
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 29 October 2018
Sec. Behavioral and Evolutionary Ecology
Volume 6 - 2018 | https://doi.org/10.3389/fevo.2018.00171
This article is part of the Research Topic How Prides of Lion Researchers are Evolving to be Interdisciplinary View all 12 articles
 Matthew Wijers1*
Matthew Wijers1* Paul Trethowan1
Paul Trethowan1 Andrew Markham2Byron du Preez1
Andrew Markham2Byron du Preez1 Simon Chamaillé-Jammes3,4Andrew Loveridge1David Macdonald1
Simon Chamaillé-Jammes3,4Andrew Loveridge1David Macdonald1Efforts to better understand patterns of animal behaviour have often been restricted by several environmental, human and experimental limitations associated with the collection of animal behavioural data. The introduction of new bio-logging technology has offered an alternative means of recording animal behaviour continuously and is being used in an increasing number of studies. Accurately calibrating these bio-loggers, however, still remains a challenge in many cases. Using lions as an example species, we test how audio recordings from animal-borne acoustic sensors can improve calibration and behaviour classification. Through a collaborative effort between computer scientists, engineers, and zoologists, custom designed acoustic bio-loggers were fitted to eight lions and recorded audio simultaneously with accelerometer and magnetometer data. Audio recordings were then used as the source of ground truth to train random forest classification models as well as to provide additional predictor variables for behaviour classification. We demonstrated near-perfect classification performance for five lion behaviour classes when all component variables were combined, with an average per-class precision of 98.5%. Using accelerometer features only, the audio-trained classifier predicted behaviours with an average per-class precision of 94.3%. On-animal audio recordings are therefore able to provide a valuable source of ground-truth for calibrating bio-loggers while also offering additional predictive features for increasing the accuracy of behaviour classification. This technological innovation has wide ranging application and provides a useful tool for behavioural ecologists wishing to collect fine scale behavioural data for animal research and conservation.
Remote data logging, also referred to as bio-logging or bio-telemetry, has evolved rapidly with new available technologies. Initially, studies focusing on animal spatial ecology were revolutionised by the introduction of GPS tracking methods in the 1980's which provide accurate and long-term location information at varying resolutions (Rutz and Hays, 2009). More recently, there has been a shift in focus to providing behavioural information in conjunction with location data using similar archival data-loggers in order to better understand the drivers of animal behaviour. To achieve this, a substantial collaborative effort between zoologists, computer scientists, and engineers has been required. The resulting technological advances have transformed the field of behavioural ecology with an increasing number of studies now relying on animal attached sensors to record behaviour (Brown et al., 2013). This rapid transition likely resulted from the need to overcome a number of difficulties associated with direct observation. These difficulties may include biases suffered as a result of observer presence (Caine, 1990; Gutzwiller et al., 1994) or the inability continuously to observe the focal animal if it is an elusive species, or a species that occurs in inaccessible habitats. In addition, direct observations require considerable time and effort on the part of the observer and thus can be heavily influenced by human physical limitations (Cagnacci et al., 2010).
While bio-loggers provide a solution to most of these challenges, they also have several drawbacks of their own. Firstly, the size of such devices may limit their use on smaller animals where it is not feasible to design a unit that weighs <2% of the animal's body mass. This is necessary to prevent behavioural changes and increases in energy expenditure (Cooke et al., 2004). Secondly, in most cases, researchers are still required to spend time in the field observing the study animal in order to calibrate the data generated by the bio-logger. This is commonly done using video cameras held by the observer with subsequent video labelling that can be matched to the corresponding bio-logger data by time stamps (Kawabata et al., 2014; McClune et al., 2014; Lush et al., 2015; Wang et al., 2015). Thirdly, and perhaps a more fundamental problem is that many types of bio-loggers do not achieve desirable results in discerning between behaviours. Recent studies still fail to differentiate accurately between more than three basic activities (Grünewälder et al., 2012; Lush et al., 2015; Wang et al., 2015).
The majority of bio-loggers used in animal behaviour studies generally rely on one or a combination of three microelectromechanical systems (MEMS) sensors: accelerometer, magnetometer and a gyroscope. An accelerometer measures the acceleration forces of the body to which it is attached (Albarbar et al., 2009) while a magnetometer measures magnetic field strength and direction (Herrera-May et al., 2016). Gyroscopes, although not as common, are used to measure angular rate of rotation (Piyabongkarn et al., 2005). In some cases, animal borne video cameras have been included to provide ground truth for directly calibrating accelerometer data but only provide visual validation for short periods due to the high power and data storage requirements for recording video (Watanabe and Takahashi, 2013; Volpov et al., 2015; Pagano et al., 2017). Audio recording can also be used to collect behavioural information as shown by Insley et al. (2008) on fur seals and Lynch et al. (2013) on deers. These studies inferred animal behaviour by visually reviewing spectrographic patterns but did not incorporate any statistical learning for automatic behaviour classification. To our knowledge, the use of audio recordings as a method of calibrating on-board movement sensors as an alternative to video footage and direct observation has not been tested. In this study, we present a novel method for calibrating bio-logger signals using simultaneously captured on-collar audio recordings from a custom designed bio-logger. In so doing, we provide suggested improvements to the issues surrounding bio-logger calibration and behaviour differentiation. We further demonstrate near-perfect (>99%) classification accuracy when we combine audio features with other sensor data, especially for behaviours which are typically misclassified using motion sensors alone (e.g., drinking water). Thus, capturing synchronised audio and multi-sensor data has not only the potential to provide detailed ground-truth, but also provides extremely accurate automatic behaviour classification.
The study took place on the privately-owned Bubye Valley Conservancy (BVC). The BVC is ~ 3,400 km2 and is located in the lowveld region of southern Zimbabwe between latitudes 21.209 and 21.851° South, and longitudes 29.798 and 30.521° East. We focused on the south-western section of BVC where an ongoing lion research project has been conducted since 2009. For a full description of the study site see du Preez et al. (2014).
Bio-loggers were custom designed through a collaborative research partnership between zoologists, computer scientists, and engineers with the overall objective of developing a device capable of recording lion behaviour continuously and accurately. The loggers were manufactured to attach onto existing lion tracking collars produced by Africa Wildlife Tracking (AWT), Pretoria, South Africa and measured ~ 50 × 20 × 30 mm with a mass of <150 g (Figure 1). Each unit comprised a triaxial accelerometer and magnetometer, with both sensors sampling at 32 Hz per axis and a mono-electret microphone sampling audio at 16 kHz with an 8-bit resolution. The microphone circuit used a compander to provide dynamic gain adjustment where more amplification is made when the ambient audio is quiet. Custom firmware was written for an 8-bit AVR microcontroller which also included a low-power 802.15.4 radio unit which was used for time-synchronising the bio-logger to a base station upon deployment. Data was logged to a 32 gigabyte micro-SD card. The bio-logger was powered by 3 CR123A lithium cells and encased in an epoxy resin reinforced housing, with a hydrophobic vent for the microphone. Table 1 shows the relative current draw for each particular sensor, including the cost of logging to the SD card. As can be seen, the audio sensor consumes nearly 100 times as much power as the accelerometer, and this is mainly due to the cost of storing the audio data into the SD card, as 16 kilobytes needs to be written per second, compared with 96 bytes per second for the accelerometer or magnetometer.

Figure 1. Image showing bio-logger bolted on to the GPS collar fitted to a lioness.
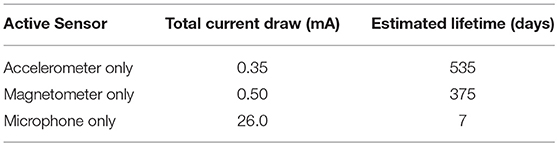
Table 1. Current draw and estimated lifetime using different sensors.
This study was carried out in accordance with the recommendations of the Use of Animals in Research, ASAB/ABS. The protocol was approved by the University of Oxford Animal Welfare and Ethical Review Board and the University Veterinary Services Department. Project staff were qualified to capture and handle the study animals by attendance at Zimbabwe's Physical and Chemical Capture of Wild Animals Course and held valid drugs licenses (Dangerous Drugs License No. 2014/16). The animals were captured with permission from the landowner and conservancy management.
In November 2014, we captured eight lions (five males and three females) that had been previously fitted with standard AWT satellite GPS collars. For a full description of the capture method see du Preez et al. (2015). Once the animals had been immobilized, the bio-loggers were bolted on to the existing GPS collars and started recorded audio (8 bit, 16 kHz mono) and three-dimensional accelerometer and magnetometer data (32 Hz) continuously until the batteries failed between 4 and 10 days later. Lions were recaptured ~ 1 month after initial capture and the loggers removed for data extraction.
In total, 80 predictor variables were calculated from the three accelerometer and magnetometer axes (40 variables for each component) for each 1 s window of data (Table 2). Many of the predictor features chosen for the movement sensor data have also been used in other studies (Gerencser et al., 2013; Wang et al., 2015). In addition to these features, 48 predictor variables were calculated from the corresponding audio recordings creating a combined feature set of 128 variables (see Table 2 for a description of each feature). Energy mean and variance were used as audio variables as they represent the zeroth and first order statistical moments of power in the 24 frequency bands as is often used in speech recognition (Kos et al., 2013). The energy mean captures whether a tone is present or not over a window, while energy variance better captures impulsive sounds such as foot falls during running.
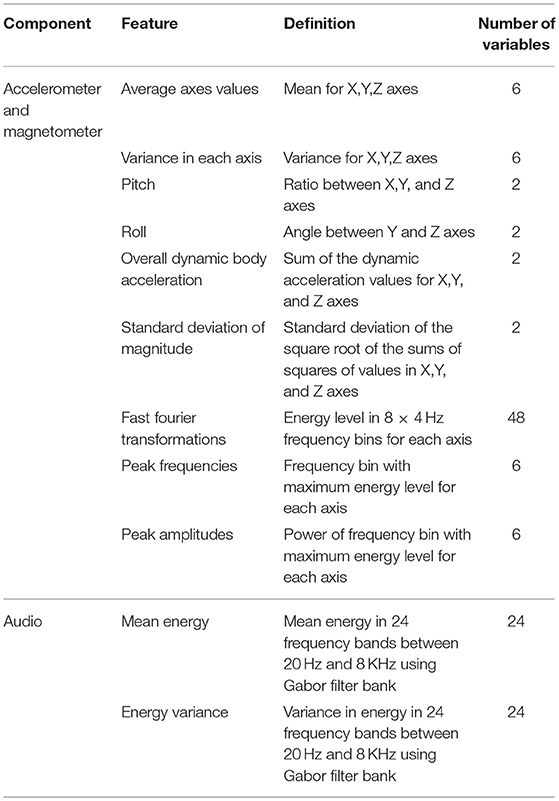
Table 2. Predictor variables calculated over each second of data, used for RF classification.
For each individual lion, random sections of audio recordings were labelled manually into one of five behavioural states (fast, slow, stationary, eat, and drink) by two lion ecologists with a minimum of 2 years of experience working on lions. We grouped running and trotting together as “fast” behaviour while walking was classed as “slow” behaviour. We were able to distinguish between these two behavioural states by the sound and pace of the lion's footfalls. Eating behaviour was discernible by the sound of chewing and bone crunching along with aggressive vocalisations that are often associated with group feeding. Drinking events were recognized by the sound of lapping water with regular swallowing (Samples of these audio recordings can be found in the Supplementary Material). We labelled a total of 20.5 h of audio which was then matched to logger measurements by corresponding time stamps. This resulted in a total labelled dataset of 73930 samples each 1 s long. We randomly subsampled this dataset to 16223 1 s samples by balancing across behavioural state and individual where possible to ensure that each individual and behaviour were sufficiently represented (Table 3). Poor representation of certain behavioural classes has been shown to reduce classification performance (Grünewälder et al., 2012). Additionally, class imbalances can result in a bias toward the over-represented classes (Stumpf and Kerle, 2011).
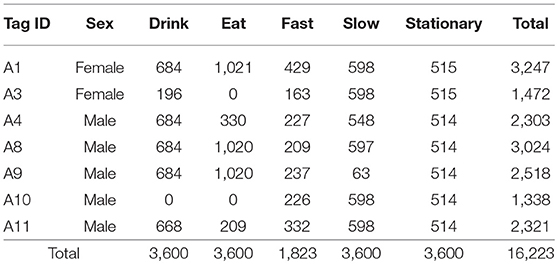
Table 3. Summary of balanced dataset showing number of seconds for each behaviour and individual.
We used the Random Forest (RF) classification method developed by Breiman (2001) to infer behaviour from bio-logger measurements. This method is advantageous as it is computationally fast, robust to outliers and noise and also offers variable importance estimates for classification (Breiman, 2001). The analysis was done using the random forest package (Breiman, 2001) in the R statistical program (R Core Team, 2016), within the R studio integrated development environment (R Studio Team, 2016). For all models, we set the number of trees (ntree) to 1,000 and used the recommended value () for the number of variables considered at each split (mtry) which has been shown to yield optimal performance (Díaz-Uriarte and Alvarez de Andrés, 2006).
We carried out a 5-fold cross-validation to train and test two RF models, one with all component features combined and another with only accelerometer features. We compared the behaviour classification performance of the models using accuracy, precision and recall. Accuracy is a measure of overall model performance and is defined as the proportion of correctly classified data. Precision is defined as the proportion of correctly predicted positive classifications for a particular behavioural state while recall (also called sensitivity) refers to the proportion of data of a particular behavioural state that is classified correctly as positive (Sokolova and Lapalme, 2009; Bidder et al., 2014). We used accuracy as the overall performance metric due to its simplicity and the fact that it takes into account all classification outcomes (Bidder et al., 2014; Wang et al., 2015). To evaluate prediction performance for each behavioural state, we used precision as the main performance metric as it is most applicable to biological inferences which generally rely on true positive classifications, as was the case in this study (Bidder et al., 2014). We included recall as recommended by Bidder et al. (2014) for novel classification methods.
We collected a total of 44 lion-days of useable data from 7 individual lions. One female lion was excluded from the final dataset as the magnetometer malfunctioned from the time of deployment. Our final subsampled dataset consisted of 16,223 data points with an hour of data for each behavioural class except “fast” for which we could only accumulate 1,823 s of data.
Combining all component features resulted in near perfect classification performance with an average per-class precision of 98.5% (Table 4A). Drink, fast, slow and stationary behaviours were predicted with ~99% precision while eating was ~3% lower with a precision of 96.2%. Training the classifier using accelerometer features only, resulted in an average per class precision of 94.3% (Table 4B) with only eating behaviour being predicted with <90% precision.
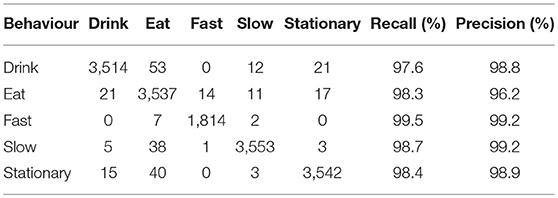
Table 4A. Confusion matrix of actual (rows) vs. predicted (columns) behaviours for audio, accelerometer, and magnetometer features combined.
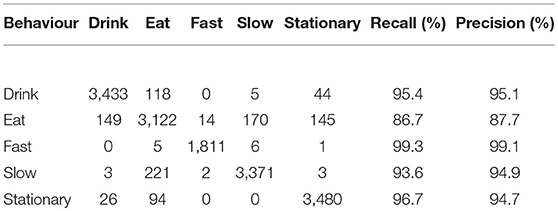
Table 4B. Confusion matrix of actual (rows) vs. predicted (columns) behaviours for accelerometer features only.
Recording the active behavioural states of African lions such as running, drinking or eating by directly observing study individuals can often be difficult as lions are mostly active at night and can be challenging to follow in areas with thick vegetation such as BVC. Adult lions, weighing in excess of 150 kg, are capable of carrying bio-logging devices attached to collars which can offer unique insights into their behavioural patterns. Using custom-designed acoustic bio-loggers, we found that audio can be used as an effective source of ground truth for training accurate behaviour classification models.
Six years ago Grünewälder et al. (2012) suggested that the collection of behavioural observations for calibrating bio-logging devices could be done remotely in the near future. Achieving this objective, however, required the integration of technological and zoological knowledge and skills both for the development and data analysis phases, which was achieved through an interdisciplinary research partnership. Our results indicate that remotely collected audio recordings can be used as a reliable source of ground truth for calibrating bio-loggers by matching audio labels to logger data following logger retrieval and thereby eliminate the need for calibration from direct observations (Figure 2 illustrates how audio and movement sensor data are synchronised).
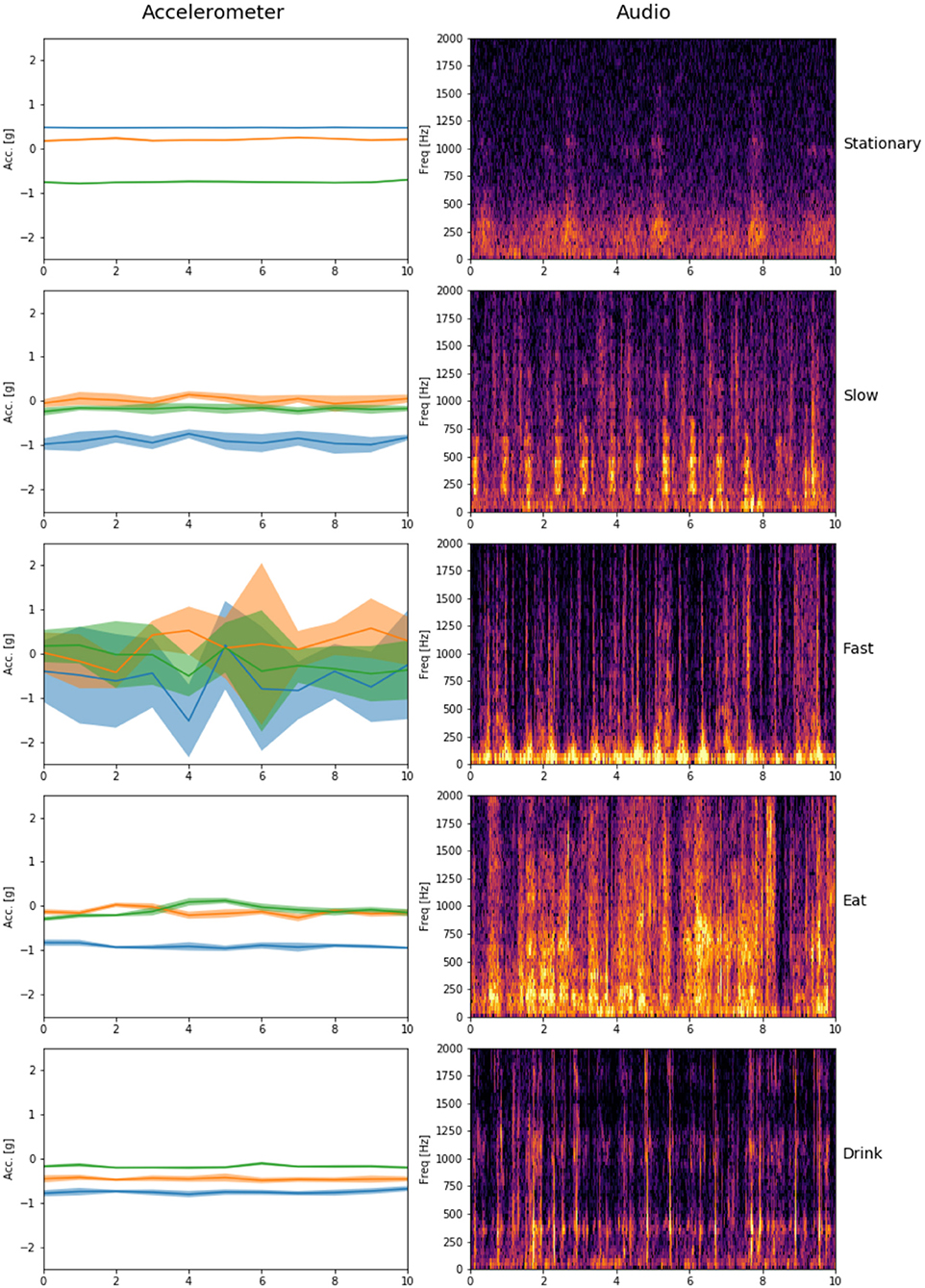
Figure 2. Synchronised accelerometer data and audio spectrograms for each behavioural state. Lines and shaded regions on accelerometer plots represent mean and standard deviation, respectively for each accelerometer axis.
It is useful to note that a relatively small number of ground truth labels (1 h per behavioural state) were required to build an accurate classifier, although it is important to ensure that all behavioural classes are sufficiently represented in the training dataset (Grünewälder et al., 2012). This requirement can be fulfilled by continuously logging audio over several days which increases the likelihood of recording rarer behavioural events.
The total of 44 lion-days of audio from 7 individuals provided considerably more data for calibration of rarer behavioural events (eat, drink, and fast) than could have been realistically achieved using video footage recorded by an observer or a video collar. In comparison, Pagano et al. (2017) recorded a total of 140 h of video for accelerometer validation from 5 ice bears fitted with video collars. Although visual determination of animal behavioural states is likely to be more objective than those which are determined audibly from sound recordings, we found that in general, the behaviour of the study animal could be determined easily from certain acoustic cues as outlined in section Data management. Insley et al. (2008) who also made use of an animal-borne acoustic recording device reported being able to clearly differentiate between resting and other active behaviours of northern fur seals. Similarly, Lynch et al. (2013) list in detail, the audible behaviours that could be captured by their animal-borne acoustic devices fitted to wild mule deer. While most behaviours may be clearly discernible using this method, short periods of ambiguous sound signals will likely be recorded and, depending on the objective of the research, may require concurrent observational data collection to confirm the behavioural state (Lynch et al., 2013). In some cases, interference from other sound sources may also make it difficult to determine behaviour. Such interference may result from self-vocalisations, vocalisations emitted by other species, anthropogenic sources (e.g., vehicles) or environmental sources (e.g., wind and rain). The use of this approach should also consider the acoustic characteristics of the species of interest as the behaviour of certain species may not be sufficiently audible, even at close range.
The results from the model built using the different component datasets showed that audio and magnetometers can also be used as additional sensor modalities for classification with high model predictive performance when accelerometer, magnetometer and audio features are combined. However, due to the considerably higher power consumption of audio recording and the battery capacity limits on current bio-loggers, it is unlikely that continuously logging raw audio would be a suitable sensor modality for long-term logger deployments. However, scheduling (e.g., sampling for only a few hours a day) could dramatically increase lifetime whilst still providing a sufficiently representative training set. Despite this drawback, we still demonstrated good model predictive performance using the audio labels and accelerometer features only. Thus, a small subset of animals can be equipped with audio and motion loggers to provide ground-truth calibration for a larger set of animals equipped only with motion loggers.
While we have primarily highlighted the use of audio for training behaviour classifiers, bio-loggers fitted with microphones may also be useful tools for other study purposes such as investigating how species respond to environmental acoustic stimuli or exploring patterns of animal vocal behaviour (Stowell et al., 2017; Wisniewska et al., 2018). The audio recordings collected from our bio-loggers often revealed the presence of other species (e.g., antelope and baboon alarm calls) and in some cases also allowed for the identification of captured prey species from the prey distress vocalisations. Such contextual information could be particularly useful where opportunities for visual observations are rare. Furthermore, we were able to identify more than 300 roar events from the 5 male lions in this study. This data alone could be used to assess vocalisation rates as well as provide insight into the spatial patterns of vocalisations when combined with GPS collar data.
In future, acoustic bio-loggers could be greatly enhanced by intelligent on-board processing functions aimed at reducing battery load by either limiting recording to sounds of interest or by storing audio variables rather than raw audio samples. Consideration must also be given to the mode of data retrieval with wireless data transmission being a preferred option. These advancements would be particularly beneficial to studies on smaller species, where battery capacity is limited, and elusive species, where logger retrieval is difficult.
Few published studies have reported the use of micro-sensors to investigate aspects of lion behaviour (Wilson et al., 2018) however, with advances in technology and the development of interdisciplinary research partnerships, opportunities to overcome previous study limitations have arisen. Gao et al. (2013) suggested that one of the main challenges associated with analysing accelerometer data from wild animals is that there is often very little observational data to generate an accurate behaviour classifier. We have demonstrated how on-animal audio recordings can be used to collect a large amount of ground truth data for training accurate classifiers. Acoustic bio-loggers have wide-ranging application and this work can inform the design and development of future bio-loggers for other animal behaviour studies.
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
SC-J, AL, and DM conceptualised the addition of audio sensors to bio-loggers. AM designed the bio-loggers and assisted in the extraction and analysis of data. Study animals were captured and fitted with bio-loggers by PT and BdP. MW carried out the data analysis and wrote the manuscript with input from all authors.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor declared a past collaboration with one of the authors, DM.
We are grateful to the John Fell Fund and the Beit Trust for funding this research and thank the management staff of the Bubye valley Conservancy for giving us access to their property. We also thank Biotrack for their assistance in the manufacturing of the bio-loggers. Finally, we thank the editor and the two reviewers for their constructive comments.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00171/full#supplementary-material
Audio 1. Eating.
Audio 2. Drinking.
Audio 3. Fast.
Audio 4. Slow.
Audio 5. Stationary.
Albarbar, A., Badri, A., Sinha, J. K., and Starr, A. (2009). Performance evaluation of MEMS accelerometers. Measurement 42, 790–795. doi: 10.1016/j.measurement.2008.12.002
Bidder, O. R., Campbell, H. A., Gómez-Laich, A., Urgé, P., Walker, J., Cai, Y., et al. (2014). Love thy neighbour: automatic animal behavioural classification of acceleration data using the K-nearest neighbour algorithm. PLoS ONE 9:e88609. doi: 10.1371/journal.pone.0088609
Brown, D. D., Kays, R., Wikelski, M., Wilson, R., and Klimley, A. P. (2013). Observing the unwatchable through acceleration logging of animal behavior. Anim. Biotelemetry 1, 1–16. doi: 10.1186/2050-3385-1-20
Cagnacci, F., Boitani, L., Roger, A., Powell, and Boyce, M. S. (2010). Animal ecology meets GPS-based radiotelemetry: a perfect storm of opportunities and challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. 365, 2157–2162. doi: 10.1098/rstb.2010.0107
Caine, N. G. (1990). Unrecognized anti-predator behavior can bias observational data. Anim. Behav. 39, 195–197. doi: 10.1016/S0003-3472(05)80741-9
Cooke, S. J., Hinch, S. G., Wikelski, M., Andrews, R. D., Kuchel, L. J., Wolcott, T. G., et al. (2004). Biotelemetry: a mechanistic approach to ecology. Trends Ecol. Evol. 19, 334–343. doi: 10.1016/j.tree.2004.04.003
Díaz-Uriarte, R., and Alvarez de Andrés, S. (2006). Gene selection and classification of microarray data using random forest. BMC Bioinformatics 7:3. doi: 10.1186/1471-2105-7-3
du Preez, B., Hart, T., Loveridge, A. J., and Macdonald, D. W. (2015). Impact of risk on animal behaviour and habitat transition probabilities. Anim. Behav. 100, 22–37. doi: 10.1016/j.anbehav.2014.10.025
du Preez, B. D., Loveridge, A. J., and Macdonald, D. W. (2014). To bait or not to bait: a comparison of camera-trapping methods for estimating leopard Panthera pardus density. Biol. Conserv. 176, 153–161. doi: 10.1016/j.biocon.2014.05.021
Gao, L., Campbell, H. A., Bidder, O. R., and Hunter, J. (2013). A web-based semantic tagging and activity recognition system for species' accelerometry data. Ecol. Inform. 13, 47–56. doi: 10.1016/j.ecoinf.2012.09.003
Gerencser, L., Vasarhelyi, G., Nagy, M., Vicsek, T., and Miklosi, A. (2013). Identification of behaviour in freely moving dogs (Canis familiaris) using inertial sensors. PLoS ONE 8:e77814. doi: 10.1371/journal.pone.0077814
Grünewälder, S., Broekhuis, F., Macdonald, D. W., Wilson, A. M., McNutt, J. W., Shawe-Taylor, J., et al. (2012). Movement activity based classification of animal behaviour with an application to data from cheetah (Acinonyx jubatus). PLoS ONE 7:e49120. doi: 10.1371/journal.pone.0049120
Gutzwiller, K. J., Wiedenmann, R. T., Clements, K. L., and Anderson, S. H. (1994). Effects of human intrusion on song occurrence and singing consistency in subalpine birds. Auk 111, 28–37. doi: 10.2307/4088502
Herrera-May, A., Soler-Balcazar, J., Vázquez-Leal, H., Martínez-Castillo, J., Vigueras-Zuñiga, M., and Aguilera-Cortés, L. (2016). Recent advances of MEMS resonators for Lorentz force based magnetic field sensors: design, applications and challenges. Sensors 16, 1–25. doi: 10.3390/s16091359
Insley, S. J., Robson, B. W., Yack, T., Ream, R. R., and Burgess, W. C. (2008). Acoustic determination of activity and flipper stroke rate in foraging northern fur seal females. Endanger. Species Res. 4, 147–155. doi: 10.3354/esr00050
Kawabata, Y., Noda, T., Nakashima, Y., Nanami, A., Sato, T., Takebe, T., et al. (2014). Use of a gyroscope/accelerometer data logger to identify alternative feeding behaviours in fish. J. Exp. Biol. 217, 3204–3208. doi: 10.1242/jeb.108001
Kos, M., Kačič, Z., and Vlaj, D. (2013). Acoustic classification and segmentation using modified spectral roll-off and variance-based features. Digit. Signal Proc. 23, 659–674. doi: 10.1016/j.dsp.2012.10.008
Lush, L., Ellwood, S., Markham, A., Ward, A. I., and Wheeler, P. (2015). Use of tri-axial accelerometers to assess terrestrial mammal behaviour in the wild. J. Zool. 298, 257–265. doi: 10.1111/jzo.12308
Lynch, E., Angeloni, L., Fristrup, K., Joyce, D., and Wittemyer, G. (2013). The use of on-animal acoustical recording devices for studying animal behavior. Ecol. Evol. 3, 2030–2037. doi: 10.1002/ece3.608
McClune, D. W., Marks, N. J., Wilson, R. P., Houghton, J. D., Montgomery, I. W., McGowan, N. E., et al. (2014). Tri-axial accelerometers quantify behaviour in the Eurasian badger (Meles meles): towards an automated interpretation of field data. Anim. Biotelemetry 2, 1–6. doi: 10.1186/2050-3385-2-5
Pagano, A. M., Rode, K. D., Cutting, A., Owen, M. A., Jensen, S., Ware, J. V., et al. (2017). Using tri-axial accelerometers to identify wild polar bear behaviors. Endanger. Species Res. 32, 19–33. doi: 10.3354/esr00779
Piyabongkarn, D., Rajamani, R., and Greminger, M. (2005). The development of a MEMS gyroscope for absolute angle measurement. IEEE Trans. Control Syst. Technol. 13, 185–195. doi: 10.1109/T.C.S.T.2004.839568
R Core Team (2016). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. Available online at: https://www.R-project.org/
R Studio Team. (2016). RStudio: Integrated Development for R. RStudio, Inc., Boston, MA Available online at: https://www.rstudio.com/
Rutz, C., and Hays, G. C. (2009). New frontiers in biologging science. Biol. Lett. 5, 289–292. doi: 10.1098/rsbl.2009.0089
Sokolova, M., and Lapalme, G. (2009). A systematic analysis of performance measures for classification tasks. Inf. Process. Manag. 45, 427–437. doi: 10.1016/j.ipm.2009.03.002
Stowell, D., Benetos, E., and Gill, L. F. (2017). On-bird sound recordings: automatic acoustic recognition of activities and contexts. IEEE/ACM Trans. Audio Speech Lang. Process. 25, 1193–1206. doi: 10.1109/TASLP.2017.2690565
Stumpf, A., and Kerle, N. (2011). Remote sensing of environment object-oriented mapping of landslides using random forests. Remote Sens. Environ. 115, 2564–2577. doi: 10.1016/j.rse.2011.05.013
Volpov, B. L., Hoskins, A. J., Battaile, B. C., Viviant, M., Wheatley, K. E., Marshall, G., et al. (2015). Identification of prey captures in Australian fur seals (Arctocephalus pusillus doriferus) using head-mounted accelerometers: field validation with animal-borne video cameras. PLoS ONE 10:e0128789. doi: 10.1371/journal.pone.0128789
Wang, Y., Nickel, B., Rutishauser, M., Bryce, C. M., Williams, T. M., Elkaim, G., et al. (2015). Movement, resting, and attack behaviors of wild pumas are revealed by tri-axial accelerometer measurements. Mov. Ecol. 3, 1–12. doi: 10.1186/s40462-015-0030-0
Watanabe, Y. Y., and Takahashi, A. (2013). Linking animal-borne video to accelerometers reveals prey capture variability. Proc. Natl. Acad. Sci. U.S.A. 110, 2199–2204. doi: 10.1073/pnas.1216244110
Wilson, A. M., Hubel, T. Y., Wilshin, S. D., Lowe, J. C., Lorenc, M., Oliver, P., et al. (2018). Biomechanics of predator - prey arms race in lion, zebra, cheetah and impala. Nat. Publ. Gr. 554, 183–188. doi: 10.1038/nature25479
Keywords: African lion, acoustic monitoring, behaviour classification, bio-logger calibration, machine learning, random forest
Citation: Wijers M, Trethowan P, Markham A, du Preez B, Chamaillé-Jammes S, Loveridge A and Macdonald D (2018) Listening to Lions: Animal-Borne Acoustic Sensors Improve Bio-logger Calibration and Behaviour Classification Performance. Front. Ecol. Evol. 6:171. doi: 10.3389/fevo.2018.00171
Received: 11 April 2018; Accepted: 08 October 2018;
Published: 29 October 2018.
Edited by:
Matt W. Hayward, University of Newcastle, AustraliaReviewed by:
Richard Anthony Peters, La Trobe University, AustraliaCopyright © 2018 Wijers, Trethowan, Markham, du Preez, Chamaillé-Jammes, Loveridge and Macdonald. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matthew Wijers, bWF0dGhldy53aWplcnNAem9vLm94LmFjLnVr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.