 Martin Heil
Martin Heil
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Ecol. Evol., 28 June 2016
Sec. Evolutionary and Population Genetics
Volume 4 - 2016 | https://doi.org/10.3389/fevo.2016.00080
This article is part of the Research TopicThe ecological and evolutionary consequences of parasitic infections through alterations of host behaviourView all 4 articles
Parasites must overcome host immunity and change hosts for dispersal. Therefore, seemingly odd behaviors of parasitized animals, like those exhibited by “Zombie ants” or the “fatal attraction” of mammals to their predators, have been explained as the extended phenotype of parasites that manipulate their hosts for transmission enhancement. Manipulation has evolved in all major phylogenetic lineages of parasites but is not ubiquitous. In fact, the real frequency and relevance of manipulation is still matter of debate. Here, I highlight some of the most pertinent questions that arise if we aim at a broader understanding of the ecological relevance and evolutionary trajectory of manipulating parasites. Why are more parasites with a trophic mode of transmission manipulating their host than horizontally transmitted parasites? Why are the causal agents of sexually transmitted diseases (STDs) so rarely reported to manipulate their host? Why do parasites of animals usually manipulate their intermediate host, whereas many pathogens of plants manipulate their final host and then cause a “fatal attraction” of herbivores to the already infected plant? How can we distinguish manipulation effects from adaptive host responses to parasitization? Does manipulation cause a cost to certain parasites that can select against the evolution of manipulation? More emphasis should be put on direct comparisons among parasites of animals, plants, and humans. Manipulation for transmission enhancement should be studied in concert with manipulation for host immunity suppression, the molecular mechanisms that control the manipulation effect need to be deciphered, and we should test for alternative explanations for the observed phenotypic changes. Finally, we should quantify transmission rates and net fitness for manipulating and non-manipulating parasites, in ecologically realistic setups, to identify the forces that select against the evolution of manipulation. Ultimately, parasite net fitness represents the relevant outcome of any manipulation effect.
Parasites represent a ubiquitous aspect in the life of multicellular organisms (Wood and Johnson, 2015) and most, if not all, of us have experienced how strongly an infection can interfere with our normal activities and well-being. Therefore, it might not appear to be too surprising when parasitized hosts exhibit behavioral alterations (see Glossary) as compared to healthy conspecifics. Nevertheless, for more than a century, naturalists have been puzzled by “odd,” seemingly pointless or counterproductive behaviors of parasitized animals, which have been described as “Zombie ants,” “lighthouse snails,” or “suicide-committing crickets” (Poulin, 2010; Hughes, 2014; Weinersmith and Faulkes, 2014). “Fatal attraction” has been observed for mammalian hosts that carry the protozoon Toxoplasma gondii: infected mammals enhance their exposure to feline predators, infected males become sexually more attractive to healthy females, and men with latent toxoplasmosis become more risk-taking and aggressive (Poulin, 2010; Hari Dass et al., 2011; Adamo, 2013; Flegr, 2013a; Weinersmith and Faulkes, 2014; Poirotte et al., 2016).
The most frequently discussed explanation for these behaviors is formulated in the “adaptive host manipulation hypothesis”: parasites can evolve to control specific aspects of their host's behavior and thereby enhance the frequency of encounters among suitable hosts and, thus, their rate of transmission (Holmes and Bethel, 1972). Thus, the altered behavior (or, in general, phenotype) of the host is assumed to be under the genetic control of the parasite and to represent its “extended phenotype” (Dawkins, 1999). This paradigm has served as a successful framework for the interpretation of multiple phenotypic alterations that would contradict the theory of adaptation if they were under the genetic control of the parasitized host itself. “Fatal attraction” is a particularly common outcome of adaptive host manipulation, because situations that represent an “encounter among suitable hosts” from the perspective of a trophically transmitted parasite are usually unfavorable “predation” events for at least one of the involved hosts (Table 1).
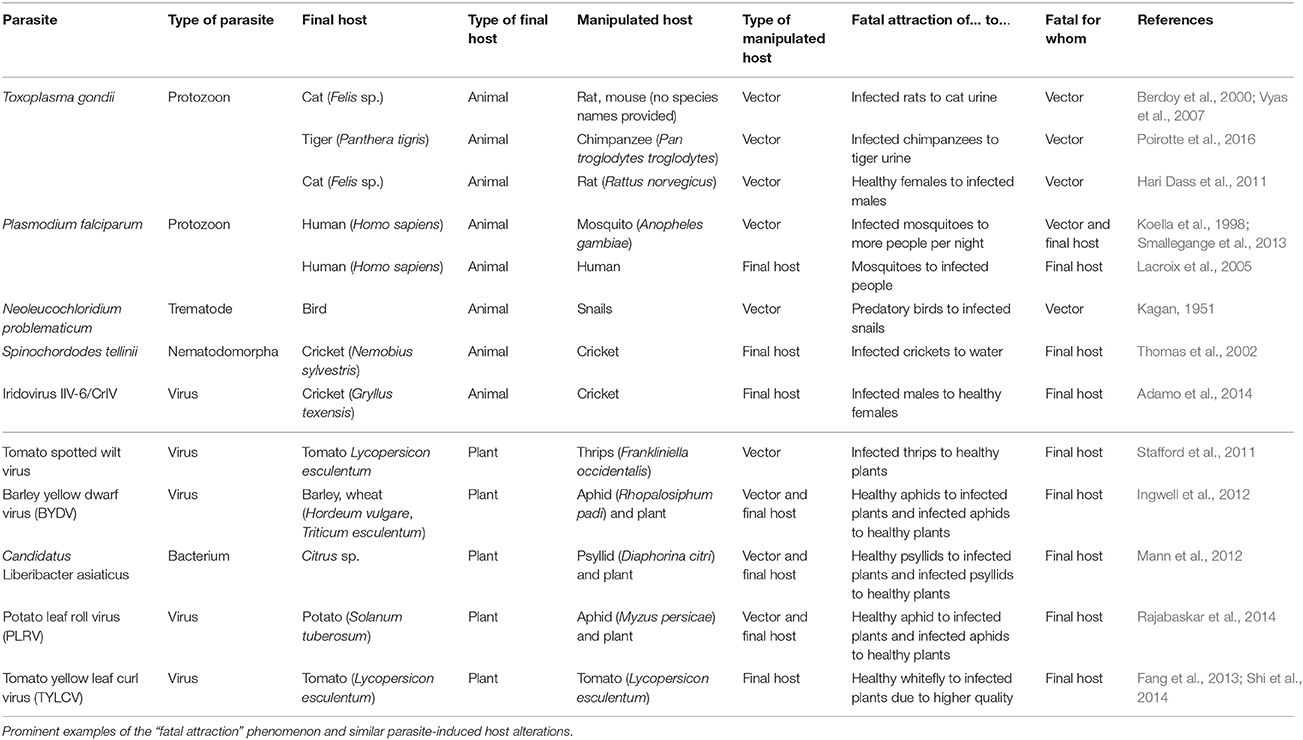
Table 1. Fatal attraction in plant and animal hosts.
Here, I discuss which parasites are most likely to manipulate their hosts, which hosts are the most prone to manipulation, and which alternative mechanisms might account for phenotypic alterations in parasitized organisms. I expand the spectrum of biological models that have been covered in other, recent reviews (Thomas et al., 2005, 2010; Libersat et al., 2009; Poulin, 2010; Flegr, 2013a; Weinersmith and Faulkes, 2014) to include plant pathogens and the causal agents of sexually transmitted diseases (STDs). A rapidly growing body of literature indicates that plant-infecting viruses, bacteria or fungi manipulate multiple traits of their hosts and vectors (van Houte et al., 2013), whereas reports on host manipulation by microbial STD agents are surprisingly scarce, in spite of abundant literature on the STDs of humans and of the obvious potential benefits that these parasites could gain from an adequate manipulation of host behavior. I argue that comparing the strategies and effects of the parasites of plants, animals and humans can help to identify general patterns in the evolution of manipulation and that the almost complete absence of STDs from the respective literature could indicate biases in research or publishing activities, but it could also mean that the effects of host manipulation by STD agents are inconspicuous.
The expression of a host phenotype that is under the genetic control of a parasite is responsible for several biological marvels. Nevertheless, it requires more than observing an “odd” behavior in a parasitized animal if we aim to unambiguously identify the parasite as the species that has genetic control over the observed phenotype. Do I sneeze or cough to get rid of the pathogens in my respiratory system, that is, do these behaviors form part of my adaptive immune response, or am I being manipulated by the microorganisms to favor their dispersal? Mammals respond to infection with sickness behavior (Hart, 1988), fatigue, fever, inflammation, and multiple cellular and molecular immune responses, such as the activation of antigen-presenting cells, priming of T-cells and the proliferation of B-cells (Janeway et al., 2001). Sickness behavior and cellular immune responses have also been shown in insects (Adamo, 2014). The equivalent responses in parasitized plants comprise a coordinated switch from primary to secondary metabolism (Heil and Baldwin, 2002; Scheideler et al., 2002), the re-allocation of nutrients and the de novo synthesis of an extremely diverse array of resistance-related structures, secondary metabolites, peptides and proteins (Karban and Baldwin, 1997; Jones and Dangl, 2006; Dangl et al., 2013). All these phenotypic alterations contribute to a successful immune response against the parasite and, thus, are reasonably assumed to be under the genetic control of the host.
Interestingly, an adaptive immune response of the host can result in phenotypic changes that could easily be interpreted as the results of a manipulation effect. For example, Malaria parasites, Plasmodium spec., are well known to change the behavior of infected mosquitoes: mosquitoes in an early stage of infection exhibit reduced attraction to human hosts, whereas those with late-stage infections (i.e., carrying infectious sporozoites) exhibit enhanced attraction to humans and tend to bite more people per night (Koella et al., 1998; Cator et al., 2013; Smallegange et al., 2013). These stage-specific changes in behavior were paralleled by changes in the responsiveness of mosquito odorant receptors, which indicates that the changed behavior has a neurophysiological basis. However, the same behavioral and neurophysiological changes were also elicited by treating the mosquitoes with heat-killed Escherichia coli (Cator et al., 2013) and were functionally linked to changes in insulin signaling in the mosquito gut (Cator et al., 2015). These results suggest that the altered mosquito phenotypes were caused by insulin signaling-dependent changes in host resource allocation patterns and, thus, likely to be under the genetic control of the host (Cator et al., 2015), rather than representing the outcome of a specific manipulation by Plasmodium.
Classical examples of manipulated phenotypes have been described as “lighthouse snails” and “Zombie ants,” and both phenotypes are elicited by trematodes (class Trematoda, Platyhelminthes). Leucochloridium spp. change the color and shape of the tentacles of snails that serve as their intermediate hosts and make them pulsate in response to light, thereby attracting predatory birds that represent the final hosts (Kagan, 1951). Dicrocoelium dendriticum-infected ants start to climb and bite into the tips of grasses, particularly at low temperatures, and thereby facilitate predation by ruminants (Moore, 2002). Interestingly, a phenotypically similar “zombie-like” behavior can also be elicited by fungal parasites such as Ophiocordyceps unilateralis. This entomopathogenic fungus infects Camponotus ants, which first search for abiotic conditions that are favorable for the fungus and then bite into a major leaf vein and remain there until their eventual death, allowing the fungus to form its fruiting body (Mongkolsamrit et al., 2012). Thus, taxonomically unrelated parasites can trigger the same biting behavior of an ant and its paralyzation in an atypical location (Libersat et al., 2009). Caterpillars that are infected with baculoviruses suffer from “Wipfelkrankheit” (“Wipfel” = German for “tree top,” “Krankheit” = German for “disease”): whereas healthy caterpillars at a certain developmental stage hide in the soil and pupate, infected individuals climb to the highest parts of their host tree where they die, liquefy and release the virions (Hofmann, 1891). Another example of “fatal attraction” is the seemingly suicidal behavior of crickets (Meconema thalassinum, Orthoptera) that carry hairworms (Spinochordodes tellinii, Nematomorpha): these animals seek water and jump into it, thereby allowing the adult hairworm to leave the dying insect, swim away and search for mating partners (Thomas et al., 2002; Biron et al., 2005; Libersat et al., 2009).
The most prominent “fatal attraction” is that of rats and other mammals to their feline predators, caused by the protozoon, T. gondii (Berdoy et al., 2000). Infected rats convert their innate fear of cat odors into attraction and exhibit enhanced general activity and decreased reaction times: all these behaviors enhance their risk of predation (Berdoy et al., 2000; Vyas et al., 2007; Flegr, 2013a). The parasite cannot achieve sexual reproduction in rodents and the described alterations increase the chances of transmission to its final, feline host. T. gondii can also infect other mammals (Poirotte et al., 2016) and is considered to be one of the most frequent infections in humans (Flegr, 2013b; Worth et al., 2013). Behavioral alterations that have been associated with latent toxoplasmosis include multiple forms of schizophrenia, personality disorder, Parkinson disease and Alzheimer disease, as well as increased aggressiveness, decreased willingness to follow social rules, and slower reaction times in men (Flegr, 2013a).
“Fatal attraction” can also be caused by human infectious diseases that are transmitted by blood-feeding insect vectors. Vector-borne disease agents can increase the frequency or duration of the contacts between vectors and hosts, suppress vector reproductive investment (an effect that is likely to increase nutrient reserves in the vector that are available for the parasite), or increase vector longevity (Hurd, 2003). For example, mosquitoes (Anopheles gambiae) that carry the malaria parasite, Plasmodium falciparum, feed longer and are more prone to bite several people per night than P. falciparum-free mosquitoes (Koella et al., 1998; Smallegange et al., 2013), whereas malaria infection in humans, mice and birds, enhances their attractiveness to the mosquitoes (Lacroix et al., 2005; Cornet et al., 2013; De Moraes et al., 2014).
With a few exceptions, such as the Caryophyllaceae, which are infected by the anther-smut fungus Microbotryum violaceum, plants as targets for manipulating parasites have only entered the literature recently. The fungus M. violaceum causes the abortion of ovaries, replaces the pollen with fungal spores, and uses insect pollinators for their transmission (Wennström et al., 2003; Antonovics, 2005; Sloan et al., 2008), whereas, quite remarkably, the rust fungus Puccinia monoica forms flower-imitating pseudoflowers on Arabis holboellii (Brassicaceae), which attract insects aiding the sexual reproduction of the fungus (Roy, 1993). Recently, many other manipulations have been reported for plant pathogens, including alterations in plant odor or in the quality and quantity of nutrients that move in the phloem. These alterations lead to a “fatal attraction” phenomenon in which plants become more attractive to their herbivores, or in which herbivores are “deceptively” attracted to host plants of inferior quality (Table 1).
Because plants are sessile, many pathogens of plants use the same strategies for their transmission as the agents of vector-borne human diseases (Guiguet et al., 2016) and, for example, change the behavior of their vectors to enhance transmission from infected to healthy plants hosts (Stafford et al., 2011; Ingwell et al., 2012; Rajabaskar et al., 2014), alter the emission of volatile organic compounds (VOCs) from the infected host (Mauck et al., 2010), or enhance the nutritional quality of the infected plant for herbivores that serve as their vectors (Fang et al., 2013; Luan et al., 2013; Shi et al., 2014). One of the most intriguing examples is Cucumber mosaic virus: the bouquet of VOCs emitted by infected plants is altered, making the plants more attractive to the aphids that vector this virus, even though the nutritional quality of infected plants is lower (Mauck et al., 2010). By contrast, other viruses enhance the nutritional quality of the infected plant for vectors such as the whitefly Bemisia tabaci (Fang et al., 2013; Luan et al., 2013; Shi et al., 2014).
In addition to the changes in the physiological parameters of their final host, several pathogens of plants also manipulate the behavior of their vector. For example, Tomato spotted wilt virus augments the frequency of all feeding behaviors in infected males of its thrips vector (Stafford et al., 2011), and Candidatus Liberibacter asiaticus makes the odor of infected citrus plants initially more attractive for its psyllid vector but then, after virus acquisition, causes psyllids to disperse to non-infected plants (Mann et al., 2012). Similarly, Tomato yellow leaf curl virus enhances the attractiveness of infected tomato plants to virus-free whiteflies (B. tabaci), whereas infected whiteflies lose the capacity to distinguish between infected and healthy host plants (Fang et al., 2013). Thus, at least some plant pathogens can manipulate both, their intermediate and their final host.
Host manipulation has been reported for species from all major taxa in which parasitism has evolved, and for hosts from the plant as well as the animal kingdom (Table 1). Nevertheless, the number of parasites for which no manipulation effect has ever been described far outweighs the number of reported cases, and most higher taxa of parasites for which manipulation has been described comprise both, manipulating and non-manipulating species (Lafferty and Kuris, 2012). Is this because no one has investigated the other cases, has host manipulation been overlooked, or is host manipulation really a specific trait of some parasites but not of others? Unfortunately, general patterns are difficult to identify. It is likely that the existing literature contains both false negative and false positive reports, and unbiased screenings of a broad number of parasite species are very difficult, due to the broad diversity of possible manipulation effects that would have to be considered. Negative reports are rare (but see, for example; Cator et al., 2012, 2013, 2015; Soh et al., 2013; Berret and Voordouw, 2015; Vantaux et al., 2015) and many cases of manipulating parasites might as yet remain undiscovered, either because they lead to inconspicuous or no alterations in the phenotype of the host, or because certain classes of parasites are seldom studied for host manipulation. By contrast, false positive reports can arise because alternative explanations for the observed phenotypic alterations are usually not excluded.
On the one hand, the number of overlooked cases is likely to be high, because successful manipulation can be inconspicuous. First, it can be the complete absence of any detectable alteration in the phenotype of a parasitized host that indicates its successful manipulation. Virtually all infected animals show some kind of sickness behavior (Hart, 1988), which usually includes a general decrease in mobility and social activities. In addition, most animals discriminate against parasitized individuals as potential mating partners (Vyas, 2013; Adamo, 2014). Both behavioral changes are under the genetic control of the host and can reduce the transmission of horizontally transmitted parasites. Therefore, the absence of sickness behavior or a lack of any discrimination against parasitized mates (Hari Dass et al., 2011; Adamo, 2014; Adamo et al., 2014) have been suggested as examples of successful host manipulation (Table 2). Second, there is clear evidence that men and women, or male and female rodents, behave differently when suffering from latent toxoplasmosis (Flegr, 2013b). Sex-specific or ontogenetic differences in the responses of hosts to certain parasites are likely to be common, and host manipulation is likely to be routinely overlooked in studies that compare parasitized and non-parasitized individuals at the population level without considering individual parameters such as sex, ontogenetic stage or nutritional status.
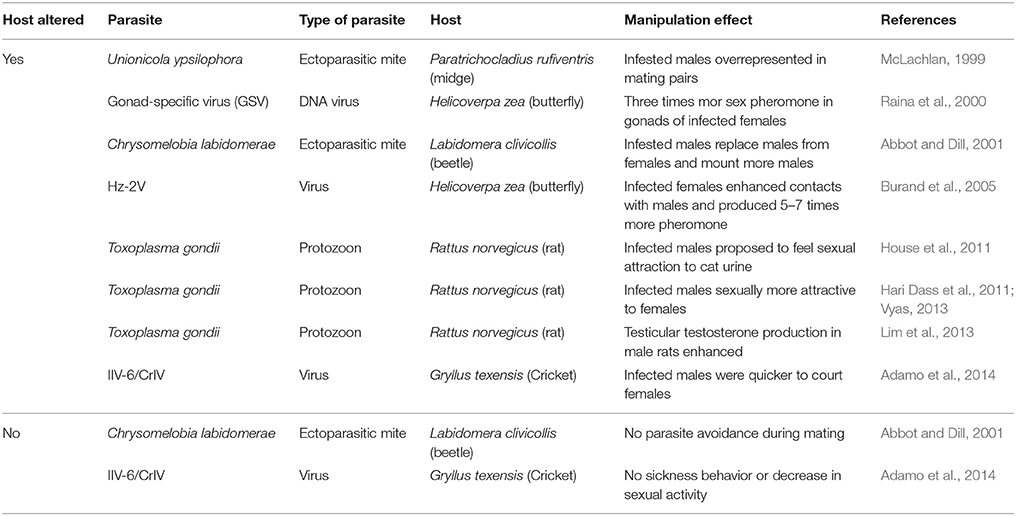
Table 2. Host manipulation by sexually transmitted parasites.
On the other hand, several authors have argued against the uncritical use of manipulation as the only explanation of alterations of host phenotypes that are elicited by parasites. For example, Poulin (2000) performed a meta-analysis and found a negative correlation of the estimated relative influence of parasites with the year of publication. As a possible explanation, he suggested publication bias, that is, an initial euphoria concerning the intensity of manipulation effects became replaced by more realistic estimates over time (Poulin, 2000). More recently, other authors have stressed that alternative explanations need to be considered—and excluded—to provide conclusive empirical support for an assumed case of manipulation (Klein, 2005; Thomas et al., 2005; James, 2010; Cator et al., 2012, 2015; Lafferty and Kuris, 2012; Worth et al., 2013). Alternatively, such alterations can: (1) represent mere side-effects of the pathology that is associated with infection; (2) form part of the successful immune response of the host to parasitization; or (3) represent traits that were inherited from ancestors but represent maladaptations in the present host–parasite combination (Klein, 2005; Thomas et al., 2005).
It seems obvious that the microbial agents of STDs and other sexually transmitted parasites would gain fitness benefits from enhancing their host's sexual promiscuity, frequency of mating behavior, or duration of the sexually active phase (Wennström et al., 2003). Nevertheless, “it is uncertain if STDs might increase mating frequency” in humans (Nesse and Foxman, 2011), and very few STDs have been reported to manipulate host behavior (Lafferty and Kuris, 2012; Berec and Maxin, 2014). A search conducted in Web of Science® Core Collection database on 10th of March 2016, produced 1000 hits when I searched for “host manipulation” AND “parasite” in “topic,” but only 66 hits when this search was combined with AND “STD* or sexual*,” and only few among these studies really reported on the manipulation of host sexual behavior by a sexually transmitted parasite (Table 2). For example, water snails infected with a trematode increased the number of mating events and the total number of different mating partners per individual (Soper et al., 2014). Males of milkweed leaf beetle, Labidomera clivicolli, or the midge, Paratrichocladius rufiventris, enhanced mating efforts and had higher mating success when they were parasitized by mites (McLachlan, 1999; Abbot and Dill, 2001). On top of that, the beetle males contacted non-parasitized males more often and for longer periods of time than mite-free males (Abbot and Dill, 2001). Parasitization can even enhance the sexual attractiveness of the host: female Helicoverpa zea moths infected with gonad-specific virus (GSV) or Hz-2V virus produced 3–7 times more sex pheromone than normal females (Raina et al., 2000; Burand et al., 2005), and T. gondii has been reported to enhance the attractiveness of male rats to females (Hari Dass et al., 2011)
Surprisingly, in several cases the reported manipulation effect consisted in the absence of alterations in the parasitized animal host. For example, no evidence was found for any discrimination during mating against parasitized L. clivicollis males (Abbot and Dill, 2001), and female H. zea butterflies as well as crickets (Gryllus texensis) of both sexes continued mating after infection with sexually transmitted viruses even though the virus rendered them essentially sterile (Burand et al., 2005; Adamo et al., 2014). Sexually transmitted parasites should enhance their fitness when they successfully suppress sickness behavior and those host phenotypic alterations that allow for a discrimination against parasitized individuals by potential mating partners. In the case of sexually or otherwise horizontally transmitted parasites, the absence of alterations in parasitized hosts can indicate a successful manipulation effect.
At the mechanistic level, reports on parasites that directly affect the neuronal system of their hosts or the levels of relevant hormones, such as testosterone, show that STDs, in principle, could easily manipulate the sexual behavior of their hosts. We only have to think of T. gondii, which enhances testicular testosterone production in male rats (Lim et al., 2013). In cats, infection with Feline Immunodeficiency Virus (FIV) and Feline Leukemia Virus (FeLV) has been correlated with enhanced plasma concentrations of sexual hormones, including testosterone (Tejerizo et al., 2012). Moreover, parasites with a horizontal mode of transmission typically enhance not only aggression, but also other forms of physical contact among the members of their host population (Klein, 2005). In summary, parasites can alter host sexual behavior, including the frequency of mating and the level of promiscuity, and potential molecular mechanisms for this type of manipulation have been described (Klein, 2003; Adamo, 2013, 2014). Nevertheless, it seems safe to conclude that STDs are underrepresented in the literature on manipulating parasites. Why are sexually transmitted parasites rarely reported to manipulate their hosts? I discuss four potential, non-exclusive explanations: first, sexual transmission per se could be less common than trophic transmission, at least among the parasites of non-human hosts; second, a sexual mode of transmission might not require manipulation; third, manipulation by sexually transmitted parasites might be particularly prone to being overlooked; or fourth, the lack of reports results from a research bias.
(1) Although STDs are common in humans, they might represent a comparably rare phenomenon among other species. For insects, Knell and Webberley (2004) list only 17 viral and no bacterial STDs at all. In plants, sexual reproduction is not determined by physical contact but rather depends on vectors, for which reason STDs in the sense of “diseases that are transmitted through physical contact during sexual activities” are not found in plants (Wennström et al., 2003), although the above-mentioned smut fungus demonstrates how plant pathogens can exploit the sexual phase of their host for transmission. By contrast, the human species alone is the primary host of several bacterial (Treponema pallidum ssp. pallidum, Neisseria gonorroeae, Chlamydia trachomatis Serovars D-K, among others) and viral (Human Immunodefficiency Virus [HIV], Human Papillome Virus [HPV], Hepatitis B Virus, Hepatitis C Virus, Herpes simplex genitalis, among others) STDs. Prerequisites for the evolution of sexual transmission are overlapping host generations and relatively long durations of the sexually active stage of the host. Most insects do not fulfill these prerequisites, because they occur as alternating larval and adult stages (Knell and Webberley, 2004). Similarly, annual plants and geophytes are likely to “escape” from sexually transmitted parasites, simply because they “disappear” from the ecosystem for extended parts of the year. In general terms, a host that exhibits only short phases of sexual activity with respect to its annual activity cycle or its overall life-time, is unfavorable for a sexual mode of disease transmission. By contrast, human reproductive behavior is characterized by overlapping generations and a prolonged adult phase with a year-round, desynchronized mating activity. Humans represent an exceptionally suitable host for the evolution of STDs indeed.
(2) The potential fitness benefit of manipulation increases when transmission represents a bottleneck for the parasite, that is, when transmission-relevant encounters among non-manipulated hosts represent a rare event (Poulin, 1995, 2010; Lafferty and Kuris, 2012). In the case of STDs “the parasite's and the host's genes have similar interests” (Flegr, 2013b) and “there is no threshold density for disease spread” (Lockhart et al., 1996). Therefore, sexually transmitted parasites might generally be under a low selective pressure to evolve manipulation, at least in terms of enhancing sexual activity over normal levels.
(3) The symptoms that characterize most viral and bacterial infections comprise fatigue and usually lead to a decrease in overall activity, including sexual activity. These symptoms are generally believed to represent a part of the host immune response. Therefore, a suppression of host immune responses might not only favor the infection process, but also aid horizontal transmission via a suppression of such secondary alterations in the social behavior of the host. How commonly are STDs reported to cause fatigue, decreased libido or sexual activity, or other symptoms that are typical for viral or bacterial infections? A search executed in the Web of Science® Core Collection database on 10th of March 2016 for reports on fatigue or sickness behavior in combination with human STDs revealed that STDs appear to be underrepresented in this literature. Whereas 0.52% (5956) of all 1,138,901 studies that I found in a search for “infection” and “human” in the field “topic” dealt with “fatigue OR sickness behavior,” the percentage of studies on “fatigue OR sickness behavior” among the studies on the major, above-mentioned STDs of humans ranged from 0.05 to 0.48%. Only few of these studies dealt with fatigue as a consequence of an STD, since terms like “treatment fatigue” or “safer sex fatigue” are abundant in the literature on HIV, whereas studies on fatigue as a consequence of vaccination are abundant in the literature on Hepatitis. It is tempting to speculate that the absence of fatigue in patients with STDs indicates a putative host manipulation by sexually transmitted parasites: an outcome of manipulation that is particularly prone to being overlooked.
(4) Finally, research bias might have contributed to the lack of reports on host manipulation by STDs, particularly when humans are overrepresented among their hosts. All early descriptions of manipulated hosts referred to trophically transmitted parasites of animals, owing to the conspicuous phenotypes that they cause. Given that studies in search of confirmation of a newly described phenomenon tend to focus on similar cases, it seems likely that little effort has been made to search for manipulating STDs of humans. As stated by Flegr (2013a), “a study of the manipulation hypothesis remains…absolutely out of the scope of interest of physicians.”
In spite of potential research biases and the likelihood of both, false positives and false negatives among the existing reports, it seems to be a real phenomenon that manipulating and non-manipulating parasites co-exist, sometimes within the same taxonomic group. Why do some parasites manipulate their hosts whereas others do not? All parasites express traits for the recognition and the invasion or occupation of a host, as well as for the manipulation or avoidance of host immunity, and traits that favor the transmission of parasites should be under an equally strong selective pressure.
If we aim to understand the circumstances under which host manipulation is likely to evolve, we must consider the benefits and costs of manipulation for the parasite. In general, parasites can gain higher selective benefits from manipulation when the frequency of transmission-relevant encounters among non-manipulated consecutive hosts is low (Poulin, 1994). In a broader sense, parasites are more likely to evolve manipulation when they do not move along the dominating pathways within the food chain, because they must shift, for example, among hosts with a diurnal vs. nocturnal activity or aquatic vs. terrestrial hosts, or from an animal intermediate host to an herbivorous final host.
A second crucial factor that determines the potential benefits of host manipulation is persistence time. The longer a parasite can persist in the respective host without suffering from any reduction in its fitness, the lower is the pressure to move to the next host (Poulin, 1994). Indeed, differing persistence times might be a major explanation for the observation that manipulation effects are more frequently reported for intermediate than for final hosts, at least as long as all hosts are animals (Holmes and Bethel, 1972). Plant pathogens, by contrast, require mobile vectors for their dispersal among their immobile final hosts and, therefore, frequently manipulate the quality of their host plant for the vectors (Belliure et al., 2010; Mauck et al., 2010, 2014a,b; Luan et al., 2013; Shi et al., 2013) as well as the feeding decisions that are taken by their vectors (Stafford et al., 2011; Ingwell et al., 2012; Mann et al., 2012; Fang et al., 2013; Rajabaskar et al., 2014). These changes can be very fine-tuned: for example, plant viruses with a persistent mode of transmission require vectors to feed for a prolonged period of time on infected hosts and, thus, usually tend to improve the quality of the host plants for the vectors. By contrast, non-persistently transmitted viruses are effectively transmitted when vectors briefly probe infected hosts and, therefore, these viruses usually tend to reduce host plant quality for vectors (Mauck et al., 2012).
In summary, the putative net benefits of host manipulation vary among different host–parasite systems. Nevertheless, manipulation should be expected to be almost ubiquitous if there were no costs that select against its evolution. Almost no empirical studies have been performed to search for such costs (Poulin, 2010); nevertheless, theoretical considerations can help to identify putative sources of costs. First, any manipulation depends on at least some kind of physical or molecular trigger that is released by the parasite. The production of these molecules would have a metabolic cost, the order of magnitude of which depends on the consumption of energy and limiting elemental resources that are required for its production or maintenance, relative to the overall metabolic demands of the parasite. It might be no coincidence that T. gondii triggers the synthesis of signaling compounds in its host, rather than producing these compounds itself (Gaskell et al., 2009; Prandovszky et al., 2011; Lim et al., 2013). Viruses depend completely on the metabolism of their host and, thus, should gain most when they manipulate the re-allocation of host compounds, or the activity of existing signaling molecules, rather than inducing costly de-novo synthesis (Hoover et al., 2011). Second, DNA itself has a metabolic cost, and viruses and intracellular bacteria in particular are under high selective pressure to reduce their genome sizes to a minimum. Therefore, the mere need to carry additional genes that encode for the manipulating trait can represent a significant cost for a parasite. Third, scaling up to the ecological level, altered host phenotypes can trigger responses in coexisting species that share the host with the parasite, or with its vector. These responses can feedback to the parasite: for example, by enhancing the competition among host and non-host predators for an infected intermediate host vector or among non-vectors for an infected final host, by depleting limiting resources within the host, or by altering the survival rates of the manipulated host in its environment. What are the net consequences for a specific parasite of an enhanced predation pressure on its intermediate host when final hosts are rare among all the predators that are currently around? (Mouritsen and Poulin, 2003) What are the consequences for plant viruses when a virus-induced enhancement in phloem quality and its “public advertisement” via odors are exploited by non-vector insects that have similar nutritional requirements to those of the vectors? (Belliure et al., 2010; Mauck et al., 2014b) Parasites might reduce these costs by limiting the manipulation of their intermediate host to the developmental phase in which they are infectious to their definitive host (Dianne et al., 2011) but still, the net success of this strategy depends on the ratio of hosts to non-hosts in the current environment. The hijacking of manipulation effects by other species (Mouritsen and Poulin, 2003) can potentially have strong negative feedback effects on the parasite.
Finally, a counter-selective force that remains to be included in the research into host manipulation is the manipulated host itself. Because manipulation usually involves a high cost for the host, it can be expected to be under a strong selective pressure to evolve resistance to manipulation. The extensive literature on the “evolutionary arms-race” between plants and their herbivores or pathogens provides us with multiple examples of the multiple possible outcomes of the “zigzag” between plant enemies evolving traits for successful attack, plants responding with the evolution of traits that allow for enemy recognition and the mounting of adequate resistance responses, and enemies responding with counter-resistance strategies (Fraenkel, 1959; Dangl and Jones, 2001; Jones and Dangl, 2006). In other words, the “lack of virulence” of a specific pathogen could either mean that the pathogen has not evolved to infect the plant species in question, or that the plant has mounted a successful resistance-strategy (Barrett and Heil, 2012). The same evolutionary forces that rule host immunity to infection are likely to act in the manipulative interactions as well. Thus, an absence of host manipulation might be a lack of the respective strategy on the side of the parasite, but it could also indicate that the host has successfully evolved to overcome the manipulation effect.
In the above paragraphs I discussed some of the circumstances that might select for, or against, host manipulation. Life-history traits that seem to be shared by almost all known manipulating parasites are the need to switch among host species and a trophic mode of transmission. Nevertheless, gaining a deeper understanding of the evolution of host manipulation crucially depends on an unambiguous identification of manipulating vs. non-manipulating parasites and, therefore, on the stringent differentiation of manipulation effects vs. induced host responses to parasitization.
Plant pathogens provide us with intriguing examples of the multiple alternative explanations that can be provided for the same phenomenon. Many of these pathogens increase the concentration of free amino acids in the phloem and enhance the attractiveness of the plant odors that are emitted to for herbivores that function as their vectors (Mann et al., 2012; Wang et al., 2012; Mauck et al., 2014a). Because most arthropods use olfactory cues for their long-distance orientation and phloem amino acids represent a limiting factor for sapsuckers, both alterations can increase the attraction of vectors to on an infected plant and benefit the transmission of the pathogen. However, the physiological mechanisms that control the described phenotypic alterations remain to be identified. In particular, we lack an explanation for a crucial feature of the successful manipulation strategy: the reliability of changes in the odor profile of a plant as indicators of its increased nutritional quality. Alternative explanations for an increase in the amino acid content in the phloem of an infected plant include: (i) a mobilization of protein-bound amino acids to rescue them from the infected tissue; (ii) the allocation of amino acids from other parts of the plant to the infected tissue to enable the local synthesis of pathogenesis-related proteins and other N-containing compounds; (iii) a sink in the infected leaves that is created by an increased need for amino acids owing to the synthesis of viral proteins; or (iv) a manipulation that the pathogen has evolved to attract its vector. The first two mechanisms would be under the control of the plant, the other two mechanisms would be under the control of the virus; however, only the last mechanism would represent a classical host manipulation that has been selected to enhance transmission of the pathogen.
Similarly, enhanced locomotion (e.g., foraging or hunting activity) of an infected animal can enhance its exposure to the predators that represent the final hosts of the parasite, but enhanced locomotion might primarily serve to fulfill the enhanced energy demands that result from the adaptive immune response in the host. The observation that Plasmodium-infected mosquitoes feed longer and on more people per night (Koella et al., 1998) could simply mean that carrying Plasmodium has a metabolic cost for which the mosquito must compensate, rather than representing a manipulation effect. This interpretation is strongly supported by the finding that similar behavioral alterations were observed in mosquitoes that had been challenged with heat-killed E. coli (Cator et al., 2013). The consecutive discovery of functional links between the behavioral alterations and altered insulin signaling in the gut of the mosquito further supports the assumption that it is the mosquito, rather than the Plasmodium, that has genetic control over the altered feeding behavior (Cator et al., 2015).
Moreover, all parasites must suppress parts of the immune system of their host, and parasites of mammals might do so by altering the levels of testosterone or of certain cytokines. For example, the causal agent of syphilis, T. pallidum, enhances the concentrations of cytokines, including several interferons, and thereby evades recognition by the immune system (Cruz et al., 2012). Specific interferons are also thought to contribute to the survival of Chlamydia in human hosts (Duell et al., 2012). Even more strikingly, T. gondii and FIV enhance the levels of testosterone in their hosts (Tejerizo et al., 2012; Flegr and Markos, 2014), a hormone that is well known for its immunosuppressive effects (Schroderus et al., 2010). Besides favoring infection in the initial phase, these molecular alterations could also suppress sickness behavior, maintain high sexual activity and enhance general aggressive behavior in males, or avoid the rejection of parasitized partners during mating: all these mechanisms would represent manipulation for infection rather than transmission, but they nevertheless can also enhance transmission rates, at least under specific environmental conditions (Ashley et al., 2009; Adamo, 2014). The equivalent of such an effect in plants could result from crosstalk among the different plant defensive signaling cascades. Many plants can either respond to chewing herbivores via the induction of genes that depend on jasmonic acid signaling, or mount resistance to infection with biotrophic pathogens via salicylic acid-dependent pathways, but not both (Thaler et al., 2012). Because the net quality of a plant as a host for an herbivore or pathogen depends on both, its content of primary nutrients and its spectrum of defensive traits (Barrett and Heil, 2012), the increased quality for herbivores that we observe in many diseased plants could be a side-effect of negative crosstalk between different defense systems of the plant, rather than the direct effect of a targeted manipulation by the parasite.
Alternative explanations have been presented for even some of the seemingly most obvious cases of host manipulation. For example, the enhanced numbers of mating events and of different mating partners reported for a water snail infected with horizontally transmitted trematodes could be the result of a successful manipulation by a sexually transmitted parasite; however, it might also represent the outcome of an adaptation of the host, because it leads to enhanced genetic variation among the offspring and, thus, enhanced resistance to the locally adapted parasite (Soper et al., 2014). Similarly, several different mechanisms could cause the frequently reported association of elevated levels of testosterone in males with latent toxoplasmosis (Flegr et al., 2008; Shirbazou et al., 2011). Rather than indicating an alteration that is elicited by T. gondii, high levels of testosterone could be a causal reason for infection and represent an intrinsic trait of certain individuals that: (i) causes more risk-taking behavior and thereby leads to higher infection risk, or (ii) simply provides the parasite with a “preferred endocrine environment in the host” (James, 2010). Even when T. gondii can unambiguously be shown to actively enhance the levels of testosterone in its host it remains an open question whether this capacity has primarily evolved for the manipulation of host immunity, in which case it would mainly serve infection, rather than transmission.
Different scenarios can be formulated as explanations for the same alteration in a parasitized host. This situation illustrates the need to identify the proximate molecular mechanisms that control these alterations and their ultimate effects on the fitness of all involved partners. Unfortunately, the mechanisms and the net fitness effects remain to be investigated for virtually all described cases of host manipulation. Multiple parasites of insects have been speculated to secrete chemical compounds for host manipulation (Libersat et al., 2009), T. gondii secretes peptides that interact in multiple ways with the host immune system (Leroux et al., 2015), and even the release of small RNA-containing vesicles has been reported for the filarial parasite Brugia malayi (Zamanian et al., 2015). However, the detailed nature of these molecules and their modes of action in the host remain to be identified.
Surprisingly, we do not even understand the physiological mechanisms that cause the association between toxoplasmosis and schizophrenia, or other behavioral alterations in T. gondii-infected humans (Flegr, 2013a; Parlog et al., 2015). Recent studies provide convincing mechanisms for at least some of the host phenotypic changes that are elicited by this specific parasite. T. gondii alters the levels of two behaviorally important molecules in the mammalian host: the neurotransmitter, dopamine, and the steroid hormone, testosterone (Flegr and Markos, 2014; Parlog et al., 2015). Interestingly, the genome of T. gondii contains two genes for tyrosine hydroxylase, which catalyzes the rate-limiting step in the synthesis of dopamine in dopaminergic neural cells (Gaskell et al., 2009). T. gondii cysts in the brain of an infected host produce large amounts of dopamine and the neurotransmitter is exported into the surrounding tissue (Prandovszky et al., 2011). The commonly observed increase in testosterone levels in infected male rats involves an increase in the expression of those receptors in the testes that regulate the synthesis of testosterone (Lim et al., 2013). This finding might explain why enhanced levels of testosterone have been observed only in infected men (Flegr et al., 2008; Shirbazou et al., 2011), and for the multiple behavioral and psychological traits in which men and women with latent toxoplasmosis exhibit opposite trends (Flegr et al., 2011; Flegr, 2013b). Recently, T. gondii has been reported to hypomethylate the arginine vasopressin promoters in the medial amygdala of rats (Hari Dass and Vyas, 2014), an epigenetic manipulation that could cause a stronger activation of vasopressinergic neurons after exposure to cat odor (Flegr and Markos, 2014). However, it remains an open question whether and to what degree these findings are also applicable to the brain of infected humans and, more importantly, the relevance of these effects in the interaction of the natural intermediate hosts with the natural final hosts of T. gondii has yet to be determined. Does, for example, the changed response of infected men to cat odor (Flegr et al., 2011) mean that the parasite elicits the correct behavior in the wrong host, or does it mean that humans and their feline predators were important hosts during the evolution of T. gondii? The last interpretation gains some support from the observation that Toxoplasma-infected chimpanzees exhibit fatal attraction to leopard urine (Poirotte et al., 2016).
A case in which domestication and other human influences are unlikely to affect the current situation is the “Wipfelkrankheit.” In elegant experiments using transformed viral strains, Hoover et al. (2011) demonstrated that a single viral gene is responsible for the seemingly “Zombie”-like behavior of caterpillars that climb to the highest points of a tree instead of going down into the soil to pupate. The viral gene ecdysteroid-uridine diphosphate-glucosyltransferase encodes an enzyme that inactivates the caterpillar's molting hormone by transferring a sugar moiety to it. Low levels of this hormone suppress molting and reduce caterpillar morbidity, thereby allowing infected individuals to continue feeding for longer time spans compared with uninfected caterpillars (Hoover et al., 2011). Recently, it has been shown that baculoviruses enhance the positive phototactic behavior in the infected caterpillars (van Houte et al., 2014), an alteration that further contributes to the phenotype of tree-top disease. In summary, it seems that parasites tend to upregulate the synthesis of endogenous signaling molecules in the host, activate or inactivate existing host-derived molecules, or change the sensitivity of existing receptors to host-derived molecules, rather than producing such compounds themselves and delivering them to the host (Adamo, 2013).
Further research efforts are required to understand the ecological, evolutionary and medical relevance of parasites that manipulate their hosts, and this research will require criteria that define how manipulated phenotypes can be identified and distinguished for example, from adaptive host responses to parasitization. I agree with (Poulin, 1995) in that particularly complex phenotypic changes can represent good indicators of a manipulation. However, challenging Anopheles stephensi females with heat-inactivated E. coli enhanced their feeding activities during the same time window in which Plasmodium would be in its infective stage (Cator et al., 2015), an observation that demonstrates how closely side effects of host immune responses can resemble the outcome of a putative host manipulation. Therefore, studies aimed at understanding the relevance of host manipulation by parasites require a clearly defined Null-hypothesis, and they should identify the molecular mechanisms that underlie the observed phenotypic change, consider alternative explanations, and quantify—under ecologically realistic conditions—the effects of the observed change on the fitness of the parasite and that of the host.
First, enhanced parasite fitness is a necessary, but not a sufficient, argument for the consideration of a certain phenotypic alteration as a manipulation effect. Hosts respond to parasitization with multiple phenotypic changes that serve to enhance resistance or tolerance and to decrease transmission rates. As many of these responses are unspecific and target large classes of parasites, they might coincidentally favor the transmission of one or a few specific parasites (Levri, 1999). These changes are under the genetic control of the host and, thus, cannot be considered a manipulation by the parasite in question. Moreover, these changes would not be counter-selected as long as the selection pressure exerted by the parasite in question is lower than the selection pressure by the overall group of parasites against which the response leads to enhanced host resistance. Phenotypically plastic hosts will always respond to parasitization, which makes it crucial to define a clear Null-hypothesis: which induced phenotypic responses to parasitization can be expected in a non-manipulated host? As mentioned above, a successful manipulation can even be indicated by the absence of any alteration when the induced host response would have diminished parasite transmission rates. Second, the adaptive host manipulation hypothesis clearly defines the observed phenotypic alterations as being under the genetic control of the parasite. Thus, the unambiguous identification of a manipulation event requires an understanding of the physiological and molecular mechanisms that cause these alterations. In other words, the observation of a manipulation effect does not suffice to characterize the manipulator as a parasite (Heil, 2015), because manipulation effects can also be exerted by mutualists (Wright et al., 2013; Heil et al., 2014; Hojo et al., 2015). Only the demonstration that a certain phenotypic alteration in the host is under the genetic control of the parasite and that it enhances the fitness of the parasite vi an enhancement of its transmission rate, while decreasing the fitness of the host, would provide clear evidence in favor of adaptive host manipulation.
Finally, the net outcome of a presumed manipulation depends crucially on the natural rates of encounters among consecutive hosts and on the probability of successful transmission during each of these events. Hence, the net fitness effects of a presumed manipulation should be quantified at natural densities of parasites, hosts, vectors, and other species that naturally co-occur with the target species in question. Making the host more suitable for competing parasites, attracting non-vectors to an infested plant that outcompete the vector, or increasing predation rates in the absence of the predator who serves as the definitive host, are all scenarios can select against the evolution of a manipulation effect. In this context, a particularly elegant approach used naturally co-existing and taxonomically related species of intermediate hosts, among which only one host can reasonably be assumed to have co-evolved with the parasite: increased transmission rates after host phenotypic alterations were observed for the native, but not the invasive intermediate host of a native parasite, an observation that represents a strong argument in favor of an adaptive manipulation (Lagrue et al., 2007). In general, a promising strategy to identify manipulators as well as the underlying mechanisms would compare the interactions of parasites that have undergone a recent host shift to the interactions among their ancestors and the original host.
Adaptive host manipulation by parasites is a fascinating phenomenon that is likely to have as-yet unknown ecological, evolutionary and medical effects. Host manipulation has been described for all major taxonomic groups of parasites, and the resulting alterations comprise some of the most spectacular examples of extended phenotypes. Although the biases in the literature make generalizations difficult, I conclude that the majority of manipulating parasites use a trophic mode of transmission and that parasites of animals frequently manipulate their intermediate hosts (i.e., vectors), whereas many plant pathogens manipulate their vectors and their (immobile) final host. I also conclude that the “silent” infection mode of most STDs might indicate an as-yet overlooked host manipulation by STDs.
More studies are required to decipher the molecular mechanisms that control host manipulation, to estimate the frequency of this phenomenon in taxonomic and ecological terms and, ultimately, to understand the evolution of host manipulation as well as its ecological (or medical) relevance for the species involved. Future research would benefit from using clearly defined criteria for the identification of manipulated phenotypes, from the integration of the putative effects of host immune responses into studies on manipulating parasites, and from cross-disciplinary approaches that apply ecological, physiological and molecular techniques to comparative studies on model systems from a diverse range of hosts and parasites.
The author confirms being the sole contributor of this work and approved it for publication.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
I thank Caroline Woods for numerous helpful and constructive comments on an earlier version of the manuscript. Financial support by Consejo Nacional de Sciencia y Técnología de México (CONACyT grant 212715) is gratefully acknowledged.
Abbot, P., and Dill, L. M. (2001). Sexually transmitted parasites and sexual selection in the milkweed leaf beetle, Labidomera clivicollis. Oikos 92, 91–100. doi: 10.1034/j.1600-0706.2001.920111.x
Adamo, S. A. (2013). Parasites: evolution's neurobiologists. J. Exp. Biol. 216, 3–10. doi: 10.1242/jeb.073601
Adamo, S. A. (2014). Parasitic aphrodisiacs: manipulation of the hosts' behavioral defenses by sexually transmitted parasites. Integrat. Compar. Biol. 54, 159–165. doi: 10.1093/icb/icu036
Adamo, S. A., Kovalko, I., Easy, R. H., and Stoltz, D. (2014). A viral aphrodisiac in the cricket Gryllus texensis. J. Exp. Biol. 217, 1970–1976. doi: 10.1242/jeb.103408
Antonovics, J. (2005). Plant venereal diseases: insights from a messy metaphor. New Phytol. 165, 71–80. doi: 10.1111/j.1469-8137.2004.01215.x
Ashley, N. T., Hays, Q. R., Bentley, G. E., and Wingfield, J. C. (2009). Testosterone treatment diminishes sickness behavior in male songbirds. Hormones Behav. 56, 169–176. doi: 10.1016/j.yhbeh.2009.04.003
Barrett, L. G., and Heil, M. (2012). Unifying concepts and mechanisms in the specificity of plant-enemy interactions. Trends Plant Sci. 17, 282–292. doi: 10.1016/j.tplants.2012.02.009
Belliure, B., Sabelis, M. W., and Janssen, A. (2010). Vector and virus induce plant responses that benefit a non-vector herbivore Basic Appl. Ecol. 11, 162–169. doi: 10.1016/j.baae.2009.09.004
Berdoy, M., Webster, J. P., and Macdonald, D. W. (2000). Fatal attraction in rats infected with Toxoplasma gondii. Proc. Roy. Soc. B 267, 1591–1594. doi: 10.1098/rspb.2000.1182
Berec, L., and Maxin, D. (2014). Why have parasites promoting mating success been observed so rarely? J. Theor. Biol. 342, 47–61. doi: 10.1016/j.jtbi.2013.10.012
Berret, J., and Voordouw, M. J. (2015). Lyme disease bacterium does not affect attraction to rodent odour in the tick vector. Parasites Vectors 8, 249. doi: 10.1186/s13071-015-0856-8
Biron, D. G., Marché, L., Ponton, F., Loxdale, H. D., Galéotti, N., Renault, L., et al. (2005). Behavioural manipulation in a grasshopper harbouring hairworm: a proteomics approach. Proc. Roy. Soc. B Biol. Sci. 272, 2117–2126. doi: 10.1098/rspb.2005.3213
Burand, J. P., Tan, W., Kim, W., Nojima, S., and Roelofs, W. (2005). Infection with the insect virus Hz-2v alters mating behavior and pheromone production in female Helicoverpa zea moths. J. Insect Sci. 5, 5. doi: 10.1673/031.005.0601
Cator, L. J., George, J., Blanford, S., Murdock, C. C., Baker, T. C., Read, A. F., et al. (2013). ‘Manipulation’ without the parasite: altered feeding behaviour of mosquitoes is not dependent on infection with malaria parasites. Proc. Roy. Soc. B 280:20130711. doi: 10.1098/rspb.2013.0711
Cator, L. J., Lynch, P. A., Read, A. F., and Thomas, M. B. (2012). Do malaria parasites manipulate mosquitoes? Trends Parasitol. 28, 466–470. doi: 10.1016/j.pt.2012.08.004
Cator, L. J., Pietri, J. E., Murdock, C. C., Ohm, J. R., Lewis, E. E., Read, A. F., et al. (2015). Immune response and insulin signalling alter mosquito feeding behaviour to enhance malaria transmission potential. Sci. Rep. 5:11947. doi: 10.1038/srep11947
Cornet, S., Nicot, A., Rivero, A., and Gandon, S. (2013). Malaria infection increases bird attractiveness to uninfected mosquitoes. Ecol. Lett. 16, 323–329. doi: 10.1111/ele.12041
Cruz, A. R., Ramirez, L. G., Zuluaga, A. V., Pillay, A., Abreu, C., Valencia, C. A., et al. (2012). Immune evasion and recognition of the Syphilis spirochete in blood and skin of secondary Syphilis patients: two immunologically distinct compartments. PLoS Negl. Trop. Dis. 6:e1717. doi: 10.1371/journal.pntd.0001717
Dangl, J. L., Horvath, D. M., and Staskawicz, B. J. (2013). Pivoting the plant immune system from dissection to deployment. Science 341, 746–751. doi: 10.1126/science.1236011
Dangl, J. L., and Jones, J. D. G. (2001). Plant pathogens and integrated defence responses to infection. Nature 411, 826–833. doi: 10.1038/35081161
De Moraes, C. M., Stanczyk, N. M., Betz, H. S., Pulido, H., Sim, D. G., Read, A. F., et al. (2014). Malaria-induced changes in host odors enhance mosquito attraction. Proc. Natl. Acad. Sci. U.S.A. 111, 11079–11084. doi: 10.1073/pnas.1405617111
Dianne, L., Perrot-Minnot, M. J., Bauer, A., Gaillard, M., Léger, E., and Rigaud, T. (2011). Protection first then facilitation: a manipulative parasite modulates the vulnerability to predation of its intermediate host according to its own developmental stage. Evolution 65, 2692–2698. doi: 10.1111/j.1558-5646.2011.01330.x
Duell, B. L., Tan, C. K., Carey, A. J., Wu, F., Cripps, A. W., and Ulett, G. C. (2012). Recent insights into microbial triggers of interleukin-10 production in the host and the impact on infectious disease pathogenesis. FEMS Immunol. Med. Microbiol. 64, 295–313. doi: 10.1111/j.1574-695X.2012.00931.x
Fang, Y., Jiao, X., Xie, W., Wang, S., Wu, Q., Shi, X., et al. (2013). Tomato yellow leaf curl virus alters the host preferences of its vector Bemisia tabaci. Sci. Rep. 3:2876. doi: 10.1038/srep02876
Flegr, J. (2013a). How and why Toxoplasma makes us crazy. Trends Parasitol. 29, 156–163. doi: 10.1016/j.pt.2013.01.007
Flegr, J. (2013b). Influence of latent Toxoplasma infection on human personality, physiology and morphology: pros and cons of the Toxoplasma–human model in studying the manipulation hypothesis. J. Exp. Biol. 216, 127–133. doi: 10.1242/jeb.073635
Flegr, J., Lenochová, P., Hodný, Z., and Vondrová, M. (2011). Fatal attraction phenomenon in humans - cat odour attractiveness increased for Toxoplasma-infected men while decreased for infected women. PLoS Neglect. Trop. Dis. 5:e1389. doi: 10.1371/journal.pntd.0001389
Flegr, J., Lindová, J., and Kodym, P. (2008). Sex-dependent toxoplasmosis-associated differences in testosterone concentration in humans. Parasitology 135, 427–431. doi: 10.1017/S0031182007004064
Flegr, J., and Markos, A. (2014). Masterpiece of epigenetic engineering - how Toxoplasma gondii reprogrammes host brains to change fear to sexual attraction. Mol. Ecol. 23, 5934–5936. doi: 10.1111/mec.13006
Fraenkel, G. S. (1959). The raison d'Être of secondary plant compounds. Science 129, 1466–1470. doi: 10.1126/science.129.3361.1466
Gaskell, E. A., Smith, J. E., Pinney, J. W., Westhead, D. R., and McConkey, G. A. (2009). A unique dual activity amino acid hydroxylase in Toxoplasma gondii. PLoS ONE 4:e4801. doi: 10.1371/journal.pone.0004801
Guiguet, A., Dubreuil, G., Harris, M. O., Appel, H. M., Schultz, J. C., Pereira, M. H., et al. (2016). Shared weapons of blood- and plant-feeding insects: surprising commonalities for manipulating hosts. J. Insect Physiol. 84, 4–21. doi: 10.1016/j.jinsphys.2015.12.006
Hari Dass, S. A., Vasudevan, A., Dutta, D., Soh, L. J. T., Sapolsky, R. M., and Vyas, A. (2011). Protozoan parasite Toxoplasma gondii manipulates mate choice in rats by enhancing attractiveness of males. PLoS ONE 6:e27229. doi: 10.1371/journal.pone.0027229
Hari Dass, S. A., and Vyas, A. (2014). Toxoplasma gondii infection reduces predator aversion in rats through epigenetic modulation in the host medial amygdala. Mol. Ecol. 23, 6114–6122. doi: 10.1111/mec.12888
Hart, B. L. (1988). Biological basis of the behavior of sick animals. Neurosci. Biobehav. Rev. 12, 123–137. doi: 10.1016/S0149-7634(88)80004-6
Heil, M. (2015). Manipulators live better, but are they always parasites? Trends Plant Sci. 20, 538–540. doi: 10.1016/j.tplants.2015.08.001
Heil, M., and Baldwin, I. T. (2002). Fitness costs of induced resistance: emerging experimental support for a slippery concept. Trends Plant Sci. 7, 61–67. doi: 10.1016/S1360-1385(01)02186-0
Heil, M., Barajas-Barron, A., Orona-Tamayo, D., Wielsch, N., and Svatos, A. (2014). Partner manipulation stabilises a horizontally transmitted mutualism. Ecol. Lett. 17, 185–192. doi: 10.1111/ele.12215
Hofmann, O. (1891). Insektentötende Pilze mit Besonderer Berücksichtigung der Nonne. Frankfurt: P. Weber Verlag.
Hojo, M. K., Pierce, N. E., and Tsuji, K. (2015). Lycaenid caterpillar secretions manipulate attendant ant behavior. Curr. Biol. 25, 2260–2264. doi: 10.1016/j.cub.2015.07.016
Holmes, J. C., and Bethel, W. M. (1972). “Modification of intermediate host behavior by parasites,” in Behavioral Aspects of Parasite Transmission, eds E. U. Canning and C. A. Wright (London: Academic Press), 123–149.
Hoover, K., Grove, M., Gardner, M., Hughes, D. P., McNeil, J., and Slavicek, J. (2011). A gene for an extended phenotype. Science 333, 1401–1401. doi: 10.1126/science.1209199
House, P. K., Vyas, A., and Sapolsky, R. (2011). Predator cat odors activate sexual arousal pathways in brains of Toxoplasma gondii infected rats. PLoS ONE 6:e23277. doi: 10.1371/journal.pone.0023277
Hughes, D. P. (2014). On the origins of parasite-extended phenotypes. Integrat. Compar. Biol. 54, 210–217. doi: 10.1093/icb/icu079
Hurd, H. (2003). Manipulation of medically important insect vectors by their parasites. Annu. Rev. Entomol. 48, 141–161. doi: 10.1146/annurev.ento.48.091801.112722
Ingwell, L. L., Eigenbrode, S. D., and Bosque-Pérez, N. A. (2012). Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2:e578. doi: 10.1038/srep00578
James, W. H. (2010). Potential solutions to problems posed by the offspring ratios of people with parasitic and viral infections. Folia Parasitol. 57, 114–120. doi: 10.14411/fp.2010.014
Janeway, C. A., Travers, P., Walport, M., and Shlomchik, M. J. (2001). Immunobiology the Immune System in Health and Disease. New York, NY: Garland Science.
Jones, J. D. G., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Kagan, I. G. (1951). Aspects in the life history of Neoleucochloridium problematicum (Magath, 1920) New Comb. and Leucochloridium cyanocittae McIntosh, 1932 (Trematoda:Brachylaemidae). Transact. Am. Microscop. Soc. 70, 281–318. doi: 10.2307/3223567
Karban, R., and Baldwin, I. T. (1997). Induced Responses to Herbivory. Chicago, IL; London: University of Chicago Press.
Klein, S. L. (2003). Parasite manipulation of the proximate mechanisms that mediate social behavior in vertebrates. Physiol. Behav. 79, 441–449. doi: 10.1016/S0031-9384(03)00163-X
Klein, S. L. (2005). Parasite manipulation of host behavior: mechanisms, ecology, and future directions. Behav. Proc. 68, 219–221. doi: 10.1016/j.beproc.2004.07.009
Knell, R. J., and Webberley, K. M. (2004). Sexually transmitted diseases of insects: distribution, evolution, ecology and host behaviour. Biol. Rev. 79, 557–581. doi: 10.1017/S1464793103006365
Koella, J. C., Sørensen, F. L., and Anderson, R. A. (1998). The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc. Roy. Soc. Lond. B 265, 763–768. doi: 10.1098/rspb.1998.0358
Lacroix, R., Mukabana, W. R., Gouagna, L. C., and Koella, J. C. (2005). Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol. 3:e298. doi: 10.1371/journal.pbio.0030298
Lafferty, K. D., and Kuris, A. M. (2012). “Eological consequences of manipulative parasites,” in Host Manipulation by PARASITES, eds D. P. Hughes, J. Brodeur, and F. Thomas (Oxford: Oxford University Press), 158–168.
Lagrue, C., Kaldonski, N., Perrot-Minnot, M. J., Motreuil, B., and Bollache, L. (2007). Modification of hosts' behavior by a parasite: field evidence for adaptive manipulation. Ecology 88, 2839–2847. doi: 10.1890/06-2105.1
Leroux, L.-P., Dasanayake, D., Rommereim, L. M., Fox, B. A., Bzik, D. J., Jardim, A., et al. (2015). Secreted Toxoplasma gondii molecules interfere with expression of MHC-II in interferon gamma-activated macrophages. Int. J. Parasitol. 45, 319–332. doi: 10.1016/j.ijpara.2015.01.003
Levri, E. P. (1999). Parasite-induced change in host behavior of a freshwater snail: parasitic manipulation or byproduct of infection? Behav. Ecol. 10, 234–241. doi: 10.1093/beheco/10.3.234
Libersat, F., Delago, A., and Gal, R. (2009). Manipulation of host behavior by parasitic insects and insect parasites. Annu. Rev. Entomol. 54, 189–207. doi: 10.1146/annurev.ento.54.110807.090556
Lim, A., Kumar, V., Hari Dass, S. A., and Vyas, A. (2013). Toxoplasma gondii infection enhances testicular steroidogenesis in rats. Mol. Ecol. 22, 102–110. doi: 10.1111/mec.12042
Lockhart, A. B., Thrall, P. H., and Antonovics, J. (1996). Sexually transmitted diseases in animals: ecological and evolutionary implications. Biol. Rev. 71, 415–471. doi: 10.1111/j.1469-185X.1996.tb01281.x
Luan, J.-B., Yao, D.-M., Zhang, T., Walling, L. L., Yang, M., Wang, Y.-J., et al. (2013). Suppression of terpenoid synthesis in plants by a virus promotes its mutualism with vectors. Ecol. Lett. 16, 390–398. doi: 10.1111/ele.12055
Mann, R. S., Ali, J. G., Hermann, S. L., Tiwari, S., Pelz-Stelinski, K. S., Alborn, H. T., et al. (2012). Induced release of a plant-defense volatile ‘deceptively’ attracts insect vectors to plants infected with a bacterial pathogen. PLoS Pathog. 8:e1002610. doi: 10.1371/journal.ppat.1002610
Mauck, K., Bosque-Perez, N. A., Eigenbrode, S. D., De Moraes, C. M., and Mescher, M. C. (2012). Transmission mechanisms shape pathogen effects on host-vector interactions: evidence from plant viruses. Funct. Ecol. 26, 1162–1175. doi: 10.1111/j.1365-2435.2012.02026.x
Mauck, K. E., De Moraes, C. M., and Mescher, M. C. (2010). Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. U.S.A. 107, 3600–3605. doi: 10.1073/pnas.0907191107
Mauck, K. E., De Moraes, C. M., and Mescher, M. C. (2014a). Biochemical and physiological mechanisms underlying effects of Cucumber mosaic virus on host-plant traits that mediate transmission by aphid vectors. Plant Cell Environ. 37, 1427–1439. doi: 10.1111/pce.12249
Mauck, K. E., Smyers, E., De Moraes, C. M., and Mescher, M. C. (2014b). Virus infection influences host plant interactions with non-vector herbivores and predators. Funct. Ecol. 29, 662–673. doi: 10.1111/1365-2435.12371
McLachlan, A. (1999). Parasites promote mating success: the case of a midge and a mite. Anim. Behav. 57, 1199–1205. doi: 10.1006/anbe.1999.1087
Mongkolsamrit, S., Kobmoo, N., Tasanathai, K., Khonsanit, A., Noisripoom, W., Srikitikulchai, P., et al. (2012). Life cycle, host range and temporal variation of Ophiocordyceps unilateralis/Hirsutella formicarum on Formicine ants. J. Inverteb. Pathol. 111, 217–224. doi: 10.1016/j.jip.2012.08.007
Mouritsen, K. N., and Poulin, R. (2003). Parasite-induced trophic facilitation exploited by a non-host predator: a manipulator's nightmare. Int. J. Parasitol. 33, 1043–1050. doi: 10.1016/S0020-7519(03)00178-4
Nesse, R. M., and Foxman, B. (2011). Evolutionary approaches to sexually transmitted diseases. Ann. N.Y. Acad. Sci. 1230, 1–3. doi: 10.1111/j.1749-6632.2011.06078.x
Parlog, A., Schlüter, D., and Dunay, I. R. (2015). Toxoplasma gondii-induced neuronal alterations. Parasite Immunol. 37, 159–170. doi: 10.1111/pim.12157
Poirotte, C., Kappeler, P. M., Ngoubangoye, B., Bourgeois, S., Moussodji, M., and Charpentier, M. J. E. (2016). Morbid attraction to leopard urine in Toxoplasma-infected chimpanzees. Curr. Biol. 26, R98–R99. doi: 10.1016/j.cub.2015.12.020
Poulin, R. (1994). The evolution of parasite manipulation of host behavior - a theoretical analysis. Parasitology 109, S109–S118. doi: 10.1017/S0031182000085127
Poulin, R. (1995). “Adaptive” changes in the behaviour of parasitized animals: a critical review. Int. J. Parasitol. 25, 1371–1383. doi: 10.1016/0020-7519(95)00100-X
Poulin, R. (2000). Manipulation of host behaviour by parasites: a weakening paradigm? Proc. Roy. Soc. Lond. B 267, 787–792. doi: 10.1098/rspb.2000.1072
Poulin, R. (2010). “Chapter 5 - Parasite manipulation of host behavior: an update and frequently asked questions,” in Advances in the Study of Behavior, eds H. J. Brockmann, T. J. Roper, M. Naguib, K. E. Wynne-Edwards, J. C. Mitani, and W. S. Leigh (Burlington: Academic Press), 151–186.
Prandovszky, E., Gaskell, E., Martin, H., Dubey, J. P., Webster, J. P., and McConkey, G. A. (2011). The neurotropic parasite Toxoplasma gondii increases dopamine metabolism. PLoS ONE 6:e23866. doi: 10.1371/journal.pone.0023866
Raina, A. K., Adams, J. R., Lupiani, B., Lynn, D. E., Kim, W. J., Burand, J. P., et al. (2000). Further characterization of the gonad-specific virus of corn earworm, Helicoverpa zea. J. Inverteb. Pathol. 76, 6–12. doi: 10.1006/jipa.2000.4942
Rajabaskar, D., Bosque-Pérez, N. A., and Eigenbrode, S. D. (2014). Preference by a virus vector for infected plants is reversed after virus acquisition. Virus Res. 186, 32–37. doi: 10.1016/j.virusres.2013.11.005
Scheideler, M., Schlaich, N. L., Fellenberg, K., Beissbarth, T., Hauser, N. C., Vingron, M., et al. (2002). Monitoring the switch from housekeeping to pathogen defense metabolism in Arabidopsis thaliana using cDNA arrays. J. Biol. Chem. 277, 10555–10561. doi: 10.1074/jbc.M104863200
Schroderus, E., Jokinen, I., Koivula, M., Koskela, E., Mappes, T., Mills, S. C., et al. (2010). Intra- and intersexual trade-offs between testosterone and immune system: implications for sexual and sexually antagonistic selection. Am. Nat. 176, E90–E97. doi: 10.1086/656264
Shi, X. B., Pan, H. P., Xie, W., Wu, Q. J., Wang, S. L., Liu, Y., et al. (2013). Plant virus differentially alters the plant's defense response to its closely related vectors. PLoS ONE 8:e83520. doi: 10.1371/journal.pone.0083520
Shi, X., Pan, H., Zhang, H., Jiao, X., Xie, W., Wu, Q., et al. (2014). Bemisia tabaci Q carrying tomato yellow leaf curl virus strongly suppresses host plant defenses. Sci. Rep. 4:5230. doi: 10.1038/srep05230
Shirbazou, S., Abasian, L., and Meymand, F. T. (2011). Effects of Toxoplasma gondii infection on plasma testosterone and cortisol level and stress index on patients referred to Sina hospital, Tehran. Jundishapur J. Microbiol. 4, 167–173. Available online at: http://jjm.ajums.ac.ir
Sloan, D. B., Giraud, T., and Hood, M. E. (2008). Maximized virulence in a sterilizing pathogen: the anther-smut fungus and its co-evolved hosts. J. Evol. Biol. 21, 1544–1554. doi: 10.1111/j.1420-9101.2008.01604.x
Smallegange, R. C., van Gemert, G.-J., van De Vegte-Bolmer, M., Gezan, S., Takken, W., Sauerwein, R. W., et al. (2013). Malaria infected mosquitoes express enhanced attraction to human odor. PLoS ONE 8:e63602. doi: 10.1371/journal.pone.0063602
Soh, L. J. T., Vasudevan, A., and Vyas, A. (2013). Infection with Toxoplasma gondii does not elicit predator aversion in male mice nor increase their attractiveness in terms of mate choice. Parasitol. Res. 112, 3373–3378. doi: 10.1007/s00436-013-3545-6
Soper, D. M., King, K. C., Vergara, D., and Lively, C. M. (2014). Exposure to parasites increases promiscuity in a freshwater snail. Biol. Lett. 10:1091. doi: 10.1098/rsbl.2013.1091
Stafford, C. A., Walker, G. P., and Ullman, D. E. (2011). Infection with a plant virus modifies vector feeding behavior. Proc. Natl. Acad. Sci. U.S.A. 108, 9350–9355. doi: 10.1073/pnas.1100773108
Tejerizo, G., Doménech, A., Illera, J. C., Silván, G., and Gómez-Lucía, E. (2012). Altered plasma concentrations of sex hormones in cats infected by feline immunodeficiency virus or feline leukemia virus. Domest. Anim. Endocrinol. 42, 113–120. doi: 10.1016/j.domaniend.2011.11.001
Thaler, J. S., Humphrey, P. T., and Whiteman, N. K. (2012). Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 17, 260–270. doi: 10.1016/j.tplants.2012.02.010
Thomas, F., Adamo, S., and Moore, J. (2005). Parasitic manipulation: where are we and where should we go? Behav. Proc. 68, 185–199. doi: 10.1016/j.beproc.2004.06.010
Thomas, F., Poulin, R., and Brodeur, J. (2010). Host manipulation by parasites: a multidimensional phenomenon. Oikos 119, 1217–1223. doi: 10.1111/j.1600-0706.2009.18077.x
Thomas, F., Schmidt-Rhaesa, A., Martin, G., Manu, C., Durand, P., and Renaud, F. (2002). Do hairworms (Nematomorpha) manipulate the water seeking behaviour of their terrestrial hosts? J. Evol. Biol. 15, 356–361. doi: 10.1046/j.1420-9101.2002.00410.x
van Houte, S., Ros, V. I. D., and van Oers, M. M. (2013). Walking with insects: molecular mechanisms behind parasitic manipulation of host behaviour. Mol. Ecol. 22, 3458–3475. doi: 10.1111/mec.12307
van Houte, S., van Oers, M. M., Han, Y., Vlak, J. M., and Ros, V. I. D. (2014). Baculovirus infection triggers a positive phototactic response in caterpillars to induce ‘tree-top’ disease. Biol. Lett. 10:20140680. doi: 10.1098/rsbl.2014.0680
Vantaux, A., De Sales Hien, D. F., Yaméogo, B., Dabiré, K. R., Thomas, F., Cohuet, A., et al. (2015). Host-seeking behaviors of mosquitoes experimentally infected with sympatric field isolates of the human malaria parasite Plasmodium falciparum: no evidence for host manipulation. Front. Ecol. Evol. 3:86. doi: 10.3389/fevo.2015.00086
Vyas, A. (2013). Parasite-augmented mate choice and reduction in innate fear in rats infected by Toxoplasma gondii. J. Exp. Biol. 216, 120–126. doi: 10.1242/jeb.072983
Vyas, A., Kim, S.-K., Giacomini, N., Boothroyd, J. C., and Sapolsky, R. M. (2007). Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proc. Natl. Acad. Sci. U.S.A. 104, 6442–6447. doi: 10.1073/pnas.0608310104
Wang, J., Bing, X.-L., Li, M., Ye, G.-Y., and Liu, S.-S. (2012). Infection of tobacco plants by a begomovirus improves nutritional assimilation by a whitefly. Entomol. Exp. Appl. 144, 191–201. doi: 10.1111/j.1570-7458.2012.01278.x
Weinersmith, K., and Faulkes, Z. (2014). Parasitic manipulation of hosts' phenotype, or how to make a Zombie - an introduction to the symposium. Integrat. Compar. Biol. 54, 93–100. doi: 10.1093/icb/icu028
Wennström, A., Ericson, L., and Garcia-Guzman, G. (2003). The concept of sexually transmitted diseases in plants: definition and applicability. Oikos 100, 397–402. doi: 10.1034/j.1600-0706.2003.12004.x
Wood, C. L., and Johnson, P. T. J. (2015). A world without parasites: exploring the hidden ecology of infection. Front. Ecol. Environ. 13, 425–434. doi: 10.1890/140368
Worth, A. R., Lymbery, A. J., and Thompson, R. C. A. (2013). Adaptive host manipulation by Toxoplasma gondii: fact or fiction? Trends Parasitol. 29, 150–155. doi: 10.1016/j.pt.2013.01.004
Wright, G. A., Baker, D. D., Palmer, M. J., Stabler, D., Mustard, J. A., Power, E. F., et al. (2013). Caffeine in floral nectar enhances a pollinator's memory of reward. Science 339, 1202–1204. doi: 10.1126/science.1228806
Zamanian, M., Fraser, L. M., Agbedanu, P. N., Harischandra, H., Moorhead, A. R., Day, T. A., et al. (2015). Release of small RNA-containing exosome-like vesicles from the human filarial parasite Brugia malayi. PLoS Negl. Trop. Dis. 9:e4069. doi: 10.1371/journal.pntd.0004069
Alteration: Differences in the phenotype of an organism that can be observed after a specific event.
Elicitation: The causing of phenotypic alterations in the phenotype of organism A by its interaction with organism B.
Final host: A host species in which a parasite can conclude its life cycle and reproduce.
Host change: As part of its dispersal behavior, the individual parasite moves from one individual host organism to another.
Host shift: During the evolution of the parasite, genetic or geographical shifts in the populations of the host or the parasite facilitate the parasite gaining positive fitness in a new host species.
Host switch: As part of its ontogeny, the individual parasite at a certain life-history stage must move from host species A to host species B.
Induction: The causing of alterations in the phenotype of organism A by its interaction with organism B that are under the genetic control of organism A.
Intermediate host: A host species that serves the dispersal of the parasite but usually does not allow for its reproduction, at least not in the sexual phase.
Manipulation: The causing of alterations in the phenotype of organism A by its interaction with organism B that are under the genetic control of organism B.
Response: An alteration in the phenotype of organism A that is under its own genetic control.
Parasite: Term used here in the widest possible sense to denominate viruses, bacteria, fungi, protozoa and animals that must colonize a living host to gain positive fitness and whose presence usually has a negative effect on the fitness of the host.
Vector: Term used here as synonym for intermediate host.
Keywords: immunity, malaria, partner manipulation, pathogen, plant–enemy interaction, STDs, toxoplasmosis
Citation: Heil M (2016) Host Manipulation by Parasites: Cases, Patterns, and Remaining Doubts. Front. Ecol. Evol. 4:80. doi: 10.3389/fevo.2016.00080
Received: 18 December 2015; Accepted: 14 June 2016;
Published: 28 June 2016.
Edited by:
Sara Goodacre, University of Nottingham, UKReviewed by:
Martin Taylor, Bangor University, UKCopyright © 2016 Heil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Heil, bWhlaWxAaXJhLmNpbnZlc3Rhdi5teA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.