido, F. (2008). Can long-distance migratory birds adjust to the advancement of spring by shortening migration distance? The response of the pied flycatcher to latitudinal photoperiodic variation. Glob. Chang. Biol. 14, 2516–2522. doi: 10.1111/j.1365-2486.2008.01668.x
Cresswell, W. (2014). Migratory connectivity of Palaearctic-African migratory birds and their responses to environmental change: the serial residency hypothesis. Ibis 156, 493–510. doi: 10.1111/ibi.12168
Dawson, A. (2008). Control of the annual cycle in birds: endocrine constraints and plasticity in response to ecological variability. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 1621–1633. doi: 10.1098/rstb.2007.0004
Duriez, O., Bauer, S., Destin, A., Madsen, J., Nolet, B. A., Stillman, R. A., et al. (2009). What decision rules might pink-footed geese use to depart on migration? An individual-based model. Behav. Ecol. 20, 560–569. doi: 10.1093/beheco/arp032
Emmenegger, T., Hahn, S., and Bauer, S. (2014). Individual migration timing of common nightingales is tuned with vegetation and prey phenology at breeding sites. BMC Ecol. 14:9. doi: 10.1186/1472-6785-14-9
Excoffier, L., and Lischer, H. E. L. (2010). Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 10, 564–567. doi: 10.1111/j.1755-0998.2010.02847.x
Fry, C. H., Ferguson-Lees, I. J., and Dowsett, R. J. (1972). Flight muscle hypertrophy and ecophysiological variation of Yellow Wagtail Motacilla flava races at Lake Chad. J. Zool. 197, 293–306. doi: 10.1111/j.1469-7998.1972.tb03113.x
Gunnarsson, T. G., Gill, J. A., Petersen, A., Appleton, G. F., and Sutherland, W. J. (2005). A double buffer effect in a migratory shorebird population. J. Anim. Ecol. 74, 965–971. doi: 10.1111/j.1365-2656.2005.00994.x
Gwinner, E. (1996a). Circadian and circannual programmes in avian migration. J. Exp. Biol. 199, 39–48.
Gwinner, E. (1996b). Circannual clocks in avian reproduction and migration. Ibis 138, 47–63. doi: 10.1111/j.1474-919x.1996.tb04312.x
Hahn, T. P., and MacDougall-Shackleton, S. A. (2008). Adaptive specialization, conditional plasticity and phylogenetic history in the reproductive cue response systems of birds. Philos. Trans. R. Soc. B Biol. Sci. 363, 267–286. doi: 10.1098/rstb.2007.2139
Hedenström, A. (2010). Extreme endurance migration: what is the limit to non-stop flight? PLoS Biol. 8:e1000362. doi: 10.1371/journal.pbio.1000362
Helm, B., and Gwinner, E. (2005). Carry-over effects of day length during spring migration. J. Ornithol. 146, 348–354. doi: 10.1007/s10336-005-0009-5
Helm, B., Schwabl, I., and Gwinner, E. (2009). Circannual basis of geographically distinct bird schedules. J. Exp. Biol. 212, 1259–1269. doi: 10.1242/jeb.025411
Hubisz, M. J., Falush, D., Stephens, M., and Pritchard, J. K. (2009). Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 9, 1322–1332. doi: 10.1111/j.1755-0998.2009.02591.x
Lisovski, S., and Hahn, S. (2012). GeoLight – processing and analysing light-based geolocator data in R. Methods Ecol. Evol. 3, 1055–1059. doi: 10.1111/j.2041-210X.2012.00248.x
Lourenço, P. M., Alves, J. A., Reneerkens, J., Loonstra, A. H. J., Potts, P. M., Granadeiro, J. P., et al. (2016). Influence of age and sex on winter site fidelity of sanderlings, Calidris alba. PeerJ 4:e2517. doi: 10.7717/peerj.2517
Mueller, T., O’Hara, R. B., Converse, S. J., Urbanek, R. P., and Fagan, W. F. (2013). Social learning of migratory performance. Science 341, 999–1003. doi: 10.1126/science.1237139
Myers, J. P. (1981). A test of three hypotheses for latitudinal segregation of the sexes in wintering birds. Can. J. Zool. 59, 1527–1534. doi: 10.1139/z81-207
Myers, J. P., Maron, J. L., and Sallaberry, M. (1985). Going to extremes: why do Sanderlings migrate to the Neotropics? Ornithol. Monogr. 36, 520–535. doi: 10.2307/40168302
Nebel, S. (2005). Latitudinal clines in bill length and sex ratio in a migratory shorebird: a case of resource partitioning? Acta Oecol. 28, 33–38. doi: 10.1016/j.actao.2005.02.002
Ouwehand, J., Ahola, M. P., Ausems, A. N. M. A., Bridge, E. S., Burgess, M., Hahn, S., et al. (2016). Light-level geolocators reveal migratory connectivity in European populations of pied flycatchers Ficedula hypoleuca. J. Avian Biol. 47, 69–83. doi: 10.1111/jav.00721
Parody-Merino, A. M. (2018). Genetics of Migration Timing in Bar-Tailed Godwits. Ph. D thesis, Massey University, Palmerston North.
Piersma, T., and Drent, J. (2003). Phenotypic flexibility and the evolution of organismal design. Trends Ecol. Evol. 18, 228–233. doi: 10.1016/S0169-5347(03)00036-3
Piersma, T., and Gill, R. E. Jr. (1998). Guts don’t fly: small digestive organs in obese bar-tailed godwits. Auk 115, 196–203. doi: 10.2307/4089124
Piersma, T., Rogers, D., González, P. M., Zwarts, L., Niles, L. J., Lima, I., et al. (2005). “Fuel storage rates before northward flights in Red Knots worldwide – facing the severest ecological constraint in tropical intertidal environments?,” in Birds of Two Worlds: The Ecology and Evolution of Migratory Birds, eds R. Greenberg and P. P. Marra (Baltimore, MD: The Johns Hopkins University Press), 262–273.
Scheiffarth, G., Wahls, S., Ketzenberg, C., and Exo, K. M. (2002). Spring migration strategies of two populations of bar−tailed godwits, Limosa lapponica, in the Wadden Sea: time minimizers or energy minimizers? Oikos 96, 346–354. doi: 10.1034/j.1600-0706.2002.960216.x
Senner, N. R., Conklin, J. R., and Piersma, T. (2015). An ontogenetic perspective on individual differences. Proc. R. Soc. Lond. B Biol. Sci. 282, 20151050. doi: 10.1098/rspb.2015.1050
Sergio, F., Tanferna, A., De Stephanis, R., Jiménez, L. L., Blas, J., Tavecchia, G., et al. (2014). Individual improvements and selective mortality shape lifelong migratory performance. Nature 515, 410–413. doi: 10.1038/nature13696
Studds, C. E., and Marra, P. P. (2005). Nonbreeding habitat occupancy and population processes: an upgrade experiment with a migratory bird. Ecology 86, 2380–2385. doi: 10.1890/04-1145
Thorup, K., Alerstam, T., Hake, M., and Kjellen, N. (2003). Bird orientation: compensation for wind drift in migrating raptors is age dependent. Proc. R. Soc. Lond. B Biol. Sci. 270, S8–S11. doi: 10.1098/rsbl.2003.0014
van Bemmelen, R. S. A., Kolbeinsson, Y., Ramos, R., Gilg, O., Alves, J. A., Smith, M., et al. (2019). A migratory divide among red-necked phalaropes in the Western Palearctic reveals contrasting migration and wintering movement strategies. Front. Ecol. Evol. 7:86. doi: 10.3389/fevo.2019.00086
Verhoeven, M. A., Van Eerbeek, J., Hassell, C. J., and Piersma, T. (2016). Fuelling and moult in Red Knots before northward departure: a visual evaluation of differences between ages, sexes and subspecies. Emu 116, 158–167. doi: 10.1071/MU15035
Keywords: geolocation, migration timing, phenology, photoperiod, Scolopacidae
Citation: Battley PF, Conklin JR, Parody-Merino ÁM, Langlands PA, Southey I, Burns T, Melville DS, Schuckard R, Riegen AC and Potter MA (2020) Interacting Roles of Breeding Geography and Early-Life Settlement in Godwit Migration Timing. Front. Ecol. Evol. 8:52. doi: 10.3389/fevo.2020.00052
Received: 31 August 2019; Accepted: 19 February 2020;
Published: 17 March 2020.
Edited by:
Yolanda E. Morbey, University of Western Ontario, CanadaReviewed by:
Kevin C. Fraser, University of Manitoba, CanadaCopyright © 2020 Battley, Conklin, Parody-Merino, Langlands, Southey, Burns, Melville, Schuckard, Riegen and Potter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Phil F. Battley, cC5iYXR0bGV5QG1hc3NleS5hYy5ueg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.