
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 24 September 2019
Sec. Conservation and Restoration Ecology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00355
This article is part of the Research Topic Understanding the Impact and Invasion Success of Aquatic Non-native Species: How they Interact with Novel Environments and Native Biota View all 11 articles
 Phillip J. Haubrock1,2,3*†‡
Phillip J. Haubrock1,2,3*†‡ Paride Balzani4†‡
Paride Balzani4†‡ Martina Azzini4‡Alberto F. Inghilesi2,4Lukáš Veselý3Wei Guo3
Martina Azzini4‡Alberto F. Inghilesi2,4Lukáš Veselý3Wei Guo3 Elena Tricarico4†
Elena Tricarico4†Interactions occurring between species in multiple invaded freshwater ecosystems are often difficult to observe and study. Studies on invasive alien species typically focus on single species, and, when the species community is exclusively composed of alien species, their interactions, eventual facilitation and hindering processes are very scarcely assessed. To investigate such a community, the species community in the Arno River (Tuscany, Italy) was examined as a model system using the combined approach of stable isotopes and dietary analyses. Established alien species have formed a pyramid shaped community with the European catfish Silurus glanis and the North American channel catfish Ictalurus punctatus in the apex position, followed by opportunistic predatory (Lepomis gibbosus) and omnivore (Alburnus alburnus, Cyprinus carpio, Barbus barbus, Pseudorasbora parva, Padogobius sp., Tinca tinca) species. These species were observed to feed on a variety of primary producers (Myriophyllum sp., Potamogeton sp., Phragmites australis) and consumers (aquatic insects, molluscs, Dikerogammarus villosus, Procambarus clarkii, Palaemon antennarius, tadpoles). Remarkably, the channel catfish Ictalurus punctatus showed an ontogenetic niche shift, with juveniles occupying a slightly higher trophic position than mature individuals. Pseudorasbora parva and A. alburnus showed a strong niche overlap, with the former having a wider niche. Such wide niches were also found for the invasive crustaceans D. villosus and P. clarkii. Outgoing from our findings, we suggest that life-history and geographic origin play a role in determining competition and interaction type among alien species, with species from the same geographic area showing a lower potential to compete than species from different areas, hence affecting their potential impact on native species.
The invasive potential as well as distribution of an alien species depend on various abiotic and biotic factors such as its life-history (Kolar and Lodge, 2001; Keller et al., 2007; Strayer, 2010; Blackburn et al., 2011), predator-prey dynamics, adaptability as well as interactions with other species (Ricciardi, 2003; Strayer et al., 2006; Ricciardi et al., 2013). Subsequent, the introduction and successive dispersal of alien species change the identity of entire species communities (Seebens et al., 2017). When multiple invasive alien species are dominating one freshwater ecosystem, they tend to occupy different trophic levels (Gamradt and Kats, 1996; Kiesecker and Blaustein, 1998), but the presence of invasive species interacting across trophic levels increases the already-existing complex threat through direct and indirect effects (Peckarsky and McIntosh, 1998; Huxel et al., 2002; Strayer, 2010), as the prevalence of omnivores (Pringle and Hamazaki, 1998), and interactions among predators (Sih et al., 1998) are unforeseeable. However, such complex interactions among invasive species are not well understood (Parker et al., 1999; Huxel et al., 2002; Strayer, 2010) but, as described in the “invasional meltdown theory,” can enhance the chance of a species to successfully establish itself and increase or add further impacts on the recipient environment (Simberloff and Von Holle, 1999; Simberloff, 2006). Otherwise, these interactions can lower the impact of predatory invasive species on native species in certain cases (Soluk and Collins, 1988a,b; Soluk, 1993; Rosenheim, 1998; Bissattini et al., 2018). Despite the utmost importance of such studies on multiple invaded ecosystems for practical management and control efforts (Polis and Strong, 1996; Rosenheim, 1998), they are still scarce (but see Johnson et al., 2009; Gherardi et al., 2011; Haubrock et al., 2019a).
The use of Stable Isotopes Analyses (SIA) enables the study of invasive species and their interactions with native biota (Vander Zanden et al., 1999; Balzani et al., 2016). In particular, SIA proved to be a useful tool in investigating the impacts of alien fish (Cucherousset et al., 2012; Britton et al., 2018). Using SIA for carbon (C) and nitrogen (N), long-term and time-mediated information of a community trophic structure can be obtained (Boecklen et al., 2011; Layman et al., 2012; Middelburg, 2014), trophic levels can be estimated (Post, 2002), and feeding ecological niches finely quantified (Newsome et al., 2007). While C signatures identify the major energy sources, N signatures relate to the trophic position of a consumer within a food web (Fry, 2006; Layman et al., 2012). This relationship relies on predictable changes in the isotopic signal from prey to consumer, being enriched by 1%0 for C and by 2.5-5%0 for N between consecutive trophic levels (Post, 2002; Vanderklift and Ponsard, 2003). Combined with the analysis of dietary contents (DA), which provide a direct but short temporal insight into the feeding habit of a species, relationships among species can be investigated with a considerably higher resolution.
Aquatic ecosystems, especially in the Mediterranean areas, have been subjected to successive invasions (Dumont et al., 2004; Marr et al., 2010). For instance, Italian freshwaters, with an ichthyofauna composed of 48 native and 41 established introduced and further 15 introduced but not yet established species, have been imperiled by invasions (Nocita et al., 2017). Many of these species are notoriously invasive, like the European catfish Silurus glanis (Copp et al., 2007, 2009; Boulêtreau et al., 2018), which has become the object of various management efforts (Gualtieri and Mecatti, 2005; Cucherousset et al., 2018), while others like the channel catfish Ictalurus punctatus have received minor attention, with only few recent studies addressing its potential impacts (Haubrock et al., 2018a,b). In the Arno River in Tuscany, species introductions occurred repeatedly, as in the case of Blicca bjoerkna, Rhodeus sericeus, I. punctatus, and S. glanis (Nocita, 2002, 2007), that are now established (Nocita and Zerunian, 2007). Moreover, anthropogenic effects combined with introductions have caused the local extinction of native predators (e.g., Esox lucius; Nocita and Zerunian, 2007). Therefore, the Arno River is a peculiar ecosystem, because almost no native fish species is currently present (Nocita, 2002), and it is thus characterized by unique species composition, with so far unknown interactions and diverse introduction histories. Moreover, several of the species present in the Arno river (e.g., P. clarkii, S. glanis, D. villosus, G. chinensis, P. antipodarum, P. parva, G. holbrooki; Gherardi et al., 2008; Nocita and Lenuzza, 2016) are listed among the worst invasive species and other comparable lists of high-profile invasive species (Vilà et al., 2009; Nentwig et al., 2018). It can be considered as a unique melting pot for alien species from Asia and North America, mainly voluntarily introduced for sport fishing and commercial fishing purposes (Nocita, 2002; Vejrík et al., 2019). Consequently, manifold interactions that can only be observed in such an invaded range are of great importance from both a theoretic and management point of view.
We, thus, hypothesize that species interactions among and across trophic levels and, especially top-down impacts, are affected not only by differing life-histories but also a shared history of co-evolution of the invading species, with invasive alien species coming from different areas being more likely to compete due to the lack of the latter.
Hence, the aims of this work were to: (i) combine SIA and DA to investigate feeding preferences and impacts of alien species in the investigated community mainly composed by alien species; (ii) compare species' trophic niches and niche overlap to infer the potential degree of feeding competition among species; (iii) unravel important functional roles of organisms (as pointed out by (Bissattini and Vignoli, 2017) for P. clarkii, linking terrestrial and aquatic energy sources), and (iv) discuss results in regard to invasive species' origins as a potential factor determining the outcome of species interactions.
The Arno River is with a length of 241 km and a drainage of more than 8.200 km2 as well as an effluent flow of 100 m2/s the second most important river in Central Italy (Nocita and Zerunian, 2007). Its discharge regime is typical of Mediterranean rivers showing extreme flow reductions during summer. It flows through the major cities of Florence and Pisa and is especially within Florence divided by various weirs and characterized by a high density of non-native aquatic vegetation during spring and early summer. During summer, the water of the Arno River in the inner-city section of Florence can reach mean temperatures of 29.6°C (min: 27.2, max: 32.4; data from 2016 to 2018 http://www.arpat.toscana.it). The historic use and cultural value have led to high anthropogenic stress on communities. Indeed, the aquatic community consists of various alien species while only a minority of them is considered native (Nocita, 2002). However, a detailed list of alien species has not been published so far (pers. comm. Gianna Innocenti).
Sampling was conducted from April to June 2018, in a very anthropogenically disturbed part of the inner-city (Florence) section of the Arno River which is highly used for recreational angling (43.765606, 11.268234; DATUM WGS84). The specific period was chosen, as these months are the ideal activity period for several fish species (samples were taken approximately 2 month after the inactive winter period, thus after species had resumed their feeding activity). Moreover, the effect of angling as a stressor that could interfere with the species trophic positioning through the input of “unusual food sources” is limited due to low angling activity (pers. Comm, Thomas Bussatto). To collect the several species, a combined approach of several methods was applied. Fish were caught with standard fishing rods and the help of local fishermen using a variety of baits and fishing techniques. Macroinvertebrates and frog tadpoles were collected using funnel traps and hand nets. Sampled species were collected under the consideration of an even spatial distribution across the sampled stretch of the river ecosystem without bias toward either section, end or distance from riverbank (Willson et al., 2010). Samples of the local aquatic vegetation were also taken as they represent the baseline of the ecosystem. Sampled specimens were immediately euthanized (fish: gills cutting; macroinvertebrates: freezing), stored on ice during transport and then preserved in the freezer at −20°C until further processing. Overall, 232 specimens belonging to nine fish species, 71 invertebrates from at least six species, four green frog tadpoles, and thirteen samples from three different plant species (two hydrophytes: Potamogeton sp. and Myriophyllum sp., and one helophyte: Phragmites australis) were collected. Due to the rarity of both Tinca tinca and Barbus barbus in the study site, only one sample of each was caught and included in the dataset to roughly indicate these species positioning in the community. However, these samples were not used for any further statistical analysis. The sampling was authorized by the Tuscan Region (“Autorizzazione alla pesca scientifica, Regione Toscana”).
For all fish species, total length (TL) was measured to infer age, as fish growth is generally continuous, presenting a link between size and age (Hopkins, 1992). Resultant, over TL; the age can be inferred and used to identify fish maturity, which can in turn be used to investigate behavioral differences (Haubrock et al., 2019b). Additionally, weight (W), and, if distinguishable, also sex were recorded. TL of fish and cephalothorax length (CTL) of crayfish were measured with a caliper and W was measured using an electronic balance. In the laboratory, stomach contents were extracted, and prey items identified to the lowest possible taxon under a stereomicroscope (Sansoni, 1998; Schultz, 2010). We estimated number of stomachs containing each food item in relation to total number of full stomachs (frequency of occurrence, F%) and number of individuals of each food item with respect to the total number of individuals (number of occurrence, N%) for each food item in each respective species (Haubrock et al., 2019b). Despite the information DA provide on the direct feeding activity, they are frequently used to supplement stable isotope analyses as priors due to their often differing results (Pacioglu et al., 2019). However, from all species analyzed, several had empty stomachs (S. glanis: 20/39; A. alburnus: 2/10; P. parva: 4/10; L. gibbosus: 7/20). For isotopic analyses, muscle tissue was cleaned of fat, skin, scales, bones and carapace (for crayfish). For plants, different leaves were cut-off from the same individual and pooled in one sample to account for eventual differences. Due to their small size, insects, little crustaceans and tadpoles were treated as whole samples while, for molluscs, only the soft body tissue was analyzed without the shell. Samples were placed on separated glass trays and dried for 48 h at 60°C. Once dry, all samples were grinded into fine powder with an agate mortar and pestle. For each sample, two replicates of 0.20–0.30 mg for animals and 1.00–1.10 mg for plants, were weighed on a Mettler Toledo AG245 microscale and enclosed in a tin capsule to be analyzed in continuous flow—isotope ratio mass spectrometry (CF-IRMS) with a Thermo FlashEA 1112 elemental analyser and a Thermo Finnigan Delta Plus isotope ratio mass spectrometer at the Istituto di Geologia Ambientale e Geoingegneria (IGAG) of the National Research Council (CNR) in Montelibretti, Rome. Isotope compositions were expressed as %0 with the δ notation (based on δ13C or δ15N = [(Rsample/Rstandard) – 1) × 1000, where R is 13C/12C or 15N/14N ratios]. Results were referred to Vienna Pee Dee Belemnite for carbon and to atmospheric N2 for nitrogen, with laboratory standards calibrated on IAEA international standards: N1, N2, and USGS25 for nitrogen and CH6, CH7, and USGS24 for carbon.
The arithmetic mean ± SE was calculated, and eventual correlations between δ15N and δ13C values and TL, CTL and W measures were tested for each species using a linear model. The trophic position (TP) of each species was estimated using the R package “tRophicPosition,” which incorporates a Bayesian model utilizing up to two baselines (Quezada-Romegialli et al., 2018). Hence, we included the combined plants (Myriophyllum sp., Potamogeton sp., Phragmites australis) as first, and molluscs (Gyraulus chinensis) as second baseline (n.adapt = 100,000; n.iter = 100,000; burnin = 10,000; n.chains = 5). These two groups were chosen due to the wide diet preferences known for alien fish species (Musil and Adámek, 2007; Almeida et al., 2009; Xu et al., 2011; Ribeiro and Leunda, 2012; Haubrock et al., 2019a,b).
To quantify intra-specific and community niche width, Layman metrics (Layman et al., 2007) were calculated with the R package “SIAR” (Stable Isotope Analysis in R; Parnell et al., 2010). Additionally, the corrected standard ellipse areas (SEAc, considering 40% of data plots) and SEAb (Bayesian standard ellipse areas, considering 95% of data plots) were calculated for all fish species with n ≥ 5 as well as Procambarus clarkii using the R package “SIBER” (Stable Isotope Bayesian Ellipses in R; Jackson et al., 2011).
The application of scatter plot for δ15N vs. δ13C of consumer tissues and food sources enabled the determination of probable prey sources and combinations of prey contributing to the diet of predators (Phillips and Gregg, 2003). Dietary analyses (DA) for catfish species (juvenile and mature I. punctatus as well as S. glanis) were previously performed (Haubrock et al., 2019b), while DA for Lepomis gibbosus, Pseudorasbora parva, and Alburnus alburnus were conducted following the same protocol (Table 1). Stable isotope mixing models (SIMM; R package “SIMMr”) were applied for all fish and crayfish without consideration of priors but considering potentially predated items, i.e., a predation on lower trophic levels. Thus, it was investigated how sampled species contribute to the isotopic signatures of each other, indicating the scenario with the highest probability (Parnell et al., 2013). Additionally, the same analyses were performed again for L. gibbosus, both life-stages of I. punctatus and S. glanis with results from DA as priors for the analyses (i.e., considering: the five most consumed prey items and then their number of occurrence N%; 4 prey items were used for L. gibbosus; Table 1). Priors from DA were not included into the mixing models for P. parva nor A. alburnus due to the low number of stomachs available (n = 10 for either species) of which the majority was empty (P. parva = 6; A. alburnus = 8). Prior based analyses were conducted with the R package “SIAR” (Parnell and Jackson, 2013). Results are presented as the average percent values with the possible range percentage for each prey item. With these, the trophic web of the studied ecosystem was reconstructed. Furthermore, using the software PRIMER (Clarke and Warwick, 2001) a Multi-Dimensional Scaling plot (MDS) was drawn and a Permutational Analysis of Variance (PERMANOVA; Resemblance: Euclidean distance; Sums of squares type: Type III (partial); Fixed effects sum to zero for mixed terms; Permutation method: Unrestricted permutation of raw data; Levels: 15; Number of permutations: 9999) was performed to test for significant differences in δ15N and δ13C among species and to eventually determine niche overlaps. Lastly, we compared the results among species to discuss the effect of an alien species origin in respect to indicated trophic interactions.
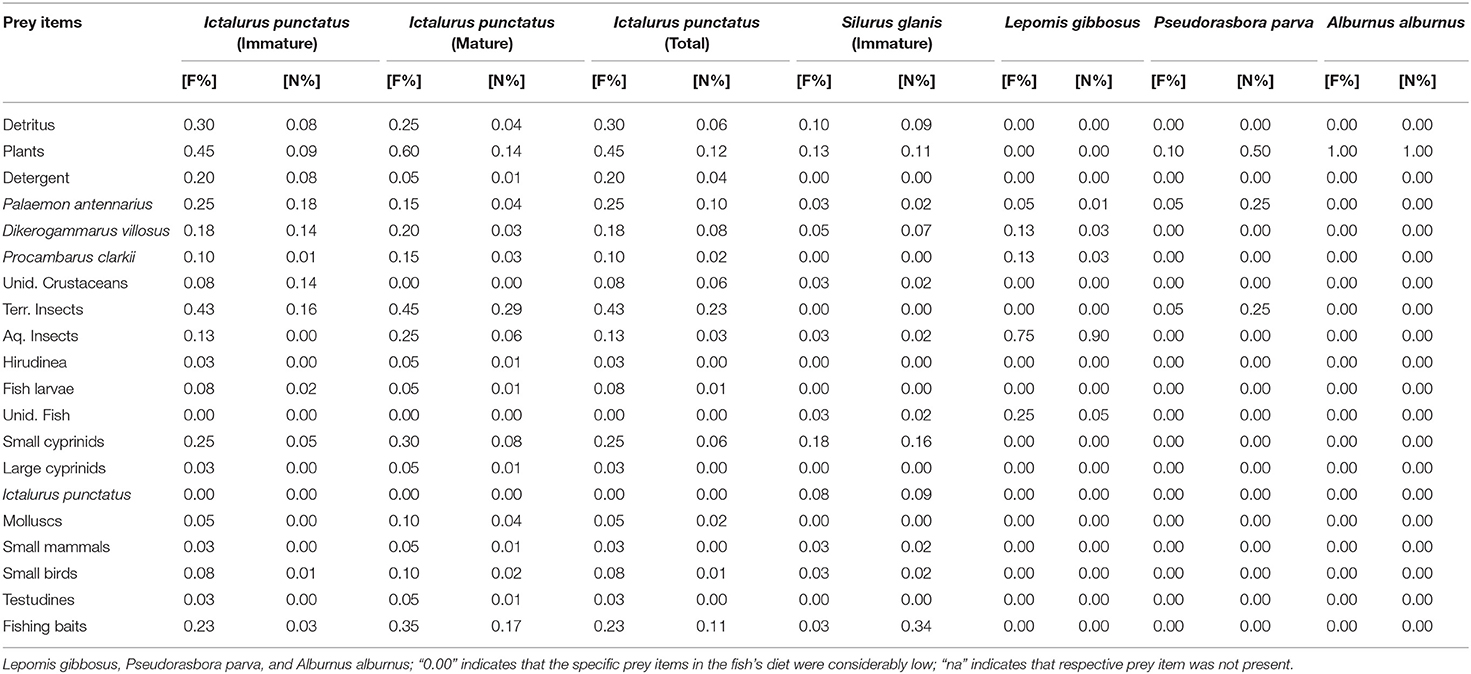
Table 1. Frequency of occurrence (%F), abundance (%N), and prominence value (PV) of food items analyzed for juvenile and mature Ictalurus punctatus, juvenile Silurus glanis.
Overall, 9 invasive but no native fish species were sampled. There were complimented by 2 invasive and 1 native crustacean as well as 1 invasive mollusc. Additionally, 2 native insect, 1 native amphibian and 3 invasive plant species were collected (Table 2).
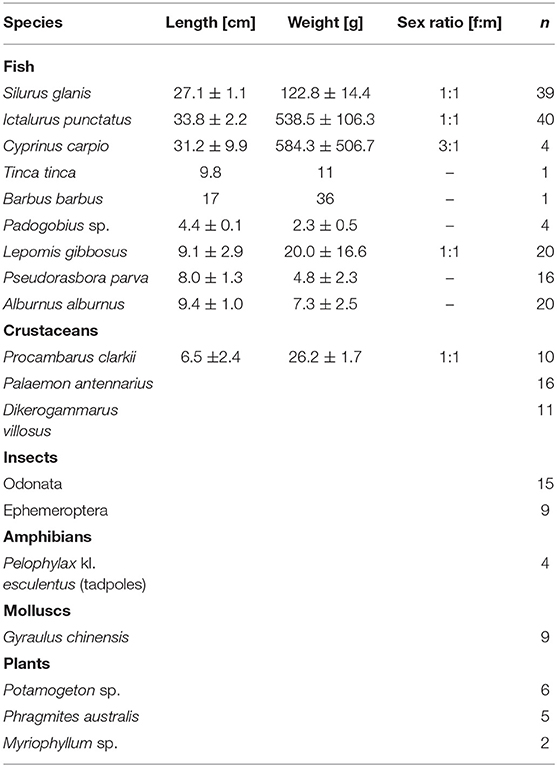
Table 2. Morphological measures (mean ± SE), sex ratio (calculated as ratio of females, f. on males, m) and number of all sampled specimens. Length measured as total length (TL) for fishes and as cephalothorax length (CTL) for crayfish; n = number of specimens caught.
From these, some species were under-sampled due to their rarity (i.e., the locally alien species Tinca tinca and Barbus barbus) or status as endangered (i.e., Padogobius sp.), while only a minimum of Cyprinus carpio and no mature S. glanis (>80 cm) individuals were sampled to meet the demands of local anglers. Additionally, the alien omnivore Squalius cephalus, which is believed to be rare or locally extinct, was not sampled. The abundant invasive pond slider Trachemys scripta as well as the occasionally found alligator snapping turtle Macrochelys temminckii were not sampled as the sampling method was not suited to collect them. Moreover, only few macroinvertebrates were found and collected. Detailed results from DA are listed in Table 1 and have been discussed previously for I. punctatus and juvenile S. glanis (Haubrock et al., 2019b). While the number of stomach contents analyzed for P. parva and A. alburnus were too low to be analyzed in depth or to be used as prior for subsequent mixing models, L. gibbosus showed a primary feeding preference for aquatic insects and secondarily for crustaceans (Table 1).
Statistically significant negative correlations were found between TL and δ13C for S. glanis (rs = −0.512; p = 0.001; n = 39), I. punctatus (rs = −0.665; p < 0.001; n = 40) as well as P. clarkii (rs = −0.693; p = 0.026; n = 10). A statistically significant positive correlation between TL and δ13C were identified for Pseudorasbora parva (rs = 0.567; p = 0.022; n = 16). Additionally, a statistically significant positive correlation between TL and δ15N was found for L. gibbosus (rs = 0.917; p < 0.001; n = 20) (Supplement 1).
Following the assumption of an isotopic enrichment of δ15N by about 3.4%0 (Post, 2002), the entire isotopic community spanned over approximately three to four trophic levels (Figure 1). Considering as baseline both molluscs and plants provided a well-structured hierarchy. The 4th apex level was constituted by S. glanis and both life stages of I. punctatus; the 3rd level by C. carpio, B. barbus, Padogobius sp., L. gibbosus, P. parva, A. alburnus, P. clarkii, and P. antennarius; the 2nd level by T. tinca, D. villosus, tadpoles, Odonata, and Ephemeroptera larvae.
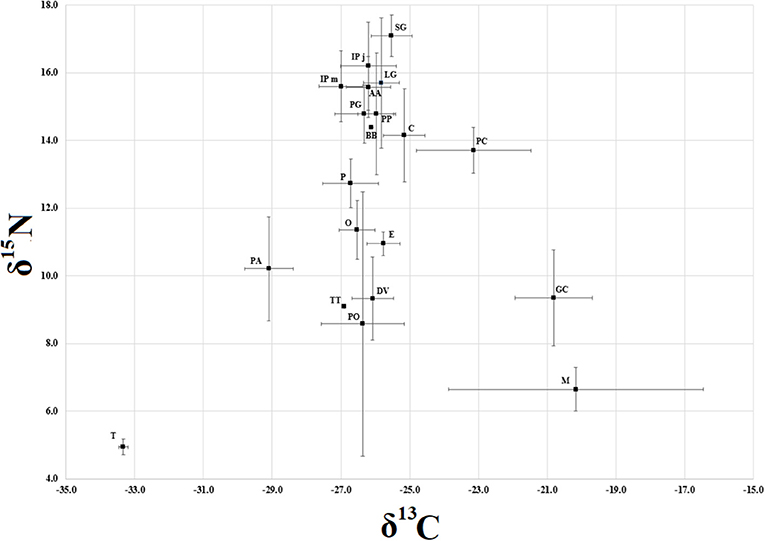
Figure 1. Isotopic space scatterplot of the community. Species IDs: SG, Silurus glanis; IPj, Ictalurus punctatus (juveniles); IPm, Ictalurus punctatus (matures); LG, Lepomis gibbosus; AA, Alburnus alburnus; PG, Padogobius sp.; PP, Pseudorasbora parva; BB, Barbus barbus; C, Cyprinus carpio; PC, Procambarus clarkii; P, Palaemon antennarius; O, Odonata; E, Ephemeroptera; DV, Dikerogammarus villosus; GC, Gyraulus chinensis; PA, Phragmites australis; T, tadpoles (Pelophylax kl. esculentus); TT, Tinca tinca; M, Myriophyllum sp.; PO, Potamogeton sp. Each species is represented as mean (centroid), with bars representing standard deviation.
Layman's metrics are shown in Table 3. Among fish species, the greatest N range (NR) was found for P. parva (7.51), followed by L. gibbosus (6.38) and adults of I. punctatus (4.60), while the narrowest values were presented by Padogobius sp. (1.79) and S. glanis (2.71). C range (CR) was largest for A. alburnus (3.24) and smallest for C. carpio (1.32). Total Area (TA, i.e., the convex hull area enclosing all individual points) was highest in P. parva (9.64), followed by I. punctatus juveniles (8.76) and L. gibbosus (7.81), and was narrowest in Padogobius sp. (0.13) and C. carpio (1.53). SEAc, which is less influenced by extreme values than TA, was highest in L. gibbosus (3.33), immediately followed by P. parva (3.27) then I. punctatus juveniles (2.20); the lowest values of SEAc were found for Padogobius sp. (0.17), S. glanis (0.87) and C. carpio (1.72). However, the small metrics for Padogobius sp. and C. carpio have to be considered with caution, due to the small number of collected individuals (n = 4 for both species). Among crustaceans, D. villosus had the highest NR (4.40), followed by Palaemon (2.11) and P. clarkii (2.07), while CR followed the exactly opposite hierarchy: P. clarkii = 3.96, Palaemon = 2.95 and D. villousus = 1.90. TA was comparable between P. clarkii and D. villosus (4.60 and 4.49, respectively), while smaller in Palaemon (3.38). In contrast, SEAc was highest for P. clarkii (3.59), followed by D. villosus (2.36), then Palaemon (1.39). Finally, tadpoles showed the lowest Layman's metrics of the whole community.
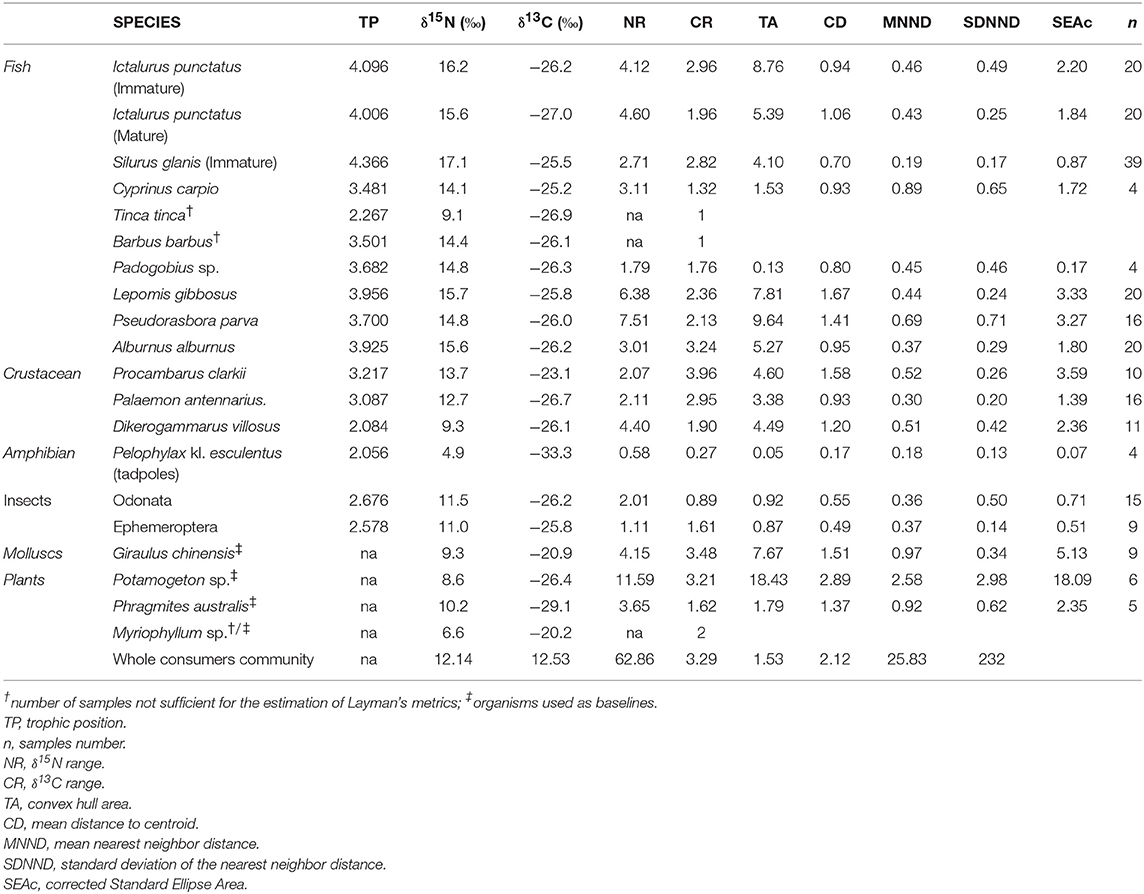
Table 3. Mean values of isotopic signatures, estimated trophic positions, Layman's metrics.
Considerable SEAc overlap was identified between juvenile I. punctatus and A. alburnus (0.50), matures I. punctatus and L. gibbosus (0.47), L. gibbosus and P. parva (0.49), and A. alburnus and P. parva (1.33). Overlap was generally low among all other fish species. P. clarkii showed particularly low overlap values with any fish species. The estimated SEAb overlaps indicated considerable overlap between life stages of I. punctatus (0.42), P. parva and L. gibbosus (0.59), and A. alburnus and P. parva (0.46) and minor overlap values between all other species. The ellipse overlaps between A. alburnus and P. parva estimated with SEAb was considerably lower than the SEAc overlap. Lastly, while the identified SEAb overlap was low between mature I. punctatus and juvenile S. glanis (0.09), juveniles I. punctatus and L. gibbosus (0.17), SEAc overlap did not indicate any overlap at all. Similarly, low SEAb overlaps between matures I. punctatus and P. parva (0.12), juvenile S. glanis and A. alburnus (0.22), and juvenile S. glanis and P. parva (0.16) were considerably higher than the respective SEAc overlap values, all being near 0 (Table 4).
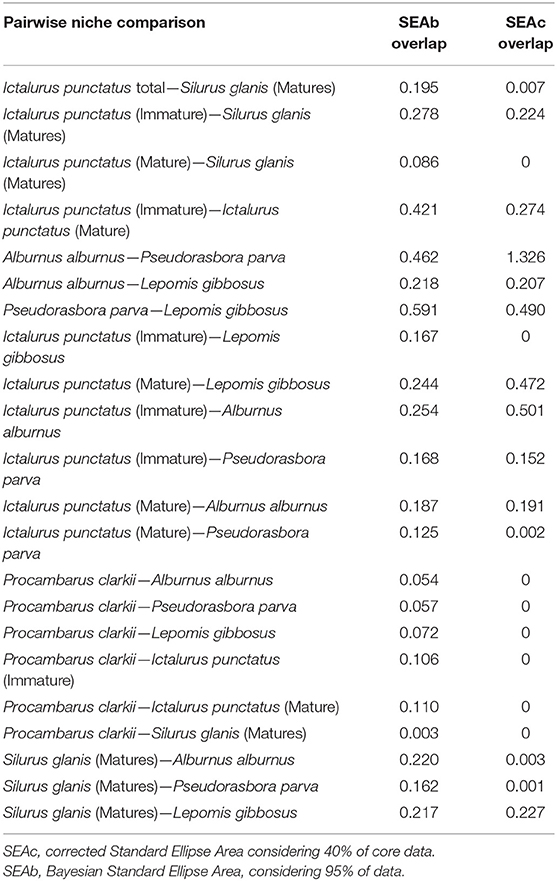
Table 4. Estimated ellipses overlap between all fish species with n ≥ 5 and the crayfish Procambarus clarkii.
Results of the applied SIMM without priors (Figure 2) indicated that both I. punctatus juveniles and adults expressed a great reliance on A. alburnus with a higher proportion in matures compared to juveniles, which consumed other items as well. Silurus glanis SIMM also showed a specialized piscivorous diet, focusing mostly on the consumption of L. gibbosus. In contrast, L. gibbosus had a more generalist diet, based almost equally on tadpoles and various invertebrates: Ephemeroptera and Odonata larvae among insects, and Palaemon sp., D. villosus and the invasive P. clarkii among crustaceans. For P. parva, the estimated diet consisted mainly of Palaemon sp., followed by Ephemeroptera and then by Odonata, D. villosus and P. clarkii. The diet of A. alburnus is similar, but more balanced and with a marked preference for Palaemon. Finally, P. clarkii showed a preference for molluscs, followed by insects (Ephemeroptera and Odonata), Palaemon, plants (Phragmites), and tadpoles.
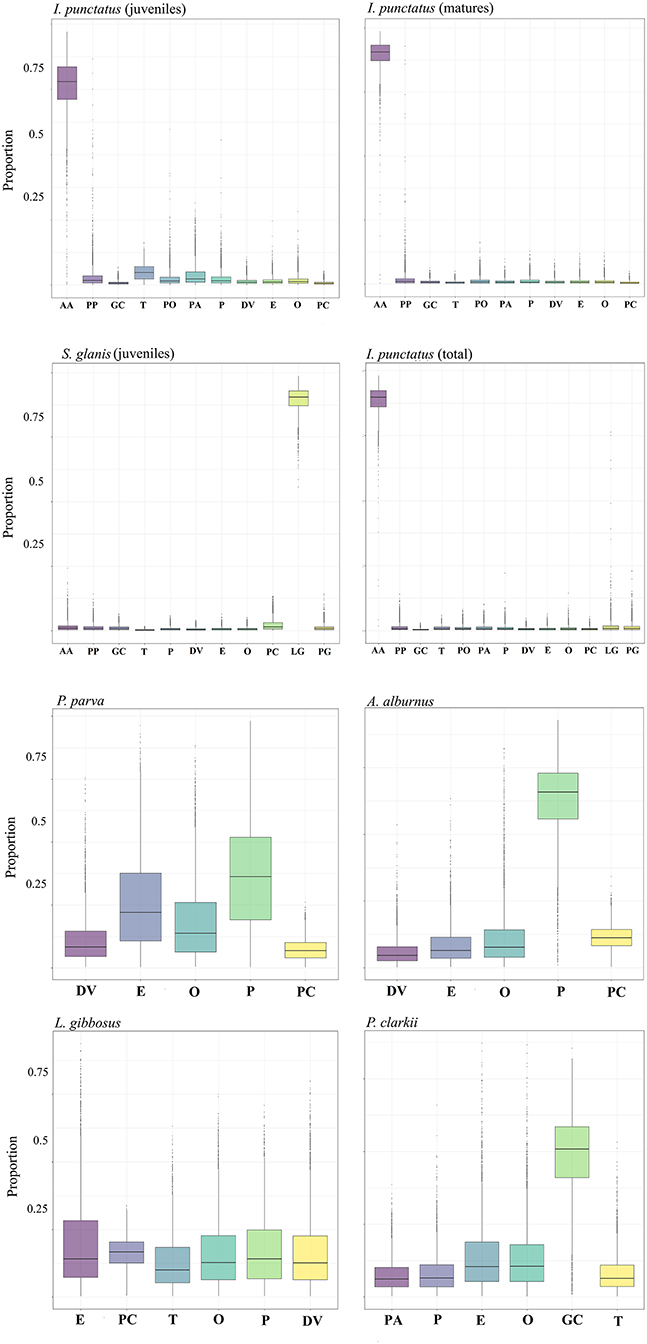
Figure 2. Estimated proportions of prey items to the diet of: Ictalurus punctatus (juveniles), Ictalurus punctatus (matures), Ictalurus punctatus (total), juvenile Silurus glanis, Pseudorasbora parva, Alburnus alburnus, Lepomis gibbosus, and Procambarus clarkii. Prey items were considered those belonging to lower trophic levels without consideration of any prior. The analysis was performed using the R-package “simmr”.
Mixing models refined using dietary priors (Figure 3) showed differing trends, highlighting a predation of I. punctatus juveniles on P. antennarius, D. villosus, as well as on small cyprinids (e.g., A. alburnus) and of I. punctatus (matures) on cyprinids, P. antennarius and D. villosus. As for S. glanis, I. punctatus and D. villosus constituted the preferred preys. Differing results were obtained for L. gibbosus; without numerical priors, Palaemonsp. and, with priors, aquatic insects majorly contributed to its trophic level. However, SIMMs including N% priors led to an increase in the estimated proportion of contributing prey items and less variability.
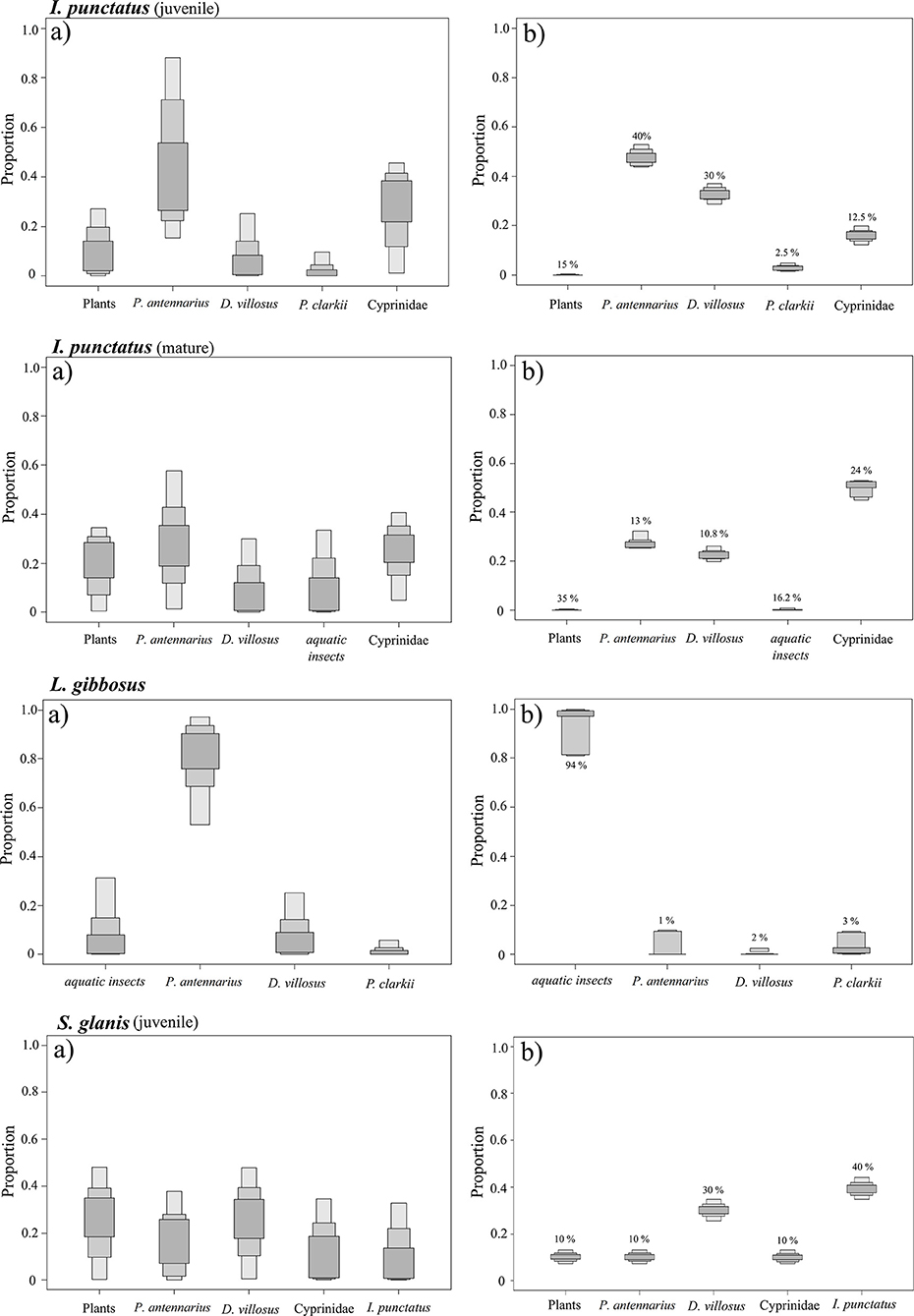
Figure 3. Estimated contribution of consumed prey items to the diet of: Ictalurus punctatus (juveniles), Ictalurus punctatus (matures), Lepomis gibbosus and juvenile Silurus glanis using dietary priors. (a) Based on the 5* most consumed prey items; (b) including the number of occurrences (N%) of the 5* most consumed prey items (indicated in %). *: 4 in the case of L. gibbosus. The prior based analysis was performed using the R-package “SIAR” with credibility intervals of 95%.
The applied PERMANOVA on carbon and nitrogen signatures highlighted an overall distinction among species [F(14, 202) = 135.34, p = 0.001; Table 5]. Nonetheless, the applied pairwise post-hoc tests (Supplement 2) indicated that some fish species had no distinct isotopic niches. In particular, overlapping niches were found between A. alburnus and juvenile I. punctatus (p = 0.054), A. alburnus and L. gibbosus (p = 0.46), A. alburnus and Padogobius sp. (p = 0.185), A. alburnus and P. parva (p = 0.085), C. carpio and L. gibbosus (p = 0.124), C. carpio and Padogobius sp. (p = 0.193), C. carpio and P. parva (p = 0.337), juvenile I. punctatus and L. gibbosus (p = 0.221), mature I. punctatus and Padogobius sp. (p = 0.146), L. gibbosus and Padogobius sp. (p = 0.35), L. gibbosus and P. parva (p = 0.145), Padogobius and P. parva (p = 0.803).
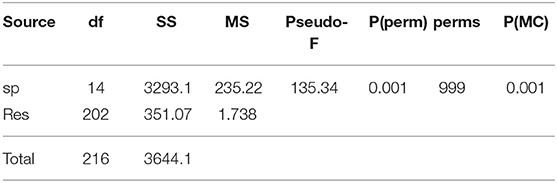
Table 5. PERMANOVA main test result on nitrogen and carbon isotopes signatures for factor SPECIES.
Multiple invaded communities are particularly interesting as most studies focus on the effects of a single introduced species (but see Preston et al., 2012; Jackson, 2015). In the Mediterranean area, to the best of our knowledge, only few studies explored the trophic interactions in highly invaded inland water communities despite the high frequency of species invasions (Galil, 2007; Di Castri et al., 2012). Furthermore, they both concern lentic waters and comprise generally fewer species. Like our study, combining dietary analyses and stable isotopes, Stellati et al. (accepted) described the trophic interactions of a pond community in Monterotondo (Rome, Central Italy), composed by almost exclusively invasive species except for top predators (Natrix natrix and Natrix tesselata. Using an identical approach, Haubrock et al. (2019a) investigated the relationships in Northern Spain in Arreo Lake dominated by invasive species. As shown by the studies above, such integrative approaches applied to a peculiar community can provide not only insights into trophic interactions among invasive species but can also improve our ability to understand their synergistic impacts onto communities. However, in the study presented here, investigations were conducted during spring season in a completely different ecosystem: a highly anthropogenically altered river with no seasonal drought and no native fish predators. While the dataset cannot be seen as enough to make a generalization on the entire year, we believe that the collected data are sufficient to analyse the interactions and subsequent interactions among alien invasive species in the Arno river during spring season.
The reconstructed community had a typical pyramidal shape, with the apex position being held by two invasive predatory fish, I. punctatus and juvenile S. glanis. Compared with the works of Stellati et al. (accepted) and Bissattini and Vignoli (2017), in which the community was composed—according to isotopic data—by approximately two trophic levels, in the Arno River it spanned over at least three trophic levels, indicating a higher complexity. However, it is interesting to note that this complexity is reached with a high number of species, while in Arreo Lake the same number of trophic levels was obtained with only four species, that had clearly separated niches, with a primary role of N isotopes signatures (and therein the trophic level) in this partitioning (Haubrock et al., 2019a). On the contrary, the Arno River community showed some overlap among trophic niches of fish species, but not a clear segregation of all niches as observed in Arreo Lake (Haubrock et al., 2019a), as the latter was the result of the low species number and a common geographic origin.
Estimations of trophic positions showed a strong top predator behavior for S. glanis, I. punctatus, especially in the juvenile stage, and also L. gibbosus, coherently with their known biology and other isotope studies (Werner et al., 1977; Vander Zanden and Rasmussen, 1999; Vander Zanden et al., 1999; Syväranta et al., 2010). Compared with Haubrock et al. (2019a), L. gibbosus was identified as a possibly secondary -not a primary- predator. This might be associated to the differences in ecosystem and community complexity. The functional category of primary predatory fish was represented by P. parva, A. alburnus, and Padogobius sp., although for the latter this position should be taken with caution due to the low number of sampled individuals (n = 4). Cyprinus carpio (n = 4) and the only specimen of B. barbus can be considered true omnivores as they had a TP intermediate between that of primary predators and herbivores. Commonly, A. alburnus and P. parva are known omnivores with a preference for zoobenthos (Politou et al., 1993; Gozlan et al., 2010). Nonetheless, they have the ability to exert wider, on higher trophic level focused feeding preferences (Vøllestad, 1985; Lammens and Hoogenboezem, 1991; Nagata et al., 2005), potentially explaining their comparatively elevated TPs (Syväranta et al., 2010; Jackson and Britton, 2013). However, the low number of individuals captured cannot allow other speculations, but rather give a rough indication of the positioning of these species trophic niche in the studied ecosystem. Among invertebrates, the invasive red swamp crayfish P. clarkii and the native P. antennarius were also included in this category. Compared with Arreo lake (Haubrock et al., 2019a), P. clarkii belonged to a higher trophic level, potentially due to the greater complexity of the Arno River community and therefore higher number of potential prey items and advanced stage of eutrophication. As discussed for B. barbus, the trophic position of the omnivorous T. tinca, cannot be considered precise; however, both these samples are still indicative also for future studies on these two rare species in this or other ecosystems. Surprisingly, the larvae of predatory insects (Ephemeroptera and Odonata) as well as the invasive amphipod D. villosus, which is known to be a predator of other aquatic macroinvertebrates and fish eggs (Dick and Platvoet, 2000), fell in the same trophic level of primary consumers. This is maybe due to predation on unsampled invertebrate consumers, with a lower N isotopes signature than mollusks, leading to potential underestimations in TPs. Moreover, due to the generally high angling activity (using commonly float and feeder fishing) in the Arno river, an unpredictable influx of unnatural food sources cannot be excluded. These items could likely affect the trophic positioning of species like T. tinca. However, the angling activity in the Arno is considerably low during spring (pers. comm. Thomas Bussatto). Additionally, if such baited fishing would occur, used bait-items would have been observed within the diet of I. punctatus (Haubrock et al., 2018b, 2019b). Interestingly, juvenile I. punctatus had a slightly higher nitrogen signature than matures, which is uncommon for catfish species as they generally express higher N values with increasing total length (Syväranta et al., 2010). Considering the high level of eutrophication and the natural accumulation of nutrients (i.e., dead matter) in the river bottom, this observation can potentially be explained by the greater consumption of enriched detritus by juveniles (Hendrix et al., 1986; Hansen and Kristensen, 1998). Another explanation could be that adult I. punctatus lower their trophic position to avoid (or is confined to a lower position as a result of) high competition with other piscivorous fish like S. glanis (Fry, 1999) as they share similar niches. Because S. glanis is not a detrivorous species, an effect of N-enriched detritus can be neglected in this species' case. Nonetheless, it should be noted that the inclusion of adult specimens of S. glanis might lead to a significant correlation between TL and N signatures (Syväranta et al., 2010).
Moreover, several species expressed a negative correlation between TL and C signatures, suggesting consumption of prey items that feed on different aquatic or terrestrial vegetation. This underlines the scenario that these species change habits as they grow, with e.g., juveniles of I. punctatus living closer to the riverbank and relying on fallen terrestrial items more than matures, the latter feeding offshore. However, although Syväranta et al. (2010) argued that in S. glanis δ15N increases with TL due to the ability to consume higher trophic prey, such an increase was only observed in L. gibbosus, indicating an ontogenetic size-based prey shift.
Among fish species, Layman's metrics (Layman et al., 2007) showed a great variability in N signatures, thus indicating the utilization of prey items from different trophic levels in P. parva, L. gibbosus, and adults of I. punctatus, while juvenile S. glanis were quite specialized in its trophic role as apex predator. Carbon variability was largest in A. alburnus, suggesting the direct or indirect consumption of prey from different origins. Niche width measures showed the widest niche for L. gibbosus, P. parva, and juveniles of I. punctatus, thus, these species express the greatest intra-specific variability. In contrast, a more specialized niche was found for S. glanis together with a niche partitioning among catfish species with the potential to compete due to the generalist diet of I. punctatus (see Haubrock et al., 2019b for a detailed discussion). Moreover, the comparison between A. alburnus and P. parva, two ecologically similar species belonging to the same trophic level and with a considerable niche overlap, showed a considerably wider niche width for P. parva based on a considerable variability in δ15N values, probably due to a higher feeding variability and adaptability to many ecosystems, a key factor in its invasion history. Indeed, Rosecchi et al. (1993), Gozlan et al. (2010) and Yalçin-Özdilek et al. (2013) stated that P. parva is prone to predate, among other items, fish larvae as well as eggs. However, while these potential preys were present during the time of the sampling, they were not observed in the diets, as previously observed (Annamaria Nocita, unpubl. data). In contrast, A. alburnus showed significantly less inter-individual variability linked to feeding activity mostly relying on macroinvertebrates (Politou et al., 1993).
Similarly, it is interesting to compare the two size-matched crustaceans D. villosus and P. antennarius. The greater variability in δ15N was found in the former, suggesting the consumption of prey items from different trophic levels. However, D. villosus unexpected low trophic position raises questions on its specialized predatory behavior, whilst P. antennarius, despite being an omnivore feeding also on detritus and algae, occupied a higher trophic position with low variability in δ15N. Moreover, as for P. parva, D. villosus had a considerably wider niche than that of P. antennarius, a factor potentially related to the success of this invasive species. Finally, P. clarkii showed a wider niche compared with the other two crustaceans, particularly due to its variability in carbon sources. This was not surprising, giving the opportunistic and generalist diet, especially in invaded areas (Souty-Grosset et al., 2016).
Permutational analysis of variance of species niches identified considerable overlaps mainly between predatory fish species, confirming SEAc and SEAb overlap estimations. The result is a community structure more packed at higher than at lower trophic levels (Figure 1). Therefore, we speculate that, in the Arno River community, a diminished predator pressure could be not caused by the presence of a keystone species (such as in Bissattini et al., 2018), but by the potential competition among predators and potentially occurring competition- avoidance mechanisms (Haubrock et al., 2018b).
Standard mixing models under the assumption of predation on lower trophic positions (Figure 2) presented limited, but representative, information, as this theoretical predation scenario is estimated based on the potential availability of prey items and the possibility of their consumption (Phillips, 2001; Phillips et al., 2005). However, including the observed prey and their occurrences in the diet in these models, considerable differences emerged (Figure 3). When using DA based priors, I. punctatus presented more precise proportions of prey contributions, while the diet of L. gibbosus is based more on aquatic insects, which is directly mirrored by DA results (Table 1). For P. parva and A. alburnus, where no suitable DA were applicable due to the low number of stomach contents, mixing models without priors showed a higher predation of P. antennarius by A. alburnus, but a wider more generalistic feeding by P. parva. In addition, using the five most commonly present prey items in the diet of S. glanis led to a considerable role of plants, which did not majorly contribute to its TP. Nonetheless, it has to be stated that only 19 of 39 S. glanis had full stomachs. In this regard, the presence of plants in the diet of S. glanis is particularly interesting, as firstly, S. glanis is only feed on plant matter during early invasion (Carol et al., 2009), thus life stages, and secondly, because symbiotic bacteria are needed for fish to be able to digest plants (Vejríková et al., 2016). However, accidental digestion of plant matter while preying on living prey may occur, the potential competition with I. punctatus cannot be excluded of having caused diet shifts within S. glanis immature life stage due to this species known generalism and adaptability to new prey sources (Vejrík et al., 2017b; Haubrock et al., 2019b).
Concluding, this latter prior-based method can be considered more accurate, due to the combination of two different time mediated methods (Moore and Semmens, 2008). Nevertheless, when information on the local diet are not available, the first method without prior can be considered as very informative (Pacioglu et al., 2019).
The study river is a peculiar ecosystem, as the majority of the inhabiting species are alien. Moreover, the communities' hierarchical order indicates a typically intact community and highlights the foregone interactions that have led to this structure (McQueen et al., 1989; Polis, 1994). The main predatory species were observed to mostly feed on other alien species, and thus, resulting in complex and manifold species interactions. These differing interactions underline the need for studies on a case by case basis. Interestingly, combining SIA and DA revealed differing interactions according to whether alien species shared a common life-history or not.
In the case of I. punctatus and L. gibbosus, as these two species co-occur in the same ecosystems in North America (Werner et al., 1977; Smith, 1981) and thus, share a common life-history and overlapping native range as well as history of co-evolution, some niche overlap was identified. Moreover, the applied PERMANOVA did not indicate a statistically significant difference between juvenile catfish and pumpkinseed trophic niches. The diet of both included a considerable percentage of aquatic insects, but the mixing models with and without prior showed a different contribution of prey items. More importantly, I. punctatus did not show any indication of predation on or direct competition with L. gibbosus. Hence, competition could be not direct, but rather based on the opportunistic nature of I. punctatus or other factors that lead to a slightly lower trophic position in mature than juvenile I. punctatus. Moreover, these species likely avoid competition with their different feeding activities (benthivorous generalist vs. pelagic opportunist). On the other hand, where potentially competing alien species came from different geographical ranges, observations were slightly different. Pseudorasbora parva and A. alburnus, two species with similar behavior and activity pattern, showed major isotopic niche overlap confirmed also by the PERMANOVA post-hoc test, suggesting that, if resources become limiting, these species are likely to compete.
As for the two apex predators, the results indicated that while both, mature and juvenile I. punctatus show a medium to high diet overlap with S. glanis, a minor isotope overlap was only observed between juvenile I. punctatus and S. glanis but not among life-stages of I. punctatus (Haubrock et al., 2019b). It has therefore to be considered that I. punctatus is a primarily benthic forager, a factor that likely limits the competition with other species. These results led to the conclusion that both life-stages of I. punctatus show a potential to compete with juvenile S. glanis but not with each other, likely due to diet- and behavioral-based intra-specific avoidance mechanisms. However, it should be noted that including mature individuals of S. glanis, as shown in Syväranta et al. (2010) and Vejrík et al. (2019), might have an effect of this species' niche occupation and the observed niche partitioning (Haubrock et al., 2019b). With the use of mixing models, different preferred prey items were identified for both species, while the use of priors, when applicable, indicated a considerable feeding activity by S. glanis on I. punctatus.
Finally, the presence of P. clarkii, often described as a keystone species (Liu et al., 2018), was shown to lower the impact of L. catesbeianus on native amphibians due to P. clarkii's availability as prey (Bissattini and Vignoli, 2017). However, in the Arno River it occurred at low densities. For this reason, the more abundant P. antennarius and D. villosus, both highly present in the diet of multiple species as well as indicated by the mixing models, probably reside in a similar function within the community. Therefore, in our study site, the presence of P. clarkii seemed not to lower impacts on present species by predatory fish, but rather contributed to it.
Not all alien species that originate from roughly the same geographic region share a common history of co-evolution. However, in this studies case, North American species indeed share the same ecosystems. As such, it was shown that not only alien species do differ in their ecological niche occupation, but the existence and type of interactions may differ according to the involved species and their respective histories of co-evolution. This is true despite the close clustering of fish species within the 4th and 3rd trophic level. Indeed, our combined SIA and DA results suggest that, when two alien species originate from the same geographical range, competition seemed to be possible due to some diet overlap, but was neglectable due to varying feeding preferences, occupied niche space and potential avoidance mechanisms (Haubrock et al., 2018b), as indicated by prior (DA) -based mixing models and SEAb/c. Nonetheless, when species originated from different geographic ranges but occupied the same trophic level, i.e., similar trophic positions, shared niche space and isotope overlap were more likely to occur. Resultant, due to the variability of occurring interactions, niche shifts or increasing competition could lead to an increased impact on native species, while predetermined interactions among species with common life-history could lead to a lowered feeding pressure. However, more studies are therefore needed in this direction to confirm these findings and to increase information on these alien species' impacts. These future sampling campaigns should also consider summer and autumn because behaviors and thus, interactions may differ (Vejrík et al., 2017a), potentially further enhancing our comprehension of mechanisms underlying biological invasions and species interactions.
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethical review and approval was not required for the animal study because this study was carried out in accordance with the principles of and under the restriction of the laws currently effective in Tuscany, Italy, following the recommendations of Dr. Annamaria Nocita. The protocol was approved by the local governmental authorities.
PH, PB, and ET designed the study. PH, PB, MA, and WG performed research. PH, MA, and AI analyzed data. LV contributed new models. PH and PB wrote the first draft. All authors contributed to the paper and approved it.
Funding was provided by the Aquainvad-ED project (2020 Marie Sklodowska-Curie ITN-2014-ETN-642197).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We wish to thank Thomas Busatto, Ivan Fribbi, and Mirko Ventrone for helping us with the sampling, and Giuliana Parisi for having allowed us to partly process the samples in her laboratory. We also want to thank Antonín Kouba for his useful comments on an earlier draft of the manuscript. We would like to acknowledge Elisabeth Jane Sainsbury for her English revision on the final manuscript. Lastly, we would like to express our thanks to two reviewers.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00355/full#supplementary-material
Almeida, D., Almodóvar, A., Nicola, G. G., and Elvira, B. (2009). Feeding tactics and body condition of two introduced populations of pumpkinseed Lepomis gibbosus: taking advantages of human disturbances? Ecol. Freshw. Fish 18, 15–23. doi: 10.1111/j.1600-0633.2008.00317.x
Balzani, P., Vizzini, S., Santini, G., Masoni, A., Ciofi, C., Ricevuto, E., et al. (2016). Stable isotope analysis of trophic niche in two co-occurring native and invasive terrapins, Emys orbicularis and Trachemys scripta elegans. Biol. Invasions 18, 3611–3621. doi: 10.1007/s10530-016-1251-x
Bissattini, A. M., Buono, V., and Vignoli, L. (2018). Field data and worldwide literature review reveal that alien crayfish mitigate the predation impact of the American bullfrog on native amphibians. Aquat. Conserv. 28, 1465–1475. doi: 10.1002/aqc.2978
Bissattini, A. M., and Vignoli, L. (2017). Let's eat out, there's crayfish for dinner: American bullfrog niche shifts inside and outside native ranges and the effect of introduced crayfish. Biol. Invasions 19, 2633–2646. doi: 10.1007/s10530-017-1473-6
Blackburn, T. M., Pyšek, P., Bacher, S., Carlton, J. T., Duncan, R. P., Jarošík, V., et al. (2011). A proposed unified framework for biological invasions. Trends Ecol. Evol. (Amst). 26, 333–339. doi: 10.1016/j.tree.2011.03.023
Boecklen, W. J., Yarnes, C. T., Cook, B. A., and James, A. C. (2011). On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 42, 411–440. doi: 10.1146/annurev-ecolsys-102209-144726
Boulêtreau, S., Gaillagot, A., Carry, L., Tétard, S., De Oliveira, E., and Santoul, F. (2018). Adult Atlantic salmon have a new freshwater predator. PLoS ONE 13:e0196046. doi: 10.1371/journal.pone.0196046
Britton, J. R., Ruiz-Navarro, A., Verreycken, H., and Amat-Trigo, F. (2018). Trophic consequences of introduced species: Comparative impacts of increased interspecific versus intraspecific competitive interactions. Funct. Ecol. 32, 486–495. doi: 10.1111/1365-2435.12978
Carol, J., Benejam, L., Benito, J., and García-Berthou, E. (2009). Growth and diet of European catfish (Silurus glanis) in early and late invasion stages. Fund. Appl. Limnol. Arch. Hydrobiol. 174, 317–328. doi: 10.1127/1863-9135/2009/0174-0317
Clarke, K. R., and Warwick, R. M. (2001). A further biodiversity index applicable to species lists: variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 216, 265–278. doi: 10.3354/meps216265
Copp, G. H., Britton, R., Cucherousset, J., García-Berthou, E., Kirk, R., Beeler, E., et al. (2009). Voracious invader or benign feline? A review of the environmental biology of European catfish Silurus glanis in its native and introduced ranges. Fish Fish. 10, 252–282. doi: 10.1111/j.1467-2979.2008.00321.x
Copp, G. H., Moffatt, L., and Wesley, K. J. (2007). Is European catfish Silurus glanis really becoming more abundant in the River Thames? Aquat. Invas. 2, 113–116. doi: 10.3391/ai.2007.2.2.5
Cucherousset, J., Boulêtreau, S., Martino, A., Roussel, J. M., and Santoul, F. (2012). Using stable isotope analyses to determine the ecological effects of non-native fishes. Fish. Manag. Ecol. 19, 111–119. doi: 10.1111/j.1365-2400.2011.00824.x
Cucherousset, J., Horký, P., Slavík, O., Ovidio, M., Arlinghaus, R., Boulêtreau, S., et al. (2018). Ecology, behaviour and management of the European catfish. Rev. Fish Biol. Fish 28, 177–190. doi: 10.1007/s11160-017-9507-9
Di Castri, F., Hansen, A. J., and Debussche, M. (eds.). (2012). Biological Invasions in Europe and the Mediterranean Basin, Vol. 65. Dordrecht: Springer Science & Business Media.
Dick, J. T., and Platvoet, D. (2000). Invading predatory crustacean Dikerogammarus villosus eliminates both native and exotic species. Proc. R. Soc. Lond. Ser. B Biol. Sci., 267, 977–983. doi: 10.1098/rspb.2000.1099
Dumont, H. J., Shiganova, T. A., and Niermann, U. (eds.). (2004). Aquatic Invasions in the Black, Caspian, and Mediterranean Seas, Vol. 35. Dordrecht: Springer Science & Business Media. doi: 10.1007/1-4020-2152-6
Fry, B. (1999). Using stable isotopes to monitor watershed influences on aquatic trophodynamics. Can. J. Fish. Aquat. Sci. 56, 2167–2171. doi: 10.1139/f99-152
Fry, B. (2006). Stable Isotope Ecology, Vol. 521. New York, NY: Springer. doi: 10.1007/0-387-33745-8
Galil, B. S. (2007). Loss or gain? Invasive aliens and biodiversity in the Mediterranean Sea. Mar. Pollut. Bull. 55, 314–322. doi: 10.1016/j.marpolbul.2006.11.008
Gamradt, S. C., and Kats, L. B. (1996). Effect of introduced crayfish and mosquitofish on California newts. Conserv. Biol. 10, 1155–1162. doi: 10.1046/j.1523-1739.1996.10041155.x
Gherardi, F., Bertolino, S., Bodon, M., Casellato, S., Cianfanelli, S., Ferraguti, M., et al. (2008). Animal xenodiversity in Italian inland waters: distribution, modes of arrival, and pathways. Biol. Invasions 10, 435–454. doi: 10.1007/s10530-007-9142-9
Gherardi, F., Britton, J. R., Mavuti, K. M., Pacini, N., Grey, J., Tricarico, E., et al. (2011). A review of allodiversity in Lake Naivasha, Kenya: Developing conservation actions to protect East African lakes from the negative impacts of alien species. Biol. Conserv. 144, 2585–2596. doi: 10.1016/j.biocon.2011.07.020
Gozlan, R. E., Andreou, D., Asaeda, T., Beyer, K., Bouhadad, R., Burnard, D., et al. (2010). Pan-continental invasion of Pseudorasbora parva: towards a better understanding of freshwater fish invasions. Fish Fish. 11, 315–340. doi: 10.1111/j.1467-2979.2010.00361.x
Gualtieri, M., and Mecatti, M. (2005). Indagine Sulla Diffusione del Siluro (Silurus glanis) nell'Arno Fiorentino. Prima relazione 2005. Technical Report, 11.
Hansen, K., and Kristensen, E. (1998). The impact of the polychaete Nereis diversicolor and enrichment with macroalgal (Chaetomorpha linum) detritus on benthic metabolism and nutrient dynamics in organic-poor and organic-rich sediment. J. Exp. Mar. Biol. Ecol. 231, 201–223. doi: 10.1016/S0022-0981(98)00070-7
Haubrock, P. J., Azzini, M., Balzani, P., Inghilesi, A. F., and Tricarico, E. (2019b). When alien catfish meet—Resource overlap between the North American Ictalurus punctatus and immature European Silurus glanis in the Arno River (Italy). Ecol. Freshw. Fish. doi: 10.1111/eff.12481. [Epub ahead of print].
Haubrock, P. J., Azzini, M., Fribbi, I., Inghilesi, A. F., and Tricarico, E. (2018b). Opportunistic alien catfish: unexpected findings in the diet of the alien species Ictalurus punctatus in Central Italy. Fish. Aquat. Life 26, 239–242. doi: 10.2478/aopf-2018-0027
Haubrock, P. J., Balzani, P., Criado, A., Inghilesi, A. F., Tricarico, E., and Monteoliva, A. P. (2019a). Predicting the effects of reintroducing a native predator (European eel, Anguilla anguilla) into a freshwater community dominated by alien species using a multidisciplinary approach. Manag. Biol. Invas. 10, 171–191. doi: 10.3391/mbi.2019.10.1.11
Haubrock, P. J., Balzani, P., Johovic, I., Inghilesi, A. F., Nocita, A., and Tricarico, E. (2018a). The diet of the alien channel catfish Ictalurus punctatus in the River Arno (Central Italy). Aquat. Invas 13, 575–585. doi: 10.3391/ai.2018.13.4.14
Hendrix, P. F., Parmelee, R. W., Crossley, D. A. Jr., Coleman, D. C., Odum, E. P., and Groffman, P. M. (1986). Detritus food webs in conventional and no-tillage agroecosystems. Bioscience 36, 374–380. doi: 10.2307/1310259
Hopkins, K. D. (1992). Reporting fish growth: a review of the basics 1. J. World Aquac. Soc. 23, 173–179. doi: 10.1111/j.1749-7345.1992.tb00766.x
Huxel, G. R., McCann, K., and Polis, G. A. (2002). Effects of partitioning allochthonous and autochthonous resources on food web stability. Ecol. Res. 17, 419–432. doi: 10.1046/j.1440-1703.2002.00501.x
Jackson, A. L., Inger, R., Parnell, A. C., and Bearhop, S. (2011). Comparing isotopic niche widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 80, 595–602. doi: 10.1111/j.1365-2656.2011.01806.x
Jackson, M. C. (2015). Interactions among multiple invasive animals. Ecology 96, 2035–2041. doi: 10.1890/15-0171.1
Jackson, M. C., and Britton, J. R. (2013). Variation in the trophic overlap of invasive Pseudorasbora parva and sympatric cyprinid fishes. Ecol. Freshw. Fish 22, 654–657. doi: 10.1111/eff.12063
Johnson, P. T., Olden, J. D., Solomon, C. T., and Vander Zanden, M. J. (2009). Interactions among invaders: community and ecosystem effects of multiple invasive species in an experimental aquatic system. Oecologia 159, 161–170. doi: 10.1007/s00442-008-1176-x
Keller, R. P., Lodge, D. M., and Finnoff, D. C. (2007). Risk assessment for invasive species produces net bioeconomic benefits. Proc. Natl. Acad. Sci. U.S.A. 104, 203–207. doi: 10.1073/pnas.0605787104
Kiesecker, J. M., and Blaustein, A. R. (1998). Effects of introduced bullfrogs and smallmouth bass on microhabitat use, growth, and survival of native red-legged frogs (Rana aurora). Conserv. Biol. 12, 776–787. doi: 10.1046/j.1523-1739.1998.97125.x
Kolar, C. S., and Lodge, D. M. (2001). Progress in invasion biology: predicting invaders. Trends Ecol. Evol. (Amst). 16, 199–204. doi: 10.1016/S0169-5347(01)02101-2
Lammens, E. H. R. R., and Hoogenboezem, W. (1991). “Diets and feeding behaviour,” in Cyprinid Fishes, eds I. J. Winfield and J. S. Nelson (Dordrecht: Springer), 353–376. doi: 10.1007/978-94-011-3092-9_12
Layman, C. A., Araujo, M. S., Boucek, R., Hammerschlag-Peyer, C. M., Harrison, E., Jud, Z. R., et al. (2012). Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol. Rev. 87, 545–562. doi: 10.1111/j.1469-185X.2011.00208.x
Layman, C. A., Arrington, D. A., Montaña, C. G., and Post, D. M. (2007). Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 88, 42–48. doi: 10.1890/0012-9658(2007)88[42:CSIRPF]2.0.CO;2
Liu, X., Wang, S., Ke, Z., Cheng, C., Wang, Y., Zhang, F., et al. (2018). More invaders do not result in heavier impacts: the effects of non-native bullfrogs on native anurans are mitigated by high densities of non-native crayfish. J. Anim. Ecol. 87, 850–862. doi: 10.1111/1365-2656.12793
Marr, S. M., Marchetti, M. P., Olden, J. D., García-Berthou, E., Morgan, D. L., Arismendi, I., et al. (2010). Freshwater fish introductions in mediterranean-climate regions: are there commonalities in the conservation problem? Divers. Distribut. 16, 606–619. doi: 10.1111/j.1472-4642.2010.00669.x
McQueen, D. J., Johannes, M. R., Post, J. R., Stewart, T. J., and Lean, D. R. (1989). Bottom-up and top-down impacts on freshwater pelagic community structure. Ecol. Monogr. 59, 289–309. doi: 10.2307/1942603
Middelburg, J. J. (2014). Stable isotopes dissect aquatic food webs from the top to the bottom. Biogeosciences 11, 2357–2371. doi: 10.5194/bg-11-2357-2014
Moore, J. W., and Semmens, B. X. (2008). Incorporating uncertainty and prior information into stable isotope mixing models. Ecol. Lett. 11, 470–480. doi: 10.1111/j.1461-0248.2008.01163.x
Musil, J., and Adámek, Z. (2007). Piscivorous fishes diet dominated by the Asian cyprinid invader, topmouth gudgeon (Pseudorasbora parva). Biologia 62, 488–490. doi: 10.2478/s11756-007-0093-5
Nagata, T., Ha, J. Y., and Hanazato, T. (2005). The predation impact of larval Pseudorasbora parva (Cyprinidae) on zooplankton: a mesocosm experiment. J. Freshw. Ecol. 20, 757–763. doi: 10.1080/02705060.2005.9664800
Nentwig, W., Bacher, S., Kumschick, S., Pyšek, P., and Vilà, M. (2018). More than “100 worst” alien species in Europe. Biol. Invas. 20, 1611–1621. doi: 10.1007/s10530-017-1651-6
Newsome, S. D., Martinez del Rio, C., Bearhop, S., and Phillips, D. L. (2007). A niche for isotopic ecology. Front. Ecol. Environ. 5, 429–436. doi: 10.1890/1540-9295(2007)5[429:ANFIE]2.0.CO;2
Nocita, A. (2002). Carta ittica della Provincia di Firenze. Prov. Firenze, Ass. Agric. Caccia e pesca – Museo di Storia Naturale della Universita di Firenze, Sezione Zoologica “La Specola”, 260.
Nocita, A., and Lenuzza, A. (2016). “Le specie ittiche aliene in Toscana e il caso del Siluro europeo,” in Conference on Codice Armonico (Castiglioncello, LI), 164–171.
Nocita, A., Tricarico, E., and Bertolino, S. (2017). Fine-scale analysis of heavily invaded Italian freshwater fish assemblages. Integr. Zool. 12, 500–511. doi: 10.1111/1749-4877.12267
Nocita, A., and Zerunian, S. (2007). L'ittiofauna aliena nei fiumi e nei laghi d'Italia. Biol. Ambient. 21, 93–96.
Pacioglu, O., Zubrod, J. P., Schulz, R., Jones, J. I., and Pârvulescu, L. (2019). Two is better than one: combining gut content and stable isotope analyses to infer trophic interactions between native and invasive species. Hydrobiologia 839, 25–35. doi: 10.1007/s10750-019-03990-8
Parker, I. M., Simberloff, D., Lonsdale, W. M., Goodell, K., Wonham, M., Kareiva, P. M., et al. (1999). Impact: toward a framework for understanding the ecological effects of invaders. Biol. Invasions 1, 3–19. doi: 10.1023/A:1010034312781
Parnell, A., and Jackson, A. (2013). siar: Stable Isotope Analysis in R. R Package Version 4.2. Available online at: http://CRAN.R-project.org/package=siar (accessed March 23, 2014).
Parnell, A. C., Inger, R., Bearhop, S., and Jackson, A. L. (2010). Source partitioning using stable isotopes: coping with too much variation. PLoS ONE 5:e9672. doi: 10.1371/journal.pone.0009672
Parnell, A. C., Phillips, D. L., Bearhop, S., Semmens, B. X., Ward, E. J., Moore, J. W., et al. (2013). Bayesian stable isotope mixing models. Environmetrics 24, 387–399. doi: 10.1002/env.2221
Peckarsky, B. L., and McIntosh, A. R. (1998). Fitness and community consequences of avoiding multiple predators. Oecologia 113, 565–576. doi: 10.1007/s004420050410
Phillips, D. L. (2001). Mixing models in analyses of diet using multiple stable isotopes: a critique. Oecologia 127, 166–170. doi: 10.1007/s004420000571
Phillips, D. L., and Gregg, J. W. (2003). Source partitioning using stable isotopes: coping with too many sources. Oecologia 136, 261–269. doi: 10.1007/s00442-003-1218-3
Phillips, D. L., Newsome, S. D., and Gregg, J. W. (2005). Combining sources in stable isotope mixing models: alternative methods. Oecologia 144, 520–527. doi: 10.1007/s00442-004-1816-8
Polis, G. A. (1994). Food webs, trophic cascades and community structure. Aust. J. Ecol. 19, 121–136. doi: 10.1111/j.1442-9993.1994.tb00475.x
Polis, G. A., and Strong, D. R. (1996). Food web complexity and community dynamics. Am. Nat. 147, 813–846. doi: 10.1086/285880
Politou, C. Y., Economidis, P. S., and Sinis, A. I. (1993). Feeding biology of bleak, Alburnus alburnus, in Lake Koronia, northern Greece. J. Fish Biol. 43, 33–43. doi: 10.1111/j.1095-8649.1993.tb00408.x
Post, D. M. (2002). Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83, 703–718. doi: 10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2
Preston, D. L., Henderson, J. S., and Johnson, P. T. (2012). Community ecology of invasions: direct and indirect effects of multiple invasive species on aquatic communities. Ecology 93, 1254–1261. doi: 10.1890/11-1821.1
Pringle, C. M., and Hamazaki, T. (1998). The role of omnivory in a neotropical stream: separating diurnal and nocturnal effects. Ecology 79, 269–280. doi: 10.1890/0012-9658(1998)079[0269:TROOIA]2.0.CO;2
Quezada-Romegialli, C., Jackson, A. L., Hayden, B., Kahilainen, K. K., Lopes, C., and Harrod, C. (2018). tRophicPosition, an R package for the Bayesian estimation of trophic position from consumer stable isotope ratios. Methods Ecol. Evol. 9, 1592–1599. doi: 10.1111/2041-210X.13009
Ribeiro, F., and Leunda, P. M. (2012). Non-native fish impacts on Mediterranean freshwater ecosystems: current knowledge and research needs. Fish. Manag. Ecol. 19, 142–156. doi: 10.1111/j.1365-2400.2011.00842.x
Ricciardi, A. (2003). Predicting the impacts of an introduced species from its invasion history: an empirical approach applied to zebra mussel invasions. Freshw. Biol. 48, 972–981. doi: 10.1046/j.1365-2427.2003.01071.x
Ricciardi, A., Hoopes, M. F., Marchetti, M. P., and Lockwood, J. L. (2013). Progress toward understanding the ecological impacts of nonnative species. Ecol. Monogr. 83, 263–282. doi: 10.1890/13-0183.1
Rosecchi, E., Crivelli, A. J., and Catsadorakis, G. (1993). The establishment and impact of Pseudorasbora parva, an exotic fish species introduced into Lake Mikri Prespa (north-western Greece). Aquat. Conserv. 3, 223–231. doi: 10.1002/aqc.3270030306
Rosenheim, J. A. (1998). Higher-order predators and the regulation of insect herbivore populations. Annu. Rev. Entomol. 43, 421–447. doi: 10.1146/annurev.ento.43.1.421
Sansoni, G. (1998). “Atlante per il riconoscimento dei macroinvertebrati dei corsi d'acqua italiani,” in Agenzia provinciale per la Protezione dell'ambiente (Trento: APR & B).
Seebens, H., Blackburn, T. M., Dyer, E. E., Genovesi, P., Hulme, P. E., Jeschke, J. M., et al. (2017). No saturation in the accumulation of alien species worldwide. Nat. Commun. 8:14435. doi: 10.1038/ncomms14435
Sih, A., Englund, G., and Wooster, D. (1998). Emergent impacts of multiple predators on prey. Trends Ecol. Evol. 13, 350–355. doi: 10.1016/S0169-5347(98)01437-2
Simberloff, D. (2006). Rejoinder to Simberloff (2006): don't calculate effect sizes; study ecological effects. Ecol. Lett. 9, 921–922. doi: 10.1111/j.1461-0248.2006.00952.x
Simberloff, D., and Von Holle, B. (1999). Positive interactions of nonindigenous species: invasional meltdown? Biol. Invasions 1, 21–32. doi: 10.1023/A:1010086329619
Smith, G. R. (1981). Late cenozoic freshwater fishes of North America. Annu. Rev. Ecol. Syst. 12, 163–193. doi: 10.1146/annurev.es.12.110181.001115
Soluk, D. A. (1993). Multiple predator effects: predicting combined functional response of stream fish and invertebrate predators. Ecology 74, 219–225. doi: 10.2307/1939516
Soluk, D. A., and Collins, N. C. (1988a). Synergistic interactions between fish and stoneflies: facilitation and interference among stream predators. Oikos 52, 94–100. doi: 10.2307/3565987
Soluk, D. A., and Collins, N. C. (1988b). A mechanism for interference between stream predators: responses of the stonefly Agnetina capitata to the presence of sculpins. Oecologia 76, 630–632. doi: 10.1007/BF00397881
Souty-Grosset, C., Anastacio, P. M., Aquiloni, L., Banha, F., Choquer, J., Chucholl, C., et al. (2016). The red swamp crayfish Procambarus clarkii in Europe: impacts on aquatic ecosystems and human well-being. Limnologica 58, 78–93. doi: 10.1016/j.limno.2016.03.003
Stellati, L., Borgiann, N., Bissattini, A. M., Buono, V., Haubrock, P. J., Balzani, P., et al. (accepted). Living with aliens: suboptimal ecological condition in semiaquatic snakes inhabiting a hot spot of allodiversity. Acta Oecol.
Strayer, D. L. (2010). Alien species in fresh waters: ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 55, 152–174. doi: 10.1111/j.1365-2427.2009.02380.x
Strayer, D. L., Eviner, V. T., Jeschke, J. M., and Pace, M. L. (2006). Understanding the long-term effects of species invasions. Trends Ecol. Evol. (Amst). 21, 645–651. doi: 10.1016/j.tree.2006.07.007
Syväranta, J., Cucherousset, J., Kopp, D., Crivelli, A., Céréghino, R., and Santoul, F. (2010). Dietary breadth and trophic position of introduced European catfish Silurus glanis in the River Tarn (Garonne River basin), southwest France. Aquat. Biol. 8, 137–144. doi: 10.3354/ab00220
Vander Zanden, M. J., Casselman, J. M., and Rasmussen, J. B. (1999). Stable isotope evidence for the food web consequences of species invasions in lakes. Nature 401:464. doi: 10.1038/46762
Vander Zanden, M. J., and Rasmussen, J. B. (1999). Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology 80, 1395–1404. doi: 10.1890/0012-9658(1999)080[1395:PCCANA]2.0.CO;2
Vanderklift, M. A., and Ponsard, S. (2003). Sources of variation in consumer-diet δ15N enrichment: a meta-analysis. Oecologia 136, 169–182. doi: 10.1007/s00442-003-1270-z
Vejrík, L., Vejríková, I., Blabolil, P., Eloranta, A. P., Kočvara, L., Peterka, J., et al. (2017b). European catfish (Silurus glanis) as a freshwater apex predator drives ecosystem via its diet adaptability. Sci. Rep. 7, 15970. doi: 10.1038/s41598-017-16169-9
Vejrík, L., Vejríková, I., Kočvara, L., Sajdlová, Z., Chung, S. H. T., Šmejkal, M., et al. (2017a). Thirty-year-old paradigm about unpalatable perch egg strands disclaimed by the freshwater top-predator, the European catfish (Silurus glanis). PLoS ONE 12:e0169000. doi: 10.1371/journal.pone.0169000
Vejrík, L., Vejríková, I., Kocvara, L., Blabolil, P., Peterka, J., Sajdlová, Z., et al. (2019). The pros and cons of the invasive freshwater apex predator, European catfish Silurus glanis, and powerful angling technique for its population control. J. Environ. Manage. 241, 374–382. doi: 10.1016/j.jenvman.2019.04.005
Vejríková, I., Vejrík, L., Syväranta, J., Kiljunen, M., Cech, M., Blabolil, P., et al. (2016). Distribution of herbivorous fish is frozen by low temperature. Sci. Rep. 6:39600. doi: 10.1038/srep39600
Vilà, M., Basnou, C., Gollasch, S., Josefsson, M., Pergl, J., and Scalera, R. (2009). “One hundred of the most invasive alien species in Europe,” in Handbook of Alien Species in Europe, ed P. Hulme (Dordrecht: Springer), 265–268. doi: 10.1007/978-1-4020-8280-1_12
Vøllestad, L. A. (1985). Resource partitioning of roach Rutilus rutilus and bleak Alburnus alburnus in two eutrophic lakes in SE Norway. Ecography 8, 88–92. doi: 10.1111/j.1600-0587.1985.tb01157.x
Werner, E. E., Hall, D. J., Laughlin, D. R., Wagner, D. J., Wilsmann, L. A., and Funk, F. C. (1977). Habitat partitioning in a freshwater fish community. J. Fish. Board Canada 34, 360–370. doi: 10.1139/f77-058
Willson, J. D., Winne, C. T., Pilgrim, M. A., Romanek, C. S., and Gibbons, J. W. (2010). Seasonal variation in terrestrial resource subsidies influences trophic niche width and overlap in two aquatic snake species: a stable isotope approach. Oikos 119, 1161–1171. doi: 10.1111/j.1600-0706.2009.17939.x
Xu, J., Zhang, M., and Xie, P. (2011). Sympatric variability of isotopic baselines influences modeling of fish trophic patterns. Limnology 12, 107–115. doi: 10.1007/s10201-010-0327-z
Keywords: stable isotopes, stomach contents, community structure, mixing models, feeding ecology, interactions
Citation: Haubrock PJ, Balzani P, Azzini M, Inghilesi AF, Veselý L, Guo W and Tricarico E (2019) Shared Histories of Co-evolution May Affect Trophic Interactions in a Freshwater Community Dominated by Alien Species. Front. Ecol. Evol. 7:355. doi: 10.3389/fevo.2019.00355
Received: 01 August 2019; Accepted: 09 September 2019;
Published: 24 September 2019.
Edited by:
Laurentiu Rozylowicz, University of Bucharest, RomaniaReviewed by:
Lucian Parvulescu, West University of Timişoara, RomaniaCopyright © 2019 Haubrock, Balzani, Azzini, Inghilesi, Veselý, Guo and Tricarico. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Phillip J. Haubrock, cGhpbGxpcC5oYXVicm9ja0BzZW5ja2VuYmVyZy5kZQ==
†ORCID: Phillip J.Haubrock orcid.org/0000-0003-2154-4341
Paride Balzani orcid.org/0000-0003-1549-7139
Elena Tricarico orcid.org/0000-0002-7392-0794
‡Joint first authors
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.