 Yan Zhou
Yan Zhou Lei Jing2†
Lei Jing2† Shaoquan Wang
Shaoquan Wang Yifei Jia
Yifei Jia- 1Co-Innovation Center for Sustainable Forestry in Southern China, College of Biology and the Environment, Nanjing Forestry University, Nanjing, China
- 2College of Forestry, Central South University of Forestry and Technology, Changsha, China
- 3School of Ecology and Nature Conservation, Beijing Forestry University, Beijing, China
Global lakes play an active role in releasing carbon into the atmosphere. However, previous research was less focused on shallow tropical and sub-tropical lakes, especially ecosystem respiration during the drawdown period. This study was designed to determine the environmental factors that determine ecosystem respiration during the drawdown period in a typical shallow sub-tropical lake, Dongting Lake in China. Ecosystem respiration from the exposed mudflat and a newly colonized meadow were investigated using a Li-8100 soil CO2 flux system in situ. The soil water content soil organic carbon (SOC), dissolved organic carbon (DOC), total nitrogen and soil C/N ratio were measured at 0–30 cm soil depth layers. No difference was found among different soil depth layers for soil properties, while the dissolved organic carbon value varied significantly among different levels of the cumulative days of the mudflat exposed to the air (CDE). Carex colonizing significantly increased soil organic carbon and DOC at the surface soil layer. Exposure to the air and Carex colonizing together strengthened the intensity of carbon dioxide (CO2) emission in the mudflat, achieving 0.716 ± 0.114 μmol m-2s-1 and 2.240 ± 0.375 μmol m-2s-1, respectively. Exposure to the air led mudflat to exceed other landscapes or different vegetation types in Dongting Lake, becoming the most active area releasing CO2 into the atmosphere, with the respiration flux reaching a peak period at around 60 days after exposure, which was enormously reinforced by Carex colonizing. Reducing the area and duration of mudflat exposure to the air during the drawdown period might be useful in reducing CO2 emissions to the atmosphere in shallow sub-tropical lakes.
1 Introduction
Global wetlands store a large proportion of the global land carbon and can play a critical role in the global carbon cycle (Mitra et al., 2005; Lal, 2008). In particular, wetlands account for 20%–30% of the global soil organic carbon (SOC) pool (Bridgham et al., 2006), and have been a net carbon sink in recent decades with an average net carbon retention rate of 118 g C m-2 yr-1 (Mitsch et al., 2013). However, changes in climate and wetland ecological features could turn them from carbon sinks into carbon sources (Oechel et al., 1993; Hadden and Grelle, 2016; Zhou et al., 2018). Freshwater bodies, such as streams, rivers and lakes, have produced a large amount of CO2 (Schrier-Uijl et al., 2011; Wang et al., 2011). It is reported that global lakes alone emit 1943 Tg CO2 to the atmosphere every year (Williamson et al., 2009). Nevertheless, most studies on wetland carbon cycles mainly focus on boreal peatlands (Fan et al., 2013; Wang et al., 2015; Karlsson et al., 2021), while research on the carbon cycle of tropical and subtropical wetlands, especially of the subtropical shallow lakes, is somewhat limited (Hu et al., 2015; Amaral et al., 2020; Calamita et al., 2021; Paranaíba et al., 2021).
Many subtropical wetlands experience seasonal cycles of water table (Carmignani and Roy, 2017). With the fluctuation of water table, some parts of the wetland soils periodically shift between an aerobic and anaerobic environment. This periodical water table fluctuation can strongly impact soil properties and the processes of subtropical wetland carbon cycle (Marcé et al., 2019; Keller et al., 2020; Machado dos Santos Pinto et al., 2020). A high water table may sustain the carbon stock of wetland by preventing aerobic respiration, whilst exposure to the air under low water table can greatly influence the CO2 emissions from the terrestrial ecosystem to the atmosphere (Couwenberg et al., 2010; Swails et al., 2022). However, the extent to which the water table fluctuation may impact soil humidity, bulk density, soil carbon and the temporal dynamics of ecosystem carbon exchanges, still remain obscure, largely due to the lack of monitoring data. Moreover, with climate change and enhanced anthropogenic activities, many subtropical wetlands have experienced substantial land transformations and land cover changes, and can permanently alter the water table level (Zhao et al., 2002; Maertens et al., 2022; Richardson et al., 2022). Such land cover changes and land transformation-induced water table level alternation, together with periodical water table cycle, have profound implications for wetland carbon balances (Hu et al., 2015; Johnson et al., 2022). In particular, ecosystem respiration, a key component of the carbon cycle, can change dramatically in response to changes in land cover and water table level (Zhou et al., 2014; Ma et al., 2022; Li et al., 2023a). Quantifying the ecosystem respiration responses to water table variations in lacustrine mudflats, therefore, is important for understanding and predicting subtropical wetland carbon cycles.
The Dongting Lake in Hunan Province, China, is a typical subtropical ephemeral lake located in the middle of the Yangtze River Basin. As one of the last two large Yangtze-connected lakes, the average monthly water level of the Dongting Lake fluctuates between 20.19 and 30.24 m (Shi et al., 2012). Consequently, the littoral areas are completely inundated in summer flood season, but are exposed to the air in autumn–winter with an annual drawdown period lasting about 4–6 months. In recent decades, due to the water volume control of the Three Gorges Dam, the hydro-condition of downstream wetlands has greatly changed (Feng et al., 2013), resulting in even longer winter drawdown periods in Dongting Lake (Lai et al., 2013). During the drawdown periods, littoral plants, such as Carex, extensively emerge and grow continuously almost throughout the drawdown period, turning mudflats into meadows (Lei et al., 2014). This mudflat-to-meadow shift during the drawdown period suggests that autotrophic respiration may also substantially contribute to the total ecosystem respiration, which has been scarcely investigated to date. The main objective of this study, therefore, is to measure and understand ecosystem respiration from both mudflats and meadows in the Dongting Lake wetlands during the winter drawdown period. The dynamics of ecosystem respiration (including both plant autotrophic respiration (Rp) and soil respiration (Rs)) can be influenced by varied biotic and abiotic factors, such as vegetation activity (Urbanova et al., 2012; Ives et al., 2013; Wang et al., 2021), the quantity and quality of soil properties (Fan et al., 2013) and soil moisture (Zhou et al., 2014). To understand how these factors may influence ecosystem respiration, we also measured soil and climate factors, including soil water content (SWC), SOC, dissolved organic carbon (DOC), total nitrogen (TN), temperature at 5 cm soil depth (T5) and air temperature in the meantime.
2 Materials and methods
2.1 Site description and site selection
Dongting Lake is the second largest freshwater lake in China located in the north of Hunan Province in central-southern China. Dongting Lake drains into the Yangtze River and is characterized with a continental subtropical monsoon humid climate. It has four distinct seasons featuring a warm and humid summer and a cold and dry winter. The long-term (1950–2012) annual mean temperature is 16.6°C–17.0°C, while the annual average precipitation is 1,200–1,400 mm (Lei et al., 2014).
We collected soil samples and measured fluxes of ecosystem respiration from 9 mudflat-sites close to the area of freshwater body during the drawdown period (Figure 1). As the area of mudflat and the spatial locations of the 9 sites vary greatly, we sampled different plots among the sites (Table 1). We collected the daily data of water level in Dongting Lake (data from website of Yangtze River hydrology, http://www.cjh.com.cn/pages/sssq.html) to determine the time of each sampling plot being exposed to the air (above the water level), and the cumulated days of exposure (CDE) at each plot were calculated. All 23 sampling plots can be divided into 3 categories based on their CDE, i.e., <30, ≈60 and >90 days. In addition, we also sampled soils and measured ecosystem respiration for seven meadow plots at the sites TED, XXH, CF and HQH (Table 1). All meadow plots were placed close to a mudflat plot, constructing one-to-one comparisons.
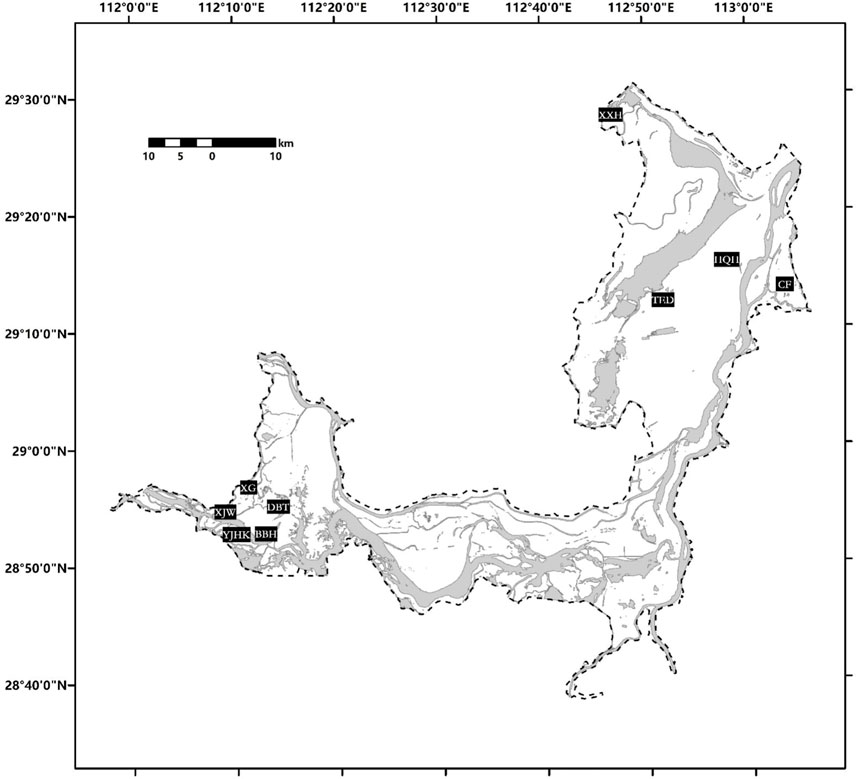
FIGURE 1. Distribution of sampling zones in Dongting Lake. The dark area shows the area of water body during the drawdown period, while the white area shows the mudflat and meadow exposing after water retreating.

TABLE 1. The replications description and the cumulated days of exposure (CDE)of sampling sites.
2.2 Ecosystem respiration measurements
Ecosystem respiration was measured on sunny days in January 2015 using a Li-8100 soil CO2 flux system with an opaque chamber (LI-COR Inc., Lincoln, NE, United States of America). We set a 20-m-long sampling line for each sampling plot, and placed three PVC collar (20cm diameter; 15 cm height) equal-distance along each sampling line with the collars installed 2–3 cm above ground. The collars were installed at least 3 days before the measurement to minimize human disturbances. Each measurement lasted 2 min, meanwhile CO2 flux is calculated using linear fit. T5 and air temperature were measured adjacent to each respiration collar with a portable temperature probe provided with the Li-8100. SWC at 5 cm depth was also measured with a portable probe provided with the Li-8100 at a point close to each chamber.
2.3 Soil properties
We also collected soil samples at 0–10, 10–20 and 20–30 cm depth at each sampling plot close to the middle collar in January 2015. In order to fully assess the impact of soil properties on ecosystem respiration, we measured the following soil properties: SWC, TN, DOC, SOC and C/N. Soil samples were divided into three sets. The first set of soil was air dried for TN and SOC measurements. The SOC of dried samples was measured with a potassium-dichromate oxidation procedure after H2SO4–HClO4 digestion (Semenov et al., 2010). Additionally, the TN of dried samples was measured using a flow injection analyzer (Star5000,FOSS, Denmark). The second set of soil was remained fresh for DOC measurements. The DOC of the fresh samples was measured with an Organic Carbon Analyzer (TOC-vwp, SHIMADZU, Japan). The third set of soil samples was weighed in the field, and then dried to a constant weight. The difference between the two weights is the weight of water, and SWC was calculated as the ratio of the weight of water content to the constant weight of the dry soil sample.
2.4 Statistical analyses
All the statistical analyses were conducted with the SPSS program version 18.0. Non-parametric statistics were used if the data failed the test for normality. The impacts of soil depth and CDE on soil properties were analyzed with two-way ANOVA. We used the Kruskal–Wallis test and Dunn’s multiple comparison test to check the change in soil properties among different soil layers and different CDE levels. We also applied the Wilcoxon signed rank test to check the differences of soil properties and ecosystem respirations between mudflats and meadows. The differences of ecosystem respiration among different CDE levels were also tested using Dunn’s multiple comparison test. It is reported that soil CO2 flux significantly correlates with soil properties of surface soil (Liu et al., 2013a), while deep soil seldom determines soil CO2 flux (Luan et al., 2011; Zhou et al., 2014). Thus, we applied a stepwise regression model to determine important surface soil properties and other environmental factors, including T5, air temperature and CDE in controlling ecosystem respiration. The statistical relationships between the natural logarithm of ecosystem respiration and T5 from the meadow, and the ecosystem respiration and DOC from the mudflat, were examined with linear regression analyses.
3 Results
3.1 Soil properties
A two-way ANOVA test showed that the soil depth and the interaction between soil depth and CDE do not significantly influence soil properties, while CDE only significantly influences DOC (Table 2).
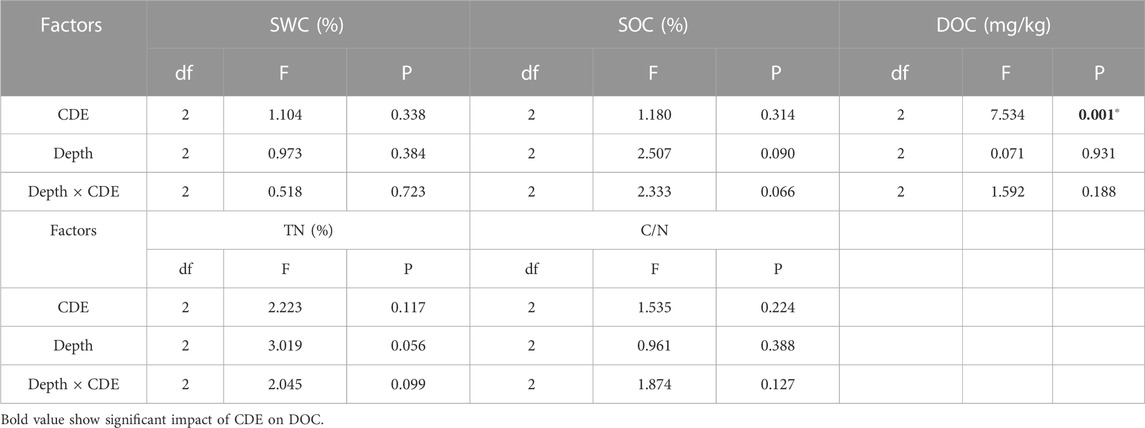
TABLE 2. Impacts of soil depth and CDE on soil properties.
No significant differences were found among the values of soil properties at different soil layers (Figure 2). Only DOC at the highest CDE level (>90 days) was significantly higher than that of lower CDE levels (<30 days and ≈60 days) (Dunn’s multiple comparison test; p < 0.01 and p < 0.01, respectively) (Figure 3).
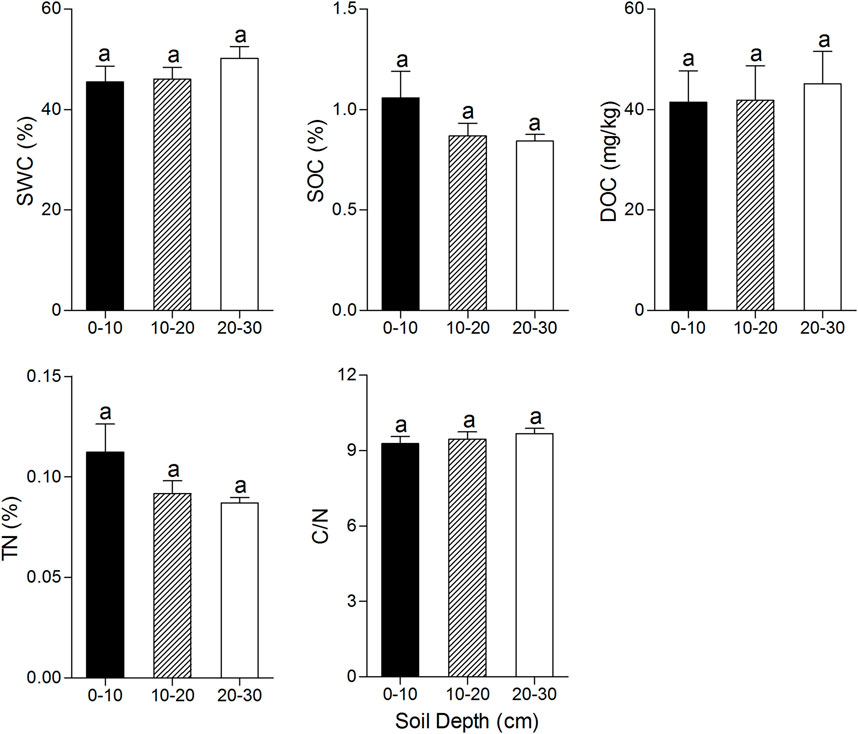
FIGURE 2. Soil properties (mean ± SE) at 3 layers of soil depth. The values of each soil property with the same lowercase letter do not differ significantly (p > 0.05) based on Dunn’s multiple comparison test.
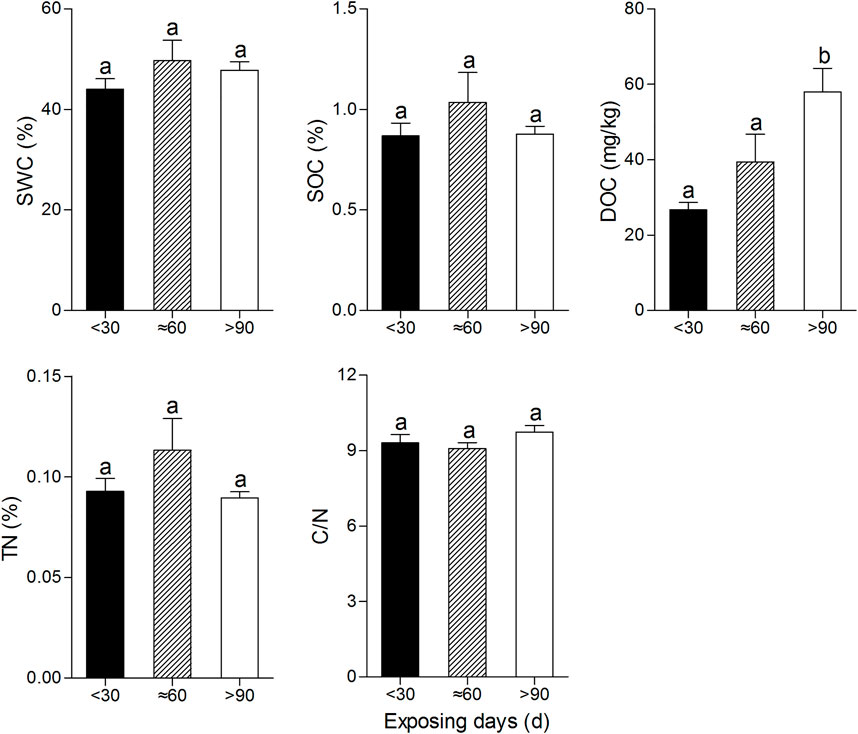
FIGURE 3. Soil properties at 3 categories of CDE. The values of each soil property with the same lowercase letter do not differ significantly (p > 0.05) based on Dunn’s multiple comparison test.
Between the paired mudflat–meadow plots, the values of SWC, SOC, DOC, TN and C/N showed no significant differences at soil layers deeper than 10 cm, while the values of SOC, DOC and C/N were significantly higher in meadows than in mudflats at the 0–10cm soil depth (Wilcoxon signed rank test; p = 0.04, p = 0.01 and p = 0.01, respectively) (Figure 4).
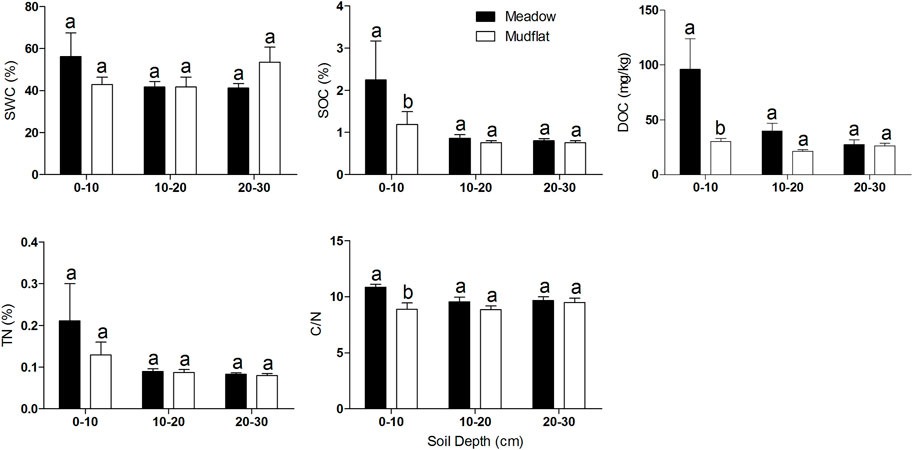
FIGURE 4. The differences of soil properties between meadow and mudflat. The values of each soil property with the same lowercase letter do not differ significantly (p > 0.05) based on the Wilcoxon signed rank test.
3.2 Ecosystem respiration
The mean value of ecosystem respiration for air-exposed mudflats in Dongting Lake is 0.716 ± 0.114 μmol m-2s-1, while it is a much higher value of 2.240 ± 0.375 μmol m-2s-1 for meadows. A further Wilcoxon signed rank test for the paired mudflat-meadow ecosystem respiration comparison confirms that ecosystem respiration at meadow plots are significantly higher than that at mudflat plots (Wilcoxon signed rank test, p = 0.01).
3.3 Environmental impact on ecosystem respiration
With the CDE increasing, the flux of ecosystem respiration from the mudflat first increases (for about 2 months) and then decreases, but without significant difference found among fluxes at different CDE levels (Figure 5).
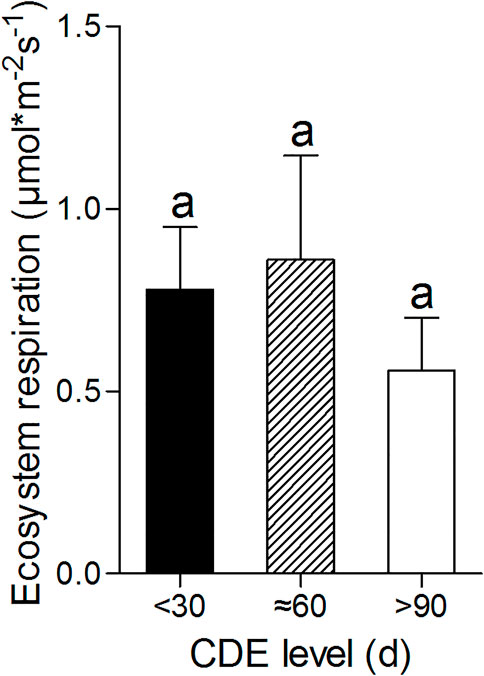
FIGURE 5. The differences of ecosystem respiration among different CDE levels. The values of fluxes with the same lowercase letter do not differ significantly (p > 0.05) based on Dunn’s multiple comparison test.
The stepwise regression analysis shows that only DOC significantly influences the ecosystem respiration of mudflats in Dongting Lake (Figure 6A,y = 0.010x+0.309,N = 23,R2 = 0.280,p < 0.01), and only T5 significantly influences the ecosystem respiration of meadows (Figure 6B, y = 0.358x-3.470,N = 7,R2 = 0.780,p < 0.01).
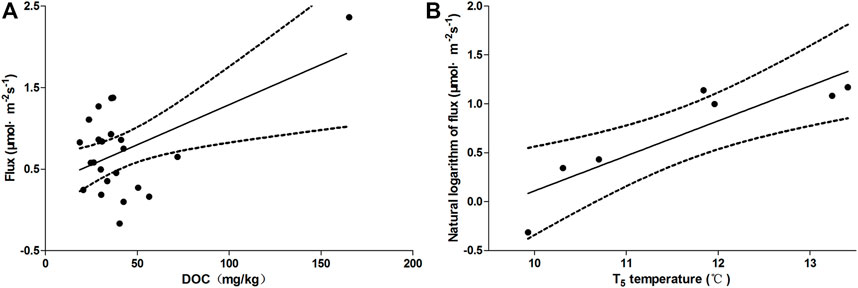
FIGURE 6. The correlation between ecosystem respiration and soil properties. (A) the correlation between ecosystem respiration from mudflat and DOC; (B) the correlation between natural logarithm of ecosystem respiration from meadow and T5 temperature.
4 Discussion
The sampling zones in this study were located close to the open water body during the drawdown period. With the water level fluctuating in Dongting Lake all the time, sampling zones exposed to the air were usually saturated during the drawdown period, which induced SWC at the surface soil that was almost same as that at the deep soil depth (Figure 2). Stable SWC among three layers of soil led to somewhat stable SOC and DOC between the surface and deep soil. It was reported that the decomposition of SOC is greatly reduced in anaerobic environments, such as marsh and swampland (Luo and Zhou, 2006). A higher value of SWC creates a more anaerobic environment, restraining soil respiration and advancing carbon sequestration in soil (Hudson, 1994). However, SWC showed the trend of a lower value at the surface soil, which is not related to the slightly higher SOC value at 0–10 cm depth soil in this study (Figure 2). Consequently, it is concluded that sedimentation during the flooded period brings external organic matter to the surface soil (Dai et al., 2005). Using the system of Surface elevation table, the height of the lake basin in the Dongting Lake area was monitored. The average annual sedimentation in the past 5 years is approximately 7–8cm (personal communication), confirming the influence of exogenous organic matter on the surface soil. TN also showed the trend of a higher value at the surface layer than the deep layer due to the deposition of exogenous nitrogen during the drawdown period.
The content of SOC and TN in the soil was positively correlated with the time of mudflat exposure in the short term due to the supplementation of exogenous organic matter. With the intensification of CO2 producing a reaction, nitrification and denitrification, the contents of SOC and TN (Gu et al., 2004) began to decline in the long term (Figure 3).
The value of C/N is one of the most important factors which determine the mineralization rate in soil. In this study, the C/N value was around nine, not significantly influenced by soil depth and CDE, which reflected that nitrogen is not the limit factor for soil mineralization in Dongting Lake. With the emergence of mudflat, soil microbial metabolic activities gradually increased, resulting in a significant increase in soil DOC content with the increase in exposure time (Figure 3).
It is believed that water content, temperature and other environmental parameters interact to determine the CO2 release from the soil (Juszczak et al., 2013; Keller et al., 2020). In this study, only DOC was found to be significantly correlated with soil respiration at the mudflat, mostly due to organic carbon dissolved in the surface soil supplying a substrate for CO2 production. When soil was saturated at the mudflats in Dongting Lake during the drawdown period, the mechanism of the CO2 cycle was limited by the amount of active substrate that could be estimated by the DOC value.
During the drawdown period in Dongting Lake, the surface soil is almost fully exposed to the CO2 reaction and consumption of DOC, making DOC at the surface soil slightly lower than in the deep soil (Figure 2). With the increase in the exposure time of the mudflat, the consumption of DOC could significantly decrease the respiratory flux from the mudflat. However, no significant correlation was found between CDE and the respiratory flux from the mudflat, which is derived from the increasing DOC induced by the organic matter deposition during the flooded period and the enhancement of microbial metabolism in the soil after mudflat exposure (Vonk et al., 2017). At the deeper soil, the DOC content increased with the increase in CDE (Figure 7), continuously supplying DOC for ecosystem respiration. Consequently, the substrate for ecosystem respiration is sufficient, intensifying ecosystem respiration shortly after exposure, then resulting in a gradual decrease in DOC at the surface soil depth after a short-term increase (Figure 7). Therefore, due to the accumulation of exogenous organic matter and the increase in soil microbial metabolism, the respiration substrate of the soil increased with the increase in CDE in the short term, resulting in a gradual increase in the respiration intensity of the sediment, which reached a high value at around 60 days. Followed by the continuous consumption of DOC, the respiratory intensity decreased gradually (Figure 5). How DOC determines the respiration process was also reflected by the significant correlation between respiration flux and DOC value (Figure 6).
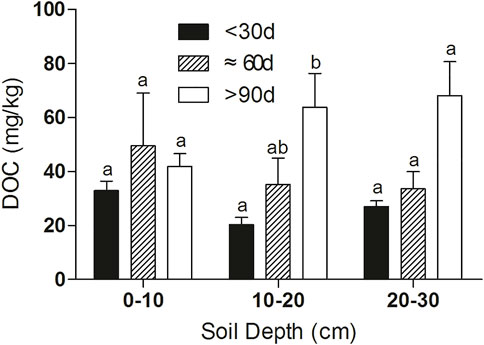
FIGURE 7. DOC value at different soil layers and CDE levels. The values of DOC with the same lowercase letter do not differ significantly (p > 0.05) based on Dunn’s multiple comparison test.
When vegetation colonized, the root system interspersed, squeezed, divided the soil and absorbed soil moisture, creating a more aerobic environment at the surface soil layer. In Dongting Lake, no significantly difference was found between the soil properties from the meadow and the mudflat at the deeper soil layer, also suggesting that vegetation colonizing only impacts soil properties at the surface layer during the drawdown period by changing the aeration and generating more external organic matter (Jobbágy and Jackson, 2000; Tuo et al., 2018). The values of SOC, DOC and C/N at the surface layer were significantly higher in the meadow than the mudflat, reflecting the fact that the dominant Carex produces more organic carbon by photosynthesis and maintains aerobic environment advancing respiration. Thus, organic carbon was not the limit factor for respiration, which was merely determined by the soil temperature instead in the meadow (Figure 6). Furthermore, due to the lower value of C/N in Dongting Lake, SOC could be consumed as a substrate for soil respiration and root respiration in the short term. Consequently, it was found that the ecosystem respiration flux from the meadow was three times greater than that from mudflat.
Due to the exposure of the mudflat and the improvement of soil ventilation, the mean value of the respiratory flux from the mudflat of Dongting Lake in this study was 0.716 ± 0.114 μmol m-2s-1. This was far higher than the CO2 flux of 0.2 μmol m-2s-1 in maximum from the terrestrial–aquatic interface in Chinese subtropical shallow lakes (Liu et al., 2013b; Xiao, 2014; Xiao et al., 2020; Li et al., 2023b). It is concluded that more SOC in the lake basin is involved in the respiration process after exposure to the air, significantly emitting more CO2 into the atmosphere.
A previous study stated that fluxes from poplars, reeds and farmlands in Dongting Lake during the drawdown period were 0.70, 0.56 and 0.45 μmol m-2s-1, respectively (Tang, 2011), much lower than that from the Carex meadow and slightly lower than that from the exposed mudflat, and this is the opposite in the flooding season. During the flooding season, fluxes from poplars, reeds and farmlands in Dongting Lake were 4.80, 8.21 and 1.27 μmol m-2s-1, respectively, significantly higher than the flux from the terrestrial–aquatic interface and the flux from water surface. Consequently, the exposed mudflat, instead of poplar, reed or farmland, became the most active area of CO2 emissions during the drawdown period. The intensity of respiration flux was strengthened after Carex colonizing. Reducing the area and duration of mudflat without littoral plants exposure to the air by manual control may be conducive to reducing CO2 emissions to the atmosphere.
5 Conclusion
With CDE increasing during the drawdown period in Dongting Lake, exposure to the air and Carex colonizing together increased the soil DOC value in exposed mudflat, strengthening the intensity of ecosystem respiration. DOC was the limit factor of ecosystem respiration that reached a peak period around 60 days after exposure of the mudflat, while only soil temperature determined respiration in the newly colonized meadow with a sufficient supply of organic carbon. The ecosystem respiration value reached 0.716 ± 0.114 μmol m-2s-1 and 2.240 ± 0.375 μmol m-2s-1 in the exposed mudflat and newly colonized meadow, respectively, both of which became the most active areas of CO2 emissions during the drawdown period.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
Investigation: YZ, LJ and YJ; data curation: YZ, LJ; writing—original draft preparation: YZ, YW, and YJ; writing—review: YZ,GL; visualization: SW; supervision: YZ and GL.
Funding
This research was funded by Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), the Primary Research & Development Plan of Jiangsu Province (BE2019773) and China National Forestry Public Welfare Fund (no. 200804005).
Acknowledgments
We thank the staff of the Management Bureau of the East Dongting Lake National Nature Reserve and the Western Dongting Lake National Nature Reserve for their support in the field work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Amaral, J. H. F., Melack, J. M., Barbosa, P. M., MacIntyre, S., Kasper, D., Cortes, A., et al. (2020). Carbon dioxide fluxes to the atmosphere from waters within flooded forests in the amazon basin. J. Geophys. Research-Biogeosciences 125 (3). doi:10.1029/2019jg005293
Bridgham, S. D., Megonigal, J. P., Keller, J. K., Bliss, N. B., and Trettin, C. (2006). The carbon balance of north American wetlands. Wetlands 26 (4), 889–916. doi:10.1672/0277-5212(2006)26[889:tcbona]2.0.co;2
Calamita, E., Siviglia, A., Gettel, G. M., Franca, M. J., Winton, R. S., Teodoru, C. R., et al. (2021). Unaccounted CO2 leaks downstream of a large tropical hydroelectric reservoir. Proc. Natl. Acad. Sci. 118 (25), e2026004118. doi:10.1073/pnas.2026004118
Carmignani, J. R., and Roy, A. H. (2017). Ecological impacts of winter water level drawdowns on lake littoral zones: A review. Aquat. Sci. 79 (4), 803–824. doi:10.1007/s00027-017-0549-9
Couwenberg, J., Dommain, R., and Joosten, H. (2010). Greenhouse gas fluxes from tropical peatlands in south-east Asia. Glob. Change Biol. 16 (6), 1715–1732. doi:10.1111/j.1365-2486.2009.02016.x
Dai, S. B., Yang, S. L., Zhu, J., Gao, A., and Li, P. (2005). The role of Lake Dongting in regulating the sediment budget of the Yangtze River. Hydrology Earth Syst. Sci. 9 (6), 692–698. doi:10.5194/hess-9-692-2005
Fan, Z. S., McGuire, A. D., Turetsky, M. R., Harden, J. W., Waddington, J. M., and Kane, E. S. (2013). The response of soil organic carbon of a rich fen peatland in interior Alaska to projected climate change. Glob. Change Biol. 19 (2), 604–620. doi:10.1111/gcb.12041
Feng, L., Hu, C., Chen, X., and Zhao, X. (2013). Dramatic inundation changes of China's two largest freshwater lakes linked to the Three Gorges Dam. Environ. Sci. Technol. 47 (17), 9628–9634. doi:10.1021/es4009618
Gu, L., Post, W. M., and King, A. W. (2004). Fast labile carbon turnover obscures sensitivity of heterotrophic respiration from soil to temperature: A model analysis. Glob. Biogeochem. Cycle 18 (1), 1022–1032. doi:10.1029/2003gb002119
Hadden, D., and Grelle, A. (2016). Changing temperature response of respiration turns boreal forest from carbon sink into carbon source. Agric. For. Meteorology 223, 30–38. doi:10.1016/j.agrformet.2016.03.020
Hu, Q., Wu, Q., Yao, B., and Xu, X. (2015). Ecosystem respiration and its components from a Carex meadow of Poyang Lake during the drawdown period. Atmos. Environ. 100, 124–132. doi:10.1016/j.atmosenv.2014.10.047
Hudson, B. D. (1994). Soil organic matter and available water capacity. J. Soil Water Conservation 49 (2), 189–194.
Ives, S. L., Sullivan, P. F., Dial, R., Berg, E. E., and Welker, J. M. (2013). CO2 exchange along a hydrologic gradient in the kenai lowlands, AK: Feedback implications of wetland drying and vegetation succession. Ecohydrology 6 (1), 38–50. doi:10.1002/eco.274
Jobbágy, E. G., and Jackson, R. B. (2000). The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 10 (2), 423–436. doi:10.1890/1051-0761(2000)010[0423:tvdoso]2.0.co;2
Johnson, A. M., Atkinson, R. B., Steven, J. C., Whiting, G., Napora, K., Sharrett, L., et al. (2022). Determining carbon dioxide emission response in soil microcosms from a shallow mid-atlantic peatland: The influence of water table restoration. J. Coast. Res. 38 (2), 261–268. doi:10.2112/jcoastres-d-21-00079.1
Juszczak, R., Humphreys, E., Acosta, M., Michalak-Galczewska, M., Kayzer, D., and Olejnik, J. (2013). Ecosystem respiration in a heterogeneous temperate peatland and its sensitivity to peat temperature and water table depth. Plant Soil 366 (1-2), 505–520. doi:10.1007/s11104-012-1441-y
Karlsson, J., Serikova, S., Vorobyev, S. N., Rocher-Ros, G., Denfeld, B., and Pokrovsky, O. S. (2021). Carbon emission from Western Siberian inland waters. Nat. Commun. 12 (1), 825. doi:10.1038/s41467-021-21054-1
Keller, P. S., Catalán, N., von Schiller, D., Grossart, H. P., Koschorreck, M., Obrador, B., et al. (2020). Global CO2 emissions from dry inland waters share common drivers across ecosystems. Nat. Commun. 11 (1), 2126. doi:10.1038/s41467-020-15929-y
Lai, X., Jiang, J., and Huang, Q. (2013). Effects of the normal operation of the three Gorges reservoir on wetland inundation in Dongting Lake, China: A modelling study. Int. Assoc. Sci. Hydrology Bull. 58 (7), 1467–1477. doi:10.1080/02626667.2013.831418
Lal, R. (2008). Carbon sequestration. Philos. Trans. Biol. Sci. 363 (1492), 815–830. doi:10.1098/rstb.2007.2185
Lei, G., Li, W., Feng, D., Hong, Z., and Lei, G. (2014). Delayed flood recession in central Yangtze floodplains can cause significant food shortages for wintering geese: Results of inundation experiment. Environ. Manage. 54 (6), 1331–1341. doi:10.1007/s00267-014-0350-7
Li, G., Xiao, K., Wang, Q. Q., Zhang, Y., Li, H. T., and Li, H. L. (2023). The regulating role of meteorology in the wetland-air CO2 fluxes at the largest shallow grass-type lake on the north China plain. Water 15 (1), 139. doi:10.3390/w15010139
Li, J., Jiang, M., Pei, J., Fang, C., Li, B., and Nie, M. (2023). Convergence of carbon sink magnitude and water table depth in global wetlands. Ecol. Lett [Epub ahead of print]. doi:10.1111/ele.14199
Liu, L. X., Ming, X., Mao, L., and Xin, Z. (2013). Spatial variability of greenhouse gas effluxes and their controlling factors in the poyang Lake in China. Pol. J. Environ. Stud. 22 (3), 749–758.
Liu, Y. S., Fan, J. W., Harris, W., Shao, Q. Q., Zhou, Y. C., Wang, N., et al. (2013). Effects of plateau pika (Ochotona curzoniae) on net ecosystem carbon exchange of grassland in the Three Rivers Headwaters region, Qinghai-Tibet, China. Plant Soil 366 (1-2), 491–504. doi:10.1007/s11104-012-1442-x
Luan, J. W., Liu, S. R., Zhu, X. L., and Wang, J. X. (2011). Soil carbon stocks and fluxes in a warm-temperate oak chronosequence in China. Plant Soil 347 (1-2), 243–253. doi:10.1007/s11104-011-0842-7
Ma, L., Zhu, G. F., Chen, B. L., Zhang, K., Niu, S. L., Wang, J. S., et al. (2022). A globally robust relationship between water table decline, subsidence rate, and carbon release from peatlands. Commun. Earth Environ. 3 (1), 254. doi:10.1038/s43247-022-00590-8
Machado dos Santos Pinto, R., Weigelhofer, G., Diaz-Pines, E., Guerreiro Brito, A., Zechmeister-Boltenstern, S., and Hein, T. (2020). River-floodplain restoration and hydrological effects on GHG emissions: Biogeochemical dynamics in the parafluvial zone. Sci. Total Environ. 715, 136980. doi:10.1016/j.scitotenv.2020.136980
Maertens, M., De Lannoy, G. J. M., Vincent, F., Massart, S., Gimenez, R., Houspanossian, J., et al. (2022). Spatial patterns of soil salinity in the central Argentinean Dry Chaco. Anthropocene 37, 100322. doi:10.1016/j.ancene.2022.100322
Marcé, R., Obrador, B., Gómez-Gener, L., Catalán, N., Koschorreck, M., Arce, M. I., et al. (2019). Emissions from dry inland waters are a blind spot in the global carbon cycle. Earth-Science Rev. 188, 240–248. doi:10.1016/j.earscirev.2018.11.012
Mitra, S., Wassmann, R., and Vlek, P. L. G. (2005). An appraisal of global wetland area and its organic carbon stock. Currentence 88 (1), 25–35.
Mitsch, W. J., Bernal, B., Nahlik, A. M., Mander, Ü., Zhang, L., Anderson, C. J., et al. (2013). Wetlands, carbon, and climate change. Landsc. Ecol. 28 (4), 583–597. doi:10.1007/s10980-012-9758-8
Oechel, W. C., Hastings, S. J., Vourlrtis, G., Jenkins, M., Riechers, G., and Grulke, N. (1993). Recent change of Arctic tundra ecosystems from a net carbon dioxide sink to a source. Nature 361 (6412), 520–523. doi:10.1038/361520a0
Paranaíba, J. R., Barros, N., Almeida, R. M., Linkhorst, A., Mendonça, R., Vale, R. d., et al. (2021). Hotspots of diffusive CO2 and CH4 emission from tropical reservoirs shift through time. J. Geophys. Res. Biogeosciences 126 (4), e2020JG006014. doi:10.1029/2020jg006014
Richardson, C. J., Flanagan, N. E., Wang, H. J., and Ho, M. C. (2022). Annual carbon sequestration and loss rates under altered hydrology and fire regimes in southeastern USA pocosin peatlands. Glob. Change Biol. 28 (21), 6370–6384. doi:10.1111/gcb.16366
Schrier-Uijl, A. P., Veraart, A. J., Leffelaar, P. A., Berendse, F., and Veenendaal, E. M. (2011). Release of CO2 and CH4 from lakes and drainage ditches in temperate wetlands. Biogeochemistry 102 (1-3), 265–279. doi:10.1007/s10533-010-9440-7
Semenov, M. V., Kravchenko, I. K., Semenov, V. M., Kuznetsova, T. V., Dulov, L. E., Udal'tsov, S. N., et al. (2010). Carbon dioxide, methane, and nitrous oxide fluxes in soil catena across the right bank of the oka river (moscow oblast). Eurasian Soil Sci. 43 (5), 541–549. doi:10.1134/s1064229310050078
Shi, X., Xiao, W. H., W, Y., and W, X. (2012). Characteristics and factors of water level variations in the Dongting Lake during the recent 50 years. South-to-North Water Transfers Water Sci. Technol. 10 (5), 18–22.
Swails, E. E., Ardon, M., Krauss, K. W., Peralta, A. L., Emanuel, R. E., Helton, A. M., et al. (2022). Response of soil respiration to changes in soil temperature and water table level in drained and restored peatlands of the southeastern United States. Carbon Balance Manag. 17 (1), 18. doi:10.1186/s13021-022-00219-5
Tang, J. (2011). Study on soil respiration characteristics for typical land-use types on the beach land of Dongting Lake. Changsha, Hunan, China: Thesis, Central South University of Forestry and Technology.
Tuo, D. F., Gao, G. Y., Chang, R. Y., Li, Z. S., Ma, Y., Wang, S., et al. (2018). Effects of revegetation and precipitation gradient on soil carbon and nitrogen variations in deep profiles on the Loess Plateau of China. Sci. Total Environ. 626, 399–411. doi:10.1016/j.scitotenv.2018.01.116
Urbanova, Z., Picek, T., Hajek, T., Bufkova, I., and Tuittila, E. S. (2012). Vegetation and carbon gas dynamics under a changed hydrological regime in central European peatlands. Plant Ecol. divers. 5 (1), 89–103. doi:10.1080/17550874.2012.688069
Vonk, J. A., Rombouts, T., Schoorl, J. C., Serne, P., Westerveld, J. W., Cornelissen, P., et al. (2017). Impact of water drawdown and rewetting on sediment nutrient-dynamics in a constructed delta-lake system (oostvaardersplassen, The Netherlands): A mesocosm study. Ecol. Eng. 108, 396–405. doi:10.1016/j.ecoleng.2017.06.017
Wang, D., Chen, J., Felton, A. J., Xia, L., Zhang, Y., Luo, Y., et al. (2021). Post-fire co-stimulation of gross primary production and ecosystem respiration in a meadow grassland on the Tibetan Plateau. Agric. For. Meteorol. 2021, 303.
Wang, D., Wu, G.-L., Liu, Y., Yang, Z., and Hao, H.-M. (2015). Effects of grazing exclusion on CO2 fluxes in a steppe grassland on the Loess Plateau (China). Ecol. Eng. 83, 169–175. doi:10.1016/j.ecoleng.2015.06.017
Wang, F., Wang, B., Liu, C.-Q., Wang, Y., Guan, J., Liu, X., et al. (2011). Carbon dioxide emission from surface water in cascade reservoirs-river system on the Maotiao River, southwest of China. Atmos. Environ. 45 (23), 3827–3834. doi:10.1016/j.atmosenv.2011.04.014
Williamson, C. E., Saros, J. E., Vincent, W. F., and Smold, J. P. (2009). Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol. Oceanogr. 54 (6), 2273–2282. doi:10.4319/lo.2009.54.6_part_2.2273
Xiao, Q. T. (2014). Study on greenhous gases (CO2, CH4 and N2O) fluxes of water-air interface in Lake Taihu. Thesis. Nanjing, Jiangsu, China: Nanjing University of Information Science and Technology.
Xiao, Q. T., Xu, X. F., Duan, H. T., Qi, T. C., Qin, B. Q., Lee, X., et al. (2020). Eutrophic Lake taihu as a significant CO2 source during 2000-2015, 170.Water Res.
Zhao, S., Fang, J., Chen, A., and Cui, H. (2002). Land use/cover changes in Dongting Lake area between 1955-1998. Resour. Environ. Yangtze Basin 11 (6), 536–542.
Zhou, Y., Jiao, S., Li, N., Grace, J., Yang, M., Lu, C., et al. (2018). Impact of plateau pikas (Ochotona curzoniae) on soil properties and nitrous oxide fluxes on the Qinghai-Tibetan Plateau. PLoS One 13 (9), e0203691. doi:10.1371/journal.pone.0203691
Keywords: dongting lake, mudflat, drawdown, ecosystem respiration, carex colonizing, soil
Citation: Zhou Y, Jing L, Wang S, Jia Y, Wang Y and Lei G (2023) Drivers for ecosystem respiration during the drawdown period in Dongting Lake, China. Front. Environ. Sci. 11:1181894. doi: 10.3389/fenvs.2023.1181894
Received: 08 March 2023; Accepted: 29 March 2023;
Published: 10 April 2023.
Edited by:
Jianghua Wu, Memorial University of Newfoundland, CanadaReviewed by:
Ming Jiang, Northeast Institute of Geography and Agroecology (CAS), ChinaDong Wang, Henan University, China
Copyright © 2023 Zhou, Jing, Wang, Jia, Wang and Lei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yan Zhou, emhvdXlhbi5lY29AZm94bWFpbC5jb20=
†These authors have contributed equally to this work