 Juan Qin
Juan Qin Ye Liu
Ye Liu- College of Resources and Environment, Anhui Agricultural University, Hefei, China
Changes in slope aspect have important effects on the C, N and P nutrient contents and stoichiometric characteristics of plants and soils. As an important forest type in the subtropical region, Pinus massoniana forests play an important role in the restoration and development of forest ecosystems. In this study, the effect of slope aspect on the C, N and P contents and stoichiometry of leaves and soils of four P. massoniana forest types (i.e., pure P. massoniana forest (PF), P. massoniana-Liquidambar formosana mixed forest (PLM), P. massoniana -Platycarya strobilacea mixed forest (PPM), and P. massoniana-Quercus acutissima mixed forest (PQM)) in north subtropical region of China were studied. The results showed that slope aspect had significant effects on soil C, N and P contents, leaf N content (LNC), soil C:P and N:P ratios, and leaf C:N and N:P ratios in four P. massoniana forests. SOC, STN and STP contents differed significantly on the two slopes, with the three mixed forests having higher contents on the sunny slope than on the shady slope. Among the four forest types, the SOC, STN and STP contents of PQM were the highest on both slopes, and their soil C:P and N:P ratios were all the highest on the sunny slope. The LNC and leaf N:P ratios were higher on shady slope than on sunny slope, while the leaf C:N ratio was higher on sunny slope than on shady slope. At the species level, leaf N content and N:P ratio in the three deciduous species were higher than in the evergreen species of P. massoniana, while leaf C content and C:N ratio were lower than in P. massoniana. The interaction between slope aspect and forest type caused a certain range of fluctuations in stoichiometry of the four forest types, but the overall ecological stoichiometry remained stable, and plant growth of the four forest types in the study area was limited by N. Slope-induced changes in soil nutrient content of the four P. massoniana forest types directly affect leaf C:N:P stoichiometry, and soil N and P contents are key factors affecting leaf stoichiometry, especially soil P content.
1 Introduction
Ecological stoichiometry is a science that studies the energy balance and the cycle of C, N, P and other chemical elements in the ecosystem, providing a new idea for the study of the relationship between the ecosystem material cycles, organisms and the environment (Sterner and Elser, 2002; Cleveland and Liptzin, 2007; Shi et al., 2016). C, N and P are the three fundamental nutrients for plant growth and play an important role in the plant function and structure in terrestrial ecosystems (Reich et al., 1997). As the key indicators of soil fertility, C, N and P in forest soils are also indispensable nutrient sources for plant growth, and their important ecological functions cannot be ignored (Fan et al., 2015). Thus, the stoichiometric characteristics of C, N and P in plants are usually closely coupled with the uptake of these elements in soils. Therefore, studying the stoichiometric characteristics of C, N and P in plants and soil can help reveal the ecological strategies of plants in response to different soil conditions, elucidate the plant-soil relationship, and understand the nutrient limitation of plants (Elser et al., 2000b).
Topography can influence the exchange of materials and energy between the soil and the environment, and affect the distribution and accumulation of soil nutrients with other soil-forming factors (Sardans et al., 2012). Slope aspect is the main topographic factor controlling the spatial distribution of solar energy, water balance, temperature, vegetation composition and distribution patterns, thus affecting almost all biological and ecological processes (Halim and Normaniza, 2015). As an important part of forest ecosystem, soil plays a key role in the growth of forest plants. The dynamic changes of soil C, N and P contents directly affect the composition and distribution of vegetation communities (Zhang et al., 2013). Therefore, the study of soil stoichiometric characteristics can reveal the nutrient availability to plants and provide a better understanding of the mechanisms of nutrient element cycling and balance (Xu et al., 2015). In recent years, great progress has been made in the study of soil and leaf C:N:P stoichiometry under different topographies in terrestrial ecosystems (Yimer et al., 2006; He et al., 2016; Heuck and Spohn, 2016). Many studies have found that the slope aspect has a strong influence on soil physical (e.g., temperature and moisture) and chemical properties (nutrient content and mineralization, etc.) (Sternberg and Shoshany, 2010; Lenka et al., 2013). Soil water content on the shady slope was significantly higher than that on the sunny slope at the same elevation (Qin et al., 2016), and soil nutrient content, C:N:P stoichiometry and plant community composition also differed significantly among different slope aspects (Yang J. et al., 2020). Ai et al. (2017) showed that slope aspect had a significant effect on soil nutrient content, and soil C, N contents and C:P, N:P ratios reaching a maximum value on the sunny slope. Gao et al. (2020) reported that soil C:P and N:P ratios of Abies Georgei var. Smithii were lower in natural forests on sunny slope than on shady slope, except for the soil C:N ratio. Niu et al. (2020) found that the slope aspect had an essential effect on leaf C:N:P stoichiometry, and leaf C content, C:P and N:P ratios on sunny slope were significantly higher than those on shade slope, while the leaf N content and C:N ratio did not differ significantly between the two slope aspects. The above studies indicated that slope aspect significantly affects the characteristics of soil nutrient cycling, and thus affects the growth and distribution of vegetation (Rong et al., 2015).
Pinus massoniana is a vital coniferous tree species with wide distribution, strong drought resistance, good timber quality, easy establishment and low requirements for soil environmental. It is often used as the preferred tree species for ecological restoration and afforestation in the low-hilly areas of southern China. It makes an important contribution to regional ecological functions and economic development (Yang Z. Q. et al., 2020). However, due to long-term large-scale multi-generation continuous cropping, P. massoniana plantations are prone to ecological problems such as decreased fertility, low productivity, loss of species diversity and soil nutrient deficiencies (Guan et al., 2015). Therefore, the introduction of native broad-leaved species to establish coniferous and broad-leaved mixed forests is one of the effective measures to improve nutrient cycling and achieve sustainable productivity in inefficiency plantations. Studies have shown that the mixing of P. massoniana and broad-leaved trees can enhance the state of forest soil fertility reduction caused by the slow decomposition rate of its conifer litter in pure P. massoniana forest, which can improve the forest ecosystem productivity and enhance the ecological benefits of forests (Zhang Y. et al., 2021). However, how do different P. massoniana mixed forests affect soil nutrient contents, and the effect of slope aspect on plant leaf and soil C, N and P nutrient contents and stoichiometric characteristics in different P. massoniana forest types in north subtropical China remain unclear.
Based on these, two slope aspects (sunny and shade slopes) were selected and the influence of slope aspect on the C, N, P contents and stoichiometric characteristics in leaves and soils of four typical P. massoniana forest types in the Dabie Mountains region were studied. Specifically, we addressed the following objectives: 1) explore the characteristics of the variation in leaf and soil C, N, P contents and stoichiometry of four P. massoniana forest types in the study area; 2) clarify the effects of different slope aspects on C, N and P contents and stoichiometry in leaves and soils; and 3) reveal the coupling relationships of C, N and P stoichiometric characteristics between leaves and soil as well as the nutrient limitation of the four P. massoniana forest types. The results provide information for the characterization of C, N and P stoichiometric characteristics in leaves and soils of four P. massoniana forest types in the Dabie Mountains region, and also help to understand the different plant adaptation strategies of P. massoniana, Liquidambar formosana, P. strobilacea and Quercus acutissima to environmental changes. This has important implications for the vegetation restoration and forest management of the P. massoniana forests in the study area.
2 Materials and methods
2.1 Study area
The study area is located in Dashan Village, Zongyang County, Anhui Province, southeastern China (30°01′-31°38′N, 117°05′-117°43′E), at the southeast edge of Dabie Mountains, which belongs to the north subtropical region. This region has a subtropical monsoon humid climate with typical north subtropical hilly land climate features. The altitude is between 160 m and 200 m. The mean annual temperature is 16.5°C. The average annual sunshine hours and annual precipitation are 2064.9 h and 1326.5 mm, respectively. Between 60% and 70% of the annual rainfall occurs in June, July, August and September. The soils are classified as yellow-brown granite with a lighter texture and the pH value of 4.5–5.5. The terrain is characterized by low hills and gentle slopes, with slopes generally above 20°, some can reach 30° in this region.
The P. massoniana mixed forests in the study area are mainly distributed at the edge of the P. massoniana natural forests, which are artificially planted and uncultivated P. massoniana forests after more than 40 years of mountain closure and afforestation, and developed by the invasion of broad-leaved species. The main tree species are P. strobilacea, Q. acutissima, L. formosana, Rhus chinensis, Cinnamomum camphor, Symplocos paniculata and Clerodendron cyrtophyllum, etc. The forest community structure is obvious, mainly divided into a tree layer, a shrub layer and an herbaceous layer. The canopy density ranges from 0.6 to 0.8.
2.2 Experimental design
According to the influence of topographic and geomorphic conditions on the distribution of P. massoniana forest communities in the study area, four typical and consistent P. massoniana forest types (pure P. massoniana forest (PF), P. massoniana-L. formosana mixed forest (PLM), P. massoniana- Platycarya strobilacea mixed forest (PPM) and P. massoniana-Q. acutissima mixed forest (PQM)) were selected. Sampling sites were arranged on two slope aspects (sunny and shady slopes). There were 3 sample plots (20 m × 20 m) for each forest type, which were randomly set up as three replicates. A total of 24 sample plots (2 slope aspects for each forest type×4 forest types×3 replicates) were set up and leaf and soil samples were collected. Table 1 shows more details.
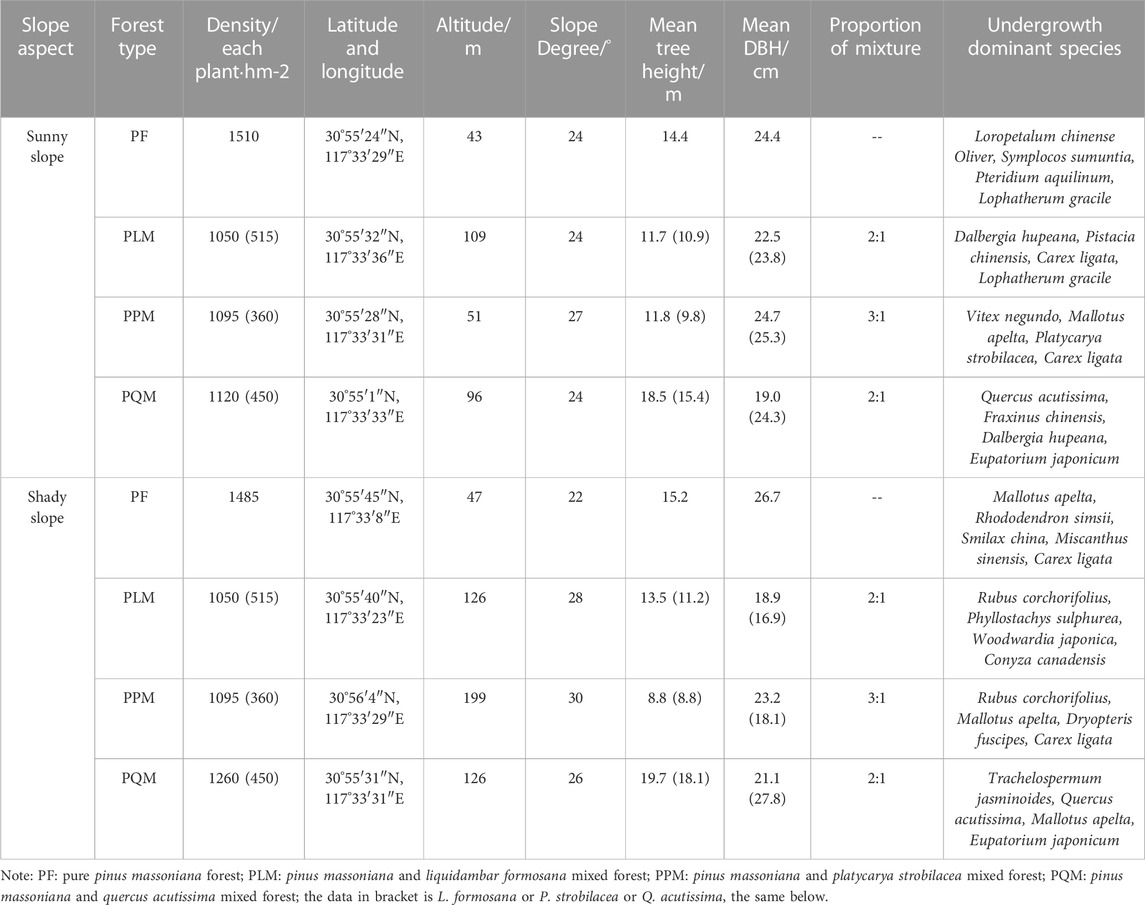
TABLE 1. Basic information of the four P. massoniana forest types in the study area.
2.3 Sampling and processing
Leaf and soil samples were taken from 24 plots in the four forest stands. In each sample plot, three to five typical plants of each tree species (P. massoniana, Quercus tissima, L. formosana, and P. strobilacea) with similar in size, robust in growth, and no pests and diseases were selected to collect leaves. For leaf sampling, high branch shears were used to collect healthy and mature leaves from the sunny side and upper or middle part of the tree crown. About 10–20 g of leaves for each tree species in each plot were collected, mixed into one sample and put into ziplock bags. After return to the laboratory, the mixed leaf samples were heat killed at 105°C for 15 min and dried at 75°C for 48 h to constant weight. The dried leaves were crushed by a high-speed crusher and passed through a 100-mesh sieve for standby. Leaf total carbon content (LCC), leaf total nitrogen content (LNC) and soil total N content (STN) were determined using an elemental analyzer (Vario EL II; Elementar, Germany) (Hu and Wan, 2019). Leaf total phosphorus content (LPC) was determined by molybdenum-antimony colorimetric method after digestion of leaf samples with H2SO4 -H2O2 solution (Kuo, 1996).
Within the range of 1 m in diameter of the sampled plants, the soil surface litter was cleaned up and the soil samples were collected at a depth of 0–20 cm using a soil auger (10 cm in diameter) in each sample plot. Three sampling sites were randomly selected for each plot. Gravel and plant debris were then removed from the soil samples and the three soil samples were mixed to form a pooled sample of approximately 1 kg from each sample plot. The fresh soil samples were then sealed in plastic bags and transported to the laboratory in ice boxes. After returning to the laboratory, the soil samples were air-dried and sieved before soil nutrient content determining. All samples were ground into a fine powder and passed through a 0.15 mm sieve, and then analyzed for soil organic C (SOC), soil total N (STN) and soil total P (STP). SOC was determined by oil bath-K2Cr2O7 titration method (Nelson and Sommers, 1982). STP was determined by the molybdate blue chloride method after digestion with H2SO4–HClO4 (Hu et al., 2023). Each sample was repeated 3 times and the average value was taken as the final result. The ratios of C:N, C:P and N:P were calculated as molar ratios for leaves and soil.
2.4 Statistical analysis
All data were described by mean ± standard deviation (SD). Statistical analyses were performed using Excel 2016 and SPSS 23.0 for Windows. Two-way ANOVA was used to determine the effects of forest type, slope aspect and their interactions on leaf and soil C:N:P stoichiometric characteristics. Significant differences were evaluated at the 0.05 level. Significance analysis was performed using the least significant difference (LSD) post hoc test. Stoichiometric characteristics of C:N:P in leaves and soil were analyzed by Pearson correlation analysis. Redundancy analysis (RDA) was used to test the relationship between leaf and soil stoichiometric characteristics on different slopes. Leaf and soil nutrient contents were expressed as g·kg-1 on a dry weight basis and all C:N:P ratios were calculated as mass ratios. The figures were drawn by Origin Pro 2021 and R 4.0.2 software.
3 Results
3.1 Variations of leaf C, N, P contents and C:N:P stoichiometry
There were significant differences in leaf C, N and P contents and stoichiometric ratios among four P. massoniana forest types (p < 0.05 or p < 0.01) (Table 2). LCC, LNC and LPC varied from 433.19 to 500.59 g·kg-1, 12.90–20.94 g·kg-1, 0.73–1.89 g·kg-1 on the sunny and shady slopes, with mean values of 481.67 g·kg-1, 15.93 g·kg-1 and 1.38 g·kg-1, respectively (Figure 1A–C). Among the four forest types, the distribution of leaf C content of P. massoniana varied from 490–501 g·kg-1, while the leaf C content of the three broad-leaved tree species ranged from 433–487 g·kg-1, and the LCC of P. massoniana was significantly higher than that of the three deciduous tree species (p < 0.05). The distribution of leaf N content of P. massoniana varied from 12.9–16 g·kg-1, while the leaf N content of the three broad-leaved tree species ranged from 13.39–20.94 g·kg-1, and the LNC of P. Strobilacea and Q. acutissima were significantly higher than that of the evergreen species of P. massoniana (p < 0.05). The distribution of leaf P content of P. massoniana varied from 0.89–1.89 g·kg-1, while the leaf P content of the three broad-leaved tree species ranged from 0.73–1.88 g·kg-1, and the LPC of P. Strobilacea, Q. acutissima and P. massoniana in PPM were significantly higher than L. formosana and P. massoniana in PF, PLM and PQM (p < 0.05).
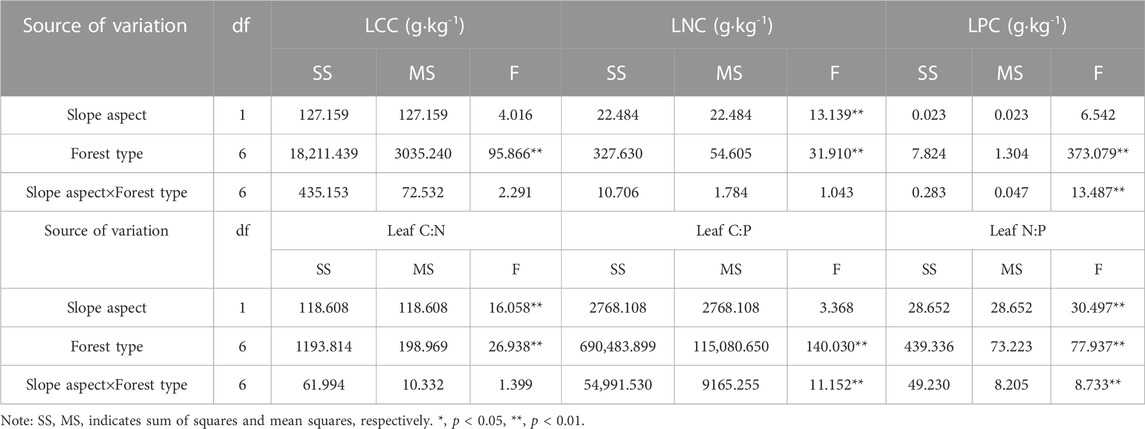
TABLE 2. Two-way ANOVA of slope aspect and forest type on leaf C, N and P contents and stoichiometry in four P. massoniana forest types.
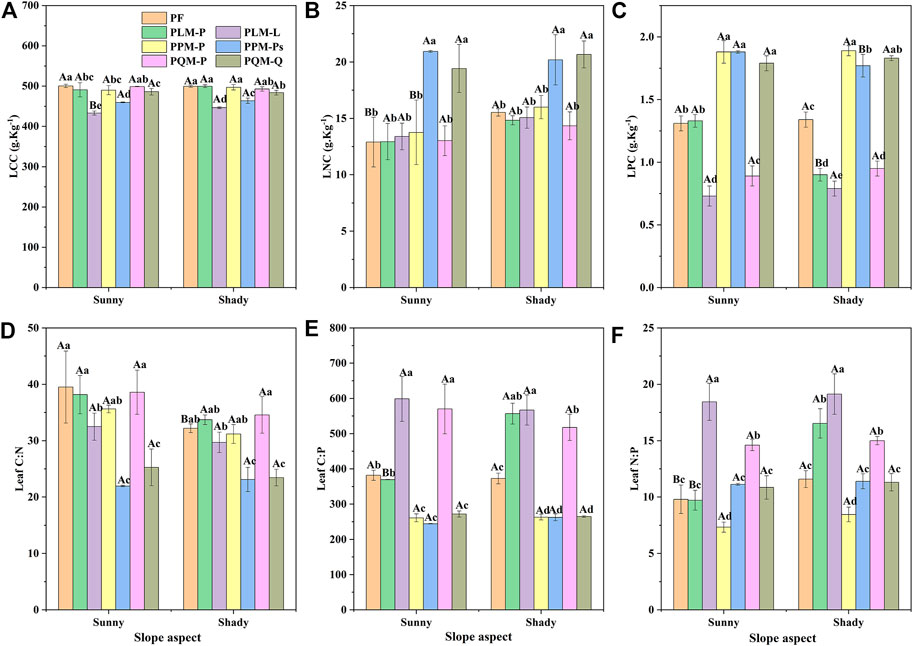
FIGURE 1. Changes in leaf C, N, and P contents and stoichiometry of four Pinus massoniana forest types on two slope aspects. LCC: leaf total carbon content; LNC: leaf total nitrogen content; LPC: leaf total phosphorus content. C:N: leaf C:N ratio; C:P: leaf C:P ratio; N:P: leaf N:P ratio. PF: Pure Pinus massoniana forest; PLM-P: Pinus massoniana in PLM; PLM-L: Liquidambar formosana in PLM; PPM-P: Pinus massoniana in PPM; PPM-Ps: Platycarya strobilacea in PPM; PQM-P: Pinus massoniana in PQM; PQM-Q: Quercus acutissima in PQM. The error bars show the standard deviations of the means. Different lowercase letters indicate significant differences at the 0.05 (p < 0.05) level among four Pinus massoniana forest types within the same slope aspect. Different capital letters indicate significant differences at the 0.05 (p < 0.05) level between two slope aspects for the same forest type, the same below.
Also, the ratios of leaf C:N, C:P and N:P of four P. massoniana forest types varied from 21.96 to 39.52, 244.42 to 598.59, and 7.33 to 19.13 on the sunny and shady slopes, with mean values of 31.40, 392.97 and 12.52, respectively (Figures 1D–F). Among the four forest types, the leaf C:N ratio of P. massoniana was distributed between 31.18–39.52, while the leaf C:N ratios of the three broad-leaved species ranged from 21.96–32.49, and the C:N of P. massoniana was significantly higher than that of the three deciduous tree species (p < 0.05). The distribution of leaf C:P and N:P ratios of P. massoniana varied from 261.1–570.02 and 7.33–16.53, respectively, while the leaf C:N and N:P ratios of the three broad-leaved species ranged from 244.42–598.59 and 10.85–19.13, respectively. The C:N and N:P ratios of L. formosana, P. massoniana in PQM were significantly higher than the other tree species (p < 0.05).
Among the three P. massoniana mixed forests, the leaf N content of the three deciduous species were significantly higher than that of the evergreen species of P. massoniana (p < 0.05) (Figure 2B), while the LCC, C:N and C:P ratios of the three deciduous species were significantly lower than that of the evergreen species of P. massoniana (p < 0.05) (Figure 2A,D,E).
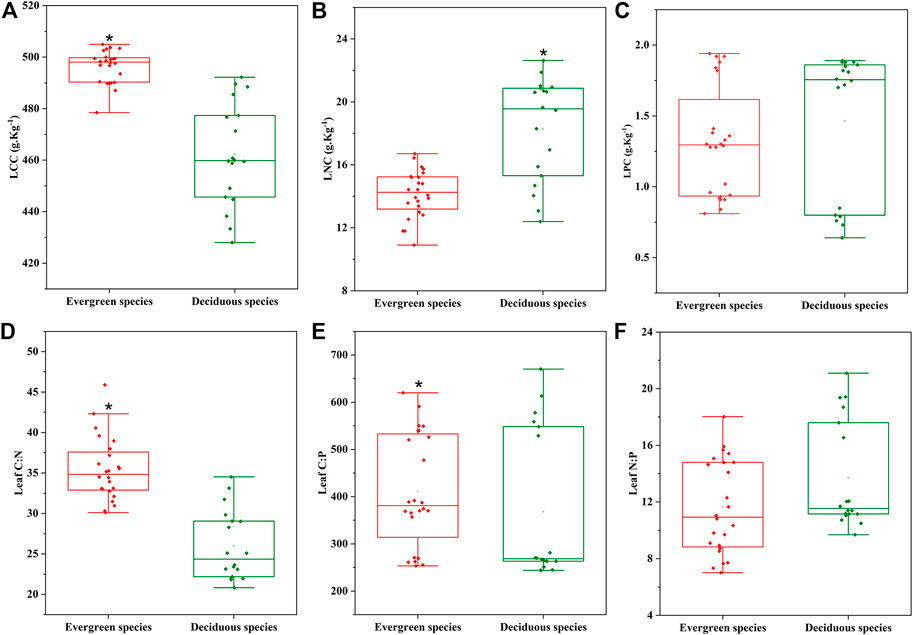
FIGURE 2. Changes in leaf C, N, and P contents and stoichiometry of evergreen and deciduous tree species in four Pinus massoniana forest types on two slope aspects. Evergreen species: Pinus massoniana; Deciduous species: L. formosana, P. strobilacea, and Q. acutissima. * p < 0.05.
The LNC, leaf C:N and N:P ratios of the four P. massoniana forest types were significantly different between the two slopes (p < 0.05 or p < 0.01) (Table 2). The results showed that LNC and N:P ratio were higher on shady slope than on sunny slope, while leaf C:N ratios were higher on the sunny slope than on the shady slope. LCC, LPC and C:P ratio showed slight variations between the two slopes.
Two-way ANOVA showed that forest type had a significant effect on LCC, LNC, LPC, N, P, C:N, C:P and N:P ratios (p < 0.01). Slope aspect also had a significant effect on LNC and C:N, N:P ratios (p < 0.01), but not on LCC, LPC and C:P ratio (p > 0.05). Also, slope aspect×forest had a significant effect on LPC, C:P and N:P ratios (p < 0.01), but not on LCC, LNC and C:N ratio (p > 0.05) (Table 2).
3.2 Variations of soil C, N, P contents and C:N:P stoichiometry
Significant differences in soil C, N, P contents and stoichiometric ratios among four P. massoniana forest types (p < 0.05 or p < 0.01) (Table 3). SOC, STN and STP contents varied from 18.09 to 42.42 g·kg-1, 0.98–2.27 g·kg-1, 0.30–0.46 g·kg-1 on the sunny and shady slopes, with an average of 26.26 g·kg-1, 1.46 g·kg-1 and 0.37 g·kg-1, respectively (Figures 3A–C). Among the four forest types, the SOC, STN and STP contents were significantly different, with a ranking of PQM > PPM > PLM > PF on the sunny slope (p < 0.05). However, the variation of SOC, STN and STP contents were slightly different on the shady slope. PQM had the highest SOC among the four forest types, significantly higher than PLM and PPM (p < 0.05), but not significantly different from PF on shady slope (p > 0.05). STN showed the same change pattern as STP, and PQM had the highest STN and STP contents, which were all significantly higher than the other three forest types on shady slope. (PPM, PLM and PF) (p < 0.05).

TABLE 3. Two-way ANOVA of slope aspect and forest type on soil C, N and P contents and stoichiometry in four P. massoniana forest types.
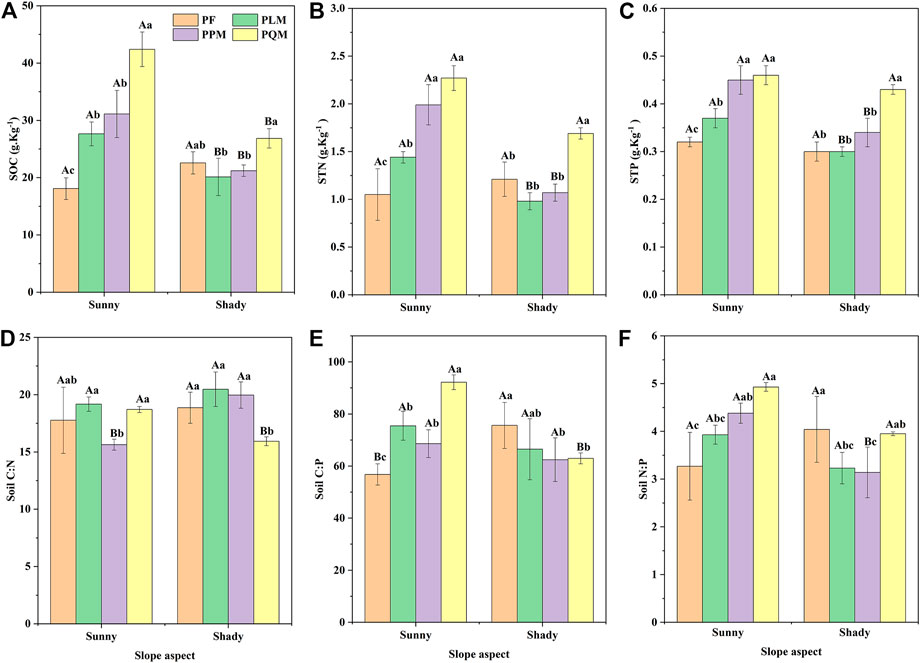
FIGURE 3. Changes in soil C, N, and P contents and stoichiometry of four Pinus massoniana forest types on two slope aspects. SOC, soil organic carbon; STN, soil total nitrogen; STP, soil total phosphorus; C:N, soil C:N ratio; C:P, soil C:P ratio; N:P, soil N:P ratio.
Soil C:N, C:P and N:P ratios of the four forest types ranged from 15.64 to 20.47, 56.79 to 92.20, and 3.14 to 4.93 on the sunny and shady slopes, and mean values of 18.32, 70.07 and 3.86, respectively (Figures 3D–F). Among the four forest types, the range of variation in soil C:N ratio was small, and PPM had the lowest soil C:N ratio on the sunny slope, which was significantly lower than the other two mixed forests (PLM and PQM) (p < 0.05). In contrast, PQM exhibited the lowest soil C:N ratio on shady slope and was significantly lower than the other three forests (PLM, PQM and PF) (p < 0.05). PQM exhibited the highest soil C:P and N:P ratios on the sunny slope, while PF had the highest soil C:P and N:P ratios on the shady slope. The range of variation of soil C:N ratios on the same slope was small, with PPM having the lowest soil C:N ratio on the sunny slope and PQM having the lowest soil C:N ratio on the shady slope.
The SOC, STN, STP and soil C:P, N:P ratios of the four forest types on different slopes were significantly different (p < 0.05 or p < 0.01) (Table 3). The results showed that the contents of SOC, STN and STP and the ratios of C:P and N:P in the three P. massoniana mixed forests were higher on the sunny slope than on the shady slope. However, the SOC, STN, C:P and N:P ratios of pure P. massoniana forest showed the opposite trend.
As seen in Table 3, forest type had a significant effect on soil C:N, C:P and N:P ratios (p < 0.05), and a highly significant effect on SOC, STN and STP contents (p < 0.01). SOC, STN and STP contents and C:P, N:P ratios were also influenced by slope aspects (p < 0.05 or p < 0.01). Slope aspect×forest type had a significant effect on SOC, STN, STP and C:N, C:P and N:P ratios (p < 0.01) (Table 3).
3.3 Correlation between leaf and soil C, N, P contents and C:N:P stoichiometry
RDA analysis showed that soil nutrient characteristics were closely related to leaf nutrient and C:N:P stoichiometry in our study (Figures 4, 5). The first axis explained 88.91% and 60.25% on the sunny and shady slopes, respectively. Among the soil factors, STN and STP had the most significant effect on leaf N, P contents and leaf N:P ratio of the four forest types on the two slopes (p < 0.05). There was a significant synergistic effect between STN and STP, which was mainly positively correlated with LNC, LPC and leaf N:P ratio and negatively correlated with LCC, leaf C:P and leaf C:N ratio. Among them, the comprehensive effect of soil N and P nutrient contents had a significant effect on the leaf stoichiometric characteristics of the four P. massoniana forest types on the sunny slope, while STP had a significant effect on the leaf stoichiometric characteristics on the shady slope.
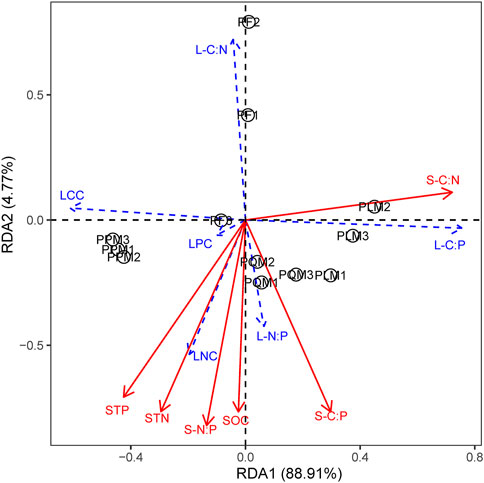
FIGURE 4. RDA sequences of C, N, and P stoichiometric characteristics of leaves and soils in four P. massoniana forest types on sunny slope. SOC, soil organic carbon; STN, soil total nitrogen; STP, soil total phosphorus; S-C:N, soil C:N ratio; S-C:P, soil C:P ratio; S-N:P, soil N:P ratio; LCC, leaf total carbon content; LNC, leaf total nitrogen content; LPC, leaf total phosphorus content; L-C:N, leaf C:N ratio; L-C:P, leaf C:P ratio; L-N:P, leaf N:P ratio.
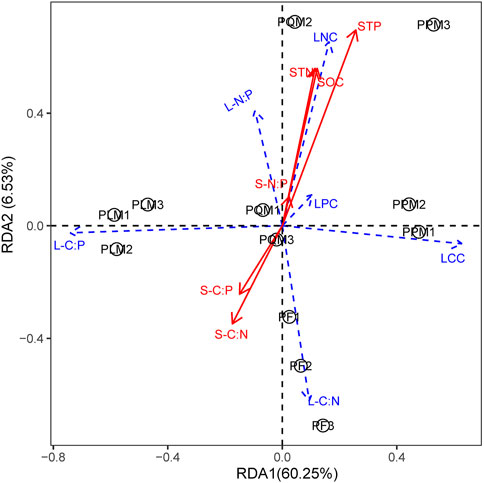
FIGURE 5. RDA sequences of C, N, and P stoichiometric characteristics of leaves and soils in four P. massoniana forest types on shady slope.
Pearson correlation analysis was conducted in four forest types on two slopes to evaluate the correlation between leaf and soil stoichiometric characteristics (Table 4). The results revealed that there was a significant positive correlation between LNC and LPC (p < 0.01), while LCC did not have a significant correlation with LNC and LPC (p > 0.05). LNC and LPC were significantly negatively correlated with leaf C:P and C:N ratios, respectively (p < 0.05 or p < 0.01). Leaf C:P ratio was significantly positively correlated with leaf C:N and N:P ratios (p < 0.05 or p < 0.01). SOC was significantly positively correlated with STN and STP (p < 0.01). Soil N:P ratio was significantly positively correlated with soil C:P ratio and negatively correlated with soil C:N ratio (p < 0.01).
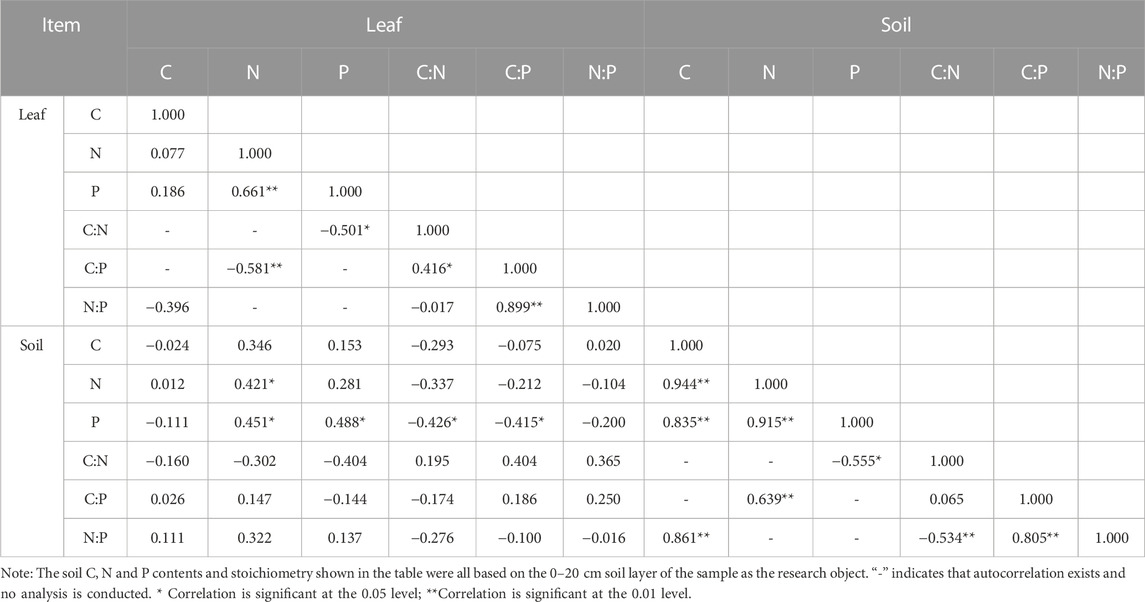
TABLE 4. Correlation coefficients of leaf and soil C, N and P contents and stoichiometry in four P. massoniana forest types.
From the correlation analysis between leaf and soil, the results showed that LCC was significantly positively correlated with STN and STP (p < 0.05). In addition, there was a significant positive correlation between LPC and STP (p < 0.05). STP content was negatively correlated with leaf C:N and C:P ratios (p < 0.05). The results indicated that the content of leaf N and P increased significantly with the increase of N and P nutrients in the soil.
4 Discussion
4.1 Influence of forest type and slope aspect on leaf nutrient contents and C:N:P stoichiometry
In this study, forest type was the main factor affecting leaf C, N, P contents and C:N:P stoichiometry. The average LCC of the four P. massoniana forest types was 481.67 g·kg-1, higher than the global average (461.6 g·kg-1) (Elser et al., 2000a) and other study areas in China (the forest ecosystem in eastern China: 480.1 g·kg-1 (Ren et al., 2012), in the Loess Plateau: 438.3 g·kg-1 (Zheng and Shangguan, 2007)), which indicates that the higher leaf organic compound content of species and stronger carbon storage capacity in the subtropical region of China. Compared with the P. massoniana forests in other regions of China, the four P. massoniana forest types were significantly higher than the average level (443.3 g·kg-1) of P. massoniana plantation in Central Guizhou region (Pan et al., 2020), and slightly lower than the P. massoniana natural forests in southern Jiangxi region (485.3–502.6 g·kg-1) (Wang and Yu, 2008), and close to the average level (478.12 g·kg-1) in the Wuling mountains area of Chongqing (Wang et al., 2022). This indicates that the leaf C content of P. massoniana in the study area was at a medium level.
Among the four forest types, PF exhibited the highest leaf C contents both on the sunny slope (500.59 g·kg-1) and shady slope (499.84 g·kg-1), and was significantly higher than the other three mixed forests (PLM, PPM and PQM). The mean values were ranked as PF > PQM > PPM > PLM. At the tree species level, the evergreen species of P. massoniana exhibited the highest leaf C contents on the two slopes, and was significantly higher than the three deciduous tree species (L. formosana, P. strobilacea and Q. acutissima) in the mixed forests. Since evergreen leaves live much longer than deciduous leaves and have higher carbon gain per unit of nitrogen than deciduous leaves throughout their life cycle (Thomas and Martin, 2012). It was found that increased C content in leaves of evergreen tree species was consistent with leaves that are poorly adapted to soil nutrient habitats, indicating increased carbon input in constitutive compounds such as lignin and phenolics (Wright et al., 2005; Bertiller et al., 2006). In addition, evergreen lanceolate leaves usually contain carbon-rich secondary products such as tannins and resins, which also relatively increased the leaf C content of P. massoniana (Song et al., 2021). Thus, this indicates that the species of P. massoniana had a strong C storage capacity and defense capability against poor soil nutrient environment in the study area.
As the most vulnerable elements to shortages in plants, N and P are the critical limiting factors in terrestrial ecosystems (Sterner and Elser, 2002). The average LNC and LPC contents in global and China were 20.1 and 1.77 g·kg-1, 20.2 and 1.46 g·kg-1, respectively (Reich and Oleksyn, 2004; Han et al., 2005). In our study, the average LNC and LPC of the four P. massoniana forest types were 15.93 g·kg-1 and 1.38 g·kg-1, respectively. They were all significantly lower than the global and Chinese averages. The lower leaf N content in north subtropical forest ecosystems may be related to the higher rainfall in this region, as plants absorb soluble nitrate and ammonia N. However, the relatively high rainfall in southeastern China may have lead to N leaching, reducing the amount of N available for plant uptake and use, resulting in lower N content in leaves in the study area. Han et al. (2005) found that compared with the global vegetation data of Elser et al. (2000a) and Reich and Oleksyn (2004), the leaf P content of Chinese plants was relatively low. This is because most of the P in plants is absorbed by the roots from the soil, while the contents of P in Chinese soil are relatively low.
The N and P contents of plant leaves vary greatly, although they both perform key functions of photosynthetic carbon, such as assimilation and transpiration (Chen et al., 2013). This is due to the differences in climate, soils, vegetation types and developing histories in different locations (Ordoñez et al., 2009; Bui and Henderson, 2013). In our study, leaf N and P contents varied significantly with forest type, and this forest type-related pattern differed in P. massoniana and three deciduous tree species (Figure 1B). Compared to the four species, two deciduous species (P. strobilacea and Q. acutissima) had higher leaf N and P contents than the evergreen species of P. massoniana (Figure 2B), especially the LNC and LPC of Q. acutissima (Figure 2). Sterner and Elser (2002) found that plants with short growth cycles and high growth rates usually had high N content, high P requirements and strong photosynthetic capacity, while plants with slow growth rates and long life spans had low N and P contents (Tian et al., 2019).
Differences in C:N:P among different forest types reflect plant responses and adaptation strategies to different environmental conditions (Zhang J. H. et al., 2021). Leaf C:N and C:P ratios can reflect carbon assimilation ability and nutrient use efficiency, which has important ecological significance (Elser et al., 2003). In general, higher leaf C:N and C:P ratios means slower plant growth rate but stronger C assimilation ability and more efficient N and P utilization (He et al., 2008). In our study, the mean C:N and C:P ratios of the four forest types were 31.40 and 392.97, respectively. The evergreen species of P. massoniana were greater than the three broad-leaved deciduous species, while the leaf C:N and C:P ratios were significantly higher than global levels (22.50 and 232, respectively) (Elser et al., 2000a), indicating a high mineral nutrient use efficiency and dry matter synthesis capacity of leaves in the study area. In addition, higher C:P ratios in subtropical forests also indicate lower P content. Previous studies have shown that high values for leaf C content and C:N ratio are strongly associated with high tolerance to physical stress, which may increase leaf longevity (Chabot and Hicks, 1982). In our study, evergreen species of P. massoniana had higher C:N and C:P ratios both in pure forest and in mixed forests. In general, higher C:N ratios seem to be consistent with a “Demand reduction strategy (use of specific morphological or histological structures to reduce plant dependence on nutrients)” (Sardans et al., 2015), as coniferous species of P. massoniana adapt to barren conditions through spikes and tapering foliage (Zhang J. H. et al., 2021). Such leaf morphology leads to reduced photosynthesis and N requirements (Fleischer et al., 2013). In addition, the lower C:N of the three broad-leaf deciduous species in the Dabie Mountains region suggests that plants tend to store N in these nutrient-poor habitats. The availability of P was determined by the decomposition rate of soil organic matter, and the lower C:P ratio, the higher P availability (Han et al., 2021). The leaf C:P ratios of two deciduous species (P. strobilacea and Q. acutissima) were lower than those of the evergreen species (P. massoniana), indicating that soil organic matter decomposition rate and soil P availability were higher in the two deciduous species than in the evergreen species. Leaf N:P ratio is an important indicator reflecting the nutrient restriction of plant growth (Koerselman and Meuleman, 1996; Güsewell, 2004). In general, N:P < 14 indicates N limitation; N:P > 16 indicates P limitation; and between 14 and 16 indicates co-limitation (Reich and Oleksyn, 2004). However, the variation of N:P in leaves varies among species and is influenced by the nutrient conditions to which the plant is exposed (Güsewell, 2004). In this study, the average leaf N:P ratio of the four forest types was 12.52, which was much lower than the average level in China (16.3) (Han et al., 2005), and slightly lower than the global average (12.7) (Elser et al., 2000a). Thus, the lower N and P contents and N:P ratios of the four P. massoniana forest types suggest that plant growth is more susceptible to N-element limitation under the combined effects of N and P in the Dabie Mountains region of north subtropical.
At the tree species level, L. formosana had the highest leaf N:P ratios (18.44 on sunny slope and 19.13 on shady slope), which were significantly higher than the other three species, with values ranked as L. formosana > P. strobilacea > Q. acutissima > P. massoniana (in PF). This suggests that the growth of L. formosana is more likely to be limited by P than the other two broad-leaved species (N:P > 16). Among the three mixed forests, leaf N:P ratios of two deciduous species (L. formosana and P. strobilacea) were significantly higher in PLM and PPM than in P. massoniana. On the contrary, leaf N:P ratio of Q. acutissima was significantly lower than that of P. massoniana in PQM. This trend was observed on both slopes. This is due to the higher leaf P content of Q. acutissima in the PQM. We also found that the leaf N:P ratio of the same tree species was higher on shady slope than on sunny slope. Many studies have found significant differences in some leaf traits among different life forms (Poorter and Bongers, 2006; Sterck et al., 2006). And it can also be seen from our study that And it is also evident from our study that even on a smaller scale, vegetation distributed in different habitats in the same area can lead to significant differences in leaf N because of their different life forms.
As an important topographic factor in small-scale spaces, slope aspect affects plant growth and physiology by regulating the combination of water, heat and light conditions. Two-way ANOVA showed that LNC and leaf C:N and N:P ratios of the four forest types differed significantly between the two slopes, indicating a significant effect of slope aspect on leaf nutrient content and stoichiometric characteristics. In our study, leaf N content and N:P ratio were higher on shady slope than on sunny slope, while leaf C:N ratio was higher on sunny slope than on shady slope. This suggests that plants have different nutrient utilization strategies and defense mechanisms when adapting to the changes in the micro-topography (different slope aspects) of the study area.
4.2 Influence of forest type and slope aspect on soil nutrient contents and C:N:P stoichiometry
The balance of soil C, N, P and other nutrient factors is related to the level of forest soil fertility, and its content and distribution affect the growth and development of forest vegetation (Wang et al., 2021). In this study, the mean soil C, N and P of the four P. massoniana forest types were 26.26 g·kg-1, 1.46 g·kg-1, and 0.37 g·kg-1, respectively. Compared to the Chinese average values of soil C (24.6 g·kg-1), N (1.88 g·kg-1) and P (0.78 g·kg-1) (Tian et al., 2010), the study area had higher C content and significantly lower N and P contents than the average. This suggests that the soil C content is relatively abundant, while soil N and P contents of four P. massoniana forest types are relatively poor in the Dabie Mountains region of north subtropical. The higher soil C content in the study area may be associated with higher precipitation, as more precipitation may stimulate plant growth and soil C input (Hui et al., 2021).
Different forest types have different feedback effects on soil C, N and P contents (Bo et al., 2020; Hui et al., 2021). There were significant differences in soil C, N and P contents among the four forest types in the study area. The results showed that the C and N contents of the three mixed forests were significantly higher than that of pure forest on sunny slope, especially PQM, and in the order of PQM > PPM > PLM > PF, which is consistent with the findings of Hui et al. (2021), since soil C and N contents increased linearly with increasing vegetation cover and litter amount. Soil C and N are important components of soil organic matter and always exhibit a coupled relationship. Hyvonen et al. (2008) found in a long-term localization experiment in 15 forests that carbon input could improve soil N accumulation to some extent, while increasing N input could reduce soil carbon mineralization. STP content differed significantly among the four forest types, but varied slightly on the two slopes. The soil P content of the three mixed forests remained higher than that of the pure P. massoniana forest, and with the highest in PQM. Due to the differences in the dominant tree species in the four P. massoniana forest types, the quality of forest litter differs, and the decomposition amount and rate of litter were also different, which leads to significant changes in soil nutrient content that are closely related to the characteristics of tree species (Vesterdal et al., 2008; Zhang J. H. et al., 2021). In terrestrial ecosystems, litter plays an important role in the nutrient cycling between plants and soils. The main sources of forest soil nutrients are litter decomposition, root exudates and biological disturbances (Rumpel and Kogel-Knabner, 2010; Chapin et al., 2011), so soil nutrients (mainly C, N and P) are closely related to cover vegetation type, growth status and litter mineralization (Lorenz and Lal, 2007; Batjes, 2014). Since evergreen trees live much longer than deciduous trees, the litter quantity of deciduous broadleaved trees is higher than that of evergreen trees (P. massoniana) in the same year, which improves the quantity and quality of exogenous inputs (litter and root deposition) and ultimately leads to an increase of soil nutrient sources in mixed forests containing deciduous trees (Cao and Chen, 2017). In addition, mixed forests usually have higher species richness and inputs of plant residues than pure forests, and high species diversity can increase the amount and type of litter, resulting in higher accumulation and nutrient content of soil organic carbon pools than in pure forest (Laganière et al., 2010). Some studies have also found that broad-leaved forest litter is more easily available to microorganisms than coniferous forest (Vesterdal et al., 2008) because decomposition of coniferous forest litter may lead to soil acidification and inhibit the growth and reproduction of soil microorganisms, whereas broad-leaved forest litter is more likely to form stable organic matter, which promotes the input of C, N and P.
Slope aspect is also an important factor affecting soil nutrient content, and plays a key role in determining temperature and moisture due to differences in sunlight hours, leading to a series of microclimates (Griffiths et al., 2009). In our study, the SOC, STN and STP contents of the three mixed forests were significantly higher on the sunny slope than on the shady slope compared to the two slopes, which is inconsistent with previous studies (Lenka et al., 2013; Huang et al., 2015). The analysis showed that this could be influenced by the slope position of the sampling sites with different slopes. The slope position of the three mixed forest plots on the shady slope was higher than the slope position on the sunny slope. Higher slopes are susceptible to strong erosion by rainfall, resulting in soil being washed to lower slopes, thus reducing the accumulation of soil material on shady slope. In addition, due to the close coupling between soil C and N, SOC also affects STN content, which increases when the available carbon source is relatively abundant on the sunny slope (Gao, et al., 2020).
Soil C:N, C:P, N:P ratios are important indicators of soil nutrient balance (Bing et al., 2016), and it is important to clarify their different responses to different forest types to reveal ecosystem function (Wu et al., 2021). Soil C:N ratio reflects the mineralization rate of soil organic matter, the higher the value, the slower the mineralization and decomposition rate of soil organic matter (Yang et al., 2019). It was found that soil C:N and C:P ratios increased when soil N and P contents were insufficient or low (Güsewell et al., 2005). In this study, soil C:N ratios of the four P. massoniana forest types ranged from 15.64–20.47 from sunny to shady slopes, with a mean C:N ratio of 18.32, significantly higher than the Chinese average of 11.9 (Tian et al., 2010). Higher soil C:N ratios indicate weaker mineralization of soil organic matter, which is related to the higher C content and lower N content of the four forest types in the study area. Soil C:N ratios had little variation among the four forest types and the two slopes, ranging from 15.64–20.47. It is believed that C and N form a relatively balanced relationship in the process of consumption and accumulation. It was found that the C:N ratio stabilized after plant residues entered the soil in the form of litter, and the C-N coupling relationship remained close (McGroddy et al., 2004). On the other hand, when litter decomposes and releases nutrients, the carbon skeleton of organic matter needs to be destroyed, and C and N are released simultaneously in this process (Olander and Vitousek, 2000). Among the four forest types, PPM (15.64) and PQM (15.93) had the lowest soil C:N ratios on sunny and shady slopes, respectively, indicating that PPM and PQM obtained more effective C during organic matter mineralization, thus accelerating the rate of mineralization (Huang et al., 2015). This may be due to the fact that P. strobilacea and Q. acutissima in PPM and PQM mixed forests can provide sufficient N sources for soil microorganisms.
Soil C:P ratio reflects the potential of soil microorganisms to release P from mineralized organic matter and measures the absorption and fixation capacity of soil P (Tian et al., 2010). In this study, the soil C:P ratio was 70.07, which was much higher than the average of 61 in China (Tian et al., 2010), indicating that the potential for P release from organic matter mineralization was significantly lower than the average. Soil C:P showed different patterns on the two slopes, with a higher mean C:P ratio on the sunny slope (73.26) than on the shady slope (66.87), and concluded that this was related to the higher soil C content on the sunny slope of the study area. In addition, soil C:P ratios of the three mixed forests (PLM, PPM and PQM) were significantly higher on sunny slope but lower on shady slope compared to pure forest. This indicates that mixed forests can alleviate the P limitation on plant growth to some extent, which is associated with more favorable environmental conditions on shady slope. In addition, because mixed forests can provide more litter, they promote soil microorganisms activity and litter decomposition capacity, thus increasing the accumulation of soil P content in mixed forests (Hu et al., 2020).
Soil N:P ratio reflects soil nutrient availability, reveals nutrient movement between plants and soil, and is a useful predictor of nutrient limitation (Fan et al., 2015; De Feudis et al., 2017). Studies have shown that vegetation growth is limited by N when the soil N:P ratio is less than 10 (Bui and Henderson, 2013). In our study, the soil N:P ratios of the four P. massoniana forest types ranged from 3.14–4.93 from sunny to shady slopes, with an average C:N ratio of 3.86, which was significantly lower than the average of 5.2 in China (Tian et al., 2010), indicating that plant growth in the four P. massoniana forest types may be limited by soil N. Among the four forest types, the soil N:P ratios of two mixed forests (PPM and PQM) were significantly higher than the PF on sunny slope, indicating that the utilization of soil N was higher in the mixed coniferous and broad-leaved forests and that the vigorous growth of above-ground vegetation in the mixed forests promoted the conversion and accumulation of soil N.
4.3 Relationships of C, N and P nutrient contents and stoichiometry between leaf and soil in the four P. massoniana forest types
Plant-soil feedback have received increasing attention as drivers of plant community composition and species coexistence (Kardol et al., 2013). Previous studies have shown that changes in the C, N and P content of plants also affect the C, N and P nutrient contents and stoichiometry of soils, as plants provide a rich substrate for soils through litter and root deposition (Peichl et al., 2012; Zhao et al., 2015). Our results showed a significant positive correlation between leaf N and P content (r = 0.661, p < 0.01) and negative correlation with the leaf C:P ratio (r = −0.581, p < 0.01). The C:P ratio was significantly influenced by leaf N and P contents due to the high positive correlation between leaf N and P contents. Except for differences between broad-leaved and coniferous species, the N:P ratio were less variable than N and P alone among different P. massoniana forest types. Such relatively stable N:P ratios of forest types may reflect the basic characteristics of leaf N and P stoichiometry (Qin and Shangguan, 2019).
Previous studies have shown that if the amount of an element in a plant organ is proportional to the amount of an element in the soil, then plant growth will be limited by that element (Güsewell, 2004). For all forest types pooled together, we found that soil nutrients would be strongly related to leaf nutrients in our study area. Leaf N content was significantly positively correlated with soil N content (r = 0.421) and soil P content (r = 0.451), and leaf P content was significantly positively correlated with soil P content (r = 0.488). As a direct source of plant nutrients, soil N and P contents affect plant C:N:P stoichiometry (Zhang et al., 2018). Our results showed that the low N and P content in the soil directly contributed to the low N and P content in the plant leaves of the four P. massoniana forest types. In addition, leaf C:N and C:P ratios of the four forest types showed a significant negative correlation with P content in the soil, i.e., the lower the P content in the soil, the lower the P content in the plant leaves. Furthermore, insufficient P supply due to insufficient PO4 concentration also affected the N fixation and C synthesis processes in plants, leading to an increase in the N:P ratio in the soil (Augusto et al., 2013). Therefore, the N:P ratio can better reflect the intrinsic relationship between plant and soil nutrients. In our study, the relationship between leaves and soil nutrients was tightly coupled. Correlation analysis revealed that slope-induced changes in soil nutrient contents of the four P. massoniana forest types directly affected the plants themselves. And RDA analysis confirmed that soil N and P contents were key factors affecting C:N:P stoichiometriy in plant leaves of the four P. massoniana forest types, especially soil P content. However, the relationship between soil C:N, C:P and N:P ratios and leaf C:N, C:P and N:P ratios was weak. Previous studies reported that the correspondence between plant stoichiometry and soil stoichiometry mainly showed an unstable relationship. In natural ecosystems, C:N:P ratios are generally regulated by environmental conditions (Chapin et al., 2011). Elevation, vegetation type, temperature, precipitation, and soil nutrients all affect the C:N:P ratios of species, leading to controversial spatial patterns (Chen et al., 2018; Hu et al., 2022). Also, slope aspect is an important environmental factor affecting plant habitat, which changes the spatial redistribution of solar radiation and precipitation. Therefore, the C:N:P ratio of P. massoniana forests at different community levels is a comprehensive parameter reflecting the relative contribution of each species to the forest community, and its spatial pattern and influencing factors may be more complex.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
YL, ZC, and BZ: methodology. JQ: software, validation, data curation, writing—original draft and writing—review and editing and project administration. YL, QB, ZC, and BZ: investigation. All authors contributed to the manuscript and approved the submitted version.
Funding
This research was supported by the Project of National Natural Science Foundation of China (No. 31901160), and the Natural Science Research Key Project of Universities in Anhui Province (No. KJ 2019A0206).
Acknowledgments
The authors express their sincere thanks to the reviewers and issue editors of the journal for their valuable comments, suggestions, and revisions to this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ai, Z., He, L., Xin, Q., Yang, T., Li, G., and Xue, S. (2017). Slope aspect affects the non-structural carbohydrates and C:N:P stoichiometry of Artemisia sacrorum on the Loess Plateau in China. Catena 152, 9–17. doi:10.1016/j.catena.2016.12.024
Augusto, L., Delerue, F., Gallet-Budynek, A., and Achat, D. L. (2013). Global assessment of limitation to symbiotic nitrogen fixation by phosphorus availability in terrestrial ecosystems using a meta-analysis approach. Glob. Biogeochem. Cycles 27 (3), 804–815. doi:10.1002/gbc.20069
Batjes, N. H. (2014). Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 65 (1), 151–163. doi:10.1111/j.1365-2389.1996.tb01386.x
Bertiller, M. B., Mazzarino, M. J., Carrera, A. L., Diehl, P., Satti, P., Gobbi, M., et al. (2006). Leaf strategies and soil N across a regional humidity gradient in Patagonia. Oecologia 148, 612–624. doi:10.1007/s00442-006-0401-8
Bing, H. J., Wu, Y. H., Zhou, J., Sun, H. Y., Luo, J., Wang, J. P., et al. (2016). Stoichiometric variation of carbon, nitrogen, and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau. J. Soils Sediments 16, 405–416. doi:10.1007/s11368-015-1200-9
Bo, F. J., Zhang, Y. X., Han, Y., Chen, H., Wang, P. G., Ren, X. M., et al. (2020). The C:N:P stoichiometry of planted and natural Larix principis-rupprechtii stands along altitudinal gradients on the Loess Plateau, China. Forests 11, 363. doi:10.3390/f11040363
Bui, E. N., and Henderson, B. L. (2013). C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 373 (1/2), 553–568. doi:10.1007/s11104-013-1823-9
Cao, Y., and Chen, Y. M. (2017). Coupling of plant and soil C:N:P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau, China. Trees 31, 1559–1570. doi:10.1007/s00468-017-1569-8
Chabot, B. F., and Hicks, D. J. (1982). The ecology of leaf life spans. Annu. Rev. Ecol. Syst. 13, 229–259. doi:10.1146/annurev.es.13.110182.001305
Chapin, F. S., Matson, P. A., and Mooney, H. A. (2011). Principles of terrestrial ecosystem ecology. New York: Springer Science and Business Media.
Chen, Y. H., Han, W. X., Tang, L. Y., Tang, Z. Y., and Fang, J. Y. (2013). Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography 36, 178–184. doi:10.1111/j.1600-0587.2011.06833.x
Chen, L. L., Deng, Q., Yuan, Z. Y., Mu, X. M., and Kallenbach, R. L. (2018). Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau of China. Ecol. Eng. 120, 14–22. doi:10.1016/j.ecoleng.2018.05.021
Cleveland, C. C., and Liptzin, D. (2007). C:N:P stoichiometry in soil: Is there a “redfield ratio” for the microbial biomass? Biogeochemistry 85, 235–252. doi:10.1007/s10533-007-9132-0
De Feudis, M., Cardelli, V., Massaccesi, L., Lagomarsino, A., Fornasier, F., Westphalen, D. J., et al. (2017). Influence of altitude on biochemical properties of European beech (Fagus sylvatica L) forest soils. Forests 8, 213. doi:10.3390/f8060213
Elser, J. J., Fagan, W. F., Denno, R. F., Dobberfuhl, D. R., Folarin, A., Huberty, A., et al. (2000a). Nutritional constraints in terrestrial and freshwater food webs. Nature 408, 578–580. doi:10.1038/35046058
Elser, J. J., Sterner, R. W., Gorokhova, E., Fagan, W. F., Markow, T. A., Cotner, J. B., et al. (2000b). Biological stoichiometry from genes to ecosystems. Ecol. Lett. 3 (6), 540–550. doi:10.1046/j.1461-0248.2000.00185.x
Elser, J., Acharya, K., Kyle, M., Cotner, J., Makino, W., Markow, T., et al. (2003). Growth rate stoichiometry couplings in diverse biota. Ecol. Lett. 6 (10), 936–943. doi:10.1046/j.1461-0248.2003.00518.x
Fan, H. B., Wu, J. P., Liu, W. F., Yuan, Y. H., Hu, L., and Cai, Q. K. (2015). Linkages of plant and soil C: N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 392, 127–138. doi:10.1007/s11104-015-2444-2
Fleischer, K., Rebel, K. T., van der Molen, M. K., Erisman, J. W., Wassen, M. J., van Loon, E. E., et al. (2013). The contribution of nitrogen deposition to the photosynthetic capacity of forests. Glob. Biogeochem. Cycles 27, 187–199. doi:10.1002/gbc.20026
Gao, T., Li, J. R., Lu, J., Zheng, W. L., Chen, J. R., Wang, J. K., et al. (2020). Soil nutrient and fertility of different slope directions in the Abie georgei var. Smithii forest in Sejila Mountain. Acta Ecol. Sin. 40 (4), 1331–1341.
Griffiths, R. P., Madritch, M. D., and Swanson, A. K. (2009). The effects of topography on forest soil characteristics in the Oregon Cascade mountains (USA): Implications for the effects of climate change on soil properties. For. Ecol. Manag. 257 (1), 1–7. doi:10.1016/j.foreco.2008.08.010
Guan, F. Y., Tang, X. L., Fan, S. H., Zhao, J. C., and Peng, C. (2015). Changes in soil carbon and nitrogen stocks followed the conversion from secondary forest to Chinese fir and Moso bamboo plantations. Catena 133, 455–460. doi:10.1016/j.catena.2015.03.002
Güsewell, S., Jewell, P. L., and Edwards, P. J. (2005). Effects of heterogeneous habitat use by cattle on nutrient availability and litter decomposition in soils of an Alpine pasture. Plant Soil 268, 135–149. doi:10.1007/s11104-004-0304-6
Güsewell, S. (2004). N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 164, 243–266. doi:10.1111/j.1469-8137.2004.01192.x
Halim, A., and Normaniza, O. (2015). The effect of plant density of Melastoma malabathricum on the erosion rate of slope soil at different slope orientations. Int. J. Sediment. Res. 30 (2), 131–141. doi:10.1016/j.ijsrc.2015.03.003
Han, W. X., Fang, J. Y., Guo, D. L., and Zhang, Y. (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 168, 377–385. doi:10.1111/j.1469-8137.2005.01530.x
Han, C., Liu, Y. J., Zhang, C. K., Li, Y., Zhou, T. R., Khan, S., et al. (2021). Effects of three coniferous plantation species on plant-soil feedbacks and soil physical and chemical properties in semi-arid mountain ecosystems. For. Ecosyst. 8 (3), 3–13. doi:10.1186/s40663-021-00281-4
He, J. S., Wang, L., Flynn, D. F. B., Wang, X., Ma, W., and Fang, J. (2008). Leaf nitrogen: Phosphorus stoichiometry across Chinese grassland biomes. Oecologia 155, 301–310. doi:10.1007/s00442-007-0912-y
He, X. J., Hou, E. Q., Liu, Y., and Wen, D. Z. (2016). Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China. Sci. Rep. 6, 24261. doi:10.1038/srep24261
Heuck, C., and Spohn, M. (2016). Carbon, nitrogen and phosphorus net mineralization in organic horizons of temperate forests:stoichiometry and relations to organic matter quality. Biogeochemistry 131 (1), 229–242. doi:10.1007/s10533-016-0276-7
Hu, M. J., and Wan, S. Q. (2019). Effects of fire and nitrogen addition on photosynthesis and growth of three dominant understory plant species in a temperate forest. J. Plant Ecol. 12 (4), 759–768. doi:10.1093/jpe/rtz014
Hu, M., Song, J., Li, S., Li, Z., Hao, Y., Di, M. M., et al. (2020). Understanding the effects of fire and nitrogen addition on soil respiration of a field study by combining observations with a meta-analysis. Agr. For. Meteorol. 292-293, 108106. doi:10.1016/j.agrformet.2020.108106
Hu, Q., Sheng, M., Bai, Y., Yin, J., and Xiao, H. (2022). Response of C, N, and P stoichiometry characteristics of Broussonetia papyrifera to altitude gradients and soil nutrients in the karst rocky ecosystem, SW China. Plant Soil 475, 123–136. doi:10.1007/s11104-020-04742-7
Hu, M. J., Wang, J. L., Lu, L. L., Shao, P. S., Zhou, Z. X., Wang, D., et al. (2023). Post-fire soil extracellular enzyme activities in subtropical–warm temperate climate transitional forests. Land Degrad. Dev. 34 1–11. doi:10.1002/ldr.4582
Huang, Y. M., Liu, D., and An, S. S. (2015). Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. Catena 125, 135–145. doi:10.1016/j.catena.2014.09.010
Hui, D. F., Yang, X. T., Deng, Q., Liu, Q., Wang, X., Yang, H., et al. (2021). Soil C:N:P stoichiometry in tropical forests on hainan island of China: Spatial and vertical variations. Catena 201 (7), 105228. doi:10.1016/j.catena.2021.105228
Hyvonen, R., Persson, T., Andersson, S., Olsson, B., Agren, G. I., and Linder, S. (2008). Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe. Biogeochemistry 89, 121–137. doi:10.1007/s10533-007-9121-3
Kardol, P., de Deyn, G. B., Laliberté, E., Mariotte, P., and Hawkes, C. V. (2013). Biotic plant-soil feedbacks across temporal scales. J. Ecol. 101, 309–315. doi:10.1111/1365-2745.12046
Koerselman, W., and Meuleman, A. F. M. (1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 33 (6), 1441–1450. doi:10.2307/2404783
Kuo, S. (1996). “Phosphorus,” in Method of soil analysis. Part 3. Chemical methods. Editors D. L. Sparks, A. I. Page, P. A. Helmke, R. H. Leoppert, and P. N. Soltanpour (Madison: Soil Science Society America Book Series), 869–919.
Laganière, J., Angers, D. A., and Paré, D. (2010). Carbon accumulation in agricultural soils after afforestation: A meta-analysis: SOC accumulation following afforestation. Glob. Change Biol. 16 (1), 439–453. doi:10.1111/j.1365-2486.2009.01930.x
Lenka, N. K., Sudhishri, S., Dass, A., Choudhury, P. R., Lenka, S., and Patnaik, U. S. (2013). Soil carbon sequestration as affected by slope aspect under restoration treatments of a degraded alfisol in the Indian sub-tropics. Geoderma 204–205, 102–110. doi:10.1016/j.geoderma.2013.04.009
Lorenz, K., and Lal, R. (2007). Stabilization of organic carbon in chemically separated pools in reclaimed coal mine soils in Ohio. Geoderma 141, 294–301. doi:10.1016/j.geoderma.2007.06.008
McGroddy, M. E., Daufresne, T., and Hedin, L. O. (2004). Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial red field type ratios. Ecology 85 (9), 2390–2401. doi:10.1890/03-0351
Nelson, D. W., and Sommers, L. E. (1982). “Total carbon, organic carbon and organic matter,” in Methods of soil analysis. Part 2. Editors A. L. Page, R. H. Miller, and D. R. Keeney 2nd ed. (Madison: American Society of Agronomy), 539–580.
Niu, Y. L., Li, K. M., Wang, X. Y., Wei, C., Wang, W. X., Su, H. H., et al. (2020). Responses of ecological stoichiometric characteristics and functional traits of Heteropappus hispidus to slope aspect. Chin. J. Ecol. 39 (6), 1946–1955. doi:10.13292/j.1000-4890.202006.029
Olander, L. P., and Vitousek, P. M. (2000). Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 49, 175–191. doi:10.1023/A:1006316117817
Ordoñez, J. C., van Bodegom, P. M., Witte, J. P. M., Wright, I. J., Reich, P. B., and Aerts, R. (2009). A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob. Ecol. Biogeogr. 18, 137–149. doi:10.1111/j.1466-8238.2008.00441.x
Pan, J. W., Guo, Q. Q., Sun, X. G., and Gao, C. (2020). Contents and stoichiometric characteristics of C, N, P and K under different stand ages of Pinus massoniana plantations. Plant nutri. Fert. Sci. 26 (4), 746–756. doi:10.11674/zwyf.19272
Peichl, M., Leava, N. A., and Kiely, G. (2012). Above-and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland. Plant Soil 350, 281–296. doi:10.1007/s11104-011-0905-9
Poorter, L., and Bongers, F. (2006). Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 87 (7), 1733–1743. doi:10.1890/0012-9658(2006)87[1733:ltagpo]2.0.co;2
Qin, J., and Shangguan, Z. P. (2019). Effects of forest types on leaf functional traits and their interrelationships of Pinus massoniana coniferous and broad-leaved mixed forests in the subtropical mountain, Southeastern China. Ecol. Evol. 9 (12), 6922–6932. doi:10.1002/ece3.5259
Qin, Y. Y., Feng, Q., Nicholas, M. H., and Cao, J. J. (2016). Variation in soil organic carbon by slope aspect in the middle of the Qilian Mountains in the upper Heihe River Basin, China. Catena 147, 308–314. doi:10.1016/j.catena.2016.07.025
Reich, P. B., and Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. PNAS 101 (30), 11001–11006. doi:10.1073/pnas.0403588101
Reich, P. B., Walters, M. B., and Ellsworth, D. S. (1997). From tropics to tundra: Global convergence in plant functioning. PNAS 94, 13730–13734. doi:10.1073/pnas.94.25.13730
Ren, S. J., Yu, G. R., Jiang, C. M., Fang, H. J., and Sun, X. M. (2012). Stoichiometric characteristics of leaf carbon, nitrogen, and phosphorus of 102 dominant species in forest ecosystems along the North-South Transect of East China. Chin. J. Appl. Ecol. 23 (3), 581–586. doi:10.13278/j.1001-9332.2012.0111
Rong, Q. Q., Liu, J. T., Cai, Y. P., Lu, Z. H., Zhao, Z. Z., Yue, W. C., et al. (2015). Leaf carbon, nitrogen and phosphorus stoichiometry of Tamarix chinensis, Lour. In the Laizhou Bay coastal wetland, China. Ecol. Eng. 76, 57–65. doi:10.1016/j.ecoleng.2014.03.002
Rumpel, C., and Kogel-Knabner, I. (2010). Deep soil organic matter—A key but poorly understood component of terrestrial C cycle. Plant Soil 338, 143–158. doi:10.1007/s11104-010-0391-5
Sardans, J., Rivas-Ubach, A., and Penuelas, J. (2012). The C: N: P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspective Plant Ecol. Evol. Syst. 14, 33–47. doi:10.1016/j.ppees.2011.08.002
Sardans, J., Janssens, I. A., Alonso, R., Veresoglou, S. D., Rillig, M. C., Sanders, T. G. M., et al. (2015). Foliar elemental composition of European forest tree species associated with evolutionary traits and present environmental and competitive conditions. Glob. Ecol. Biogeogr. 24, 240–255. doi:10.1111/geb.12253
Shi, S. W., Peng, C. H., Wang, M., Zhu, Q. A., Yang, G., Yang, Y. Z., et al. (2016). A global meta-analysis of changes in soil carbon, nitrogen, phosphorus and sulfur, and stoichiometric shifts after forestation. Plant Soil 407 (1/2), 323–340. doi:10.1007/s11104-016-2889-y
Song, S. M., Zhou, Y., and Zhang, J. (2021). Ecological stoichimetry of leaf carbon, nitrogen and phosphorus of Cunninghamia lanceolata plantations in different site types. Nat. Sci. Ed. 49 (6), 38–53. doi:10.13207/j.cnki.jnwafu.2012.06.005
Sterck, F. J., Poorter, L., and Schieving, F. (2006). Leaf traits determine the growth survival trade-off across rain forest tree species. Am. Nat. 167 (5), 758–765. doi:10.1086/503056
Sternberg, M., and Shoshany, M. (2010). Influence of slope aspect on mediterranean woody formations: Comparison of a semiarid and an arid site in Israel. Ecol. Res. 16 (2), 335–345. doi:10.1046/j.1440-1703.2001.00393.x
Sterner, R. W., and Elser, J. J. (2002). Ecological stoichiometry: The biology of elements from molecules to the biosphere. Princeton: Princeton University Press.
Thomas, S. C., and Martin, A. R. (2012). Carbon content of tree tissues: A synthesis. Forests 3, 332–352. doi:10.3390/f3020332
Tian, H. Q., Chen, G. S., Zhang, C., Melillo, J. M., and Hall, C. A. S. (2010). Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 98, 139–151. doi:10.1007/s10533-009-9382-0
Tian, D., Kattge, J., Chen, Y. H., Han, W. X., Luo, Y. K., He, J. S., et al. (2019). A global database of paired leaf nitrogen and phosphorus concentrations of terrestrial plants. Ecology 100, e02812. doi:10.1002/ecy.2812
Vesterdal, L., Schmidt, I. K., Callesen, I., Nilsson, L. O., and Gundersen, P. (2008). Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 255 (1), 35–48. doi:10.1016/j.foreco.2007.08.015
Wang, S. Q., and Yu, G. R. (2008). Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements. Acta Ecol. Sin. 28 (8), 3937–3947.
Wang, K., Zhang, R. S., Song, L. N., Yan, T., and Na, E. (2021). Comparison of C:N:P stoichiometry in the plant-litter-soil system between poplar and elm plantations in the Horqin Sandy Land, China. Front. Plant Sci. 12, 655517. doi:10.3389/fpls.2021.655517
Wang, Y. H., Zhou, J. G., and Fu, Y. H. (2022). Effects of stand ages on C, N and P stoichiometry of Pinus massoniana secondary forests in Wuling Mountain area of Chongqing. Acta Ecol. Sin. 42 (23), 9537–9547. doi:10.5846/stxb202201100093
Wright, I. J., Reich, P. B., Cornelissen, J. H. C., Falster, D. S., Garnier, E., Hikosaka, K., et al. (2005). Assessing the generality of global leaf trait relationships. New Phytol. 166, 485–496. doi:10.1111/j.1469-8137.2005.01349.x
Wu, X., Niu, Y., Xun, M., Jin, J., Tang, Y., and Chen, Y. (2021). Soil carbon, nitrogen, and phosphorus storages and their stoichiometry due to mixed afforestation with Hippophae rhamnoides in the Loess hilly region, China. Forests 12, 1718. doi:10.3390/f12121718
Xu, X. F., Hui, D. F., King, A. W., Song, X., Thornton, P. E., and Zhang, L. H. (2015). Convergence of microbial assimilations of soil carbon, nitrogen, phosphorus, and sulfur in terrestrial ecosystems. Sci. Rep. 5, 17445. doi:10.1038/srep17445
Yang, D. X., Song, L., and Jin, G. Z. (2019). The soil C:N:P stoichiometry is more sensitive than the leaf C:N:P stoichiometry to nitrogen addition: A four-year nitrogen addition experiment in a Pinus koraiensis plantation. Plant Soil 442 (5), 183–198. doi:10.1007/s11104-019-04165-z
Yang, J., El-Kassaby, Y. A., and Guan, W. B. (2020). The effect of slope aspect on vegetation attributes in a mountainous dry valley, southwest China. Sci. Rep. 10, 16465. doi:10.1038/s41598-020-73496-0
Yang, Z. Q., Xia, H., Tan, J. H., Feng, Y. H., and Huang, Y. L. (2020). Selection of superior families of Pinus massoniana in southern China for large-diameter construction timber. J. For. Res. 31, 475–484. doi:10.1007/s11676-018-0815-2
Yimer, F., Ledin, S., and Abdelkadir, A. (2006). Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetation in the Bale Mountains, Ethiopia. Geoderma 135, 335–344. doi:10.1016/j.geoderma.2006.01.005
Zhang, Z. S., Song, X. L., Lu, X. G., and Xue, Z. S. (2013). Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: Influences of vegetation coverage, plant communities, geomorphology, and seawalls. J. Soil Sediment. 13 (6), 1043–1051. doi:10.1007/s11368-013-0693-3
Zhang, J. H., Zhao, N., Liu, C. C., Yang, H., Li, M. L., Yu, G. R., et al. (2018). C:N:P stoichiometry in China’s forests: From organs to ecosystems. Funct. Ecol. 32 (1), 50–60. doi:10.1111/1365-2435.12979
Zhang, J. H., Li, M. X., Xu, L., Zhu, J. X., Dai, G. H., and He, N. P. (2021). C:N:P stoichiometry in terrestrial ecosystems in China. Sci. Total Environ. 795, 148849. doi:10.1016/j.scitotenv.2021.148849
Zhang, Y., Li, X., Song, S., Zhou, Y., and Zhang, J. (2021). Characteristics of microbial biomass during the decomposition of mixed foliage litter from Pinus massoniana and broadleaved tree species. Ecol. Environ. Sci. 30 (4), 681–690. doi:10.16258/j.cnki.1674-5906.2021.04.003
Zhao, F. Z., Sun, J., Ren, C. J., Kang, D., Deng, J., Han, X. H., et al. (2015). Land use change influences soil C, N, and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci. Rep. 5, 10195. doi:10.1038/srep10195
Keywords: slope aspect, C, N, and P stoichiometry, Pinus massoniana, mixed forest, north subtropical
Citation: Qin J, Liu Y, Bi Q, Chen Z and Zhang B (2023) Response of leaf and soil C, N and P stoichiometry in different Pinus massoniana forest types to slope aspect in the Dabie mountains region of North subtropical, China. Front. Environ. Sci. 11:1148986. doi: 10.3389/fenvs.2023.1148986
Received: 20 January 2023; Accepted: 02 May 2023;
Published: 15 May 2023.
Edited by:
Dafeng Hui, Tennessee State University, United StatesReviewed by:
Dong Wang, Henan University, ChinaJianping Li, Ningxia University, China
Kaibo Wang, Chinese Academy of Sciences, China
Copyright © 2023 Qin, Liu, Bi, Chen and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan Qin, cWpqODE0QDEyNi5jb20=