 Fabrice G. W. A. Ottburg
Fabrice G. W. A. Ottburg Edgar A. van der Grift
Edgar A. van der Grift
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 12 February 2019
Sec. Urban Ecology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00023
This article is part of the Research TopicIntegrating Transport Infrastructures with Living LandscapesView all 16 articles
Roads and traffic may have major impacts on amphibian populations, primarily as a result of amphibian road mortality. A variety of measures have been developed to prevent road mortality of amphibians, such as the construction of fences to keep the animals off the road and amphibian tunnels to provide them a safe passage. We carried out a capture-mark-recapture study to evaluate the performance of two tunnels and permanent drift fences for common toads at a local road in the Netherlands. We found that of the marked toads only 31% used the tunnels to cross the road. We assessed four possible explanations for the fact that a proportion of the toads did not use the tunnels: for toad groups that used the tunnels, as compared to toad groups that did not use the tunnels, (1) the mean distance between the location of first capture and the nearest tunnel was significantly smaller; (2) the mean movement distance along the fence was significantly larger; (3) the number of toad groups that walked in the wrong direction after encountering the drift fence was lower; (4) the mean number of nights between first and last capture of the toad group was significantly higher. Over all study years 28% of the migrating toads—marked and unmarked—that attempted to cross the road ended up on the road pavement, despite the mitigation. Migrating population numbers decreased with about 75% after the mitigation measures were installed. We emphasize that better baseline studies on where toads cross before mitigation and improved knowledge on effects of tunnel design and the distances the animals move along a drift fence are vital to mitigate road impacts properly and maintain viable toad populations. We recommend to base tunnel densities on the mean movement distance of the toads that move only small distances and spent relatively little time along the drift fence, install drift fences that go well beyond the location where toads cross the road, take appropriate measures at entrance roads and at fence ends and consider alternatives to tunnels and fences, such as the creation of breeding waters on both sides of the road.
Roads and traffic have major impacts on animal populations (Forman et al., 2003; van der Ree et al., 2015). The impacts of roads and traffic on amphibian populations is primarily the result of amphibian road mortality (Fahrig et al., 1995; Carr and Fahrig, 2001; Hels and Buchwald, 2001; Gibbs and Shriver, 2005; Andrews et al., 2008; Glista et al., 2008; Beebee, 2013). This is particularly obvious in species such as the common toad (Bufo bufo), that usually migrate in large numbers between their wintering and breeding habitat and need to cross roads during these movements (see van Gelder, 1973; Santos et al., 2007; Cooke, 2011). A variety of measures have been developed to prevent road mortality of amphibians, of which the construction of fences to keep the animals off the road and amphibian tunnels to provide them a safe passage are the most common ones (Iuell et al., 2003; Schmidt and Zumbach, 2008; Jackson et al., 2015). Although such measures are frequently applied across the world, only a few studies have evaluated their effectiveness in reducing road-kill and facilitating safe movements across roads (Woltz et al., 2008; see overview in Glista et al., 2009; Niemi et al., 2014).
High road-kill numbers among common toads have been observed on a local road near Ede city in the central parts of Netherlands. Road-kill occurred particularly during spring migration, as the animals had to cross the road to migrate from their wintering habitat (south of the road) to their breeding ponds (north of the road). Until 2010 volunteers put up temporary drift fences and pitfall traps to capture the migrating animals and transport them manually across the road. In 2010 the temporary measures were replaced by two tunnels and permanent drift fences. During the spring migrations of 2011 and 2012 use of the tunnels by toads was frequently observed. However, it was also noted that not all toads were successful in finding the tunnels. In addition, a proportion of the migrating toads were still killed by traffic despite the mitigation. Further, no inferences could be made on the effectiveness of the mitigation on preserving the toad population as population estimates after mitigation were lacking. Our objective is to evaluate the performance of these mitigation measures for common toads and assess (1) what proportion of the toads that approach the road make use of the tunnels, (2) the possible reasons that some of the toads do not use the tunnels, (3) what proportion of the toads that approach the road end up on the tarmac, despite the mitigation, and (4) whether size of the migrating toad population differs before and after the installation of the mitigation measures.
Our study site is a 1-km road stretch of a local road—the “Horalaan”—on the outskirts of the city of Ede, The Netherlands. The road provides access to a business site and private properties situated in a forested area. The road is paved, five meters wide and traffic volume is relatively low (< 5,000 vehicles/day). The road crosses mixed-forests on sandy soils, which are poor in nutrients and deposited during glacial periods. The area is about 17.5–50 m above sea level. These forests host one of the four largest known common toad populations in the country (Ottburg and van Blitterswijk, 2009). Land habitat of the toads is mainly found south of the road. The vegetation exists here of Oak-Beech and Oak-Birch forests and plantations of Pine (e.g., Pinus sylvestris) and Spruce (e.g., Picea abies). Dominant tree species are Common oak (Quercus robur), Sessile oak (Quercus petraea), Beech (Fagus sylvatica), Downy birch (Betula pubescens), and Silver birch (Betula pendula). The undergrowth mainly consists of Alder buckthorn (Rhamnus frangula), Elder (Sambucus nigra), Honeysuckle (Lonicera periclymenum), Calluna (Calluna vulgaris), Bracken (Pteridium aquilinum), Blueberry (Vaccinium myrtillus), and Blackberry (Rubus fruticosus). Breeding habitat is situated north of the road and consists of three ponds. Distance between road and ponds varies between 140 and 200 m. The ponds are up to 1.5 m deep and are, respectively, about 220, 700, and 720 square meters in size. The aquatic vegetation consists mainly of Western waterweed (Elodea nuttallii), Mare's tail (Hippurus vulgaris), Fringed waterlily (Nymphoides peltata) and the invasive New Zealand pigmyweed (Crassula helmsii). The pond banks are about 3–5 m wide. The vegetation on the banks consists mainly of Willow (Salix spec.), Blackberry (Rubus fruticosus), Common rush (Juncus effusus), Common cattail (Typha latifolia), Slender tufted-sedge (Carex acuta), Purple loosestrife (Lythrum salicaria), and Reed (Phragmites australis). The ponds were excavated in 2010 because, at that time, four existing breeding waters were lost due to urban development. The new ponds were situated about 100 meters south of the existing breeding waters.
In 2010 two amphibian tunnels and permanent drift fences were installed (Figure 1). The tunnels are two-way concrete tunnels (type ACO), 8.6 m long, 0.50 m wide, 0.30 m high, with an open roof (grid). The tunnels are situated about 100 m apart in the central part of the mitigated road stretch. The drift fences are smooth black barriers, 0.40 m high and made of high density poly-ethylene. The fence on the south side of the road is 900 m long. The fence on the north side of the road is 1,000 m long. The fence ends include a 25 m-long drift fence perpendicular to the road. Where access roads cross the fences, small cattle guards have been installed to prevent the toads from accessing the road corridor.

Figure 1. Amphibian tunnel and drift fence at the study site. Photo: F. Ottburg.
In 2013, 2014, and 2015 we carried out a capture-mark-recapture study. We installed 36 pitfall traps, 25 m apart, along the drift fence south of the road. Pitfall traps were also installed at both tunnel exits, north of the road (Figure 2). Immediately after the start of spring migration toad surveys were conducted (1) along the drift fence on the south side of the road, (2) on the road and (3) at the northern tunnel exits.
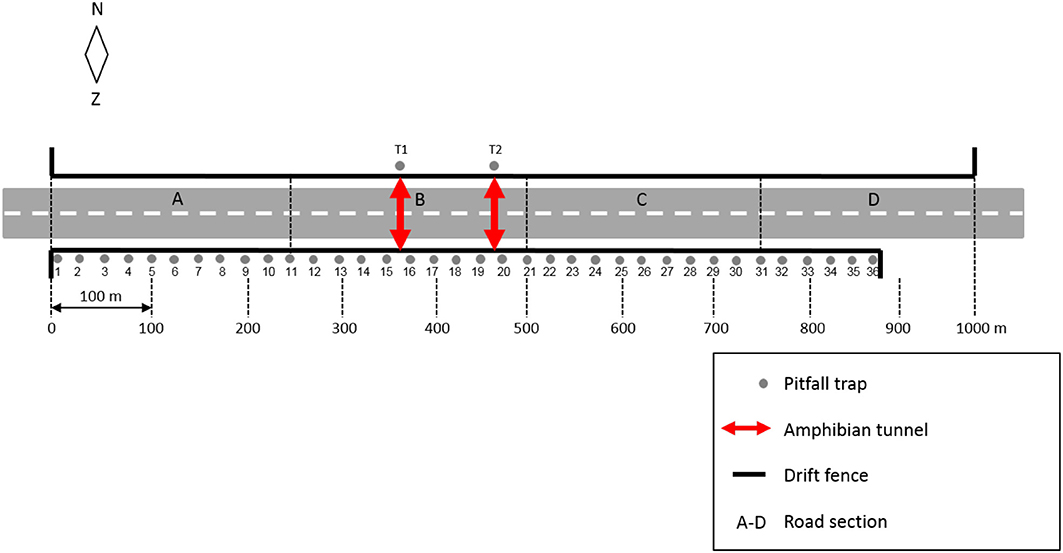
Figure 2. Schematic overview of study site and experimental set-up.
During the surveys along the drift fence all toads that were captured in the pitfall traps or that were found moving along the fence were individually marked. For marking we used a small sticker with a unique number (hereafter referred to as “ID number”) that was attached to the back of the toad with the help of a biodegradable glue (Figure 3). If toads were in amplexus, i.e., the mating embrace during which eggs will be shed into the water by the female and there fertilized by the male, we only attached a sticker to the back of the male toad. Based on re-sightings of marked toads in the breeding ponds, we estimate that the markings stayed on for at least 1 week. After marking we released the toads on the same spot where they had been found along the fence or at a distance of 1–2 m from the pitfall trap where they had been captured. We recorded for each captured toad, date, time of capture, sex, life stage (sub-adult, adult), and capture location. Capture location was registered as either pitfall trap number (1–36) or fence-stretch indication (1–2, 2–3, 3–4, etc.). During data entry these location descriptions were transformed into distances (in m), measured from the most western pitfall trap (number 1). If the capture location was in between two pitfall traps, the central spot of that particular fence stretch was used. This approach results in a spatial resolution of 1 m for the pitfall traps (exact locations) and 25 m for toads that were captured in between the pitfall traps. In 2013 we used all 36 pitfall traps. In 2014 and 2015 we used trap 1, 5, 9, 13, 17, 21, 25, 29, 33, and 36. The others were closed, aiming to reduce the number of trap encounters for a migrating toad during one night.

Figure 3. Toads with marking moving along the drift fence. Photo: F. Ottburg.
During the road surveys all toads found on the road, dead or alive, were counted. We recorded date, time of discovery, sex and life stage (if recognizable), capture location and, if present, their ID number. For capture location we distinguished four 250 m-road sections (A–D; see Figure 2). Dead toads were removed from the pavement to avoid double counts. Living toads were collected and released in one of the breeding ponds.
During the surveys of the pitfall traps at the tunnel exits, all toads were counted. We recorded date, time of capture, sex, life stage, capture location (tunnel 1 or 2) and, if present, their ID number. All toads were collected and released in one of the breeding ponds.
The surveys took place in the morning (6:00–11:00), incidentally in the afternoon (14:00–18:00), and in the evening (19:00–24:00), on days that weather conditions were assumed favorable for toad activity (air temperature >8°C and preferably some rain). During one survey the pitfalls, drift fence, and road was checked two times. The second check ended more or less when the toads became inactive, i.e., were no longer found moving along the drift fence. That “inactivity” could be judged adequately, is supported by the observation that, over all study years, only about 5% of all toads captured along the drift fence, have been captured during morning surveys. The survey period ended when no more toads were found along the drift fence for at least five consecutive days. Table 1 provides an overview of the survey characteristics per study year.
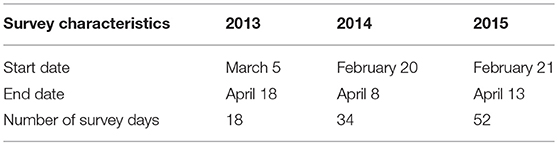
Table 1. Survey characteristics for each study year.
The proportion of toads that approach the road and make use of the tunnels was estimated by dividing the number of marked toads that did use the tunnels by the total number of marked toads. Herein “marked toads” is defined as the sum of (1) all individually marked toads (male or female), (2) all males that are part of a marked amplexus and (3) all females that are part of a marked amplexus. A crossing index (CI) was calculated through dividing the number of marked toads that did use the tunnels by the number of marked toads that did not use the tunnels. This implies that if CI = 1, the number of toads that did or did not use the tunnels is equal; if CI < 1, the number of toads that used the tunnels is lower than the number of toads that did not; if CI > 1, the number of toads that used the tunnels is higher than the number of toads that did not. A road permeability index (RPI) was calculated through dividing the number of marked and unmarked toads that did use the tunnels by the total number of toads that attempted to cross the road to reach the breeding ponds. This total number was estimated by adding up (1) the number of marked toads, (2) the number of unmarked toads on the road, dead or alive, and (3) the estimated number of missed toads along the drift fence, calculated through multiplying the number of unmarked toads in the pitfalls at the tunnel exits with 1/CI.
To explore the reasons why some of the toads do not use the tunnels we tested five hypotheses. If toads that do not use the tunnels are compared with toads that do use the tunnels, we expected the first to (1) arrive at the drift fence further away from a tunnel and, hence, these toads will have to cover more distance; (2) move less distance along the fence; (3) pass tunnels more often without using them; (4) walk more often in the wrong direction, i.e., away from the tunnels, after encountering the drift fence; (5) spend less time along the drift fence. Our null hypothesis was that there is no difference in these variables for toads that do and do not use the tunnels. In testing these hypotheses we used “toad group” as unit. A toad group is either an individual toad or two (or three) toads in amplexus. We chose this approach—instead of using “toad” as unit—because the movements of the toads that are part of an amplexus are not independent.
We compared the mean distance between the location of first capture and the nearest tunnel (test 1), the mean movement distance along the fence, derived from the location of first capture and the location of last re-capture (test 2), the number of toad groups that walked past a tunnel (test 3), the number of toad groups that walked in the wrong direction after encountering the drift fence (test 4), and the mean number of nights between first and last moment the toad group was captured (test 5) for toad groups that did and did not use the tunnels over all study years. Marked toad groups found on the road (n = 2) were excluded from the analysis as the spatial resolution of the road survey data was insufficient. Marked toad groups for which the location and time of first capture was not known (n = 10), were excluded from the analyses as for these animals no inferences on movement distances or time spent along the drift fence could be made. Marked toad groups that were not recaptured (n = 243) or did not move between first capture and consecutive re-captures (n = 14) were excluded from the analysis in test 4 as no inferences could be made on their direction of movement. In test 2, the calculated mean for toad groups that did not use the tunnels can be seen as a minimum value as toad groups that are captured only once result in a movement distance of 0 m, while it is likely that the animals did move along the fence for a while but left it before the next survey was carried out. In test 1, 2, and 5 we tested for significance between the means with the use of the non-parametric Mann-Whitney U-Test (p < 0.05 significance level) as normal distribution of the data could not be assumed and the number of possible values was limited. We used the Chi-square test of association (p < 0.05 significance level) to test for independence between the categorical variables “walking past one or more tunnels” and “eventual use of the tunnels” as well as “toad group movement direction after encountering the drift fence” and “eventual use of the tunnels.” We explored whether sex affected tunnel use probability for toads that moved along the drift fence individually by logistic regression. The same analysis was performed to explore whether toads in amplexus were more likely to use the tunnels than individual toads.
The proportion of the migrating toads that ended up on the road was estimated by dividing the number of toads that were found on the road—dead or alive—by the estimated population size (see below) per study year. These proportions were averaged to calculate an estimate over all study years.
For the years 2013–2015, the size of the toad population that had to cross the road to reach the breeding ponds, was estimated by adding up (1) the number of toads found along the drift fence that were not marked but immediately transported to the breeding ponds, (2) the number of marked toads, (3) the number of unmarked toads on the road, dead or alive, and (4) the estimated number of missed toads along the drift fence. The first group of toads, which were immediately transported to the breeding ponds, concerned toads that were discovered by volunteers prior to the start date of the surveys. The estimated number of missed toads along the drift fence was calculated through multiplying the number of unmarked toads in the pitfalls at the tunnel exits with 1/CI. Estimates of the size of the toad population that had to cross the road to reach the breeding ponds in the pre-mitigation situation were derived from a previous survey, carried out in 2007 and 2008 (Ottburg and van Blitterswijk, 2009). In this earlier survey all toads that were found along temporary drift fences—which were of the same length as the later as part of the mitigation installed permanent drift fences—and on the road were counted by volunteers after which the animals were captured and released in the breeding ponds. Similar to our study, these surveys lasted for the entire period of spring migration, which was, respectively 28 days (2007) and 59 days (2008).
Over all study years a total of 722 toads were marked (Table 2), either individually (43%) or as part of an amplexus (57%). 61% of the marked toads were males, 38% were females, and for 1% sex was not identified. All marked toads were adults. Of the marked toads 31% used the tunnels to cross the road, 68% were not detected using the tunnels to get across, and 1% ended up on the road. For all years together the CI is 0.46, indicating that the number of toads that did use the tunnels is less than half the number of toads that did not. The RPI is 0.26, indicating that only about one quarter of all toads that attempted to cross the road, managed to find and use the tunnels. Logistic regression showed that male toads were more likely to use the tunnels than female toads (B = −1.13, s.e. = 0.155, p < 0.001). Twenty four percent of all marked males, excluding the ones that were part of an amplexus, were detected to use the tunnels vs. 8% of all marked females. Amplexuses were more likely to use the tunnels than individually moving toads (B = −0.95, s.e. = 0.156, p < 0.001). Twenty eight percent of all marked amplexuses were detected to use the tunnels vs. 20% of all marked individual toads (male or female).
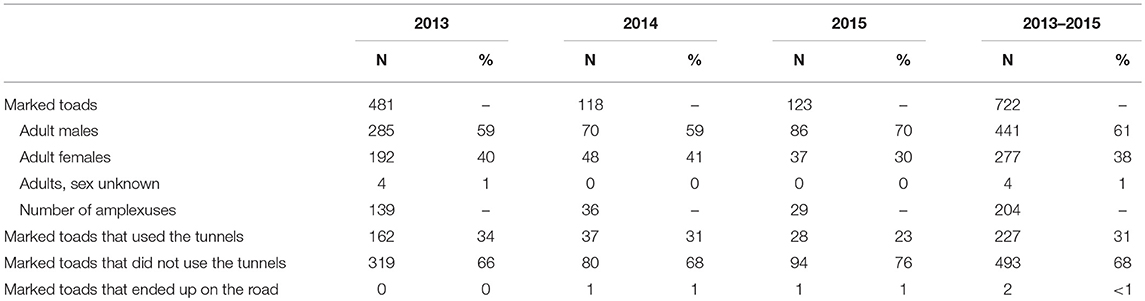
Table 2. Number and proportion of marked toads that did and did not use the amphibian tunnels per study year and over all study years.
Over all study years a total of 504 toad groups were marked. Of these, 118 used the tunnels and 386 did not. The toad groups were not evenly distributed over the drift fence; about 90% of all toad groups were first captured along the 400-m road stretch between kilometer-post 0.4 and 0.8 (Figure 4).
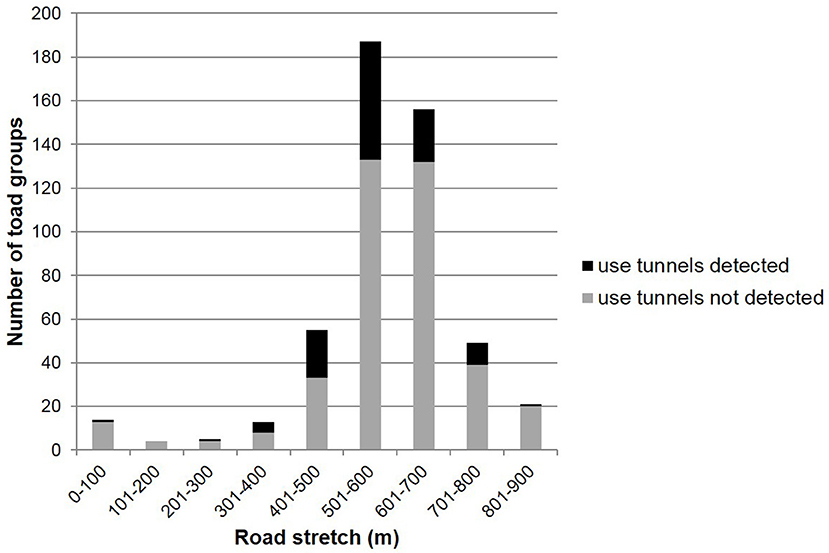
Figure 4. Distribution of toad groups that eventually did and did not use the tunnels along the drift fence during first capture over all study years.
The mean distance between the location of first capture and the nearest tunnel was significantly smaller (U = 16608, z = 4.45, P < 0.001) for toad groups that did use the tunnels as compared to toad groups that did not use the tunnels (Table 3). On average toad groups that used the tunnels encountered the drift fence almost 50 m closer to the nearest tunnel than toad groups that did not use the tunnels.
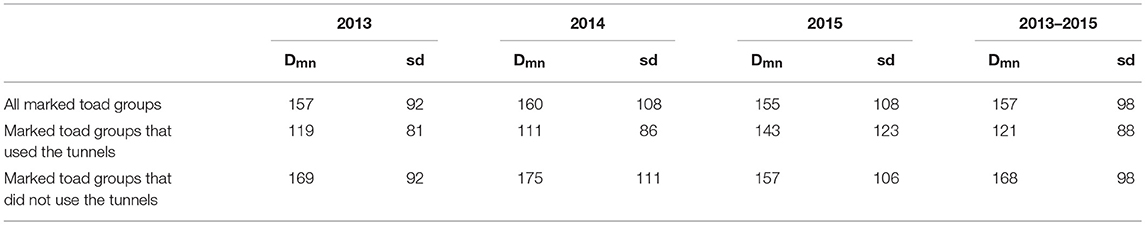
Table 3A. Mean distance (Dmn) and standard deviation of the mean (sd) between location of first capture and nearest tunnel per toad group per study year and over all study years.
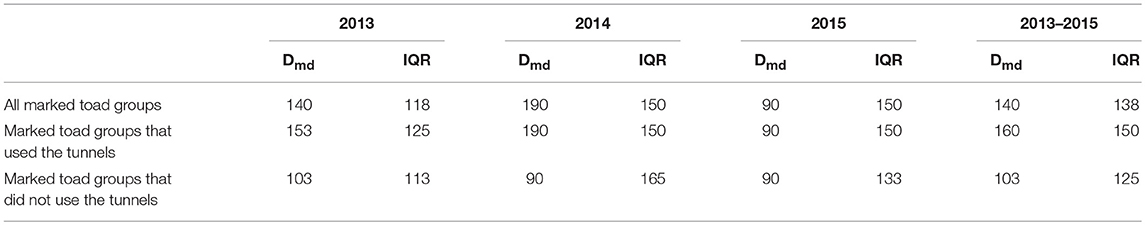
Table 3B. Median distance (Dmd) and interquartile range (IQR) between location of first capture and nearest tunnel per toad group per study year and over all study years.
The mean movement distance along the fence was significantly larger (U = 5993, z = 12.12, P < 0.001) for toad groups that did use the tunnels as compared to toad groups that did not use the tunnels (Table 4). On average toad groups that used the tunnels moved 120 m further along the drift fence than toad groups that did not use the tunnels.
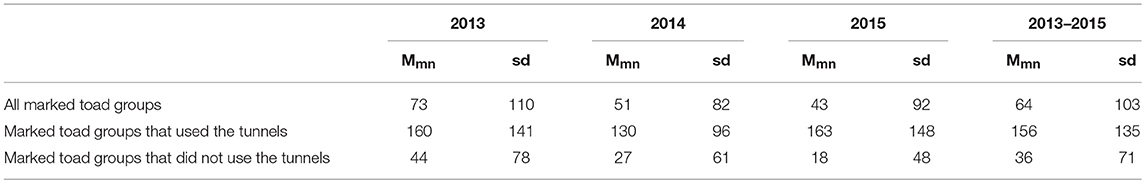
Table 4A. Mean movement distance (Mmn) and standard deviation of the mean (sd) along the drift fence per toad group per study year and over all study years.
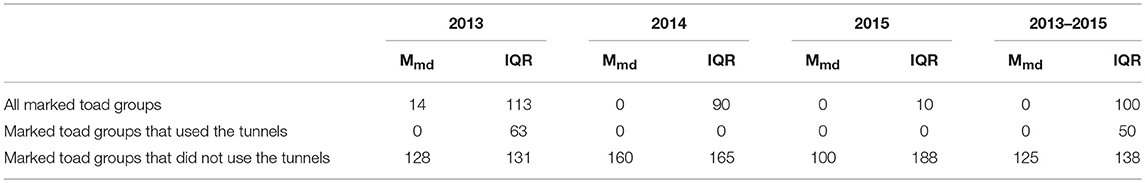
Table 4B. Median movement distance (Mmd) and interquartile range (IQR) along the drift fence per toad group per study year and over all study years.
The number of toad groups that walked past a tunnel was higher for toad groups that did use the tunnels as compared to toad groups that did not use the tunnels (Table 5). On average 12% of the toad groups that eventually used the tunnels first walked past a tunnel vs. 2% of toad groups that did not use the tunnels. A chi-square test showed there was an association between tunnel use and walking past one or more tunnels (χ2 = 18.86, d.f. = 1, P < 0.001).
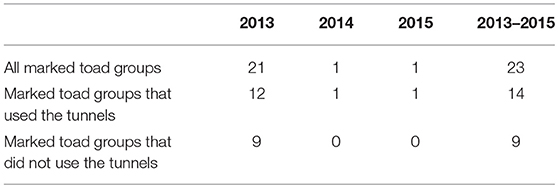
Table 5. Number of toad groups that walked past a tunnel per toad group and per study year and over all study years.
For 247 toad groups the movement direction along the drift fence is known after first capture, of which, respectively 129 and 118 toad groups did and did not use the tunnels eventually. The number of toad groups that walked in the wrong direction after encountering the drift fence was lower for toad groups that did use the tunnels as compared to toad groups that did not use the tunnels (Table 6). On average 13% of the toad groups that eventually used the tunnels first walked in the wrong direction vs. 37% of toad groups that did not use the tunnels. A chi-square test showed there was an association between tunnel use and the direction of movement after encountering the drift fence (χ2 = 19.46, d.f. = 1, P < 0.001).
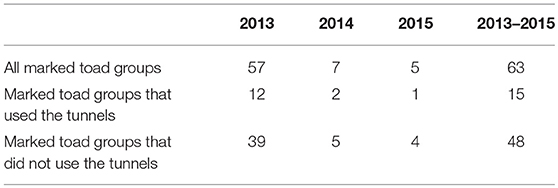
Table 6. Number of times that the wrong direction was chosen after encountering the drift fence per toad group and per study year and over all study years.
The mean number of nights between first and last capture of the toad group was significantly higher (U = 10433, z = 8.91, P < 0.001) for toad groups that did use the tunnels as compared to toad groups that did not use the tunnels (Table 7). On average, toad groups that used the tunnels spent two days more along the drift fence than toad groups that did not use the tunnels.
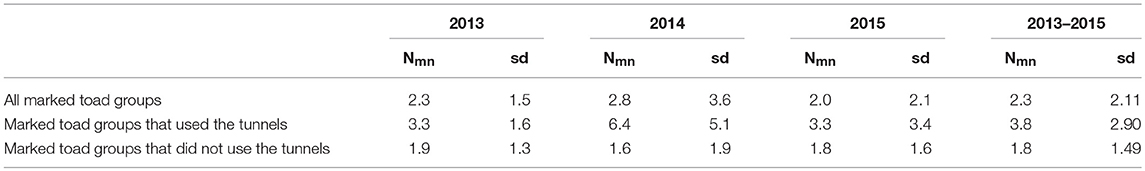
Table 7A. Mean number of nights (Nmn) and standard deviation of the mean (sd) between first and last capture per toad group per study year and over all study years.
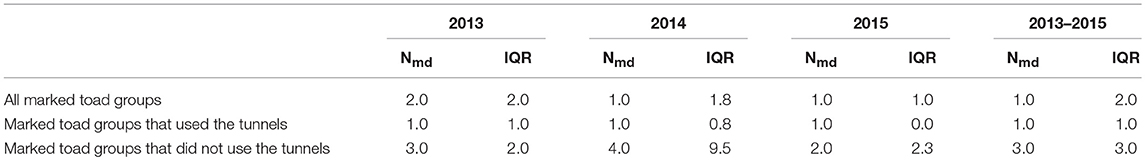
Table 7B. Median number of nights (Nmd) and interquartile range (IQR) between first and last capture per toad group per study year and over all study years.
Over all study years 28% of the migrating toads—marked and unmarked—that attempted to cross the road ended up on the road pavement, despite the mitigation (Figure 5). Of these 20% were found dead and 80% were found alive. The number of toads per road section steeply increases from west to east with <50 toads in section A to over 250 toads in section D (Figure 5).
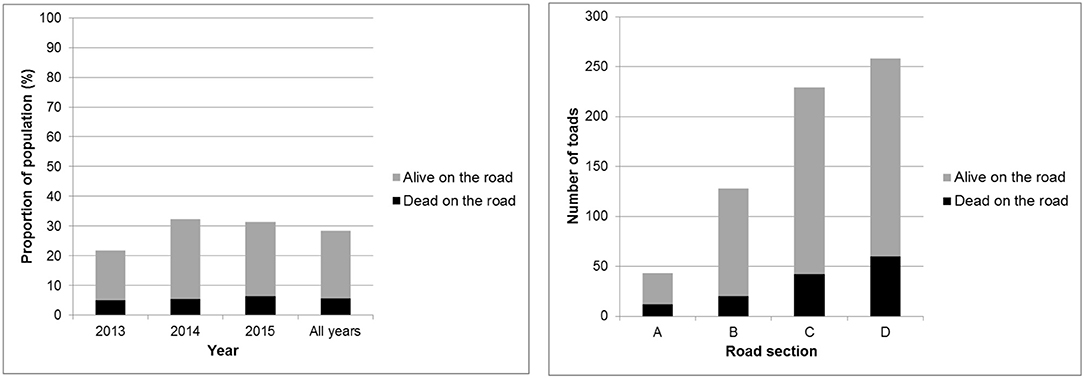
Figure 5. Proportion of the migrating toads that were found—dead or alive—in the road corridor (left) and distribution of these toads over the road sections (right).
Migrating population numbers decreased with about 75% after the mitigation measures were installed (Figure 6). Before mitigation the toad population that had to cross the road to reach the breeding ponds, was estimated at about 3,000 individuals. After mitigation, this population size was estimated at about 800 individuals.
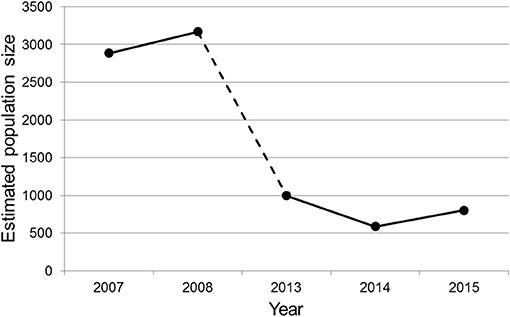
Figure 6. Trend in population size before (2007–2008) and after (2013–2015) the road mitigation measures were installed.
We found that a relatively small proportion of the toads that approached the road managed to get across via the two amphibian tunnels. The main reason behind this seems to be the low tunnel density (2.2/km) which may present too large a distance for most toads to cover. Mean movement distance along the fence for all toad groups is about 60 meters, while the mean distance to the nearest tunnel after encountering the drift fence is about 160 meters. Installing more tunnels will likely allow more toads to reach and use them to get across.
We found a relatively low CI and RPI. Both indices will be underestimated if toads that used the tunnels successfully were not captured in the pitfall traps at the tunnel exits or when marked toads lost their markings along the way. As the traps at the tunnel exits covered the full width of the tunnels, it was impossible for the toads to exit the tunnels on the north side without being captured. Some toads may have turned around after encountering the trap, however, we expect these numbers to be low, as a sudden drop into the traps was created at the tunnel exits. Although the loss of markings cannot be excluded, we expect this to have happened only occasionally. No lost markings were found during the surveys along the drift fence, in the tunnels or in the pitfall traps. Moreover, many marked toads were observed in the breeding ponds, more than a week after they had been marked along the drift fence.
Besides the distance between tunnels, design features of the tunnels may play a role in tunnel acceptance and use. If, for example, the tunnels are perceived by the toads as too small, too long or with an inhospitable micro-climate, they may avoid them. The fact that a proportion of the marked toad groups walked past a tunnel may indicate the existence of such an effect. However, as only 14% of all marked toad groups have been found to walk past a tunnel, we conclude that design features can only partially explain the low crossing rates. To gain more insight in the effect of tunnel design we recommend to carry out a study with the use of motion-triggered camera traps. This will allow to study the behavior of individual toads that arrive at the tunnel entrances and will help to assess what proportion of the toads that enter a tunnel pass it fully and what proportion turns around somewhere halfway.
The data show that the behavior of individual toads differs considerably. At one side of the spectrum there are toads that move large distances and spent considerable time along the drift fence in an attempt to get across the road (“runners”). On the other side there are toads that move only small distances and spent relatively little time along the drift fence (“strollers”). The runners are more successful in using the tunnels, even despite the fact that they walk past tunnels more often than the strollers. In decisions on the number of tunnels needed, the demands of the strollers should preferably be leading, as they have the greatest demands in respect to the maximum distance between tunnels.
If desired tunnel density is based on the mean movement distance (about 40 m; Table 4) of the toad groups that did not use the tunnels, 23 tunnels should be installed along the 0.9 kilometer-road stretch (tunnel density: 25/km). It should be noted, however, that the mean movement distance of the toad groups that did not use the tunnels is likely underestimated. Many non-crossing toad groups (n = 202; 52%) were not recaptured and hence their movement distance was set as 0 m. It is likely, however, that these toads did move along the fence after being released, but left the fence before the next survey was carried out. Hence, we recommend the use of the mean movement distance for all toads (about 60 m; Table 4) as rule-of-thumb for the maximum distance between tunnels. In that case 15 tunnels should be installed along the 0.9 kilometer-road stretch (tunnel density: 16.6/km). For future studies, we recommend to increase chances for recapturing toads, carrying out additional checks during both the morning and evening surveys, as this may reduce the number of “0 m movements.” We also recommend increasing the spatial resolution of the data through applying 1 m-markings on the drift fence. This will allow for better inferences about both capture locations and distances moved.
Another issue in relation to the desired number of tunnels is that the toads did not arrive evenly distributed over the fence length. About 90% of all toads arrived within a stretch of 400 m (Figure 4). If desired tunnel density is based on the mean movement distance of all toad groups (60 m) and is limited to this 400 meter-road stretch where most toads arrive, in total 7 tunnels should be installed. Hence, for future mitigation projects we recommend to explore the spatial distribution of locations where toads cross the road in detail prior to the installation of the mitigation measures. This will allow for tailor-made solutions and decrease the cost-benefit ratio.
Over all study years about 28% of the migrating toads ended up on the road. Most of these toads were still alive when found, but it seems reasonable to assume that a large proportion of them would have been killed eventually if they had not been captured and moved to the ponds as part of our study. Three possible explanations for these relatively high number of toads in the road corridor are (1) toads walk around the fence ends, (2) toads are able to jump over the drift fence, (3) toads use breaches in the drift fence, and (4) toads are able to pass the cattle guards at side roads. The first explanation certainly applies as we frequently observed toads entering the road surface beyond the fence ends during our surveys. This particularly occurred beyond the eastern fence end, which seems reflected in the high numbers of toads found in road section D (Figure 5). The southern drift fence is here 100 m shorter than the northern one, which means that toads that surpass the southern barrier will encounter the fence on the north side of the road and likely end up wandering on the pavement between the two fences. Some toads also appeared to be able to pass the cattle guards at five side roads, which are all located in road sections B and C. During our surveys we repeatedly observed how toads were able to balance themselves on the grid and enter the road corridor. No evidence has been found for toads jumping over the fence or using breaches in the fence. Before the surveys took place the drift fence was carefully checked for breaches and, if needed, repaired. Both before and during the surveys conditions that could possibly help toads jumping the fence, e.g., fallen branches or accumulation of litter, were removed. Table 2 shows that a relatively small proportion of marked toads ended up on the road. One possible explanation is that the markings can be easily missed after marked toads have been killed on the pavement, which may imply that the number of marked toads on the road is underestimated. Another explanation is that the number of toads that enter the road corridor at the fence ends is considerably higher than the number of toads that enter the road across the cattle guards at side roads. At the fence end the proportion of entering toads that is marked is likely lower, as these not only include toads that surpass the fence but also toads that approach the road beyond the fence end, where no capturing took place. The latter explanation is supported by the relatively high number of toads found in road section D (Figure 5). For future mitigation projects we recommend (1) apply drift fences over a length that goes well beyond the road stretch where the toads cross, as crossing locations may shift between years, (2) apply drift fences on both sides of the road that are equal in length, and (3) install toad-proof cattle guards in the road at both fence ends and at all side roads to prevent toads from entering the road corridor between the fences (Figure 7).

Figure 7. Example of a toad-proof cattle guard (left) and one that can still be bridged by toads (right). The opening between the road and the first steel bar in the toad-proof cattle guard is 6 vs. 4 cm in the one that can be bridged by toads. Photos: F. Ottburg.
The toad population that had to cross the road to reach the breeding ponds, decreased significantly after the mitigation measures were installed (Figure 6). One explanation may be the removal of the historic breeding waters, as the total size of all breeding waters was reduced from about 6,775 m2 in 2007–2008 (pre-mitigation) to 1,640 m2 in 2013–2015 (post-mitigation). Another plausible explanation for this steep decline is the relatively small number of toads that manages to get across the road and take part in breeding. Even more so if we assume that something similar applies to the migration of adults and juveniles back to the land habitat after breeding. Considering that—simultaneously with the installation of the mitigation—land habitat north of the road was lost or became inaccessible for toads due to city expansion, one could expect increasing numbers of toads that migrate to land habitat south of the road. However, the opposite has been observed. Instead of, or besides, building more tunnels to increase the proportion of toads that make it across the road and take part in breeding, the creation of breeding waters south of the road may be considered. Currently, toads that fail to use the tunnels will have no part in breeding as no ponds occur south of the road. Additional tunnels will increase the likelihood that toads will get across safely, but there may always be a proportion of the migrating toads that do not manage to reach or use the tunnels. With the creation of ponds south of the road these toads are offered an alternative breeding place. When such a measure is applied, we recommend the careful monitoring of the acceptance and use of the new ponds by toads and evaluate the effectiveness of the measure to increase participation in breeding and eventually population numbers.
During spring migration the road mitigation measures at our study site provide a safe passage across the road for a relatively small proportion of the migrating toads. Most toads do not manage to get across through the tunnels and consequently take no part in breeding. In addition toads frequently end up on the road where they run the risk of being killed by passing cars. This all seems to have seriously affected population numbers and may even threaten the survival of the population. Our research emphasizes that better baseline studies on where toads cross before mitigation and improved knowledge on the distances the animals move along a drift fence are vital to mitigate road impacts properly and maintain viable toad populations. We recommend to base tunnel densities on the mean movement distance of the toads that move only small distances and spent relatively little time along the drift fence, and install drift fences that go well beyond the location where toads cross the road. At entrance roads and at fence ends tailor-made measures are needed to prevent toads from entering the road corridor and start wandering between the fences. Alternative mitigation measures to tunnels and fences should always be considered, such as the creation of breeding waters on both sides of the road.
This study was carried out in accordance with the recommendations and approval of N. van den Brink, licensed coordinator animal experiments at Wageningen Environmental Research.
FO and EvdG: study design and data collection; EvdG: data analyses and writing manuscript; FO: reviewing manuscript.
The funding for the research was provided by Grondbank Bennekomseweg, Ede Municipality, and the CEDR Transnational Road Research Programme Call 2013: Roads and Wildlife, which was supported by the national road administrations of Austria, Denmark, Germany, Ireland, Norway, Sweden, Netherlands, and UK. Funding for the publication fee was provided by IENE.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank Carlo van Rijswijk (Ede Municipality) for his practical help in organizing and conducting the research as well as reviewing an earlier draft of this manuscript. We thank Wies van der Reijden, René van der Reijden, Wiebe Verbaan, and their team of volunteers (NGO Hora ést) for their tremendous help in preparing the experiment and collecting the data.
Andrews, K. M., Gibbons, J. W., and Jochimsen, D. M. (2008). Ecological effects of roads on amphibians and reptiles: a literature review. Herpetol. Conser. 3, 121–143.
Beebee, T. J. (2013). Effects of road mortality and mitigation measures on amphibian populations. Conser. Biol. 27, 657–668. doi: 10.1111/cobi.12063
Carr, L. W., and Fahrig, L. (2001). Effect of road traffic on two amphibian species of differing vagility. Conser. Biol. 15, 1071–1078. doi: 10.1046/j.1523-1739.2001.0150041071.x
Cooke, A. S. (2011). The role of road traffic in the near extinction of common toads (Bufo bufo) in Ramsey and Bury. Nat. Cambridge. 53, 45–50.
Fahrig, L., Pedlar, J. H., Pope, S. E., Taylor, P. D., and Wegner, J. F. (1995). Effect of road traffic on amphibian density. Biol. Conser. 73, 177–182. doi: 10.1016/0006-3207(94)00102-V
Forman, R. T. T., Sperling, D., Bissonette, J. A., Clevenger, A. P., Cutshall, C. D., Dale, V. H., et al. (2003). Road Ecology: Science and Solutions. Washington, DC: Island Press.
Gibbs, J. P., and Shriver, W. G. (2005). Can road mortality limit populations of pond-breeding amphibians? Wetlands Ecol. Manage. 13, 281–289. doi: 10.1007/s11273-004-7522-9
Glista, D. J., DeVault, T. L., and DeWoody, J. A. (2008). Vertebrate road mortality predominantly impacts amphibians. Herpetol Conser. Biol. 3, 77–87.
Glista, D. J., DeVault, T. L., and DeWoody, J. A. (2009). A review of mitigation measures for reducing wildlife mortality on roadways. Landscape Urban Plan. 91, 1–7. doi: 10.1016/j.landurbplan.2008.11.001
Hels, T., and Buchwald, E. (2001). The effect of road kills on amphibian populations. Biol. Conser. 99, 331–340. doi: 10.1016/S0006-3207(00)00215-9
Iuell, B., Bekker, G. J., Cuperus, R., Dufek, J., Fry, G., Hicks, C., et al. (eds.). (2003). “Cost Action 341. Habitat fragmentation due to transportation infrastructure,” in Wildlife and Traffic: A European Handbook for Identifying Conflicts and Designing Solutions (Utrecht: KNNV Publishers).
Jackson, S. D., Smith, D. J., and Gunson, K. E. (2015). “Mitigating road effects on small animals,” in Roads and Ecological Infrastructure - Concepts and Applications for Small Animals, eds K. M. Andrews, P. Nanjappa and S. P. D. Riley Johns (Baltimore, MD: Hopkins University Press), 177–207.
Niemi, M., Jääskeläinen, N. C., Nummi, P., Mäkel,ä, T., and Norrdahl, K. (2014). Dry paths effectively reduce road mortality of small and medium-sized terrestrial vertebrates. J. Env. Manage. 144, 51–57. doi: 10.1016/j.jenvman.2014.05.012
Ottburg, F. G. W. A., and van Blitterswijk, H. (2009). The Common Toad Population of Hoekelum, Horapark and ENKA-Site; Monitoring Population Size During the Spring Migration of 2007 and 2008. Alterra-report 1798. Wageningen, Alterra.
Santos, X., Llorente, G. A., Montori, A., Carretero, M. A., Franch, M., Garriga, N., et al. (2007). Evaluating factors affecting amphibian mortality on roads: the case of the Common Toad (Bufo bufo), near a breeding place. Animal Biodiver. Conser. 30, 97–104.
Schmidt, B. R., and Zumbach, S. (2008). Amphibian road mortality and how to prevent it: a review. Urban Herpetology, 157–167. doi: 10.5167/uzh-10142
van der Ree, R., Smith, D. J., and Grilo, C. (eds.). (2015). Handbook of Road Ecology. Chichester, UK: Wiley Blackwell.
van Gelder, J. V. (1973). A quantitative approach to the mortality resulting from traffic in a population of Bufo bufo L. Oecologia 13, 93–95. doi: 10.1007/BF00379622
Keywords: habitat fragmentation, road mortality, road mitigation, amphibian tunnel, amphibian fence, toad, population effect
Citation: Ottburg FGWA and van der Grift EA (2019) Effectiveness of Road Mitigation for Common Toads (Bufo bufo) in the Netherlands. Front. Ecol. Evol. 7:23. doi: 10.3389/fevo.2019.00023
Received: 06 June 2018; Accepted: 23 January 2019;
Published: 12 February 2019.
Edited by:
Clara Grilo, Brazilian Center for Road Ecology Studies (CBEE), BrazilReviewed by:
Tom Langen, Clarkson University, United StatesCopyright © 2019 Ottburg and van der Grift. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Edgar A. van der Grift, ZWRnYXIudmFuZGVyZ3JpZnRAd3VyLm5s
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.