
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 24 September 2019
Sec. Behavioral and Evolutionary Ecology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00356
This article is part of the Research Topic Cooperation and Coordination in the Family View all 26 articles
 Léa Lejeune1*
Léa Lejeune1* James L. Savage2,3
James L. Savage2,3 Aisha C. Bründl1,4
Aisha C. Bründl1,4 Alice Thiney1
Alice Thiney1 Andrew F. Russell1,4
Andrew F. Russell1,4 Alexis S. Chaine1,5†
Alexis S. Chaine1,5†In bi-parental care systems each parent shares benefits with its unrelated partner from the common investment in offspring, but pays an individual cost of providing that care, leading to sexual conflict. However, several recent empirical studies have shown that coordinating behaviours like synchronisation (e.g., arriving at similar times) and alternation (taking turns in providing care) at the nest lead to increased investment overall, presumably to reduce conflict through policing or synergistic benefits. Ecological conditions should impact the costs and benefits of bi-parental care, yet there exists a gap in research on the relationship between ecological conditions and patterns of parental care behaviour beyond visitation rate. Here we provide an examination of how bi-parental provisioning behaviours, i.e., pair feeding rate and feeding consistency, and the degree to which parents synchronise or take turns, differ under contrasting ecological conditions in populations of blue tits (Cyanistes caeruleus) spanning a 1,000 m altitudinal gradient. We found that blue tit pairs synchronised and alternated more than expected by chance, and that care patterns were modified by ecology. Pairs synchronised more in woodland-pasture edges than in woodland interiors, and alternated more and fed more frequently at lower altitude compared to higher altitude nests. Variation in bi-parental coordination behaviours did not have a significant impact on fledging success but more synchronous nests had heavier chicks in woodland habitats. Taken as a whole, our results show that patterns of care are influenced by ecological conditions and that their interplay may change the outcome of sexual conflict.
In 1972, Trivers' landmark paper (Trivers, 1972) suggested that the current-future reproduction trade-off should lead to sexual conflict between parents, as each parent benefits when the other provides a greater share of the resources needed by offspring. Using this framework, many experimental and theoretical studies have since examined the outcome of sexual conflict over feeding decisions in bi-parental systems, at both evolutionary and behavioural time scales (Lessells, 2013). Theory generally suggests that the best strategy for parents is to incompletely compensate for changes in their partner's feeding rate (sealed bid models: Houston and Davies, 1985; negotiation models: McNamara et al., 1999, 2003; Houston et al., 2005; Ewald et al., 2007; Lessells and McNamara, 2011; but see Jones et al., 2002), at least when parents have similar information about the brood need (Johnstone and Hinde, 2006). This prediction has been generally supported by empirical results, although considerable variation in parental strategies remains to be explained (review: Harrison et al., 2009; Meade et al., 2011). The role of ecology in shaping parental care decisions is one potential source of this variation, but relatively little attention has focused on the impact of ecology on conflict over bi-parental care.
Existing theoretical work on parental care has focused on the amount of care that each parent provides as a continuous variable, ignoring variation in the environment in order to maximise simplicity and generality. The realities of care are usually more complex: in the case of feeding behaviour parents often provide care to offspring in discrete units (e.g., food items), and offspring may benefit from both the consistency of care as well as the total amount of prey delivered. Consistent care (i.e., low variance in provisioning) is more efficient for rearing offspring, but high-variance, risk-prone provisioning could be adaptive when offspring are in poor condition or in poor quality habitats (Ydenberg, 2008; Westneat et al., 2012; Mathot et al., 2017). With discrete events, costs of sexual conflict are predicted to be reduced by alternating care from the parents (Johnstone et al., 2014). Likewise, synchronisation of parental visits at the nest could decrease costs of sexual conflict. As with overall care levels, such behaviours could also be influenced by ecology. For example, the distribution of food resources could influence the consistency of care (Ydenberg, 2008; Westneat et al., 2012; Mathot et al., 2017) and the degree of alternation between parents. Likewise, elevated predation risk could favour synchronisation of nest visits. If these patterns of care are important for the overall costs and benefits of care, then the outcome of sexual conflict should depend on ecological conditions such as spatial and temporal availability of food or the risk of predation.
The view that patterns of offspring care influence sexual conflict and the success of parental care has gained support through detailed studies of feeding behaviour in bi-parental and cooperative breeding species. Bebbington and Hatchwell (2015) found a positive relationship between turn-taking and total feeding rate in bi-parental (pairs without helpers) long-tailed tits (Aegithalos caudatus), as predicted by Johnstone et al.'s (2014) model, showing that alternation might reduce the costs of sexual conflict for offspring and parents through a sort of policing mechanism (Trivers, 1971; Axelrod and Hamilton, 1981). Alternation also occurs more often than expected by chance in the cooperative breeding species chestnut-crowned babbler (Pomatostomus ruficeps) (Savage et al., 2017), and nest visit synchrony has also been documented in several species with bi-parental and cooperative care (Krebs et al., 1999; Masello et al., 2006; Lee et al., 2010; van Rooij and Griffith, 2013; but see Gray and Hamer, 2001). Synchronously visiting the nest can be adaptive by reducing predation risk for offspring (Raihani et al., 2010), by facilitating equal partitioning of food among nestlings as fed simultaneously (Shen et al., 2010), or by improving assessment of chick need (McDonald et al., 2008). Synchrony at the nest has been associated with more regular feed visits, without involving a higher overall visitation rate or greater equity between partners' visit rates, but is associated with increased offspring mass and number in bi-parental species (Mariette and Griffith, 2012, 2015). These results suggest that in addition to feeding rate, studying consistency of care, and synchrony and alternation of parental visits might be crucial for understanding the outcome of sexual conflict. It remains unclear how these parameters relate to each other or are modified by ecological conditions.
While models of bi-parental care have generally ignored ecological variation, substantial empirical work has explored the impact of ecology on individual foraging behaviour and total amounts of care delivered. Studies have shown clear habitat-related differences in foraging distances and feeding rates of breeding passerines, related to the abundance of food available around the nest (blue tits: Blondel et al., 1991; Tremblay et al., 2005; great tits: Naef-Daenzer et al., 2000), and individuals in rich habitats feed more frequently and are better able to match their feeding rate to the size of the brood than individuals in poor habitats (Tremblay et al., 2003, 2005; Stauss et al., 2005). Food availability can be difficult to measure, but habitat feature such as at the edge versus interior of forests and species diversity can provide good proxies. Variation in food abundance and other ecological patterns such as forest-edge effects impact fitness among woodland passerines (Murcia, 1995; Wilkin et al., 2007). While reproductive output is often reduced at forest edges (but see Lahti, 2001 and Wilkin et al., 2007), it is unclear if such effects result from increased competition with conspecifics (Huhta et al., 1999), higher predation exposure (Batàry and Bàldi, 2004), or reduced food availability (Huhta et al., 1999). Likewise, tree diversity is often related to insect availability and therefore can impact parental care and fitness (Gering and Crist, 2000; Sobek et al., 2009). Whether parents also respond to these different conditions by altering their patterns of care provisioning, and whether these tactics impact fitness outcomes for parents or offspring is still unknown. In this context altitudinal gradients provide good model systems, as they cause a sharp change in ecology as increasing elevations have colder temperatures, greater seasonality, shorter breeding seasons, and greater fluctuations in food availability (Abdusalyamov, 1964), which can have downstream effects on parental care strategies and fitness (see Badyaev and Ghalambor, 2001 for an inter-species review; Boyle et al., 2016 for a population review). Studies focusing on feeding behaviour along altitudinal gradients have found increases in feeding rates within species at higher elevation, either for both parents (Johnson et al., 2007; Lee et al., 2011) or just males (Badyaev, 1997). Sex-limited effects of ecology on care should impact sexual conflict, yet it is unclear whether patterns of care are modified under such environmental variation.
We examined how variation in ecology and altitude impacted patterns of parental care in blue tits, Cyanistes caeruleus, in a nest box population in the French Pyrenees. The blue tit is a short-lived passerine, socially monogamous, in which females and males contribute to offspring provisioning. Blue tits are almost wholly insectivorous in the breeding season and may bring arthropods to the nest at the rate of almost one a minute during much of a 16-h day (Perrins, 1991). First, we fully describe the feeding behaviour patterns of blue tits in the Pyrenees by examining the rate of food delivered, its consistency, the synchrony and alternation of feeding in breeding pairs, and how these parameters are related to each other. Secondly, we explore the influence of climatic conditions, habitat characteristics (tree diversity, forest edges vs. interiors), and altitude on these patterns, to build a more complete picture of feeding behaviour and how it relates to local ecology. Finally, we examine the fitness consequences of variation in care patterns seen across these contrasting environments.
This study was conducted during three breeding seasons from 2015 to 2017, near the research Station for Theoretical and Experimental Ecology of Moulis (SETE, UMR 5321; 42°5729N, 1°0512E), in the French Pyrenees. Our study area comprised 14 woodlots divided into 5 sites situated along an altitudinal gradient ranging from 430 to 1,530 m. Our sites each span altitudinal ranges: 430–593, 555–774, 818–1,108, 1,230–1,530, and 945–1,193 m. All sites are composed of mixed deciduous woodland interspersed to varying degrees with open areas of rough pasture or bog. More than 20 tree species have been recorded in our sites, but primarily oak (Quercus robur), ash (Fraxinus excelsior), hazel (Corylus avellana), and beech (Fagus sylvatica) have been observed, with beech more common at higher elevations and oak at lower elevations. In total the study area contained on average 600 Woodcrete nestboxes (2M entrance hole 32mm; Schwegler, Schorndorf, Germany) spaced at ca. 50 m intervals within each woodlot. The occupancy of blue tits varies from 18% of nest boxes at low altitude, and declines to 4% at high altitude (unpublished data), leading to lower densities at higher altitudes, similar to general patterns of occupancy across most tit species in our study sites.
We characterised the habitat within a 25 m radius of each nestbox in order to estimate the fine-scale habitat structure. We focused on two measures of habitat characteristics that have been shown to impact food resources and predation risk: forest edge versus interior (Huhta et al., 1999; Batàry and Bàldi, 2004) and tree diversity (Gering and Crist, 2000; Sobek et al., 2009). First, we determined whether a nest was on the edge of the woodland, based on whether or not it was within 5 m of pasture or bog. Most of the other nests were in woodland interiors. Very few nestboxes were on a solitary tree within a pasture or bog, therefore to achieve similar sample sizes between categories we removed the 7 nestboxes in these habitats from the dataset and leading to two categories: woodland edge and interior. Second, we attempted to provide a qualitative estimate of tree diversity around each nest using a qualitative approach. Tree diversity was denoted as being low if one or two species of tree made up at least 80% of the species within a 20 m radius of a nest box, while it was denoted as high if no species dominated. Finally, temperature and humidity were also measured every half hour during the breeding season using eight remote loggers (TinytagTM types TGP-4500 and TGP-4505) positioned at altitudes of 430, 565, 604, 847, 1,110, 1,002, 1,334, and 1,522 m, allowing the climatic conditions during the collection of provisioning behaviour to be estimated for each nest.
Nestboxes were checked at least twice a week for breeding activity throughout the breeding season (Mid-March to July). Nests were checked daily when fully built and lined (with feather, hair and/or wool), toward clutch completion (from the 6th egg) and from 11 days after the start of incubation, to collect data on lay date and hatch date with a 1-day precision, and on clutch size and the number of hatchlings. Nestling mass and number of nestlings were determined when broods were 15 days old and the number of fledgings was determined as the number of nestlings minus any chicks found dead in the nest after fledging. In total 471 chicks were weighed at 84 nests.
We recorded parental feeding behaviour for 3 h at least once for each nest, between 08:45 a.m. and 04:10 p.m. (mean hour = 10:50 a.m.), during peak offspring demand (brood age 11–16 d). Feeding behaviour was recorded using digital video cameras (Sony HDR-CX220E Handycam R Camcorders) positioned 5–10 m from each nest at a 45° angle to the entrance to enable identification of the provisioning adult from its unique colour-ring combination. Adult birds were individually marked with a metal ring bearing a unique number, and with a unique combination of coloured rings at least 24 h prior to the videos to enable identification. We then transcribed the arrival and departure times of each parent during 2 h of each video, with the first hour excluded to ensure that all pairs had time to habituate to the presence of the camera. The nest box opening is small enough such that only one parent can enter at a time, thereby excluding perfectly simultaneous visits, and different parents visiting within a few seconds was rare (46 intervals of less than or equal to 2 s on 15,663 visits in total). We extracted data from 131 videos of 84 nests as two videos, taken 2 days apart, were recorded on most nests. Non-feeding visits are rare at this stage in the nestling period (Nur, 1984); as such, we assumed that nest visits were feeding visits, except for when we could visibly see parents leaving the nestbox with the food item or entering without any food in its beak (considering that we estimated that non-feeding visits represented on average 1.1% of the visits for each pair in our dataset).
Bird capture was carried out under permits to ASC from the French bird ringing office (CRBPO; program 576) and breeding monitoring under permits from the state of Ariége (Préfecture de l'Ariége, Protection des Populations, no A09-4) and the Région Midi-Pyrenées (DIREN, no 2013-02).
To describe the pair's feeding behaviour we assessed how often parents visited the nest, how consistently they visited, to what degree they took turns, and whether they visited the nest together. To do so we estimated the following four parameters using the data extracted from the videos: pair mean inter-visit time interval (IVI), standard deviation of the pair IVI, and indices of nest visit synchrony and alternation by the pair, using the methods described in Savage et al. (2017) and Savage and Ouyang (2019).
Pair inter-visit time interval (IVI) is the time between two consecutive entries into the nestbox regardless of which parent entered, which is a measure inversely related to the brood feeding rate. An advantage of using brood IVI over feeding rate is that we can calculate an index of variation in IVI (standard deviation here), which provides added information on the consistency with in which food is delivered to the brood.
The synchrony index of pairs was a measure of how likely the two parents were seen together at the nest, relative to that expected by chance. We estimated the actual synchrony of breeding pairs using cross-correlations of kernel-density estimates calculated for the visits of each parent. We then randomised the inter-visit-intervals of each parent 1,000 times to generate disassociated provisioning data, and calculated the synchrony score for each randomisation to generate a distribution of synchrony scores expected by chance. Using the mean and standard deviation of this distribution we calculated the z-score of the natural data as a measure of whether pairs were more or less synchronous than expected given their particular provisioning rates and inter-visit time intervals. Using this method, positive z-scores indicate that pairs are more synchronous than expected by chance given the observed mean pair inter-visits interval, and vice versa for negative z-scores. To be clear, a high synchrony score does not necessarily correspond to a very short time interval, but rather a shorter time interval than expected from the distribution of observed visits if parents arrive at random.
The alternation (or turn-taking) index of pairs was a measure of how often parents visited following their partner versus sequential visits by the same parent. To quantify whether the observed alternation differed from that expected from parents visiting independently, we used the Wald-Wolfowitz runs test (Wald and Wolfowitz, 1940), a non-parametric test for independence of elements in a two-valued data sequence [in this case, visits by either the male (M) or the female (F)]. This test is based on the null hypothesis that elements of the sequence are independent and identically distributed (although not necessarily equally common), and generates a z-score for the observed data, which is significantly positive when there are more alternated visits than expected by chance (e.g., MFMFMFMFMF) and significantly negative when there are fewer (e.g., FFFFFMMMMM). The calculation is based on the observed number of visits from each parent such that differences in visit rate between the sexes are taken into account.
Statistical analyses were performed in R 3.4.1 (R Core Team, 2017) using the lme4 package (Bates et al., 2015). In all models explanatory variables were mean-centered to facilitate comparisons between the effect of each variable. We used AIC corrected for the sample size (AICc) to select the most parsimonious model, using the dredge function in the package MuMIn (Bartoń, 2016). This function uses the all-subset approach in which all possible combinations of fixed variables are run, the random structure is kept constant. Models that are better supported by the data while retaining fewer explanatory variables achieve lower AICc values. All rejected terms were added singly to the most parsimonious models to confirm non-significance, and reported effect sizes are derived from final models. Multi-collinearity was tested using the variance inflation factor (Fox and Monette, 1992). Residuals of our models were normally distributed (after transformation of the response variable where necessary) and independent.
The complete dataset included 91 observations from 73 pairs across the entire altitudinal gradient. High diversity habitats were in higher proportion at low altitudes (differences between both diversities = −93.70 ± SD = 30.58, p < 0.01). Therefore, to avoid confounding habitat effects with altitudinal effects, we performed the analyses testing the effect of tree diversity and habitat type on feeding behaviour on a restricted dataset, including 57 observations from 48 pairs nesting at low altitude. The analysis examining the effect of altitude and weather were performed on the whole dataset. Missing values led to smaller samples in some cases.
We investigated which environmental variables significantly explained each of the pair's feeding behaviour parameters by using four sets of general linear mixed models, one for each model. All parameters were fitted with a Gaussian error distribution, identity-link function, and maximum likelihood (ML) estimation. Our fixed effects of interest were altitude, habitat type, tree diversity and weather conditions (see below). We ran models on each pair feeding behaviour separately and did not include other parental feeding behaviours as covariates. This is because parental behaviours were correlated (see section Results) with each other as well as with environmental parameters which would lead to collinearity between predictors that could mask effects of interest in the exploratory analyses presented here. Altitude was included as a continuous variable in our models. As feeding rate can be influenced by food availability at short time scales (Arlettaz et al., 2010), we included mean temperature and mean humidity during the video as fixed effects. No correlation between climate variables and altitude were found, so we used the whole dataset to test the effect of climatic conditions. However, as mean temperature and mean humidity during the feeding video were correlated, we used a principal component analysis to create a combined “weather” variable. The first principal component, our measure of “weather,” accounted for 80.41% of the total variance in these variables. High values of this meteorological variable correspond to high humidity (loading: 0.71) and low temperature (loading: −0.71) and vice versa. This meteorological variable was split into extreme values and mid values (between percentile 15 and 85%) and the analyses were conducted for mid values, to avoid bias due to extreme values.
Additional variables were included in the models to account for their separate confounding effect on feeding behaviour traits, or to investigate whether they modified the effect of ecological variables on parental behaviour by including interaction terms. As we recorded two videos for some pairs, within a few days, with a brood swap experiment in-between, we included the order of the videos in the model to account for an effect of a possible experimental stimulation of feeding behaviour. We included brood size as a covariate to control for potential variation in visitation pattern with the number of chicks. As brood size variations can lead to variations in food demand we also tested the interactions between brood size and tree diversity, habitat type and altitude. Brood age varied from day 11 to 16 (mean = 13.6 ± SD = 1.5 days) across the videos; we therefore included age in the models as a continuous fixed effect. As laying date may indicate whether parents are synchronous with the food supply, and hence whether food is available during the rearing period, we included it in the covariates. Birds at high elevation breed later (Bründl, 2018) and as laying date can vary between years we standardised the laying date within site and year to not confound effects from late versus early breeding with altitude or year effects.
As some nestboxes had two different videos of parental care, we included nestbox identity as a random intercept, to account for non-independence of these data. Year and julian date of the video were also fitted as random intercepts to account for date-specific random variations in parental care.
To explore the ecological and parental coordination correlates of reproductive success, we conducted analyses asking whether parental feeding coordination and ecological parameters were related to fitness proxies. Mean brood mass (brood age = 15.1 days ± SD = 0.4 days) and number of fledglings were modelled using a LMM with Gaussian errors. As alternation and synchronisation may allow a better distribution of food within a brood, we also explored the relationship between these coordination behaviours and the variance of brood mass, measured by the standard deviation in chick mass near fledging. The primary terms of interest in each analysis were habitat type, altitude, and metrics of nest visit synchrony and alternation, as well as the interaction between synchrony and habitat type. Nests from the whole altitudinal gradient were included in the analysis. Tree diversity was not considered as previous analyses had shown no effect of this environmental parameter on coordination behaviours, avoiding the model to be biased by a correlation between tree diversity and altitude. IVI, lay date and the number of hatchlings were fitted as covariates in the three analyses, while the average mass analysis additionally controlled for the average tarsus length of the brood, in order to control for variation in brood age and body size, and the standard deviation mass analysis controlled for the mean brood mass. Finally, year and site were fitted as random terms for both models.
Parents visited the nest every 84.1 s on average during the brood ages considered (SD = 26.1 s), leading to a brood-level nest visitation rate of 46 times (± 11 SD) per hour. Mean synchrony z-score was 0.5 ± SD = 1.1 (range = − 2 to +3), mean alternation z-score was 1.1 ± SD = 1.3, and standard deviation in IVI was 81.7 s ± SD = 29.6 s. In our data 41% of pairs alternated significantly more than expected by chance and only 3% alternated less. Overall 66% of the breeding pairs had a positive z-score for synchrony, indicating that many pairs were more synchronous than expected by chance based on visit intervals. More synchronous pairs also alternated more, although the correlation was relatively weak (r = 0.17, p = 0.04; Figure 1).
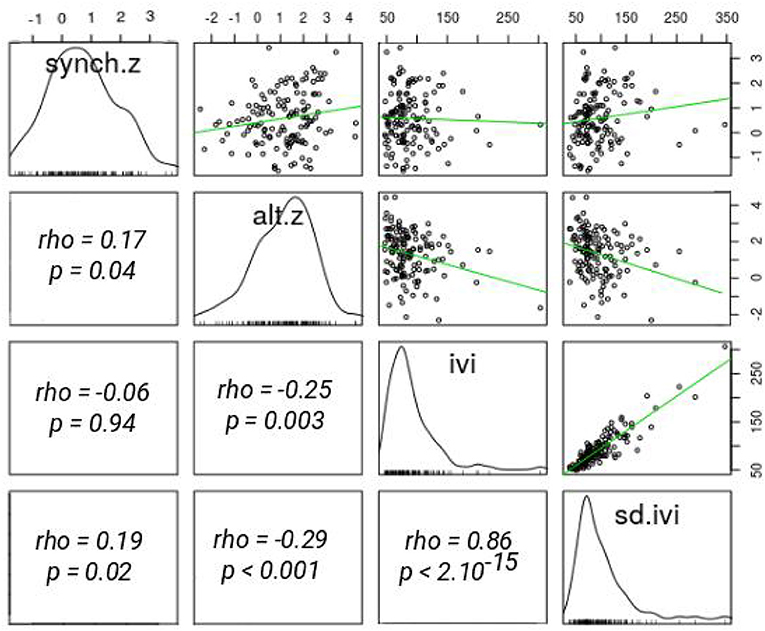
Figure 1. Correlations between pairs feeding behaviour parameters. alt.z, alternation z-score; synch.z, synchrony z-score; ivi, mean inter-visits time interval; sd.ivi, ivi standard deviation. Correlation coefficients and p-values are from Spearman correlation tests. No. of obs = 87, No. of pairs = 71.
Mean pair IVI and standard deviation in IVI were significantly positively correlated (r = 0.86, p < 2.10−15; Figure 1), suggesting that pairs with more heterogenous feeding rate also fed offspring less. Pairs that fed more also alternated more (r = −0.25, p < 0.01; Figure 1). Less synchronous pairs did not work less overall, because pair visit rate did not vary with visit synchrony (p = 0.94; Figure 1). However, less synchronous pairs had a less regular visit pattern (r = 0.19, p = 0.02; Figure 1).
The variation in the degree of nest visit synchrony varied with some, but not all ecological variables examined (Table 1). We found no association between either altitude or weather on visit synchrony (Figure 2A), although we found a significant interaction between the two variables (Table 1). Specifically pairs synchronised significantly more when the weather was cold and humid at high altitude (estimate = 0.88, SE = 0.33, p < 0.01) but not at low altitude (p = 0.32). In addition, pairs were 24% less synchronous in forest habitats than in edge habitats (Table 1, Figure 2A). By contrast, we found no evidence to suggest that the degree of nest visit synchrony was influenced by tree diversity (Table 1).
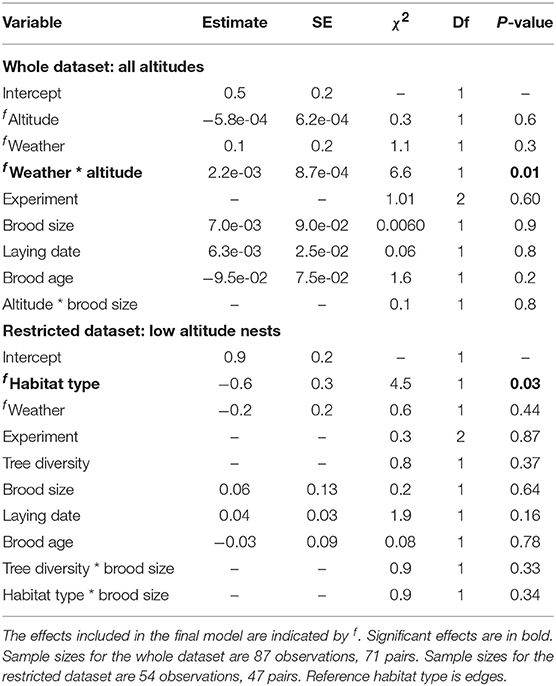
Table 1. Results of a LMM examining synchrony.
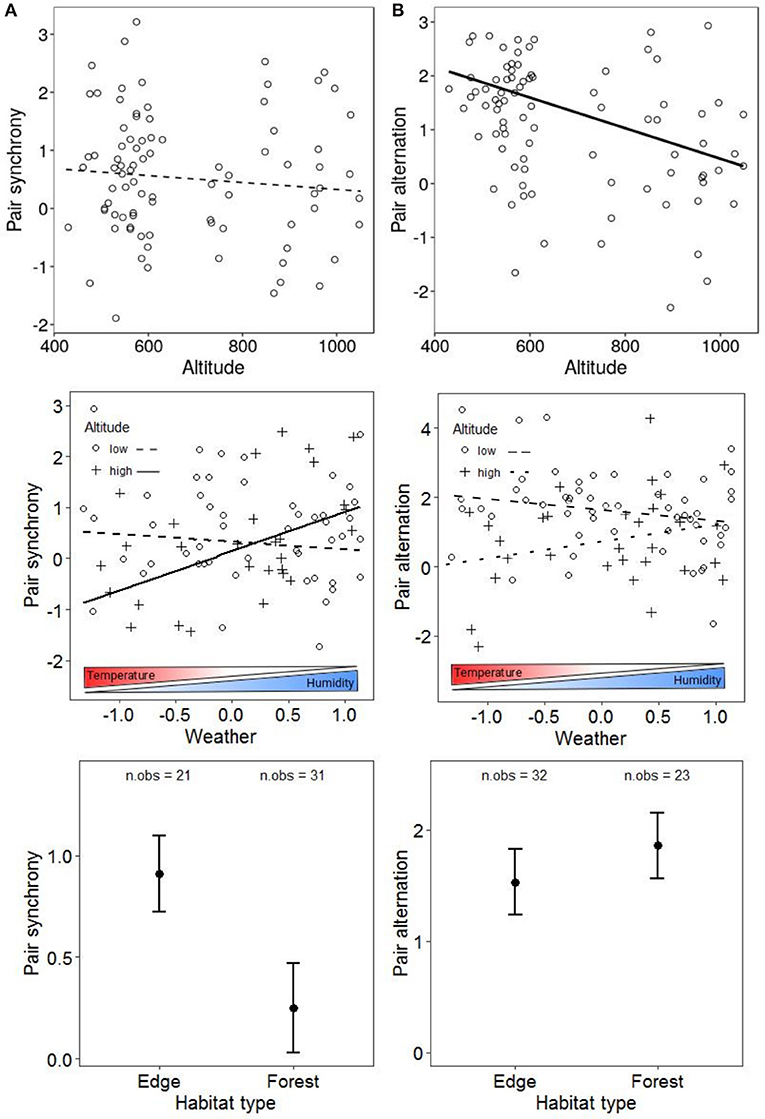
Figure 2. Pair synchrony (A) and pair alternation (B) in relation to altitude, habitat type, and weather. Lines show the predictions of the models (plain when significant effect, dotted if non-significant) and circles are observed data. Black points are predicted means and error bars are standard errors. Weather and altitude effects are from the analysis with the whole dataset (No. of obs/No. of pairs = 87/71 for synchrony, 91/73 for alternation), habitat type effect is from the analysis with the low altitude nests only (No. of obs/No. of pairs = 52/45 for synchrony and 55/46 for alternation).
The primary ecological predictor of nest visit alternation was altitude, with a 23% reduction in the degree of alternation for every 100 m increase in altitude. The magnitude of the relationship between weather and alternation tended to be modified by altitude but was non-significant at both low (estimate = −0.31, t = −1.48) and high altitude (estimate = 0.53, t = 1.76, Figure 2B). However, unlike with synchrony, alternation showed a non-significant tendency to be higher within woodland habitats than on woodland edges (Table 2, Figure 2B).
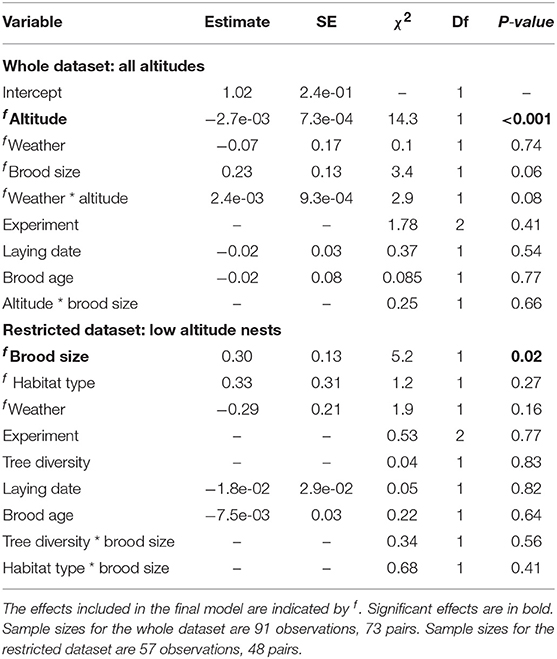
Table 2. Results of a LMM examining alternation.
The only significant ecological predictor of inter-visit interval was altitude which had a negative effect (Table 3). Pairs at high altitude fed significantly less frequently than pairs at low altitude, with a decrease of 5% for each 100 m (Table 3, Figure 3A). Parents fed slightly more frequently in habitats with high tree diversity but this effect was not significant (Table 3, Figure 3A).
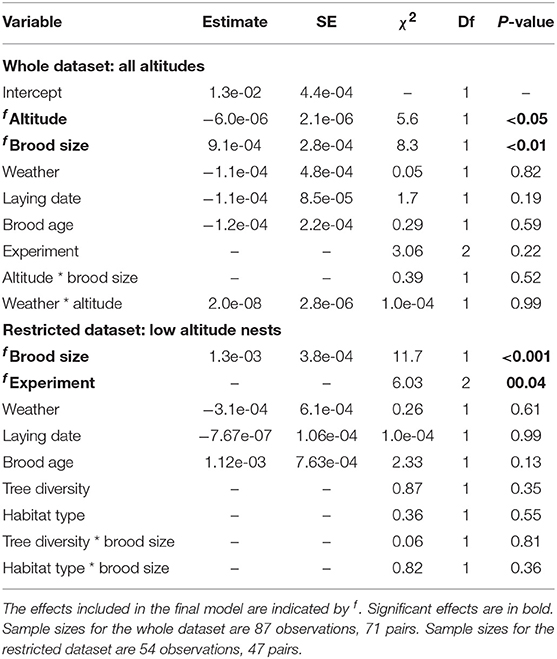
Table 3. Results of a LMM examining IVI, after reciprocal transformation.
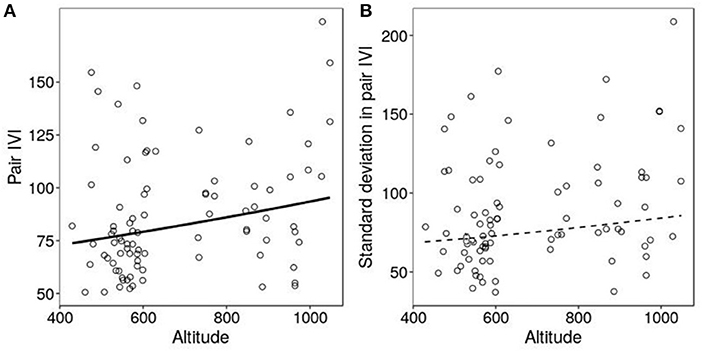
Figure 3. Pair mean inter-visists interval (A) and standard deviation in inter-visits interval (B) in relation to altitude. Lines show the predictions of the models (plain when significant effect, dotted if non-significant) and circles are observed data. Analysis were performed on 87 observations, from 71 pairs.
No ecological effects were correlated with the standard deviation in IVI. Only brood size was significantly correlated with it, with parents that had bigger brood sizes having less variation in IVI (Table 4, Figure 3B).
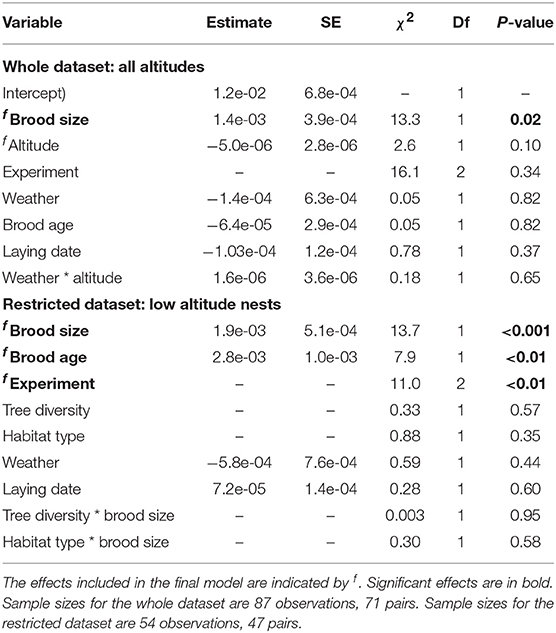
Table 4. Results of a LMM examining standard deviation in IVI, after reciprocal transformation.
The number of fledglings varied from 0 to 9 (mean = 6 ± SD = 1.7), with an average fledging success of 85% of chicks hatching (range = 5–9, mean = 7 ± SD = 1.5). Variation in the number of fledglings was not predicted by any of the feeding behaviour and ecological variables tested (Table 5, Figure 4A).
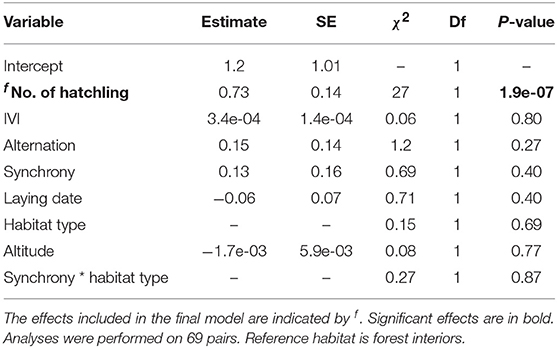
Table 5. Results of a LMM examining the number of fledglings.
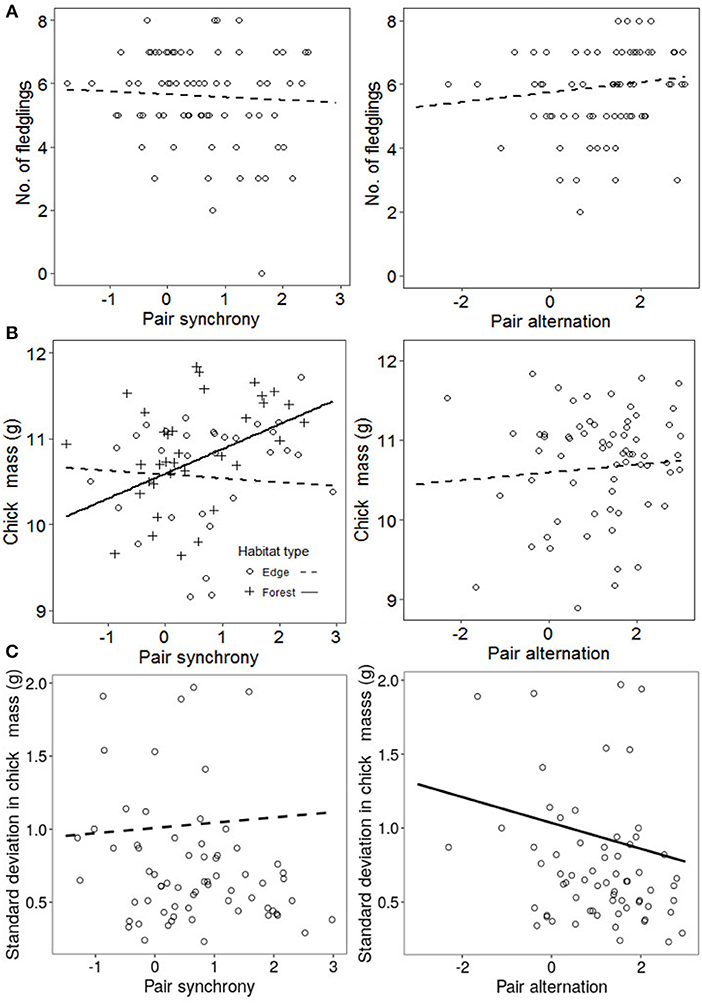
Figure 4. No. of fledglings (A), mean nestling mass (B) and standard deviation in nestling mass (C) in relation to pairs alternation and synchrony. Lines show the predictions of the models (plain when significant effect, dashed if non-significant) and points are observed data. Analyses were performed on 69 pairs.
The average mass of nestlings in broods on days 14–16 varied from 8.7 to 11.8 g (mean = 10.6 g ± SD = 0.77 g). After controlling for the significant effects of tarsus, we found a significant interaction between nest synchrony and habitat, with chick mass increased in more synchronous nests in woodland (estimate = 0.28, t = 2.7) but not edge habitats (estimate = −0.04, t = −0.42, Figure 4B). However, we found no effect of alternation on brood mass (Table 6, Figure 4B).
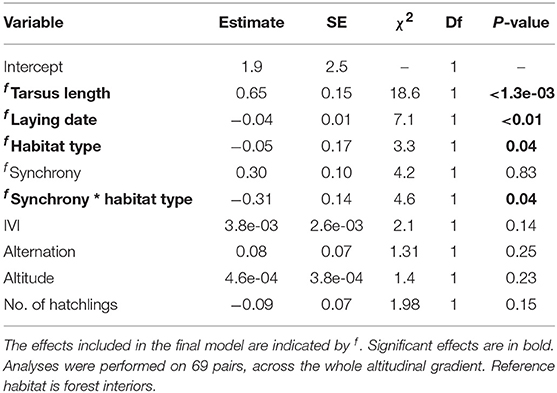
Table 6. Results of a LMM examining mean fledging mass in a brood.
The standard deviation of within brood nestling mass near fledging ranged from 0.2 to 1.9 g. After controlling for the significant effects of the average mass and the IVI, we found a significant relationship between alternation and the standard deviation in chick mass, with greater homogeneity of brood mass in pairs where parents alternated more (Table 7, Figure 4C). Broods were more heterogeneous in mass in forest edges but there was no relationship between synchrony and the standard deviation in chick mass, regardless of habitat (Table 7, Figure 4C).
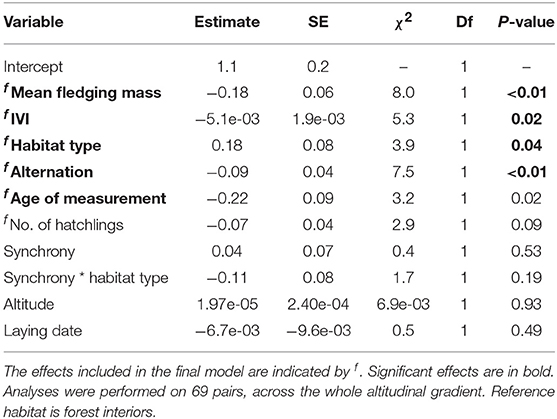
Table 7. Results of a LMM examining standard deviation in fledging mass in a brood.
Habitat characteristics can have an important impact on avian life histories including parental care (Martin, 1995) and breeding performance (Suorsa et al., 2004; Arriero et al., 2006) through its effect on predation and food availability (e.g., Zanette et al., 2000; Suorsa et al., 2004), yet studies of bi-parental care have rarely explored how such ecological variation influences parental visitation patterns. We found that Pyrenean blue tit pairs synchronised and alternated their nest visits much more often than expected by chance given their visit intervals and these patterns were influenced by ecology. In particular, nest visits were more synchronous in edge habitats, and parents synchronised more at high elevations on cold and wet days. The fact that ecology has contrasting effects on the patterns of nest visit synchrony and turn-taking suggests that the two coordination behaviours are largely influenced by different factors and may serve different functions in bi-parental care. Similarly, while inter-visit intervals decreased with altitude, variance in inter-visit interval was not influenced by any ecological factors we measured. Finally, variation in patterns of parental care had only limited effects on fitness as patterns of care were unrelated to fledgling success despite more synchronous parents within woodlands producing heavier young and pairs that alternated more had a lower variance in within brood mass. Overall, our results suggest that ecology can indeed influence the amount of parental care provided and also patterns of care visits, and that the interplay between environment and coordination behaviours may impact chick growth, but not the number of fledglings.
Increases in elevation coincide with changes in many critical ecological variables and, as a result, elevational gradients have been used to better understand the effects of ecology on parental care. Cooler temperatures, shorter breeding seasons and lower food availability at high altitude are often associated with lower annual fecundity (Badyaev, 1997; Sandercock et al., 2005; Bears et al., 2008; Boyle et al., 2016) and longer post-hatching care (Badyaev, 1997; Badyaev and Ghalambor, 2001), perhaps to compensate for a lower quantity or quality of food (Schöll et al., 2016). Indeed, dwarf hamsters have maternal-only care at low altitude, but bi-parental care at high altitude (Wynne-Edwards, 1998). Such increased costs of reproduction related to ecology could influence sexual conflict. Johnstone et al. (2014) argued that the costs of sexual conflict can be reduced through conditional cooperation (Keser and Van Winden, 2000; Gächter, 2006), where individuals refuse to come and feed the brood until their partner has fed the young. In this case theory predicts that alternation of feeding trips (i.e., conditional cooperation) would result in a higher total parental investment, which is closer to the optimal feeding rate that maximises the fitness of both parents (Johnstone et al., 2014; Johnstone and Savage, 2019). Therefore, under harsher care conditions expected at high altitude and assuming costs of waiting are not excessively high, we would expect to see increased parental coordination. However, we found the opposite result in our population: pairs that breed at low altitudes feed and alternate significantly more than couples that breed at high altitudes. Our results could be explained by environmental constraints at high elevations; with low food availability in these habitats (Abdusalyamov, 1964; Kovshar, 1981; Schöll et al., 2016) parents may have difficulty finding food and therefore can not afford to adopt a coordination strategy. The increased variance of the IVI at high altitude confirms the idea that there may be less regularity in the success of foraging, as can be expected with increasing difficulty finding food. Further, pairs are not likely to compensate for prey scarcity by prioritising high quality prey items at high altitude, as high quality caterpillar prey are rarely provided in our population (around 10% of feeding visits; Bründl et al., 2019).
Many different ecological factors change along altitudinal gradients and examining each factor separately may help us better understand which ecological parameters are most likely to affect the different components of parental coordination. Low temperatures can reduce food activity and caterpillar growth, resulting in low food abundance (Topp and Kirsten, 1991; Ayres, 1993; Schöll et al., 2016). Likewise, persistent rainfall negatively affects caterpillar abundance during cold period due to increased risk of caterpillar infections and diseases (Dennis and Sparks, 2007; Tamburini et al., 2013) and reduced arthropod movement (Tamburini et al., 2013), resulting in fewer caterpillar hatching or surviving (see study by Bale et al., 2002; Schöll et al., 2016). We found no effect of weather on the mean IVI and standard deviation in IVI, but weather did have a complex influence on coordination behaviour. Synchrony between parents increased on colder and wetter days, but only at higher elevation, and alternation had the same tendency, suggesting that parental coordination behaviour does increase under more extreme weather conditions. Birds living at higher elevations may in general be more sensitive to meteorological conditions and their impact on food availability. Under harsh conditions it could be advantageous to monitor partner's investment, as this ensures that offspring are fed sufficiently. Synchrony at the nest may also be due to synchrony of foraging, possibly improving the probability of finding food in patchy environments expected in harsh environments (Mariette and Griffith, 2012, 2015). Short-term changes in weather might also cause correlated changes in the visit rates of both parents, which are not accounted for by randomisations, and hence are included in coordination scores (Ihle et al., 2019; Santema et al., 2019).
In addition to weather, habitat tree diversity may also influence the quantity and quality of food available which in turn could impact patterns of parental feeding. Birds in high quality habitats travel shorter distances between nest visits, which may increase feeding rates (European starlings Sturnus vulgaris: Wright et al., 1998; blue tits C. caeruleus: Tremblay et al., 2005) and increase fledging mass (Santema et al., 2019). Tree species richness is positively related to the abundance and diversity of certain insect groups in temperate forests (Gering and Crist, 2000; Sobek et al., 2009). Also, the presence of tree species differing in growth rates and foliage structure may result in less horizontal canopy space used and, thus, a more open canopy, making prey more visible (Lang et al., 2012; Muiruri et al., 2016) and then reduce search time (Arvidsson and Klaesson, 1986; Mason, 1997). However, we found no effect of tree diversity on patterns of feeding behaviour. The lack of an effect on the feeding rate suggests that parents feed their offspring at the same rate in habitats of low trees diversity, where food is assumed to be less abundant so that adults would have to compensate for the decline in local food abundance by increasing their foraging distances. It is possible that an effect of habitat diversity on feeding rate is masked by an effect on prey quality if parents in poorer quality habitats are less selective in the type of prey they choose (Bańbura et al., 1994) in order to feed at similar rates. Interestingly, some studies have shown that the effect of plant species richness on herbivore predators abundance and richness was not strong, especially when phylogenetic diversity of plant species was low (Dinnage et al., 2012). Direct measurements of food abundance and analysis of the type of prey brought to the nest would allow us to better disentangle the effects of forest diversity and insect abundance on patterns of parental care.
Edges are defined as the interface between the relatively stable environment of the forest interior and the highly variable external environment (Saunders et al., 1991; Wilkin et al., 2007). They can provide a great diversity of resources much like mixed forests although their impact on within-species reproductive output is mixed (Murcia, 1995; Lahti, 2001; Wilkin et al., 2007, 2009). Reduced reproductive success could result from increased exposure to predation in edge communities (Wilcove et al., 1986; Andrén and Anglestam, 1988; Hartley and Hunter, 1998; Batàry and Bàldi, 2004). It has been shown that feeding rate was plastic in response to predation and that some birds reduce their feeding rate when exposed to predation at the nest (Eggers et al., 2008; Peluc et al., 2008; Ghalambor et al., 2013). We expect that increased predation risk in open habitats at forest edges could favour increased synchronisation of parents when visiting the nest to reduce the exposure to predators and potentially the risk of predation to parents (Raihani et al., 2010). Indeed, pairs nesting on the edge were significantly more synchronous than pairs nesting in the forest interior as expected if predation risk is higher in ecotones. However, edge habitats also provide a broader diversity of food if individuals forage in both habitats (woodland and pastures) (Huhta et al., 1999) or decreased food if they only forage in one habitat which each could alter patterns of care. Increased synchronisation of nest visits could allow parents to better distribute food among chicks (Shen et al., 2010) or may result from adults feeding together to increase foraging efficiency (Ward and Zahavi, 1973; Beauchamp, 1998; Mariette and Griffith, 2012, 2015). Since tits rarely forage in open habitats (Perrins, 1991), we expected that such edge habitats would have less food available relative to the forest interior which, as for higher predation risk, should increase synchrony. Direct measures of both predation risk and food abundance in edges relative to forest interiors would help to distinguish between these two alternative explanations for changes in synchrony of parental care. It should be noted, however, that we considered the nests to be on the edge when they were located in the forest, up to 5 meters away from open habitats. Most researchers have found that the edge effects on species presence and success persists up to 150m into forest fragments (Laurance and Cochrane, 2001). It is therefore possible that our definition has biased the results by softening the contrasts between what is considered to be interiors and edges. Defining the edge by environmental factors such as light, height, density, and vegetation diversity could provide a better understanding of the edge effect on parental care (Paton, 1994; Batàry and Bàldi, 2004).
While we found considerable variation in the degree of alternation and synchrony both between pairs and across environments, these parental strategies were less clearly tied to increases in offspring fitness. Indeed, there was no link between fledging success and parental coordination behaviours in our population. These results contrast with other studies that have found a relationship between alternation or synchrony and fledging success (Raihani et al., 2010; Bebbington and Hatchwell, 2015). Patterns of parental care did, however, have more subtle effects on chick mass which could influence post-fledging success. Both a higher feeding rate and increased synchrony at nests in woodland interiors was associated with larger chicks on average, but had no effect on the within-brood variance in chick mass. In contrast, alternation was unrelated to average chick mass, but was positively related to the variance in chick mass, suggesting that this behaviour may be involved in a better distribution of food within brood. Other studies show mixed results for the impact of parental coordination behaviours on chick mass. Synchrony seems to influence both chick mass and the distribution of food to nestlings in zebra finches (Mariette and Griffith, 2015) but how synchrony influences average chick mass but not variance in chick mass in our population remains unclear. Likewise, why alternation of parents per seinfluences variance in chick mass above and beyond feeding rate remains unclear but could result from differences in feeding rules between parents (Lessells, 2002; Shizuka and Lyon, 2013). Weather a relationship between coordination behaviours and other measures of offspring fitness such as post-fledging survival and recruitment remains to be determined. If patterns of parental care have a greater influence on chick condition than fledging success, evaluating the fitness impacts of parental coordination behaviours would require following broods through recruitment.
Overall, our results suggest that blue tits have “active” alternation and synchrony when provisioning nestlings. Ecological conditions are related to the different feeding behaviour parameters we studied, but different ecological conditions affect each one, suggesting that alternation and synchrony may serve different functions in bi-parental care. The finding of a positive relationship between synchrony at the nest and the nestling mass in woodland habitats supports the contention that coordination can mitigate the costs of sexual conflict for offspring. Taken together, these results suggest that other studies of parental care patterns should likewise examine the interaction between the environment and coordinative behaviour for feeding offspring. Experimental studies will be needed to study how this interaction affects the outcome of sexual conflict and these will require short-term manipulations of offspring demand or parental costs to tease apart the potential drivers of parental behaviour.
The datasets analysed for this study can be found in the figshare repository here: https://figshare.com/articles/pcare_csv/8153762.
The animal study was reviewed and approved by Birds capture was carried out under permits to AC from the French bird ringing office (CRBPO; program 576) and breeding monitoring under permits from the state of Ariége (Préfecture de l'Ariége, Protection des Populations, no A09-4) and the Région Midi-Pyrenées (DIREN, no 2013-02).
LL, AC, JS, and AR conceived research. AT, AB, AC, and LL coordinated and performed the fieldwork. LL and AB coordinated the video analyses. JS created the method to estimate the feeding behaviour parameters. LL compiled the data and performed the statistical analyses. LL wrote the manuscript with important contributions from AC, AR, and JS. All coauthors contributed to revisions, gave final approval for publication and agree to be held accountable for the work performed therein.
This work was supported by The Region Midi-Pyrenees grants to AC and AR, a MESR (Ministére de l'Enseignement supérieur et de la Recherche) Ph.D. scholarship to LL, fellowship grants from the Région Midi-Pyrénées and the Centre national de la Recherche scientifique and a Natural Environment Research Council to the University of Exeter to AB, an IRC Government of Ireland Postdoctoral Fellowship and Ulysses grant to JS, and ANR-JCJC Net- Select and HFSP RGP0006/2015 grants to AC. This work was supported by the Laboratoire d'Excellence (LABEX) TULIP (ANR-10-LABX-41) and IAST through ANR grant ANR-17-EURE-0010 (Investissements d'Avenir program).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are very grateful to the following interns for helping with data collection field and from video recordings: Amandine Sieper, Kilian David, Laurie Abou-Cajal, Aourell Lanfrey, Kiara L'herpinière, Meidhi Khelifi, Baptiste Averly, Louis Bliard, Maeliss Hoarau, Virginie Mercier, Shirley Laurent, Aurélie Nouri, Sebastian Bekker, Purabi Deshpande, Christel Blot, Pauline Clin, Quentin Carbonnier, Clément Rul, Robin Bhattacharyya-Dickson, Madeleine Barr, Manon Broadribb, Amy Gresham, Ben Murphy, Sophie Porta, Solenn Auquiére, Nils Serventis, Camille Paillard, Lauréne Michel, Émeline Garnier, Jessica Mulvey, Murielle Vergniol, Benjamin Begou, Thomas Vulvin, and Noé Rehspringer. We would like to thank the two reviewers Kat Bebbington and Amelie Fargevieille for their comments and suggestions that greatly improved the quality of this manuscript.
Andrén, H., and Anglestam, P. (1988). Elevated predation rates as an edge effect in habitat islands: experimental evidence. Ecology 69, 544–547. doi: 10.2307/1940455
Arlettaz, R., Schaad, M., Reichlin, T. S., and Schaub, M. (2010). Impact of weather and climate variation on hoopoe reproductive ecology and population growth. J. Ornithol. 151, 889–899. doi: 10.1007/s10336-010-0527-7
Arriero, E., Sanz, J. J., and Romero-Pujante, M. (2006). Habitat structure in mediterranean deciduous oak forests in relation to reproductive success in the blue tit parus caeruleus. Bird Study 53, 12–19. doi: 10.1080/00063650609461411
Arvidsson, B., and Klaesson, P. (1986). Territory size in a willow warbler phylloscopus trochilus population in mountain birch forest in Swedish lapland. Ornis Scand. 17, 24–30. doi: 10.2307/3676749
Axelrod, R., and Hamilton, W. D. (1981). The evolution of cooperation. Science 211, 1390–1396. doi: 10.1126/science.7466396
Ayres, M. P. (1993). “Plant defense, herbivory, and climate change,” in Biotic Interactions and Global Change, eds P. M. Kareiva, J. G. Kingsolver, and R. B. Huey (Sunderland, MA: Sinauer Associates Inc.), 75–94.
Badyaev, A. V. (1997). Avian life history variation along altitudinal gradients: an example with cardueline finches. Oecologia 111, 365–374. doi: 10.1007/s004420050247
Badyaev, A. V., and Ghalambor, C. K. (2001). Evolution of life histories along elevational gradients: trade-off between parental care and fecundity. Ecology 82, 2948–2960. doi: 10.1890/0012-9658(2001)082[2948:EOLHAE]2.0.CO;2
Bale, J. S., Masters, G. J., Hodkinson, I. D., Awmack, C., Bezemer, T. M., Brown, V. K., et al. (2002). Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Global Change Biol. 8, 1–16. doi: 10.1046/j.1365-2486.2002.00451.x
Bańbura, J., Blondel, J., de Wilde-Lambrechts, H., Galan, M.-J., and Maistre, M. (1994). Nestling diet variation in an insular mediterranean population of blue tits parus caeruleus: effects of years, territories and individuals. Oecologia 100, 413–420. doi: 10.1007/BF00317863
Batàry, P., and Bàldi, A. (2004). Evidence of an edge effect on avian nest success. Conserv. Biol. 18, 389–400. doi: 10.1111/j.1523-1739.2004.00184.x
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Bears, H., C. Drever, M., and Martin, K. (2008). Comparative morphology of dark-eyed juncos junco hyemalis breeding at two elevations: a common aviary experiment. J. Avian Biol. 39, 152–162. doi: 10.1111/j.2008.0908-8857.04191.x
Beauchamp, G. (1998). The effect of group size on mean food intake rate in birds. Biol. Rev. 73, 449–472. doi: 10.1017/S0006323198005246
Bebbington, K., and Hatchwell, B. J. (2015). Coordinated parental provisioning is related to feeding rate and reproductive success in a songbird. Behav. Ecol. 27, 652–659. doi: 10.1093/beheco/arv198
Blondel, J., Dervieux, A., Maistre, M., and Perret, P. (1991). Feeding ecology and life history variation of the blue tit in mediterranean deciduous and sclerophyllous habitats. Oecologia 88, 9–14. doi: 10.1007/BF00328397
Boyle, A. W., Sandercock, B. K., and Martin, K. (2016). Patterns and drivers of intraspecific variation in avian life history along elevational gradients: a meta-analysis. Biol. Rev. 91, 469–482. doi: 10.1111/brv.12180
Bründl, A. C. (2018). Parental investment across an altitudinal gradient in blue tits (Cyanistes caeruleus) (Ph.D. thesis). Universities of Exeter and Toulouse Paul Sabatier III, Falmouth, Cornwall, United Kingdom.
Bründl, A. C., Sorato, E., Sallé, L., Thiney, A. C., Kaulbarsch, S., Chaine, A. S., et al. (2019). Experimentally induced increases in fecundity lead to greater nestling care in blue tits. Proc. R. Soc. B. 286:20191013. doi: 10.1098/rspb.2019.1013
Dennis, R. L., and Sparks, T. H. (2007). Climate signals are reflected in an 89 year series of british lepidoptera records. Eur. J. Entomol. 104:763. doi: 10.14411/eje.2007.097
Dinnage, R., Cadotte, M. W., Haddad, N. M., Crutsinger, G. M., and Tilman, D. (2012). Diversity of plant evolutionary lineages promotes arthropod diversity. Ecol. Lett. 15, 1308–1317. doi: 10.1111/j.1461-0248.2012.01854.x
Eggers, S., Griesser, M., and Ekman, J. (2008). Predator-induced reductions in nest visitation rates are modified by forest cover and food availability. Behav. Ecol. 19, 1056–1062. doi: 10.1093/beheco/arn063
Ewald, C.-O., McNamara, J., and Houston, A. (2007). Parental care as a differential game: A dynamic extension of the houston–davies game. Appl. Math. Comput. 190, 1450–1465. doi: 10.1016/j.amc.2007.02.060
Fox, J., and Monette, G. (1992). Generalized collinearity diagnostics. J. Am. Statist. Assoc. 87, 178–183. doi: 10.1080/01621459.1992.10475190
Gächter, S. (2006). Conditional Cooperation: Behavioral Regularities from the Lab and the Field and Their Policy Implications. Technical report, CeDEx Discussion Paper Series.
Gering, J. C., and Crist, T. O. (2000). Patterns of beetle (coleoptera) diversity in crowns of representative tree species in an old-growth temperate deciduous forest. Selbyana 21, 38–47.
Ghalambor, C. K., Peluc, S. I., and Martin, T. E. (2013). Plasticity of parental care under the risk of predation: how much should parents reduce care? Biol. Lett. 9:20130154. doi: 10.1098/rsbl.2013.0154
Gray, C. M., and Hamer, K. C. (2001). Food-provisioning behaviour of male and female manx shearwaters, puffinus puffinus. Ani. Behav. 62, 117–121. doi: 10.1006/anbe.2001.1717
Harrison, F., Barta, Z., Cuthill, I., and Szekely, T. (2009). How is sexual conflict over parental care resolved? a meta-analysis. J. Evol. Biol. 22, 1800–1812. doi: 10.1111/j.1420-9101.2009.01792.x
Hartley, M. J., and Hunter, M. L. (1998). A meta-analysis of forest cover, edge effects, and artificial nest predation rates. Conserv. Biol. 12, 465–469. doi: 10.1046/j.1523-1739.1998.96373.x
Houston, A. I., and Davies, N. B. (1985). “The evolution of cooperation and life history in the dunnock, Prunella modularis,” in Behavioural Ecology: The Ecological Consequences of Adaptive Behaviour, eds R. M. Sibly and R. H. Smith (Oxford: Blackwell Scientific), 471–487.
Houston, A. I., Székely, T., and McNamara, J. M. (2005). Conflict between parents over care. Trends Ecol. Evol. 20, 33–38. doi: 10.1016/j.tree.2004.10.008
Huhta, E., Jokimäki, J., and Rahko, P. (1999). Breeding success of pied flycatchers in artificial forest edges: the effect of a suboptimally shaped foraging area. Auk 116, 528–535. doi: 10.2307/4089385
Ihle, M., Pick, J. L., Winney, I. S., Nakagawa, S., and Burke, T. (2019). Measuring up to reality: null models and analysis simulations to study parental coordination over provisioning offspring. Front. Ecol. Evol. 7:142. doi: 10.3389/fevo.2019.00142
Johnson, L. S., Brubaker, J. L., Ostlind, E., and Balenger, S. L. (2007). Effect of altitude on male parental expenditure in mountain bluebirds (sialia currucoides): Are higher-altitude males more attentive fathers? J. Ornithol. 148, 9–16. doi: 10.1007/s10336-006-0092-2
Johnstone, R. A., and Hinde, C. A. (2006). Negotiation over offspring care—how should parents respond to each other's efforts? Behav. Ecol. 17, 818–827. doi: 10.1093/beheco/arl009
Johnstone, R. A., Manica, A., Fayet, A. L., Stoddard, M. C., Rodriguez-Gironés, M. A., and Hinde, C. A. (2014). Reciprocity and conditional cooperation between great tit parents. Behav. Ecol. 25, 216–222. doi: 10.1093/beheco/art109
Johnstone, R. A., and Savage, J. L. (2019). Conditional cooperation and turn-taking in parental care. Front. Ecol. Evol. 7:335. doi: 10.3389/fevo.2019.00335
Jones, K. M., Ruxton, G. D., and Monaghan, P. (2002). Model parents: is full compensation for reduced partner nest attendance compatible with stable biparental care? Behav. Ecol. 13, 838–843. doi: 10.1093/beheco/13.6.838
Keser, C., and Van Winden, F. (2000). Conditional cooperation and voluntary contributions to public goods. Scand. J. Econ. 102, 23–39. doi: 10.1111/1467-9442.00182
Kovshar, A. (1981). Peculiarities of the bird reproduction in subalpine: on the material of passeriformes in the tien shan. Acad. Sci. Kazakh SSR Alma-Ata.
Krebs, E. A., Cunningham, R. B., and Donnelly, C. F. (1999). Complex patterns of food allocation in asynchronously hatching broods of crimson rosellas. Anim. Behav. 57, 753–763. doi: 10.1006/anbe.1998.1029
Lahti, D. (2001). The “edge effect on nest predation” hypothesis after twenty years. Biol. Conserv. 99, 365–374. doi: 10.1016/S0006-3207(00)00222-6
Lang, A. C., Härdtle, W., Bruelheide, H., Kröber, W., Schröter, M., von Wehrden, H., et al. (2012). Horizontal, but not vertical canopy structure is related to stand functional diversity in a subtropical slope forest. Ecol. Res. 27, 181–189. doi: 10.1007/s11284-011-0887-3
Laurance, W. F., and Cochrane, M. A. (2001). Synergistic effects in fragmented landscapes. Conserv. Biol.
Lee, J.-W., Kim, H.-Y., and Hatchwell, B. J. (2010). Parental provisioning behaviour in a flock-living passerine, the vinous-throated parrotbill paradoxornis webbianus. J. Ornithol. 151, 483–490. doi: 10.1007/s10336-009-0484-1
Lee, J. K., Chung, O.-S., and Lee, W.-S. (2011). Altitudinal variation in parental provisioning of nestling varied tits (poecile varius). Wilson J. Ornithol. 123, 283–288. doi: 10.1676/10-106.1
Lessells, C. (2002). Parentally biased favouritism: why should parents specialize in caring for different offspring? Philos. Trans. R. Soc. Lond. B Biol. Sci. 357, 381–403. doi: 10.1098/rstb.2001.0928
Lessells, C., and McNamara, J. M. (2011). Sexual conflict over parental investment in repeated bouts: negotiation reduces overall care. Proc. R. Soc. B Biol. Sci. 279, 1506–1514. doi: 10.1098/rspb.2011.1690
Lessells, C. M. (2013). “Sexual conflict,” in The Evolution of Parental Care, eds N. J. Royle, P. T. Smiseth, and M. Kölliker (Oxford, UK: Oxford University Press).
Mariette, M. M., and Griffith, S. C. (2012). Nest visit synchrony is high and correlates with reproductive success in the wild zebra finch taeniopygia guttata. J. Avian Biol. 43, 131–140. doi: 10.2307/41477965
Mariette, M. M., and Griffith, S. C. (2015). The adaptive significance of provisioning and foraging coordination between breeding partners. Am. Nat. 185, 270–280. doi: 10.1111/j.1600-048X.2012.05555.x
Martin, T. E. (1995). Avian life history evolution in relation to nest sites, nest predation, and food. Ecol. Monogr. 65, 101–127. doi: 10.2307/2937160
Masello, J. F., Pagnossin, M. L., Sommer, C., and Quillfeldt, P. (2006). Population size, provisioning frequency, flock size and foraging range at the largest known colony of psittaciformes: the burrowing parrots of the north-eastern patagonian coastal cliffs. Emu-Austral Ornithol. 106, 69–79. doi: 10.1071/MU04047
Mason, C. (1997). Association between willow warbler phylloscopus trochilus territories and birch in woodlands in southeastern england. Ibis 139, 411–412. doi: 10.1111/j.1474-919X.1997.tb04648.x
Mathot, K. J., Olsen, A.-L., Mutzel, A., Araya-Ajoy, Y. G., Nicolaus, M., Westneat, D. F., et al. (2017). Provisioning tactics of great tits (parus major) in response to long-term brood size manipulations differ across years. Behav. Ecol. 28, 1402–1413. doi: 10.1093/beheco/arx083
McDonald, P. G., Te Marvelde, L., Kazem, A. J., and Wright, J. (2008). Helping as a signal and the effect of a potential audience during provisioning visits in a cooperative bird. Anim. Behav. 75, 1319–1330. doi: 10.1016/j.anbehav.2007.09.005
McNamara, J. M., Gasson, C. E., and Houston, A. I. (1999). Incorporating rules for responding into evolutionary games. Nature 401:368. doi: 10.1038/43869
McNamara, J. M., Houston, A. I., Barta, Z., and Osorno, J.-L. (2003). Should young ever be better off with one parent than with two? Behav. Ecol. 14, 301–310. doi: 10.1093/beheco/14.3.301
Meade, J., Nam, K.-B., Lee, J.-W., and Hatchwell, B. J. (2011). An experimental test of the information model for negotiation of biparental care. PLoS ONE 6:e19684. doi: 10.1371/journal.pone.0019684
Muiruri, E. W., Rainio, K., and Koricheva, J. (2016). Do birds see the forest for the trees? scale-dependent effects of tree diversity on avian predation of artificial larvae. Oecologia 180, 619–630. doi: 10.1007/s00442-015-3391-6
Murcia, C. (1995). Edge effects in fragmented forests: implications for conservation. Trends Ecol. Evol. 10, 58–62. doi: 10.1016/S0169-5347(00)88977-6
Naef-Daenzer, L., Naef-Daenzer, B., and Nager, R. G. (2000). Prey selection and foraging performance of breeding great tits parus major in relation to food availability. J. Avian Biol. 31, 206–214. doi: 10.1034/j.1600-048X.2000.310212.x
Nur, N. (1984). The consequences of brood size for breeding blue tits ii. nestling weight, offspring survival and optimal brood size. J. Anim. Ecol. 53, 497–517. doi: 10.2307/4530
Paton, P. W. (1994). The effect of edge on avian nest success: how strong is the evidence? Conserv. Biol. 8, 17–26. doi: 10.1046/j.1523-1739.1994.08010017.x
Peluc, S. I., Sillett, T. S., Rotenberry, J. T., and Ghalambor, C. K. (2008). Adaptive phenotypic plasticity in an island songbird exposed to a novel predation risk. Behav. Ecol. 19, 830–835.
Perrins, C. (1991). Tits and their caterpillar food supply. Ibis 133, 49–54. doi: 10.1111/j.1474-919X.1991.tb07668.x
R Core Team (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Raihani, N. J., Nelson-Flower, M. J., Moyes, K., Browning, L. E., and Ridley, A. R. (2010). Synchronous provisioning increases brood survival in cooperatively breeding pied babblers. J. Anim. Ecol. 79, 44–52. doi: 10.1111/j.1365-2656.2009.01606.x
Sandercock, B. K., Martin, K., and Hannon, S. J. (2005). Life history strategies in extreme environments: comparative demography of arctic and alpine ptarmigan. Ecology 86, 2176–2186. doi: 10.1890/04-0563
Santema, P., Schlicht, E., and Kempenaers, B. (2019). Testing the conditional cooperation model: What can we learn from parents taking turns when feeding offspring? Front. Ecol. Evol. 7:94. doi: 10.3389/fevo.2019.00094
Saunders, D. A., Hobbs, R. J., and Margules, C. R. (1991). Biological consequences of ecosystem fragmentation: a review. Conserv. Biol. 5, 18–32. doi: 10.1111/j.1523-1739.1991.tb00384.x
Savage, J. L., Browning, L. E., Manica, A., Russell, A. F., and Johnstone, R. A. (2017). Turn-taking in cooperative offspring care: by-product of individual provisioning behavior or active response rule? Behav. Ecol. Sociobiol. 71:162. doi: 10.1007/s00265-017-2391-4
Savage, J. L., and Ouyang, J. Q. (2019). Short communication: Coordinated great tit (parus major) parents have more similar hormone levels. NA.
Schöll, E. M., Ohm, J., Hoffmann, K. F., and Hille, S. M. (2016). Caterpillar biomass depends on temperature and precipitation, but does not affect bird reproduction. Acta Oecol. 74, 28–36. doi: 10.1016/j.actao.2016.06.004
Shen, S.-F., Chen, H.-C., Vehrencamp, S. L., and Yuan, H.-W. (2010). Group provisioning limits sharing conflict among nestlings in joint-nesting taiwan yuhinas. Biol. Lett. 6, 318–321. doi: 10.1098/rsbl.2009.0909
Shizuka, D., and Lyon, B. E. (2013). Family dynamics through time: brood reduction followed by parental compensation with aggression and favouritism. Ecol. Lett. 16, 315–322. doi: 10.1111/ele.12040
Sobek, S., Tscharntke, T., Scherber, C., Schiele, S., and Steffan-Dewenter, I. (2009). Canopy vs. understory: Does tree diversity affect bee and wasp communities and their natural enemies across forest strata? Forest Ecol. Manage. 258, 609–615. doi: 10.1016/j.foreco.2009.04.026
Stauss, M. J., Burkhardt, J. F., and Tomiuk, J. (2005). Foraging flight distances as a measure of parental effort in blue tits parus caeruleus differ with environmental conditions. J. Avian Biol. 36, 47–56. doi: 10.1111/j.0908-8857.2005.02855.x
Suorsa, P., Helle, H., Koivunen, V., Huhta, E., Nikula, A., and Hakkarainen, H. (2004). Effects of forest patch size on physiological stress and immunocompetence in an area-sensitive passerine, the eurasian treecreeper (certhia familiaris): an experiment. Proc. R. Soc. Lond. B Biol. Sci. 271, 435–440. doi: 10.1098/rspb.2003.2620
Tamburini, G., Marini, L., Hellrigl, K., Salvadori, C., and Battisti, A. (2013). Effects of climate and density-dependent factors on population dynamics of the pine processionary moth in the southern alps. Clim. Change 121, 701–712. doi: 10.1007/s10584-013-0966-2
Topp, W., and Kirsten, K. (1991). Synchronisation of pre-imaginal development and reproductive success in the winter moth, operophtera brumata l. J. Appl. Entomol. 111, 137–146. doi: 10.1111/j.1439-0418.1991.tb00304.x
Tremblay, I., Thomas, D., Blondel, J., Perret, P., and Lambrechts, M. M. (2005). The effect of habitat quality on foraging patterns, provisioning rate and nestling growth in corsican blue tits parus caeruleus. Ibis 147, 17–24. doi: 10.1111/j.1474-919x.2004.00312.x
Tremblay, I., Thomas, D. W., Lambrechts, M. M., Blondel, J., and Perret, P. (2003). Variation in blue tit breeding performance across gradients in habitat richness. Ecology 84, 3033–3043. doi: 10.1890/02-0663
Trivers, R. L. (1971). The evolution of reciprocal altruism. Q. Rev. Biol. 46, 35–57. doi: 10.1086/406755
Trivers, R. L. (1972). “Parental investment and sexual selection,” in Sexual Selection & the Descent of Man, 1871–1971, Vol. 136, ed B. Campbell (Chicago, IL: Aldine).
van Rooij, E. P., and Griffithc, S. C. (2013). Synchronised provisioning at the nest: parental coordination over care in a socially monogamous species. PeerJ. 1:e232. doi: 10.7717/peerj.232
Wald, A., and Wolfowitz, J. (1940). On a test whether two samples are from the same population. Ann. Math. Statist. 11, 147–162. doi: 10.1214/aoms/1177731909
Ward, P., and Zahavi, A. (1973). The importance of certain assemblages of birds as “information-centres” for food-finding. Ibis 115, 517–534. doi: 10.1111/j.1474-919X.1973.tb01990.x
Westneat, D. F., Schofield, M., and Wright, J. (2012). Parental behavior exhibits among-individual variance, plasticity, and heterogeneous residual variance. Behav. Ecol. 24, 598–604. doi: 10.1093/beheco/ars207
Wilcove, D. S., McLellan, C. H., and Dobson, A. P. (1986). Habitat fragmentation in the temperate zone. Conserv. Biol. 6, 237–256.
Wilkin, T. A., Garant, D., Gosler, A. G., and Sheldon, B. C. (2007). Edge effects in the great tit: analyses of long-term data with gis techniques. Conserv. Biol. 21, 1207–1217. doi: 10.1111/j.1523-1739.2007.00767.x
Wilkin, T. A., King, L. E., and Sheldon, B. C. (2009). Habitat quality, nestling diet, and provisioning behaviour in great tits Parus major. J. Avian Biol. 40, 135–145. doi: 10.1111/j.1600-048X.2009.04362.x
Wright, J., Both, C., Cotton, P., and Bryant, D. (1998). Quality vs. quantity: energetic and nutritional trade-offs in parental provisioning strategies. J. Anim. Ecol. 620–634.
Wynne-Edwards, K. E. (1998). Evolution of parental care in phodopus: conflict between adaptations for survival and adaptations for rapid reproduction. Am. Zool. 38, 238–250. doi: 10.1093/icb/38.1.238
Ydenberg, R. C. (2008). “Provisioning,” in Foraging: Behavior and Ecology, eds D. W. Stephens, J. L. Brown, and R. C. Ydenberg (Chicago, IL: University of Chicago Press), 273–303.
Keywords: alternation, bi-parental system, Cyanistes caeruleus, environmental variation, feeding behaviour, parental care, sexual conflict, synchrony
Citation: Lejeune L, Savage JL, Bründl AC, Thiney A, Russell AF and Chaine AS (2019) Environmental Effects on Parental Care Visitation Patterns in Blue Tits Cyanistes caeruleus. Front. Ecol. Evol. 7:356. doi: 10.3389/fevo.2019.00356
Received: 18 June 2019; Accepted: 09 September 2019;
Published: 24 September 2019.
Edited by:
Rita Covas, University of Porto, PortugalReviewed by:
Kat Bebbington, University of Groningen, NetherlandsCopyright © 2019 Lejeune, Savage, Bründl, Thiney, Russell and Chaine. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Léa Lejeune, bGVhQGxqbi5uYW1l
†ORCID: Alexis S. Chaine orcid.org/0000-0003-3346-551X
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.