 Raymond L. Bernor
Raymond L. Bernor Omar Cirilli
Omar Cirilli Advait M. Jukar
Advait M. Jukar Richard Potts2
Richard Potts2 Maia Buskianidze
Maia Buskianidze Lorenzo Rook
Lorenzo Rook
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 29 May 2019
Sec. Paleontology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00166
This article is part of the Research Topic Examining Evolutionary Trends in Equus and its Close Relatives from Five Continents View all 14 articles
We report here ecological and morphological characterization of the main Old World Equus in North America, Asia, Europe, and Africa, by comparing the studied fossil forms with the living Equus grevyi zebra. Equus simplicidens from North America, Equus livenzovenzis, Equus stenonis, and Equus stehlini from Italy, Equus sivalensis from India, Equus cf. stenonis and a small Equus from Georgia (Caucasus), Equus oldowayensis, Equus koobiforensis, and Equus cf. tabeti from Kenya and the extant Equus grevyi are described in their cranial and dental features and are compared in morphological postcranial dimensions by means of log10 ratio analysis. The occurrence of the two horses at the Dmanisi Homo site in Georgia is reported here for the first time. Our comparative analyses allow to confirm the primitive lineage of the ancient zebras as derived from Equus simplicidens, and the successive evolution of the stenonine horses in Asia, South Asia, and Europe during the Plio—Pleistocene. The morphological analysis has reveals a clear trend in third metacarpals and third metatarsals of E. simpicidens, the small Equus from Dmanisi and E. grevyi, suggesting a close relationship between these species. The trend of the stenonine Equus from Europe and Asia confirms the possible derivation from the North America Equus simplicidens. The description of all the Old World Equus is integrated with an overview of their paleoecological context, with a referred section for each locality where these fossils were found. This contribution represents a comprehensive review of the present knowledge of the Old World Equus evolutionary history, with some new important data in deciphering the deep origin and evolution of ancient and living zebras.
The genus Equus includes extant species of horses, asses (and donkeys) and zebras. Wild Equus species live in Africa and Asia. A fundamental subdivision of Equus is caballine horses (including the domesticated horse) and non-caballine horses that include zebras and asses. African species include the zebras Equus grevyi (Ethiopia and Kenya), Equus quagga (south of Ethiopia to southern Africa), Equus zebra (southern Africa), and the African ass Equus asinus (Ethiopia, Somali, Horn of Africa) from which the domestic donkey is derived. Asian horses include Equus hemionus (Asiatic ass ranging from Iran to China) and Equus kiang (Tibetan Plateau). Equus ferus is the wild (caballine) horse of Eurasia. Bernor et al. (2010) recently reviewed the fossil and extant African horses and followed closely Groves (2002) taxonomy of living horses citing extensive conflicts between ethological, morphological, and molecular interpretations of Equus taxonomy and phylogeny. We follow traditional morphological criteria herein. Bernor et al. (2010) further drew extensively upon Churcher and Richardson's (1978) as well as Eisenmann's (1983) taxonomy of African Equus and herein we also rely upon the Equus phylogenetic interpretations of Azzaroli last presented by him in 2003 and again in Bernor et al. (2018).
Skeletal elements studied herein originate from the Smithsonian Institution (USNM), University of Florence, Italy (IGF), the Georgian National Museum (Dmanisi, D numbers), Natural History Museum, London (NHMUK), Panjab University (PU), and the National Museums of Kenya (KNM numbers). Anatomical nomenclature, osteological landmarks, and ligament attachment interpretations are according Bernor et al. (1997). Measurements are all given in millimeters and rounded to 0.1 mm. Measurement numbers (M1, M2, M3, etc.) refer to those published by Eisenmann et al. (1988) for the skulls and postcrania, reiterated and augmented for dentition by Bernor et al. (1997). In various studies, Eisenmann (see Eisenmann, 1995 for a comprehensive summary) has used log 10 ratio diagrams to evaluate differences in equid long bone proportions, in particular, as a basis for recognizing taxa and their evolutionary relationships. (Bernor et al., 2003a,b), Bernor et al. (2017, 2018), Bernor and Harris (2003), and Sun et al. (2018) have used log10 ratio diagrams for analyses of postcranial proportions which we apply herein for comparing metacarpal III (MCIII) and metatarsal (MTIII) proportions of Equus species under consideration. We use the mean values for the Hoewenegg Horse Quarry sample of Hippotherium primigenium (Bernor et al., 1997) for these log10 comparisons.
We undertake a brief description and comparison of the following Equus species which are relevant to our study of early Old World Equus. From North America we provide observations and measurements on the 3.3 Ma sample of Equus simplicidens from the Hagerman Horse Quarry, Idaho. From Eurasia and Africa: Equus cf. livenzovensis, E. stenonis, and E. stehlini from Italy with comments on the earliest small Equus from Italy; Equus sivalensis from India with comments on the earliest small Equus from the Indian Subcontinent; Equus cf. stenonis from Dmanisi, Georgia; Equus “oldowayensis,” E. koobiforensis and E.cf. tabeti from Kenya; extant African Equus grevyi. Figure 1 includes the localities under consideration. Figure 2 is a chronology of the localities under consideration.
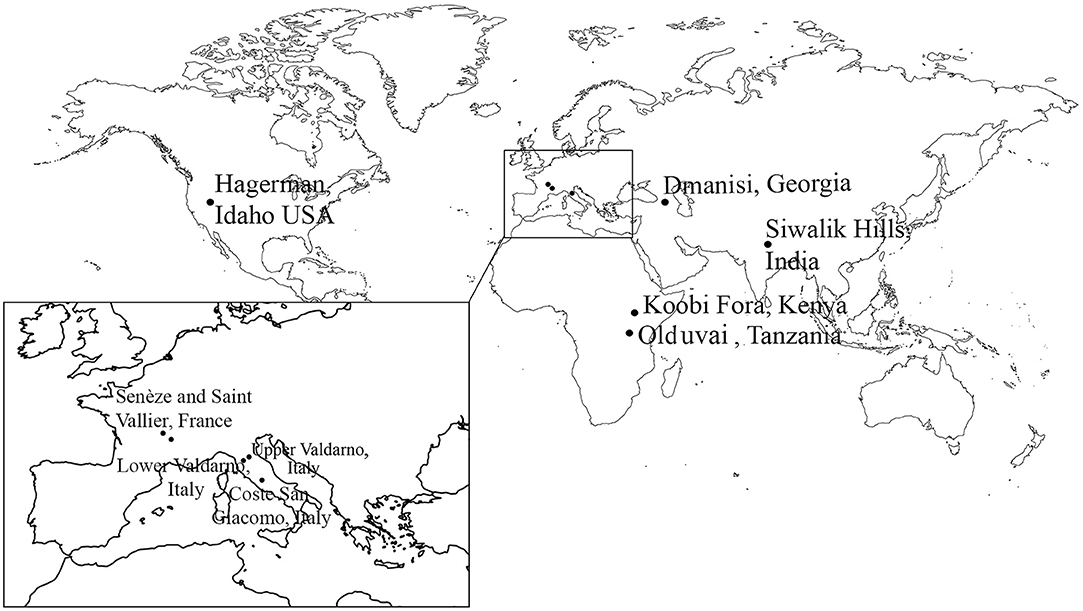
Figure 1. Map of the localities discussed in the text.
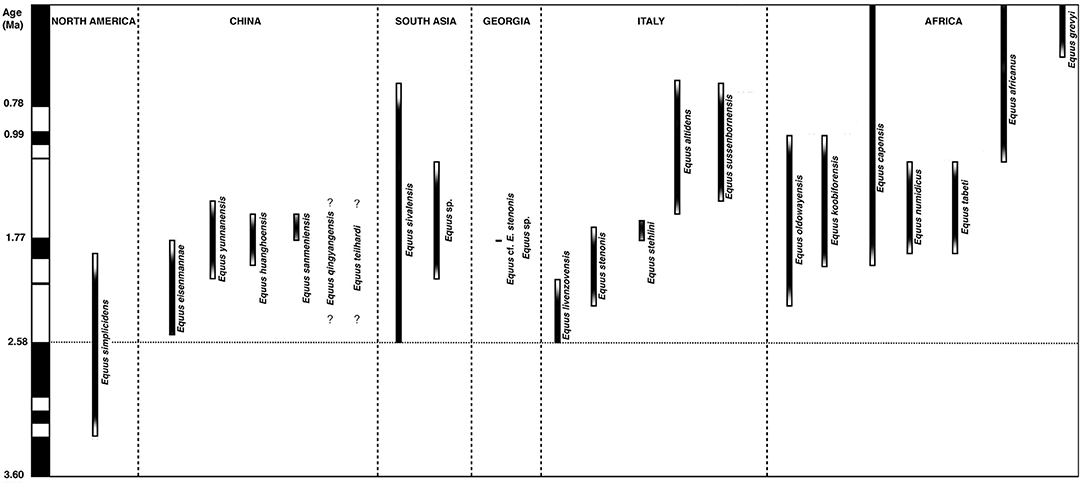
Figure 2. Chronology of the taxa in North America, Asia, South Asia, Georgia, Italy, and Africa.
The Hagerman Horse Quarry, Glenns Ferry Formation, is within the Hagerman Fossil Beds National Monument located west of Hagerman, Idaho (McDonald, 1996). The Hagerman is an extremely rich deposit discovered by a local rancher, Elmer Cook in the late 1920s. He reported this find to Dr. Harold T. Steam of the United States Geological survey who in turn reported it to Dr. James W. Gidley of the Smithsonian Institution. The Smithsonian Institution's field crew excavated three quarry beds during 1929–1931 and 1934 (McDonald, 1996; Richmond et al., 2002). Gidley (1930) published the first description of the Hagerman Horse Quarry sample as Plesippus shoshonensis and followed by Gazin (1936) who described and illustrated samples of the dentitions of various ontogenetic stages. Skinner and Hibbard (1972) assigned the Hagerman horse to the taxon Equus simplicidens which we follow herein. McDonald (1996) undertook a demographic study of the Hagerman Horse quarry. The Smithsonian Institution accumulated the largest sample of the Hagerman horse which is currently under study by us.

Figure 3. Equus simplicidens. (A) Skull in lateral view; (B) Skull in ventral view; (C) Mandible in lateral view; (D) Upper tooth row in occlusal view; (E) Lower tooth row in occlusal view.
Figures 3A–C is of an adult male skull and mandible of Equus simplicidens, USNM12573. In lateral view (Figure 3A) the skull is somewhat deformed but would appear to have a preorbital fossa well anterior to the orbit; nasal notch is retracted to mesostyle of P2, infraorbital foramen is placed high on the maxilla reflecting the horses great cheek tooth crown height. In ventral view (Figure 3B) the snout is elongate, with arcuate incisor arcade, canine is large indicative of a male individual. The maxillary cheek teeth (Figure 3D) have P2 with relatively short anterostyle; M3 is just coming into wear; protocones are elongate, mostly flattened lingually and on P2-M1 are connected to the protoloph; mesostyles are pointed. Figure 3C is the mandible in lateral view. Figure 3E renders the left cheek tooth row in occlusal view with the following salient features: p2 anterostyle is short; metaconids and metastylids are predominantly rounded; linguaflexid on premolars is a shallow V-shape, being deeper V to U-shape on the molars; ectoflexids are shallow on p2 and p3, deeper on p4 and extend very deeply into the metaconid-metastylid isthmus on m1-m3. Azzaroli and Voorhies (1993) have proposed that North American Equus simplicidens is the source for first occurring Old World Equus.
Bernor et al. (2018) reported the first occurring Equus in Italy at the locality of Montopoli, ca. 2.6 Ma which they referred to E. cf. livenzovensis. This sample is represented by very large anterior and posterior 1st and 2nd phalanges III closely comparable to E. livenzovensis from Khapry, near the Azov Sea and a sand pit at Livensovka near Rostov on Don (Bajgusheva, 1971, 1978). Azzaroli (1982) noted that the skulls of Equus livenzovensis are somewhat larger than E. namadicus and E. stenonis, but otherwise similar. Azzaroli (1982) has reported that the Khapry skull has a well developed preorbital fossa (POF). Bernor et al. (2018) described a large anterior and posterior 1st and 2nd phalanges III Equus cf. livenzovensis from the 2.6 Ma locality of Montopoli and reported that they were larger than a large suite of Eurasian, African and North American Equus, validating the species identification.
Equus stenonis type skull (IGF 560) comes from Terranova (Arezzo, Tuscany), a 1.8 Ma fossil locality in the Upper Valdarno basin in Central Italy, housed in the Natural History Museum in Florence. Cocchi (1867) named the new species and the holotype was described by Forsyth Major (1877–1880) and later, in much detail, by Azzaroli (1964). The oldest occurrence of Equus stenonis in Europe is recorded in France, from the fossil locality of Saint Vallier, ca. 2.2 Ma (Eisenmann, 2004); its chronologic range is known until Tasso FU, 1.6 Ma. The Upper Valdarno basin yielded a relevant number of specimens referable to Equus stenonis, from various fossil localities; one of these, Matassino, located near the town of Figline (Arezzo), has been calibrated as being 1.8 Ma (Napoleone et al., 2001), and the fossil horse collection from this site shows features and proportions of Equus stenonis (Mazza et al., 2004). This locality, both with the type skull from Terranova (Azzaroli, 1964) and the other specimens from Olivola (Val di Magra basin; De Giuli, 1972), demonstrate the presence of Equus stenonis in central Italy during the Early Pleistocene (Alberdi and Palombo, 2013). According to several researchers (Azzaroli, 1982, 2003; Alberdi et al., 1998; Forsten, 1999; Palombo and Alberdi, 2017; Palombo et al., 2017), the primitive lineage of Equus livenzovenzis co-occurs with Equus stenonis in Europe. Equus stenonis has some morphological features close to Equus livenzovenisis, but is a slightly smaller in size. The European Middle to Late Villafranchian horses referred to Equus stenonis differ in size and body proportions, even if they show a similar bauplan. For this reason, several subspecies have been proposed, including: Equus stenonis livenzovenzis, E. s. stenonis, E. s. olivolanus, E. s. vireti, E. s. pueblensis, E. s. guthi, E. s. senezensis, E. s. mygdoniensis. The validity of these subspecies is matter of debate (re: Forsten, 1999).

Figure 4. Equus stenonis. (A) Skull in lateral view; (B) Skull in ventral view; (C) Mandible in lateral view; (D) Upper tooth row in occlusal view; (E) Lower tooth row in occlusal view.
The type specimen of Equus stenonis is IGF560, a skull with an associated mandible. The original mandible is missing and in its place is a plaster cast painted to match the color of the original skull. The skull is viewed from its left side (Figure 4A) which is the least distorted, but nevertheless crushed in the maxillary region. The skull has an elongate snout with a large canine indicative of a male individual. The nasal bones are retracted to the level of P3 mesostyle. There is a faintly delimited preorbital fossa depression on the postero-superior aspect of the maxilla. The facial maxillary crest is strongly developed. In ventral view (Figure 4B) the snout is elongate with an arcuate incisor arcade. In occlusal view the incisors have well demarcated infundibula that are open. The maxillary cheek teeth (Figure 4D), are inclusive of dP1-M3 and M3 is in wear; dP1 is remarkably still in place being well in wear. The maxillary cheek teeth have the following salient features in occlusal view as seen on the left side row: dP1 is small and rounded; P2 has a short anterostyle; all cheek teeth have protocone short with linkage to the protoloph; pli caballins are single on all cheek teeth; fossettes are well developed on P2–P4 and M2, lesser developed on M1 and M3; hypoglyph is moderately deep on P2–M2 and not expressed on M3.
The mandible (cast) is complete on the left side (Figure 4C). In occlusal view (Figure 4E) the incisors are complete, worn, have infundibula, are slightly procumbent and form an arcuate arcade. The mandibular canine is placed very close to i3 and is large as in the maxilla. The left side occlusal view of the cheek teeth reveals the following salient features: p2 has a short anterostyle; whereas metaconids are rounded, metastylids exhibit some distal squaring; linguaflexids are uniformly V-shaped as is diagnostic of stenonine horses; ectoflexid is shallowest on p2, somewhat deeper in p3 and p4, and deeper on the molars, but not separating metaconid and metastylid; pre- and postfossettes for the most part have simple enamel margins.
Equus stehlini is the second most common horse occurring in the Upper Valdarno deposits after Equus stenonis. The species is represented by the type skull housed in the Natural History Museum in Florence (IGF 563); it comes from the same level as the Equus stenonis Type skull (IGF 560). The occurrence of a smaller species associated with E. stenonis in the Upper Valdarno was recognized for the first time by Stehlin and Graziosi (1935) basing their observation on a jaw found in the locality of Terranova (and kept in the Basel Naturhistorisches Museum collection), which they ascribed to Equus cf. hydruntinus. Later, Azzaroli (1964) formalized the species Equus stehlini based on the IGF 563 skull and on several postcranial specimens. This taxon is less common than Equus stenonis in Upper Valdarno even if is the only horse species that characterize the Casa Frata locality, which is correlative with the Tasso Faunal Unit, 1.6 Ma (De Giuli and Masini, 1986). The presence of Equus stehlini is documented in the Upper Valdarno basin (Azzaroli, 1964, 1982; De Giuli and Masini, 1986; Alberdi and Palombo, 2013), its phylogenetic relationships are not yet fully understood. Azzaroli (1964) recognized some asinine and stenonine features on the type skull and interpreted this species as derived from Equus stenonis. Later (Azzaroli, 1990), noted that Equus stehlini shares many cranial specializations with Equus senezensis from the French fossil locality of Senèze. Alberdi et al. (1998) considered this horse as a subspecies of Equus senezensis. De Giuli (1972) and Delson et al. (2006) both reported the presence of Equus stehlini from Senèze in association with Equus senezensis.

Figure 5. Equus stehlini. (A) Skull in lateral view; (B) Skull in ventral view; (C) MCIII in dorsal view; (D) Forelimb in lateral view.
Recently, Palombo et al. (2017) described a small horse from the 2.1 Ma locality of Coste San Giacomo, Central Italy (Bellucci et al., 2012, 2014) as Equus senezensis aff. Equus senezensis stehlini. The origin and evolution of small horses in Europe, including Equus stehlini, is not yet well understood. The recent findings at Coste San Giacomo extends back in time the presence of these small horses, suggesting a different interpretation of their evolution, in contrast to the old theories that they originated from the “Equus stenonis group.”
IGF563 is the type skull of Equus stehlini, which is an old adult individual. In lateral view (Figure 5A), the skull has a complete snout and orbit and is missing the posterior braincase. The nasal notch is as in E. stenonis being retracted to P3 mesostyle. The maxilla is crushed and may have had a faintly developed pre-orbital fossa high and posteriorly placed on the maxilla. In lateral view, the cheek teeth can be seen to be very worn and the cusps are blunt from wear. In ventral view (Figure 5B) the snout is relatively short and broad. The incisor arcade is arcuate and incisors are heavily worn not preserving infundibula; right and left I1 are severely worn with broken buccal margins. On both the right and left side, the occlusal surfaces are so worn as to not preserve any meaningful morphological details except on M2 and M3 that have simple fossette ornamentation.
The MCIII (Figure 5C) is elongate and slender, as is the anterior first phalanx. Figure 5D is an image of a forelimb (lateral view) referred to Equus stehlini.
Upper Valdarno small horses are found from Tasso FU (1.6 Ma), where Equus stehlini has previously been recognized (Azzaroli, 1964; De Giuli and Masini, 1986). The recent discovery of a small horse in the site of Coste San Giacomo (ca. 2.1 Ma) reopened speculation about the origin and evolution of these small horses (Palombo et al., 2017). Our ongoing review of the Villafranchian horses housed in the collection of the Natural History Museum in Florence, is revealing some important information in this regard. An important discovery by OC is an incomplete third metatarsal (IGF 566) from Montecarlo (a site dated 2.4–2.2 Ma; Ghinassi et al., 2005), reported here for the first time. The Montecarlo locality has yielded few fossil vertebrates specimens, fishes, small mammals ascribed to Mimomys polonicus and gomphothere remains (Anancus arvernensis). This new discovery allows us to increase the faunal assemblage from this locality and share new insights into the distribution of early middle Villafranchian horses. This specimen has been catalogued as an MTIII and referred to “Equus stenonis.” Its metric dimensions and morphological comparison reveals that it is similar to the Equus stehlini sample of Casa Frata, and smaller than Equus stenonis from Olivola and Matassino (M3 = 30.8; M4 = 30; M5 = 46.7; M6 = 39; M7 = 32.7; M8 = 11.1. Eisenmann et al., 1988; Bernor et al., 1997). The attribution of IGF 566 is not yet formalized and more data from Equus stenonis and Equus stehlini are needed to compare this specimen with these two Italian Villafranchian species, however this new finding is congruent with the occurrence of a small horse from Coste San Giacomo (Palombo et al., 2017).
The first occurrence of these small horses seems to be earlier than 1.6, in contrast with the classical view of Equus stehlini as the horse indicator of Tasso Faunal Unit (1.6 Ma); this hypothesis needs to be tested with more data and analysis of coeval localities, such as Senèze and Saint Vallier.
Equus sivalensis is a large species of horse found in the Siwaliks of the Indian Subcontinent, ranging from the Potwar Plateau in the west to the Nepal Siwaliks in the east. It was first discovered by Hugh Falconer and Proby Cautley in the foothills of the Himalayas between the rivers Yamuna and Sutlej in the 1830s (Falconer and Cautley, 1849). The type series, which includes 27 cranial and postcranial specimens housed at the Natural History Museum, London (Falconer and Cautley, 1849; Lydekker, 1886). Other referred material is housed at the American Museum of Natural History (Colbert, 1935b), Naturalis Biodiversity Center (Hooijer, 1951), Wadia Institute of Himalayan Geology (Nanda, 2015), Panjab University (Sahni and Khan, 1988), Indian Museum (Lydekker, 1885), Saketi Fossil Park (Mishra et al., 2013), Geological Survey of Pakistan (Dennell, 2004), Science and Art Museum, Dublin (Lydekker, 1884), and the Museum of Natural History, Ludlow (Baker, 1850).

Figure 6. Equus sivalensis. (A) Skull in lateral view; (B) Skull in ventral view; (C) Mandible in lateral view; (D) Upper tooth row in occlusal view; (E) Lower tooth row in occlusal view.
Falconer and Walker (1859) described a few specimens housed in museum of the Asiatic Society of Bengal (collection now housed in the Indian Museum) but never formally described the species. Lydekker (1882, 1886) described these species in the Palaeontologia Indica and Catalogue of Fossil Mammalia in the British Museum (Natural History). When Falconer and Cautley documented Equus from the Subcontinent, they named three species: Equus sivalensis from the Siwaliks, and Equus namadicus and Equus palaeonus from the Narmada Valley. Because the holotype of Equus palaeonus was a series of lower milk molars, Lydekker (1882) synonymized this species with E. namadicus. Matthew (1929) provisionally considered E. namadicus as a junior synonym of E. sivalensis or derived from it, because the distinguishing character used by Lydekker (longer protocones on both premolars and molars) was too variable, an observation reiterated by Hooijer (1951). Lydekker (1882) further stated that the small protocones on the premolars were indicative of a relationship with E. hemionus. Later, Lydekker (1891) stated that E. sivalensis was intermediate between E. stenonis and E. caballus, and was similar to early Pleistocene horses recovered from Yunnan. Similarly, Forsyth Major (1885) also closely compared E. sivalensis with E. stenonis from the Val d'Arno. In contrast, Hopwood (1936) suggested that the lectotype of E. sivalensis was a caballine horse, but erected a new species E. cautleyi based on a large partial mandibular ramus with p2-m3 (NHMUK PV OR 23107), which showed typical zebrine features (see description below). However, this view was rejected by Azzaroli (1966). Azzaroli studied the then known skulls of E. sivalensis and concluded that in the features of the orbits and zygomatic arches were typical of hippotigrine zebras rather than caballine horses. A skull (PU A/639) and mandibular rami (PU A/601 and PU A/602) described by Sahni and Khan (1961) show similar zebrine characteristics as well. These morphological observations led researchers like Badam and Tewari (1974), Sahni and Khan (1961), and Azzaroli (1966) to conclude that extinct species of Pleistocene Equus from South Asia were exclusively zebrines.
The exact stratigraphic range of E. sivalensis was ascertained several decades after the original discovery. Pilgrim (1910, 1913) determined that Equus is found in the Upper Siwaliks, and later Colbert (1935a) demonstrated that E. sivalensis would have made its first appearance in the Siwaliks during the Pleistocene in the Pinjor faunal zone. With the advent of modern paleomagnetic studies in the region, the biostratigraphy of Siwalik mammals was further refined. It is now known that Equus sivalensis first appears just above the Gauss-Matuyama boundary in the early Pleistocene (ca. 2.58 Ma), and ranges through until about 0.6 Ma, the upper end of the fossiliferous Siwalik depositional sequence (Patnaik, 2013).
NHMUK PV M.16160 is the Lectotype of Equus sivalensis Falconer and Cautley. It is a skull lacking the snout. Figure 6A shows that this specimen preserves the nasal notch that is incised to the mesostyle of P3 and there is a faint depression high dorsally and posteriorly on the maxilla for a weakly defined preorbital fossa. Figure 6B reveals that NHMUK PV M.16160 is an old individual with P3–M1 occlusal surfaces severely worn. Of these, P3–M3 preserve protocones with P4, M2, and M3 having weak connections to the protoloph and P3 and M1 having broader connections due to their more advanced wear; protocones are moderately elongate on P4–M1, longer on M2 and M3; hypoglyph is preserved on P3, P4, and M2 and is only moderately deeply incised; P4, M2, and M3 preserve fossettes that are slightly to moderately ornamented; all mesostyles are pointed buccalward. Figure 6C is a right mandible, NHMUK PV OR 23107, labial view. Figure 6E is the mandible's occlusal view and reveals the following salient features: premolars have elongate metaconid and squared metastylid; p3 and p4 have a prominent mesiolabial pli directed labialward on the preflexid; premolar linguaflexids are a deep narrow V-shape; premolar ectoflexids are all shallow, not invading the space between opposing borders of the pre- and postflexids; molar teeth likewise have rounded metaconids and squared metastylids with deep V-shaped linguaflexids; ectoflexids are as in the premolars not being deep or invading the pre- and postflexids; the premolar and molar teeth all have distinct to diminuitive pli caballinids.
Within the stratigraphic range of E. sivalensis, remains of a smaller horse with slender metapodials have also been found. Gaur and Chopra (1984a) were the first to document a smaller subspecies of equid from the Siwaliks of South Asia which they named Equus sivalensis minor, owing to its small size, but overall morphological similarity with Equus sivalensis. These fossils were found ~3.0 km N.E. of the town of Mirzapur in northwestern India in the Pinjor Formation. The exact age of the Pinjor Formation at this site is unknown, therefore the age of these specimens can only be constrained to the entire range of the formation, 2.58–0.6 Ma. However, given the age of the Pinjor Formation ~13 km away at a parallel location along the Patiali stream (Gaur and Chopra, 1984b; Ranga Rao et al., 1995), this locality is likely from the middle-upper Pinjor Formation. Subsequently, other remains of small horses were discovered on the Mangla-Samwal anticline (Hussain et al., 1992), and in the Pabbi Hills (Dennell, 2004; Dennell et al., 2006). Hussain et al. (1992) document both a large horse, which they refer to as Equus cf. sivalensis which ranges in duration from 2.35 to 1.9 Ma, and a small horse at locality HGSP 4460 (ca. 2.1 Ma), which is referred to as Equus sp. A. The Pabbi Hills specimens are all referred to as Equus cf. sivalensis and the metapodials are metrically similar to the smaller horses from the Mangla-Samwal anticline (Dennell, 2004; Dennell et al., 2006). Small equid remains are also present in the Falconer and Cautley collection at the Natural History Museum, which includes the type series of E. sivalensis (Lydekker, 1882; Forsten and Sharapov, 2000). However, a detailed morphometric analysis (beyond the scope of this paper) is necessary to determine the taxonomic identity and relationships of this smaller taxon.
The site of Dmanisi is located about 65 km southwest of Tbilisi (Georgia, South Caucasus), in the Kvemo Kartli region (Gabunia et al., 2000). Today the site is situated at an elevation of 910 m MSL on a promontory that is isolated on two sides by the deeply entrenched Mashavera and Pinesauri rivers. Radiometric ages and paleomagnetic data provide a chronology for all the deposits at Dmanisi bracketing the age of the fossiliferous deposits in a chronologic range between 1.85 and 1.77 Ma (Ferring et al., 2011). Well-preserved bones have been recovered from throughout the succession although the great majority are from stratum “B1.” The latter has a complex, very brief, depositional history and has yielded an exceptionally well-preserved fauna occurring as dense accumulations. In addition to its unique paleoanthropological record, Dmanisi is one of the richest Early Pleistocene palaeontological sites of Eurasia's temperate zone. More than three decades of excavation have uncovered a highly diverse fossil vertebrate fauna with 49 taxa, inclusive of Homo. The large mammal record from Dmanisi indicates a biochronology positioned at the very early Late Villafranchian (Vekua, 1995; Lordkipanidze et al., 2007; Rook and Martínez-Navarro, 2010), while the occurrence of the water vole Mimomys pliocaenicus correlates with the Late Villanyan of the micromammal biochronology (Fejfar et al., 1998). The Equus collection is sizeable and growing annually from an active excavation program. Our observations herein are strictly preliminary and intended to give some early insight into the equids present at Dmanisi.

Figure 7. Dmanisi Equus. (A) Skull in lateral view; (B) Skull in ventral view; (C) Mandible in lateral view; (D) Upper tooth row in occlusal view; (E) Lower tooth row in occlusal view.
D353 (Figure 7A) is a partial skull lacking the snout, nasals, posterior orbit, and posterior cranium. One cannot determine the nasal notch position for this individual. There is no discernible preorbital fossa for this individual. Figure 7B illustrates an occlusal view of specimen D66, an adult individual with M3 in early wear. This specimen has a very small, worn right dP1 with a socket still present for a missing right dP1. Salient features of the remaining cheek teeth include: P2 has an elongate, bucco-lingually constricted anterostyle; protocone on P2 is short and rounded, whereas P3 and P4 are progressively longer; M1 protocone is as in P4, whereas M2 is longer and M3 is long but labiolingually compressed; protocones are lingually flattened on all cheek teeth and P3–M2 prefossettes have a distinct lingually directed mesial pli; protocone has a weak connection to the protoloph on P2, P3, M1, and M3, but not so on P4, M2; pli caballin is only clearly expressed on M1; hypoglyph is deeply incised on all cheek teeth encircling the hypocone on P3 and P4; mesostyles are pointed on M2 and M3 and more rounded on P2-M1.
Churcher and Hooijer (1980) reviewed the taxonomy of E. oldowayensis which Bernor et al. (2010) closely followed. Hopwood (1937: Figures 1, 2) designated a lower jaw from an animal about 2 years old (Catalogue Number VIII, 353 m in the Bayerische Palaeontologische Staatsammlung, Munich) as the holotype of E. oldawayensis. Hopwood (1937) also designated a lower incisive region with left incisors and right i1 (BMNH14199) as the paratype. The type was destroyed by bombing in World War II leaving the paratype to represent the species; this in itself, is an inadequate element for species recognition in horses.
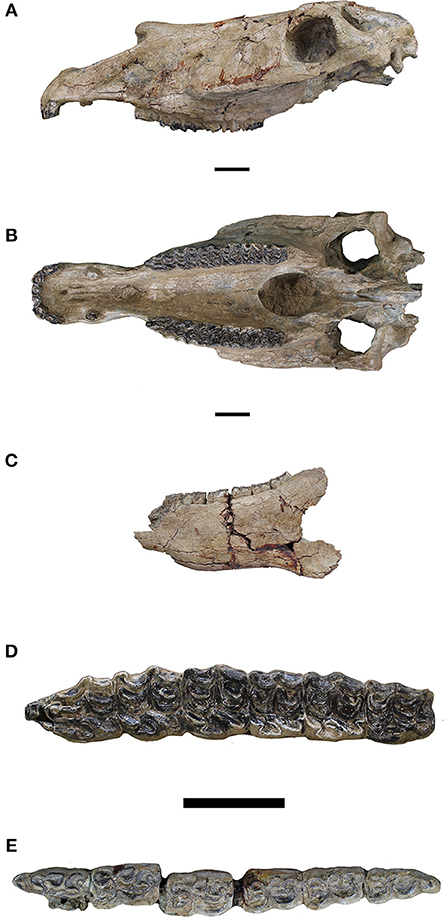
Figure 8. Equus oldowayensis. (A) Skull in lateral view; (B) Skull in ventral view; (C) Mandible in lateral view; (D) Upper tooth row in occlusal view; (E) Lower tooth row in occlusal view.
Cooke (1963: Table 4) assigned a large equid from the Omo deposits to E. oldowayensis (Hopwood, 1937), or possibly Equus grevyi, but without discussion. Hooijer (1976) reported a large Equus from the Omo that he considered to be indeterminate. Churcher and Richardson (1978) identified Equus (Dolichohippus) oldowayensis from the Omo Shungura F through J explicitly recognizing it as being a member of the Grevy's zebra clade. Coppens (1971) earlier recognized that the earliest Equus occurred in Omo Shungura G (not as early as member F), ca. 2.3 Ma. which is our current understanding (J.-R. Boissierie, personal communication to Bernor) Churcher and Hooijer (1980) reported an extensive Equus material currently on loan to the Rijksmuseum van Natuurlijk Historie Leiden and nominated a mandible, BMNHM14184, as a Neotype. Eisenmann (1983) noted the likely similarity of Koobi Fora Equus koobiforensis to Equus oldwayensis. Bernor et al. (2010) followed these arguments and in review again here we find that while E. koobiforensis and E. oldowayensis may overlap in their morphology and temporal distribution, Equus oldowayensis presents a very confusing and uncertain taxonomy whereas Equus koobiforensis is clearly characterized and defined and brings stability to our understanding of this large East African horse. We agree with Eisenmann's decision to characterize and define Equus koobiforensis as a legitimate species. Clearly, the Olduvai and Omo Equus material needs a comprehensive revision with the benefit of Eisenmann's (1983) study of the Koobi Fora Equus assemblage.
Our clearest understanding of the morphological basis for Equus oldowayensis comes from Churcher and Hooijer's (1980) diagnosis of Equus oldowayensis, the salient features are summarized after them briefly herein: Equus (Dolichohippus) oldowayensis is a large horse overlapping in size with Equus grevyi; with a large, broad, incisor arcade; infundibula present on I1 and I2, possibly absent on I3; canine large in males, may be absent in females; P2 and P3 protocones shorter and rounder than on P4 and persistently longer on molars; protocone strongly linked to protoloph; pli caballins vestigial or absent; fossette plications simple. Mandibular incisors as in maxillary incisors having distinct infundibula; lower cheek teeth have vestigial metaconid on p2 and rounded or slightly elongate on p3-m3; metastylids are rounded to square shaped; linguaflexids very shallow on p2 and V-shaped on p3-m3. Metapodial IIIs reportedly variable in length and slenderness. Bernor et al. (2010) reviewed these characters and particularly in the case of metapodial III variability, suggested that there is likely more than one Equus species represented in this sample. Eisenmann (1983) recognized species specific variability in the Koobi Fora equid assemblage so it is reasonable to expect similar variability in the Olduvai and Omo assemblages. A relatively complete cranium (Figure 8) and mandibular specimens from upper Member 1 Olorgesailie Formation, ca. 1.0 Ma, have also been assigned provisionally to E. oldowayensis based large overall size, a broad incisor arcade, and a P2–M3 tooth row longer than in male E. grevyi (Potts, 1998; Potts et al., 2018).
Olorgesailie, Kenya has yielded a 1.0 Ma skull referred to herein as Equus “oldowayensis” (KNMOG22833; Figures 8A–C). The skull is in pristine condition and in lateral view (Figure 8A) exhibits a moderately long snout with nasal notch retracted to P2 mesostyle, preorbital fossa is lacking; broken canines are large revealing that this individual was male; P2–M3 are clearly worn and all paracone cusps are extremely low and blunt suggesting an abrasive diet likely composed largely of grass. In ventral view (Figure 8B) the incisor arcade is arcuate with right I1 having a broken buccal margin and all incisors and canines being heavily worn; incisors have round infundibula occlusally; no dP1 is present. Figure 8D provides a closeup of P2–M3; P2 has an elongate anterostyle; P2, P3, M1, and M2 have protocone strongly connected to protoloph, P4 and M3, less so; hypoglyphs range from being slightly incised on P2–M2, being more strongly incised on M3; all protocones are flattened lingually; protocones show slight serial increase in length from P2–M3; all pre- and postfossettes have slightly to non-plicated margins; mesostyle is rounded on P2–M2 and pointed labially on M3; pli caballins are not apparent on P2–M3. Figure 8C is a partial mandible in lateral view, and discovered within the same stratum as the cranium, ~80 m away Figure 8E is a closeup of p2-m3 with the following salient features: the cheek teeth are typically stenonine/zebrine with p2-m3 having rounded metaconid, squared, and often distolingually pointing metastylid; shallow linguaflexid on p2 and V-shaped on p3-m3; preflexid has a strong, labially directed pli on the mesiolabial border; pre- and postflexids are distinct and separated except for the p4 postflexid which is highly complex labially, connecting mesially with the metaconid; ectoflexid is shallow on p2-p4 and deep separating metaconid from metastylid on m1-m3; pli caballinids are absent on all cheek teeth.
Eisenmann (1983: 178) named Equus koobiforensis for a skull of a young mare, KNM-ER1484 originating from the Notochoerus scotti zone, below the KBS Tuff, in Area 130. Other specimens were reported from the Notochoerus scotti and Metridiochoerus andrewsi zones of Koobi Fora and questionably from the Omo Shungura Formation and Olduvai Gorge.

Figure 9. Equus koobiforensis. (A) Skull in lateral view; (B) Skull in ventral view; (C) Upper tooth row in occlusal view.
The skull is large with P2–M3 length being 202 mm. In lateral view (Figure 9A) the skull has a moderate length snout, nasal notch is retracted to mesostyle of P3 and there is no apparent depression for a preorbital fossa. The lacrimal bone is clearly defined anteriorly by its suture. The facial-maxillary crest is modestly developed. In ventral view (Figure 9B) the right half of the snout is broken away with only left I2 and I3 in place with open infundibula. The left premaxilla has a very small alveolus for the diminutive female canine. The maxillary cheek tooth dentition has the following salient features: P2 with a short anterostyle and short ovate protocone not connected to protoloph; P3 is triangular shape with a slight connection to protoloph and protocone is likewise triangular shaped with no connection to the protoloph; P2 has a labially squared mesostyle, while P3 and P4 have mesiodistally constricted and labially pointed mesostyles; all premolars have weakly developed fossette ornamentation; P2 has a moderately deeply incised hypoglyph and P3 and P4 have deeply incised hypoglyphs; M1–M3 have elongate, lingually flattened protocones with only M1 having a clear connection to the protoloph; all molars have labially pointed mesostyles and weakly developed fossette ornamentation.
Eisenmann (1983, p. 179–180) reported one lower cheek tooth series that she found matched the holotype cranium, KNM-ER4051, which she characterized as having teeth like those of E. stenonis with rounded metaconids and metastylids, deep and pointed lingual grooves (= linguaflexids), short vestibular grooves (= ectoflexids) on at least some of the molars, and a relatively long p2. The presence of a protostylid on p2 evokes a comparison with E. grevyi but the shallow nature of the vestibular grooves is rare in the later species.
Eisenmann (1983, p. 188) referred a smaller Equus skull, KNM-ER 1211 of a young adult (Figure 10 herein) to Equus cf. tabeti. As she pointed out: “E. tabeti is a North African early Pleistocene species of moderate size, mostly characterized by slender limb bones, asinine upper teeth and stenonine lowers.” In lateral view (Figure 10A) the skull is seen to lack the pre-maxilla and nasals, has a broken distal right maxilla, partial orbit, and almost complete brain case. The lateral view also shows that M3 is not yet fully emerged from its crypt and hence this individual is a sub-adult. Figure 10B is a ventral view of KNM-ER 1211. Eisenmann (1983, p. 188) reports that h P2–M3 tooth row length is 169 mm. The salient features of the upper cheek teeth are: P2 with a short anterostyle, rounded protocone with lingually flattened surface and weak connection to the protoloph; P3 with longer, lingually flattened protocone, and weak connection to protoloph; P4 with still longer and flattened protocone and no connection to the protoloph. M1-M3 show increased bucco-lingual flattening serially, with M1 having a weak connection to the protoloph while M2 and M3 have no connection to the protoloph; m3 protocone lenticular shaped; hypoglyph is deep on P2, P3, and M1, being very deep on P4 and M2 and not discernible on M3 due to its very early wear stage; pli caballins are not developed on any of the cheek teeth; mesostyle is labially squared on P2 and pointed labially on P3–M3; cheek tooth fossettes are mostly weakly ornamented on all cheek teeth.

Figure 10. Equus cf. tabeti. (A) Skull in lateral view; (B) Skull in ventral view; (C) Upper tooth row in occlusal view.
Equus grevyi is the largest living zebra, exceeding the size of the plains and mountain zebras. It is the sole living member of the subgenus Dolichohippus.
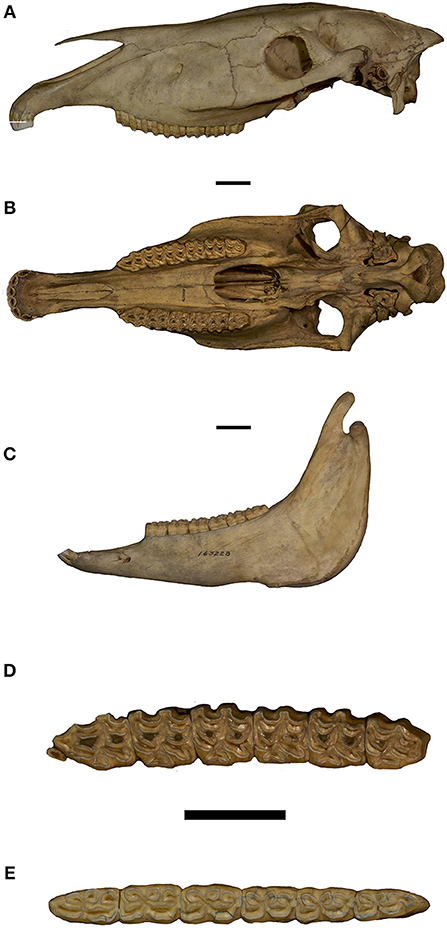
Figure 11. Equus grevyi. (A) Skull in lateral view; (B) Skull in ventral view; (C) Mandible in lateral view; (D) Upper tooth row in occlusal view; (E) Lower tooth row in occlusal view.
We characterize Equus grevyi using a pristine adult female skull with associated mandible, USNM163228. The skull has a P2–M3 dimension of 185.7 mm. The skull in lateral view (Figure 11A) has nasal notch reflected to P2 metastyle. The infraorbital foramen is prominent, placed slightly higher than midway between the occlusal level of distal P4 and the superior surface of the posterior nasal bone. The lacrimal bone is large, square shaped, and extends from the anterior orbit to the level of the mesostyle of M3. Paracone cusps of P4–M2 are low and blunt and reflect a grazing diet. In ventral view (Figure 11B) the snout is elongate with a rounded incisor arcade; all incisors have distinct, open infundibulae; canines are small and just emerging from their crypts. The left cheek tooth series (Figure 11D) with dP1 small and worn; P2–M2 have mesostyles squared labially; all maxillary cheek teeth have moderately to poorly developed pre- and postfossette plis; pli caballins are absent on all cheek teeth; P2 has a moderately elongate, mesially rounded anterostyle; P2 protocone is the shortest, P3-M3 being longer; P3-M3 protocones are longer than P2 and have a flattened lingual border with a buccally-directed inflection midway along its margin.
As seen in the lateral view (Figure 11C) the mandible is pristine, incisors are moderately procumbent, canine is lacking and symphysis is moderately long. The left p2-m3 cheek tooth series (Figure 11E) has p2 with short anterostylid; premolars have rounded metaconids, and square metastylids, all mandibular premolars have a V-shaped linguaflexid; ectoflexids are very shallow on p2, and deeper on p3-4, but not coursing between the pre- and postflexids; p2-p4 have pli caballinids that become reduced serially through the premolar series; pre-and postflexid margins are mostly not plicated; p3 and p4 have a labially directed pli on their mesio-labial margin. Mandibular molars have metaconids and metastylids as in the premolars; linguaflexid is U-shaped on m1 but as in the premolars V-shaped for m2 and m3; ectoflexids are deeply inflected separating metaconid and metastylid on all of the molars; the molars lack pli caballinids; pre- and postflexids are smaller, have simple margins and are more labio-lingually compressed than in the premolars and have a weakly developed mesio-buccal pli. The lower cheek teeth retain the so-called stenonine occlusal pattern, especially with regards to metaconid-metastylid and linguaflexic morphologies.
Figures 12A,B are log10 ratio diagrams of metacarpal IIIs (MCIII) and metatarsal IIIs (MTIII), respectively, of many of the Equus species we have discussed in this contribution; the Hoewenegg Hippotherium primigenium log10 mean is used as our comparative sample because of its large sample size and clear proportional differences with our Equus sample.
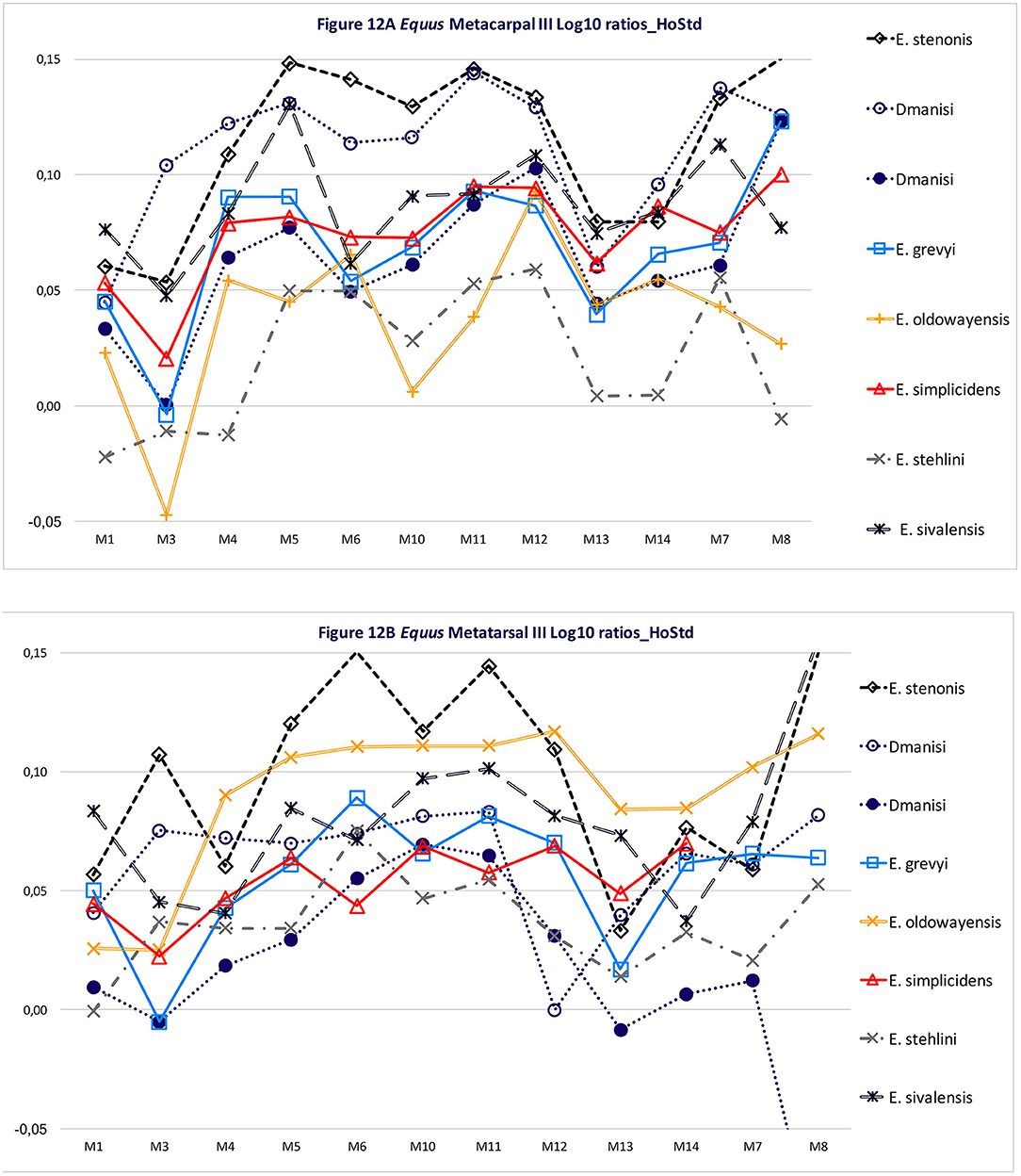
Figure 12. Log10 ratio diagrams; the specimens are analyzed on the Hoewenegg Hippotherium primigenium sample. (A) Log10 ratio diagram of third metacarpal; (B) Log10 ratio diagram of third metatarsal.
Figure 12A MCIII shows that all Equus species under consideration herein have longer MCIIIs than Hoewenegg (Ho) Hippotherium primigenium except for Equus stehlini, which is shorter. The largest (M5 and M6 proximal articular dimensions and M10 distal supra-articular dimension) horse in our sample is Equus stenonis. The large Dmanisi Equus plots closely to E. stenonis having similar midshaft (M3 and M4) dimensions, and slightly smaller proximal articular facet (M5 and M6) and trans- distal suprarticular (M10) dimensions; this supports our referral of the large Dmanisi form to Equus cf. stenonis. Equus sivalensis has the longest MCIII (M1), has strongly contrasting large midshaft width (M3) and depth (M4) dimensions as in E. stenonis and narrower trans- supraarticular (M10) dimensions but otherwise plots with species in the central portion of the figure. Equus simplicidens, E. grevyi, and the smaller Dmanisi Equus sp. are in the center of the plot and have close trajectories to one another. Equus “oldowayensis” likewise plots close to this group except for the narrow midshaft (M3) and supraarticular (M10) dimensions. Equus stehlini is the shortest MCIII and overall and is the smallest Equus in this sample but generally has a plot trajectory similar to the E. simplicidens, E. grevyi, and smaller Dmanisi Equus triad.
Figure 12B again shows that all Equus species under consideration herein have longer MTIIIs than Hoewenegg (Ho) Hippotherium primigenium except for Equus stehlini and the smaller Equus from Dmanisi which are approximately the same as each other and the Hoewenegg mean sample. The most remarkable feature of this plot, again is how close the log10 ratios are between Hagerman E. simplicidens, extant E. grevyi and the smaller form of Equus sp. from Dmanisi, except for the short length of the Dmanisi small form. Equus stenonis has a similar length as a majority of specimens, but its log10 trajectory shows large size peaks for midshaft width (M3), proximal articular depth (M6) and distal articular width (M11). Equus “oldowayensis” length (M1) and midshaft width (M3) is as in other specimens, but is larger approaching E. stenonis for M4-M12 measurements and has the largest relative dimensions for M12–14. Equus sivalensis is the longest MTIII of our sample and its log10 trajectory largely resembles E. simplicidens, E. grevyi and the large and small Dmanisi Equus. Equus stehlini and the smaller Dmanisi Equus have the shortest MCIIIs. This log10 ratio diagram on MTIII supports the interpretation than E. stehlini is in fact a different species than E. stenonis, and that our referral of the larger Dmanisi Equus to E. cf. stenonis is warranted on skull, dental and now MCIII and MTIII grounds. Our limited sample of Equus “oldowayensis” suggests that this “species” is somewhat shorter (M1), has wider midshaft (M3), and distal width (both M10 and M11) than E. grevyi, E. simplicidens and both Dmanisi Equus sample; a survey of a larger sample of E. “oldowayensis” is needed.
The fossils from the Hagerman beds are best known from the Hagerman Fossil Beds National Monument, Idaho (McDonald et al., 1996). These beds are located on the Snake River Plain, and are currently under the protection of the U.S. National Park Service. The beds are part of the Glenns Ferry Formation (Repenning et al., 1995), and consist of fluvial, flood plain, and lacustrine deposits (Ruez Jr., 2009a). The age of the beds exposed in the National Monument range from 4.18 to 3.11 Ma (Ruez Jr., 2009a), and corresponds to the Blancan Land Mammal Age (Bell et al., 2004). The Hagerman Horse Quarry is arguably the most famous fossil locality in the beds, where scores of Equus simplicidens remains have been recovered (Gazin, 1936). This quarry is roughly 3.19 Ma in age. It is thought that either a flood killed a herd of horses and subsequently buried them (McDonald, 1996), or drought stricken horses died at a dwindling watering hole and were then subsequently buried by a flash flood (Richmond and McDonald, 1998). The paleoclimate at the National Monument is interpreted to have been warm and wet, with a short cool interval between 3.6 and 3.2 Ma (Ruez Jr., 2006). The climate of the Hagerman Horse Quarry is interpreted to be very warm, providing some support for the drought hypothesis (Ruez Jr., 2006). A diverse assemblage of mammals has been recovered from the Hagerman beds, including a mix of small and large taxa from various guilds (Ruez Jr., 2009b).
Palaeoecological data, from the comparison of Montopoli fauna (the site recording the Equus Datum in Italy) with other Plio-Pleistocene mammal assemblages of Europe, show that the Equus event is related to the faunal turnover and palaeoenvironmental change that occurred at the early–to–middle Villafranchian transition, chronologically corresponding to 2.6 Ma. Furthermore this time coincides with one of the major shifts in δ18O isotopic oscillations (cf. Miller et al., 2005), and the setting of 40 Ka obliquity-forced thermal glacial/interglacial cyclicity (Bertini, 2010) with strong effects on seasonality, and the first record of cool winters and drier climates as testified by the diffusion of herbaceous steppe vegetation (Fortelius et al., 2006).
Several paleoecological analyses (Pradella and Rook, 2007; Rivals and Athanassiou, 2008; Valli and Palombo, 2008; Strani et al., 2015, 2018) stress a trend toward more open environments, both in terms of taxonomical composition (at family and genus levels), ungulate hypsodonty, and dietary categories. This is consistent with the faunal turnover known as “Elephant-Equus event” (Azzaroli, 1983), that occurred at 2.5 Ma in Europe, an event that produced a faunal composition with predominance of grazing species and grass-dominated mixed feeders, among which the most representative taxa are the monodactyl horse, Equus cf. livenzovensis, and the first true elephant, the primitive Mammuthus gromovi, and the disappearance of the extinction of numerous woodland dwellers like Mesopithecus monspessulanus, Mammut borsoni, Tapirus arvernensis, Sus minor, and Ursus minimus (Azzaroli et al., 1986; Eronen and Rook, 2004; Pradella and Rook, 2007).
The large mammal record from Dmanisi includes several of the herbivore faunal elements also known from Middle Villafranchian contexts of Western Asia and Europe, but a significant number of genera, especially of artiodactyls, reflect the early Late Villafranchian faunal renewal, the composition of the carnivore guild also points to an early Late Villafranchian age (Vekua, 1995; Lordkipanidze et al., 2007), correlative with a geochronologic age range between 1.85 and 1.77 Ma (Ferring et al., 2011). Overall the large mammal assemblage is characterized by a preeminence of open woodland adapted herbivores. The small mammal association is dominated by steppic or dry-adapted rodents, while woodland with fluviatile elements are very rare (Lordkipanidze et al., 2007; Furió et al., 2010). The herpetofaunal assemblage from Dmanisi indicates an important water stress environment suggesting a period of increased aridity and supports a reconstruction of open, relatively dry environments and climate as dry and warm as the Mediterranean type (Blain et al., 2014). The Dmanisi mammal fauna points to a mosaic-like landscape with remarkable differences in humidity and vegetation across its terrain. Although the immediate vicinity of the fossil site was situated in a more or less forested valley, the wider region was largely made up of drier open tree savanna and grassland and by mountainous to semi-arid rocky terrains.
Siwalik Equus is known to be a grazer based on the flat occlusal surfaces (also, low and blunt paracones of P4–M2 mesowear terminology) and enamel isotopic data (Patnaik, 2015). Carbon isotopes from dental enamel and soil carbonates show the presence of grasslands in the Siwaliks during the Pleistocene, albeit with patches of woodland within grasslands (Quade et al., 1993; Quade and Cerling, 1995; Patnaik, 2015). Equus appears in the Siwaliks along with other hypsodont grazers such as the alcelaphine Damalops palaeindicus, the reduncine Sivacobus palaeindicus, and bovines Bos acutifrons, Bubalus platyceros, Leptobos falconeri, and Hemibos triqueticornis and the elephantid Elephas hysudricus and mesodont mixed feeders such as cervids and rhinocerotids (Jukar, 2018). Equus also replaces the smaller and less hypsodont hipparionines across the Plio-Pleistocene boundary (Jukar, 2018). The causes of this ecological replacement are still unclear since both hipparionines and Equus were adapted to a grazing lifestyle in open habitats.
The first record of East African Equidae, the C4-grazing Eurygnathohippus, coincided with a sharp 3.1%0 enrichment in δ13C values connoting C4 grassland expansion at ca.10 Ma, based on n-C35 alkane terrestrial plant wax biomarkers recovered from Somali Basin and Red Sea drill cores (Uno et al., 2016). Pedogenic carbonate δ13C values across sub-Saharan African fossil sites suggest, however, that C4 grasses were a minor component of the environment until the early Pleistocene (Ségalen et al., 2007). The earliest appearance of Equus in East Africa, currently dated at 2.3 Ma in the base of Member G Shungura Formation of the Omo-Turkana basin, and its subsequent numerical increase are associated with further expansion of C4 grass-dominated habitats (Cerling et al., 2015). Omo-Turkana equids, as represented by Equus and Eurygnathohippus, range from 1.5 to 11% of all large mammal specimens between 2.3 and 1.35 Ma (Bobe and Eck, 2001; Hernández Fernández and Vrba, 2006). Between 2.3 and 1.8 Ma, Equus accounts for roughly 30% of all Omo-Turkana equids, increasing to around 50% of all equids from 1.8 to 1.35 Ma (Bobe and Leakey, 2009). The appearance of Equus in the Omo-Turkana record coincided with an overall transition in mammalian herbivore diets as a C4-grazer-dominated fauna supplanted an herbivore community of C3-C4 mixed-feeders by 2.3 Ma. Early Pleistocene Equus of Turkana was a C4 grazer with an average δ13C value of −0.1 ± 1.1%0 (n = 39), only slightly less enriched than that of extant East African Equus with an average δ13C value of 1.3 ± 1.4%0 (n = 147) (Cerling et al., 2015).
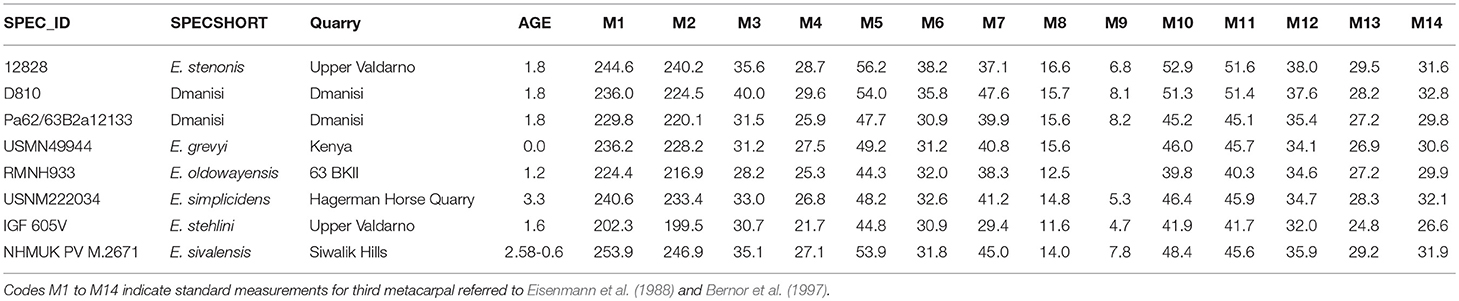
Table 1. Measurements of the specimens reported in Figure 12A.
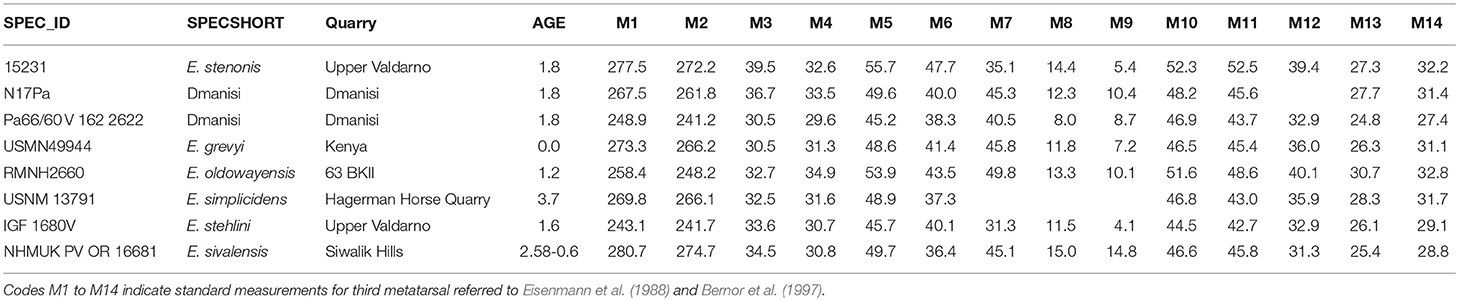
Table 2. Measurements of the specimens reported in Figure 12B.
In 2 Ma sediments at Kanjera South, Kenya, Equus accounts for 6–10% of the excavated large mammal specimens from three stratigraphic levels, all of which are associated with pedogenic carbonate δ13C values indicative of >75% C4 grassland habitat (Plummer et al., 2009; Ferraro et al., 2013). In the excavated fauna of Bed I Olduvai Gorge, Tanzania, dated ~1.9–1.8 Ma, Equus ranges from 3 to 14% of the large-bodied mammals (percent minimum number of individuals); the genus increases in representation relative to all herbivores after 1.8 Ma in the faunas of Bed II Olduvai, associated with increased pedogenic δ13C values and open habitat (Leakey, 1971; Cerling and Hay, 1986; Potts, 1988). Although the spread of grasslands has often been attributed to increased aridity, East African vegetation and moisture varied independently; instead, the transition to open habitat favoring Equus and other grazing herbivores now appears to have been more closely related to decreasing atmospheric CO2 concentration rather than climatic aridification (Blumenthal et al., 2017; Faith et al., 2018).
After 1.35 Ma, the best-preserved evidence of fossil Equus in East Africa comes from the Olorgesailie Basin, southern Kenya. Comprising 44% of the large mammals, Equus is the dominant taxon in the bovid-depauperate fauna dated ca. 1.0 Ma, and is associated with a nearly 100% C4 grassland. The relative abundance of Olorgesailie Equus decreases to 10% of the herbivore fauna after this time (Sikes et al., 1999; Potts, 2007). In modern East African game reserves, Equus typically comprises 10–25% of the large mammal fauna (data compiled in Potts, 1988). Based on faunal representation from the Omo-Turkana, Olduvai, and Olorgesailie basins, the number of non-ruminant C4-grazing ungulate genera ranged between five and ten during the interval between 2.3 and 0.6 Ma. However, most species in those large ungulate clades either became extinct or switched to browsing and mixed-feeding such that only four non-ruminant C4-grazing taxa, including Equus, currently survive in East Africa (Potts, 1988; Cerling et al., 2015; Potts et al., 2018). While a major ecological shift in African mammal communities occurred after 0.6 Ma, the resilience of Equus appears to relate to a dietary strategy that involves C4 grazing and the ability to adopt a mixed-feeding mode in situations where graze is scarce (Kaiser and Franz-Odendaal, 2004).
We have provided a brief review of Equus evolution including samples of North American Hagerman Equus simplicidens, Italian Equus stenonis, and Equus stehleni, two apparent species of Equus from Dmanisi, Georgia, Equus sivalensis from the Indian Sub-Continent and Kenyan Equus koobiforensis, Equus “oldowayensis” and Equus cf. tabeti.in comparison to extant Grevy's zebra. We consider this an initial evaluation of a restricted sample of these species skulls and dentitions meant to evaluate the plausibility of their evolutionary relatedness. We find that traditional views on the relationships of stenonine horses and zebras are upheld: that Azzaroli and Voorhies (1993) were correct that Hagerman Quarry E. simplicidens is a likely phylogenetic source for Old World stenonine horses; that McDonald (1996) was justified in referring to Hagerman horse Equus simplicidens as a zebra; that Equus grevyi and, more broadly, African zebras are the descendants of stenonine horses. We have reported remarkable similarities in skull, tooth and metapodial III log10 proportions between E. simplicidens, smaller Dmanisi Equus sp. and Equus grevyi. Both Equus simplicidens and its likely descendant Equus stenonis exhibit the basic bauplan of other Eurasian early Equus species and Equus grevyi. The revelation herein is that African zebras would appear to have a genuine deep-time origin in North American Equus through the Eurasian Equus stenonine lineage. This hypothesis needs future testing with large sample sizes of E. simplicidens, Eurasian and African fossil and extant Equus. We refigure (Figure 13) herein Azzaroli (2003) last version of his phylogeny of Equus for ready reference.
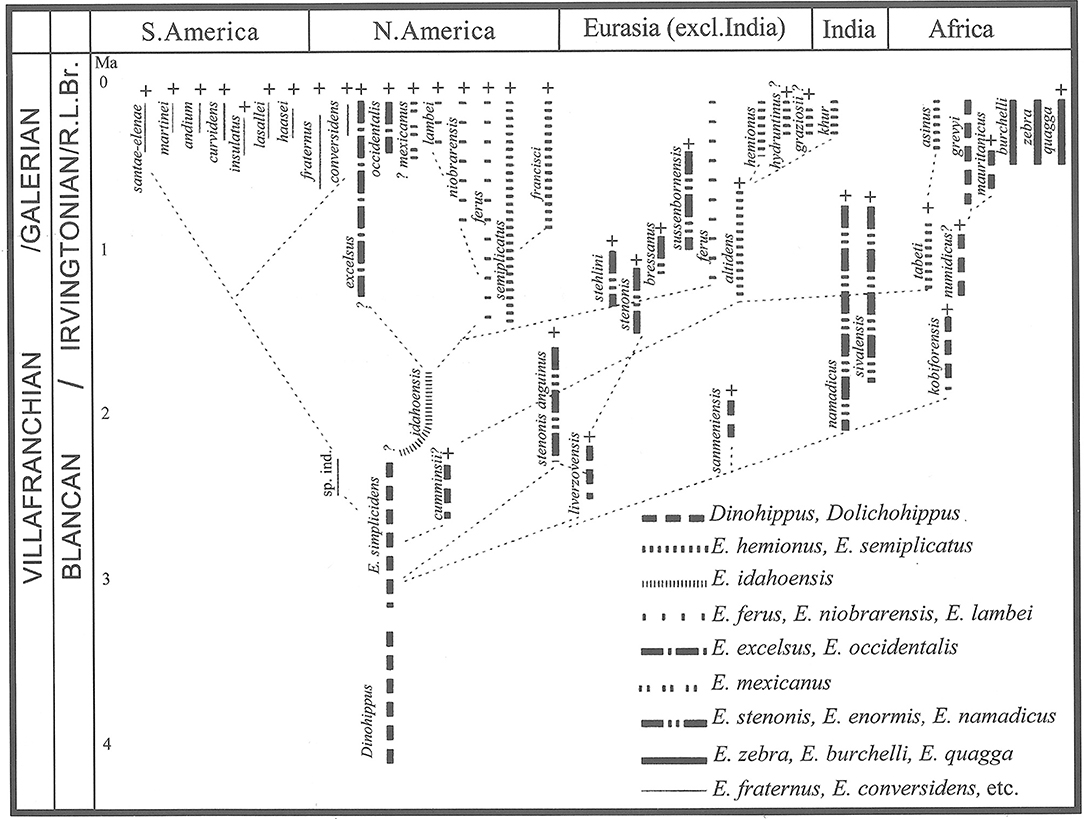
Figure 13. Phylogeny of the genus Equus with species geographical and stratigraphical distribution from Azzaroli (2003).
The datasets generated for this study are available on request to the corresponding author.
RB and OC conceived the paper. RB, OC, AJ, RP, and MB provided data. RB, OC, and AJ carried out analyses. RB wrote the manuscript with support from OC, AJ, RP, and LR. All authors equally contributed to discussion and conclusions.
RB's research has been supported by a Visiting Professor fellowship at the Earth Sciences Department of the University of Florence (May-June 2017 and 2018), NSF grants EAR0125009, 1113175, 1138908, and 1558586 for the study of fossil horses and NSF:DBI:ABI Innovation 1759882 FuTRES. Background work crucial for the results here presented has been supported by grants to LR by the University of Florence (Fondi di Ateneo) and by the Italian Ministry for Foreign Affairs and International Cooperation (MAECI DGSP-VI).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor is currently organizing a Research Topic with one of the authors, RB, and confirms the absence of any other collaboration. The reviewer, RS, declared a past co-authorship with one of the authors, LR, to the handling Editor.
Alberdi, M. T., Ortiz Jaureguizar, E., and Prado, J. L. (1998). A quantitative review of European stenonoid horses. J. Paleontol. 72, 371–387. doi: 10.1017/S0022336000036350
Alberdi, M. T., and Palombo, M. R. (2013). The late Early to early Middle Pleistocene stenonoid horses from Italy. Q. Int. 288, 25–44. doi: 10.1016/j.quaint.2011.12.005
Azzaroli, A. (1964). The two Villafranchian Horses of the Upper Valdarno. Palaeontogr. Ital. 59, 1–12.
Azzaroli, A. (1966). Pleistocene and living horses of the Old World, an essay of a classification based on skull characters. Palaeontogr. Ital. 61, 1–15.
Azzaroli, A. (1982). On Villafranchian Palaearctic Equids and their allies. Palaeontogr. Ital. 72, 74–97.
Azzaroli, A. (1983). Quaternary mammals and the “end-Villafranchian” dispersal event – a turning point in the history of Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 44, 117–139. doi: 10.1016/0031-0182(83)90008-1
Azzaroli, A. (1990). “The genus Equus in Europe,” in European Neogene Mammal Chronology (NATO ASI Series, Series A: Life Sciences, vol. 180), eds E. H. Lindsay, V. Fahlbusch, and P. (New York, NY: Mein Plenum Press), 339–355. doi: 10.1007/978-1-4899-2513-8_20
Azzaroli, A., De Giuli, C., Ficcarelli, G., and Torre, D. (1986). Mammal succession of the Plio-Pleistocene of Italy. Memorie Soc. Geol. Ital. 31, 213–218.
Azzaroli, A., and Voorhies, M. R. (1993). The Genus Equus in North America. The Blancan species. Palaeontogr. Ital. 80, 175–198.
Badam, G. L., and Tewari, B. S. (1974). On the zebrine affinities of the Pleistocene horse Equus sivalensis Falconer and Cautley. Bull. Deccan Coll. Res. Institute 34, 7–11.
Bajgusheva, V. S. (1971). Iskopaemaja Teriofauna Livenzovskogo Karera (Severo-Vostochnoe Priazovije) SSSR. Trudy Zool. Institute 49, 5–23.
Bajgusheva, V. S. (1978). Krupnaja Loshad Khaprovskogo Kompleksa is alluvija SeveroVostochnoto Priazovija. Invesija Servo-Kavkazkogo Nauchnogo Zentra Vychei Shkoly 1, 98–102.
Baker, W. E. (1850). Memoir by Major W. E. Baker, Bengal Engineers, on the Fossil Remains Presented by Himself and Colonel Colvin, C.B., to the Museum of Natural History at Ludlow. Ludlow: E. J. Partridge.
Bell, C. J., Lundelius, E. L., Barnosky, A. D., Graham, R. W., Lindsay, E. H., et al. (2004). “The Blancan, Irvingtonian, and Rancholabrean Mammal Ages,” in Late Cretaceous and Cenozoic Mammals of North America, ed M. O. Woodburne (New York, NY: Columbia University Press), 232–314. doi: 10.7312/wood13040-009
Bellucci, L., Bona, F., Corrado, P., Magri, D., Mazzini, I., Parenti, F, et al. (2014): Evidence of late Gelasian dispersal of African fauna at Coste San Giacomo (Anagni Basin, central Italy): Early Pleistocene environments the background of early human occupation in Europe. Q. Sci. Rev. 96, 72–85. doi: 10.1016/j.quascirev.2013.10.011.
Bellucci, L., Mazzini, I., Scardia, G., Bruni, L., Parenti, F., Segre, A. G., et al. (2012). The site of Coste San Giacomo (Early Pleistocene, central Italy): palaeoenvironmental analysis and biochronological overview. Q. Int. 267, 30–39. doi: 10.1016/j.quaint.2011.04.006
Bernor, R. L., Armour-Chelu, M., Gilbert, H., Kaiser, T. M., and Schulz, E. (2010). “Equidae,” in Cenozoic mammals of Africa, eds L. Werdelin and W. L. Sanders (Berkeley, CA: University of California Press), 685–721. doi: 10.1525/california/9780520257214.003.0035
Bernor, R. L., Armour-Chelu, M., Kaiser, T. M., and Scott, R. S. (2003a). An evaluation of the late MN 9 (Late Miocene, Vallesian Age), hipparion assemblage from Rudabánya (Hungary): systematic background, functional anatomy and paleoecology. Coloquios Paleontol. 1, 1132–1660.
Bernor, R. L., Cirilli, O., Wang, S. Q., and Rook, L. (2018). Equus cf. livenzovenzis from Montopoli, Italy (early Pleistocene; MN16b; ca. 2.6 Ma). Boll. Soc. Paleontol. Ital. 57, 203–216.
Bernor, R. L., Goehlic, U., Harzhauser, M., and Semprebon, G. (2017). Stratigraphy, chronology, systematics and paleoecology of the Vienna Basin Pannonian C hipparions (basal MN9, late Miocene). Palaeoclimatol. Palaeogeogr. Palaeoecol. 476, 28–41. doi: 10.1016/j.palaeo.2017.03.026
Bernor, R. L., and Harris, J. M. (2003). “Systematics and evolutionary biology of the Late Miocene and Early Pliocene hipparionine horses from Lothagam, Kenya,” in Lothagam – The Dawn of Humanity in Eastern Africa, eds J. M. and M. G. Leakey (New York, NY: Columbia University Press), 387–438. doi: 10.7312/leak11870-018
Bernor, R. L., Scott, R. S., Fortelius, M., Kappelman, J., and Sen, S. (2003b). “Systematics and evolution of the Late Miocene Hipparions from Sinap, Turkey,” in The Geology and Paleontology of the Miocene Sinap Formation, Turkey, eds M. Fortelius, J. Kappelman, S. Sen and R. L. Bernor (New York, NY: Columbia University Press), 220–281.
Bernor, R. L., Tobien, H., Hayek, L. A. C., and Mittmann, H. W. (1997). Hippotherium primigenium (Equidae, Mammalia) from the late Miocene of Höwenegg (Hegau, Germany). Andrias 10, 1–230.
Bertini, A. (2010). Pliocene to Pleistocene palynoflora and vegetation in Italy: state of the art. Q. Int. 225, 5–24. doi: 10.1016/j.quaint.2010.04.025
Blain, H. A., Agustí, J., Rook, L., Lordkipanidze, D., and Delfino, M. (2014). Paleoclimatic and paleoenvironmental context of the Early Pleistocene hominins from Dmanisi (Georgia, Lesser Caucasus) inferred from the herpetofaunal assemblage. Q. Sci. Rev. 105, 136–150. doi: 10.1016/j.quascirev.2014.10.004
Blumenthal, S. A., Levin, N. E., Brown, F. H., Brugal, J. P., Chritz, K. L., Harris, J. M., et al. (2017). Aridity and hominin environments. Proc. Natl. Acad. Sci. U.S.A. 114, 7331–7336. doi: 10.1073/pnas.1700597114
Bobe, R., and Eck, G. G. (2001). Responses of African bovids to pliocene climatic change. Paleobiology 27, 1–47. doi: 10.1666/0094-8373(2001)0270001:ROABTP2.0.CO;2
Bobe, R., and Leakey, M. G. (2009). “Ecology of Plio-Pleistocene mammals in the Omo-Turkana Basin and the emergence of Homo,” in The First Humans: Origin and Early Evolution of the Genus Homo, eds F. E. Grine, J. G. Fleagle, and R. E. Leakey (New York, NY: Springer), 173–184. doi: 10.1007/978-1-4020-9980-9_15
Cerling, T. E., Andanje, S. A., Blumenthal, S. A., Brown, F. H., Chritz, K. L., Harris, J. M., et al. (2015). Dietary changes of large herbivores in the Turkana Basin, Kenya from 4 to 1 Ma. Proc. Natl. Acad. Sci. U.S.A. 112, 11467–11472. doi: 10.1073/pnas.1513075112
Cerling, T. E., and Hay, R. L. (1986). An isotopic study of paleosol carbonates from Olduvai Gorge. Q. Res. 25, 63–78. doi: 10.1016/0033-5894(86)90044-X
Churcher, C. S., and Hooijer, D. A. (1980). The Oldovay zebra (Equus oldowayensis) from the later Omo beds, Ethiopia. Zool. Med. 55, 265–280.
Churcher, C. S., and Richardson, M. L. (1978). “Equidae” in Evolution on African Mammals, eds V. J. Maglio and H. S. B. Cooke (Cambridge, MA: Harvard University Press), 379–422.
Colbert, E. H. (1935a). Distributional and Phylogenetic Studies on Indian Fossil Mammals II: the relation of the Siwaliks of India as inferred by the migrations of Hipparion and Equus. Am. Museum Novitates 797, 1–15.
Colbert, E. H. (1935b). Siwalik mammals in the American Museum of Natural History. Trans. Am. Philos. Soc. 26:i-401. doi: 10.2307/1005467
Cooke, H. B. S. (1963). “Pleistocene mammal faunals of Africa, with particular references to South Africa,” in African Ecology and Human Evolution, eds F. C. Howell and F. Bourlière (New York, NY: Viking Fund Publications in Anthropology), 65–116. doi: 10.4324/9781315082943-5
Coppens, Y. (1971). Les vèrtébres Villafranchiens de Tunisie: Gisements nouveaux, signification. Comptes Rendus l'Acadèmie de Sciences. Paris, Série D, Science Naturelles 273, 51–54.
De Giuli, C., and Masini, F. (1986). Late Villafranchian faunas in Italy: the Casa Frata Local Fauna (Upper Valdarno, Tuscany). Palaeontogr. Ital. 74, 1–9.
Delson, E., Faure, M., Guérin, C., Aprile, L., Blackwell, B. A. B., Debard, E., et al. (2006). Franco-American renewed research at the Late Villafranchian locality of Senèze (Haute-Loire, France). Courier Forschungsinstitut Senckenberg 256, 275–290.
Dennell, R., Coard, R., and Turner, A. (2006). The biostratigraphy and magnetic polarity zonation of the Pabbi Hills, northern Pakistan: an Upper Siwalik (Pinjor Stage) Upper Pliocene–Lower Pleistocene fluvial sequence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 234, 168–185. doi: 10.1016/j.palaeo.2005.10.008
Dennell, R. W. (2004). Early Hominin Landscapes in Northern Pakistan: Investigations in the Pabbi Hills. Oxford: John and Erica Hedges Ltd and Archaeopress.
Eisenmann, V. (1983). “Family Equidae,” in Koobi Fora Research Project; Volume 2: The Fossil Ungulates: Proboscidea, Perissodactyla, and Suidae, ed J. M. Harris (Oxford, United Kingdom: Clarendon Press), 156–214.
Eisenmann, V. (1995). “What metapodial morphometry has to say about some Miocene hipparions,” in Paleoclimate and Evolution: With Emphasis on Human Origins, eds E. S. Vrba, G. H. Denton, T. C. Partridge, and L. H. Burckle (New Haven, CT: Yale University Press), 148–163.
Eisenmann, V (2004): Les Equidés (Mammalia, Perissodactyla) de Saint-Vallier (Drôme, France) et les Equidés Plio-Pleistocènes d'Europe. Geobios 37, 279–305. doi: 10.1016/S0016-6995(04)80019-6.
Eisenmann, V., Alberdi, M. T., De Giuli, C., and Staesche, U. (1988). “Studying Fossil Horses. Volume I – Methodology,” in Collected Papers After the New York International Hipparion Conference 1981, eds M. O. Woodburne and P. Y. Sondaar (Leiden: Brill), 1–71.
Eronen, J. T., and Rook, L. (2004). The Mio-Pliocene European primate fossil record: dynamics and habitat tracking. J. Hum. Evol. 47, 323–341. doi: 10.1016/j.jhevol.2004.08.003
Faith, J. T., Rowan, J., Du, A., and Koch, P. L. (2018). Plio-Pleistocene decline of African megaherbivores: no evidence for ancient hominin impacts. Science 362, 938–941. doi: 10.1126/science.aau2728
Falconer, H., and Cautley, P. T. (1849). Fauna Antiqua Sivalensis. Being the Fossil Zoology of the Sewalik Hills in the North of India. Illustrations-Part IX Equidae, Camelidae, Sivatherium. London: Smith, Elder and Co.
Falconer, H., and Walker, H. (1859). Descriptive Catalogue of the Fossil Remains of Vertebrata From the Siwalik Hills, the Nerbudda, Perim Island, etc in the Museum of the Asiatic Society of Bengal. Calcutta: C. B. Lewis, Baptist Mission Press. doi: 10.5962/bhl.title.112301
Fejfar, O., Heinrich, W. D., and Lindsay, E. H. (1998). “Updating the Neogene rodent biochronology in Europe,” in The Dawn of the Quaternary, eds T. van Kolfschoten and P.L. Gibbard (Mededelingen, Nederlands:Instituut voor Toegepaste Geowetenschappen TNO 60), 533–554.
Ferraro, J. V., Plummer, T. W., Pobiner, B. L., Oliver, J. S., and Bishop, L. C. (2013). Earliest archaeological evidence of persistent hominin carnivory. PLoS ONE 8:e62174. doi: 10.1371/journal.pone.0062174
Ferring, R., Oms, O., Agust,í, J., Berna, F., Nioradze, M., Shelia, T., et al. (2011). Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma. Proc. Natl. Acad. Sci. U.S.A. 108, 10432–10436. doi: 10.1073/pnas.1106638108
Forsten, A (1999): A review of Equus stenonis Cocchi (Perissodactyla, Equidae) and related forms. Q. Sci. Rev. 18, 1373–1408. doi: 10.1016/S0277-3791(98)00073-0.
Forsten, A., and Sharapov, S. (2000). Fossil equids (Mammalia, Equidae) from the Neogene and Pleistocene of Tadzhikistan. Geodiversitas 22, 293–314.
Forsyth Major, C. J. (1885). On the Mammalian Fauna of the Val d'Arno. Q. J. Geol. Soc. London 41, 1–8. doi: 10.1144/GSL.JGS.1885.041.01-04.2
Fortelius, M., Eronen, J., Liu, L., Pushkina, D., Tesakov, A., Vislobokova, I., et al. (2006). Late Miocene and Pliocene large land mammals and climatic changes in Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 238, 219–227. doi: 10.1016/j.palaeo.2006.03.042
Furió, M., Agustí, J., Mouskhelishvili, A., Sanisidro, Ó., and Santos-Cubedo, A. (2010). The paleobiology of the extinct venomous shrew Beremendia (Soricidae, Insectivora, Mammalia) in relation to the geology and paleoenvironment of Dmanisi (Early Pleistocene, Georgia). J. Vertebrate Paleontol. 30, 928–942. doi: 10.1080/02724631003762930
Gabunia, L., Vekua, A., Lordkipanidze, D., Swisher, C. C., Ferring, R., Justus, A., et al. (2000). Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: taxonomy, geological setting, and age. Science 288, 1019–1025. doi: 10.1126/science.288.5468.1019
Gaur, R., and Chopra, S. R. K. (1984a). On a new subspecies of Equus from Pinjor Formation of Upper Sivaliks - with remarks on Sivalik Equus. J. Palaeontol. Soc. India 29, 19–25.
Gaur, R., and Chopra, S. R. K. (1984b). Taphonomy, fauna, environment and ecology of Upper Sivaliks (Plio-Pleistocene) near Chandigarh, India. Nature 308, 353–355. doi: 10.1038/308353a0
Gazin, C. L. (1936). A study of the fossil horse remains from the Upper Pliocene of Idaho. Proc. U.S.Natl. Museum 83, 281–320. doi: 10.5479/si.00963801.83-2985.281
Ghinassi, M., Abbazzi, L., Esu, D., Gaudant, J., and Girotti, O. (2005). Facies analysis, stratigraphy and paleontology (molluscs and vertebrates) in the upper Pliocene sandy flood-basin deposits of the Upper Valdarno basin (Northern Apennines). Riv. Ital. Paleontol. Stratigr. 111, 463–483. doi: 10.13130/2039-4942/6332
Gidley, J. W. (1930). Continuation of the fossil horse round-up on the Old Oregon Trail. Smithsonian Explor. Fieldwork 3111, 33–40.
Groves, C. P. (2002). Taxonomy of living Equidae, Report 8:94-107. Gland, Switzerland, and Cambridge, IUCN/SCC Equid Specialist Group, IUCN.
Hernández Fernández, M., and Vrba, E. S. (2006). Plio-Pleistocene climatic change in the Turkana Basin (East Africa): Evidence from large mammal faunas. J. Hum. Evol. 50, 595–626. doi: 10.1016/j.jhevol.2005.11.004
Hooijer, D. A. (1951). Observations on a calvarium of Equus sivalensis Falconer et Cautley from the Siwaliks of the Punjab, with craniometrical notes on recent Equidae. Arch. Néerlandaises Zool. 8, 243–266. doi: 10.1163/187530151X00036
Hooijer, D. A. (1976). The late pliocene equidae of langebaanweg, Cape Province, South Africa. Zool. Verhandelingen 148, 3–39.
Hopwood, A. T. (1936). The Former Distribution of Caballine and Zebrine Horses in Europe and Asia. Proc. Zool. Soc. London 106, 897–912. doi: 10.1111/j.1469-7998.1936.tb06291.x
Hopwood, A. T. (1937). Die fossilen Pferde von Oldoway. Wissenschaft. Ergebnisse Oldoway Expedition 4, 112–136.
Hussain, S. T., van den Bergh, G., Steensma, K. J., de Visser, J. A., de Vos, J., Mohammad, A., et al. (1992). Biostratigraphy of the Plio-Pleistocene continental sediments (Upper Siwaliks) of the Mangla-Samwal anticline, Azad Kashmir, Pakistan. Proc. Koninklijke Nederlandse Akademie Wetenschappen 95, 65–80.
Jukar, A. M. (2018). Temporal Turnover in Late Neogene and Quaternary Mammal Assemblages. [dissertation]. Fairfax, VA: George Mason University.
Kaiser, T. M., and Franz-Odendaal, T. A. (2004). A mixed-feeding Equus species from the Middle Pleistocene of South Africa. Q. Res. 62, 316–323. doi: 10.1016/j.yqres.2004.09.002
Lordkipanidze, D., Jashashvili, T., Vekua, A., Ponce de León, M. S., Zollikofer, C. P., Rightmire, G. P., et al. (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 449, 305–310. doi: 10.1038/nature06134
Lydekker, R. (1882). Siwalik and Narbada Equidae. Memoirs Geol. Soc. India Palaeontol. Indica Ser. X 2, 67–98.
Lydekker, R. (1884). Catalogue of Vertebrate Fossils From the Siwaliks of India in the Science and Art Museum, Dublin. Dublin: The Royal Dublin Society.
Lydekker, R. (1885). Catalogue of the Remains of Siwalik Vertebrata Contained in the Geological Department of the Indian Museum, Calcutta. Calcutta: Government Printing, India.
Lydekker, R. (1886). Catalogue of the fossil Mammalia in the British Museum (Natural History) Part III. Containing the order Ungulata, suborders Perissodactyla, Toxodontia, Condylartha, and Ambylopoda. London: Taylor and Francis.
Lydekker, R. (1891). On a collection of mammalian bones from Mongolia. Records Geol. Survey India 24, 207–211.
Matthew, W. D. (1929). Critical observations upon Siwalik mammals (exclusive of Proboscidea). Bull. Am. Museum Nat. History 56, 437–560.
Mazza, P., Bertini, A., and Magi, M. (2004). - The Late Pliocene site of Poggio Rosso (Central Italy): taphonomy and paleoenvironment. Palaios 19, 227–248. doi: 10.1669/0883-1351(2004)019<0227:TLPSOP>2.0.CO;2
McDonald, H. (1996). “Population structure of the late Pliocene (Blancan) zebra Equus simplicidens (Perissodactyla: Equidae) from the Hagerman Horse Quarry, Idaho,” in Paleoecology and Paleoenvironments of Late Cenozoic Mammals: Tributes to the Career of CS (Rufus) Churcher, eds K. M. Stewart and K. L. Seymour. (Toronto: University of Toronto Press), 134–155.
McDonald, H. G., Link, P. K., and Lee, D. E. (1996). An overview of the geology and paleontology of the Pliocene Glenns Ferry Formation, Hagerman Fossil Beds National Monument. Northwest Geol. 26, 16–45.
Miller, K. G., Kominz, M. A., Browning, J. V., Wright, J. D., Mountain, G. S., Katz, et al. (2005). The phanerozoic record of global sea-level change. Science 310, 1293–1298. doi: 10.1126/science.1116412
Mishra, V. P., Singh, J., and Chandel, R. S. (2013). Catalogue of Fossils in the Museum at Siwalik Fossil Park, Saketi, Himachal Pradesh. Kolkata: Geological Survey of India.
Nanda, A. C. (2015). Siwalik mammalian faunas of the Himalayan foothills with reference to biochronology, linkages and migration. WIGH Monogr. Ser. 2, 1–341.
Napoleone, G., Albianelli, A., Azzaroli, A., and Mazzini, M. (2001). The Poggio Rosso locality calibrated to the end-Pliocene and its significance for dating the late Villafranchian mammal faunas of the Upper Valdarno, Central Italy. Riv. Ital. Paleontol. Stratigr. 107, 287–296. doi: 10.13130/2039-4942/5436
Palombo, M. R., and Alberdi, M. T (2017): Light shadows in the evolution of South European stenonoid horses. Fossil Imprint 73, 115–140. doi: 10.2478/if-2017-0006
Palombo, M. R., Alberdi, M. T., Bellucci, L., and Sardella, R (2017): An intriguing middle-sized horse from Coste San Giacomo (Anagni Basin, central Italy). Q. Res. 87, 347–362. doi: 10.1017/qua.2017.6
Patnaik, R. (2013). “Indian Neogene Siwalik mammalian biostratigraphy: an overview,” in Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology, eds X. Wang, L. J. Flynn and M. Fortelius. (New York, NY: Columbia University Press), 423–444. doi: 10.7312/columbia/9780231150125.003.0017
Patnaik, R. (2015). Diet and habitat changes among Siwalik herbivorous mammals in response to Neogene and Quaternary climate changes: an appraisal in the light of new data. Q. Int. 371, 232–243. doi: 10.1016/j.quaint.2014.11.025
Pilgrim, G. E. (1910). Preliminary note on a revised classification of the Tertiary freshwater deposits of India. Rec. Geol. Survey India 40, 185–205.
Pilgrim, G. E. (1913). The correlation of the Siwaliks with mammal horizons of Europe. Rec. Geol. Survey India 43, 264–326.
Plummer, T. W., Ditchfield, P. W., Bishop, L. C., Kingston, J. D., and Ferraro, J. V. (2009). Oldest evidence of toolmaking hominins in a grassland-dominated ecosystem. PLoS ONE 4:e7199. doi: 10.1371/journal.pone.0007199
Potts, R. (1998). Variability selection in hominid evolution. Evol. Anthropol. 7, 81–96. doi: 10.1002/(SICI)1520-6505(1998)7:3<81::AID-EVAN3>3.0.CO;2-A
Potts, R. (2007). “Environmental context of Pliocene human evolution in Africa,” in Hominin Environments in the East African Pliocene: An Assessment of the Faunal Evidence, eds R. Bobe, Z. Alemseged, and A.K. Behrensmeyer, (New York, NY: Springer), 25–48.
Potts, R., Behrensmeyer, A. K., Faith, J. T., Tryon, C. A., Brooks, A. S., Yellen, J. E., et al. (2018). Environmental dynamics during the onset of the Middle Stone Age in eastern Africa. Science 360, 86–90. doi: 10.1126/science.aao2200
Pradella, C., and Rook, L. (2007). Mesopithecus (Primates, Cercopithecoidea) from Villafranca d'Asti (Early Villafranchian; NW Italy) and palaeoecological context of its extinction. Swiss J. Geosci. 100, 145–152. doi: 10.1007/s00015-007-1208-2
Quade, J., and Cerling, T. E. (1995). Expansion of C4 grasses in the Late Miocene of Northern Pakistan: evidence from stable isotopes in paleosols. Palaeogeogr. Palaeoclimatol. Palaeoecol. 115, 91–116. doi: 10.1016/0031-0182(94)00108-K
Quade, J., Cerling, T. E., Bowman, J., and Jah, A. (1993). “Paleoecologic reconstruction of floodplain environments using palaeosols from Upper Siwalik Group sediments, northern Pakistan,” in Himalaya to the Sea: Geology, Geomorphology and the Quaternary, ed J. F. Shroder Jr. (London: Routledge), 213–226. doi: 10.4324/9780203414637_chapter_10
Ranga Rao, A., Nanda, A. C., Sharma, U. N., and Bhalla, M. S. (1995). Magnetic polarity stratigraphy of the Pinjor Formation (Upper Siwalik) near Pinjore, Haryana. Curr. Sci. 68, 1231–1236.
Repenning, C. A., Weasma, T. R., and Scott, G. R. (1995). The early Pleistocene (latest Blancan-earliest Irvingtonian) Froman Ferry fauna and history of the Glenns Ferry Formation, southwestern Idaho. U. S. Geol. Survey Bull. 2105, 1–86.
Richmond, D. R., and McDonald, H. G. (1998). “The Hagerman Horse Quarry: death and deposition,” in National Park Service Paleontological Research 3. Technical Report NPS/NRGRD/GRDTR-98/1, eds V.L. Santucci and L. McClelland. (Lakewood, CO: National Park Service), 103–104.
Richmond, D. R., McDonald, H. G., and Bertog, J. (2002). Stratigraphy, Sedimentology and Taphonomy of the Hagerman Horse Quarry, Hagerman, Idaho. Hagerman: Hagerman Fossil Beds National Monument.
Rivals, F., and Athanassiou, A. (2008). Dietary adaptations in an ungulate community from the late Pliocene of Greece. Palaeogeogr. Palaeoclimatol. Palaeoecol. 265, 134–139. doi: 10.1016/j.palaeo.2008.05.001
Rook, L., and Martínez-Navarro, B. (2010). Villafranchian: the long story of a Plio-Pleistocene European large mammal biochronologic unit. Q. Int. 219, 134–144. doi: 10.1016/j.quaint.2010.01.007
Ruez Jr D. R. (2006). Middle Pliocene paleoclimate in the Glenns Ferry Formation of Hagerman Fossil Beds National Monument, Idaho: a baseline for evaluating faunal change. J. Idaho Acad. Sci. 42, 1–17.
Ruez Jr D. R. (2009a). Framework for stratigraphic analysis of Pliocene fossiliferous deposits at Hagerman Fossil Beds National Monument, Idaho. Rocky Mountain Geol. 44, 33–70. doi: 10.2113/gsrocky.44.1.33
Ruez Jr D. R. (2009b). Revision of the Blancan (Pliocene) mammals from Hagerman Fossil Beds National Monument, Idaho. J. Idaho Acad. Sci. 45, 1–144.
Sahni, M. R., and Khan, E. (1961). Recent finds of shivalik vertebrates. 5. On a skull and ramii of Equus sivalensis with observations on the remains of Equus so far collected from the shivaliks. Res. Bull. Panjab Univ. Sci. 12, 265–268.
Sahni, M. R., and Khan, E. (1988). Pleistocene Vertebrate Fossils and Prehistory of India. New Delhi: Books and Books.
Ségalen, L., Lee-Thorp, J. A., and Cerling, T. (2007). Timing of C4 grass expansion across sub-Saharan Africa. J. Hum. Evol. 53, 549–559. doi: 10.1016/j.jhevol.2006.12.010
Sikes, N. E., Potts, R., and Behrensmeyer, A. K. (1999). Early Pleistocene habitat in Member 1 Olorgesailie based on paleosol stable isotopes. J. Hum. Evol. 37, 721–746. doi: 10.1006/jhev.1999.0343
Skinner, M. P., and Hibbard, C. W. (1972). Early Pleistocene pre-glacial and glacial rocks and faunas of north-central Nebraska. Bull. Am. Museum Natural History 148, 1–148.
Stehlin, H. G., and Graziosi, P (1935): Ricerche sugli Asinidi fossili d'Europa (Studies on the fossil European asses). Memoires Soc. Paléontol. Suisse 56, 1–73.
Strani, F., DeMiguel, D., Sardella, R., and Bellucci, L. (2015). Paleoenvironments and climatic changes in the Italian Peninsula during the Early Pleistocene: evidence from dental wear patterns of the ungulate community of Coste San Giacomo. Q. Sci. Rev. 121, 28–35. doi: 10.1016/j.quascirev.2015.05.008
Strani, F., DeMiguel, D., Sardella, R., and Bellucci, L. (2018). Resource and niche differentiation mechanisms by sympatric Early Pleistocene ungulates: the case study of Coste San Giacomo. Q. Int. 481, 157–163. doi: 10.1016/j.quaint.2017.08.064
Sun, B., Zhang, X., Liu, Y., and Bernor, R. L. (2018). Sivalhippus ptychodus and Sivalhippus platyodus (Perissodactyla, Mammalia) from the late Miocene of China. Riv. Ital. Paleontol. Stratigr. 124, 1–22. doi: 10.13130/2039-4942/9523
Uno, K. T., Polissar, P. J., Jackson, K. E., and deMenocal, P. B. (2016). Neogene biomarker record of vegetation change in eastern Africa. Proc. Natl. Acad. Sci. U.S.A. 113, 6355–6363. doi: 10.1073/pnas.1521267113
Valli, A. M. F., and Palombo, M. R. (2008). Feeding behaviour of middle-size deer from the Upper Pliocene site of Saint-Vallier (France) inferred by morphological and micro/mesowear analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 257, 106–122. doi: 10.1016/j.palaeo.2007.09.006
Keywords: Equus, evolution, African zebras, North America, old world, Plio–Pleistocene, Villafranchian
Citation: Bernor RL, Cirilli O, Jukar AM, Potts R, Buskianidze M and Rook L (2019) Evolution of Early Equus in Italy, Georgia, the Indian Subcontinent, East Africa, and the Origins of African Zebras. Front. Ecol. Evol. 7:166. doi: 10.3389/fevo.2019.00166
Received: 11 February 2019; Accepted: 25 April 2019;
Published: 29 May 2019.
Edited by:
Leonardo Dos Santos Avilla, Universidade Federal do Estado do Rio de Janeiro, BrazilReviewed by:
Raffaele Sardella, Sapienza University of Rome, ItalyCopyright © 2019 Bernor, Cirilli, Jukar, Potts, Buskianidze and Rook. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raymond L. Bernor, cmxiZXJub3JAZ21haWwuY29t; cmJlcm5vckBob3dhcmQuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.