
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci. , 01 August 2024
Sec. Ecosystem Restoration
Volume 12 - 2024 | https://doi.org/10.3389/fenvs.2024.1401513
This article is part of the Research Topic The Sustainable Management of Land Systems View all 24 articles
 Kirsten Jung1*†
Kirsten Jung1*† Miriam Teuscher2†Stefan Böhm1,3†
Miriam Teuscher2†Stefan Böhm1,3† Konstans Wells4†
Konstans Wells4† Manfred Ayasse1Markus Fischer5†Wolfgang W. Weisser6†Swen C. Renner7†‡Marco Tschapka1,8†‡
Manfred Ayasse1Markus Fischer5†Wolfgang W. Weisser6†Swen C. Renner7†‡Marco Tschapka1,8†‡In modified production landscapes, biodiversity faces unprecedented pressures from human actions, resulting in significant species declines of plant and animal taxa, including birds. Understanding the underlying mechanisms responsible for such declines is essential to counteract further loss and support practitioners in conserving biodiversity and associated ecosystem function. In this study, we used standardized bird monitoring data collected over 6 years in managed forest and grassland areas across different regions in Germany, Central Europe. We combined these data with morphometric, ecological, behavioral, and acoustic trait data and detailed information on local land use management practices to understand how management decisions affect species and functional diversity, as well as ecological processes shaping local species composition. Our results reveal that the ecosystem and regional context must be considered to understand how management practices affect bird diversity aspects and composition. In forests, regional management decisions related to tree species and stand age affected bird diversity, as well as community and functional composition, and indicated environmental sorting due to ecological and behavioral requirements, biotic interactions, and morphometric constraints. In grasslands, independent of local management practices, increased intensity of land use resulted in an overall loss in bird species richness and functional diversity. Predominantly, constraints due to ecological or behavioral requirements affected bird species assemblage composition. In addition, our results indicated the importance of woody vegetation near managed grasslands and of considering environmental conditions beyond the local scale to support bird diversity and associated ecosystem functions. Our results highlighted that local management decisions can support bird diversity and maintain ecological function. However, this needs a view beyond the local scale of management units. It also demands a joint effort of biologists and land managers to integrate targeted conservation actions into regional management practices and create a network of habitats within production landscapes to protect nature, guard against biotic and functional homogenization, and prevent further degradation of ecosystems in production landscapes.
Centuries of human land use have shaped the current landscapes in central Europe and created mosaics of different habitats for wildlife (Ellis et al., 2021). However, especially during the past decades, biodiversity has faced increasing pressures from human actions (Newbold et al., 2015; Joppa et al., 2016), including the intensification and industrialization of land use practices and land conversion (Foley et al., 2005). Altogether, this caused significant transformations from natural or near-natural to extreme anthropogenic landscapes (IPBES, 2019) and includes environmental changes from the local to the landscape scale (Tilman et al., 2017).
These environmental transformations are accompanied with an unprecedented global loss in biodiversity (IUCN 2019) across a wide array of plant and animal species groups (Ceballos et al., 2015; IPBES, 2019), including birds.
Among vertebrates, birds are one of the functionally most diverse taxa inhabiting different habitats across all climatic regions of the earth. They are integral parts of ecosystems in almost all terrestrial and aquatic ecosystems, and in addition to playing essential key roles in ecosystems, such as seed dispersal and predation, the acoustic diversity of bird song (Hedblom et al., 2014) has been shown to positively influence human empathy toward nature (Methorst et al., 2021).
Approximately 60% of the bird species globally are, however, reported to be declining by the IUCN (e.g., Johnson et al., 2011). Negative developments over the past decades are also reported for many European populations (e.g., Bowler et al., 2019; Burns et al., 2021), where declines have been linked to increased intensity and industrialization of local land use management practices across different ecosystems, including agricultural areas (Reif and Vermouzek, 2019; Rigal et al., 2023) and production forests (Reif et al., 2023).
Although land use and its effect on overall population trends may vary regionally (Bowler et al., 2021) between groups of species (Rigal et al., 2023) and across different spatial scales (Leroy et al., 2023), responses of individual bird species have been linked to functional traits such as size and mobility (Concepción et al., 2015), as well as resource and habitat specialization (e.g., Bowler et al., 2019; Reif and Vermouzek, 2019), key characteristics that have been suggested to determine the tolerance to environmental change (Tews et al., 2004; Öckinger et al., 2011) across various taxa.
Trait-based approaches provide more meaningful insights into species response to environmental change (Ali et al., 2022) and are more suited to understand and predict environmental effects on species occurrence and assembly than species numbers and species diversity measures alone (Mouchet et al., 2010; Wells et al., 2012; Carmona et al., 2021; Schleuning et al., 2022; Mouchet et al., 2010). Moreover, patterns of functional diversity (Villéger et al., 2008; Laliberté and Legendre, 2010) and trait composition (Lisboa et al., 2014) may thus help to understand the drivers of species occurrences and assemblage composition along environmental gradients (e.g., Mouchet et al., 2010; Schleuning et al., 2022), including the relative strength of environmental filtering and biotic interaction processes in differently managed environments (Mason et al., 2005; Gámez-Virués et al., 2015; Kraft et al., 2015). A true understanding of such processes is important to counteract further species loss and support practitioners in maintaining bird diversity and associated ecosystem function and services.
In birds, functional traits reflect distinct associations with environment, lifestyle, and diet (Tobias et al., 2022) and, thus, shape ecological niches (Pigot et al., 2020) and function (Ali et al., 2022). Bird traits are well-documented (Tobias et al., 2020), including morphometric parameters, ecological and behavioral attributes, and acoustic characteristics. Morphometric traits predominantly reflect the physical performance, such as aerodynamic abilities (e.g., mobility and maneuverability) or resource accessibility (beak size and strength) of species. Morphometric traits are correlated with ecological niches across the global bird phylogeny (Pigot et al., 2020); however, in regional datasets, morphometric traits alone may hamper the identification of environmental challenges due to niche-based processes and biotic interactions (Cadotte and Tucker, 2017; Rigal et al., 2022). Such challenges may be better described using ecological and behavioral trait classifications, such as habitat preferences, trophic level, dietary resource type, and foraging and migratory behavior (Tobias et al., 2022). In addition, acoustic traits, which are crucial for bird species communication to find mates and defend territories, may better reflect environmental challenges to ensure the acoustic recognition of a species identity, territorial quality, and individual fitness in differently structured habitats.
In this study, we used a large-scale dataset of birds collected in forest and grassland production landscapes across three regions in Germany, Central Europe. The data include the two major land cover types in Germany, grassland (21%–39% land cover depending on the federal state) and forest (39% land cover across Germany) ecosystems (Federal Statistical Office, 2022), both being under pressure through increased land conversion and representing environments where anthropogenic disturbance is relatively limited compared to industrialized agricultural and urban areas and, thus, are highly valuable ecosystems to support biodiversity.
Bird species observations were collected during 6 years using standardized audio–visual point counts and used to calculate alpha diversity and assess species composition per site. These data were combined with species trait data, including morphometric, ecological and behavioral, and acoustic traits and very detailed information about local land use management practices and land use intensity (LUI).
Our aim was to shed light on how the management of forest and grassland is linked to changes in species diversity and functional diversity aspects of bird assemblages. We thus assessed regional and ecosystem-specific differences in species and functional diversity metrics and composition and investigated the local effects of different management practices and land use intensity. To address the importance of different ecological processes shaping local species composition, we distinguished between morphometric, ecological and behavioral, and acoustic trait dimensions, which represent distinct environmental challenges due to management practices that may cause ecological sorting or biotic homogenization and can help evaluate regional- and ecosystem-specific differences in management effects. Finally, we investigated whether the strength of ecological processes may differ due to land management practices and/or change due to land use intensity.
Our study was conducted within the long-term research platform of the Biodiversity Exploratories (www.biodiversity-exploratories.de), a large-scale project for functional biodiversity research, which encompasses three regions along a north–south axis in Germany: 1) the Schorfheide-Chorin Biosphere Reserve, a young glacial landscape characterized by moraines, lakes, and marshes in the lowlands (3–140 m asl) of northeast Germany; 2) the Hainich-Dün area situated in the hilly lands (300–400 m asl) of central Germany; and 3) the Schwäbische Alb plateau in the low Swabian Jura mountain ranges (720–840 m asl) of southwest Germany. All three regions are shaped by the anthropogenic land management of grassland and forest areas and harbor 100 permanently marked experimental sites in differently managed forest (n = 50; 100 * 100 m) and grassland systems (n = 50; 50*50 m), totaling 300 sites (Fischer et al., 2010). The average annual temperature decreases from north to south and reaches 8.1°C–9.6°C in the Schorfheide-Chorin region, 7.5°C–9.2°C in the Hainich-Dün region, and 6.9°C–8.6°C on the Schwäbische Alb plateau. In contrast, the average annual precipitation increases from north to south, with 450–790 mm in the Schorfheide-Chorin region, 350–860 mm in the Hainich-Dün region, and 810–1,100 mm in the Schwäbische Alb plateau, according to project-based records from 2008 to 2018.
In all three exploratories, experimental sites differed in management practices, which we classified into different management categories. Management categories in forests include old unmanaged beech, oak, and spruce stands in different developmental stages, mixed cultures, and intensely managed beech, pine, and spruce monocultures (Supplementary Table S1). Grasslands are managed either as meadows, mown pastures, and pastures (cattle, horse, and sheep grazing), where the management intensity varies from unfertilized and extensive grazing to high fertilizer input, mowing frequency, and grazing pressure on production grasslands (Supplementary Table S2). Annual information from local foresters and farmers allows us to quantify the management intensity of individual sites and was used to calculate the quantitative indices of silvicultural management intensity (SMI) in forests following the procedures outlined by Schall and Ammer (2013) and LUI for grasslands following the procedures outlined by Blüthgen et al. (2012). To reflect the environmental heterogeneity of the grassland neighborhood, we additionally quantified the proportion of woody vegetation coverage in a 100-m radius from the center of each site based on a land cover map provided by Schug et al. (2020). This map was originally designed to discriminate between woody, non-woody, and build-up surface at a 10-m resolution for all of Germany, using Sentinel-2 satellite data. We thus cross-checked the extracted data with digital aerial orthophotos from the three exploratory regions (License CC BY 4.0) and confirmed that the extracted wood vegetation from this map corresponds to single trees, hedges, and forests in the direct vicinity of our grassland sites.
Bird surveys were conducted for 5 consecutive years between 2008 and 2012 and again in 2018. Sampling per year included five visits to each of the 300 sites in the grasslands and forests during the main breeding season from March to June. Observers conducted standardized audio–visual point counts for 5 min during the morning chorus between sunrise and 11:00 a.m. Surveys were conducted by experienced ornithologists or trained and tested student helpers. To reduce observer bias, the personnel did not change during the season, and changes in personnel between years were kept to a minimum. All visible and audible birds exhibiting territorial, breeding, or feeding behavior on site were counted as a record. For later analysis, we, however, only considered the maximum number of individuals observed during one of the five visits per year as a measure for the relative abundance of a bird species per site. In addition, we considered a presence-only count of a species per year and site as a measure for a species occurrence and used the accumulated occurrence counts of a species over a total of 6 years (based on 30 visits to a site) to determine local bird species assemblage composition.
All bird species were characterized by a suite of functional traits including 1) morphometric measurements, which reflect physical limitations to individual species for resource use and movement; 2) ecological and behavioral traits, which reflect the environmental and interaction-based niches of a species; and 3) acoustic features of bird song, which reflect acoustic challenges for social communication and optimal signal design in differently structured environments (Table 1).

Table 1. Functional traits used to calculate functional diversity measures in morphometric, ecological and behavioral, as well as acoustic trait space, and community weighted means of trait composition. Given are trait variable names, challenges associated with the three trait groups, and individual trait descriptions. Description of morphometric, ecological and behavioral traits follow Tobias et al. 2021 (Avonet) and are slightly Descriptions of morphometric, as well as ecological and behavioral traits follow Tobias et al. (2022) (Avonet), but are slightly adapted to the current study system; acoustic trait description represents definitions for feature extraction during acoustic analysis.
Morphometric, ecological, and behavioral traits were extracted from the online data repository AVONET (Tobias et al., 2022). For morphometric traits, we included measurements of the wings (wing length and Kipp’s distance), tarsus and tail lengths, body mass, beak length, and beak width, which determine the aerodynamic ability of a species, e.g., mobility and maneuverability (Rayner, 1988), as well as resource specializations. Ecological and behavioral traits included habitat preferences, including the relative use of different habitat types and environmental conditions with respect to habitat density, foraging style, trophic position and trophic niche, migratory behavior, and the centroid of the latitudinal distribution of the respective species, reflecting ecological or behavioral requirements and ecological niches (Tobias et al., 2020). In addition, we assessed acoustic characteristics of the song of a species, which determines the acoustic identity of a species and the occupied acoustic space (Luther, 2009) within bird assemblages. Acoustic trait data were extracted from original audio data of bird songs available from the online repository xeno-canto (https://xeno-canto.org). We restricted downloads of recordings to those of the highest-quality ranking (“A”) and an origin from Germany. We then selected recordings for further analysis based on a good signal-to-noise ratio (minimum 15 db) and unambiguity of the species identification. In very few cases, where xeno-canto did not provide sufficient recordings, we used the online sound archive of the Naturkundemuseum Berlin and supplemented the acoustic data with recordings of a commercially available CD. All recordings considered for analysis were made with a minimum sample rate of 32 kHz and a maximum of 48 kHz (resulting in a resolution of 32–41 Hz and 2.8–4 ms).
As we were interested in the species divergence of acoustic features, which allow partitioning of the acoustic space to improve communication with conspecific receivers, we focused the acoustic analysis on the complete song, rather than song components or elements (notes), following the procedure used by Tobias et al. (2010). The song structure and characteristics were then quantified using temporal and spectral measures, i.e., the duration of song, number of elements (notes), the maximum and minimum frequencies, and mean frequency of the entire song. In addition, we calculated the bandwidth and pace of the song (number of elements per song).
All acoustic measurements were obtained using Avisoft SASLab Pro 5.3.02 (Raimund Specht Berlin, Germany) and obtained from a spectrogram generated with an FFT of 1,024, a flat-top window, and 87% overlap. Songs were automatically detected using a threshold of −35 db, and the start and end of the song were determined by –6 db relative to the detection threshold. To ensure that the features of complete phrases were extracted, we set the hold time to 500 ms. For sub-element detection, the hold time was set to 10 ms. All measurements were visually inspected for acoustic disturbance effects and filtered if necessary to ensure that the extracted features are representative for the respective species.
Based on the relative abundance of bird species per site, we calculated the effective number of species (ENS, vegan package; Oksanen, 2019) and considered it a measure of species diversity (Jost, 2006). To further quantify the similarity in species assembly across sites, we used non-metric multidimensional scaling (NMDS; vegan package). NMDS scores were based on the Hellinger-transformed accumulated presence counts of a species across all 6 years, and Bray–Curtis similarity was used to assess the similarity between site-specific bird assemblages.
Using the assembled functional traits described above, we calculated functional diversity metrics, including functional richness, functional evenness (Villéger et al., 2008), and functional dispersion (Laliberté and Legendre, 2010). Here, functional richness is defined as the “volume” in functional trait space occupied within an assemblage (Villéger et al., 2008) and increases with an increase in functionally distinct species (independent of species abundances). Functional evenness represents the homogeneity of trait abundance distribution in the functional space (Villéger et al., 2008). Functional evenness approaches zero if only a narrow part within the functional trait space is occupied (e.g., in species-poor assemblages or assemblages with high dominance of single species). A decrease in functional evenness thus indicates an abundance shift toward specific traits (Mason et al., 2005), which are, for example, more suitable under certain environmental conditions, suggesting environmental sorting. In addition, functional dispersion reflects the mean distance in the multidimensional trait space of a species to the centroid of all species within a community, weighted by its abundance (Laliberté and Legendre, 2010), and reflects functional dissimilarity in traits within assemblages. To interpret the relative importance of individual traits contributing to the three different functional diversity metrics, namely, functional richness, functional dispersion, and functional evenness, we used random forest (function: random forest, 1,000 permutations; (Breimann 2001; Supplementary Figure S1.1)).
To further investigate the similarity in trait composition across species assemblages from different sites, we also calculated the community weighted means of traits, weighted by the relative abundance of individual bird species per site package FD (Laliberté et al., 2022), and then used NMDS (vegan package) to compute the similarity in trait composition between sites.
Both functional diversity metrics and community weighted means of traits were based on Gower’s distances, and a “Cailliez” correction was applied.
Functional diversity metrics and multidimensional trait space were calculated first across all traits combined and then separately for 1) morphometric, 2) ecological and behavioral, and 3) acoustic subsets of traits to evaluate the importance of different trait dimensions as the drivers of species assemblage composition.
We assessed regional and ecosystem-specific differences in species and functional diversity metrics using generalized linear mixed-effect models (package: glmmTMB), including an interaction term between region and habitat types (forest and grassland; formula: glmmTMB (diversity metrics ∼ region * habitat type). The models were fitted using a negative binominal distribution, and sites were included as random factors due to repetitive sampling. Between-group effects were evaluated using a type II Wald chi-squared test (ANOVA; package car) and EM means (package emmeans).
Within each habitat type, we then assessed whether species diversity and functional diversity metrics correlate using the Pearson product moment correlation and tested correlative significance using linear regression models.
We then explored the effects of land management practices and land use intensity on bird diversity (ENS) by fitting generalized additive models (gams; package mgcv), including the respective management intensity indices (SMI/LUI) as smooth terms and management practice categories as factors. For this, we used a hierarchical approach and fitted a global model across regions {setting region as a random factor; formula: gam [ENS∼s (SMI/LUI) + management practice + s (region, bs = “re”)]} followed up by a separate regional model in which the region was included as a moderating effect for either silvicultural management intensity or local land use intensity in grasslands {Formula: gam [ENS∼s (SMI/LUI, by = region) + management practice]}. In all models, geographic coordinates were included as random effects to account for geographic autocorrelation. Models were fit either with negative binominal (grasslands) or Gaussian distribution (forests) after the visual inspection of model diagnostics (function appraise; package: gratia).
We further evaluated patterns of species and trait composition. We first assessed whether species composition and functional compositions (across all traits and separately for morphometric, ecological, behavioral, and acoustic trait space) differed between regions, habitat types, and management types using permutational multivariate analysis of variance (function adonis, vegan package, 1,000 permutations).
To assess possible relationships between the similarities in species composition and functional composition across sites, we then performed pairwise Procrustes correlations (functions: procrustes) and tested the significance using procrustean randomization tests (function: protest, 999 randomizations, vegan package) based on the extracted NMDS scores with symmetric rotation. Strong correlations, indicating a high interdependence of species occurrences and trait composition, suggest that traits play an essential role in determining species occurrence and structuring local assemblages. Pairwise Procrustes tests were conducted across all traits combined and then separately for 1) morphometric, 2) ecological and behavioral, and 3) acoustic subsets of traits to evaluate the relative importance of different trait dimensions.
To further understand how management decisions may affect the interdependence of species and functional composition, we then extracted procrustean residuals (PAMs) as a measure of the correlative strength between species and functional composition (all traits combined and individual trait dimensions). Here, smaller residuals reflect a stronger correlation and interdependence between species composition and trait composition, while larger residuals reflect lower interdependence (Lisboa et al., 2014). We then assessed the variation in procrustean residuals (PAMS) due to land use management practices using ANOVA (function aov and Tukey’s HSD post hoc test) and silvicultural management intensity or local grassland management intensity using gams (package mgcv). We included region as a factor and fitted the respective management intensity as a smooth [formula: PAMres ∼ EXPLO + s (management intensity)]. For grasslands, we also included the amount of woody vegetation as an additional smooth term [Formula: PAMres ∼ EXPLO + s (Land use intensity) + s (Woody vegetation)]. In all models did we include geographic coordinates as random effects to account for potential geographic autocorrelation.
All statistical analyses were carried out using R software (version 4.31.2, R Core Team, 2023) in the RStudio environment (2023.09.1).
A technical framework diagram and an overview of the different research variables are given in Supplementary Figure S1.2, Table S3.
In total, we observed 100 bird species across all three exploratories and habitat types, with 70–83 species recorded in each region (Schorfheide-Chorin: 83; Hainich-Dün: 82; and Schwäbische Alb plateau: 70). Across the monitored years, the effective number of bird species varied only slightly, with a decrease in bird observations and species diversity from 2008 to 2009 and a slight recovery in 2010–2011 and 2012 (Supplementary Figure S1.3). Only very few species were exclusively observed within one of the three regions (Supplementary Figure S1.4).
Species diversity and functional richness exhibited similar expected average numbers and variation between the three regions (Table 2), whereas considerably less species diversity (GLMM: χ2 = 1,539.58; p < 0.001) and lower functional richness (GLMM: χ2 = 8.15; p < 0.01) (Figure 1) were found in grasslands than in forest sites. Similar results were also obtained when considering morphometric, ecological and behavioral, and acoustic richness separately.
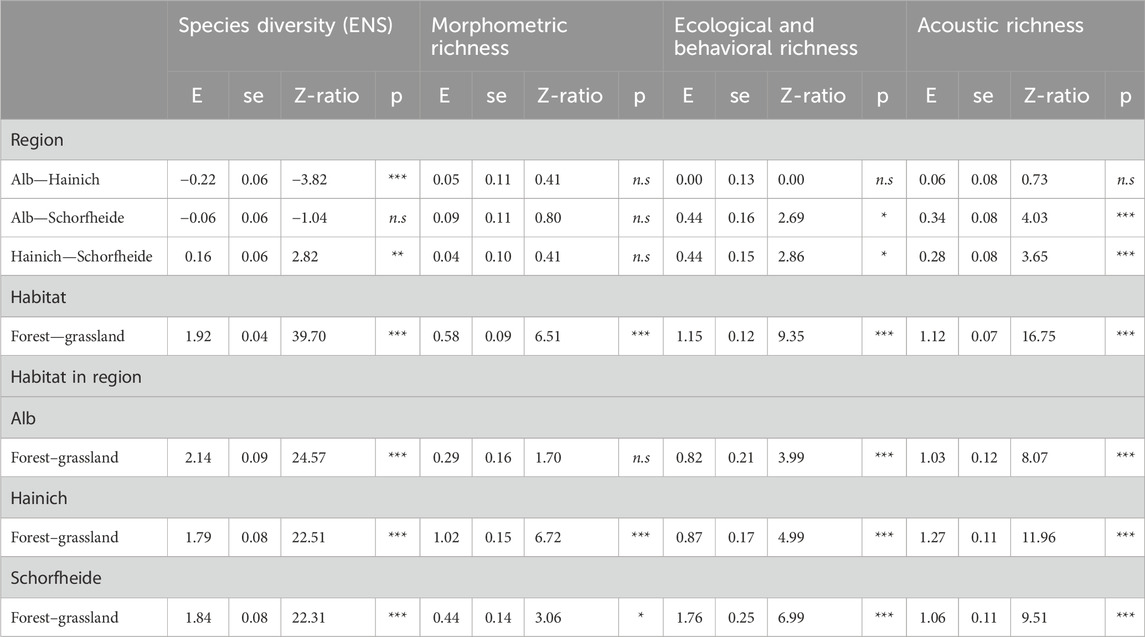
Table 2. Region- and habitat-specific differences in species diversity and morphometric, ecological and behavioral, as well as acoustic richness, assessed with generalized linear mixed-effect models (package: glmmTMB). Presented here are pairwise differences (based on EM means) between exploratory regions, habitats, and habitat types within each region.
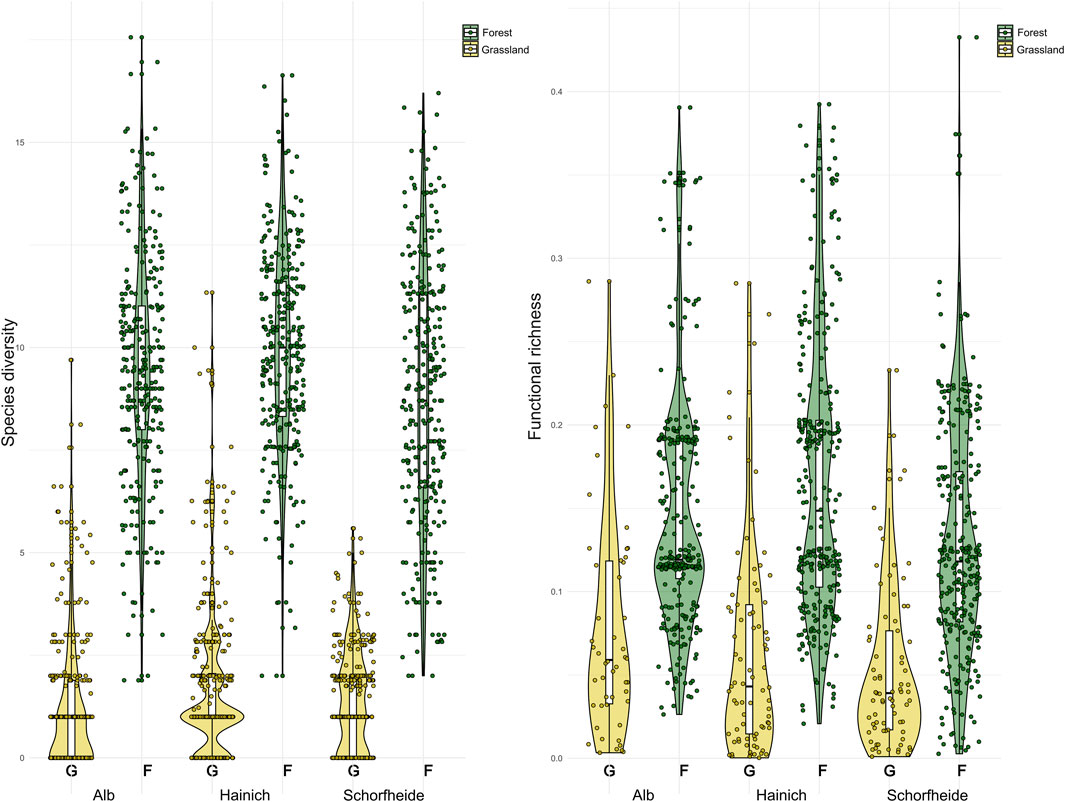
Figure 1. Bird diversity and functional richness and its variation across trait spaces in grasslands (G, yellow) and forests (F, green) of the three different Biodiversity Exploratory regions.
In both forests and grasslands, bird species diversity was correlated positively with bird functional richness (forest: R2 = 0.35, F = 474.81, p < 0.001; grassland: R2 = 0.45, F = 166.46, p < 0.001) and functional dispersion (forest: R2 = 0.15, F = 1,898 = 165.44, p < 0.001; grassland: R2 = 0.56, F1 = 603 = 776.14, p < 0.001) (Figure 2). However, in forests, the overall functional evenness across trait spaces decreased with species diversity (R2 = 0.01, F1 = 898 = 7.02, p < 0.05) but increased with increasing species diversity in grasslands (R2 = 0.1, F1 = 206 = 18.31, p < 0.001).
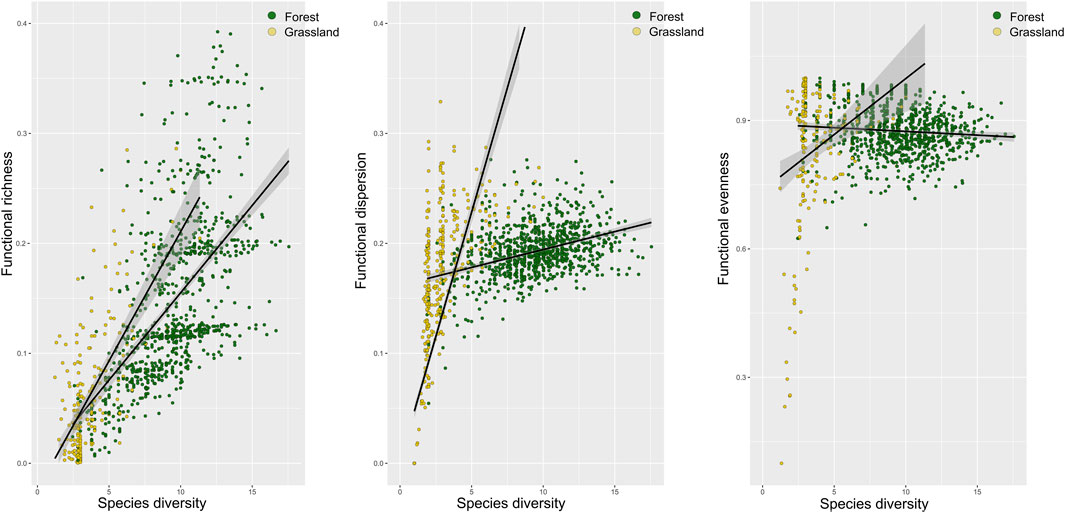
Figure 2. Functional richness, dispersion (trait dissimilarity; Laliberté and Legendre, 2010), and evenness (homogeneity of trait abundances) across trait dimensions with increasing species diversity in grasslands (yellow) and forests (green).
Bird diversity differed significantly between forest management categories (explained deviance = 37.3%, chi-square = 174, p < 0.001), with lower species diversity in younger forest stands (e.g., beach and pine pole wood) and higher species diversity in old-growth mature beech and mature spruce and structurally more heterogeneous forest stands (e.g., beech thicket with shelterwood; Figure 3). Across regions, silvicultural management intensity did not reveal a significant effect on species diversity (F = 2.5; p > 0.05); however, in the Schorfheide-Chorin region, species diversity decreased significantly with silvicultural management intensity (edf = 1.21, F = 13.9, p < 0.001), while it tended to increase with increasing silvicultural management intensity in the Hainich-Dün area (edf = 1.41, F = 6.0, p = 0.05).
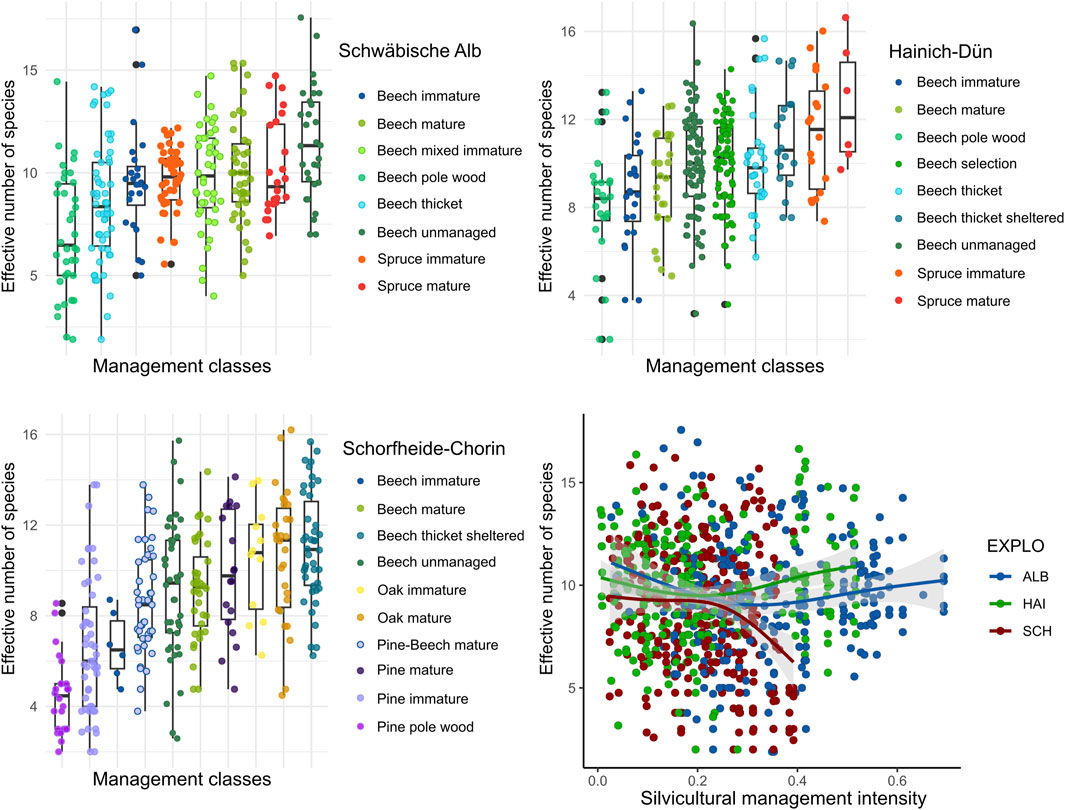
Figure 3. Differences in species diversity between forest management classes within each of the three regions and in response to silvicultural management intensity across regions.
Across regions, bird diversity in grasslands (Figure 4) was slightly higher in pastures and mown pastures than that in meadows; however, independent of local management practices, species diversity decreased significantly with increasing local land use intensity (explained deviation = 35%, χ2: 8.01, p < 0.001). The strength of this effect, however, differed between regions. At the Schwäbische Alb plateau, we found an increasing loss in bird diversity, especially from extensive to intermediate land use intensities, and only a slightly negative response from intermediate to high land use intensities (edf = 2.28, χ2 = 11.65, p < 0.01) (Figure 4). Bird diversity in the Hainich-Dün region tended to linearly decrease with increasing land use intensity (edf = 1, χ2 = 2.875, p = 0.08), whereas we found no significant effect of land use intensity in the Schorfheide-Chorin region (edf: 1, χ2 = 0.48, n.s.).
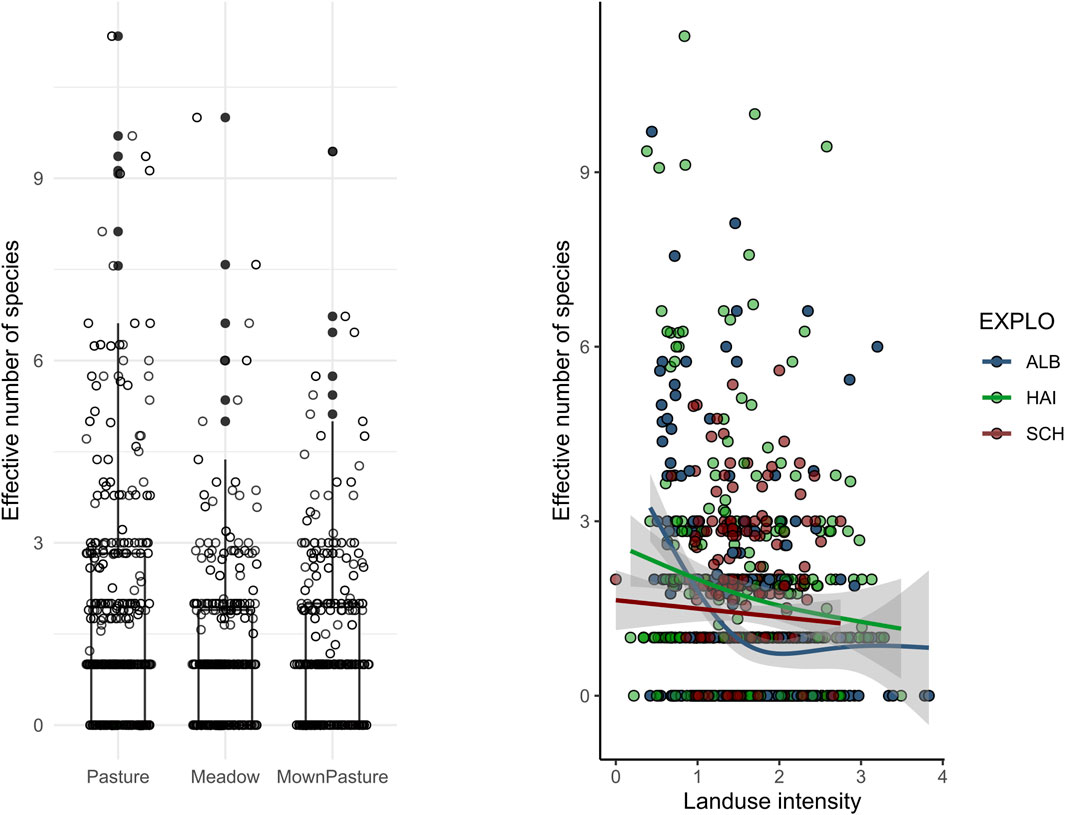
Figure 4. Species diversity patterns in grassland areas, between different management types, and in response to increased management intensity using an index combining mowing frequency, fertilizer input, and grazing pressure.
Non-metric multidimensional scaling clearly separated bird assemblages between sites (NMDS final stress: 0.12; R2 = 0.98, Figure 5). Permutational multivariate analysis of variance confirmed significant differences in species assemblages between regions (adonis R = 13.9, p < 0.001), grassland and forest sites (R = 232.6, p < 0.001), and between different management categories (R = 2.24, p < 0.001) in forests and grasslands. A plot-based permutation test for the homogeneity of multivariate dispersions further indicated a significant turn-over of species (F = 164.18, p < 0.001) between forest and grassland areas in all three exploratories, indicating clearly separated bird assemblages between ecosystems.
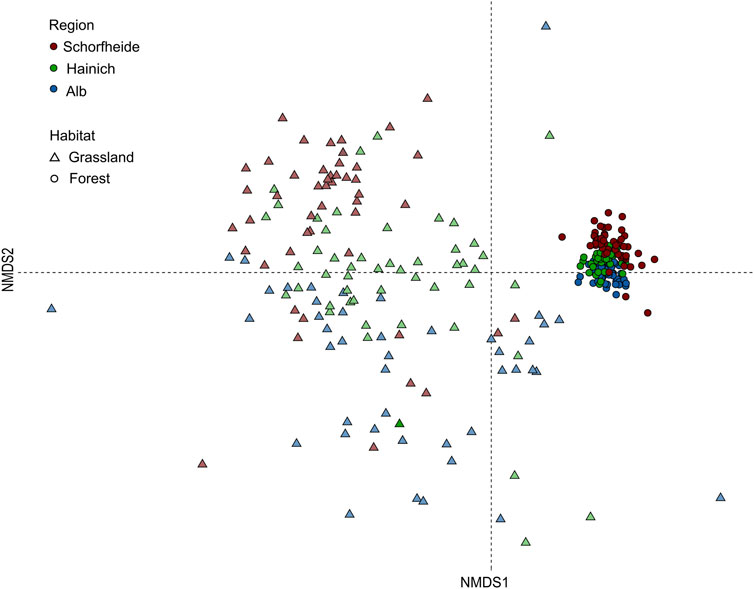
Figure 5. Non-metric multidimensional scaling (Bray–Curtis dissimilarities) of bird assemblages based on differences in the accumulated presence/absence counts of species observations across the 6-year monitoring period on grassland (triangles) and forest (dots) sites. Different regions are represented in different colors (red = Schorfheide-Chorin; green = Hainich-Dün; blue = Schwäbische Alb).
Across the exploratories, differences in taxonomic and functional composition between assemblages (across trait spaces) correlated stronger in forests (Procrustes, m2 = 0.3465, r = 0.8084, p = 0.001) than in grasslands (m2 = 0.7341, r = 0.5156, p < 0.001), suggesting that local environmental conditions via environmental sorting or a biotic interaction based on trait features play a greater role in structuring bird assemblages in differently managed forests than in grasslands.
Habitat-specific analysis, investigating the similarity in species composition, further confirmed significant differences in species assemblages between forest management categories [adonis (species): F = 8.63; p < 0.001] and also indicated significant differences in functional composition [adonis (traits): F = 5.66; p < 0.001; Figure 6], indicating that local forest management decisions affect both bird species composition and ecological function.

Figure 6. Non-metric multidimensional scaling of bird assemblages based on differences in trait composition (across trait dimensions) and between differently managed forests (final stress: 0.14; non-metric fit R2 = 0.98) in the three different regions. Color reflects different management categories in forests, shape reflects different regions, i.e., Alb = circle, Hainich = triangle, and Schorfheide = square, and the size of points is proportional to the average log-scaled occurrence of species at a given site.
Changes in bird species composition were correlated with changes in trait composition for ecological and behavioral (70%, m2 = 0.5005, p = 0.001), acoustic (76%, m2 = 0.42, p < 0.001), and morphometric (59%, m2 = 0.6386, p < 0.001) trait dimensions, pointing toward ecological sorting, especially with respect to ecological requirements and communication with con- and hetero-specifics and also, but to a lesser degree, with respect to morphological characteristics.
The residual correlation strength of species and functional composition across trait dimensions, however, differed significantly between management categories (Figure 7; ecological and behavioral: F = 8.08, p < 0.001; acoustic: F = 8.33, p < 0.001; and morphometric: F = 3.18, p < 0.01) and decreased with increasing silvicultural management intensity for acoustic (explained deviance = 30.7%, F = 6.1, p < 0.001) and morphometric trait dimensions (explained deviance = 15.9%, edf = 1.68, F = 5.63, p < 0.05), while the residual correlation strength for ecological and behavioral traits remained unaffected by increased silvicultural management intensity (explained deviance = 26.7%, edf = 1.9, F = 0.779, p > 0.05).
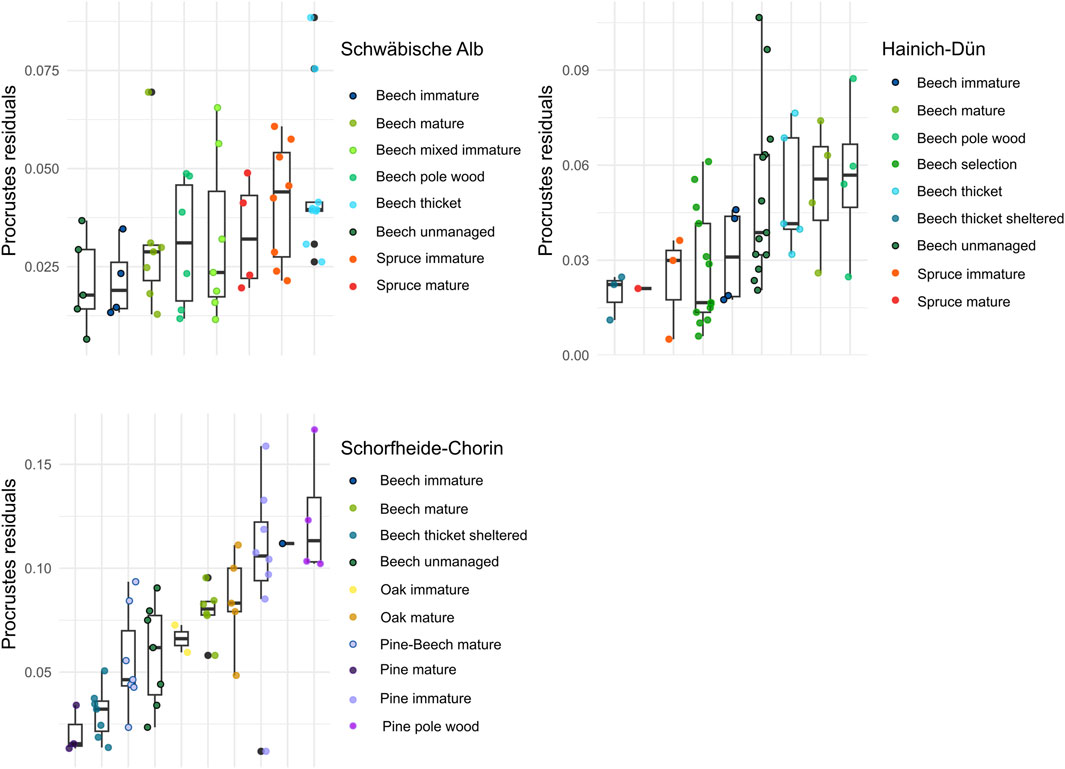
Figure 7. Differences in Procrustes residuals for ecological and behavioral trait dimension between forest management classes in the three different regions. Smaller residuals reflect a stronger interdependence of species and trait composition.
In grasslands, bird species compositional changes (adonis F = 2.09, p < 0.001) differed significantly between grassland sites managed as pastures, meadows, and mown pastures (Figures 5, 8). Furthermore, the overall functional composition revealed significant differences between grassland management categories [adonis (traits): F = 2.17, p < 0.05]; however, this overall result only reflected significant differences in ecological and behavioral trait composition (R = 2.62, p < 0.001), while acoustic (R = 1.71, p > 0.05) and morphological composition (R = 2.51, p > 0.05) did not significantly differ between management categories.
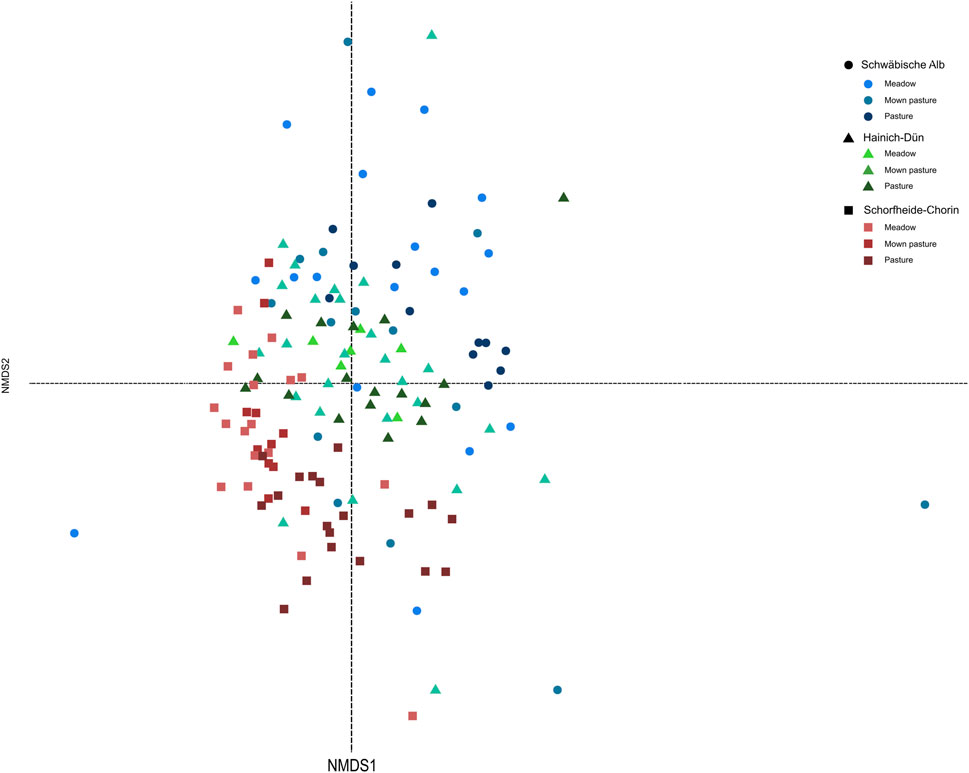
Figure 8. Non-metric multidimensional scaling of bird assemblages in grassland based on differences in species composition. Color reflects different management categories meadows, mown pastures, and pastures; shape reflects different regions, i.e., Alb = circle, Hainich = triangle, and Schorfheide = square.
Changes in bird species composition were highly correlated with changes in the composition of ecological and behavioral traits (84%, Procrustes m2 = 0.42, p < 0.001), highlighting that constraints due to ecological or behavioral requirements predominantly affect species assemblage composition in grasslands. Meanwhile, changes in acoustic composition (67%, m2 = 0.56, p < 0.001) and morphological composition were less correlated (26%, m2 = 0.9301, p < 0.01).
The residual correlation strength derived from the procrustean correlation between species and functional composition in grasslands did not differ between regional management categories (ANOVA: morphology: F = 1.4, p > 0.05; eco–behavioral: F = 1.8, p > 0.05; and acoustic: F = 1.6, p > 0.05). The local land use intensity neither revealed a significant effect on the interdependence of species composition and functional composition, indicating that changes in species composition due to local land use intensity (predominantly by reducing species diversity) in agricultural grasslands are followed by a similar change in functional composition across trait spaces. However, the residual correlation strength decreased with an increase in woody vegetation located near our grassland sites (Table 3), and this was consistent across ecological and behavioral (explained deviance = 24.9%, R2: 0.19; woody vegetation: F = 5.2, p < 0.001), acoustic (explained deviance = 22.3%, R2: 0.17; woody vegetation: F = 2.99, p < 0.05), and morphometric trait dimensions (explained deviance = 23.7%, R2: 0.18, edf: 1.5, woody vegetation: F = 3.58, p < 0.05), indicating an increased variance in bird and functional trait composition.
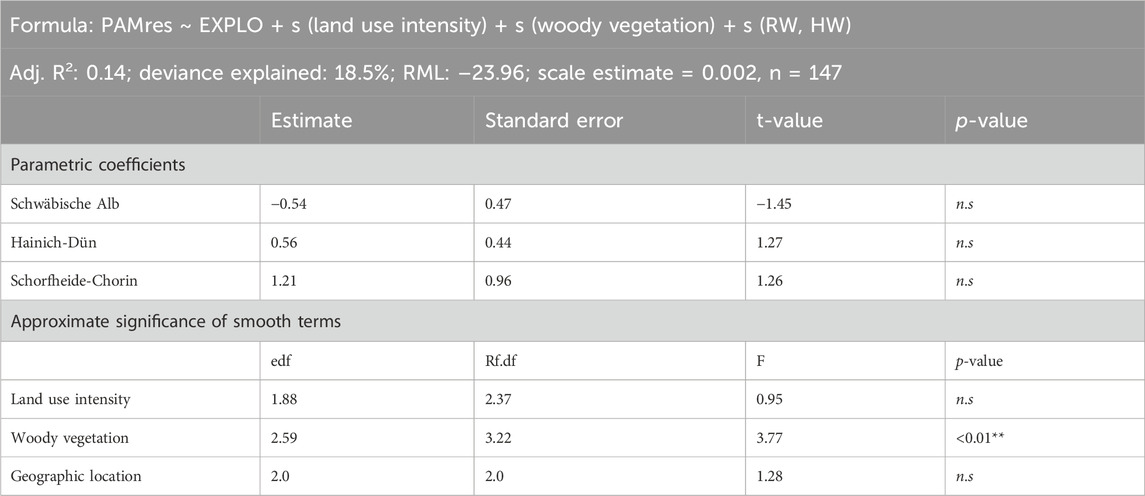
Table 3. Model specifications and results for grasslands, assessing the relative importance of land use intensity and the amount of woody vegetation on the correlative strength of species and functional composition across trait dimensions.
Birds are known to react sensitively to environmental pressures, and their population numbers and distribution patterns can reflect environmental changes in ecosystems (Morelli et al., 2021; Reif et al., 2023), including changes in other animal and plant populations. Understanding the ecological mechanisms responsible for bird species response to land use can thus help support biodiversity and ecosystem function in anthropogenic landscapes.
Our study identified how local land management affects the diversity and functional diversity of bird assemblages, whether local land management causes ecological sorting, and if so, which processes drive bird diversity and assemblage composition across different regions and the two major habitats, grassland and forest ecosystems. Such knowledge is essential to target conservation aims effectively and address respective stakeholders on how to maintain bird diversity and associated ecosystem function in anthropogenic landscapes.
Our data are based on an extensive dataset of bird species occurrence from 6 years in differently managed forest and grassland areas across three regions in Germany, which differ in climate and land use practices, and where bird occurrence data could be linked to very detailed and annually updated environmental information for management practices and local land use intensity. This is of particular importance as management applications and intensity of land use, especially in grasslands, may change annually, and neglecting such changes may mask the effects of local land use as an environmental driver for bird species occurrence in open land.
Our results highlight that the ecosystem (grassland/forest) and regional context must be considered to understand how management practices affect bird diversity and composition and emphasize that the importance of drivers for bird diversity differs between forest and grassland areas. This is in accordance with the results obtained by Vaccaro and Filloy (2022), who demonstrated that factors determining bird diversity and assemblage composition very much depend on the biome.
In forests, our results on species diversity patterns and functional diversity metrics suggested that within species-rich assemblages of differently managed forests, individual species tended to increasingly share functional traits, pointing toward ecological sorting. Interestingly, this pattern was especially pronounced for ecological and behavioral traits, while acoustic traits did not reveal an increased trait convergence, pointing toward the need to maintain acoustic identity and ensure biotic interactions in species-rich assemblages.
In contrast, assemblages in grasslands were rather species-poor or dominated by a single species and very limited functional diversity. An increase in species here led to an increase in functional dispersion and evenness, indicating a cumulative population of potentially available niches across trait dimensions and thus suggesting at the same time that in the managed grassland areas, higher bird diversity is limited by the availability of different potentially available niche spaces.
Within both forests and grasslands, bird diversity was significantly affected by local management.
In forests, bird diversity differed significantly between forest management classes, with generally lower species diversity in younger forests (e.g., beach and pine pole wood) and higher species diversity in mature and more structurally heterogeneous forest stands. The overall silvicultural management intensity across regions, however, did not reveal a significant effect on bird diversity. We argue that this is due to regional differences in forest management practices with intensively managed and homogenous pine pole wood stands in the Schorfheide-Chorin region, and the fine-grained uneven-aged but relatively intense management practices in beech stands of the Hainich-Dün region (Schall et al., 2018). This corroborates with previous findings and emphasizes that increased within-stand heterogeneity (Heidrich et al., 2020) promotes bird diversity and increases overall species richness by providing a high variety of different resources and potential nesting sites.
Bird diversity in grasslands did not differ between regional management categories but decreased significantly (across management categories) with local land use intensity. The strength of this effect, however, differed between regions and revealed stronger negative effects of local land use intensity in the Schwäbische Alb plateau and the Hainich-Dün region but not in the Schorfheide-Chorin region, where grassland sites are all located within long-established biosphere reserves, barely fertilized or mown and predominantly used for cattle grazing (Vogt et al., 2019), highlighting that regional differences in management practices can mask land use intensity effects on biodiversity patterns.
Habitat-specific analysis confirmed significant differences in species and functional composition between forest management categories and pointed toward ecological sorting, especially with respect to ecological requirements and biotic interactions. These results highlight that local forest management decisions with respect to tree composition and forest structure strongly affect both bird species composition and ecological functions. Similar results were reported by Leidinger et al. (2021), who showed that admixing tree species determines forest diversity by combining habitat heterogeneity effects and tree species-specific associations. This seems to be especially true for birds, which very much rely on local environmental conditions for nesting and feeding (Charbonnier et al., 2016) and where acoustic cues are used to attract mating partners and defend territories.
Our results further indicated that the correlative strength between species and functional composition varies between forest management classes and weakened with higher silvicultural management intensity, suggesting that bird assemblages in more intensely managed younger forest stands such as pine and beech pole wood are more randomly assembled compared to heterogeneous and/or mature forests. This is likely a combined effect of reduced availability of necessary resources at the local scale and spill-over effects from nearby mature stands, where optimal mating territories after remigration in spring are already occupied by more successful conspecifics. This corroborates with previous findings that demonstrated that younger forest stands reveal a greater year-to-year turnover of species, and heterogeneous forest management in the surrounding forest matrix positively affects locally determined results on species richness in younger stands (Wells et al., 2011).
In grasslands, constraints due to ecological or behavioral requirements of individual bird species greatly affected species assemblage composition and indicated the importance of considering environmental conditions beyond the local scale. This is of particular importance for grassland birds, which integrate habitat conditions over a wider area (Concepción et al., 2015). Although increased local land use intensity resulted in an equal change in species and functional composition, predominantly by reducing species diversity, our results suggested a positive effect of nearby woody vegetation such as single trees and hedgerows, maintaining ecological function. This supports the general call to enhance landscape heterogeneity (Stanton et al., 2018) by incorporating structural landscape elements such as hedges, paddock trees, or live fence to benefit the overall bird diversity (Concepción et al., 2015) and ecosystem service provisioning (e.g., Smith et al., 2022) and to counteract biotic homogenization (Gámez-Virués et al., 2015) within agricultural areas. However, we emphasize that focusing conservation efforts in grasslands only on increasing bird species diversity could neglect ecological requirements of open grassland specialists such as the European lapwing (Vanellus vanellus), the Eurasian skylark (Alauda arvensis), and yellow wagtail (Motacilla flava), which are all exclusive ground-nesting birds that prefer open landscapes (Borges et al., 2017; Püttmanns et al., 2021; Buschmann et al., 2023) and, thus, are directly affected by management applications (e.g., mowing and fertilizing), which often temporally overlap with their breeding times. This emphasizes that one single strategy for bird conservation does not exist, and ensuring both overall diversity and the persistence of grassland specialists needs to incorporate local conservation efforts for individual species into overall biodiversity conservation aims.
The European Biodiversity Strategy for 2030 aims to put Europe’s biodiversity on a path to recovery by 2030 by protecting nature and reversing the degradation of ecosystems (EC 2020), of which more than half are managed as agricultural and forested areas. Bird populations and assemblage composition reflect environmental conditions and changes in other animal and plant populations across different spatial scales and provide important ecosystem function and services. Understanding the mechanisms driving bird diversity and assemblage composition in managed ecosystems may thus benefit overall biodiversity conservation, help target conservation aims, and address respective stakeholders on how to maintain bird diversity and associated ecosystem functions in anthropogenic landscapes.
Our results emphasize that the ecosystem and regional context must be considered to understand how management practices affect bird diversity and composition. By including functional traits and understanding ecological mechanisms, however, local management decisions can be directed toward supporting bird diversity and ecological function. In particular, local forest management decisions with respect to tree composition and forest structure strongly affected both bird species composition and ecological function. In addition, higher structural heterogeneity generally benefitted the taxonomic and functional diversity of birds also in nearby, intensely managed, younger forest stands, suggesting that a mosaic of differently managed forests (e.g., Penone et al., 2019) with respect to tree species and age classes supports bird diversity and ecosystem functions within production forests. In grasslands, predominantly, constraints due to ecological or behavioral requirements of individual bird species greatly affect the bird species occurrence and species assemblage composition. Our results thus highlight the importance of maintaining extensively managed areas for grassland specialist birds [e.g., Rigal et al. (2023)] and support the general call for increased landscape heterogeneity by maintaining landscape elements such as trees and hedgerows, which benefit the overall bird diversity.
Finally, we argue that supporting bird diversity and ecological function in managed grassland and forest systems needs an integrative approach considering regional differences in species distributions and management applications and local consequences of management strategies affecting species-specific habitats and resource requirements. In particular, we emphasize on a view beyond the local scale of management units and a joint effort of biodiversity conservationists and land managers to create a connected network of different habitats within production landscapes, to ensure both the conservation of overall biodiversity and individual species with very specialized requirements in designated areas. We argue that a regionally focused management strategy for biodiversity is important to protect nature, guard against biotic and functional homogenization, and prevent the degradation of ecosystems in production landscapes.
This work is based on data elaborated by the Vertebrate (193945537) and Local Management project (193990583) of the Biodiversity Exploratories program (DFG Priority Program 1374). Most datasets are publicly available in the Biodiversity Exploratories Information System (http://doi.org/10.17616/R32P9Q) and listed in the References section (Weisser et al., 2016, 2018; Tschapka et al., 2018, 2020a, 2020b, 2020c; Jung et al., 2020; Fischer, 2023; Lorenzen et al. 2023; Magdon and Wöllauer, 2023; Ostrowski and Petzold, 2023). The raw trait data-sets for morphometric and ecological and behavioral available from Avonent (2022), acoustic audio files are publicly available under a creative common license agreement from Xeno canto (https://xeno-canto.org/), and the Animal Sound archive- at the Museum für Naturkunde Berlin (https://www.tierstimmenarchiv.de).
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because passive monitoring surveys were used to obtain bird occurrence data.
KJ: conceptualization, data curation, formal analysis, funding acquisition, investigation, writing–original draft, writing–review and editing, and methodology. MT: conceptualization, data curation, and writing–review and editing. SB: conceptualization, data curation, validation, and writing–review and editing. KW: conceptualization, data curation, and writing–review and editing. MA: funding acquisition, resources, writing–review and editing. MF: funding acquisition, resources, writing–review and editing. WWW: funding acquisition, resources, and writing–review and editing. SR: conceptualization, data curation, formal analysis, validation, and writing–review and editing. MT: conceptualization, funding acquisition, resources, validation, and writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was funded by the DFG Priority Program 1374 “Biodiversity Exploratories” (DFG Project numbers: 193945537; 433326865; and 193990583).
The authors dedicate this work to the memory of Elisabeth Kalko and her passion for birds and bats and their acoustic diversity. The authors thank the field assistants for their hard work. The authors thank the managers of the three exploratories: Andreas Hemp, Martin Gorke (Sch), Sonja Gockel, Kerstin Wiesner, Katrin Lorenzen, Juliane Vogt (Hai), Iris Steitz, Sandra Weitmann, and Florian Straub (Alb) and the local management team members for their constant support and their work in maintaining the plot and project infrastructure. The authors thank Simone Pfeiffer, Maren Gleisberg, Christiane Fischer, and Jule Mangels for their support through the central office; Jens Nieschulze, Michael Owonibi, and Andreas Ostrowski for managing the central database; and Juliane Balmer and Andreas Ostrowski for special care with the bird data. The authors thank Eduard Linsenmair, Dominik Hessenmöller, Daniel Prati, Ingo Schöning, François Buscot, Ernst-Detlef Schulze, and the late Elisabeth Kalko for their role in setting up the Biodiversity Exploratories project. The authors thank the administration of the Hainich National Park, the UNESCO Biosphere Reserve Swabian Alb, and the UNESCO Biosphere Reserve Schorfheide-Chorin, as well as all land owners for the excellent collaboration. Field work permits were issued by the responsible state environmental offices of Baden-Württemberg, Thüringen, and Brandenburg.
At the end of this study, Author SB was employed by Sieber Consult.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2024.1401513/full#supplementary-material
Ali, J. R., Blonder, B. W., Pigot, A. L., and Tobias, J. A. (2022). Bird extinctions threaten to cause disproportionate reductions of functional diversity and uniqueness. Funct. Ecol. 29, 162–175. doi:10.1111/1365-2435.14201
Blüthgen, N., Dormann, C. F., Prati, D., Klaus, V. H., Kleinebecker, T., Hölzel, N., et al. (2012). A quantitative index of land-use intensity in grasslands: integrating mowing, grazing and fertilization. Basic Appl. Ecol. 13 (3), 207–220. doi:10.1016/j.baae.2012.04.001
Borges, F., Glemnitz, M., Schultz, A., and Stachow, U. (2017). Assessing the habitat suitability of agricultural landscapes for characteristic breeding bird guilds using landscape metrics. Environ. Monit. Assess. 189, 166. doi:10.1007/s10661-017-5837-2
Bowler, D. E., Heldbjerg, H., Fox, A. D., de Jong, M., and Böhning-Gaese, K. (2019). Long-term declines of European insectivorous bird populations and potential causes. Conserv. Biol. 33 (5), 1120–1130. doi:10.1111/cobi.13307
Bowler, D. E., Richter, R. L., Eskildsen, D., Kamp, J., Moshøj, C. M., Reif, J., et al. (2021). Geographic variation in the population trends of common breeding birds across central Europe. Basic Appl. Ecol. Prepr. 56, 72–84. doi:10.1016/j.baae.2021.07.004
Burns, F., Eaton, M. A., Burfield, I. J., Klvaňová, A., Šilarová, E., Staneva, A., et al. (2021). Abundance decline in the avifauna of the European Union reveals cross-continental similarities in biodiversity change. Biodivers. change’ 11, 16647–16660. doi:10.1002/ece3.8282
Buschmann, C., Böhner, H. G., and Röder, N. (2023). The cost of stabilising the German lapwing population: a bioeconomic study on lapwing population development and distribution using a cellular automaton. J. Nat. Conservation 71, 126314. doi:10.1016/j.jnc.2022.126314
Cadotte, M. W., and Tucker, C. M. (2017). Should Environmental Filtering be Abandoned? Trends Ecol. Evol. 32 (6), 429–437. doi:10.1016/j.tree.2017.03.004
Carmona, C. P., Tamme, R., Pärtel, M., de Bello, F., Brosse, S., Capdevila, P., et al. (2021). Erosion of global functional diversity across the tree of life. Sci. Adv. 7 (13), eabf2675. doi:10.1126/sciadv.abf2675
Ceballos, G., Ehrlich, P. R., Barnosky, A. D., García, A., Pringle, R. M., and Palmer, T. M. (2015). Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 1 (5), e1400253. doi:10.1126/sciadv.1400253
Charbonnier, Y. M., Barbaro, L., Barnagaud, J. Y., Ampoorter, E., Nezan, J., Verheyen, K., et al. (2016). Bat and bird diversity along independent gradients of latitude and tree composition in European forests. Oecologia 182, 529–537. doi:10.1007/s00442-016-3671-9
Concepción, E. D., Moretti, M., Altermatt, F., Nobis, M. P., and Obrist, M. K. (2015). Impacts of urbanisation on biodiversity: the role of species mobility, degree of specialisation and spatial scale. Oikos 124 (12), 1571–1582. doi:10.1111/oik.02166
Ellis, E. C., Gauthier, N., Klein Goldewijk, K., Bliege Bird, R., Boivin, N., Díaz, S., et al. (2021). People have shaped most of terrestrial nature for at least 12,000 years. PNAS 17 (118), e2023483118. doi:10.1073/pnas.2023483118
Federal Statistical Office (2022). Data on land cover in Germany. Statistisches Bundesamt (Destatis).
Fischer, M., Bossdorf, O., Gockel, S., Hänsel, F., Hemp, A., Hessenmöller, D., et al. (2010). Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 11, 473–485. doi:10.1016/j.baae.2010.07.009
Fischer, M. (2023). Bird survey and trait data on all grassland and forest EPs 2018. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/31521?version=2.
Foley, J. A., DeFries, R., Asner, G. P., Barford, C., Bonan, G., Carpenter, S. R., et al. (2005). Global Consequences of Land Use. Science 309 (5734), 570–574. doi:10.1126/science.1111772
Gámez-Virués, S., Perović, D. J., Gossner, M. M., Börschig, C., Blüthgen, N., de Jong, H., et al. (2015). Landscape simplification filters species traits and drives biotic homogenization. Nat. Commun. 6 (1), 8568. doi:10.1038/ncomms9568
Hedblom, M., Heyman, E., Antonsson, H., and Gunnarsson, B. (2014). Bird song diversity influences young people’s appreciation of urban landscapes. Urban For. Urban Green. 13, 469–474. doi:10.1016/j.ufug.2014.04.002
Heidrich, L., Bae, S., Levick, S., Seibold, S., Weisser, W., Krzystek, P., et al. (2020). Heterogeneity–diversity relationships differ between and within trophic levels in temperate forests. Nat. Ecol. Evol. 4 (9), 1204–1212. doi:10.1038/s41559-020-1245-z
IPBES (2019) Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Bonn, Germany: IPBES Secretariat. doi:10.5281/zenodo.3831673
Johnson, R. J., Jedlicka, J. A., Quinn, J. E., and Brandle, J. R. (2011). Global Perspectives on Birds in Agricultural Landscapes. Integrating Agric. Conservation Ecotourism Examples Field 1, 55–140. doi:10.1007/978-94-007-1309-3_3
Joppa, L. N., O'Connor, B., Visconti, P., Smith, C., Geldmann, J., Hoffmann, M., et al. (2016). Filling in biodiversity threat gaps. Science 352 (6284), 416–418. doi:10.1126/science.aaf3565
Jung, K., Renner, S., and Tschapka, M. (2020). ‘Bird survey data 2012, all 300 EPs’. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/24690?version=2.
Kraft, N. J. B., Adler, P. B., Godoy, O., James, E. C., Fuller, S., and Levine, J. M. (2015). Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 29 (5), 592–599. doi:10.1111/1365-2435.12345
Laliberté, E., and Legendre, P. (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology 91 (1), 299–305. doi:10.1890/08-2244.1
Laliberté, E., Legendre, P., and Shipley, B. (2022). Package FD: measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0-12.3.
Leidinger, J., Blaschke, M., Ehrhardt, M., Fischer, A., Gossner, M. M., Jung, K., et al. (2021). Shifting tree species composition affects biodiversity of multiple taxa in Central European forests. For. Ecol. Manag. 498, 119552. doi:10.1016/j.foreco.2021.119552
Leroy, F., Reif, J., Storch, D., and Keil, P. (2023). How has bird biodiversity changed over time? A review across spatio-temporal scales. Basic Appl. Ecol. 69, 26–38. doi:10.1016/j.baae.2023.03.004
Lisboa, F. J. G., Peres-Neto, P. R., Chaer, G. M., Jesus, E. d. C., Mitchell, R. J., Chapman, S. J., et al. (2014). Much beyond Mantel: Bringing Procrustes Association Metric to the Plant and Soil Ecologist’s Toolbox. PLOS ONE 9 (6), e101238. doi:10.1371/journal.pone.0101238
Lorenzen, K., Juliane, V, Miriam, T., Andreas, O., and Jan, T. (2023). Input Data for LUI Calculation Tool of all grassland plots since 2006 - revised 2019. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/25086?version=11.
Luther, D. (2009). The influence of the acoustic community on songs of birds in a neotropical rain forest. Behav. Ecol. 20 (4), 864–871. doi:10.1093/beheco/arp074
Magdon, P., and Wöllauer, S. (2023). Digital orthophotos (DOP-20) of all Biodiversity Exploratories regions. Available at: https://www.bexis.uni-jena.de.
Mason, N. W. H., Mouillot, D., Lee, W. G., and Wilson, J. B. (2005). Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos 111, 112–118. doi:10.1111/j.0030-1299.2005.13886.x
Methorst, J., Rehdanz, K., Mueller, T., Hansjürgens, B., Bonn, A., and Böhning-Gaese, K. (2021). The importance of species diversity for human well-being in Europe. Ecol. Econ. 181, 106917. doi:10.1016/j.ecolecon.2020.106917
Morelli, F., Reif, J., Díaz, M., Tryjanowski, P., Ibáñez-Álamo, J. D., Suhonen, J., et al. (2021). Top ten birds indicators of high environmental quality in European cities. Ecol. Indic. 133, 108397. doi:10.1016/j.ecolind.2021.108397
Mouchet, M. A., Villéger, S., Mason, N. W. H., and Mouillot, D. (2010). Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 24, 867–876. doi:10.1111/j.1365-2435.2010.01695.x
Newbold, T., Hudson, L. N., Hill, S. L. L., Contu, S., Lysenko, I., Senior, R. A., et al. (2015). Global effects of land use on local terrestrial biodiversity. Nature 520 (7545), 45–50. doi:10.1038/nature14324
Öckinger, E., Schweiger, O., Crist, T. O., Debinski, D. M., Krauss, J., Kuussaari, M., et al. (2010). Life-history traits predict species responses to habitat area and isolation: a cross-continental synthesis: Habitat fragmentation and life-history traits. Ecol. Lett. 13, 969–979. doi:10.1111/j.1461-0248.2010.01487.x
Oksanen, J., Simpson, G., Blanchet, F., Kindt, R., Legendre, P., Minchin, P., et al. (2022). Vegan: Community Ecology Package. R package version 2.6-4. Available at: https://CRAN.R-project.org/package=vegan.
Ostrowski, A., and Petzold, E. (2023). Coordinates of all meteorological stations of all EP plots. Available at: https://www.bexis.uni-jena.de.
Penone, C., Allan, E., Soliveres, S., Felipe-Lucia, M. R., Gossner, M. M., Seibold, S., et al. (2019). Specialisation and diversity of multiple trophic groups are promoted by different forest features. Ecol. Lett. 22, 170–180. doi:10.1111/ele.13182
Pigot, A. L., Sheard, C., Miller, E. T., Bregman, T. P., Freeman, B. G., Roll, U., et al. (2020). Macroevolutionary convergence connects morphological form to ecological function in birds. Nat. Ecol. Evol. 4 (2), 230–239. doi:10.1038/s41559-019-1070-4
Püttmanns, M., Böttges, L., Filla, T., Lehmann, F., Martens, A. S., Siegel, F., et al. (2021). Habitat use and foraging parameters of breeding Skylarks indicate no seasonal decrease in food availability in heterogeneous farmland. Ecol. Evol. 12, 8461. doi:10.1002/ece3.8461
Rayner, J. M. V. (1988). “Form and Function in Avian Flight,” in Current Ornithology. Editor R. F. Johnston (Boston, MA: Springer US), 1–66. doi:10.1007/978-1-4615-6787-5_1
Reif, J., Koleček, J., Morelli, F., and Benedetti, Y. (2023). Population trends of ground-nesting birds indicate increasing environmental impacts from Eastern to Western Europe: different patterns for open-habitat and woodland species. Front. Environ. Sci. 11, 1156360. doi:10.3389/fenvs.2023.1156360
Reif, J., and Vermouzek, Z. (2019). Collapse of farmland bird populations in an Eastern European country following its EU accession. Conserv. Lett. 12, e12585. doi:10.1111/conl.12585
Rigal, S., Dakos, V., Alonso, H., Auniņš, A., Benkő, Z., Brotons, L., et al. (2023). Farmland practices are driving bird population decline across Europe. Proc. Natl. Acad. Sci. 120 (21), e2216573120. doi:10.1073/pnas.2216573120
Rigal, S., Devictor, V., Gaüzère, P., Kéfi, S., Forsman, J. T., Kajanus, M. H., et al. (2022). Biotic homogenisation in bird communities leads to large-scale changes in species associations. Oikos 2022 (3). doi:10.1111/oik.08756
Schall, P., and Ammer, C. (2013). How to quantify forest management intensity in Central European forests. Eur. J. For. Res. 132, 379–396. doi:10.1007/s10342-013-0681-6
Schall, P., Gossner, M. M., Heinrichs, S., Fischer, M., Boch, S., Prati, D., et al. (2018). The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 55 (1), 267–278. doi:10.1111/1365-2664.12950
Schleuning, M., García, D., and Tobias, J. A. (2022). Animal functional traits: Towards a trait-based ecology for whole ecosystems. Funct. Ecol. 37, 4–12. doi:10.1111/1365-2435.14246
Schug, F., Frantz, D., Okujeni, A., van der Linden, S., and Hostert, P. (2020). Mapping urban-rural gradients of settlements and vegetation at national scale using Sentinel-2 spectral-temporal metrics and regression-based unmixing with synthetic training data. Remote Sens. Environ. 246, 111810. doi:10.1016/j.rse.2020.111810
Smith, O. M., Kennedy, C. M., Echeverri, A., Karp, D. S., Latimer, C. E., Taylor, J. M., et al. (2022). Complex landscapes stabilize farm bird communities and their expected ecosystem services. J. Appl. Ecol. 59, 927–941. doi:10.1111/1365-2664.14104
Stanton, R. L., Morrissey, C. A., and Clark, R. G. (2018). Analysis of trends and agricultural drivers of farmland bird declines in North America: A review. Agric. Ecosyst. Environ. 254, 244–254. doi:10.1016/j.agee.2017.11.028
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann, M. C., Schwager, M., et al. (2004). Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J. Biogeogr. 31 (1), 79–92.
Tilman, D., Clark, M., Williams, D. R., Kimmel, K., Polasky, S., and Packer, C. (2017). Future threats to biodiversity and pathways to their prevention. Nature 546 (7656), 73–81. doi:10.1038/nature22900
Tobias, J. A., Ottenburghs, J., and Pigot, A. L. (2020). Avian Diversity: Speciation, Macroevolution, and Ecological Function. Annu. Rev. Ecol. Evol. Syst. 51 (1), 533–560. doi:10.1146/annurev-ecolsys-110218-025023
Tobias, J. A., Seddon, N., Spottiswoode, C. N., Pilgrim, J. D., Fishpool, L. D. C., and Collar, N. J. (2010). Quantitative criteria for species delimitation. Ibis 152 (4), 724–746. doi:10.1111/j.1474-919X.2010.01051.x
Tobias, J. A., Sheard, C., Pigot, A. L., Devenish, A. J. M., Yang, J., Sayol, F., et al. (2022). AVONET: morphological, ecological and geographical data for all birds. Ecol. Lett. 25 (3), 581–597. doi:10.1111/ele.13898
Tschapka, M., Renner, S., and Jung, K. (2018). ‘Bird survey data 2008, all 300 EPs’. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/21446?version=2.
Tschapka, M., Renner, S., and Jung, K. (2020a). ‘Bird survey data 2009, all 300 EPs’. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/21447?version=2.
Tschapka, M., Renner, S., and Jung, K. (2020b). ‘Bird survey data 2010, all 300 EPs’. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/21448?version=2.
Tschapka, M., Renner, S., and Jung, K. (2020c). ‘Bird survey data 2011, all 300 EPs’. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de/ddm/data/Showdata/21449?version=2.
Vaccaro, A. S., and Filloy, J. (2022). Factors underlying bird community assembly in anthropogenic habitats depend on the biome. Sci. Rep. 12, 19804. doi:10.1038/s41598-022-24238-x
Villéger, S., Mason, N. W. H., and Mouillot, D. (2008). New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89 (8), 2290–2301. doi:10.1890/07-1206.1
Vogt, J., Klaus, V., Both, S., Fürstenau, C., Gockel, S., Gossner, M., et al. (2019). Eleven years’ data of grassland management in Germany. Biodivers. Data J. 7, e36387. doi:10.3897/BDJ.7.e36387
Weisser, W., Ayasse, M., and Fischer, M. (2016). ‘Grassland survey for landuse identification on all EPs, 2006 - 2009’. Biodiversity Exploratories Information System. Available at: https://www.bexis.uni-jena.de.
Weisser, W., Ayasse, M., and Fischer, M. (2018). Grassland survey for landuse identification on all EPs 2010 - 2016. Biodivers. Explor. Inf. Syst. Available at: https://www.bexis.uni-jena.de.
Wells, K., Böhm, S. M., Boch, S., Fischer, M., and Kalko, E. K. (2011). Local and landscape-scale forest attributes differ in their impact on bird assemblages across years in forest production landscapes. Basic Appl. Ecol. 12 (2), 97–106. doi:10.1016/j.baae.2011.01.002
Keywords: birds, functional traits, land use effects, grasslands and forest management, biodiversity conservation
Citation: Jung K, Teuscher M, Böhm S, Wells K, Ayasse M, Fischer M, Weisser WW, Renner SC and Tschapka M (2024) Supporting bird diversity and ecological function in managed grassland and forest systems needs an integrative approach. Front. Environ. Sci. 12:1401513. doi: 10.3389/fenvs.2024.1401513
Received: 18 March 2024; Accepted: 02 July 2024;
Published: 01 August 2024.
Edited by:
Chong Jiang, Guangdong Academy of Science (CAS), ChinaReviewed by:
Yixin Wang, Hohai University, ChinaCopyright © 2024 Jung, Teuscher, Böhm, Wells, Ayasse, Fischer, Weisser, Renner and Tschapka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kirsten Jung, a2lyc3Rlbi5qdW5nQHVuaS11bG0uZGU=
†ORCID: Kirsten Jung, orcid.org/0000-0002-9449-2215; Miriam Teuscher, orcid.org/0000-0002-8701-4477; Stefan Böhm, orcid.org/0009-0005-7416-6606; Konstans Wells, orcid.org/0000-0003-0377-2463; Manfred Ayasse, orcid.org/0000-0001-9914-6269; Markus Fischer, orcid.org/0000-0002-5589-5900; Wolfgang Weisser, orcid.org/0000-0002-2757-8959; Swen Renner, orcid.org/0000-0002-6893-4219; Marco Tschapka, orcid.org/0000-0001-9511-6775
‡These authors share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.