 Jiří Reif
Jiří Reif Petr Chajma
Petr Chajma Lenka Dvořáková1,4
Lenka Dvořáková1,4 Pavel Marhoul
Pavel Marhoul Tomáš Kadlec
Tomáš Kadlec- 1Faculty of Science, Institute for Environmental Studies, Charles University, Prague, Czechia
- 2Department of Zoology, Faculty of Science, Palacký University in Olomouc, Olomouc, Czechia
- 3Department of Ecology, Faculty of Environmental Sciences, Czech University of Life Sciences Prague, Prague, Czechia
- 4Beleco, Z.S, Prague, Czechia
Introduction: Abandoned military training areas are biodiversity strongholds, and this is particularly true for open-habitat and threatened species in Central Europe. Such species benefited from a specific disturbance regime created by military activities that maintained small-grained environmental heterogeneity. However, the disturbance regime no longer occurs after abandonment and the biodiversity is at risk due to forest and shrub encroachment if the areas are left unmanaged. To combat these adverse changes, several management options are used. As these options are not always applied for conservation purposes and substantially differ in their implementation, it is essential to assess their impacts on biodiversity.
Methods: We performed repeated standardized surveys (first in 2009–2010, second in 2020–2022) of vascular plants, grasshoppers, butterflies and birds in 42 abandoned military training areas in Czechia, a Central European country. We calculated changes of species richness and abundance between periods for each taxon and related these changes to six different management types (woody plant cutting, mowing for conservation, mowing for agriculture, grazing for conservation, grazing for agriculture, vehicle movement) performed in these areas between periods.
Results: Vascular plants and grasshoppers showed generally positive changes, whereas the reverse was true for butterflies, and birds experienced mixed changes. Although beta-diversity increased between periods in plants, grasshoppers and butterflies, this increase was driven by extirpation of common species. Management impacts greatly different between respective types and between taxa. Woody plant cutting showed solely positive impacts (on plants and grasshoppers), while the impacts of both types of grazing were mixed (positive on plants and birds, negative on butterflies, mixed on grasshoppers). Mowing for agriculture supported plants and birds but had negative effects on grasshoppers. Mowing for conservation and vehicle movement were linked solely to negative biodiversity changes (former in plants, latter in butterflies).
Discussion: Some components of biodiversity, i.e. plants and grasshoppers, indicate that abandoned military training areas still serve as their strongholds and the management most likely contributes to this favourable state. In contrast, the pattern found for butterflies is worrying since the management performed up to now apparently does not meet their requirements, likely because they are based on smaller-scale habitat mosaic than currently occurs in the areas. Our results may serve as a guide for future prioritization of environmental management, and we urge for development of more nuanced approaches to save the butterflies.
Introduction
Biodiversity loss is one of the most serious global environmental problems (Chase et al., 2020). Such losses, e.g., the “insect apocalypse” (Goulson, 2019), could have pertinent impacts on the functioning of trophic relationships, with the overlap into human economy (Cardoso et al., 2020). In European lowlands, this loss is particularly severe in open habitats that have been maintained by extensive agriculture for millennia (Ellis et al., 2021). Such habitats are a part of European cultural landscape that is protected under the EU’s Habitats Directive in some regions (Maes et al., 2012). Traditional management maintained high species diversity in these landscapes including some Red List taxa (Spulerova et al., 2017). Nowadays, open habitats are rapidly deteriorating due to eutrophication, intensive agricultural use or, if not managed, they become encroached by shrubland or forest (Bardgett et al., 2021). It is therefore important to find solutions on how to stop the further deterioration of the biodiversity of the open habitats in Europe.
One solution is offered by the areas with exceptionally high biodiversity of open habitat species. Such areas may serve as regional biodiversity refuges—they maintain the level of regional diversity in a plausible state and provide propagules for the potential re-colonization of the surrounding landscape (Habel et al., 2013; Tropek et al., 2013; Šálek et al., 2022). In this respect, abandoned military training areas play an important role (Zentelis and Lindenmayer, 2015). They have been widely recognized as regional open habitat biodiversity refuges, supporting a high number of threatened species and showing species richness values comparable to strictly protected nature reserves (Reif et al., 2011; Čížek et al., 2013; Bušek and Reif, 2017). Conservation of these biodiversity refuges is therefore of the utmost importance.
At the same time, abandoned military training areas are exposed to various threats that have arisen after the end of military activity (Herčik et al., 2014; Ellwanger and Reiter, 2019). Although these areas were saved from transformation to intensive forms of cultural landscape for a long time, they may recently face habitat loss due to construction activities or conversion into intensively cultivated farmland, and the quality of their open habitats may deteriorate due to changes driven by ecological succession in the absence of management (Gaertner et al., 2010; Dvořáková et al., 2023). Therefore, to secure the key role of abandoned military training areas in supporting European open habitat biodiversity, it is important to implement the appropriate active management in these areas.
Up to now, abandoned military training areas have experienced several different management approaches. These include traditional management measures such as cutting of shrubs and trees to create open habitats and slow down the encroachment, as well as mowing and cattle grazing to mitigate grassland eutrophication and suppress nutrient-demanding competitive superior species (Ellwanger and Reiter, 2019). In some areas, these management approaches are carried out to obtain economic profit rather than for conservation purposes (Wang et al., 2014), but it is possible that biodiversity may still enjoys some benefits. Besides traditional management types, novel approaches have recently been introduced. These include semi-wild horse or cattle rewilding (Konvička et al., 2021; Dvorský et al., 2022), and prescribed or free-range rides of off-road and military vehicles (Jentsch et al., 2009). They may provide different types of conservation benefits but their impact on different taxa occurring in abandoned military areas remains unclear. Therefore, knowledge of ecological impacts of respective management types on the temporal biodiversity dynamics (sensu Volery et al., 2023) is urgently needed.
To fill this knowledge gap, we explore a unique dataset based on repeated biodiversity surveys in 42 abandoned military training areas in Czechia, a country in Central Europe. The surveys focused simultaneously on multiple taxa that differ in their lifestyle and trophic position: vascular plants—primary producers with low vagility, being highly sedentary and responding slowly to management (Diekmann, 2003); grasshoppers (species of the insect order Orthoptera)—primary and secondary consumers and dietary generalists, indicators of small-scale habitat changes (Fartmann et al., 2012); butterflies—primary consumers and important pollinators, short-lived species highly sensitive to changes in habitat quality (Warren et al., 2021); birds—secondary consumers, long-lived habitat generalists with high dispersal capacity (Fraixedas et al., 2020). Simultaneous focus on such different taxa can provide a complex insight into biodiversity responses to different types of management (Hilty and Merenlender, 2000).
In the focal abandoned military training areas, we conducted a baseline survey of the above-mentioned taxa in 2009–2010 (Reif et al., 2011; Čížek et al., 2013) and repeated the survey using the same methodology in 2020–2022. While the areas were unmanaged at time of the first survey, the above-listed management approaches were applied at multiple sites during the period between surveys. In this study, we express the change in representation of the respective taxa between surveys and relate these changes to the respective management types. Since each of the focal taxa contains numerous species with different ecological characteristics that may influence these relationships, we define several ecological groups of species within each taxon.
The aim of this study is to examine the relationships between biodiversity and different management approaches across taxa and species groups. We predict that species richness of all taxa will increase between survey periods because management application should create specific habitats that host unique species that elevate species richness. We further predict that the management should be most beneficial for species associated with open habitats across all taxa because creating and conserving such habitats is a general management target. In addition, we predict that the management types conducted for conservation purposes should result in more positive outcomes than the management types conducted for economic profit. Finally, we predict that invertebrate taxa, i.e., grasshoppers and butterflies, will be more responsive to the management types applied in the focal military training areas than plants and birds because invertebrates have faster life styles.
Materials and methods
Study area and study sites
We focused on 42 abandoned military training areas scattered throughout Czechia (Figure 1), a central European country situated in the northern temperate zone at the transition from Atlantic to continental climate. The size of the areas varied from 21 to 351 ha (mean 91 ha), and all were located at low to mid elevations (200–625 m asl., mean 364 m). Our selection of the 42 areas used in this study was based on their regional availability and forms a representative sample of all abandoned military training areas in Czechia (Reif et al., 2011).
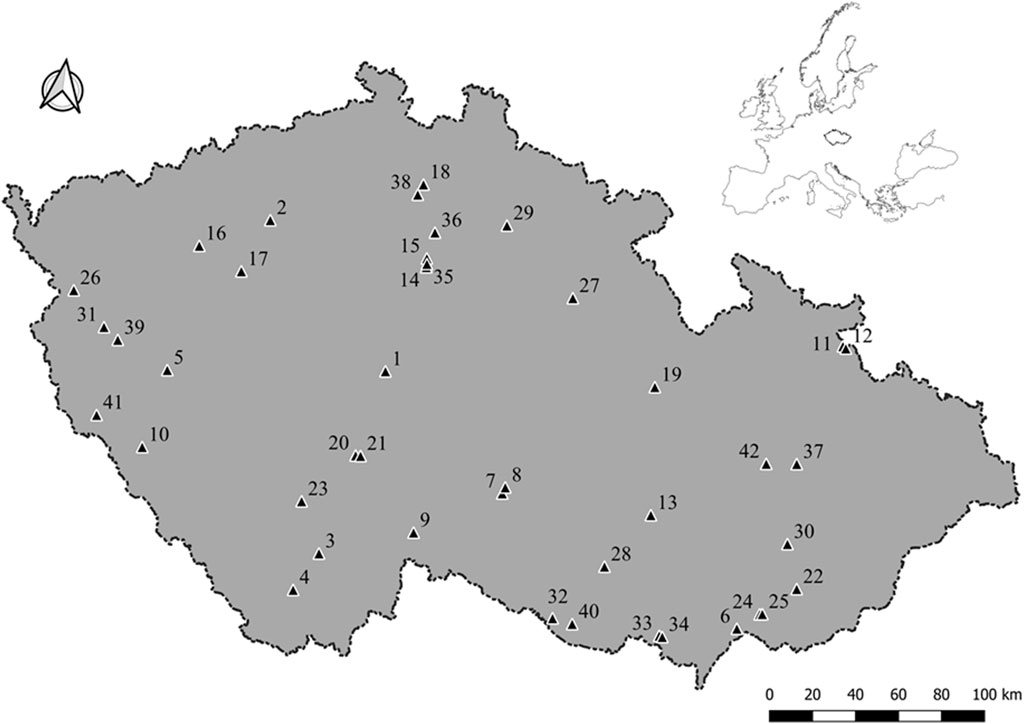
FIGURE 1. Map of the surveyed abandoned military training areas in Czechia. Inset shows position of Czechia in Europe.
The areas were used by the army until 1990s and then gradually abandoned due to the downsizing of troops after the end of the Cold War (Tagarev, 2004). Military activities created a heterogeneous fine-scale mosaic of different habitats that were maintained by disturbances (shell explosions, fires, rides of tanks and other vehicles, movement of troops) that were variable in space and time (Warren et al., 2007). After abandonment, the disturbance regime ceased, and the habitats underwent changes due to ecological succession (Skokanová et al., 2017). However, as previous disturbances varied greatly within the respective areas, habitat heterogeneity was still considerable even in 2009 (Čížek et al., 2013) and included a wide range of successional stages from sparsely vegetated bare ground (on the impacted sites being most intensively used by the army) through grassland with varying degrees of shrub encroachment, to woodland (on sites already covered by woody vegetation at the time when the areas were actively used for military training). Therefore, the areas retained considerable biodiversity even 10–15 years after abandonment (Reif et al., 2011; Čížek et al., 2013).
We mapped the habitat composition of each area prior to the respective biodiversity surveys were conducted, i.e., in early 2009 and 2020, respectively, discriminating five major habitat types: bare ground (sites where bare ground dominates over herbaceous vegetation), grassland (sites dominated by herbaceous vegetation without woody plants), sparse shrubland (sites where grassland is encroached by shrubs and small trees but the woody vegetation is discontinuous), dense shrubland (heavily encroached sites consisting of continuous shrub stands), and forest (tall trees with a continuous canopy). These habitat maps (1:2600) were used in subsequent biodiversity surveys.
Biodiversity surveys
We surveyed four major taxa, i.e., vascular plants (hereafter referred to as “plants”), species of the insect order Orthoptera (“grasshoppers”), diurnal butterflies and burnet moths of Zygaenidae family (“butterflies”), and birds in the respective areas in 2009–2010 (first period) and 2020–2022 (second period). The spread of the respective periods over several years was dictated by our capacity to cover all taxa in all areas, with no annually repeated surveys within each period. The respective taxa were surveyed by trained experts to ensure correct identification of individual species.
In both periods, survey techniques were kept the same to facilitate comparability. At the same time, survey techniques varied between taxa corresponding to their biological characteristics:
Plants were surveyed by visiting the respective major habitat types (see their definitions above) in each area and recording the presence of individual plant species (Čížek et al., 2013). The effort was proportional to the size of the respective abandoned military training areas (areas up to 50 ha were surveyed for 4 h, areas 50–100 ha for 8 h, areas 100–200 ha for 16 h, and areas larger than 200 ha for 24 h), and the time spent by surveying the respective habitat types in a given area corresponded to their relative coverage in that area. For plant surveys, each area was visited once during each period between June and August.
Grasshoppers were counted at regularly spaced points in each area, and the number of points corresponded to the size of the area (from 21 in the smallest area to 116 in the largest). Point locations remained the same in both periods. In each of both periods, every point was visited once between the mid-July and the end of August at the time of the highest grasshopper diurnal activity (between 10:00 and 17:00) under suitable weather conditions (warm sunny days with no wind). Surveys combined two methods widely used for grasshopper monitoring, i.e., sweeping (e.g., Racz et al., 2013) and acoustic detections (e.g., McNeil and Grozinger, 2020). Specifically, at each point, the observer made exactly 20 sweeps with a sweep net followed by a 2-min long acoustic detection of stridulating individuals. The records of both types of detections were summed to obtain the number of individuals of each species at a given point. These point-level data were then summed over the entire abandoned military training areas in the respective periods.
Butterflies were recorded using repeated, time-limited visits (sensu Kadlec et al., 2012). The effort during one visit corresponded to the size of the abandoned military training area (areas up to 25 ha were surveyed for 60 min, areas of 25–50 ha for 90 min, areas of 50–100 ha for 120 min, areas of 100–200 ha for 240 min, and areas larger than 200 ha for 300 min). In each period, the visits were repeated five times in each area covering the entire butterfly flying season from the end of April to the mid-September, to record species with different phenology and activity phases. Visits were conducted under suitable weather conditions (warm sunny days with no wind) between 9:00 and 16:00. During each visit, the observer surveyed all five major habitat types mentioned above present in a given abandoned military training area by a slow zigzag walk, and the time spent in the respective habitat types corresponded to their relative coverage in that area. During this walk, all observed species and their abundances were recorded. In the case of less distinguishable species, individuals were captured with an insect net, identified in hand, and released. Only in the case of species complexes indeterminable based on external morphology (accounting for a maximum of five individuals per habitat type and visit), the captured individuals were euthanised and identified later by genital dissection in laboratory.
Birds were surveyed during their breeding season using the spot mapping technique (Bibby et al., 2000), in which the observer walked slowly through the entire abandoned military training area early in the morning (between 5:00 and 10:00) and recorded every bird individual on a map. In each period, every area was visited twice (firstly from the late April to mid-May, secondly from mid-May to early June, with at least a two-week break between visits in an area) to cover both early and late breeders. The maximum count of a given species across the visits was considered to be its abundance in a given abandoned military training area in a given period.
Ecological group definition for the respective taxa
The perception of management approaches is likely to be different for species with different ecological traits. To address this issue, we defined several ecological groups of species for each taxon based on literature information on species ecology (plants—Chytrý et al., 2018; grasshoppers—Kočárek et al., 2013; butterflies—Beneš et al., 2002; birds—Hudec and Šťastný, 2005; Šťastný and Hudec, 2011). The groups were non-exclusive, i.e., a species classified in one group can also be classified in another. These ecological groups were delimited based on the position of species along successional and wetness gradients (for all taxa) and along a nutrient gradient (for plants) as follows:
Plants were sorted into 11 groups (Supplementary Table S1): shade-dwelling species, light-dwelling species, and generalists in respect to light conditions; hygrophilic species, mesophilic species, xerophilic species, and generalists in respect to wetness; nutrient-undemanding species, nutrient-demanding species, nutrient-intensive species, and generalists in respect to nutrients.
Grasshoppers were sorted into 5 groups (Supplementary Table S2): open-habitat hygrophilic species, open-habitat mesophilic species, semi-open/woodland-habitat mesophilic species, open-habitat xerophilic species, and semi-open/woodland-habitat xerophilic species.
Butterflies were sorted into 7 groups (Supplementary Table S3): open-habitat hygrophilic species, semi-open/woodland-habitat hygrophilic species, open-habitat mesophilic species, semi-open-habitat mesophilic species, woodland-habitat mesophilic species, open-habitat xerophilic species, semi-open/woodland-habitat xerophilic species.
Birds were sorted into 6 groups (Supplementary Table S4): species of open-wetter habitats, species of semi-open-wetter habitats, species of woodland-wetter habitats, species of open-drier habitats, species of semi-open-drier habitats, species of woodland-drier habitats.
In addition to the ecologically defined groups, we defined the group of threatened species for each taxon as the species listed in the currently valid national Red List (plants—Grulich, 2017; grasshoppers and butterflies—Hejda et al., 2017; birds—Šťastný et al., 2017) in the categories Near Threatened, Vulnerable, Endangered or Critically Endangered.
Processing of biodiversity data
The data were processed at the level of individual military training areas. For each taxon, we expressed the total number of species recorded in each area in the respective periods. Similarly, we expressed the abundance of the threatened species and the abundance of the respective ecological groups for each animal taxon. We decided to focus on the abundance data due to the low number of species in some groups and due to a higher sensitivity of abundance to environmental factors compared to the species richness (e.g., Sutcliffe et al., 2014; Dornelas et al., 2023). For plants, we used the number of species instead of abundance because data on the number of individuals were not available for this taxon (see above).
Although the identity of the abandoned military training areas and the effort devoted to the surveys were kept the same in both periods, the raw data were not fully comparable due to the loss of some parts of several areas between periods. These parts were inaccessible due to private construction activities without the possibility to collect biodiversity data. To solve this problem, we divided the raw biodiversity data (i.e., the number of species or individuals of the respective taxa or their groups) by the size of the surveyed area in each period in every abandoned military training area. Therefore, all numbers of species or individuals are further expressed as relative per hectare values. Since we were interested in possible impact of the management on the change of biodiversity, we calculated the change in these relative values between periods for each abandoned military training area. The change was calculated by subtracting the value in the first period from the value in the second period. The resulting positive value indicates an increase in the relative number of species or individuals between periods, while the negative value indicates a decrease. These changes between periods for the respective taxa and species groups within taxa were used as response variables for further analyses.
Management data
We distinguished the following six types of management that were carried out in the focal abandoned military training areas between the periods:
Woody plant cutting—removal of shrubs or trees from grasslands or shrublands to reduce encroachment on open habitats or to create new open habitat patches.
Mowing for conservation—extensive mowing of grasslands to improve their quality for biodiversity (preventing encroachment by woody plants, limiting nutrient demanding highly competitive plant species, and promoting less competitive, stress tolerant plant species and animals associated with extensively used grasslands).
Mowing for agriculture—mowing of grasslands to extract biomass for hay or silage.
Grazing for conservation—extensive grazing by domestic (sheep, cows) or semi-wild animals (Exmoor pony, European bison, aurochs-like cattle) to improve the quality of open habitats for biodiversity (creating bare ground patches, limiting nutrient-demanding highly competitive plant species and promoting less competitive stress-tolerant plant species and animals associated with extensively used grasslands).
Grazing for agriculture—grazing by domestic animals (sheep, cows) for agricultural production (meat, milk).
Vehicle movement—movement of civilian (motor bikes, off-road cars, trucks) or military (tanks, troop carriers) vehicles to reduce woodland encroachment on open habitats, to create bare ground patches and to promote stress tolerant plant species and animals associated with regrowth on such sites. This management type included both prescribed (for conservation purposes) and free-range rides due to their unclear distinction (e.g., some rides were free under the supervision of conservation managers) and the relatively small number of abandoned military training areas that experienced this type of management.
The influence of each management type on each abandoned military training area was expressed by two measures: i) the size of the area where it was applied and ii) the number of years of application (Table 1). The information on these measures was obtained from local stakeholders, nature conservation authorities and a public land use database (LPIS: https://eagri.cz/public/app/lpisext/lpis/verejny2/plpis/). We then converted these numbers into relative values (from 0 to 1), i.e., the coverage of a given abandoned military training area by a given management type and the proportion of years when this management type was applied (relative to the number of years between periods). For each abandoned military training area, these proportions were multiplied by each other to obtain a single measure of “management effort” for each management type, ranging from 0 to 1. The higher the value, the larger the area where that management type has been applied or the longer the period of its application in a given abandoned military training area. These management effort scores were used as explanatory variables in further analyses.
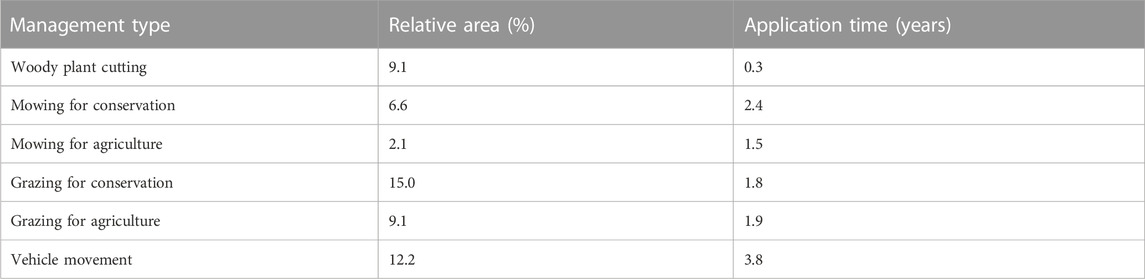
TABLE 1. Management effort in the surveyed abandoned military training areas. The effort is expressed as the mean relative area (%) and the mean number of years of application (application time) across all areas.
Statistical analysis
To assess changes in biodiversity between periods in the focal abandoned military training areas, we fitted intercept-only linear models (LM) for the change in the relative number of species or individuals of respective taxa or species groups. These changes illustrate the development of biodiversity over time (see above). Since the abandoned military training areas are rarely located randomly, and thus the location of the area may affect the species distribution, we used Moran’s test using the ‘Moran.I’ function from the ‘ape’ package (Paradis and Schliep, 2019) to test the residuals of each model for spatial autocorrelation. When significant, we remodelled the relationship using generalized least squares models (GLS) using the ‘gls’ function from the ‘nlme’ package (Pinheiro and Bates, 2022). The spatial autocorrelation structure was selected from linear, exponential, Gaussian, spherical or ratio quadratic correlation structures, based on the Akaike information criterion corrected for small sample sizes (AICc) calculated by the ‘AICc’ function from the ‘MuMIn’ package (Bartoń, 2022).
We then tested the effects of respective management types on the observed biodiversity changes between periods. For each biodiversity variable, i.e., the change in the relative number of species or individuals of a given taxon or a given group of species within a taxon, we fitted a separate LM or GLS relating this variable to the respective management types (quantified as “management effort”, see above) applied in individual abandoned military training areas. In addition to the influence of management, changes in biodiversity could be driven by the habitat structure of the area, or by interactions between management and habitat structure. Habitat structure was expressed as the relative cover of two habitat types, grassland and sparse shrubland. Information on the relative cover of these types was obtained by mapping of habitat composition in the abandoned military training areas prior to the 2009 biodiversity survey (see above). Although five habitat types were recognized during this mapping, only the two above-mentioned types were selected for the analysis based on results of the pilot modelling investigating their non-independence and representation.
Ideally, all possible combinations of management type, habitat structure and their interactions would be considered for each response variable. However, this approach was not feasible due to the limited sample size, i.e., the number of abandoned military training areas relative to the number of possible variable combinations, and the resulting potential statistical power issues. Therefore, to keep the model composition simple and comprehensive, we constructed three types of models:
i) models containing only the main effect of each management type;
ii) models containing the main effect of each management type and the main effects of habitat structure;
iii) models containing the main effect of each management type, the main effects of habitat structure, and their relevant two-way interactions. We considered relevant interactions to be those involving a habitat type in which a given management was most likely conducted assuming that the relative area of that habitat type may influence the impact of that management approach on biodiversity. Specifically, we included the following interactions: sparse shrubland x woody plant cutting, grassland x mowing for conservation, grassland x mowing for agriculture, grassland x grazing for conservation, and grassland x grazing for agriculture.
To make inference about the effect of management on a given response variable, we selected the best-fitting model from the three above mentioned possibilities based on the AICc, using ΔAICc < 2 as the criterion for model selection. However, we always used only the main effects of respective management types for interpretation because we were interested in the “pure” effect of management and not in the effect of habitat structure or its interaction. All models were tested for spatial autocorrelation and adjusted if necessary; using the same procedure as for the intercept-only models described above.
All model assumptions were checked visually using residuals vs. fitted plots, and Q-Q plots of standardized residuals. All statistical analyses were performed in R 4.2.1 (R Core Team, 2022).
To gain further insights into biodiversity dynamics in the focal areas and to provide a general overview of the processes involved in the observed changes, we calculated temporal beta-diversity for the respective taxa following Tatsumi et al. (2021). Temporal beta-diversity informs about spatiotemporal biodiversity changes by decomposing the presence-absence data collected at spatial replicates (i.e., individual abandoned military training areas in our case) and at different temporal occasions (i.e., our two survey periods) into the processes of extirpation and colonization (Tatsumi et al., 2021). Therefore, for each taxon, we can learn whether extirpation or colonization leads to homogenization or heterogenization of the ecological communities in the focal areas between survey periods, which can be translated into losses and gains of common and rare species, respectively. For this purpose, we quantified for each taxon (i) extirpation leading to homogenization, (ii) extirpation leading to heterogenization, (iii) colonization leading to homogenization, (iv) colonization leading to heterogenization, (v) total extirpation, (vi) total colonization, and (vii) total beta-diversity change. These measures could only be expressed for all abandoned military training areas together, and thus could not be related to management data at the level of individual areas.
Results
Combining data from both periods and all abandoned military training areas, we recorded 1089 species of plants, 64 species of grasshoppers, 121 species of butterflies and 86 species of birds. Of these species, 244 plants, 17 grasshoppers, 58 butterflies and 24 birds were listed in the actual national Red List.
Biodiversity changes between periods
In plants, biodiversity changes were generally positive. Overall, the number of plant species recorded per hectare of an abandoned military training area increased by 0.29 (±SE = 0.13) and the same pattern applied to Red List species, shade-dwelling species and nutrient-demanding species (Figure 1). Furthermore, increases were observed in all three groups of generalists, i.e., species indifferent to light conditions, wetness and nutrient gradients (Figure 2). None of the plant species groups showed a significant decrease in species richness (Figure 2).
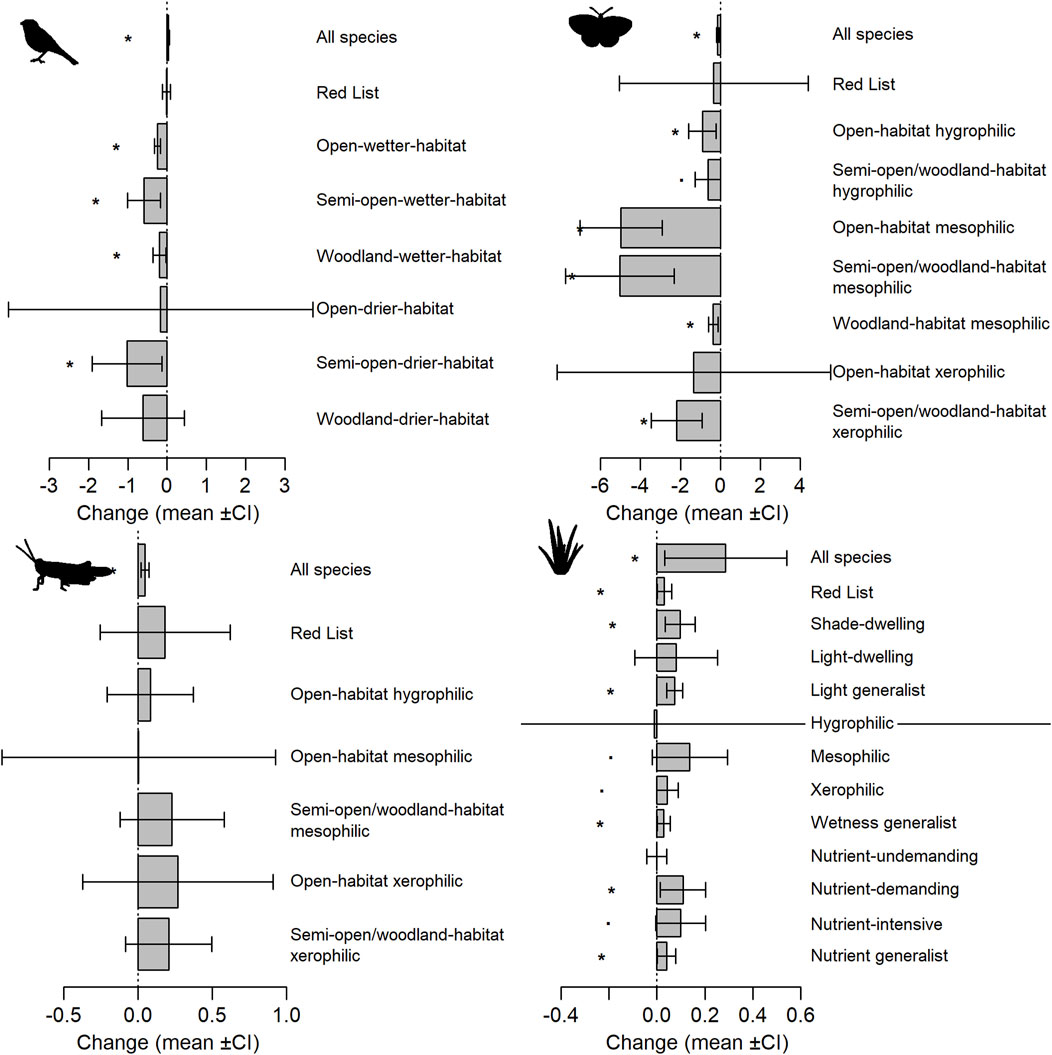
FIGURE 2. Changes in biodiversity of respective taxa (plants, grasshoppers, butterflies, and birds) in the surveyed abandoned military training areas between periods (2009–2010 and 2020–2022) estimated by linear or generalized least squares models. Mean changes (grey bars) together 95% confidence intervals (whiskers) in species richness (for plants groups and for all species in grasshoppers, butterflies and birds) or abundance (for respective groups of grasshoppers, butterflies and birds) are shown. Asterisk marks a significant change. Change is a relative value based on a difference in the per hectare number of species (or individuals) between periods (see the Materials and methods section for details).
For grasshoppers, the total number of species recorded per hectare increased significantly, similar to plants (Figure 2), but to a lesser extent, i.e., by 0.05 (±0.01). Focusing on changes in the abundance of respective species groups, we observe a general pattern of increases across the groups defined by various combinations of habitat openness and wetness, as well as in the Red List species (Figure 2). However, none of these changes was statistically significant (Figure 2).
Butterflies, in contrast to plants and grasshoppers, showed remarkably different patterns of change in species richness and abundance per hectare. Specifically, their total species richness significantly decreased by 0.14 (±0.03), and five out of seven groups defined by various combinations of habitat openness and wetness showed significant decreases in abundance (Figure 2). These decreases included semi-open/woodland-habitat xerophilic species, open-, semi-open- and woodland-habitat mesophilic species, and open-habitat hygrophilic species (Figure 2). The remaining groups, as well as Red List species, showed a tendency to decline, but this was not statistically significant (Figure 2).
Bird biodiversity expressed a mixed pattern of change. The total number of bird species recorded per hectare of an abandoned military training area slightly (0.03 ± 0.01), but significantly increased (Figure 2). However, the changes in abundance per hectare of the respective species groups were exclusively negative. Specifically, significant decreases were observed in all groups on wetter habitats regardless of their position along the open-woodland habitat gradient, and in semi-open-drier-habitat species (Figure 2). The abundance of the remaining groups including the Red List species also tended to decrease, but not significantly (Figure 2).
Relationships between biodiversity changes and management types
For plants (Supplementary Table S5), the change in richness of Red List species was positively related to woody plant cutting and mowing for agriculture (Figure 3A). Mowing for conservation was negatively related to the change in richness of generalists with respect to wetness (Figure 3A). Changes in the number of xerophilic and nutrient undemanding species were both positively related to woody plant cutting (Figure 3A). The latter group also responded positively to grazing for agriculture (Figure 3A).
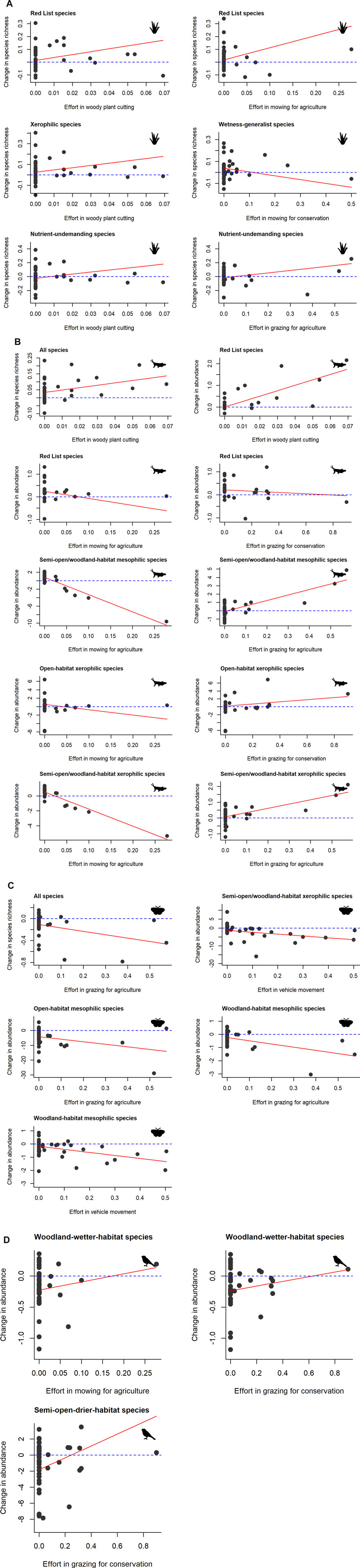
FIGURE 3. Effects of respective management types on changes in biodiversity of respective taxa in the surveyed abandoned military training areas between periods (2009–2010 and 2020–2022) estimated by linear or generalized least squares models: (A) plants, (B) grasshoppers, (C) butterflies, (D) birds. Change is a relative value based on a difference in the per hectare number of species (or individuals) between periods (see the Materials and methods section for details). Only the significant relationships are shown (see Supplementary Tables S5–S8 for full results).
Grasshoppers showed partly different relationships to the respective management types than plants (Supplementary Table S6). Here we also observed a positive response to woody plant cutting, as indicated by the increase of species richness of all species and abundance of Red List species (Figure 3B). However, we detected four negative relationships to mowing for agriculture: in the case of abundance of Red List species, open-habitat xerophilic species, semi-open/woodland habitat xerophilic and mesophilic species (Figure 3B). Grazing for conservation was positively related to the change in abundance of open-habitat xerophilic species, but negatively related to the change in abundance of Red List species (Figure 3B). Abundance of semi-open/woodland-habitat xerophilic and mesophilic species increased with increasing effort devoted to grazing for agriculture (Figure 3B).
For butterflies (Supplementary Table S7), the change in their total species richness was negatively related to grazing for agriculture, as was the change in abundance of open- and woodland-habitat mesophilic species (Figure 3C). In addition, the change in abundance of semi-open/woodland-habitat xerophilic and woodland-habitat-mesophilic species was negatively related to vehicle movement (Figure 3C).
For birds, the change in their total species richness was unrelated to any of the management types, and the same applied to the abundance of Red List species (Supplementary Table S8). The change in abundance of woodland-wetter-habitat species was positively related to both mowing for agriculture and grazing for conservation (Figure 3D). Finally, the abundance of semi-open-drier-habitat species increased with increasing effort in grazing for conservation (Figure 3D).
In summary across all taxa and species groups (Table 2), woody plant cutting was associated with an increase in biodiversity in five cases (three for plants, two for grasshoppers) and it did not show any negative relationships. Mowing for conservation showed a single negative relationship (in plants), whereas mowing for agriculture two positive (one for plants, and one for birds) and four negative relationships (all in grasshoppers). Grazing for conservation showed three positive (one for grasshoppers, two for birds) and one negative relationship (for grasshoppers), while grazing for agriculture showed three positive (one for plants and two for grasshoppers) and three negative (all in butterflies) relationships. Vehicle movement was linked to two negative responses (both in butterflies).

TABLE 2. Summary of the impacts of respective management types on individual taxa in the surveyed abandoned military training areas: A) plants, B) grasshoppers, C) butterflies, D) birds. ↓↑ denote significantly positive and negative relationship, respectively.
Temporal change in beta-diversity
In plants, grasshoppers and butterflies we observed an increase in beta-diversity, i.e., their communities across abandoned military training areas became more dissimilar between periods, although the increase was modest in grasshoppers (Table 3). In birds, beta-diversity decreased, i.e., bird communities became increasingly similar between periods, but the value was close to zero, meaning the change was small (Table 3).
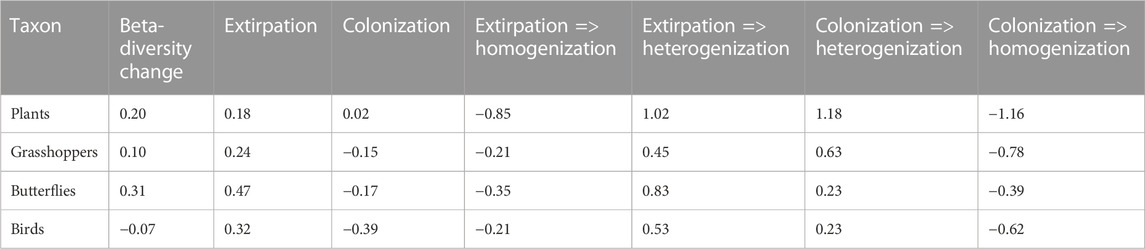
TABLE 3. Decomposition of temporal changes in beta-diversity for respective taxa (plants, grasshoppers, butterflies, and birds) in the surveyed abandoned military training areas. Temporal change in beta-diversity (first column) can be expressed as the sum of extirpation and colonization processes (second and third columns) among focal areas between survey periods. Extirpation is the sum of local extinction of rare species leading to homogenization (fourth column) and local extinction of common species leading to heterogenization (fifth column). Colonization is the sum of the spread of rare species leading to heterogenization (sixth column) and the spread of common species leading to homogenization (seventh column). The sign of beta-diversity change shows its increase (positive values, i.e., higher dissimilarity of assemblages represented by a given taxon across areas) or decrease (negative values) over time. The sign of extirpation and colonization shows the dominance of homogenization (negative values, i.e., assemblages became increasingly homogenous in terms of species composition, leading to a decrease in beta-diversity) or heterogenization (positive values, leading to an increase in beta-diversity) in a given process.
The extirpation of common species (leading to heterogenization) was always greater than the extirpation of rare species (leading to homogenization) and it was the main driver of the overall beta-diversity change in plants, grasshoppers and butterflies (Table 3). However, this does not mean that colonization did not occur in these taxa. In fact, both plants and grasshoppers showed high levels of colonization (Table 3), but since the colonization processes were symmetric between common and rare species (Table 3), their net impact on beta-diversity change was negligible in these taxa. In butterflies, colonization of common species was much greater than colonization of rare species (Table 3). However, extirpation of common butterflies was even higher, so that it outweighed the effect of colonization on beta-diversity change in this taxon (Table 3). The situation was somewhat different in birds, where colonization was slightly greater than extirpation for both common and rare species (Table 3). As colonization of common bird species dominated among the processes, total bird beta-diversity decreased between survey periods (Table 3).
Discussion
Four major taxa selected to represent the biota of abandoned military training areas showed strikingly divergent trajectories of biodiversity change according to our standardized-effort surveys repeated after 10–12 years. Plant species richness generally increased, and the increase typically involved ecological generalists and nutrient-demanding and shade-dwelling species. However, national Red Listed plant species increased richness as well, and none of the plant ecological groups declined. Positive changes were also observed in grasshoppers whose total species richness increased and abundance of all species groups tended to increase, although not significantly. This optimistic picture contrasts deeply with the patterns found for butterflies. Their total species richness decreased, as did the abundance of most of their species groups, regardless of their position along gradients of habitat openness and wetness. Birds showed a mixed pattern containing increase in total species richness and decreases in the abundance of species associated with semi-open drier habitats and various types of wetter habitats. Beta-diversity increased between periods in all but one taxon (birds), driven by extirpation of common species. In contrast, colonization of common species was responsible for the decrease in beta-diversity in birds.
Regarding management effort, we found a complex picture of highly variable biodiversity impacts of different approaches. The cutting of woody plants was beneficial for national Red Listed, xerophilic and nutrient undemanding plants, as well as for overall grasshopper richness and for grasshoppers listed in the national Red List. Mowing for conservation showed only negative relationship with biodiversity change, but this may not necessarily be a sign of conservation failure, as we discuss below. Mowing and grazing for agriculture provided both conservation benefits (for plants, birds and some grasshoppers) and costs (for butterflies and some other grasshoppers). Grazing for conservation had positive effects on open-habitat xerophilic grasshoppers and woodland-wetter-habitat birds, but was negatively related to abundance changes in national Red Listed grasshoppers. Vehicle movement was negative for butterflies of later successional stages.
The patterns observed in plants, i.e., the increase in the richness of shade-dwelling and nutrient-demanding species, indicate some progress in ecological succession (sensu Poorter et al., 2023) leading to habitat closure and eutrophication (e.g., Pruchniewicz, 2017). On the other hand, these processes do not seem to endanger the sensitive plant species, since no decrease in biodiversity was observed in any of the plant species groups. Moreover, the increase in richness of Red Listed plant species suggests that abandoned military training areas maintain their position as strongholds of plant biodiversity in the region (Čížek et al., 2013). One reason for these patterns may be the positive effects of management. The cutting of woody plants supported Red List species, nutrient undemanding and xerophilic species, i.e., groups that should be negatively affected by shrub or forest encroachment and eutrophication (Teleki et al., 2020). Grazing, although for agricultural purposes, could have also contributed to mitigating these impacts (Pykälä, 2003; Elias et al., 2018), as it supported nutrient undemanding species, too.
Mowing is another management tool that is considered to counteract successional changes and eutrophication (Lepš, 1999). However, its effects on plants were mixed in our study areas. Interestingly, plant species (those listed in the national Red List) seem to benefit from mowing for agriculture, but suffer from mowing for conservation (generalists to wetness). Although surprising, these patterns can be explained by species’ ecological characteristics. Many species listed in the Red List originally occur in steppe habitats that are highly exposed to natural disturbances (Chytrý et al., 2007). Therefore, disturbance represented by mowing is beneficial for them as it keeps their habitats open. In contrast, generalists, that include many woody plants and expansive species (such as Calamagrostis epigeios) in our data, are often targeted for reduction by management (Těšitel et al., 2018) because they trigger deterioration of open habitats (Somodi et al., 2008). Thus, the negative impact of mowing for conservation on these species can be perceived as a positive outcome.
Similar to plants, grasshoppers are a taxon showing predominance of positive biodiversity changes, and management accounts for at least part of them. Cutting of woody plants proved to be beneficial for total species richness of grasshoppers and abundance of national Red List species. Since grasshoppers generally prefer open habitats (Latchininsky et al., 2011) and the threatened species like grasshopper Dociostaurus brevicollis or bush-cricket Montana montana are typically those with the highest habitat specialization (Engelhardt et al., 2022), reducing woody plant encroachment seems to be a vital strategy to support them. Xerophilic and mesophilic species in semi-open and woodland habitats were supported by grazing for agriculture that may maintain the conditions created by the cutting of woody plants, even though it is unlikely to improve the quality of the habitat per se. Instead, habitat quality can be improved by grazing for conservation that was associated with increased abundance of open-habitat xerophilic grasshopper species like grasshopper Calliptamus italicus and bush-cricket Platycleis albopunctata. The surprisingly negative impact of grazing for conservation on Red List grasshopper species was driven by large increases of these species in localities where this management type was absent and their modest increases in localities with a high effort devoted grazing for conservation. Such a difference resulted in an apparently negative relationship, but it would be premature to consider this management type as harmful.
In contrast to plants, where we found both positive and negative effects of mowing, this management approach had solely negative effects on grasshoppers. The abundance of open-habitat xerophilic species, semi-open/woodland habitat xerophilic and mesophilic species, as well as of Red List species decreased with increasing effort devoted to mowing for agriculture. Due to mowing, plant individuals are cut which results in high mortality of grasshoppers (Humbert et al., 2010). If mowing occurs too often, as may be the case for this management type in some of our localities, such frequent mortality events limit population recovery and result in population declines. Mortality is probably less severe in the case of grazing, even though it is performed for agricultural purposes as it had a positive effect on semi-open/woodland-habitat xerophilic grasshoppers. We suggest that the benefit may lie in preventing woodland habitat closure. Although these species are associated with woody plants (Latchininsky et al., 2011), they require open and not closed woodland (Rösch et al., 2019) which may be maintained by grazing.
Butterflies are organisms that seem to be at particular risk in abandoned military training areas. Their total species richness and the abundance of several groups show negative changes over time, and we did not detect any positive effects of management. Instead, two types of management showed negative effects. Specifically, butterflies suffered from grazing for agriculture and from vehicle movement. Management by intensive and long-term grazing results in uniform and low sward height which reduces butterfly breeding and shelter opportunities (Bussan, 2022), as well as in destruction of plant-pollinators interactions by reduction of nectar food supply for butterflies (Rakosy et al., 2022). These results suggest that butterflies may be negatively affected by the management type that simultaneously provides benefits to some other taxa (i.e., plants and grasshoppers in our dataset). Such sensitivity of butterflies can be explained by the high ecological specialization of many species, such as Euphydryas aurinia or hairstreaks (species of subfamily Theclinae), coupled with their complex habitat requirements—species often require the simultaneous presence of several different kinds of ecological conditions on a small area (e.g., both tall and short herbs, both grassland and woodland) that are very difficult to create or even support by a single management type (e.g., Hůla et al., 2004). The fine mosaic of variable habitat types in military training areas a few years after their abandonment was most likely the reason of the high richness and abundance of butterflies recorded at time of our first survey (Čížek et al., 2013) and it seems that subsequent homogenization of this mosaic was not prevented by management despite considerable effort. This probably resulted in the observed butterfly biodiversity decline. Therefore, butterflies appear to require a more nuanced approach to the management of abandoned military training areas.
Butterflies bound to the later successional stages were also negatively affected by vehicle movement. This result is driven by the extensive top-soil removal by military vehicles carried out as part of planned management on several abandoned military training ranges, which occurred just before our second survey period (Dvořáková et al., 2002a; Dvořáková et al., 2002b). Such an extensive disturbance inevitably resulted in habitat loss for many butterfly species, particularly those associated with forest-steppe or light forests, as Callophrys rubi or Lasiommata maera, which show significant population decreases. However, this apparent negative effect may not be permanent, since the experience from other areas that underwent such kind of disturbance indicates that, if these disturbances are not too frequent, butterfly populations recover after a few years (Zografou et al., 2017) and the community may become even more diverse compared to pre-disturbance conditions (Tropek et al., 2012). Therefore, although it is unclear whether such recovery will occur in our study areas, the observed negative impacts of vehicle movement may not be as fatal as our results suggest.
Birds showed a mixture of positive and negative changes as their total species richness increased, while the abundance of several species groups decreased between study periods. Some of these increases can be attributed to management options. Such options were represented by grazing for conservation to which birds associated with semi-open, drier habitat types responded positively. This type of disturbance blocks successional changes and maintains habitat openness, i.e., creates conditions that exactly match the habitat preferences of these bird species (Reif et al., 2013; Aunins and Avotis, 2018), such as the Stonechat (Saxicola rubicola) or Woodlark (Lullula arborea) in our dataset. At the same time, woodland birds in wetter habitats responded positively to both mowing for agriculture and grazing for conservation. We suggest that these associations can be attributed to their foraging habits—even though these species breed in woodland, they forage on the ground in grasslands (Hudec and Šťastný, 2005; Šťastný and Hudec, 2011), and reduction of herb height makes their food more accessible (Hoste-Danylow et al., 2010). In addition, large mammalian herbivores used for conservation grazing serve as a bird food source per se, attracting various kinds of flying insects (e.g., species of the order Diptera) and hosting their larvae (Musitelli et al., 2016).
Regarding the changes in beta-diversity, it is interesting that common species drove the pattern in all taxa, regardless of whether their species richness increased or decreased. This is consistent with the important role of common species in the ecosystems which has recently been recognized from both theoretical (Sizling et al., 2009) and applied perspectives (Gaston, 2010). In this respect, it is particularly worrying that the increasing beta-diversity of plants, grasshoppers and butterflies in the studied abandoned military training areas was driven by the extirpation of the common species. Such species may suffer from being overlooked by the current management efforts, which are typically prescribed on the basis of the needs of the most threatened species (Scheele et al., 2018) which are usually also rare (Mace et al., 2008). Birds contrasted somewhat with the other taxa, as their beta-diversity decreased due to a slight dominance of colonization by common species over extirpation. As the most common bird species prefer later successional stages (Bystricky et al., 2023), it is possible that they benefited from the lack of management in some areas.
Although at least some of the observed changes in biodiversity can be explained by management, there may be other drivers that could not be included in our analysis given the data we have. In this respect, climate change is the hottest candidate. Indeed, the increase in species richness due to climate change has been documented in Central European grasshoppers (Fartmann et al., 2022) and birds (Leroy et al., 2023), as well as the decrease in plant (Wesche et al., 2012) and butterfly richness (Eskildsen et al., 2015). However, the effects of this global driver are usually recorded at large scales, whereas the local-level data we focus on here reflect the effects of local habitat changes (see e.g., Jandt et al., 2022). Thus, we do not think that it would be strong enough to affect the reported relationships with management types at the study sites.
Our simultaneous focus on four different taxa uncovered various taxon-specific patterns in biodiversity-management relationships. If we had studied only a single taxon, e.g., birds which are often considered reliable state-of-nature indicators (Fraixedas et al., 2020), we would have obtained a seriously biased view of the system showing increasing species richness (not true for butterflies), decreasing beta-diversity (not observed in plants, grasshoppers and butterflies), and the lack of response to several management options, such as woody plant cutting (supporting plants and grasshoppers) and vehicle movement (negatively affecting butterflies). Therefore, we recommend conducting multitaxonomic studies to gain complex insights into the effects of various management types on biodiversity in open habitats. Considering such complexity is important for setting appropriate conservation targets (e.g., Boetzl et al., 2021).
Conclusions and conservation implications
Our study demonstrated a dynamic change of biodiversity in abandoned military training areas, sites considered to be biodiversity strongholds. At least some of these changes can be explained by the management. To that end, we propose the following recommendations for practitioners:
1) We observed both positive and negative biodiversity responses to the focal management types across taxa. It often happens that the benefits of one management approach for a given group of organisms are offset by costs to another one. Environmental management of abandoned military training areas thus must be based on setting the priorities for conservation.
2) As one such priority that can be considered the most urgent, we propose the conservation of butterflies. Butterflies are the only taxon to have a consistent pattern of biodiversity loss, negative management impacts and no benefits. We believe that these patterns arise from the specific ecology of this taxon, requiring smaller-scale habitat heterogeneity than any other taxa studied. To reverse the negative trends, we suggest that butterflies need combination of different management approaches that would increase small-scale habitat heterogeneity. Finding the optimal management combinations for the most declining butterflies should be the most urgent research priority.
3) The cutting of woody plants seems to be widely positive with benefits for both plants and grasshoppers and did not have any negative impacts on the other species groups. Therefore, it seems that it is an important management tool for blocking woody plant encroachment and it is currently applied in biologically plausible. Its current form can be further supported.
4) The effectiveness of mowing for conservation should be thoroughly investigated. According to our data, it did not increase the biodiversity of any groups of organisms. Although the observed negative effect can be perceived as advantageous for biodiversity (i.e., suppression of generalist plants), the absence of positive relationships is worrying. More detailed studies with an experimental design are needed to identify the aspects of this type of management that should be improved.
5) Both mowing and grazing for agriculture showed a mixture of biodiversity benefits and costs. Overall, plants and birds were more likely to enjoy the benefits, while grasshoppers and butterflies were more likely to experience the costs. We suggest that these different effects may be explained by the differences in the lifestyle of these organisms. As both types of agricultural management are relatively intensive, the organisms that could perceive their benefits should also exhibit some form of ecological generalisation (birds) or resilience (plants). In contrast, both invertebrate taxa are relatively specialized and thus more sensitive to the negative consequences of this management approach which particularly concerns butterflies. Therefore, we suggest that mowing and grazing for agriculture can be applied on some abandoned military training areas, but not on sites where they can harm the sensitive taxa. This particularly applies to mowing for agriculture in grasshoppers and to grazing for agriculture in butterflies.
6) Grazing for conservation brought more benefits than costs, which seems encouraging. On the other hand, the benefits are modest at best, since none were observed for plants and butterflies. At the same time, these taxa are often considered to be those for which this management type is most frequently prescribed (Bubová et al., 2015). We therefore urge detailed experimental studies (such as Konvička et al., 2021) in areas of application to identify the aspects of this management approach that can be improved to maximize its positive influence.
7) Vehicle movement resulted in costs for butterflies. Although this effect may be temporary as we discuss above, we suggest that vehicle movement should be applied in a more spatially restricted manner that would prevent such negative impacts.
8) Extirpation of common species was a main driver of biodiversity changes in plants, grasshoppers and butterflies. Such species do not appear to be supported by current management efforts, and the development of specific management types to satisfy their requirements may be needed.
In conclusion, our study is the first to investigate the consequences of various managements approaches on different representatives of biodiversity in abandoned military training areas. Based on this investigation, we identified several knowledge gaps and provide management recommendations and suggestions for further research. It is important to recognize that our data collected at the level of whole areas cannot reveal more detailed relationships that require experimental design. Such experimental studies should be a research priority in the future.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://osf.io/9j6ya/.
Ethics statement
Data were obtained by standardly used methodology conforming to the national legislation. No specific permissions for manipulation with living objects were required given the observational nature of the study.
Author contributions
PM, JR, JK, OČ, and TK conceived the idea. TK, JR, PM, JK, OČ, and LD designed the research. LD, JK, PM, TK, OČ, and JR collected the data. PC conducted the analysis. JR led writing with inputs from all co-authors. All authors contributed to the article and approved the submitted version.
Funding
The study was supported by the Technology Agency of the Czech Republic (program “Prostředí pro život”, project SS03010162).
Acknowledgments
We wish to thank following colleagues who significantly contributed to fieldwork: Miroslav Bažant, Václav Bystřický, Jaroslav Chloupek, Štěpánka Čížková, Jiří Darebník, Libor Dvořák, Tomáš Dvořák, Květoslav Fryšták, Jan Hájek, Petr Heřman, Jaroslav Holuša, David Hořák, Záboj Hrázský, Jan Hrnčíř, Lucie Hrůzová, Kamil Hromádko, Věra Hromádková, Vladimír Hula, Lada Jakubíková, Petr Kočárek, Vojtěch Kodet, Jaroslav Koleček, Karel Konečný, Vojtěch Kubelka, David Musiolek, Stanislav Rada, Dušan Rossi, Libor Schröpfer, Jiří Skala, Robert Stejskal, Vlasta Škorpíková, David Storch, Jiří Švarc, Roman Vacík, Jakub Vlček, Robert Vlk, Ondřej Volf, Petr Voříšek, Jakub Vrána and Jaroslav Zámečník. Martin Konvička and Marta Kotecká Misíková were involved in designing this study.
Conflict of interest
Authors LD, JK, and PM are employed by Beleco, Z.S.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2023.1243568/full#supplementary-material
References
Aunins, A., and Avotins, A. (2018). Impact of military activities on bird species considered to benefit from disturbances: an example from an active military training area in Latvia. Ornis Fenn. 95, 15–31.
Bardgett, R. D., Bullock, J. M., Lavorel, S., Manning, P., Schaffner, U., Ostle, N., et al. (2021). Combatting global grassland degradation. Nat. Rev. Earth Environ. 2, 720–735. doi:10.1038/s43017-021-00207-2
Bartoń, K. (2022). MuMIn: multi-model inference. R package. version 1.47.1 https://CRAN.R-project.org/package=MuMIn.
Beneš, J., Konvička, M., Dvořák, J., Fric, Z., Havelda, Z., Pavlíčko, A., et al. (2002). Butterflies of the Czech republic: distribution and conservation I, II. Praha: SOM.
Bibby, C. J., Hill, D. A., Burgess, N. D., and Mustoe, S. H. (2000). Bird census techniques. 2nd edn. London: Academic Press.
Boetzl, F. A., Krauss, J., Heinze, J., Hoffmann, H., Juffa, J., Konig, S., et al. (2021). A multitaxa assessment of the effectiveness of agri-environmental schemes for biodiversity management. Proc. Natl. Acad. Sci. U. S. A. 118, e2016038118. doi:10.1073/pnas.2016038118
Bubová, T., Vrabec, V., Kulma, M., and Nowicki, P. (2015). Land management impacts on European butterflies of conservation concern: a review. J. Insect Conserv. 19, 805–821. doi:10.1007/s10841-015-9819-9
Bušek, O., and Reif, J. (2017). The potential of military training areas for bird conservation in a central European landscape. Acta Oecol 84, 34–40. doi:10.1016/j.actao.2017.08.005
Bussan, S. K. (2022). Can cattle grazing benefit grassland butterflies? J. Insect Conserv. 26, 359–374. doi:10.1007/s10841-022-00373-8
Bystricky, V., Dvorakova, L., and Reif, J. (2023). Conservation status of birds in habitats along a successional gradient from bare ground to forest. Comm. Ecol. doi:10.1007/s42974-023-00158-2
Cardoso, P., Barton, P. S., Birkhofer, K., Chichorro, F., Deacon, C., Fartmann, T., et al. (2020). Scientists' warning to humanity on insect extinctions. Biol. Conserv. 242, 108426. doi:10.1016/j.biocon.2020.108426
Chase, J. M., Blowes, S. A., Knight, T. M., Gerstner, K., and May, F. (2020). Ecosystem decay exacerbates biodiversity loss with habitat loss. Nature 584, 238–243. doi:10.1038/s41586-020-2531-2
Chytrý, M., Kočí, M., Šumberová, K., Sádlo, J., Krahulec, F., Hájková, P., et al. (2007). Vegetation of the Czech republic 1. Grassland and shrubland communities. Praha: Academia.
Chytrý, M., Tichý, L., Dřevojan, P., Sádlo, J., and Zelený, D. (2018). Ellenberg-type indicator values for the Czech flora. Preslia 90, 83–103. doi:10.23855/preslia.2018.083
Čížek, O., Vrba, P., Beneš, J., Hrázsky, Z., Koptík, J., Kučera, T., et al. (2013). Conservation potential of abandoned military areas matches that of established reserves: plants and butterflies in the Czech Republic. PLoS ONE 8, e53124. doi:10.1371/journal.pone.0053124
Diekmann, M. (2003). Species indicator values as an important tool in applied plant ecology - a review. Basic Appl. Ecol. 4, 493–506. doi:10.1078/1439-1791-00185
Dornelas, M., Chase, J. M., Gotelli, N. J., Magurran, A. E., McGill, B. J., Antao, L. H., et al. (2023). Looking back on biodiversity change: lessons for the road ahead. Phil. Trans. R. Soc. Lond. B 378, 20220199. doi:10.1098/rstb.2022.0199
Dvořáková, L., Bušek, O., Hernová, J., and Reif, J. (2023). Relationships between bird species richness and different facets of landscape heterogeneity – insights from a military area. J. Vert. Biol. 72, 23012. doi:10.25225/jvb.23012
Dvořáková, L., Marhoul, P., Koptík, J., and Kotecká Misíková, M. (2022a). Restoration of sandy steppes in Pánov – case study of the Military LIFE for Nature project. Praha: Beleco.
Dvořáková, L., Marhoul, P., Koptík, J., and Kotecká Misíková, M. (2022b). Restoration of steppe habitats in načeratický kopec special area of conservation – case study of the military LIFE for nature project. Praha: Beleco.
Dvorský, M., Mudrák, O., Doležal, J., and Jirků, M. (2022). Reintroduction of large herbivores restored plant species richness in abandoned dry temperate grassland. Plant Ecol. 223, 525–535. doi:10.1007/s11258-022-01225-w
Elias, D., Hölzel, N., and Tischew, S. (2018). Goat paddock grazing improves the conservation status of shrub-encroached dry grasslands. Tuexenia 38, 215–233. doi:10.14471/2018.38.017
Ellis, E. C., Gauthier, N., Goldewijk, K. K., Bird, R. B., Boivin, N., Diaz, S., et al. (2021). People have shaped most of terrestrial nature for at least 12,000 years. Proc. Natl. Acad. Sci. U.S.A. 118, e2023483118. doi:10.1073/pnas.2023483118
Ellwanger, G., and Reiter, K. (2019). Nature conservation on decommissioned military training areas – German approaches and experiences. J. Nat. Conserv. 49, 1–8. doi:10.1016/j.jnc.2019.02.003
Engelhardt, E. K., Biber, M. F., Dolek, M., Fartmann, T., Hochkirch, A., Leidinger, J., et al. (2022). Consistent signals of a warming climate in occupancy changes of three insect taxa over 40 years in central Europe. Glob. Chang. Biol. 28, 3998–4012. doi:10.1111/gcb.16200
Eskildsen, A., Carvalheiro, L. G., Kissling, W. D., Biesmeijer, J. C., Schweiger, O., and Hoye, T. T. (2015). Ecological specialization matters: long-term trends in butterfly species richness and assemblage composition depend on multiple functional traits. Divers. Distrib. 21, 792–802. doi:10.1111/ddi.12340
Fartmann, T., Kramer, B., Stelzner, F., and Poniatowski, D. (2012). Orthoptera as ecological indicators for succession in steppe grassland. Ecol. Ind. 20, 337–344. doi:10.1016/j.ecolind.2012.03.002
Fartmann, T., Poniatowski, D., and Holtmann, L. (2022). Effects of land-use and climate change on grasshopper assemblages differ between protected and unprotected grasslands. Biodiv. Conserv. 63, 83–92. doi:10.1016/j.baae.2022.06.005
Fraixedas, S., Linden, A., Piha, M., Cabeza, M., Gregory, R., and Lehikoinen, A. (2020). A state-of-the-art review on birds as indicators of biodiversity: advances, challenges, and future directions. Ecol. Ind. 118, 106728. doi:10.1016/j.ecolind.2020.106728
Gaertner, M., Konold, W., and Richardson, D. M. (2010). Successional changes on a former tank range in eastern Germany: does increase of the native grass species Molinia caerulea cause decline of less competitive Drosera species? J. Nat. Conserv. 18, 63–74. doi:10.1016/j.jnc.2009.03.001
Goulson, D. (2019). The insect apocalypse, and why it matters. Curr. Biol. 29, R967–R971. doi:10.1016/j.cub.2019.06.069
Grulich, V. (2017). Red List of the threatened species of the Czech Republic. Vascular plants. Příroda 35, 75–132.
Habel, J. C., Dengler, J., Janisova, M., Torok, P., Wellstein, C., and Wiezik, M. (2013). European grassland ecosystems: threatened hotspots of biodiversity. Biodiv. Conserv. 22, 2131–2138. doi:10.1007/s10531-013-0537-x
Hejda, R., Farkač, J., and Chobot, K. (2017). Red list of the threatened species of the Czech republic. Invertebrates. Příroda 36, 8–554.
Herčík, J., Šimáček, P., Szczyrba, Z., and Smolová, I. (2014). Military brownfields in the Czech Republic and the potential for their revitalisation, focused on their residential function. Quaest. Geogr. 33, 127–138. doi:10.2478/quageo-2014-0021
Hilty, J., and Merenlender, A. (2000). Faunal indicator taxa selection for monitoring ecosystem health. Biol. Conserv. 92, 185–197. doi:10.1016/s0006-3207(99)00052-x
Hoste-Danyłow, A., Romanowski, J., and Żmihorski, M. (2010). Effects of management on invertebrates and birds in extensively used grassland of Poland. Agric. Ecosyst. Environ. 139, 129–133. doi:10.1016/j.agee.2010.07.009
Hůla, V., Konvička, M., Pavličko, A., and Fric, Z. (2004). Marsh Fritillary (Euphydryas aurinia) in the Czech Republic: monitoring, metapopulation structure, and conservation of the endangered butterfly. Entomol. Fenn. 15, 231–241. doi:10.33338/ef.84226
Humbert, J. Y., Ghazoul, J., Richner, N., and Walter, T. (2010). Hay harvesting causes high orthopteran mortality. Agric. Ecosyst. Environ. 139, 522–527. doi:10.1016/j.agee.2010.09.012
Jandt, U., Bruelheide, H., Jansen, F., Bonn, A., Grescho, V., Klenke, R. A., et al. (2022). More losses than gains during one century of plant biodiversity change in Germany. Nature 611, 512–518. doi:10.1038/s41586-022-05320-w
Jentsch, A., Friedrich, S., Steinlein, T., Beyschlag, W., and Nezadal, W. (2009). Assessing conservation action for substitution of missing dynamics on former military training areas in Central Europe. Restor. Ecol. 17, 107–116. doi:10.1111/j.1526-100x.2007.00339.x
Kadlec, T., Tropek, R., and Konvička, M. (2012). Timed surveys and transect walks as comparable methods for monitoring butterflies in small plots. J. Insect Conserv. 16, 275–280. doi:10.1007/s10841-011-9414-7
Kočárek, P., Holuša, J., Vlk, R., and Marhoul, P. (2013). Grasshoppers of the Czech republic. Praha: Academia.
Konvička, M., Ričl, D., Vodičková, V., Beneš, J., and Jirků, M. (2021). Restoring a butterfly hot spot by large ungulates refaunation: the case of the Milovice military training range, Czech Republic. BMC Ecol. Evol. 21, 73. doi:10.1186/s12862-021-01804-x
Latchininsky, A., Sword, G., Sergeev, M., Cigliano, M. M., and Lecoq, M. (2011). Locusts and grasshoppers: behavior, ecology, and biogeography. Psyche 2011, 1–4. doi:10.1155/2011/578327
Lepš, J. (1999). Nutrient status, disturbance and competition: an experimental test of relationships in a wet meadow. J. Veget. Sci. 10, 219–230. doi:10.2307/3237143
Leroy, F., Reif, J., Storch, D., and Keil, P. (2023). How has bird biodiversity changed over time? A review across spatio-temporal scales. Basic Appl. Ecol. 69, 26–38. doi:10.1016/j.baae.2023.03.004
Mace, G. M., Collar, N. J., Gaston, K. J., Hilton-Taylor, C., Akcakaya, H. R., Leader-Williams, N., et al. (2008). Quantification of extinction risk: IUCN's system for classifying threatened species. Conserv. Biol. 22, 1424–1442. doi:10.1111/j.1523-1739.2008.01044.x
Maes, J., Paracchini, M. L., Zulian, G., Dunbar, M. B., and Alkemade, R. (2012). Synergies and trade-offs between ecosystem service supply, biodiversity, and habitat conservation status in Europe. Biol. Conserv. 155, 1–12. doi:10.1016/j.biocon.2012.06.016
McNeil, D. J., and Grozinger, C. M. (2020). Singing in the suburbs: point count surveys efficiently reveal habitat associations for nocturnal Orthoptera across an urban-to-rural gradient. J. Insect Conserv. 24, 1031–1043. doi:10.1007/s10841-020-00273-9
Musitelli, F., Romano, A., Moller, A. P., and Ambrosini, R. (2016). Effects of livestock farming on birds of rural areas in Europe. Biodiv. Conserv. 25, 615–631. doi:10.1007/s10531-016-1087-9
Paradis, E., and Schliep, K. (2019). Ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528. doi:10.1093/bioinformatics/bty633
Pinheiro, J., and Bates, D. (2022). Nlme: linear and nonlinear mixed effects models–157. R package version 3.1 https://CRAN.R-project.org/package=nlme.
Poorter, L., Amissah, L., Bongers, F., Hordijk, I., Kok, J., Laurance, S. G. W., et al. (2023). Successional theories. Biol. Rev. doi:10.1111/brv.12995
Pruchniewicz, D. (2017). Abandonment of traditionally managed mesic mountain meadows affects plant species composition and diversity. Basic Appl. Ecol. 20, 10–18. doi:10.1016/j.baae.2017.01.006
Pykälä, J. (2003). Effects of restoration with cattle grazing on plant species composition and richness of semi-natural grasslands. Biodiv. Conserv. 12, 2211–2226. doi:10.1023/A:1024558617080
Racz, I. A., Deri, E., Kisfali, M., Batiz, Z., Varga, K., Szabo, G., et al. (2013). Early changes of orthopteran assemblages after grassland restoration: a comparison of space-for-time substitution versus repeated measures monitoring. Biodiv. Conserv. 22, 2321–2335. doi:10.1007/s10531-013-0466-8
Rakosy, D., Motivans, E., Ştefan, V., Nowak, A., Świerszcz, S., Feldmann, R., et al. (2022). Intensive grazing alters the diversity, composition and structure of plant-pollinator interaction networks in Central European grasslands. PLoS ONE 17, e0263576. doi:10.1371/journal.pone.0263576
R Core Team (2022). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/.
Reif, J., Marhoul, P., Čížek, O., and Konvička, M. (2011). Abandoned military training sites are an overlooked refuge for at-risk open habitat bird species. Biodiv. Conserv. 20, 3645–3662. doi:10.1007/s10531-011-0155-4
Reif, J., Marhoul, P., and Koptík, J. (2013). Bird communities in habitats along a successional gradient: divergent patterns of species richness, specialization and threat. Basic Appl. Ecol. 14, 423–431. doi:10.1016/j.baae.2013.05.007
Rösch, V., Hoffmann, M., Diehl, U., and Entling, M. H. (2019). The value of newly created wood pastures for bird and grasshopper conservation. Biol. Conserv. 237, 493–503. doi:10.1016/j.biocon.2019.07.036
Šálek, M., Kalinová, K., and Reif, J. (2022). Conservation potential of semi-natural habitats for birds in intensively-used agricultural landscapes. J. Nat. Conserv. 66, 126124. doi:10.1016/j.jnc.2021.126124
Scheele, B. C., Legge, S., Armstrong, D. P., Copley, P., Robinson, N., Southwell, D., et al. (2018). How to improve threatened species management: an Australian perspective. J. Environ. Manage. 223, 668–675. doi:10.1016/j.jenvman.2018.06.084
Sizling, A. L., Sizlingova, E., Storch, D., Reif, J., and Gaston, K. J. (2009). Rarity, commonness, and the contribution of individual species to species richness patterns. Am. Nat. 174, 82–93. doi:10.1086/599305
Skokanová, H., Havlíček, M., Klusáček, P., and Martinát, S. (2017). Five military training areas – five different trajectories of land cover development? Case studies from the Czech Republic. Geogr. Cassoviensis 11, 201–213.
Somodi, I., Virágh, K., and Podani, J. (2008). The effect of the expansion of the clonal grass Calamagrostis epigejos on the species turnover of a semi-arid grassland. Appl. Veg. Sci. 11, 187–192. doi:10.3170/2008-7-18354
Spulerova, J., Bezak, P., Dobrovodska, M., Lieskovsky, J., and Stefunkova, D. (2017). Traditional agricultural landscapes in Slovakia: why should we preserve them? Landsc. Res. 42, 891–903. doi:10.1080/01426397.2017.1385749
Šťastný, K., Bejček, V., and Němec, M. (2017). Red list of the threatened species of the Czech republic. Birds. Příroda 34, 107–154.
Sutcliffe, P. R., Mellin, C., Pitcher, C. R., Possingham, H. P., and Caley, M. J. (2014). Regional-scale patterns and predictors of species richness and abundance across twelve major tropical inter-reef taxa. Ecography 37, 162–171. doi:10.1111/j.1600-0587.2013.00102.x
Tagarev, T. (2004). “From downsizing to modernising defence in C&E Europe: opportunities for SME’s,” in Defence related SME’s: analysis and description of current conditions. Editor F. D. Carvalho (Amsterdam: NATO advanced science institutes series).
Tatsumi, S., Iritani, R., and Cadotte, M. W. (2021). Temporal changes in spatial variation: partitioning the extinction and colonisation components of beta diversity. Ecol. Lett. 24, 1063–1072. doi:10.1111/ele.13720
Teleki, B., Sonkoly, J., Erdős, L., Tóthmérész, B., Prommer, M., and Török, P. (2020). High resistance of plant biodiversity to moderate native woody encroachment in loess steppe grassland fragments. Appl. Veget. Sci. 23, 175–184. doi:10.1111/avsc.12474
Těšitel, J., Mládek, J., Fajmon, K., Blažek, P., and Mudrák, O. (2018). Reversing expansion of Calamagrostis epigejos in a grassland biodiversity hotspot: hemiparasitic Rhinanthus major does a better job than increased mowing intensity. Appl. Veget. Sci. 21, 104–112. doi:10.1111/avsc.12339
Tropek, R., Černá, I., Straka, J., Čížek, O., and Konvička, M. (2013). Is coal combustion the last chance for vanishing insects of inland drift sand dunes in Europe? Biol. Conserv. 162, 60–64. doi:10.1016/j.biocon.2013.03.027
Tropek, R., Hejda, M., Kadlec, T., and Spitzer, L. (2012). Local and landscape factors affecting communities of plants and diurnal Lepidoptera in black coal spoil heaps: implications for restoration management. Ecol. Eng. 57, 252–260. doi:10.1016/j.ecoleng.2013.04.024
Volery, L., Vaz Fernandez, M., Wegmann, D., and Bacher, S. (2023). A general framework to quantify and compare ecological impacts under temporal dynamics. Ecol. Lett. 26, 1726–1739. doi:10.1111/ele.14288
Wang, G., Murphy, D., Oller, A., Howard, H. R., Anderson, A. B., Rijal, S., et al. (2014). Spatial and temporal assessment of cumulative disturbance impacts due to military training, burning, haying, and their interactions on land condition of Fort Riley. Environ. Manage. 54, 51–66. doi:10.1007/s00267-014-0284-0
Warren, M. S., Maes, D., van Swaay, C. A. M., Goffart, P., Van Dyck, H., Bourn, N. A. D., et al. (2021). The decline of butterflies in Europe: problems, significance, and possible solutions. Proc. Natl. Acad. Sci. U.S.A. 118, e2002551117. doi:10.1073/pnas.2002551117
Warren, S. D., Holbrook, S. W., Dale, D. A., Whelan, N. L., Elyn, M., Grimm, W., et al. (2007). Biodiversity and the heterogeneous disturbance regime on military training lands. Restor. Ecol. 15, 606–612. doi:10.1111/j.1526-100x.2007.00272.x
Wesche, K., Krause, B., Culmsee, H., and Leuschner, C. (2012). Fifty years of change in Central European grassland vegetation: large losses in species richness and animal-pollinated plants. Biol. Conserv. 150, 76–85. doi:10.1016/j.biocon.2012.02.015
Zentelis, R., and Lindenmayer, D. (2015). Bombing for biodiversity–enhancing conservation values of military training areas. Conserv. Lett. 8, 299–305. doi:10.1111/conl.12155
Keywords: vascular plants, grasshoppers, butterflies, birds, ecological succession, environmental management, threatened species, Europe
Citation: Reif J, Chajma P, Dvořáková L, Koptík J, Marhoul P, Čížek O and Kadlec T (2023) Biodiversity changes in abandoned military training areas: relationships to different management approaches in multiple taxa. Front. Environ. Sci. 11:1243568. doi: 10.3389/fenvs.2023.1243568
Received: 20 June 2023; Accepted: 23 October 2023;
Published: 09 November 2023.
Edited by:
Purabi Saikia, Central University of Jharkhand, IndiaReviewed by:
Alfredo Ramírez-Hernández, Instituto Potosino de Investigación Científica y Tecnológica (IPICYT), MexicoFrederico Neves, Federal University of Minas Gerais, Brazil
Copyright © 2023 Reif, Chajma, Dvořáková, Koptík, Marhoul, Čížek and Kadlec. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tomáš Kadlec, a2FkbGVjdEBmenAuY3p1LmN6